Embed Size (px)
Citation preview

J. Exp. Mar. Biol. EcoL, 159 (1992) 1-13 © 1992 Elsevier Science Publishers BV. All rights reserved 0022-0981/92/$05.00
JEMBE 01767
Alkaline phosphatase activity in Porphyra umbilicalis (L.) Kutzing
I. Hernfindez a, F.X. Niell a and J.A. Fern f indez b
" Departamento de Ecologia; b Departamento de Fisiologia Vegetal, Facultad de Ciencias, Universidad de MMaga. Campus Universitario de Teatinos. MMaga. Spain
(Received 24 January 1992; accepted 6 February 1992)
Abstract: External alkaline phosphatase activity (APA) has been measured in Porphyra umbilicalis (L.) Kutzing over an ecological range of temperature, pH, salinity, photon irradiance and external phosphate. Temperature, pH, salinity and external phosphate caused significant changes in APA. On the contrary, photon irradiance caused no significant effect. In the range of temperature assayed, maximum APA was 249 izmol pNP g dry wt - ' h - ~ at 30 ° C. With regard to pH, maximum activity occurred at pH 8.8 (249/~mol pNP g dry wt- i h - I), whereas varying salinity, maximum enzymatic activity (270/~mol pNP g dry wt- ~ h - z)
o/ took place at 43.75/°°. As expected, there was an inverse relationship between APA and external phosphate concentration. There is a minimum constant activity at phosphate concentration over 14/~M and a final inhibition of 71 ~o in enzymatic activity.
Key words: Alkaline phosphatase; pH; Inorganic phosphate; lrradiance; Organic phosphate; Salinity; Tem- perature
INTRODUCTION
A dynamic model of the transformation that phosphorus undergoes in aquatic ec- osystems includes dissolved organic phosphorus (DOP) as one of the most important fractions (Taft et al., 1977; Siuda et al., 1981; Francko, 1984; Boavida & Heath, 1986; Heath, 1986; Heath & Edinger, 1990). By means of external enzymatic hydrolysis, seaweeds can make use of phosphate monoesters as a source of orthophosphate.
Alkaline phosphatase is an enzyme that cleaves inorganic phosphate from organic phosphomonoesters, a fraction of the external DOP pool, so algae can have an addi- tional source of orthophosphate (Reichardt et al., 1967; Fitzgerald & Nelson, 1966; Berman, 1970; Reichardt, 1971; Heath & Cooke, 1975; Siuda et al., 1981; Boavida & Heath, 1986; Pick, 1987; Matavulj et al., 1989). This enzyme could play a relevant role in the phosphorus supply during phc, sphate depletion. The enzyme is generally believed to be located on the outer surface of the cell (Kuenzler, 1965; Kuenzler & Perras, 1965; Cembella et al., 1984), either at the cell wall (Brandes & Elston, 1956), at the plasma membrane (Patni et al., 1984) or at the periplasmic region (Matagne et al., 1976, Marco
Correspondence address: I. He:nfindez, Departamento de Ecologifi, Facultad de Ciencias, Universidad de Mfilaga, Campus UniversitaEo de Teatinos, 29071 Malaga, Spain.

2 I. HERNANDEZ ET AL.
& Orris, 1988). High phosphatase activity is generally thought to indicate conditions of phosphate deficiency (Fitzgerald & Nelson, 1966; Perry, 1972; Jones, 1972; Healey, 1973; Sakshaug et al., 1984; Boavida & Heath, 1986).
Alkaline phosphatase activity in aquatic macrophytes is scarcely mentioned, espe- cially in the marine environment. Since Walther & Fries (1976) identified alkaline phosphatase from macroalgae, Atkinson (1987) measured activities in some benthic algae and communities of reef organisms, Lapointe & O'Connell (1988) measured al- kaline phosphatase activity of some macroalgae in Bermuda's inshore waters, whereas Weich & Gran~li (1989) detected extracellular alkaline phosphatase activity in Ulva lactuca, studying the dependence of alkaline phosphatase activity on the P status, ex- ternal phosphate concentration, light, pH and bacteria. Nevertheless, at the moment there is no clear characterization of the enzyme in aquatic macrophytes. It is essen- tial to design an adequate assay method which permits incubation of the whole plant under controlled conditions, in order to plot enzymatic activities versus environmen- tal variables and discuss the results from the standpoint of the ecology of aquatic macrophytes.
In the present study, external alkaline phosphatase activity was investigated in Porphyra umbilicalis (L.) Kutzing. This seaweed was selected because it is a model for the assay of APA in different conditions. The alga lives in the supralittoral zone and may be subjected to a broad range of variation of environmental factors (temperature, pH, salinity, light and external orthophosphate). The influence of these factors on APA were studied, agreeing with their probable range of variations in natural conditions in order to deterr, line the dependence of the enzyme on these factors.
MATERIALS AND METHODS
Collection and pretreatment. This study was carried out during the period December 1989 to April 1990, the season of presence of Porphyra umbilicalis on the coast of southern Spain. The algae live on flat rocks, in the supralittoral zone which usually receives seawater only from wave splashes. The plants are frequently exposed to the atmosphere in the case of a calm sea, since no tidal variation exists in this area of the Mediterranean Sea, and just occasional storms could cover such populations. After collection, plants were kept cool and transported to the laboratory in an icebox (30 min). The algae were maintained (5 days maximum) in renewed cultures of aerated, filtered natural seawater (pH 8.2) at a constant temperature of 15 °C and photon flux density of 150 #tool photons m -l s -1 (Sylvania, F 20W/D-RS).
Measurement of alkaline phosphatase activity. Alkaline phosphatase activity (APA) was determined in three replicate samples using a modification of the method of Reichardt et al. (1967), which uses p-nitrophenyl phosphate (pNPP)as a dissolved organic sub- strate. 50ml of 10-3M pNPP was mixed with an equal volume of 0.1 M

APA IN PORPHYRA UMBILICALIS 3
Tris-hydroxymethyl-aminomethan hydrochloric acid buffer, pH 8.3. Both solutions were made up using artificial seawater, of 35~oo ~a!inity (Kalle, 1945), filtered through Whatman GF-F. APA was assayed at 25 ° C, placing the wh~.!e macrophyte (0.25-0.5 g wet wt) in an incubation chamber "vidra FOC E-12563", a spherical flask of 135 ml in volume with a water jacket which permits thermoregulation. Air was bubbled con- tinuously during the assay to oxygenate and stir the medium. Samples were taken at 0, 20, 40 and 60 min and the absorbance read at 410 nm against a blank (buffer and substrate solutions without alga) in a Beckman DU-7 Spectrophotometer. Changes of color in the incubation solution were directly related to the enzymatic hydrolysis of pNPP. The molar coefficient of pNP was determined in the range of pH 7-9, the ex- pected range of variation of seawater pH. In all cases solutions were prepared in the range from 2.5 10 -7 to 2.5 10-SM of paranitrophenol. Enzymatic activity was ex- pressed as #mol of paranitrophenol (pNP) released g dry wt -1 h -l
This assay method permits the incubation of the whole macrophyte, so it is possi- ble to estimate the overall enzymatic activity. With artificial seawater as the experi- mental medium it is possible to reproduce the osmolarity of natural seawater and avoid undesired deviations from natural conditions.
Experimental set-up. To estimate the kinetic parameters of Porphyra umbilicalis, pNPP concentrations ranging from 5 10-6 to 2.5 10-4 M were used. A Hanes transformation (S/v vs. S) (Hanes, 1932) of the Michaelis-Menten equation was used in order to calculate the Michaelis constant (KM) and maximum velocity (Vm) of the enzyme. These values were computed graphically by linear regression on a line constructed with the values obtained in the assay.
Para~neters concerning the influence of tcmpcrature il~ enzymatic reactions were calculated from the Arrhenius equation (Price & Stevens, 1982) in a temperature range of 10-30 °C.
In order to elucidate the influence of pH on enzymatic activity, different Tris-hydroxymethyl-aminomethan hydrochloric acid buffers were prepared in the range of 7-9.
Different salinities were obtained by making up solutions in water from 0%0 salin- ity (distilled water) to 52.5%0, varying quantities of all reagents in the artificial seawa- ter.
Influence of photon irradiance on APA was studied by illuminating the incubation chamber with a slide projector in which different filters to vary the light intensity were inserted. Photon irradiance ranged from dark to 1500 #mol photons m-l s-
The influence of P enrichment on APA was studied by pre-incubating the algae for 5 days in 400 ml of natural seawater enriched with a different amount of K2HPO 4 as a source of inorganic orthophosphate (Pi). The final concentration of Pi added ranged from 0 to 28.8 #M. Water was renewed in the cultures twice daily.
Differences between treatments have been statistically tested for significance (test for equality of slopes, Sokal & Rohlf, 1981), ~ = 0.05.
4 I. HERNANDEZ ET AL.
16
14
8'
6'
4"
2 ~
0 0
i i i i i i
50 100 150 200 250 300
S (I~M)
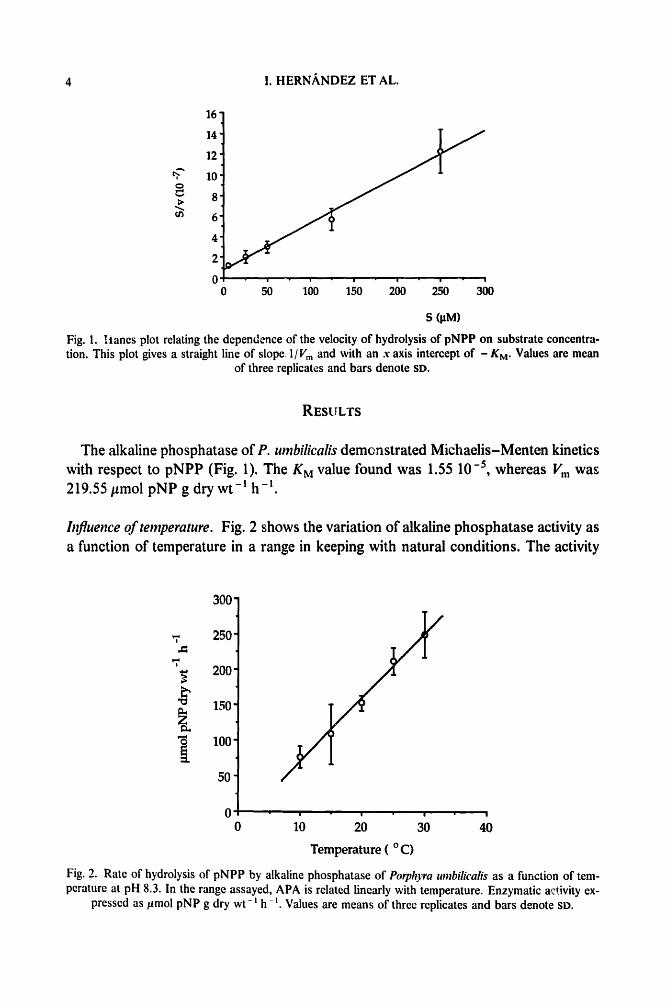
Fig. 1. Iianes plot relating the dependence of the velocity of hydrolysis of pNPP on substrate concentra- tion. This plot gives a straight line of slope, 1/Vm and with an x axis intercept of - KM. Values are mean
of three replicates and bars denote SD.
RESULTS
The alkaline phosphatase of P. umbilicalis demonstrated Michaelis-Menten kinetics with respect to pNPP (Fig. 1). The KM value found was 1.55 10 -5, whereas Vm was 219.55 #mol pNP g dry wt -m h -t.
h~fluence of temperature. Fig. 2 shows the variation of alkaline phosphatase activity as a function of temperature in a range in keeping with natural conditions. The activity
>
300
250
200
150
100"
50
| g | |
0 10 20 30 40
Temperature (°C)
Fig. 2. Rate of hydrolysis of pNPP by alkaline phosphatase of Porpto,ra umbilicalis as a function of tem- perature at pH 8.3. in the range assayed, APA is related linearly with temperature. Enzymatic activity ex-
pressed as ~lmol pNP g dry wt-~ h-~. Values are means of three replicates and bars denote SD.

APA IN PORPHYRA UMBILICALIS 5
increased linearly in the range analyzed, so no inhibition was observed at temperature values up to 30 °C. Maximum activity (249/~mol pNP g dry wt -I h -l) was found at 30 °C, the maximum temperature tested. We found a linear regression between APA and temperature. The regression line fitted ( r2 -0 .91)was A P A = - 19.80+8.95 T, with 0.97 and 20.64 being the standard errors of the slope and the y intercept, respec- tively. The Arrhenius plot (log Vm- Ea/R x 1/T+ cte) revealed an activation energy (Ea) of 43.31 kJ mol -l. That gives a Qlo value of 1.78.
Influence ofpH. Fig. 3 shows that the rate at which P. umbilicalis hydrolyzed pNPP was markedly pH dependent. Maximum activity occurred at pH 8.8 and resulted in 249 #mol pNP g dry wt-~ h -l. However activity existed in all the range of pH assayed (pH 7-9).
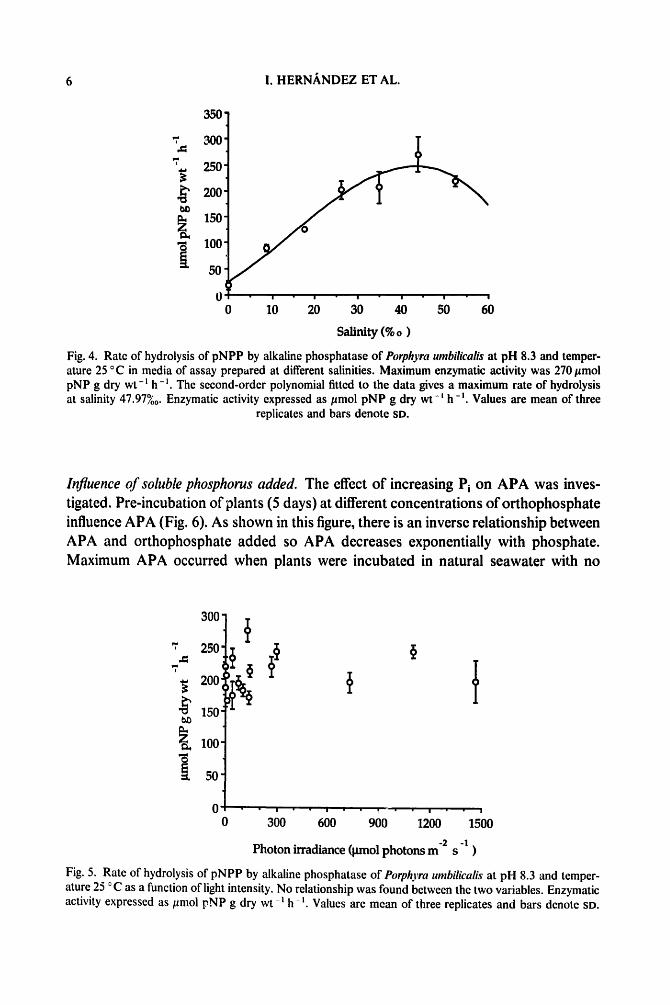
h~uence of salinity. Artificial seawater used in the enzymatic assays (salinity 35%0) avoids large deviations from natural conditions. Salinity significantly changed alkaline phosphatase activity in P. umbilicalis (Fig. 4). APA increased with salinity until reach- ing a maximum activity of 270/~mol pNP g dry wt-l h - ~ around 45?/00 whilst higher salinity decreased enzymatic activity. Distilled water and low salinities inhibited the enzymatic activity drastically.
Influence of photon irradiance. High APA was found in the whole range of photon irradiance in which enzymatic activity was assayed, but no correlation between APA and photon irradiance was observed (Fig. 5), so it seems that in P. umbilicalis APA is not influenced significantly by light intensity.
300"
'7 250'
|
200'
150
100
50"
. . . . ! . . . . | . . . . | . . . . | • . . ~ . . . . !
6,5 7,0 7,5 8,0 8,5 9,0 9,5 pH
Fig. 3. Rate of hydrolysis of pNPP by alkaline phosphatase of Porphyra umbilicalis as a function of pH at 25 °C. Enzymatic activity expressed as #mol pNP g dry wt -~ h- ' . Maxi:~ am rate is 260/lmol pNP g dry
wt -~ h -~ at pH 8.3. Values are mean of three replicates and bars denote sD.
350
0 0
-r, 300
250
200
150
100
5O
! ! | ! | i
10 20 30 40 50 60
6 I. HERNANDEZ ET AL.
Salinity (%0)
Fig. 4. Rate of hydrolysis of pNPP by alkaline phosphatase of Porphyra umbilicalis at pH 8.3 and temper- ature 25 °C in media of assay prepared at different salinities. Maximum enzymatic activity was 270 #mol pNP g dry wt -~ h -~. The second-order polynomial fitted to the data gives a maximum rate of hydrolysis at salinity 47.97%0. Enzymatic activity expressed as/~mol pNP g dry wt- t h- '. Values are mean of three
replicates and bars denote SD.
Influence of soluble phosphorus added. The effect of increasing Pi on APA was inves- tigated. Pre-incubation of plants (5 days) at different concentrations of orthophosphate influence APA (Fig. 6). As shown in this figure, there is an inverse relationship between APA and orthophosphate added so APA decreases exponentially with phosphate. Maximum APA occurred when plants were incubated in natural seawater with no
300"
" 250'
'7,
200:
150'
~, 100"
5o
Fig. 5. Rate of hydrolysis of pNPP by alkaline phosphatase of Porphyra umbilicalis at pH 8.3 and temper- ature 25 -'C as a function of light intensity. No relationship was found between the two variables. Enzymatic activity expressed as/mlol pNP g dry wt- ~ h- ~. Values are mean of three replicates and bars denote so.
! ! . ! |
0 300 600 900 1200 1500
-2 -1 Photon irradiance (lamol photons m s )

APA IN PORPHYRA UMBILICALIS 7
400'
'7, 3501
-, 3OO
,~, 250
200" ,
I~)" I ,
100 ", ,~
5O t
X
1 } I . . . . . . . , . - , . . , • • , . . ,
0 3 6 9 12 15 18 21 24 27 30 3-
PO 4 added (pM)
Fig. 6. Inhibitory effect of increasing inorganic phosphate concentration upon APA in Porphyra umbilicalis. Seaweeds were incubated for :3 days in different regimes of external orthophosphate. Maximum rate of hydrolysis of pNPP takes place when no phosphorus was added at all. Final APA was ~71~o of the maximum rate. Enzymatic activity expressed as/~mol pNP g dry wt-~ h- ~. Values are mean of three rep-
licates and bars denote SD.
addition of phosphate. The data shown in the figure were fitted to a "single exponential decay plus offset" model:
APA = APAe + APAm • e - he, (1)
where APAc (85.85 jumol pNP g dry wt -~ h -~, SE 37.55) is the APA not affected by the inorganic phosphate addition, APAm is the maximum phosphatase activity (213.23/~mol pNP g dry wt -~ h -Z, SE 43.91), b is the decay coefficient (0.25 #M -1, SE 0.14) and P is the amount of inorganic phosphate added. There is a strong inhibi- tion of the enzymatic activity for the range of 0-3 #M added, and then a smooth de- cline in activity to 28.8 i tM. A final inhibition of 71% in APA (external phosphate concentration ~ 14/tM) was observed, whereas for 50% inhibition ~4.8/~M of or- thophosphate addition was required.
DISCUSSION
Alkaline phosphatase from P. umbilicalis seems to be well adapted for the physical and chemical environment of the organism, so it seems necessary to study how the enzyme responded to different environmental changes.
The Hanes plot showed that the APA of P. umbilicalis, as expected, follows Michaelis-Menten kinetics with respect to pNPP. The Ku (1.55 10 -5 M) is within the range of values given by Chrost & Overveck (1987) for freshwater algae (10 -6 tO
10 -4 M). Also it is very similar to other values found in some phytoplankton and

8 I. HERNANDEZ ET AL.
bacterioplankton species, e.g., 1.6 10-SM for Oscillatoria sp. (Marco & Orfs, 1988) and 4.34 10 -5 M for Calothrix par,~tina (Grainger et al., 1989).
Temperature influences alkaline phosphatase activity significantly (Fig. 2). The ac- tivity increased over the range assayed (10-30 °C)without loss of catalytic activity. A possible explanation is based on the fact that P. umbilicalis lives on rocks, from De- cember to April. Algae can remain exposed directly to the sunlight for several hours, only slightly splashed by waves and reaching temperatures up to 25 °C or even more (average of maximum from 16.2 to 24.2 o C, data from the Meteorological Institute of Mfilaga). The experiments shown in this paper were performed without previous ad- aptation of the specimens to the assay temperature. In no case was a decrease of APA caused by thermal stress observed, and no change in the APA was detected during the time course of experiments after changing the algae from the culture room (15 ° C) to the assay medium. We found a Q~0 value of <2, which suggests an adaptation to cold conditions so that essential metabolic rea~.tions related with phosphate metabolism are not slowed down too much when temperature decreases (Price & Stevens, 1982). Porphyra is able to resist high temperatures and prevent metabolic inhibition at low temperatures since APA is just reduced <50% when temperature decreases 10 °C (Fig. 2). The value of Qt0 found in P. umbilicalis is in the middle of the range between the 1.6 found by Pettersson & Jansson (!978) in unfiltered lake water and 2.0, reported by Healy & Hendzel (1979) for Anabaena variabilis. A previous reference of Reichardt et al. (1967) gives a temperature optimum of 27 °C and Ea of 69 kJ mol -t also for alkaline phosphatase dissolved in lake water. This is equivalent to a Qto value ~2.5.
The rate at which P. umbilicalis hydrolyzed pNPP was markedly dependent on pH. Maximum APA takes place at pH 8.8 (Fig. 3). This value may be easily reached in the case of direct sunlight exposure, when active photosynthesis is occurring; that means a maximum phosphorus supply in cases of high rates of carbon fixation. In P. umbil- icalis, tl:e external pH range may be variable, reaching values of ~ 9 in conditions of high irradiance and direct exposure to sunlight, and values ~ 7 in the case of strong rainfall; conditions typical of Meditei'ranean coast during the winter. This dramatic effect might be compensated by relatively ' high APA at pH values of ~ 7. Most stud- ies in fresh and seawater have been made on alkaline phosphatases with pH optima between 8 and 9 (Pettersson & Jansson, 1978; Reichardt et al., 1967; Reichardt, 1971; Siuda, 1984). In P. umbilicalis the optimum pH is less alkaline than the optimum found in most of the phytoplankton and bacterioplankton species in which optimum pH for alkaline phosphatase has been reported (Kuenzler & Perras, 1965; Patni et al., 1977; Flynn et al., 1986; Gibson & Whitton, 1987; Chan & Dean, 1987; Marco & Orris, 1988; Mahasneh et al., 1990).
Salinity significantly influenced APA (Fig. 4). Due to the features mentioned above, P. umbilicalis is subjected to strcag and unpredictable salinity fluctuations. These fluc- tuations extend from high values when there is desiccation (direct exposure), to strong dilution when torrential downpours occur. Owing to the brief time of assay (1 h), low

APA IN PORPHYRA UMBILICALIS 9
values of enzymatic activity at low salinities are probably attributed to inhibition rather than suppression of enzyme synthesis.
High values of salinity decreased APA (Fig. 4). In the case of exposure and high irradiance this effect could be compensated by an increase of external pH by photo- synthesis (which determines a greater APA, Fig. 3), due to OH-disposal (Lucas & Berry, 1985).
It is known that APA increases with the ionic strength of the test solution (Wilson et al., 1964). A future study will elucidate whether the enhancement in APA with sa- linity is due t, the iL~fluencc of salinity itself or there is a significant influence of some divalent cations on the enzymatic activity. Mg 2+ is added as an activator of the en- zyme reaction (Kuenzler & Perras, 1965; Reichardt ctal., 1967; Francko, 1984; At- kinson, 1987; Boavida & Heath, 1988; Wynne & Rhee, 1988) and could be the divalent cation responsible for the observed increase in APA. Hino (1988) reported Mg 2+ ion as an activator in Melossira ambigua. Ca 2 + has also been considered as an activator of the alkaline phosphatase (Hino, 1988; Mahasneh et al., 1990).
The fact that the enzymatic activity is dependent on salinity could be of ecological significance in cases of exposure and high irradiance. In those conditions, an extra supply of phosphorus from the hydrolysis of phosphomonoesters may balance states of low nutrient concentration as stress could force an additional demand of nutrients.
Alkaline phosphatase activity has been assayed in a broad range of photon irradiancc (Fig. 5). The enzymatic activity was high in all the range assayed, but no significant influence of photon irradiance was found, so in P. umbilicalis APA seems to be inde- pendent of photon irradiance. These results are in agreement with Huber & Hamel (1985a,b) whose study with Nodularia spumigena cultures led them to the conclusion that APA was not light related. On the contrary, Wynne & Rhee (1988) found that intracellular and extracellular APA varied with intensity and quality of light in Dunaliella tertiolecta, Thalassiosira pseudonana, Phaeodactylum tricornutum and Prymnesium parvum. Evidence of influence of light on APA has also been postulated in Selenastrum capricornutum (Klotz, 1985), Pyrocystis noctiluca (Rivkin & Swift, 1979) and Ulva lactuca (Weich & Gran61i, 1989). As P. umbilicalis lives on fiat rocks and is usually exposed to sunlight, algae receive a broad range of photon irradiance. This fact could be a factor of selection of the enzyme to be independent of photon irradiance.
Fig. 6 shows an inverse relationship between inorganic phosphorus and APA. This relationship has been demonstrated by other workers (Perry, 1972; Pettersson, 1980; Siuda, 1984; Siuda& Chrost, 1987; Hirayama et al., 1989; Veldhuis & Admiraal, 1987), with Pi as a competitive inhibitor (Reichardt et al., 1967; Pick, 1987). A negative ex- ponential curve fits the data shown in Fig. 5. APA in P. umbilicalis is partially sup- pressed by addition of phosphorus. A final inhibition of ~ 71% in 5 days of culture was found but this occurs with an orthophosphate concentration probably higher than maximum soluble phosphorus levels present "in situ". We previously found that var- ious days of pre-incubation were necessary to detect any inhibition in APA. Similar results have been found by Lapointe & O'Connell (1988) in Cladophora prolifera. In this

10 !, I-IERNANDEZ ET AL.
species, enrichment with 16 ~uM Pi during 6 days caused a decrease of ~ 357~, in APA. Reichardt et al. (1967) reported that the phosphatase activity of some cyanobacteria was inhibited by 3.1 llM of orthophosphate, while Huber & Hamcl (1985b), working wilh Nodularia spumigena noticed a very sharp drop in APA in the range of 0-0.5 I~M Pi added and then a linear decline in activity to 10.6/~M. In U. lacmca, Weich & Gran61i (1989), demonstrated that the addition of 50 #M phosphate reduced but did not inhibit APA, suggesting an internal regulation of APA activity in this species. In natural waters, some of the levels found by other workers (Reichardt et al., 1967; Perry, 1972; Jones, 1972; Siuda ct al., 1981: Pick, 1987; Davies & Smith, 1988) vary greatly; e.g. I ILM Pi addition required for 50~o inhibition in Lake Ontario (Pick, 1987) or 32 pM necessary for 800~ inhibition in Lake District (Jones, 1972). This may be due to the different composition in planktonic species, because APA in some of them, specially C/trysophytes and Cryptomonadv is not easily inhibited by additions of ortho- phosphate (Kuenzler, 1965; Aaronson & Patni, 1976; Stewart & Wetzel, 1982; Pick, 1987). Disparities in the surface areas of the species also have been proposed as an explanation for this fact (Aaronson & Patni, 1976). The long-term effect of Pi upon APA in P. umbilicalis seems to be a suppression of enzyme synthesis rather than in- hibition, but this suppression is not total. This fact suggests the existence of two phosphatases working at pH 8.3: one adaptative phosphatase whose synthesis rate is regulated by changes in the orthophosphate concentration, and a constitutive one, independent of the external phosphate levels ~see Fig. 6).
Research to compare different species is being done, because APA seems to play different specific roles in each species or group ~,,f species. For instance, Lapoinic & O'Connell (1989) Ibund values of APA up to 100-fi, ld lower in some tropical species, and Atkinson {1987)reported values of APA for some benthic algae in the same order ol" tnagnilttde that! the data presented here fi~t' P. tmlhilicalis.
The results obtained in this paper suggested good correlation belween the variables assayed and APA, and il seems highly probable Ihat there is some synergislic effect between them. These etfccts must be studied in optimal and realistic combinations in ~.rder to delermine simultaneous influence on the enzymatic activity.
ACKNOWLEDGEM ENrS
This work has been financially supported by grant i1" PB86/1)677 of the Spanish Commission for Science and Technology (CICYT). I. Hcrn~indez has received a grant fiom the Spanish Ministry of Educalion and Science, We wish to thank R,T. Heath for valuable criticism on earlier versions of the manuscript.

APA IN PORPHYRA UMBILICALIS 11
REFERENCES
Aaronson, S. & J. Patni, 1976. The role of surface and extracellular phosphatases in the phosphorus requirement of Ochromonas. Lironoi. Oceanogr., Vol. 21, pp. 838-845.
Atkinson, M.J., 1987. Alkaline phosphatase activity of coral reef benthos. Coral Reefs, Vol. 6, pp. 59-62. Berman, T., 1970. Alkaline phosphatase and phosphorus availability in Lake Kinneret. Limnol. Oceanogr.,
Vol. 15, pp. 663-674. Boavida, M.J. & R.T. Heath, 1986. Phosphatase activity on Chlamydomonas acidophyla Negoro (Volvocales,
Chiorophyceae). Phycologia, Vol. 25, pp. 400-404. Boavida, M.J. & R.T. Heath, 1988. Is alkaline phosphatase always important in phosphate regeneration?.
Arch. Hydrobiol., Vol. 111, pp. 507-518. Brandes, D. & R.N. Elston, 1956. An electron microscopical study of the histochemical localization of
alkaline phosphatase in the cell wall of Chlorella vulgaris. Nature, Vol. 177, pp. 274-275. Cembella, A.D., N.J. Antia & P.J. Harrison, 1983. The utilization of inorganic and organic phosphorous
compounds as nutrients by eukaryotic microalgae: A multidisciplinary perspective: Part 1. C. R. C. Crit. Rev. Microbiol., Vol. 10, pp. 317-391.
Chan, K. & A. C. R. Dean, 1987. Growth, phosphatase activity and antibiotic resistance of a cadmium re- sistant marine bacterium Pseudomonas marina growing in a chemostat culture. Microbios Lett., Vol. 35, pp. 57-65.
Chrost, R.J. & J. Overbeck, 1987. Kinetics of alkaline phosphatase activity and phosphorus availability for phytoplankton and bacterioplankton in Lake Plubsee (north German eutrophic lake). Microb. Ecol., Vol. 13, pp. 229-248.
Davies, A.G. & M.G. Smith, 1988. Alkaline phosphatase activity in the Western English Channel. J. Mar. Biol. Assoc. U.K., Vol. 68, pp. 239-250.
Fitzgerald, G.P. & T.C. Nelson, 1966. Extractive and enzymatic analysis for limiting of surplus phospho- rus in algae. J. Phycol., Vol. 2, pp. 32-37.
Flynn, K.J., H. Opik & P.J. Syrett, 1986. Localization of the alkaline phosphatase and 5'-nucleotida~e activities of the diatom Phaeodactylum tricornutum. J. Gen. Microbiol., Vol. 132, pp. 289-298.
Francko, D.A., 1984. Relationships between phosphorus functional classes and alkaline phosphatase ac- tivity in reservoir lakes. J. Freshwater Ecol., Vol. 2, pp. 541-547.
Gibson, M.T. & B.A. Whitton, 1987. Hairs, phosphatase activity and environmental chemistry in Stigeo- clonium, Chaetophora and Draparnaldia (Chaetophorales). Br. Phycol. J., Vol. 22, pp. 11-22.
Grainger, S.L.J., A. Peat, D.N. Tiwari & D.A. Whitton, 1989. Phosphomonoesterase activity of the cyanobacterium (blue-green alga) Caiothrix parietina. Microbios, Vol. 59, pp. 7-17.
Hanes, C.S., 1983. The effect of starch concentration upon the velocity of hydrolysis by the amylase of germinated barley. Biochem. J., Vol. 26, pp. 1406-1421.
Healey, F.P., 1973. Characteristics of phosphorus deficiency in Anabaena. J. Phycol., Vol. 9, pp. 383-394. Healey, F.P. & L.L. Hendzel, 1979. Fluorometric measurement of alkaline phosphatase activity in algae.
Freshwater BioL, Vol. 9, pp. 429-439. Heath, R.T., 1986. Dissolved organic phosphorus compounds: do they satisfy planktonic phosphate demand
in summer? Can. J. Fish. Aquat. Sci., Vol. 43, pp. 343-350. Heath, R.T. & G.D. Cooke, 1975. The significance of alkaline phosphatase in a eutrophic lake. Verh. hlt.
Ver. LimnoL, Vol. 19, pp. 959-965. Heath, R.T. & A.C. Edinger, 1990. Uptake of 32p-phosphoryl from glucose-6-phosphate by plankton in an
acid bog lake. Verh. h~t. Ver. LimnoL, Vol. 24, pp. 210-213. Hino, S., 1988. Fluctuations of algal alkaline phosphatase activity and the possible mechanisms of hydrolysis
of dissolved organic phosphorus in Lake Barato. Hydrobiologia, Vol. 157, pp. 77-84. Hirayama, K., T. Doma, N. Hamamura & T. Muramatsu, 1989. Role of alkaline phosphatase activity in
the growth of red tide organisms. In, Red tides: biology, environmental science and toxicology, edited by Okaichi et al., Elsevier Science Publishing, pp. 317-320.

12 I. HERNANDEZ ET AL.
Huber, A.L. & K. S. Hamel, 1985a. Phosphatase activities in relation to phosphorus nutrition in Nodularia spumigena (Cyanobacteriaceae). 2. Laboratory studies. Hydrobiologia, Vol. 123, pp. 81-88.
Huber, A. L. & K.S. Hamel, 1985b. Phosphatase activities in relation to phosphorus nutrition in Nodularia spwnigena (Cyanobacteriaceae). 1, Field studies. Hydrobiologia, Voi. 123, pp. 145-152.
Jones, J.G., 1972. Studies of freshwater micro-organisms: phosphatase activity in lakes of differing degrees of ~atriphication. J. Ecol., Vol. 60, pp. 777-791.
Kalle, K. (1945). In Chemicaloceanography, Vol. 2, edited by J. P. Riley & G. Skirrow, 1975, Academic Press, London.
Klotz, R.L., 1985. Influence of hght on the alkaline phosphatase activity of Selenastrum capricornutum (Clorophyceae) in streams. Can. J Fish. Aquat. Sci., Vol. 42, pp. 384-388.
Kuenzler, E.J., 1965. Glucose-6 phosphate utilization by marine algae. J. Phycol., Vol. 1, pp. 156-164. Kuenzler, E.J. & J.P. Perras, 1965. Phosphatases of marine algae. Biol. Bull., Vol. 128, pp. 277-284. Lapointe, B.E. & J. O'Connell, 1989. Nutrient-enhanced growth of Ciadophora prolifera in Harrington
Sound, Bermuda: Eutrophication of a confined, phosphorus limited marine ecosystem. Estuarine Coastal Shelf Sci., Vol. 28, pp. 347-360.
Lucas, W.J. & J.A. Berry, 1985. Inorganic carbon transport in aquatic photosynthetic organisms. Physiol. Plant., Vol. 65, pp. 539-543.
Mahasneh, 1. A., S.L.J. Grainger & B.A. Whitton, 1990. Influence on hair formation and phosphatase activities of the blue-green alga (C:/anobacterium) Calothrix viguieri D253. Br. Phycol. J., Vol. 25, pp. 25- 32.
Marco, E. & M.I. Orris, 1988. Alkaline phosphatase activity in two Cyanobacteria. Oyton, Vol. 48, pp. 27- 32.
Matagne, R. F., ~. Loppes & R. Deltour, 1976. Phosphatases of Chlamydomonas reinhardi: Biochemical and cytochemical approach with specific mutants. J. Bacteriol., Vol. 126, p. 937.
Matavulj, M., S. Gajin, M. Ebeznik, M. Bokorov & O. Petrovic, 1989. Phosphatase activity of water as a parameter of the river tisa water monitoring. Tiscia (Szeged), Vol. 23, pp. 29-36.
Patni, N.J., S.W. Dhawade & S. Aaronson, 1977. Extracellular phosphatase of Cldamydomonas remhardi and their regulation. J. Bacteriol., Vol. 130, pp. 205-211.
Perry, M.J., 1972. Alkaline phosphatase activity in subtropical Central North Pacific waters using a sen- sitive fluoromctric method. Mar. Biol., Vol. 15, pp. 113-119.
Pettersson, K., 1980. Alkaline phosphatase activity and algal surplus phosphorus as phosphorus-deficiency indicators in Lake Erken. Arch. Hydrobiol., Voi. 89, pp. 54-87.
Pettersson, K, & M. Janssc~n, 1978. Determination of phosphatase activity in lake water. A study of methods. I/erh. ltzt. Per. Lira,el., Vol. 20, pp. 1226-1230.
Pick, F.R., 1987. Interpretations of alkaline phosphatase activity in Lake Ontario. Can. J. Fish. Aquat. Sci., Vol, 44, pp. 2087-2094.
Price, N.C. & L. Stevens, 1982 Fundamentals ofenzymology. Oxford University Press, Oxford, 454 pp. Reichardt, W., 1971. Catalytic mobilization of phosphate in lake water and by Cyanophyta. Hydrobiologia,
Vol. 38, pp. 337-394. Rcichardt, W,, J. Overbeck & L. Steubing, 1967. Free dissolved enzymes in lake waters. Nature, Voi. 216,
pp. 1345-1347. Rivkin, R.B. & E. Swift, 1979. Diel and vertical patterns of alkaline phosphatase activity in oceanic
dinoflagellate Pj,racystis ,octiluca. Limnol. Oceanogr., Vol. 24, pp. 107-116. Sakshaug, E., E. Gran61i, M. Elbrachter & K. Hermann, 1984. Chemical composition and alkaline phos-
phatase activity of nutrient-saturated and P-deficient cells of four marine dinoflagellates. J. Exp. Mar. Biol. Ecol., Vol. 77, pp. 241-254.
Siuda, W., 1984. Phosphatases and their role in organic phosphorus transformations in natural waters. A review. Pol. Arch. Hydrobiol., Vol. 31, pp. 207-233.
Siuda, W. & R.J. Chr6st, 1987. The relationship between alkaline phosphatase activity and phosphate availability for phytoplankton and bacteria in eutrophic lakes. Acta Microbiol. Pol., Vol. 36, pp. 247-257.

APA IN PORPHYRA UMBILICALIS 13
Siuda, W., J. Chr6st, R. Wcislo & M. Krupka, 1982. Factors affecting alkaline phosphatase activity in a lake (short-term experiments). Acta Hydrobiol., Vol. 24, pp. 3-20.
Sokal, R.R. & F. J. Rohlf, 1981. Biometry. W.H. Freeman and Co., San Francisco, California, second edition, 859 pp.
Stewart, A.J. & R.G. Wetzel, 1982. Influence of dissolved humic materials on carbon assimdetion and alkaline phosphatase activity in natural algal-bacterial assemblages. Freshwater Biol., Vol. 12, pp. 369-380.
Taft, J. L., M. E. Loftus & W. R. Taylor, 1977. Phosphate uptake from phosphomonoesters by phytoplankton in the Chesapeake Bay. Limnol. Oceanogr., Vol. 22, pp. 1012-1021.
Walther, C. & L. Fries, 1976. Extracellular alkaline phosphatase in multicellular algae and their utilisation of glycerophosphate. Physiol. Plant., Vol. 36, pp. 118-122.
Weich, R.G. & E. Gran61i, 1989. Extracellular alkaline phosphatase activity in Ulva lactuca L. J. Exp. Mar. Biol. Ecol., Vol. 129, pp. 33-44.
Veldhuis, J.W. & W. Admiraal, 1987. Influence of phosphate depletion on the growth and colony forma- tion of Phaeocystis pouchetii. In, The eco-physiology of the colonial alga Phaeocysfis pouchetii, edited by M. J. W. Veldhuis, Drukkerij Van Dendcren, Groningcn, pp. 43-61.
Wilson, I.B., J. Dayan & K. Cyr, 1964. Some properties of alkaline phosphatase from Escherichia coli. J. Biol. Chem., Vol. 239, pp. 4182-4185.
Wynne, D. & G.-Y. Rhee, ! 988. Changes in alkaline phosphatase activity and phosphate uptake in P-limited phytoplankton, induced by light intensity and spectral quality. Hydrobiologia, Vol. 160, pp. 173-178.





























