Embed Size (px)
Citation preview

Amino Acid Catabolism: N
Copyright © 1999-2008 by Joyce J. Diwan. All rights reserved.
Molecular Biochemistry II
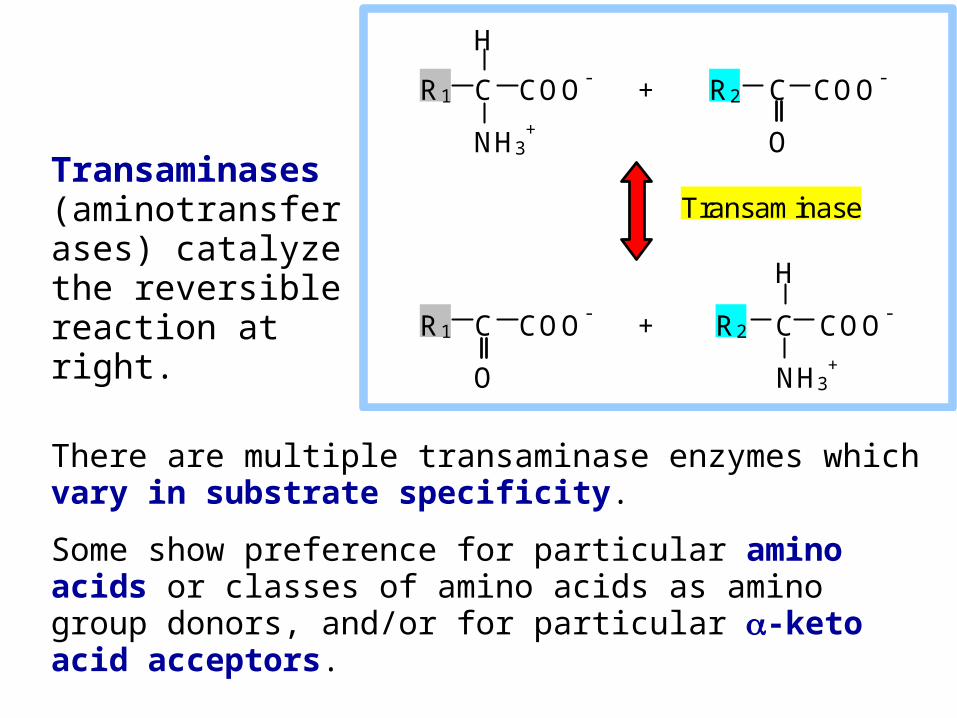
There are multiple transaminase enzymes which vary in substrate specificity.
Some show preference for particular amino acids or classes of amino acids as amino group donors, and/or for particular -keto acid acceptors.
H
R1 C COO- + R2 C COO-
NH3+ O
Transaminase
H
R1 C COO- + R2 C COO-
O NH3+
Transaminases (aminotransferases) catalyze the reversible reaction at right.
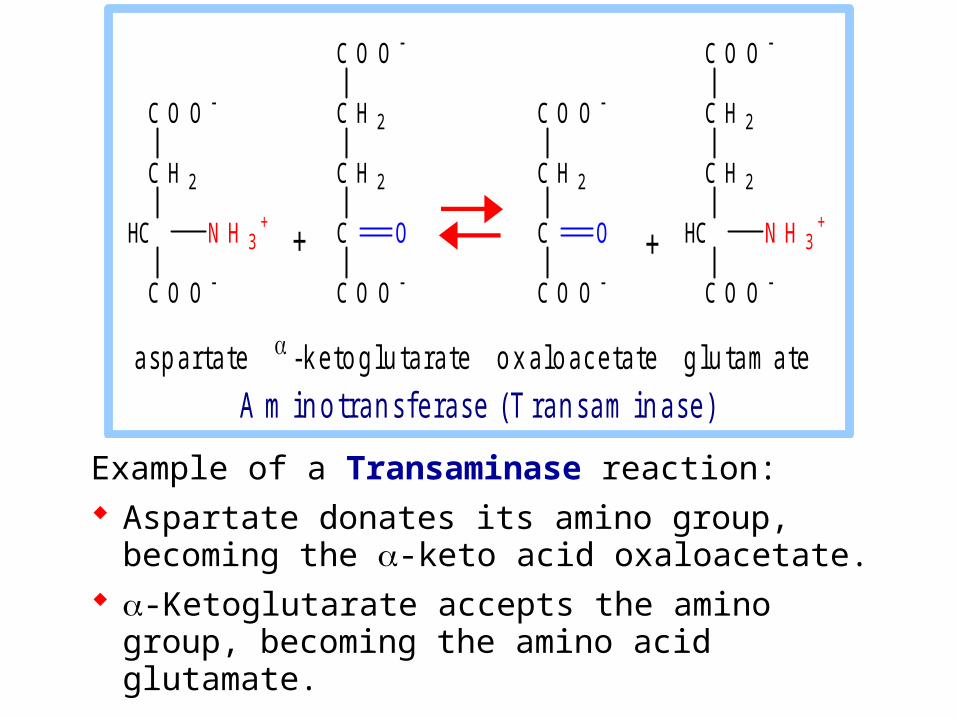
Example of a Transaminase reaction: Aspartate donates its amino group, becoming the
-keto acid oxaloacetate. -Ketoglutarate accepts the amino group,
becoming the amino acid glutamate.
a s p a r t a t e - k e t o g l u t a r a t e o x a l o a c e t a t e g l u t a m a t e
A m i n o t r a n s f e r a s e ( T r a n s a m i n a s e )
C O O
C H 2
C H 2
C
C O O
O
C O O
C H 2
HC
C O O
N H 3+
C O O
C H 2
C H 2
HC
C O O
N H 3+
C O O
C H 2
C
C O O
O + +
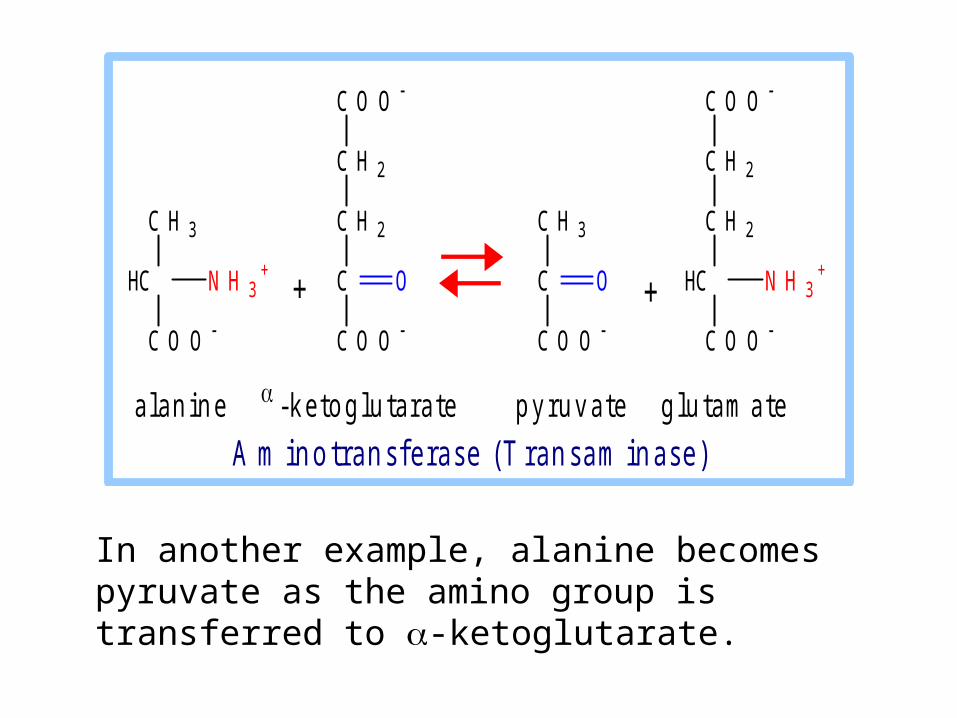
In another example, alanine becomes pyruvate as the amino group is transferred to -ketoglutarate.
a l a n i n e - k e t o g l u t a r a t e p y r u v a t e g l u t a m a t e
A m i n o t r a n s f e r a s e ( T r a n s a m i n a s e )
C O O
C H 2
C H 2
C
C O O
O
C H 3
HC
C O O
N H 3+
C O O
C H 2
C H 2
HC
C O O
N H 3+
C H 3
C
C O O
O + +

Transaminases equilibrate amino groups among available -keto acids.
This permits synthesis of non-essential amino acids, using amino groups from other amino acids & carbon skeletons synthesized in a cell.
Thus a balance of different amino acids is maintained, as proteins of varied amino acid contents are synthesized.
Although the amino N of one amino acid can be used to synthesize another amino acid, N must be obtained in the diet as amino acids (proteins).

Essential amino acids must be consumed in the diet.
Mammalian cells lack enzymes to synthesize their carbon skeletons (-keto acids). These include:
Isoleucine, leucine, & valine
Lysine
Threonine
Tryptophan
Phenylalanine (Tyr can be made from Phe.)
Methionine (Cys can be made from Met.)
Histidine (Essential for infants.)
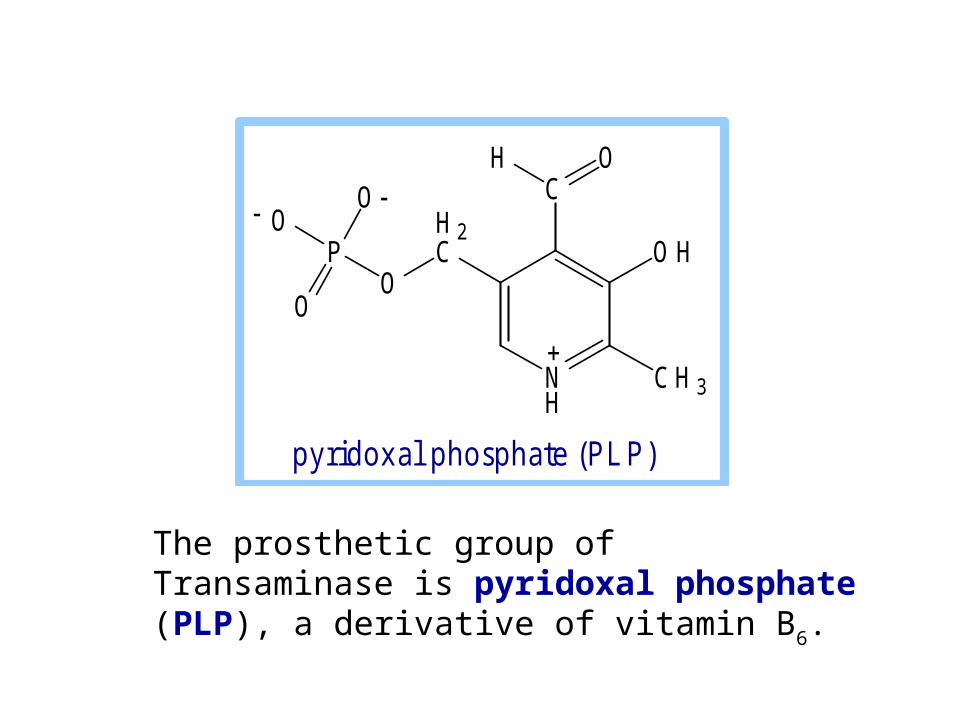
The prosthetic group of Transaminase is pyridoxal phosphate (PLP), a derivative of vitamin B6.
p y rid o x a l p h o sp h a te (P L P )
NH
CO
P
O O
O
O H
C H 3
CH O
H 2
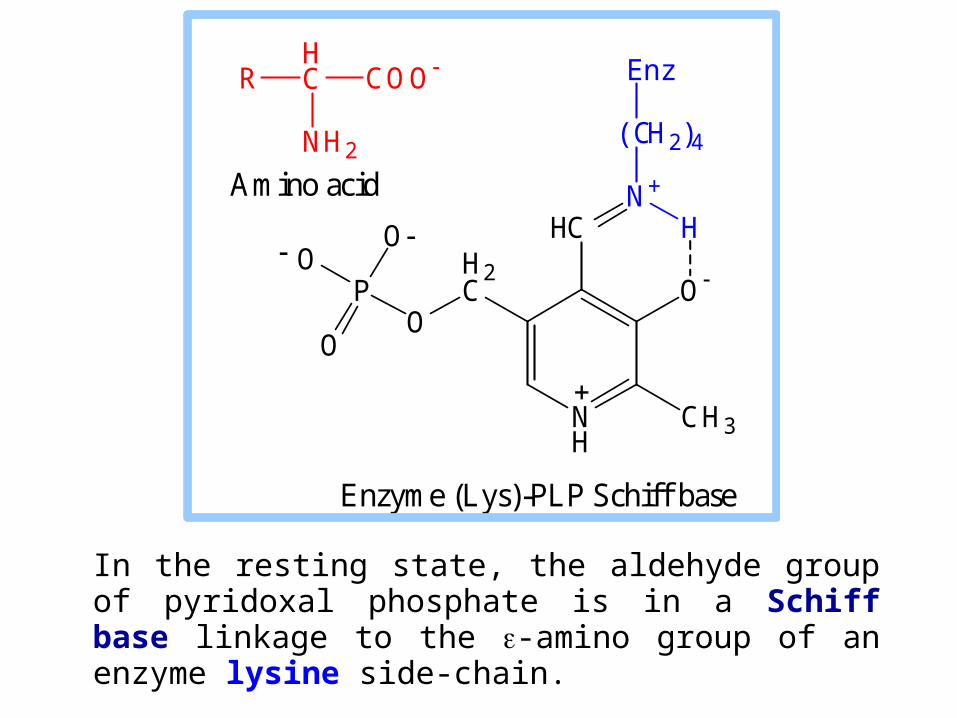
In the resting state, the aldehyde group of pyridoxal phosphate is in a Schiff base linkage to the -amino group of an enzyme lysine side-chain.
NH
CO
P
OO
O
O
CH3
HC
H2
N
(CH2)4
Enz
H
+
RHC COO
NH2
Enzyme (Lys)-PLP Schiff base
Amino acid
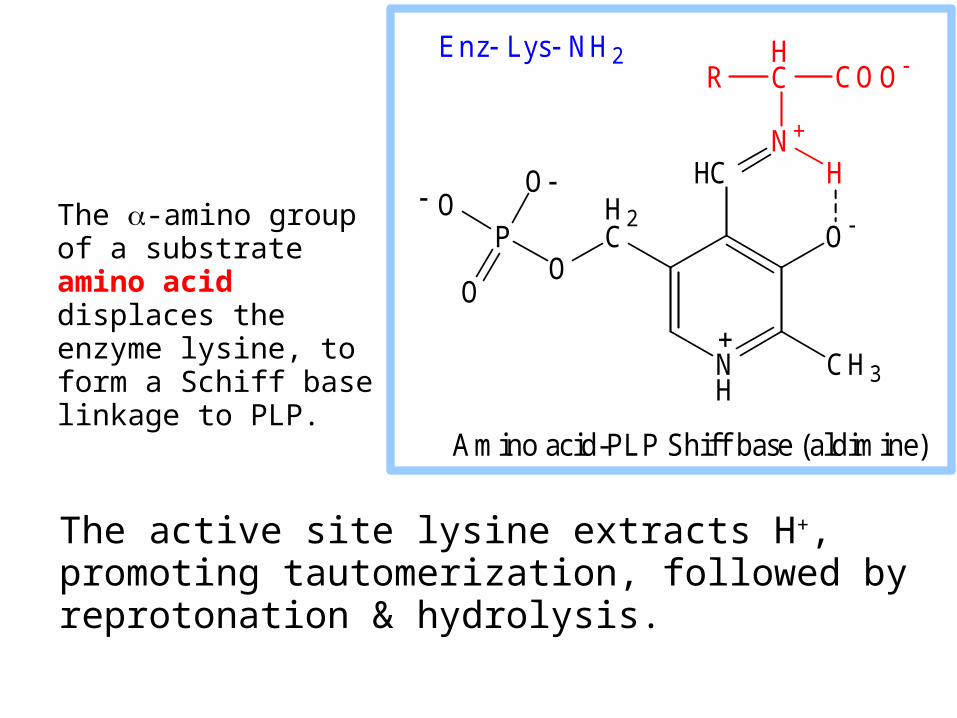
The active site lysine extracts H+, promoting tautomerization, followed by reprotonation & hydrolysis.
NH
CO
P
OO
O
O
CH3
HC
H2
N
HC
H
+
R COO Enz Lys NH2
Amino acid-PLP Shiff base (aldimine)
The -amino group of a substrate amino acid displaces the enzyme lysine, to form a Schiff base linkage to PLP.
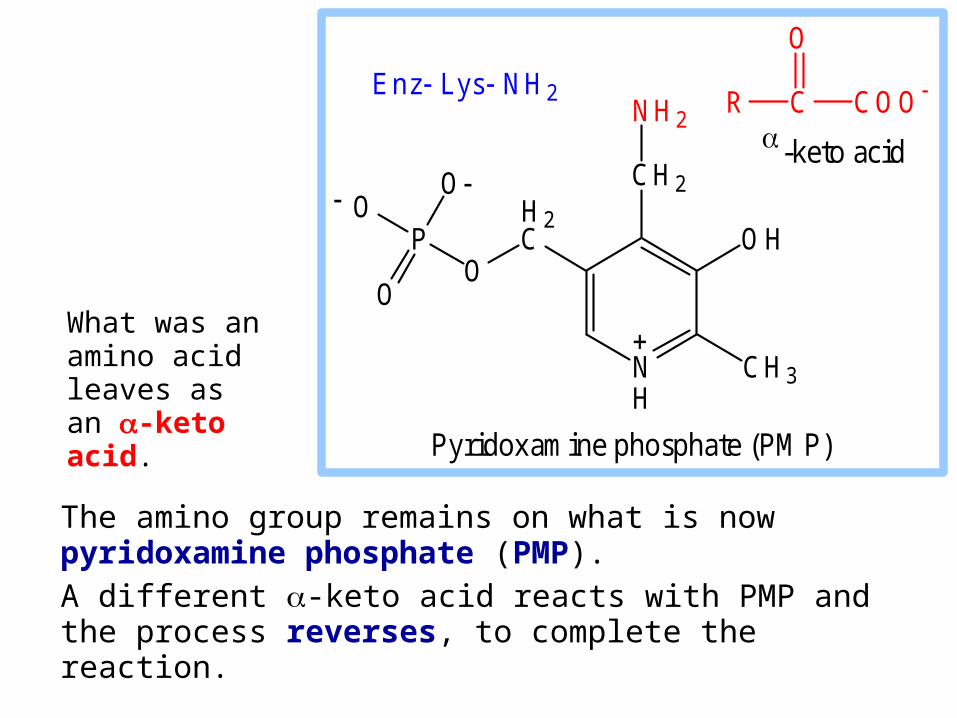
The amino group remains on what is now pyridoxamine phosphate (PMP).
A different -keto acid reacts with PMP and the process reverses, to complete the reaction.
NH
CO
P
O O
O
OH
CH3
CH2
NH2
H2
R C COO
O
Enz Lys NH2
Pyridoxamine phosphate (PM P)
-keto acid
What was an amino acid leaves as an -keto acid.
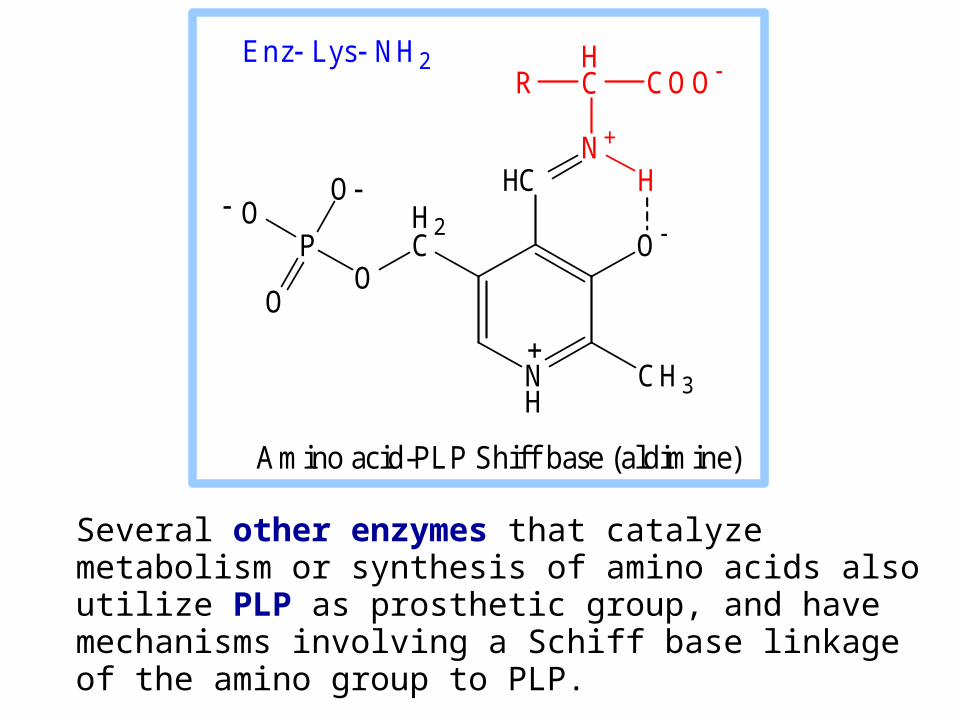
Several other enzymes that catalyze metabolism or synthesis of amino acids also utilize PLP as prosthetic group, and have mechanisms involving a Schiff base linkage of the amino group to PLP.
NH
CO
P
OO
O
O
CH3
HC
H2
N
HC
H
+
R COO Enz Lys NH2
Amino acid-PLP Shiff base (aldimine)

Chime Exercise
Two neighboring students or student groups should team up, each displaying one of the following:
Transaminase with PLP in Schiff base linkage to the active site lysine residue.
Transaminase in the PMP form, with glutarate, an analog of -ketoglutarate, at the active site.
Students should then show and explain the structure displayed by them to the neighboring student or student group.

In addition to equilibrating amino groups among available -keto acids, transaminases funnel amino groups from excess dietary amino acids to those amino acids (e.g., glutamate) that can be deaminated.
Carbon skeletons of deaminated amino acids can be catabolized for energy, or used to synthesize glucose or fatty acids for energy storage.
Only a few amino acids are deaminated directly.
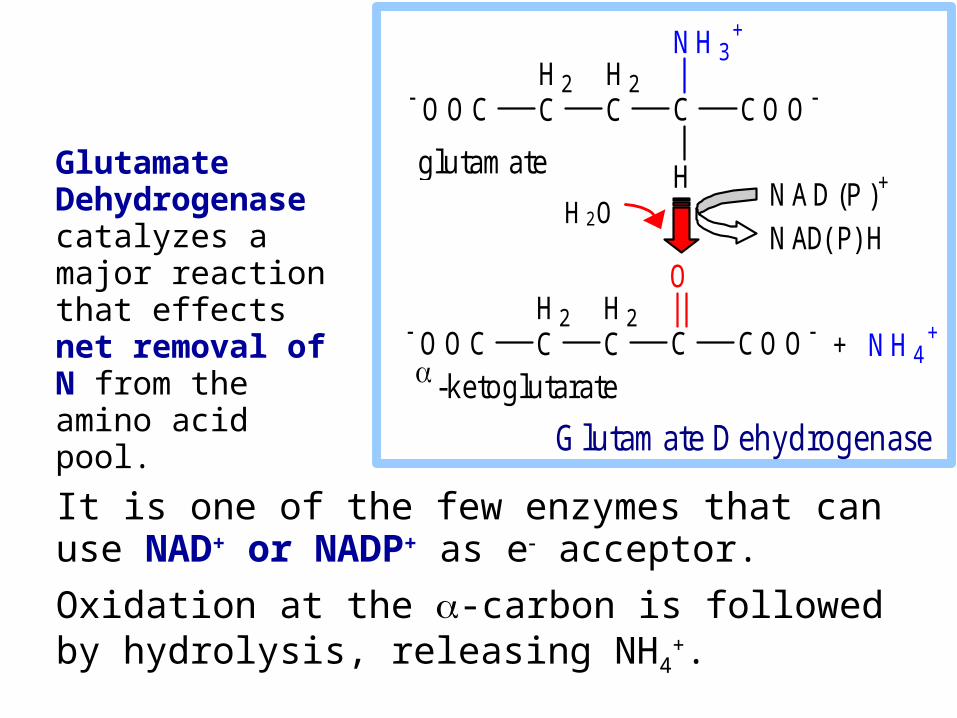
It is one of the few enzymes that can use NAD+ or NADP+ as e acceptor.
Oxidation at the -carbon is followed by hydrolysis, releasing NH4
+.
O O CH 2C
H 2C C C O O
O
+ N H 4+
N A D (P )+
N AD(P)H
O O CH 2C
H 2C C C O O
N H 3+
Hglu tam ate
-ke toglu tara te
G lu tam ate D ehydrogenase
H 2O Glutamate Dehydrogenase catalyzes a major reaction that effects net removal of N from the amino acid pool.
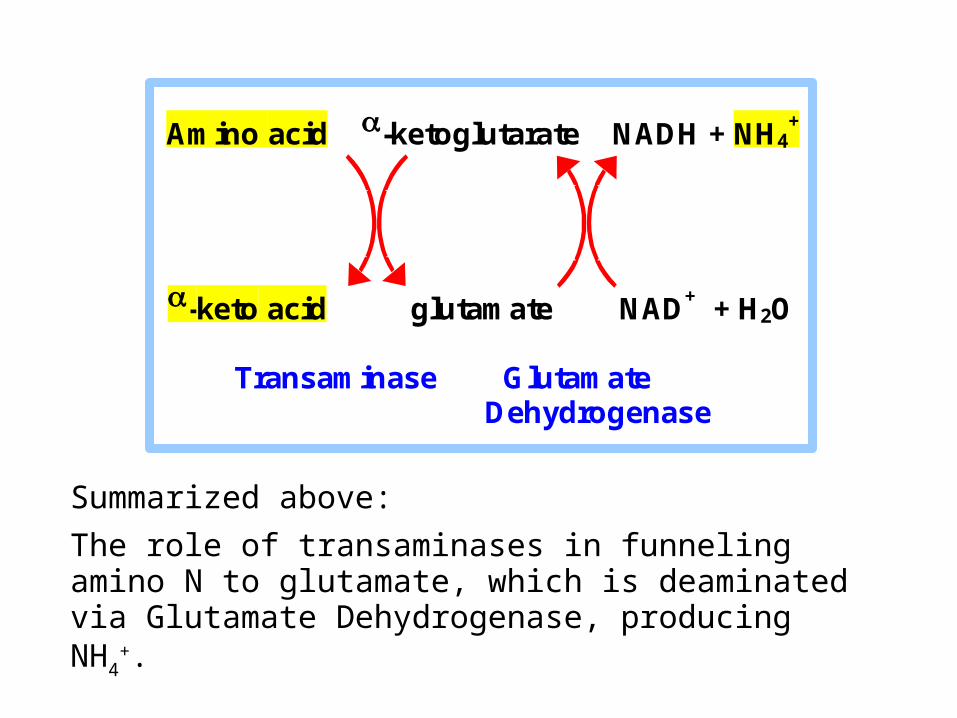
Summarized above:
The role of transaminases in funneling amino N to glutamate, which is deaminated via Glutamate Dehydrogenase, producing NH4
+.
Amino acid -ketoglutarate NADH + NH4+
-keto acid glutamate NAD+ + H2O
Transaminase Glutamate Dehydrogenase
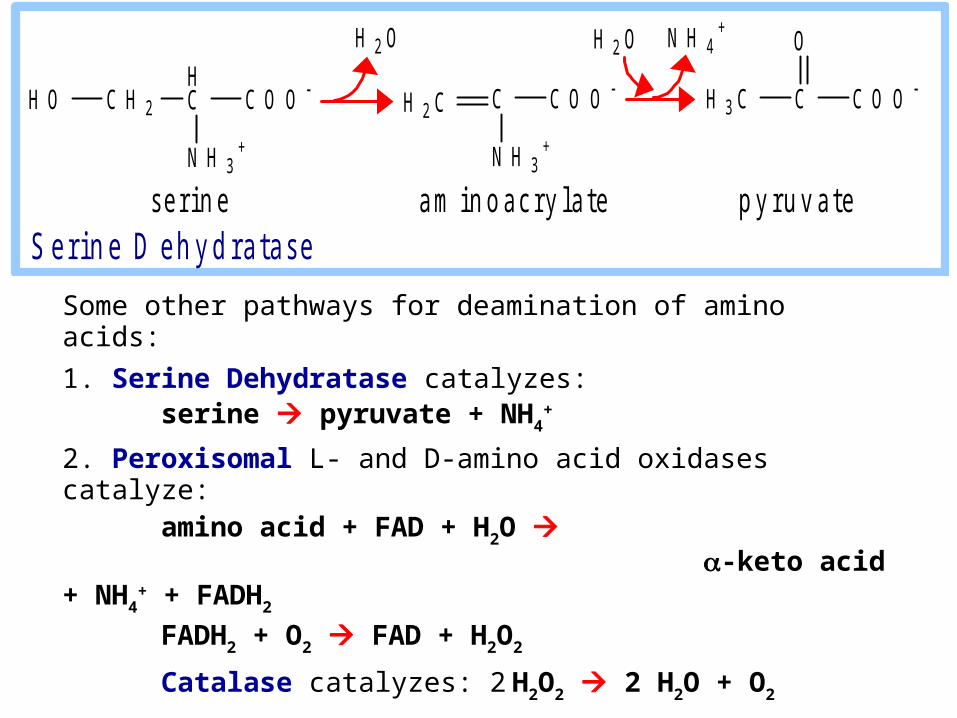
Some other pathways for deamination of amino acids:
1. Serine Dehydratase catalyzes: serine pyruvate + NH4
+
2. Peroxisomal L- and D-amino acid oxidases catalyze: amino acid + FAD + H2O -keto acid + NH4
+ + FADH2
FADH2 + O2 FAD + H2O2
Catalase catalyzes: 2 H2O2 2 H2O + O2
H O C H 2
HC C O O
N H 3+
C C O O
OH 2 O N H 4+
C C O O
N H 3+
H 2 C H 3 C
H 2 O
s e r i n e a m i n o a c r y l a t e p y r u v a t e S e r i n e D e h y d r a t a s e

Most terrestrial land animals convert excess nitrogen to urea, prior to excreting it.
Urea is less toxic than ammonia.
The Urea Cycle occurs mainly in liver.
The 2 nitrogen atoms of urea enter the Urea Cycle as NH3 (produced mainly via Glutamate Dehydrogenase) and as the amino N of aspartate.
The NH3 and HCO3 (carbonyl C) that will be part of urea
are incorporated first into carbamoyl phosphate.
H 2 N C
O
N H 2
u r e a
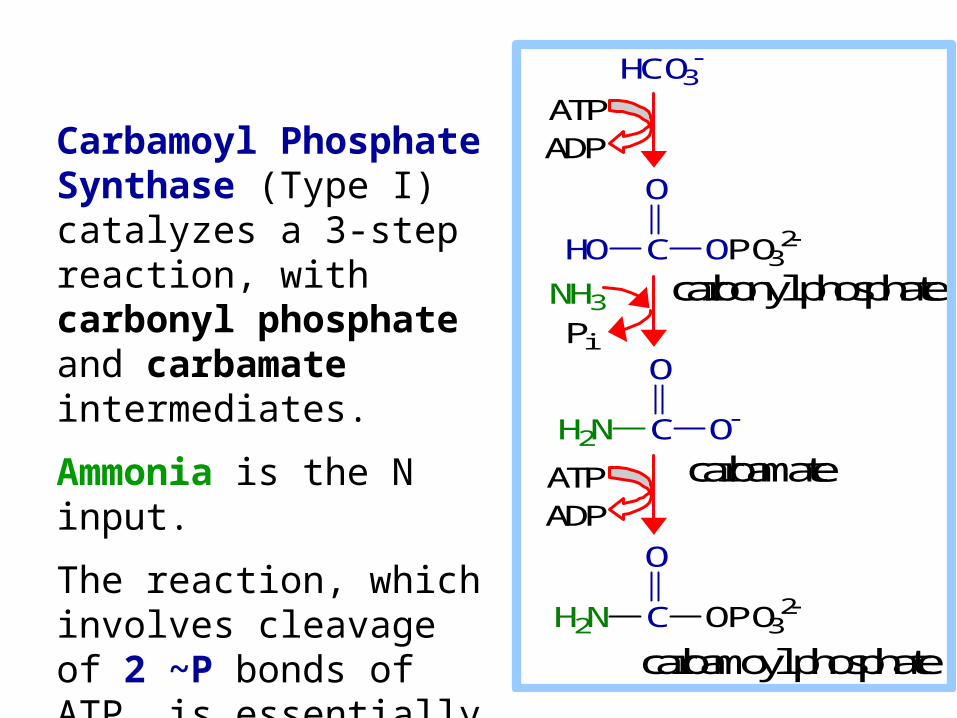
Carbamoyl Phosphate Synthase (Type I) catalyzes a 3-step reaction, with carbonyl phosphate and carbamate intermediates.
Ammonia is the N input.
The reaction, which involves cleavage of 2 ~P bonds of ATP, is essentially irreversible.
H2N C OPO32
O
H2N C O
O
HO C
O
OPO32
HCO3
ATP
NH3
ADP ATP
Pi
ADP
carbonyl phosphate
carbamate
carbamoyl phosphate
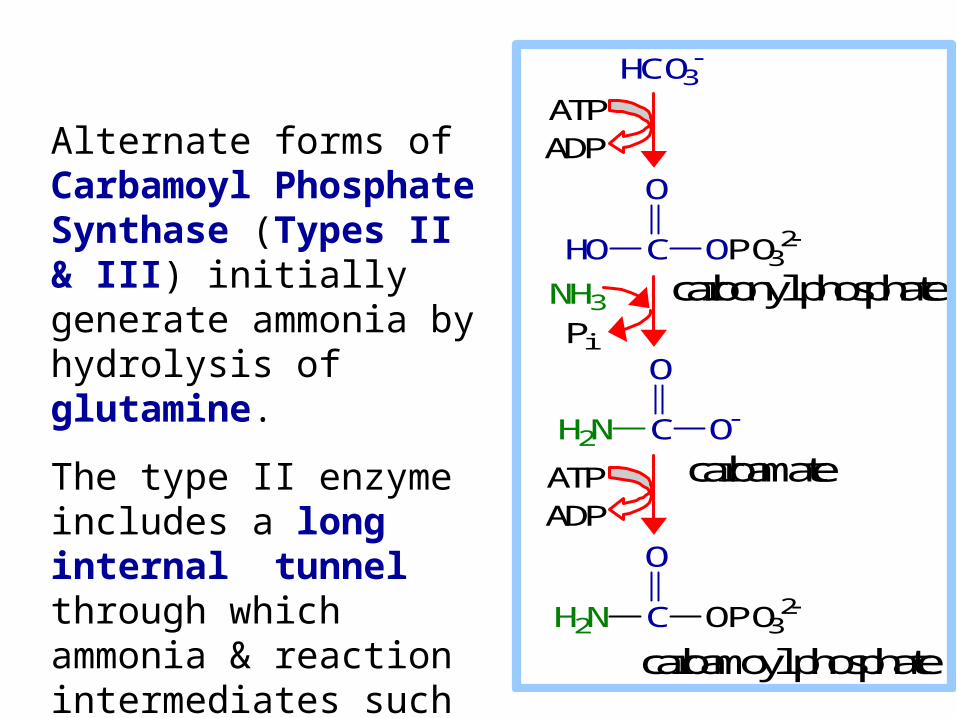
Alternate forms of Carbamoyl Phosphate Synthase (Types II & III) initially generate ammonia by hydrolysis of glutamine.
The type II enzyme includes a long internal tunnel through which ammonia & reaction intermediates such as carbamate pass from one active site to another.
H2N C OPO32
O
H2N C O
O
HO C
O
OPO32
HCO3
ATP
NH3
ADP ATP
Pi
ADP
carbonyl phosphate
carbamate
carbamoyl phosphate
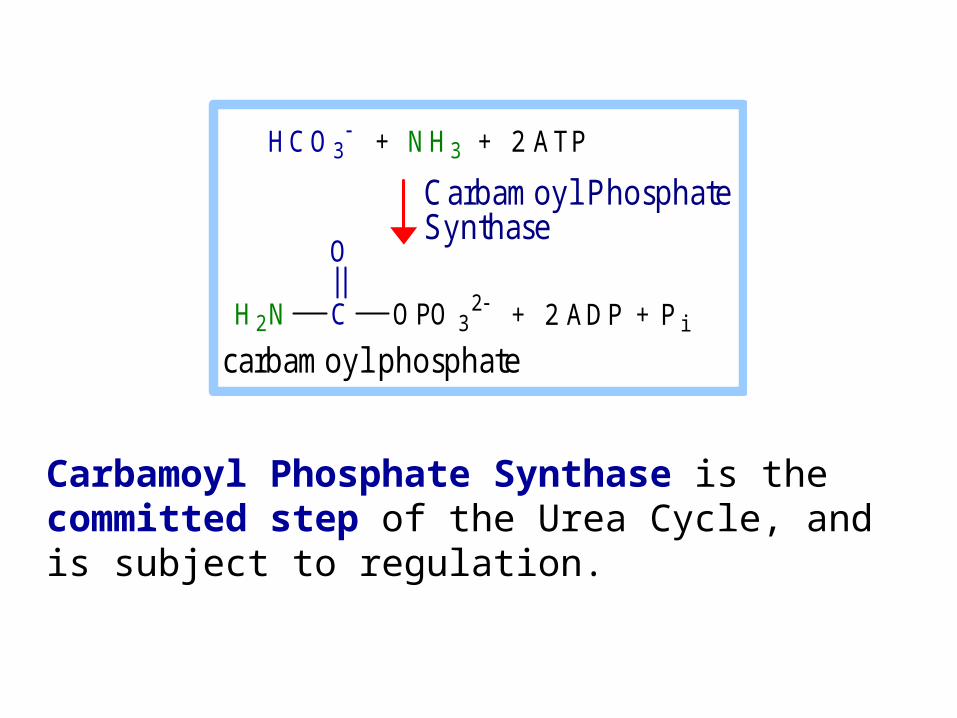
Carbamoyl Phosphate Synthase is the committed step of the Urea Cycle, and is subject to regulation.
H 2N C O PO 32
O
HC O 3 + N H 3 + 2 A TP
+ 2 AD P + P i
C arbam oyl Phosphate Synthase
carbam oyl phosphate
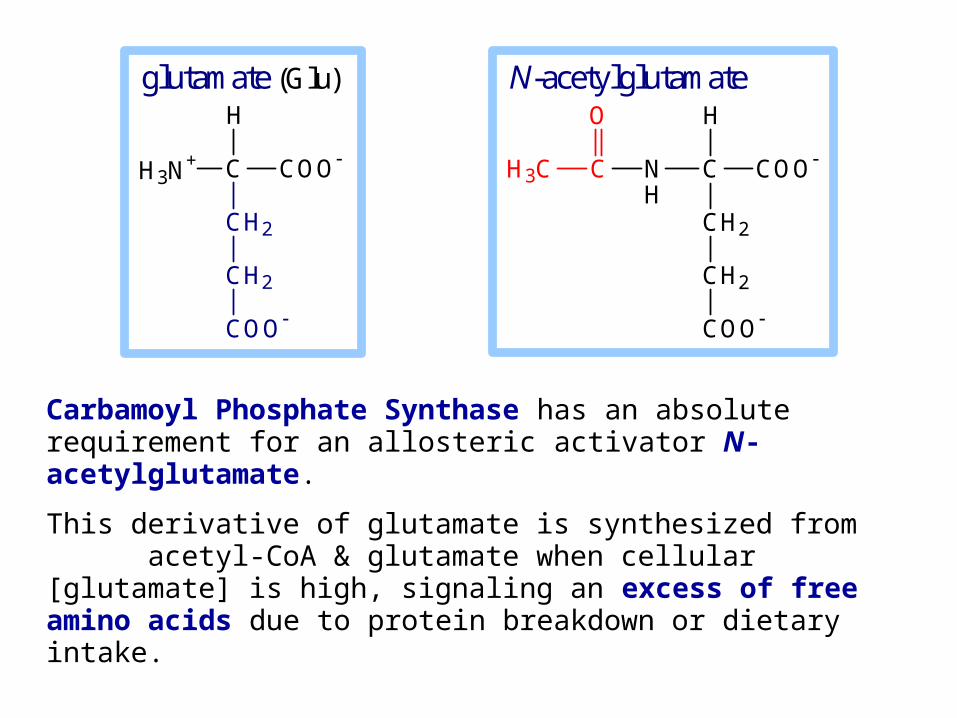
Carbamoyl Phosphate Synthase has an absolute requirement for an allosteric activator N-acetylglutamate.
This derivative of glutamate is synthesized from acetyl-CoA & glutamate when cellular [glutamate] is high, signaling an excess of free amino acids due to protein breakdown or dietary intake.
H3N+ C COO
CH2
CH2
COO
H
glutamate (Glu)
NH
C COO
CH2
CH2
COO
H
CH3C
O
N-acetylglutamate
H2N C OPO32
O
CH2
CH2
CH2
HC
COO
NH3+
NH3+
CH2
CH2
CH2
HC
COO
NH3+
NH
CO NH2
COO
CH2
HC
COO
NH2
CH2
CH2
CH2
HC
COO
NH3+
NH
C NH2+
COO
CH2
HC
COO
HN
AMP + PPi
ATP
CH2
CH2
CH2
HC
COO
NH3+
NH
C
NH2+H2N
COO
HC
CH
COO
C NH2H2N
O H2O
Pi
ornithine
urea
citrulline
aspartate
arginino- succinate
fumarate
arginine
carbamoyl phosphate
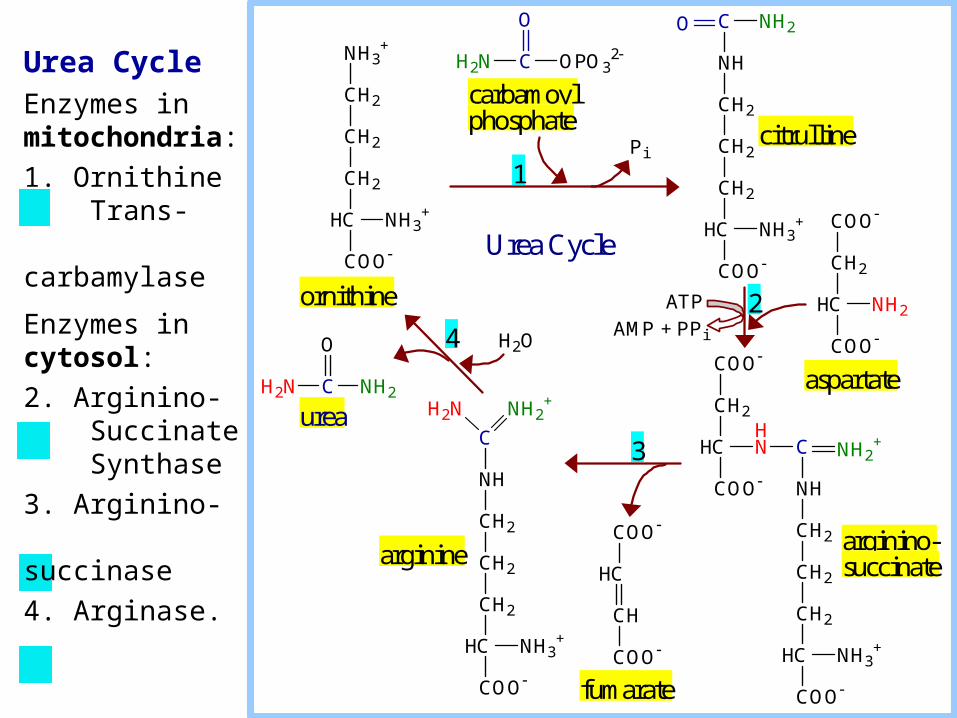
Urea Cycle
1
2
3
4
Urea CycleEnzymes in mitochondria:
1. Ornithine Trans- carbamylase
Enzymes in cytosol:
2. Arginino- Succinate Synthase
3. Arginino- succinase
4. Arginase.
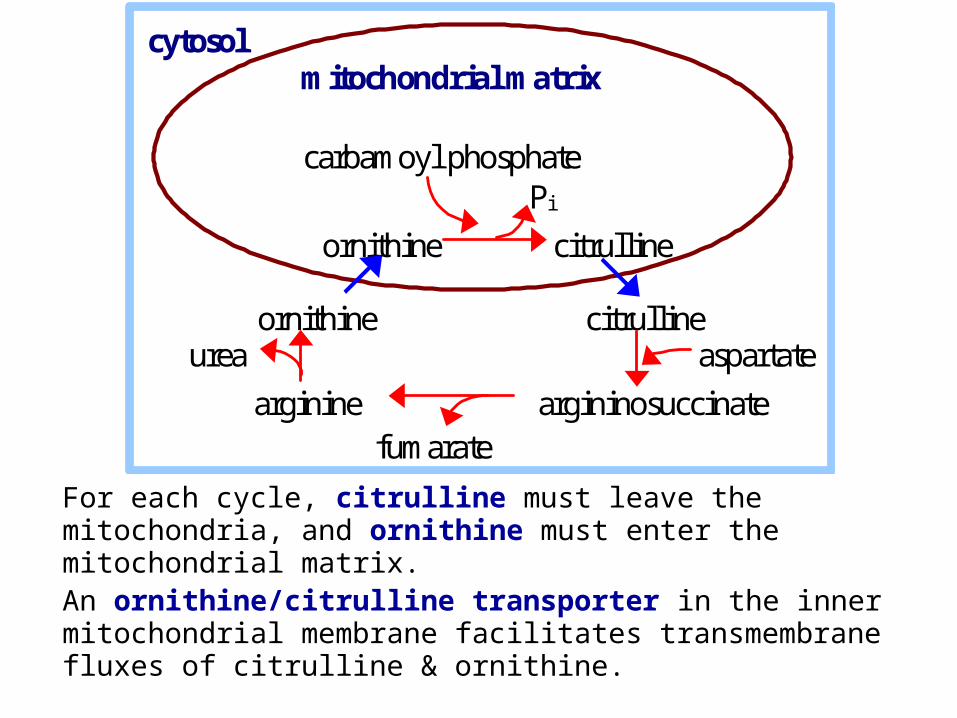
For each cycle, citrulline must leave the mitochondria, and ornithine must enter the mitochondrial matrix. An ornithine/citrulline transporter in the inner mitochondrial membrane facilitates transmembrane fluxes of citrulline & ornithine.
cytosol mitochondrial matrix
carbamoyl phosphate Pi
ornithine citrulline
ornithine citrulline urea aspartate
arginine argininosuccinate fumarate
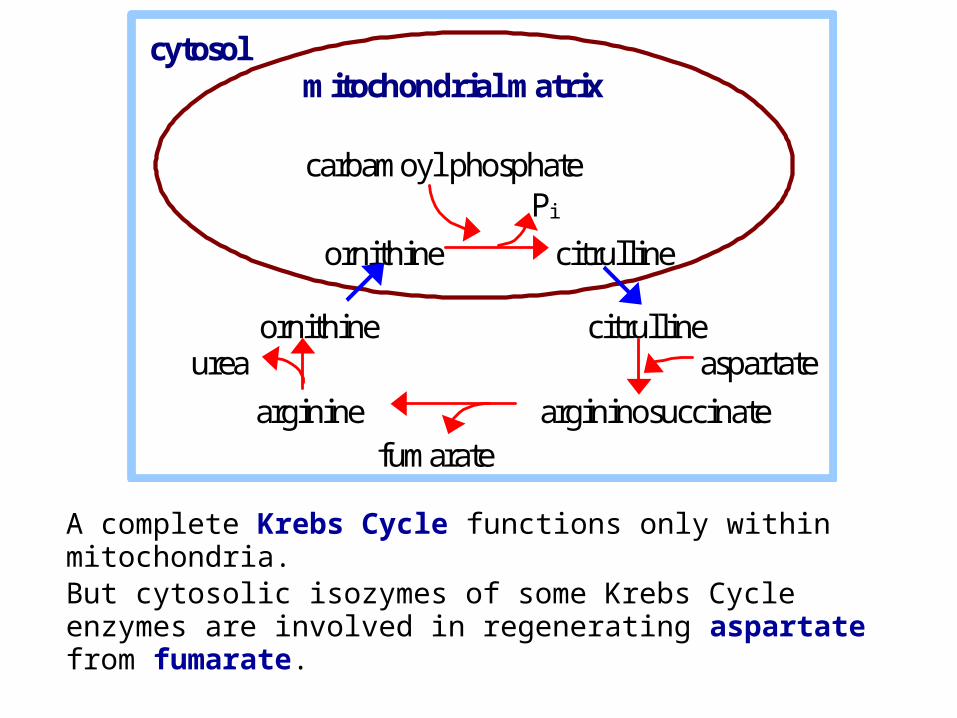
A complete Krebs Cycle functions only within mitochondria. But cytosolic isozymes of some Krebs Cycle enzymes are involved in regenerating aspartate from fumarate.
cytosol mitochondrial matrix
carbamoyl phosphate Pi
ornithine citrulline
ornithine citrulline urea aspartate
arginine argininosuccinate fumarate

Fumarate is converted to oxaloacetate via Krebs Cycle enzymes Fumarase & Malate Dehydrogenase.
Oxaloacetate is converted to aspartate via transamination (e.g., from glutamate).
Aspartate then reenters Urea Cycle, carrying an amino group derived from another amino acid.
a s p a r t a t e - k e t o g l u t a r a t e o x a l o a c e t a t e g l u t a m a t e
A m i n o t r a n s f e r a s e ( T r a n s a m i n a s e )
C O O
C H 2
C H 2
C
C O O
O
C O O
C H 2
HC
C O O
N H 3+
C O O
C H 2
C H 2
HC
C O O
N H 3+
C O O
C H 2
C
C O O
O + +

Hereditary deficiency of any of the Urea Cycle enzymes leads to hyperammonemia - elevated [ammonia] in blood.
Total lack of any Urea Cycle enzyme is lethal.
Elevated ammonia is toxic, especially to the brain.
If not treated immediately after birth, severe mental retardation results.

Postulated mechanisms for toxicity of high [ammonia]:
1. High [NH3] would drive Glutamine Synthase:
glutamate + ATP + NH3 glutamine + ADP + Pi
This would deplete glutamate – a neurotransmitter & precursor for synthesis of the neurotransmitter GABA.
2. Depletion of glutamate & high ammonia level would drive Glutamate Dehydrogenase reaction to reverse: glutamate + NAD(P)+ -ketoglutarate + NAD(P)H + NH4
+
The resulting depletion of -ketoglutarate, an essential Krebs Cycle intermediate, could impair energy metabolism in the brain.

Treatment of deficiency of Urea Cycle enzymes (depends on which enzyme is deficient):
limiting protein intake to the amount barely adequate to supply amino acids for growth, while adding to the diet the -keto acid analogs of essential amino acids.
Liver transplantation has also been used, since liver is the organ that carries out Urea Cycle.
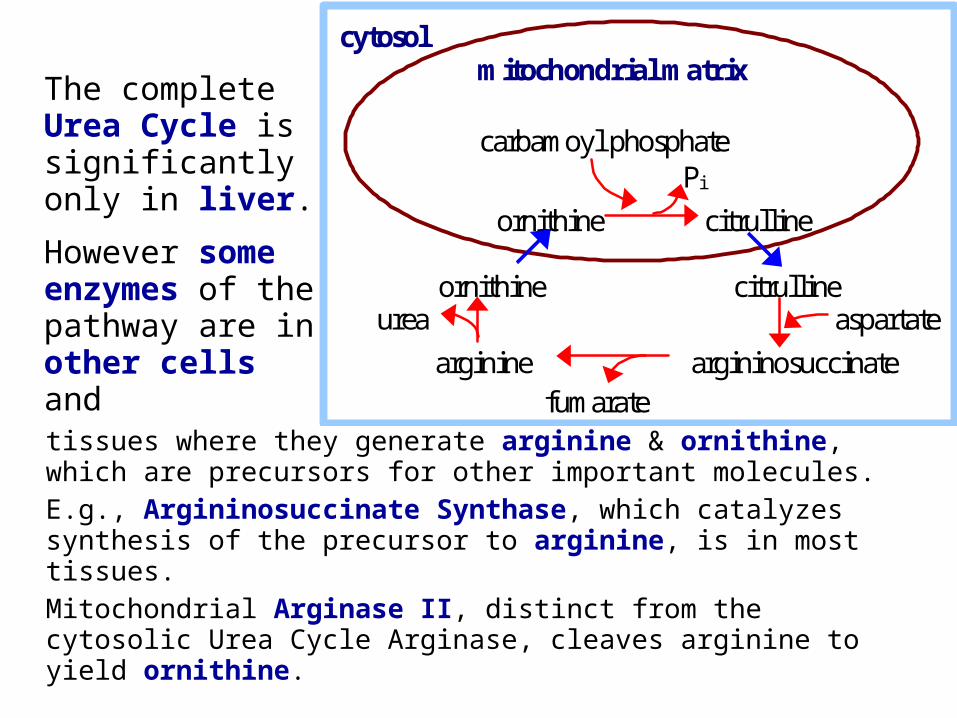
tissues where they generate arginine & ornithine, which are precursors for other important molecules.
E.g., Argininosuccinate Synthase, which catalyzes synthesis of the precursor to arginine, is in most tissues.
Mitochondrial Arginase II, distinct from the cytosolic Urea Cycle Arginase, cleaves arginine to yield ornithine.
cytosol mitochondrial matrix
carbamoyl phosphate Pi
ornithine citrulline
ornithine citrulline urea aspartate
arginine argininosuccinate fumarate
The complete Urea Cycle is significantly only in liver.
However some enzymes of the pathway are in other cells and
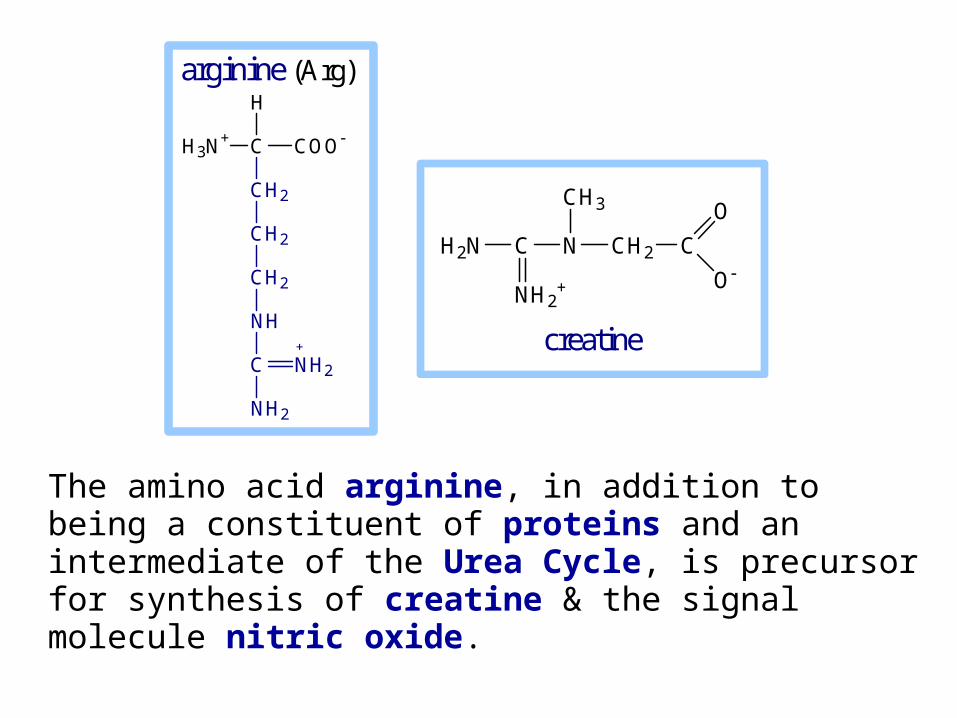
The amino acid arginine, in addition to being a constituent of proteins and an intermediate of the Urea Cycle, is precursor for synthesis of creatine & the signal molecule nitric oxide.
H3N+ C COO
CH2
CH2
CH2
NH
C
NH2
NH2
H
arginine (Arg)
H2N C N
NH2+
CH2
CH3
C
O
O
creatine

Synthesis of the radical species nitric oxide (·NO) from arginine is catalyzed Nitric Oxide Synthase, a distant relative of cytochrome P450.
Different isoforms of Nitric Oxide Synthase (e.g., eNOS expressed in endothelial cells and nNOS in neuronal cells) are subject to differing regulation.
+H3N CH COO
CH2
CH2
CH2
NH
C
NH2
NH2+
NADPH NADP+
O2 H2O O2 H2O
+H3N CH COO
CH2
CH2
CH2
NH
C
NH2
N OH
+H3N CH COO
CH2
CH2
CH2
NH
C
NH2
O
1/2 NADPH 1/2 NADP+
+ NO
Nitric Oxide Synthase
arginine hydroxyarginine citrulline

·NO is a short-lived signal molecule with diverse roles in different cell types, including regulation of smooth muscle contraction, gene transcription, metabolism, and neurotransmission.
Many of the regulatory effects of ·NO arise from its activation of a soluble cytosolic Guanylate Cyclase enzyme that catalyzes synthesis of cyclic-GMP (analogous in structure to cyclic-AMP).
Cytotoxic effects of ·NO observed under some conditions are attributed to its non-enzymatic reaction with superoxide (O2·) to form the strong oxidant peroxynitrite (ONOO).

Polyamines include putrescine, spermidine, spermine.
Ornithine is a major precursor for synthesis of polyamines.
Conversion of ornithine to putrescine is catalyzed by Ornithine Decarboxylase.
+H3N CH2 CH2 CH2 CH2 NH3
+
+H3N CH2 CH2 CH2 NH CH2 CH2 CH2 CH2 NH3+
putrescine
spermidine
H3N+ C COO
CH2
CH2
CH2
NH3
H
ornithine

The cationic polyamines have diverse roles in cell growth & proliferation.
Disruption of polyamine synthesis or metabolism leads to disease in animals & humans.
+H3N CH2 CH2 CH2 CH2 NH3
+
+H3N CH2 CH2 CH2 NH CH2 CH2 CH2 CH2 NH3+
putrescine
spermidine

However, Ca++-activated Peptidylarginine Deiminases convert arginine residues within proteins to citrulline as a post-translational modification.
H3N+ C COO
CH2
CH2
CH2
NH
C
NH2
NH2
H
H3N+ C COO
CH2
CH2
CH2
NH
C NH2
H
O
arginine citrulline
There is no tRNA for citrulline & this amino acid is not incorporated translationally into proteins.

is essential to terminal differentiation of skin cells.
Excessive protein citrullination, with production of antibodies against citrullinated proteins, is found to be a factor in the autoimmune diseases such as rheumatoid arthritis and multiple sclerosis.
H3N+ C COO
CH2
CH2
CH2
NH
C
NH2
NH2
H
H3N+ C COO
CH2
CH2
CH2
NH
C NH2
H
O
arginine citrulline
Substitution of citrulline, which lacks arginine's positive charge, may alter structure & properties such as binding affinities of a protein.
E.g., citrullination of certain proteins, including keratin intermediate filament proteins,





























