Embed Size (px)
Citation preview
AMINO ÁCIDOS
PÉPTIDOS Y
PROTEÍNAS
C
COO
H3N H
R
+
- 2 -
AMINO ÁCIDOS, PÉPTIDOS Y PROTEÍNAS
1.-DEFINICIONES
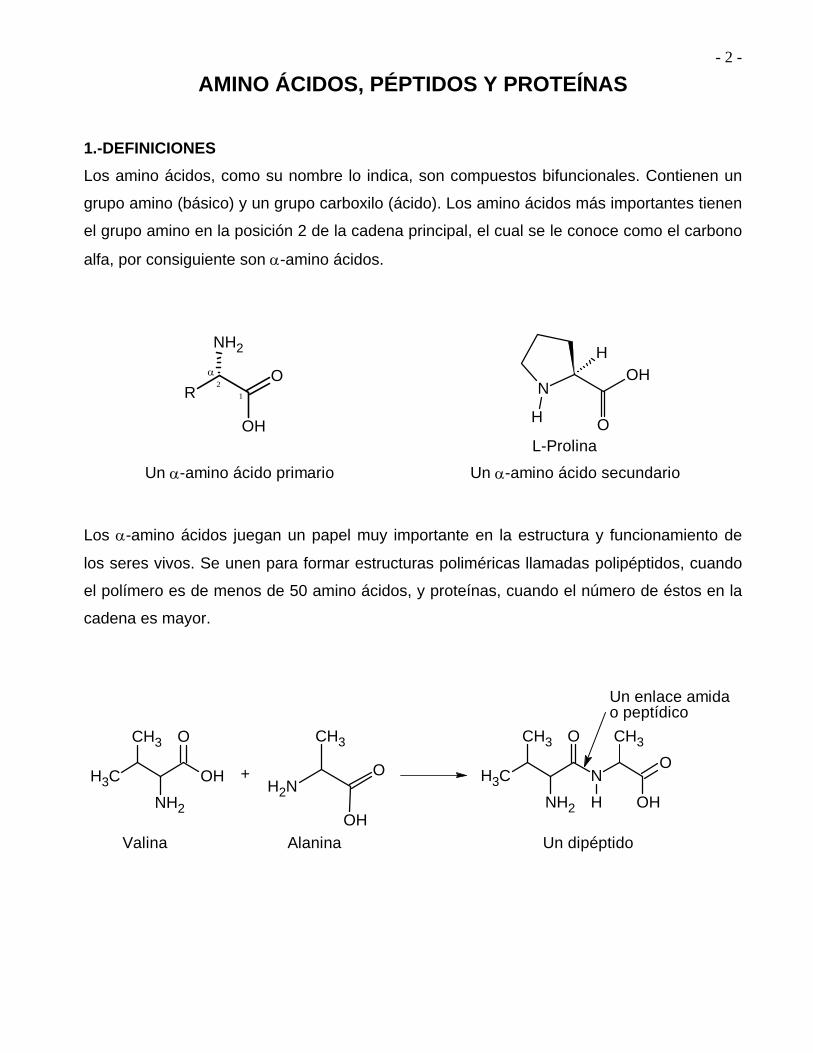
Los amino ácidos, como su nombre lo indica, son compuestos bifuncionales. Contienen un
grupo amino (básico) y un grupo carboxilo (ácido). Los amino ácidos más importantes tienen
el grupo amino en la posición 2 de la cadena principal, el cual se le conoce como el carbono
alfa, por consiguiente son -amino ácidos.
O
NOH
H
H
O
OH
NH2
R
Un -amino ácido primario
L-Prolina
Un -amino ácido secundario
1
2
Los -amino ácidos juegan un papel muy importante en la estructura y funcionamiento de
los seres vivos. Se unen para formar estructuras poliméricas llamadas polipéptidos, cuando
el polímero es de menos de 50 amino ácidos, y proteínas, cuando el número de éstos en la
cadena es mayor.
ONH2
CH3
OH
O
NH2
N
H
O
OH
CH3
CH3
CH3
NH2
OH
OCH3
CH3 +
AlaninaValina Un dipéptido
Un enlace amidao peptídico
- 3 -
NH
NH
NH
NH
NH
R1
R2
R3
R4
R5
O
O
O
O
O
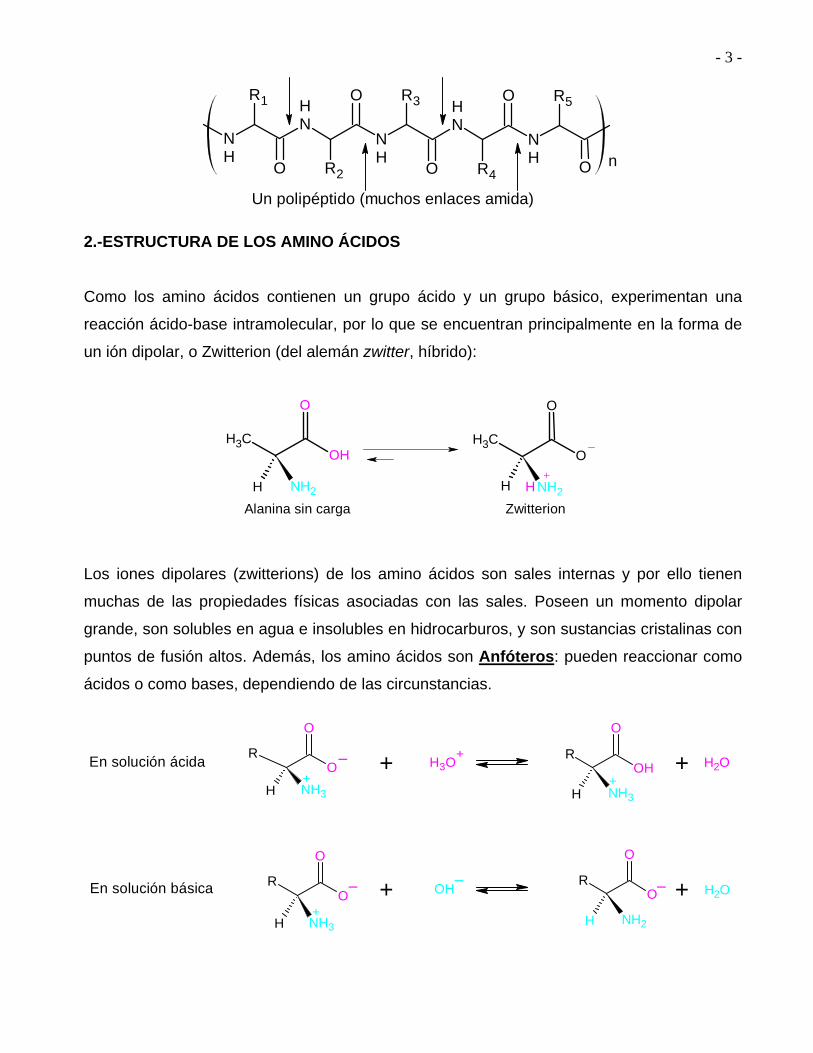
Un polipéptido (muchos enlaces amida)
n
2.-ESTRUCTURA DE LOS AMINO ÁCIDOS
Como los amino ácidos contienen un grupo ácido y un grupo básico, experimentan una
reacción ácido-base intramolecular, por lo que se encuentran principalmente en la forma de
un ión dipolar, o Zwitterion (del alemán zwitter, híbrido):
O
OH
H
CH3
NH2
O
O
H
CH3
NH2
Alanina sin carga Zwitterion
-
H+
Los iones dipolares (zwitterions) de los amino ácidos son sales internas y por ello tienen
muchas de las propiedades físicas asociadas con las sales. Poseen un momento dipolar
grande, son solubles en agua e insolubles en hidrocarburos, y son sustancias cristalinas con
puntos de fusión altos. Además, los amino ácidos son Anfóteros: pueden reaccionar como
ácidos o como bases, dependiendo de las circunstancias.
R
H
O
O
NH3
+ H3OR
H
OH
O
NH3
OH2+En solución ácida
En solución básica R
H
O
O
NH3
+ OHR
H
O
O
NH2
+ OH2-
+
+
- -
-
++

- 4 -
3.-ACIDEZ, BASICIDAD Y pKa 3.1.-Ácidos y Bases de Brøsnted-Lowry Ácido: cede protones y forma la base conjugada Base: recibe protones y forma el ácido conjugado
H2O H3OA+
-H A+ +-
Ácido Base Ácido Baseconjugado conjugada
3.2.-Constante de acidez y pKa
Cuando un ácido, como el ácido acético, se disuelve en agua se lleva a cabo una reacción
ácido-base hasta llegar a un equilibrio que se representa, de acuerdo a la ley de acción de
masas, como:
H2O H3O+
-H + + -
Ácido Base Ácido Baseconjugado conjugada
CH3COO CH3COOKeq.
En donde Keq. es la constante de equilibrio, la cual se determina precisamente cuando se
establece el equilibrio y está dada por la siguiente ecuación:
Keq. = [CH3COO -] [H3O+]
[CH3COO H] [H2O]
[CH3COO-] = es la concentración molar de CH3COO-] en el equilibrio.
[H3O+] = es la concentración molar de [H3O
+] en el equilibrio.
[CH3COOH] = es la concentración molar de CH3COOH en el equilibrio.
Para fines prácticos, la concentración molar del agua se considera constante e igual a 55.6
M a 25 C.
Si se multiplica la Keq. por otra constante, la concentración del agua en el equilibrio, se
obtiene una nueva constante (Ka) llamada constante de acidez, ionización o disociación en
agua:

- 5 -
Keq. [CH3COO -] [H3O
+]
[CH3COO H] [H2O] = Ka = = 1.75 x 10-5
Para el caso de una base protonada:
RNH2H H2O RNH2+ +
+ Keq.
Keq. =
H3O+
Por consiguiente Keq. = Ka = = 1 x 10-10
[H3O]
[H2O]
[RNH2]
[RNH3]
+
+
[H2O][H3O] [RNH2]
[RNH3]
+
+
Con el propósito de manejar números enteros y comparar la acidez de las sustancias más
fácilmente, se toma el logaritmo negativo de la Ka y se obtiene la constante pKa:
-log Ka = pKa Para el ácido acético: pKa = -log (1.75 x 10-5) = 5-0.2430 = 4.7570 ~5
Para el caso de las bases (aminas con hibridación sp3) pKa = -log (1 x 10-10) = 10
Según su valor de pKa los ácidos se consideran muy fuertes, moderadamente fuertes,
débiles y muy débiles, como se aprecia en la Tabla 1.
Tabla 1. Constantes de acidez y valores de pKa de algunos ácidos Fórmula Ka pKa Fuertes: HClO4 HCl H2SO4
~1010 ~107
~105
~-10 ~-7 ~-5
Moderadamente fuertes: H3PO4 ClCH2COOH
7.52 x 10-3
2.12 2.9
Débiles: CH3COOH CH3CH2COOH
1.75 x 10-5 1.34 x 10-5
4.75 4.87
Muy débiles: HCN
4.93 x 10-10
9.31

- 6 -
H2O 2 x 10-16 15.7 Con estos datos se aprecia que: A menor valor de pKa más fuerte es el ácido. A mayor valor de pKa más débil es el ácido.
3.3.-Ecuación de Henderson–Hasselbalch
La ecuación de Henderson–Hasselbalch es de mucha utilidad, ya que permite conocer la
forma protonada o no protonada de un ácido o una base débil a un pH determinado. La
ecuación se deriva de la forma siguiente:
Para un ácido débil
[CH3COO -] [H3O+]
[CH3COOH] -logKa = -log o bien
[CH3COO -]
[CH3COOH] pKa = -log [H3O
+] -log
[CH3COO -]
[CH3COOH] pKa = -logpH-log [H3O
+] = pH Por consiguiente
pKa = -logpH[Ácido no disociado]
[Ácido disociado]
Ecuación de Henderson-Hasselbach
Para una base débil protonada
pKa = -logpH pKa = -logpH[Base ionizada]
[Base no ionizada][RNH2]
[RNH3]+
La ecuación de Henderson–Hasselbalch se puede entonces escribir de manera general,
para un ácido o una base, de la siguiente forma:

- 7 -
pKa = -logpH[Protonado]
[No protonado]
Si el ácido y su sal se encuentran en igual proporción, entonces:
pKa = -logpH[Protonado]
[No protonado]
50%
50%pKa = -logpH 1 pKa = pH-0 pKa = pH
Por lo que se puede definir al pKa como el pH en donde el ácido y su sal o la base y su sal
se encuentran en igual proporción, también se podrá apreciar con esto que:
Un ácido en solución acuosa a pH por abajo de su pKa se encuentra esencialmente
protonado (no disociado).
Un ácido en solución acuosa a pH por arriba de su pKa se encuentra esencialmente no
protonado (disociado).
Una base en solución acuosa a pH por abajo de su pKa se encuentra esencialmente
protonada (disociada).
Una base en solución acuosa a pH por arriba de su pKa se encuentra esencialmente no
protonada (no disociada).
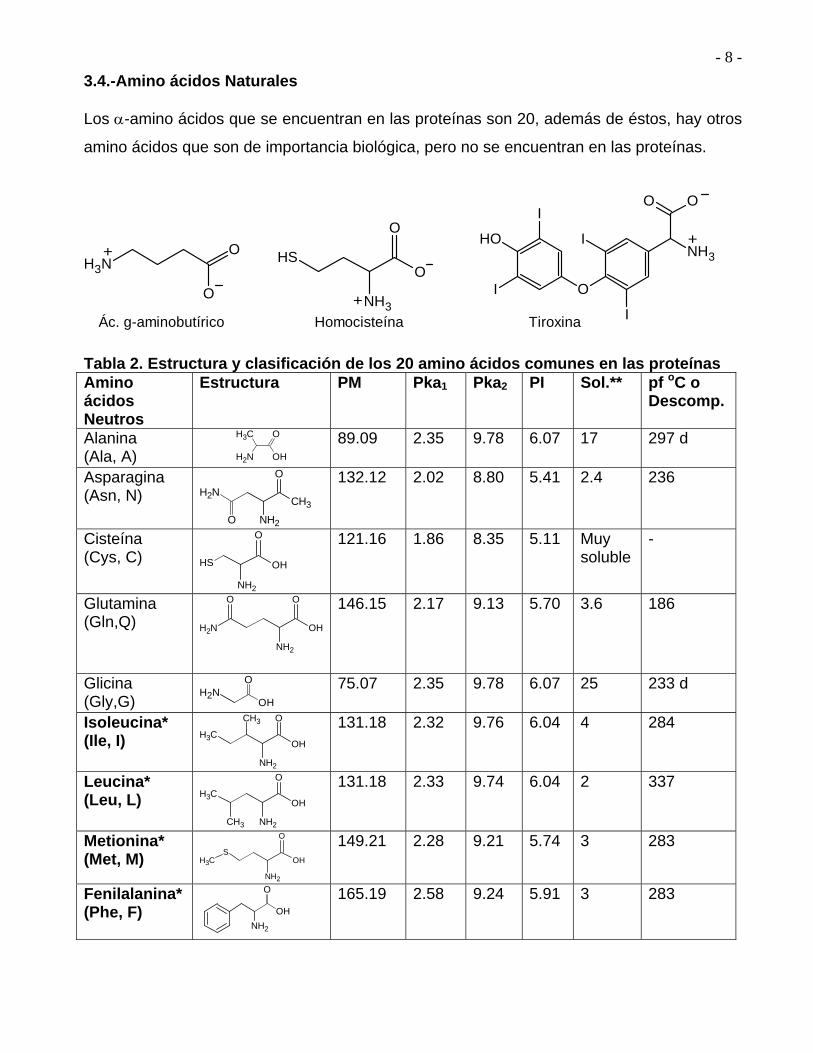
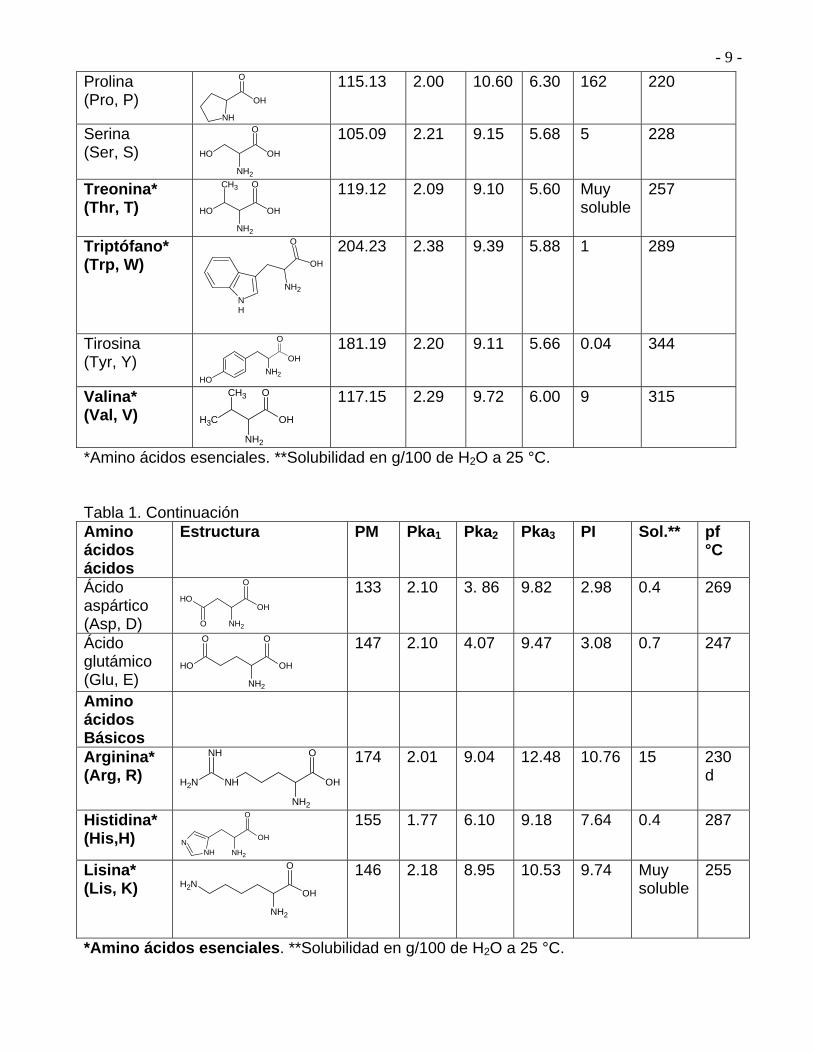
En la Tabla 2 se representa la estructura de los 20 amino ácidos que se encuentran en las
proteínas. Todos son -amino ácidos, 19 de ellos son aminas primarias, RNH2, y sólo
difieren en la naturaleza del sustituyente R, llamado Cadena lateral, unido al carbono .
CR
H3N
O
O
H
Cadena lateral
Estructura general de un -aminoácido
-+
El carbono es el carbono alcual se une un grupo funcional
CR
NH2
OH
O
H
- 8 -
3.4.-Amino ácidos Naturales Los -amino ácidos que se encuentran en las proteínas son 20, además de éstos, hay otros
amino ácidos que son de importancia biológica, pero no se encuentran en las proteínas.
O
O
H3N
O
NH3
SHO
O
I
I
I
I
OHNH3
O O
Ác. g-aminobutírico Homocisteína Tiroxina
+
+
+
Tabla 2. Estructura y clasificación de los 20 amino ácidos comunes en las proteínas Amino ácidos Neutros
Estructura PM Pka1 Pka2 PI Sol.**
pf oC o Descomp.
Alanina (Ala, A)
CH3
NH2 OH
O
89.09 2.35 9.78 6.07 17 297 d
Asparagina (Asn, N)
O
NH2CH3
O
NH2
132.12 2.02 8.80 5.41 2.4 236
Cisteína (Cys, C) SH OH
O
NH2
121.16 1.86 8.35 5.11 Muy soluble
-
Glutamina (Gln,Q) OH
O
NH2
NH2
O
146.15 2.17 9.13 5.70 3.6 186
Glicina (Gly,G)
O
OHNH2
75.07 2.35 9.78 6.07 25 233 d
Isoleucina* (Ile, I) OH
O
NH2
CH3
CH3
131.18 2.32 9.76 6.04 4 284
Leucina* (Leu, L) OH
O
NH2
CH3
CH3
131.18 2.33 9.74 6.04 2 337
Metionina* (Met, M) OH
O
NH2
SCH3
149.21 2.28 9.21 5.74 3 283
Fenilalanina* (Phe, F)
O
OH
NH2
165.19 2.58 9.24 5.91 3 283
- 9 -
Prolina (Pro, P)
NH
OH
O
115.13 2.00 10.60 6.30 162 220
Serina (Ser, S)
NH2
OH
O
OH
105.09 2.21 9.15 5.68 5 228
Treonina* (Thr, T)
NH2
OH
O
OH
CH3
119.12 2.09 9.10 5.60 Muy soluble
257
Triptófano* (Trp, W)
NH
OH
O
NH2
204.23 2.38 9.39 5.88 1 289
Tirosina (Tyr, Y)
O
OH
NH2OH
181.19 2.20 9.11 5.66 0.04 344
Valina* (Val, V)
NH2
OH
OCH3
CH3
117.15 2.29 9.72 6.00 9 315
*Amino ácidos esenciales. **Solubilidad en g/100 de H2O a 25 °C. Tabla 1. Continuación Amino ácidos ácidos
Estructura PM Pka1 Pka2 Pka3 PI Sol.** pf °C
Ácido aspártico (Asp, D)
OHOH
O
NH2O
133 2.10 3. 86 9.82 2.98 0.4 269
Ácido glutámico (Glu, E)
OH
O
NH2
OH
O
147 2.10 4.07 9.47 3.08 0.7 247
Amino ácidos Básicos
Arginina* (Arg, R) OH
O
NH2
NHNH2
NH 174 2.01 9.04 12.48 10.76 15 230 d
Histidina* (His,H) OH
O
NH2NHN
155 1.77 6.10 9.18 7.64 0.4 287
Lisina* (Lis, K) OH
O
NH2
NH2
146 2.18 8.95 10.53 9.74 Muy soluble
255
*Amino ácidos esenciales. **Solubilidad en g/100 de H2O a 25 °C.

- 10 -
Problema 1. ¿Cuántos de los amino ácidos que se muestran en la Tabla 2 contienen anillos
aromáticos? ¿Cuántos contienen azufre? ¿Cuántos contienen alcoholes? ¿Cuántos
contienen cadenas laterales hidrocarbonadas?
Con excepción de la glicina, los carbonos alfa de los -amino ácidos son quirales; por lo
tanto, son posibles dos formas enantioméricas. En la naturaleza sólo se encuentra uno de
los dos enantiómeros, el que estructuralmente está relacionado con el L-gliceraldehído. Por
esta razón se ha aceptado que los -amino ácidos que se encuentran en forma natural se
les denominen amino ácidos L o de la familia L.
En las proyecciones de Fischer, los amino ácidos que se encuentran en forma natural se
representan colocando el grupo COOH en la parte superior, la cadena lateral abajo, y el
grupo –NH2 a la izquierda, como se dibuja un carbohidrato.
CHO
CH2OH
H OH
COOH
R
H NH2
CHO
CH2OH
OH H
COOH
R
NH2 H
D-Gliceraldehído(Natural)
L-Gliceraldehído(No Natural)
D--Aminoácido(No Natural)
L--Aminoácido( Natural)
Problema 2. Dieciocho de los 19 L-amino ácidos tienen configuración S en el carbono . La
cisteína es el único L-aminoácido con una configuración R. Explique la razón.
Problema 3. El aminoácido Treonina, ácido (2S, 3R)-2-amino-3-hidroxibutanoico tiene dos
centros quirales. Dibuje una proyección de Fischer para la Treonina. ¿Cuántos
estereoisómeros podrían existir? Dibuje las proyecciones de Fischer e identifique los centros
quirales como R o S.
Problema 4. Dibuje formulas con enlaces de cuña (tetraédricas) y proyecciones de fischer
que representen la configuración de los átomos de carbono de la L-valina, L-leucina y L-
isoleucina.

- 11 -
3.5.-Amino ácidos esenciales Los -amino ácidos que el organismo no puede sintetizar, se les conoce como “Esenciales”.
En la tabla 1 son los 12 amino ácidos en negritas. 3.6.-Fenilcetonuria La fenilcetonuria o PKU es una alteración del metabolismo que ocasiona que no se pueda metabolizar la fenilalanina en el hígado. Es un padecimiento de origen genético que se manifiesta por la carencia de las enzimas fenilalanina hidroxilasa (FAOH) o tirosina hidroxilasa (DHPR). La carencia de alguna de estas enzimas eleva la concentración sanguínea de fenilalanina que se metaboliza por la reacción de transaminación generando un incremento de los metabolitos fenilpiruvato, fenilactato y fenilacetato. Los síntomas son el retraso psicomotor, cuadros psicóticos de tipo autista, convulsiones y eczema facial.
O
NH2
OH
O
NH2OH
OHEnzimas: FAOH o DHPR
Fenilalanina Tirosina REACCIÓN DE TRANSAMINACIÓN Un interesante el papel de las iminas como intermediarios es el de la reacción de transaminación, de importancia biológica. La transaminación es un proceso por el cual se transfiere un grupo amino de una molécula a otra. En los sistemas biológicos el grupo amino de un aminoácido se transfiere al grupo carbonilo de otra molécula. La secuencia, es promovida por una enzima, una aminotransferasa, es un método de formación de nuevos amino ácidos.
+
-cetoglutarato glutamatoUn -amino ácido
CO2
NH3H
R O2C CO2
O
Glutamato
Transaminasa CO2
O
R
Un -ceto ácido
O2C CO2
NH3H
+
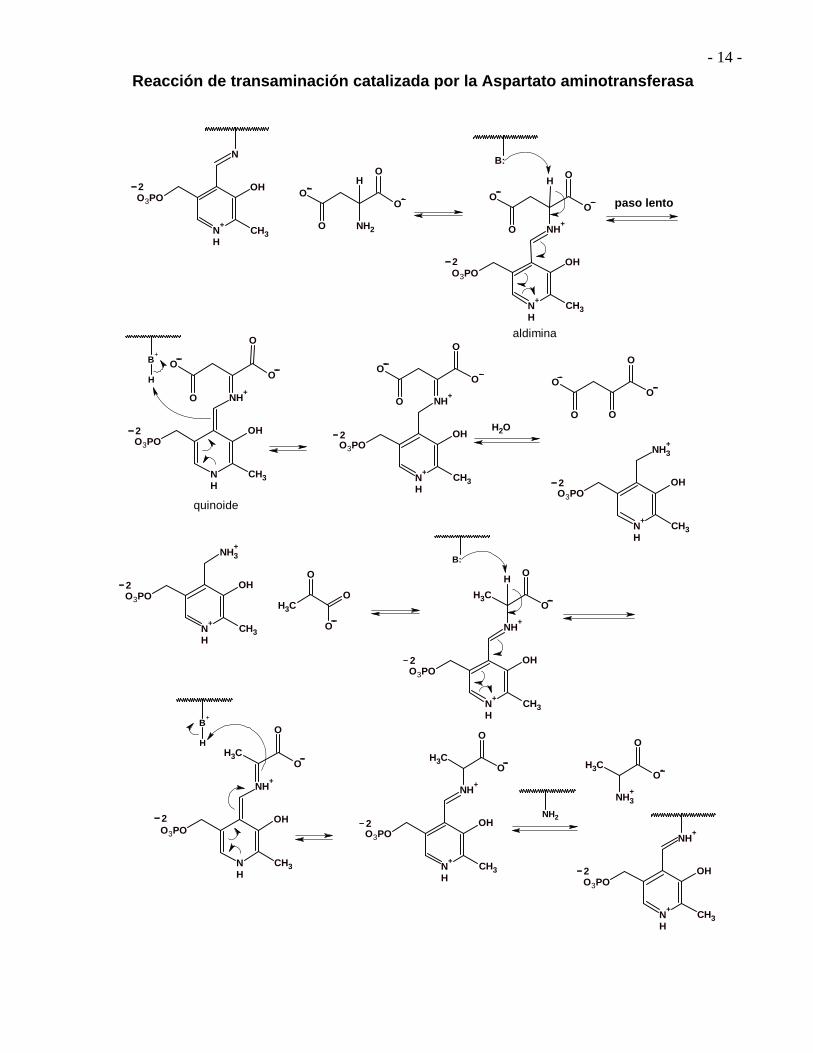
Todas las transaminasas importantes parecen compartir la misma coenzima, el fosfato de piridoxal (PLP). Las coenzimas son pequeños constituyentes no proteínicos de las enzimas, con frecuencia indispensables para la actividad enzimática.

- 12 -
N
H
OHO3PO
CH3
O
H
2
Fosfato de piridoxal (PLP)
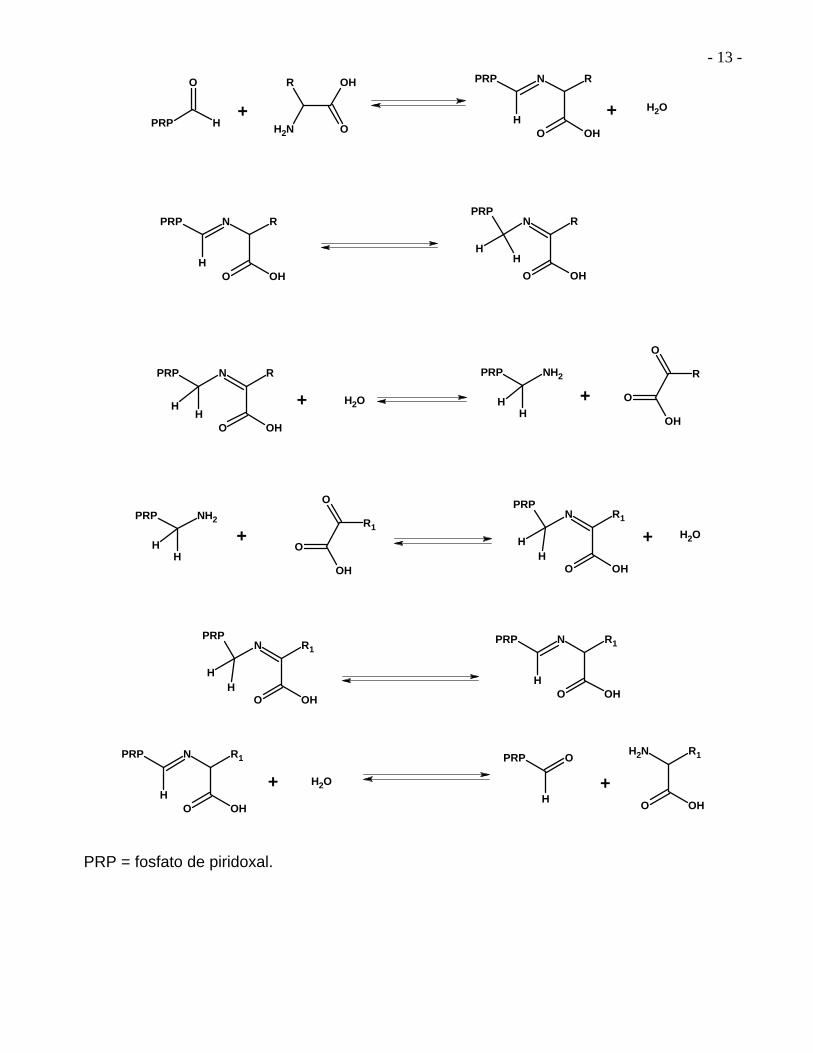
La transaminación biológica no es la simple transferencia de un grupo amino de una molécula a otra. El mecanismo que explica una reacción de transaminación se aprecia mejor por pasos.
Mecanismo general de una reacción de transaminación
- 13 -
O
PRP HO
OH
NH2
R PRP
HO OH
N R
OH2
PRP
HO OH
N RPRP
H
O OH
N R
H
PRP
HO OH
N R
H OH2
PRP
H
NH2
H O
OH
R
O
O
OH
R1
O
PRP
H
NH2
H
PRP
H
N
H
O OH
R1
OH2
PRP
H
N
H
O OH
R1PRP N
HO OH
R1
PRP N
HO OH
R1
OH2
NH2
O OH
R1PRP O
H
+ +
+ +
+ +
+ +
PRP = fosfato de piridoxal.
- 14 -
Reacción de transaminación catalizada por la Aspartato aminotransferasa
N
NH
+
OHO3PO
CH3
O-
O-
NH2O
OH
NH+
NH
+
OHO3PO
CH3
O-O
-
O
OH
B:
NH+
NH
OHO3PO
CH3
O-
O-
O
O
NH+
NH
+
OHO3PO
CH3
O-O
-
O
O
NH3+
NH
+
OHO3PO
CH3
O
O-O
-
O
O
paso lento
CH3 CH3
B+
H
OH2
aldimina
quinoide
NH3+
NH
+
OHO3PO
CH3
O
O-
CH3
O
NH+
NH
+
OHO3PO
CH3
O-
O
CH3
H
NH+
NH
OHO3PO
CH3
O-
O
CH3
NH+
NH
+
OHO3PO
CH3
O-
O
CH3
NH+
NH
+
OHO3PO
CH3
NH3+
O-
O
CH3
CH3 CH3
B+
H
CH3 CH3
NH2
2
2
2 2
2
2
2
2 2
2
CH3 CH3
B:
- 15 -
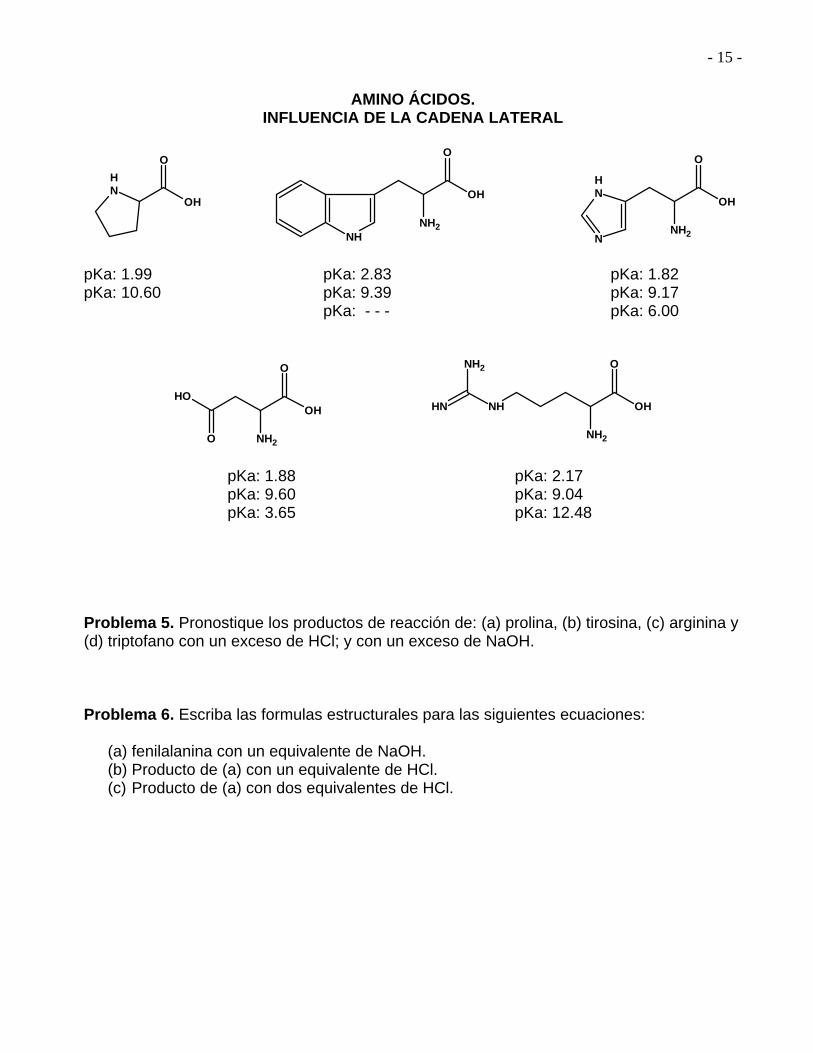
AMINO ÁCIDOS.
INFLUENCIA DE LA CADENA LATERAL
O
NH
OH
O
NH2
NH
N
OH
O
NH2NH
OH
pKa: 1.99 pKa: 2.83 pKa: 1.82 pKa: 10.60 pKa: 9.39 pKa: 9.17 pKa: - - - pKa: 6.00
OHOH
NH2O
O
NH
O
NH
NH2
NH2
OH
pKa: 1.88 pKa: 2.17 pKa: 9.60 pKa: 9.04 pKa: 3.65 pKa: 12.48 Problema 5. Pronostique los productos de reacción de: (a) prolina, (b) tirosina, (c) arginina y (d) triptofano con un exceso de HCl; y con un exceso de NaOH. Problema 6. Escriba las formulas estructurales para las siguientes ecuaciones:
(a) fenilalanina con un equivalente de NaOH. (b) Producto de (a) con un equivalente de HCl. (c) Producto de (a) con dos equivalentes de HCl.
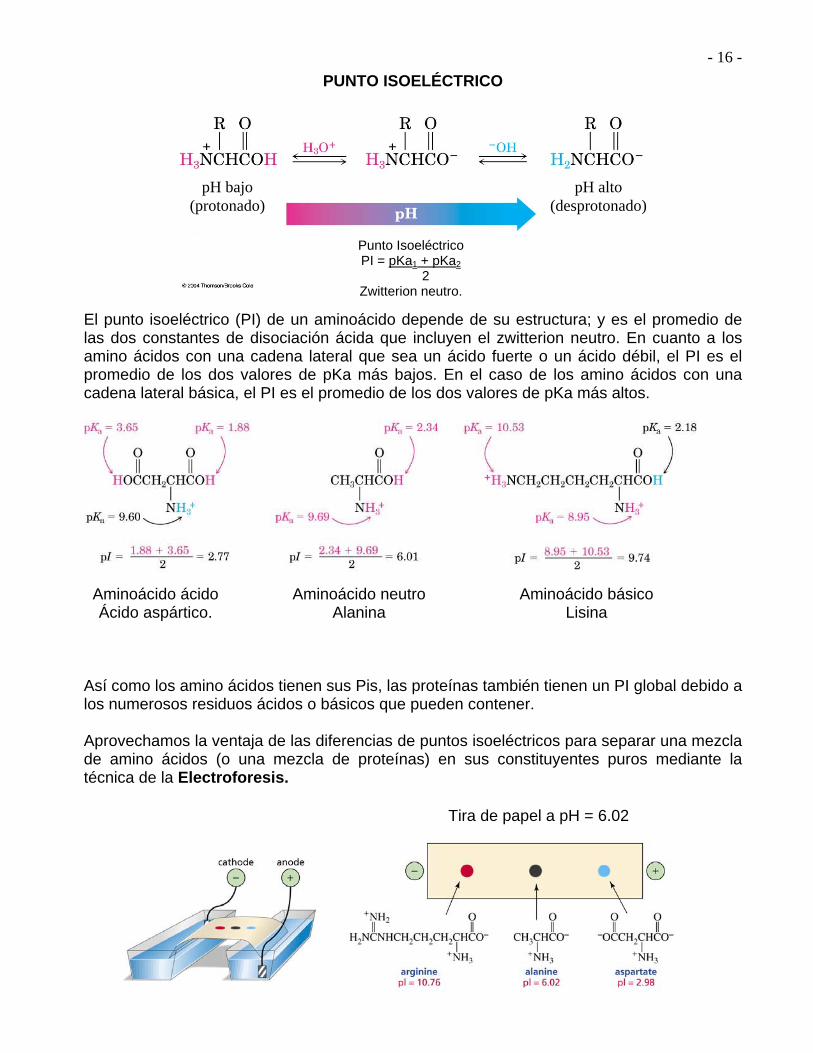
- 16 -
PUNTO ISOELÉCTRICO El punto isoeléctrico (PI) de un aminoácido depende de su estructura; y es el promedio de las dos constantes de disociación ácida que incluyen el zwitterion neutro. En cuanto a los amino ácidos con una cadena lateral que sea un ácido fuerte o un ácido débil, el PI es el promedio de los dos valores de pKa más bajos. En el caso de los amino ácidos con una cadena lateral básica, el PI es el promedio de los dos valores de pKa más altos.
Así como los amino ácidos tienen sus Pis, las proteínas también tienen un PI global debido a los numerosos residuos ácidos o básicos que pueden contener. Aprovechamos la ventaja de las diferencias de puntos isoeléctricos para separar una mezcla de amino ácidos (o una mezcla de proteínas) en sus constituyentes puros mediante la técnica de la Electroforesis.
pH bajo (protonado)
pH alto (desprotonado)
Punto Isoeléctrico PI = pKa1 + pKa2
2 Zwitterion neutro.
Aminoácido ácido Ácido aspártico.
Aminoácido neutro Alanina
Aminoácido básico Lisina
Tira de papel a pH = 6.02

- 17 -
Cuando se aplica un potencial eléctrico, los amino ácidos con cargas negativas (los que se han desprotonado a causa de que el pH del buffer es más alto que sus PI) migra lentamente hacia el electrodo positivo. Al mismo tiempo que los amino ácidos con cargas positivas (los que se han protonado porque el pH del buffer es menor que su PI) migran hacia el electrodo negativo. Los diferentes amino ácidos migran a diferentes velocidades, lo que depende de sus puntos isoeléctricos y del pH del buffer acuoso. Así es posible separar una mezcla de diversos amino ácidos. Problema 7. Para las siguientes mezclas de amino ácidos, prediga la dirección y la velocidad relativa de migración de cada componente:
(a) valina, ácido glutámico e histidina a pH= 7.6 (b) glicina, fenilalanina y serina a pH= 5.7 (c) glicina, fenilalanina y serina a pH= 5.5 (d) glicina, fenilalanina y serina a pH= 6.0
Problema 8. La glicina, al igual que la alanina tiene un punto isoeléctrico de 6.0. Trace estructuras de las formas predominantes de la glicina a pH= 2.0, 6.0 y 10.0. Problema 9. ¿Cómo podría explicar el hecho de que el triptófano tenga menor punto isoeléctrico que la histidina, pese a que ambos tienen átomos de nitrógeno en el anillo de cinco miembros? ¿Cual nitrógeno del anillo de cinco miembros de la histidina es más básico? Un aminoácido reacciona con dos equivalentes del hidrato de ninhidrina, para dar el púrpura de Ruhemann, un producto azul-violeta. Esta reacción se emplea como ensayo para detectar la presencia de amino ácidos en una muestra desconocida.
O
O
OH
OH
O
O
O
OH2
O
O-
NH3+
R
+
O
R
H
CO2 OH2
O
O
N
O-
O
+ + +
ninhidrina
púrpura de Ruhemann.

- 18 -
O
OH
NH3+
CH3
O
O-
NH3+
CH3
O
O-
NH3+
CH3
O
O-
NH2
CH3
OH2
OH2
OH3+
OH3+
+
+ +
+
Si se conocen los valores exactos de pKa para los sitios ácidos de un aminoácido, se puede calcular los porcentajes de las formas protonadas, neutra y desprotonadas en una solución a un pH dado utilizando la estuación de Henderson-Hasselbalch:
pH – pKa = log A-
HA
pH = pKa + log A- HA
Para ver cómo se utiliza la ecuación de Henderson-Hasselbalch, encontremos las especies que están presentes en una solución 1M de alanina a pH= 9.00 de acuerdo a los valores antes mencionados, la alanina protonada tiene un pKa1 de 2.34, y la alanina neutra tiene un pKa2 de 9.69: pKa1= 2.34 pKa2= 9.69
Como el pH de nuestra solución está mucho más cerca del pKa2 que del pKa1, necesitamos usar pKa2 para nuestros cálculos. De acuerdo a la ecuación de Henderson-Hasselbalch, tenemos:
log A- = pH – pKa = 9.0 – 9.69 = - 0.69 HA De tal modo que A- = antilog (-0.69) = 0.20 HA Además, sabemos que: A- + HA = 100% (1.0 M) HA= 83% y A- = 17% Se pueden efectuar cálculos similares a otro pH, con lo que se llega a la curva de titulación siguiente:
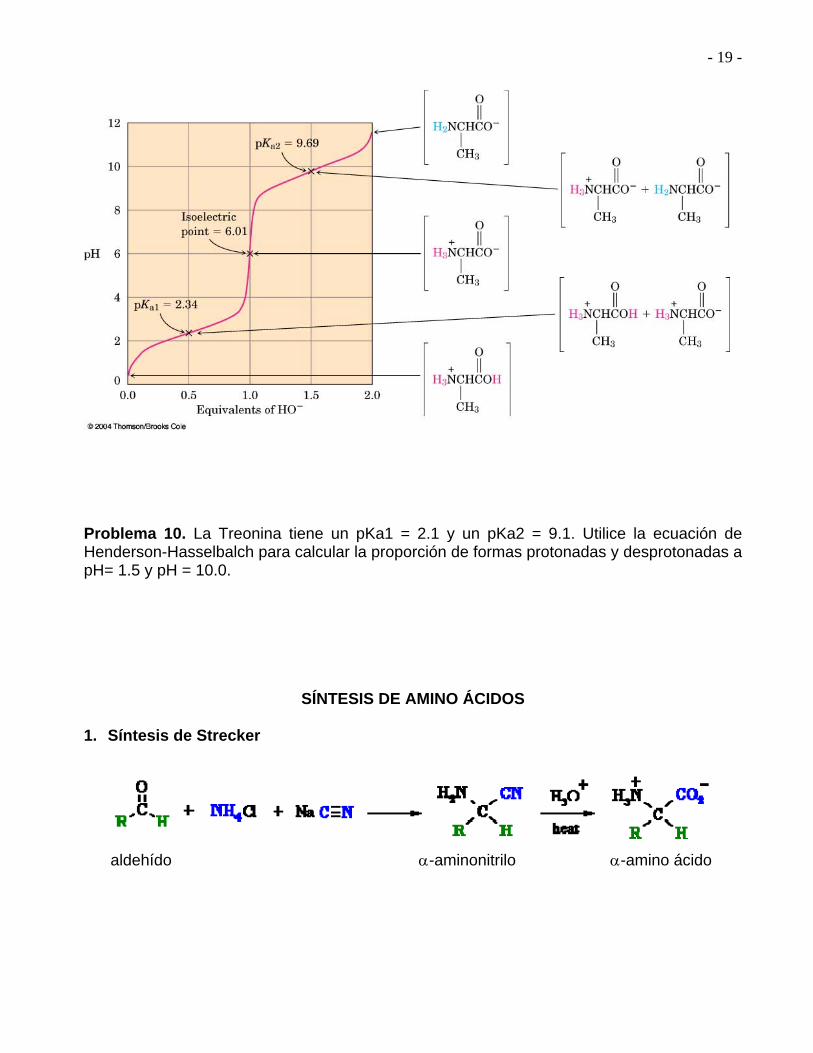
- 19 -
Problema 10. La Treonina tiene un pKa1 = 2.1 y un pKa2 = 9.1. Utilice la ecuación de Henderson-Hasselbalch para calcular la proporción de formas protonadas y desprotonadas a pH= 1.5 y pH = 10.0.
SÍNTESIS DE AMINO ÁCIDOS 1. Síntesis de Strecker
aldehído -aminonitrilo -amino ácido
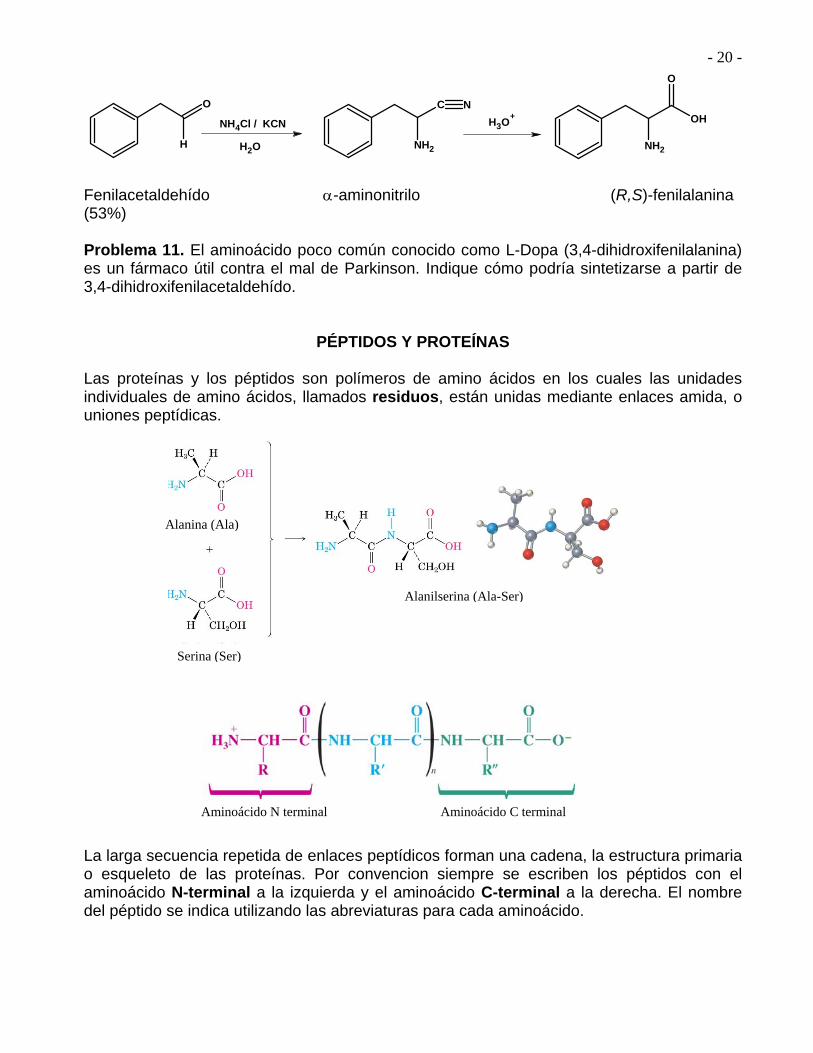
- 20 -
O
H NH2
O
OH
NH2
C N
NH4Cl / KCN
H2O
H3O+
Fenilacetaldehído -aminonitrilo (R,S)-fenilalanina (53%) Problema 11. El aminoácido poco común conocido como L-Dopa (3,4-dihidroxifenilalanina) es un fármaco útil contra el mal de Parkinson. Indique cómo podría sintetizarse a partir de 3,4-dihidroxifenilacetaldehído.
PÉPTIDOS Y PROTEÍNAS Las proteínas y los péptidos son polímeros de amino ácidos en los cuales las unidades individuales de amino ácidos, llamados residuos, están unidas mediante enlaces amida, o uniones peptídicas. La larga secuencia repetida de enlaces peptídicos forman una cadena, la estructura primaria o esqueleto de las proteínas. Por convencion siempre se escriben los péptidos con el aminoácido N-terminal a la izquierda y el aminoácido C-terminal a la derecha. El nombre del péptido se indica utilizando las abreviaturas para cada aminoácido.
Alanina (Ala)
Serina (Ser)
Alanilserina (Ala-Ser)
Aminoácido N terminal Aminoácido C terminal
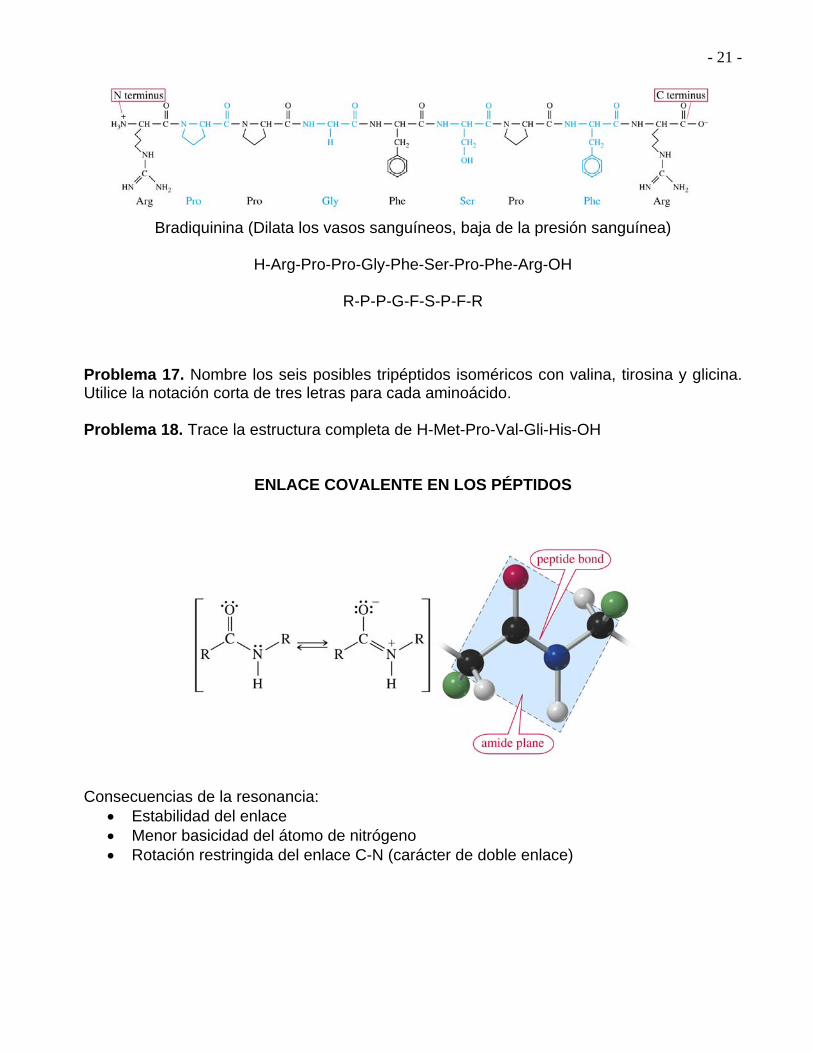
- 21 -
Bradiquinina (Dilata los vasos sanguíneos, baja de la presión sanguínea)
H-Arg-Pro-Pro-Gly-Phe-Ser-Pro-Phe-Arg-OH
R-P-P-G-F-S-P-F-R Problema 17. Nombre los seis posibles tripéptidos isoméricos con valina, tirosina y glicina. Utilice la notación corta de tres letras para cada aminoácido. Problema 18. Trace la estructura completa de H-Met-Pro-Val-Gli-His-OH
ENLACE COVALENTE EN LOS PÉPTIDOS Consecuencias de la resonancia:
Estabilidad del enlace Menor basicidad del átomo de nitrógeno Rotación restringida del enlace C-N (carácter de doble enlace)
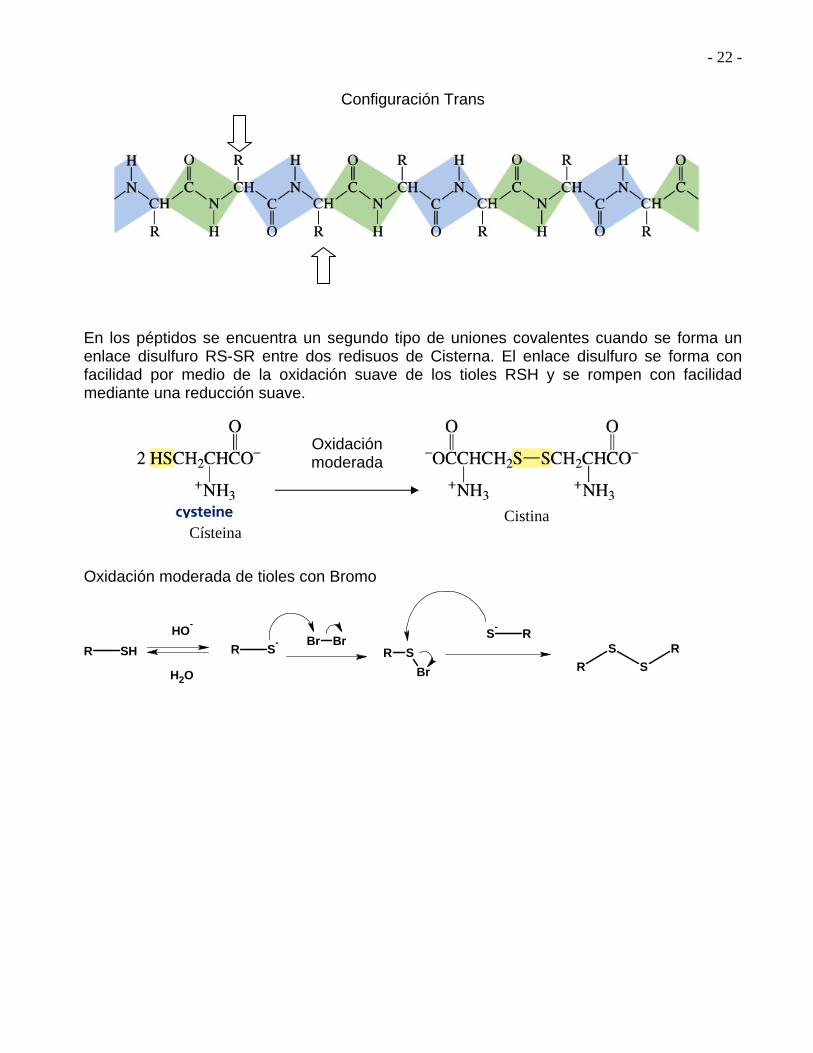
- 22 -
Configuración Trans
En los péptidos se encuentra un segundo tipo de uniones covalentes cuando se forma un enlace disulfuro RS-SR entre dos redisuos de Cisterna. El enlace disulfuro se forma con facilidad por medio de la oxidación suave de los tioles RSH y se rompen con facilidad mediante una reducción suave. Oxidación moderada de tioles con Bromo
R SH
OH-
OH2
R S- Br Br
R S
Br
RS-
R
S
S
R
Oxidación moderada
Císteina Cistina

- 23 -
Los enlaces disulfuro contribuyen a la forma (estructura) de un polipéptido.
Oxitocina
DETERMINACIÓN DE LA ESTRUCTURA DE LOS PÉPTIDO: ANÁLISIS DE LOS AMINO ÁCIDOS
¿Cuáles amino ácidos están presentes? ¿Cuántos hay de cada uno? ¿Cuál es la secuencia de amino ácidos de la cadena peptídica? Las respuestas a las dos primeras preguntas se obtienen con un Analizador de amino ácidos. William Stein y Standford Moore.
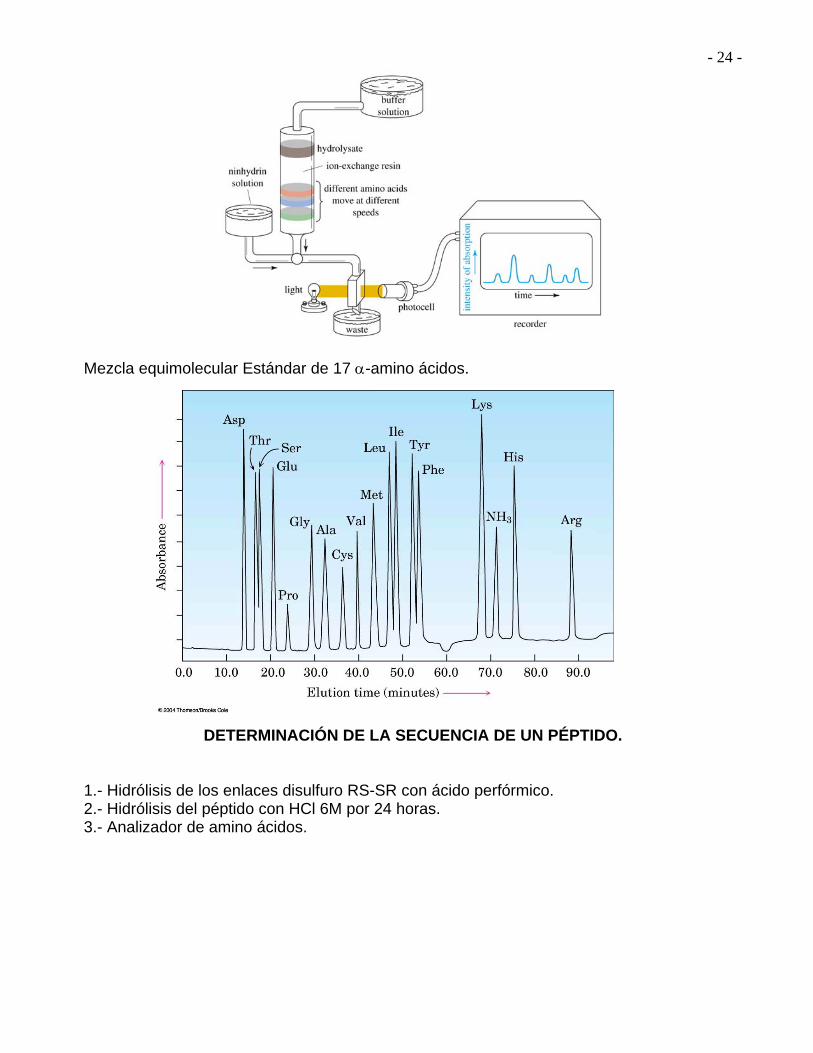
- 24 -
Mezcla equimolecular Estándar de 17 -amino ácidos.
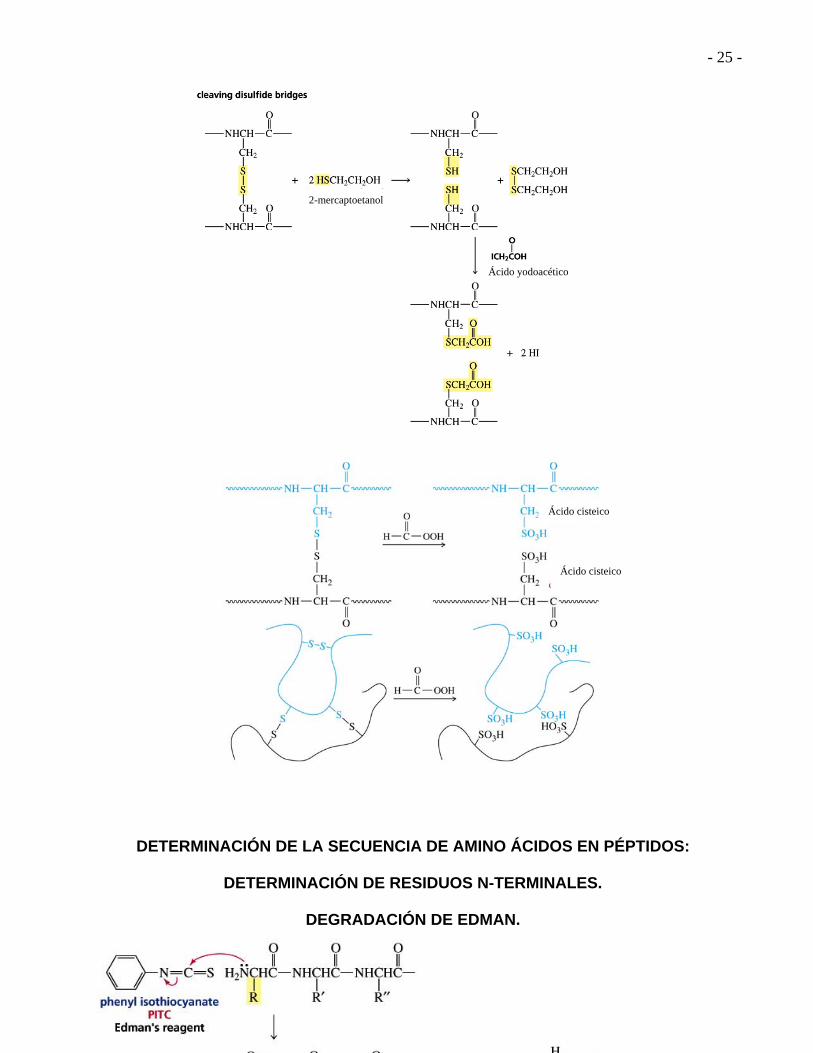
DETERMINACIÓN DE LA SECUENCIA DE UN PÉPTIDO. 1.- Hidrólisis de los enlaces disulfuro RS-SR con ácido perfórmico. 2.- Hidrólisis del péptido con HCl 6M por 24 horas. 3.- Analizador de amino ácidos.
- 25 -
DETERMINACIÓN DE LA SECUENCIA DE AMINO ÁCIDOS EN PÉPTIDOS:
DETERMINACIÓN DE RESIDUOS N-TERMINALES.
DEGRADACIÓN DE EDMAN.
2-mercaptoetanol
Ácido yodoacético
Ácido cisteico
Ácido cisteico
- 26 -
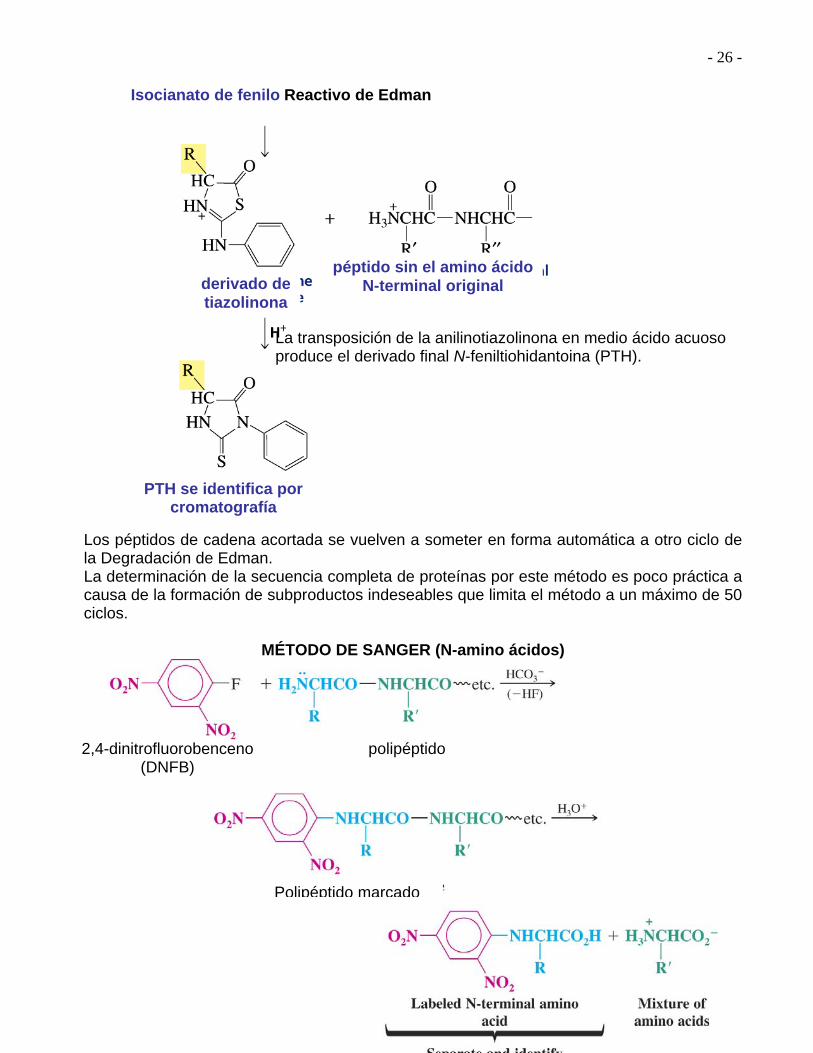
La transposición de la anilinotiazolinona en medio ácido acuoso produce el derivado final N-feniltiohidantoina (PTH). Los péptidos de cadena acortada se vuelven a someter en forma automática a otro ciclo de la Degradación de Edman. La determinación de la secuencia completa de proteínas por este método es poco práctica a causa de la formación de subproductos indeseables que limita el método a un máximo de 50 ciclos.
MÉTODO DE SANGER (N-amino ácidos)
Isocianato de fenilo Reactivo de Edman
derivado de tiazolinona
péptido sin el amino ácido N-terminal original
PTH se identifica por cromatografía
2,4-dinitrofluorobenceno (DNFB)
polipéptido
Polipéptido marcado

- 27 -
El método de Sanger no es tan utilizado como el de Edman. Dadas las limitaciones de los métodos de Edman y Sanger; primero se rompe una cadena peptídica larga por medio de una hidrólisis parcial en varios fragmentos más pequeños, se determina la secuencia de cada fragmento y éstos se ajustan haciendo coincidir los extremos que se traslapan.
HIDRÓLISIS ENZIMÁTICAS. Enzima Especificidad Tripsina hidrólisis carboxilo de amino ácidos básicos: Arginina y Lisina. Quimiotripsina hidrólisis carboxilo de amino ácidos arilo: Fenilalanina, Tirosina y Triptofano. Elastasa hidrólisis carboxilo de amino ácidos pequeños: Glicina y alanina. Carboxipeptidasa A Remueve el aminoácido C-terminal Carboxipeptidasa B Remueve los amino ácidos C-terminal Arginina y Lisina solamente.
Val – Phe – Leu – Met – Tyr – Pro – Gly – Trp – Cys – Glu – Asp – Ile – Lys – Ser – Arg – His
Quimiotripsina rompe estos enlaces Tripsina rompe estos enlaces.
HIDRÓLISIS PARCIAL 1. Un análisis de los amino ácidos de la angiotensina II revelaría la presencia de ocho
amino ácidos diferentes: Arg, Asp, His, Ile, Fen, Pro, Tir y Val en cantidades equimoleculares.
2. Un análisis de extremos N-terminales por el método de Edman indicaría que la
angiotensina II tiene un residuo de ácido aspártico en el extremo N-terminal.
Aminoácido N-terminal marcado.
Separado e identificado
Mezcla de resto de
aminoácidos

- 28 -
3. La hidrólisis parcial de la angiotensina II con ácido clorhídrico diluido produciría los siguientes fragmentos, cuya secuencia puede ser determinada por Degradación de Edman:
a. H-Asp-Arg-Val-OH b. H-Ile-His-Pro-OH c. H-Arg-Val-Tir-OH d. H-Pro-Fen-OH e. H-Val-Tir-Ile-OH
4. Traslapando las regiones que se superponen de los fragmentos se obtiene la secuencia
completa de amino ácidos en la angiotensina II:
a. H-Asp-Arg-Val-OH c. H-Arg-Val-Tir-OH e. H-Val-Tir-Ile-OH b. H-Ile-His-Pro-OH d. H-Pro-Fen-OH
H-Asp-Arg-Val-Tir-Ile-His-Pro-Fen-OH Problema 19. ¿Cual es la estructura de un pentapéptido que da los siguientes tripéptidos cuando se hidroliza parcialmente?
Gli-glu-arg, glu-arg-gli, arg-gli-fen Problema 20. ¿Qué fragmentos resultarían si se rompiera la Angiotensina II con tripsina en un caso, y con quimiotripsina en otro?. Problema 21. Dibuje la estructura del derivado PTH que se puede formar por la degradación de Edman de la Angiotensina II. Problema 22. Indique la secuencia de los hexapéptidos que producen los siguientes fragmentos por hidrólisis parcial con ácido:
(a) Arg,Gli,Ile,Leu,Pro,Val forman H-Pro-Leu-Gli-OH, H-Arg-Pro-OH, H-Gli-Ile-Val-OH (b) Asp,Leu,Met,Trp, Val2 forman H-Val-Leu-OH, H-Val-Met-Trp-OH, H-Trp-Asp-Val-OH

- 29 -
DETERMINACIÓN DE RESIDUOS C-TERMINALES
Problema 23. Se encuentra que un hexapéptido con la composición Arg, Gli, Leu, Pro tiene prolina en los extremos C y N. La hidrólisis parcial produce los siguientes fragmentos:
H-Gli-Pro-Arg-OH H-Arg-Pro-OH H-Pro-Leu-Gli-OH ¿Cuál es la estructura del hexapéptido? Problema 24. Proponga dos estructuras para un tripéptido que por hidrólisis se descompone en Leu, Ala, y Fen pero que no reacciona con carboxipeptidasa ni con fenilisotiocianato. El bromuro de cianógeno hidroliza el enlace peptídico de la Metionina.
H – Ala – Lys – Phe – Asp – Met – Val – Arg – Trp – OH
BrCN rompe este enlace. Mecanismo de la hidrólisis del enlace peptídico por bromuro de cianógeno.
carboxipeptidasa

- 30 -
SÍNTESIS DE PÉPTIDOS. Aunque las amidas simples suelen formarse por la reacción entre aminas y cloruros de ácidos, la síntesis de los péptidos es más difícil porque se deben de formar muchos enlaces amida diferentes en orden específico y no aleatoriamente. 2 sitios de unión + 2 sitios de unión = 4 productos distintos.
- 31 -
O
OR
O
OCl
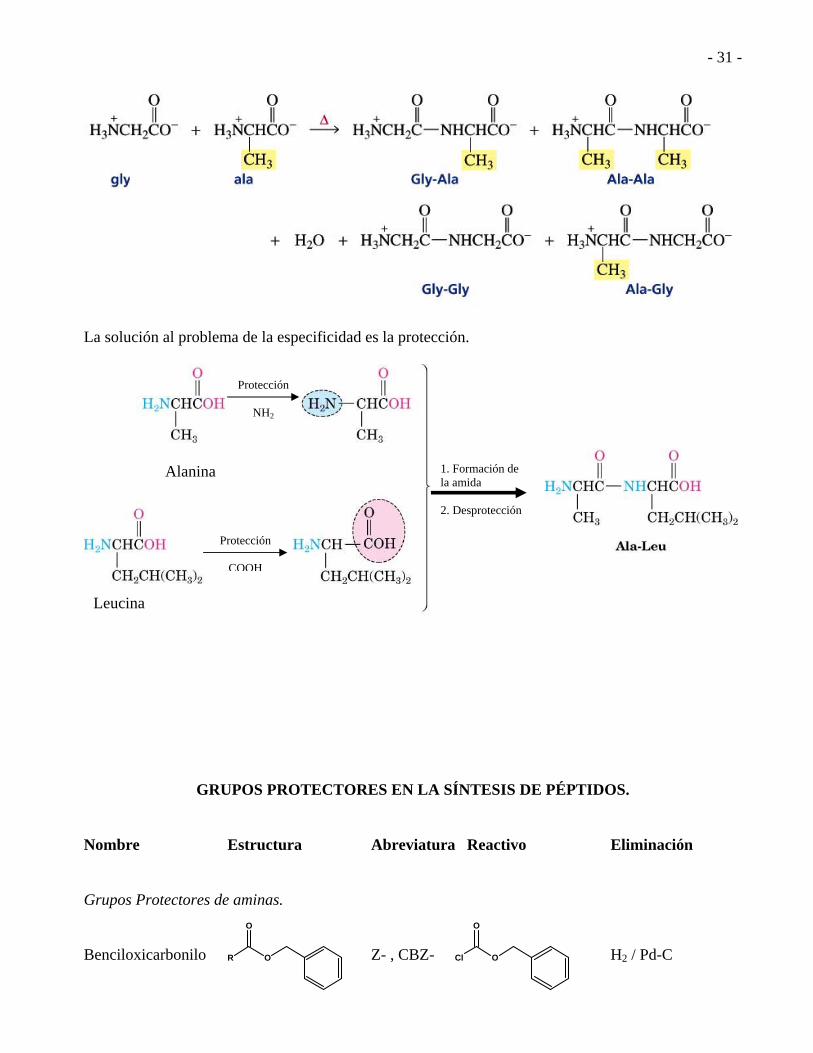
La solución al problema de la especificidad es la protección.
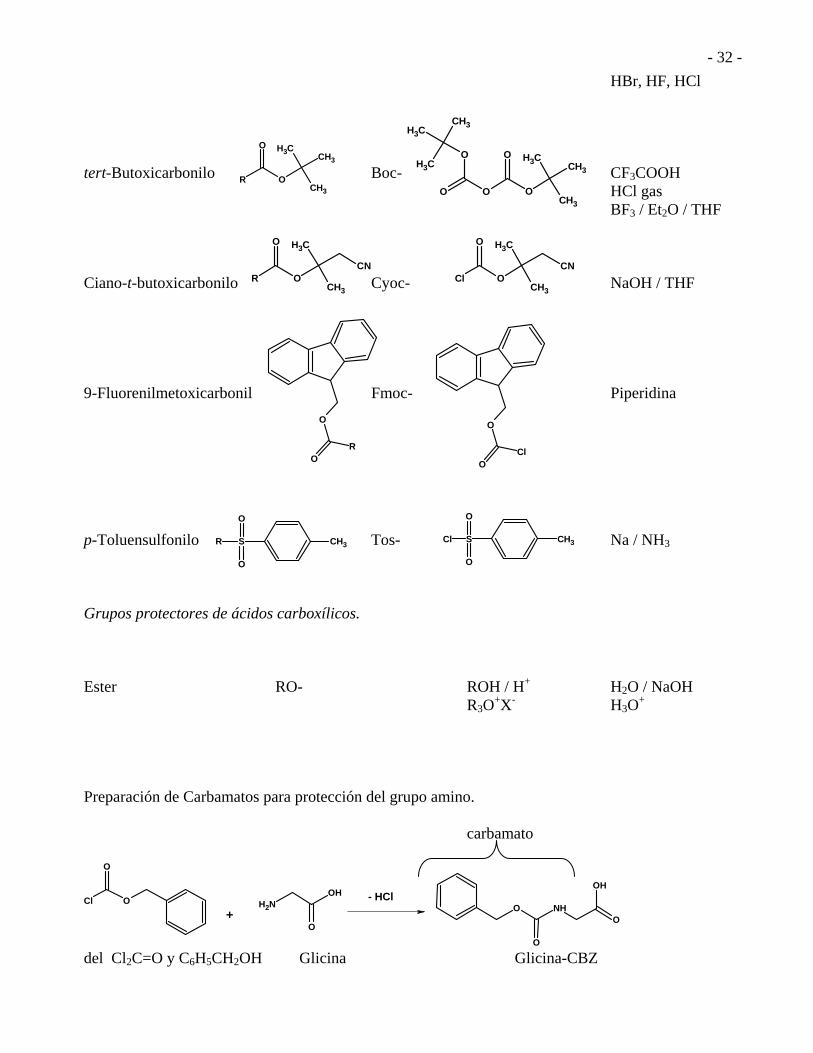
GRUPOS PROTECTORES EN LA SÍNTESIS DE PÉPTIDOS. Nombre Estructura Abreviatura Reactivo Eliminación Grupos Protectores de aminas. Benciloxicarbonilo Z- , CBZ- H2 / Pd-C
Alanina
Leucina
Protección
NH2
Protección
COOH
1. Formación de la amida 2. Desprotección
- 32 -
O
O CH3CH3
CH3
R
O
O
CH3
CH3
CH3
O O
O CH3CH3
CH3
O
O CH3
CH3
CNR O
O CH3
CH3
CNCl
O
OR
O
OCl
S
O
O
CH3R S
O
O
CH3Cl
HBr, HF, HCl tert-Butoxicarbonilo Boc- CF3COOH HCl gas BF3 / Et2O / THF Ciano-t-butoxicarbonilo Cyoc- NaOH / THF 9-Fluorenilmetoxicarbonil Fmoc- Piperidina p-Toluensulfonilo Tos- Na / NH3 Grupos protectores de ácidos carboxílicos. Ester RO- ROH / H+ H2O / NaOH R3O
+X- H3O+
Preparación de Carbamatos para protección del grupo amino.
carbamato
O
OCl
O
OHNH2
O
OO
OH
NH+
- HCl
del Cl2C=O y C6H5CH2OH Glicina Glicina-CBZ
- 33 -
+
O
OO
O
NH
O
O
CH3
O
NH2
OH
O
OO
NH
NH
O
OH
O
OCl CH3
ZBC
O
OHNH
+- HCl ZBC
O
ONH
O
O
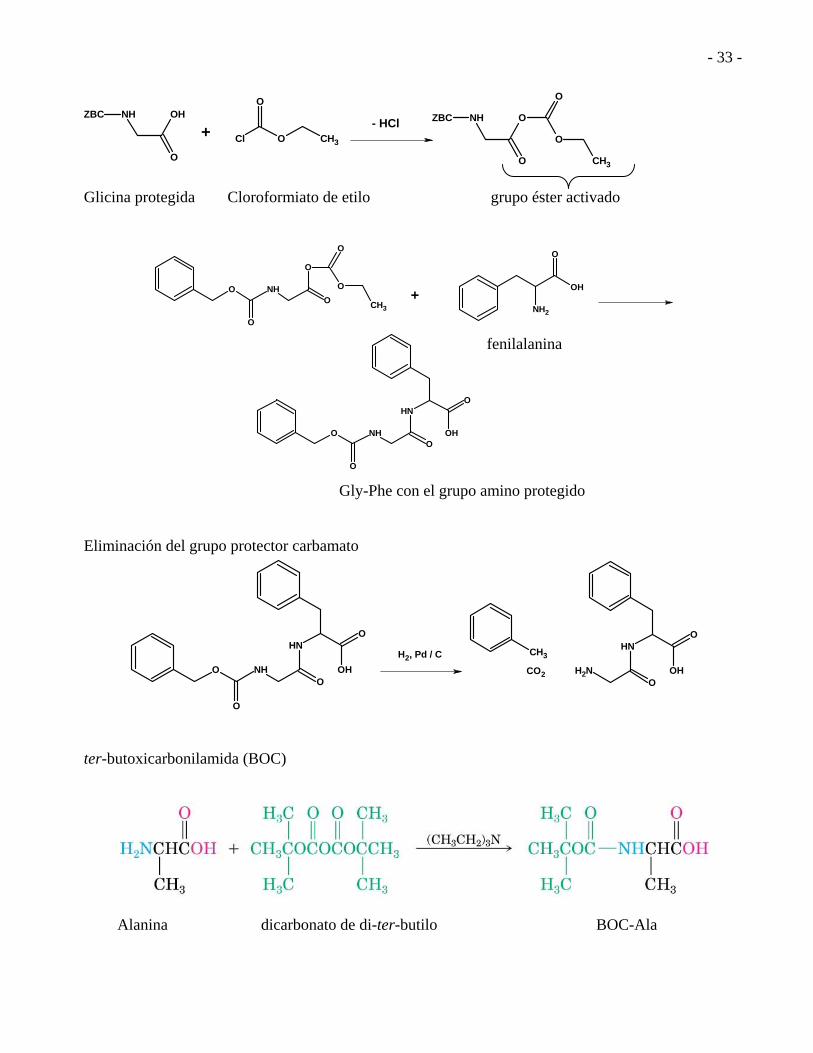
CH3 Glicina protegida Cloroformiato de etilo grupo éster activado
fenilalanina Gly-Phe con el grupo amino protegido Eliminación del grupo protector carbamato
O
OO
NH
NH
O
OH
H2, Pd / C CH3
CO2O
NH
NH2
O
OH
ter-butoxicarbonilamida (BOC)
Alanina dicarbonato de di-ter-butilo BOC-Ala
- 34 -
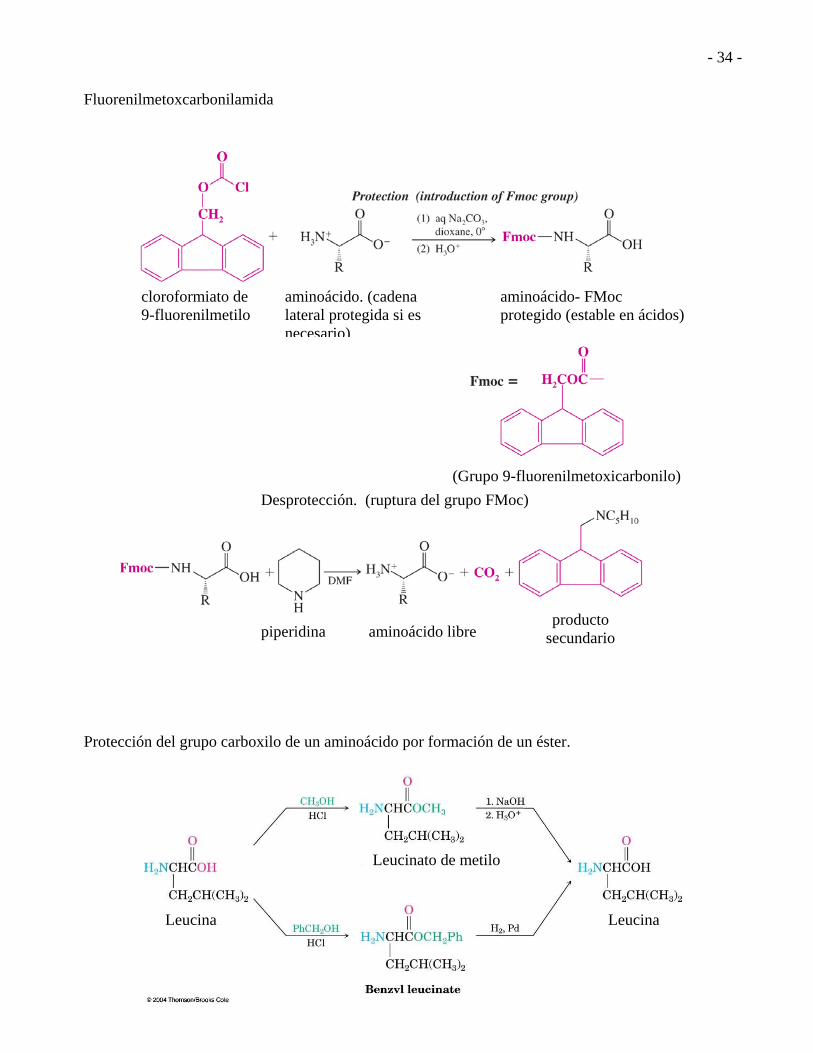
Fluorenilmetoxcarbonilamida Protección del grupo carboxilo de un aminoácido por formación de un éster.
Leucina
Leucinato de metilo
Leucina
cloroformiato de 9-fluorenilmetilo
aminoácido. (cadena lateral protegida si es necesario)
aminoácido- FMoc protegido (estable en ácidos)
piperidina
aminoácido libre
producto secundario
(Grupo 9-fluorenilmetoxicarbonilo)
Desprotección. (ruptura del grupo FMoc)
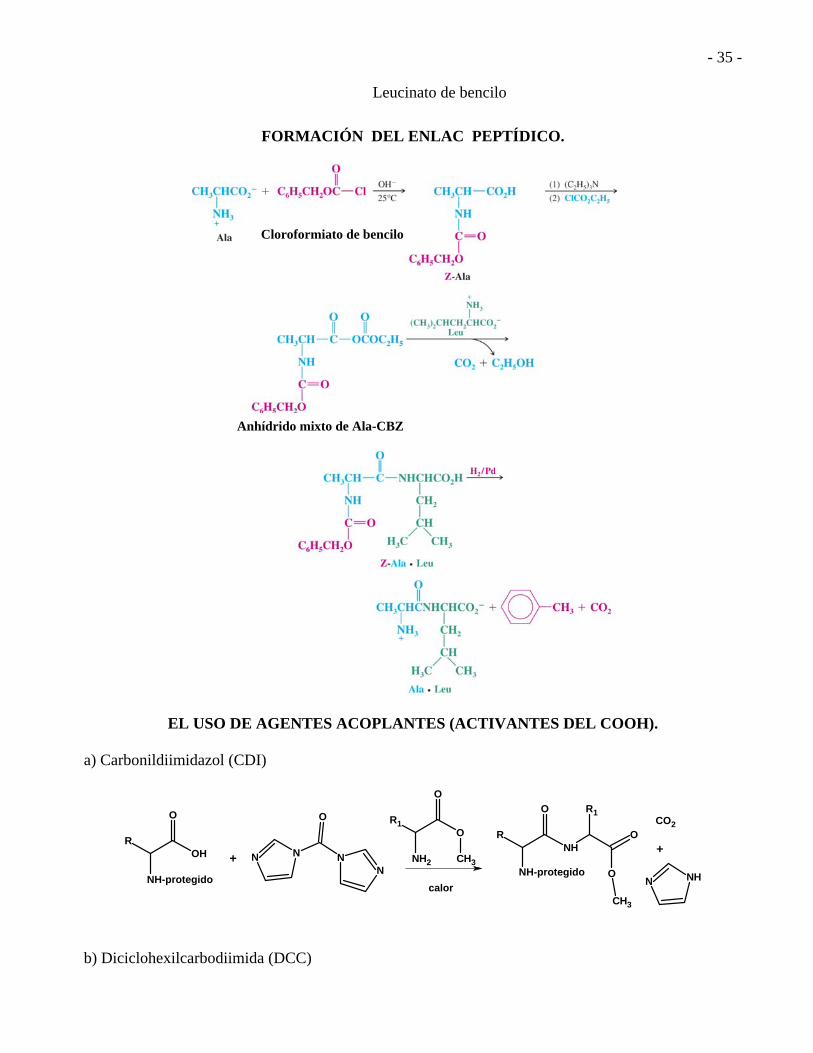
- 35 -
FORMACIÓN DEL ENLAC PEPTÍDICO.
EL USO DE AGENTES ACOPLANTES (ACTIVANTES DEL COOH). a) Carbonildiimidazol (CDI)
O
OH
NH-protegido
R
++
calor
N N NN
O CO2
N NH
O
O
NH2
R1
CH3
O
NH
NH-protegido
R O
O
R1
CH3 b) Diciclohexilcarbodiimida (DCC)
Leucinato de bencilo
Cloroformiato de bencilo
Anhídrido mixto de Ala-CBZ
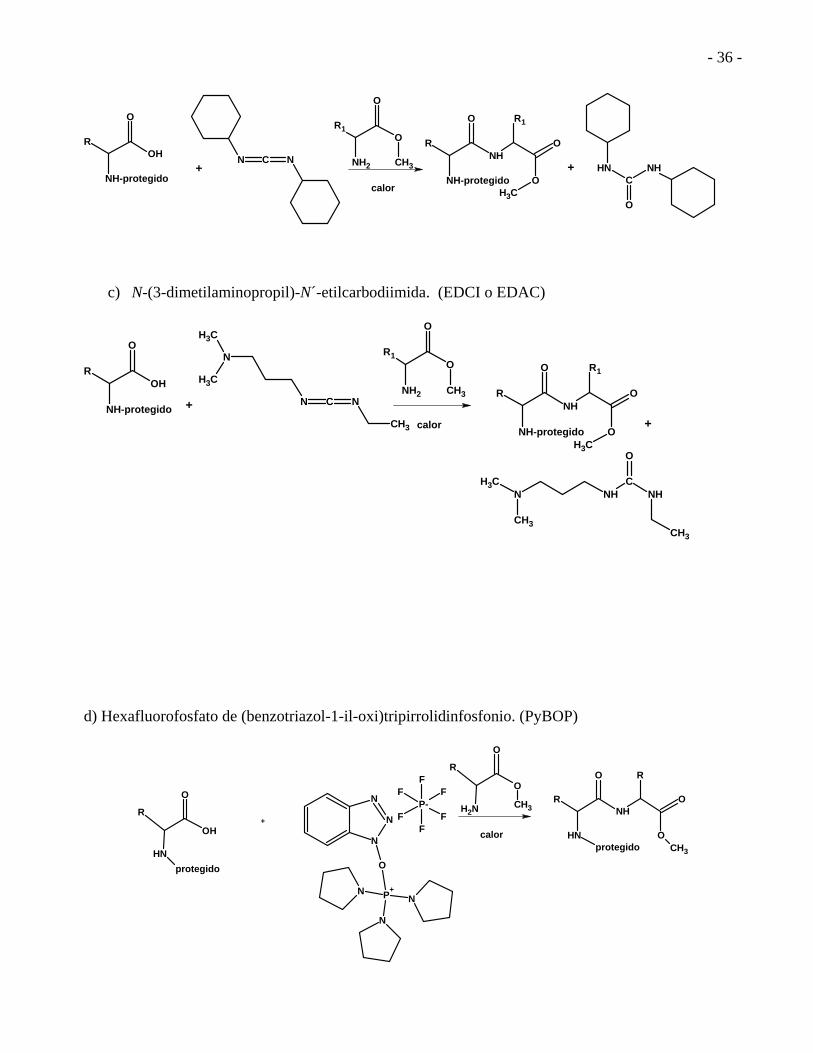
- 36 -
+ +
calor
N C NNH
CNH
O
O
OH
NH-protegido
R
O
O
NH2
R1
CH3
O
NH
NH-protegido
R O
O
R1
CH3
c) N-(3-dimetilaminopropil)-N´-etilcarbodiimida. (EDCI o EDAC)
+
+calor
N
N C
CH3
N
CH3
CH3
N NHC
CH3
NHCH3
CH3
O
O
OH
NH-protegido
R
O
O
NH2
R1
CH3
O
NH
NH-protegido
R O
O
R1
CH3
d) Hexafluorofosfato de (benzotriazol-1-il-oxi)tripirrolidinfosfonio. (PyBOP)
O
OH
R
NH
protegido
+
calor
N
N
N
O
P+
N
NN
P-
F
FF
FF
F
O
O
R
NH2CH3
O
NHR
NHprotegido
O
O
R
CH3

- 37 -
Resumen de 5 pasos para la síntesis de H-Ala-Leu-OH 1. Proteger el grupo amino de la leucina como su derivado BOC
H-Ala-OH + bicarbonato de di-tert-butilo BOC-Ala-OH 2. Proteger el grupo carbonilo de la leucina como su éster metílico
H-Leu-OH + CH3OH H-Leu-OCH3 3. Acoplar los dos amino ácidos protegidos usando DCC
BOC-Ala-OH + H-Leu-OCH3 BOC-Ala-Leu-OCH3 4. Retirar el grupo protector BOC por tratamiento ácido
BOC-Ala-Leu-OCH3 + CF3COOH H-Ala-Leu-OCH3 5. Retirar el éster metílico por hidrólisis básica:
H-Ala-Leu-OCH3 + NaOH / H2O H-Ala-Leu-OH Problema 25. Indique el mecanismo de la formación de un derivado BOC para la reacción de Tirosina con bicarbonato de di-ter-butilo. Problema 26. Escriba los cinco pasos que se requieren para la síntesis de H-Leu-Ala-OH a partir de alanina y leucina. Problema 27. ¿Cómo podrían prepararse los siguientes tripéptidos?
(a) H-Leu-Ala-Gli-OH (b) H-Gli-Leu-Ala-OH (c) H-Ala-Gli-Leu-OH

- 38 -
La insulina esta compuesta por dos cadenas que suman un total de 51 amino ácidos unidos pos dos puentes disulfuro. Frederick Sanger determinó su estructura y recibió el premio Nobel en Química en 1958 por su trabajo.
SÍNTESIS DE PÉPTIDOS AUTOMATIZADA: TÉCNICA DE FASE SÓLIDA DE MERREFIELD.
La síntesis de péptidos se efectúa sobre esferas sólidas de poliestireno, preparado de un modo tal que por cada 100 anillos de benceno uno lleve un grupo clorometilo:
CH
CH3
CH3
CH2 HC CH2 CH CH2 CH CH2 CH
CH3
CH3
Cl Cl
Síntesis de un tripéptido utilizando la Fase sólida de Merrifield.
Aminoácido N-protegido
cadena A 21 aminoácidos
cadena B 30 aminoácidos
insulina
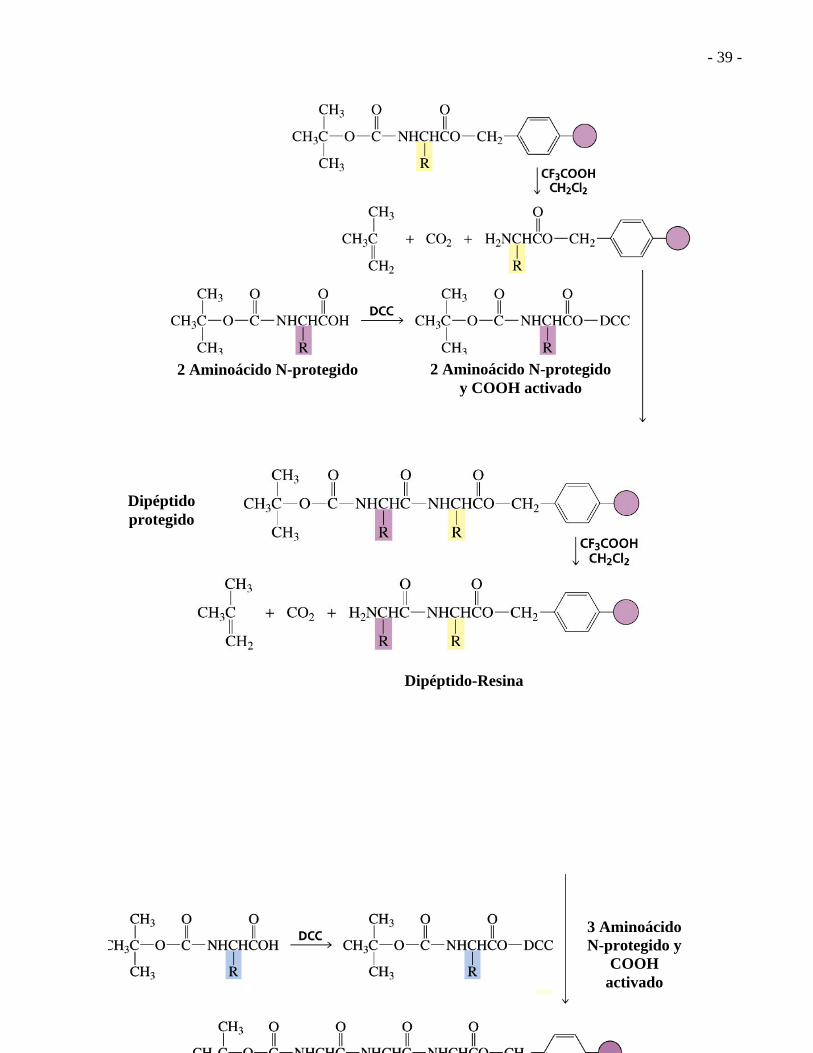
- 39 -
2 Aminoácido N-protegido 2 Aminoácido N-protegido y COOH activado
Dipéptido protegido
Dipéptido-Resina
3 Aminoácido N-protegido y
COOH activado
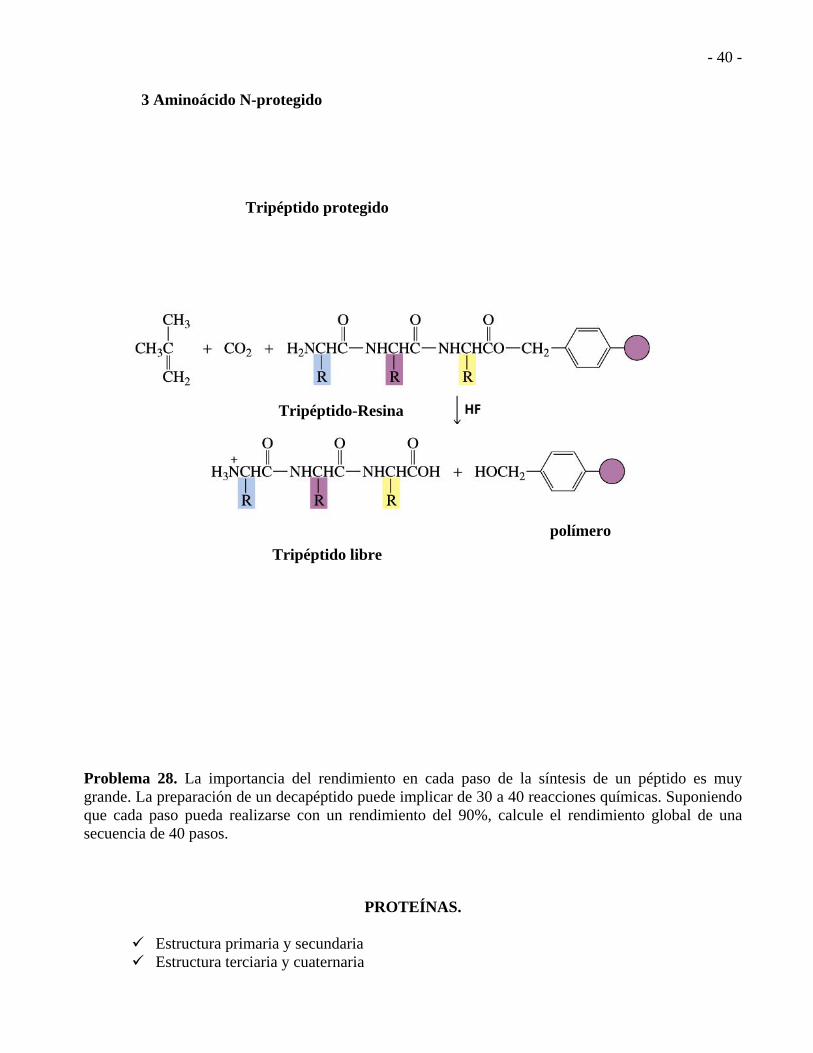
- 40 -
Problema 28. La importancia del rendimiento en cada paso de la síntesis de un péptido es muy grande. La preparación de un decapéptido puede implicar de 30 a 40 reacciones químicas. Suponiendo que cada paso pueda realizarse con un rendimiento del 90%, calcule el rendimiento global de una secuencia de 40 pasos.
PROTEÍNAS.
Estructura primaria y secundaria Estructura terciaria y cuaternaria
3 Aminoácido N-protegido
Tripéptido protegido
Tripéptido-Resina
Tripéptido libre
polímero
- 41 -
Clasificación y funciones de proteínas Enzimas Desnaturalización de proteínas
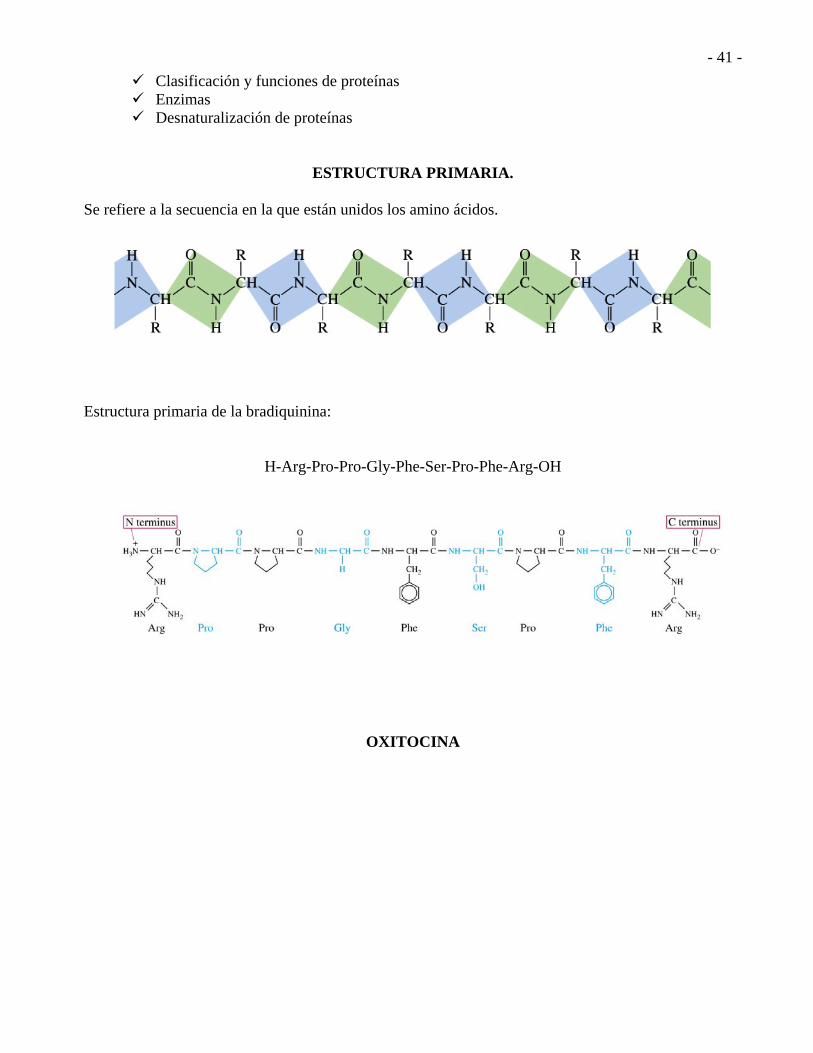
ESTRUCTURA PRIMARIA. Se refiere a la secuencia en la que están unidos los amino ácidos.
Estructura primaria de la bradiquinina:
H-Arg-Pro-Pro-Gly-Phe-Ser-Pro-Phe-Arg-OH
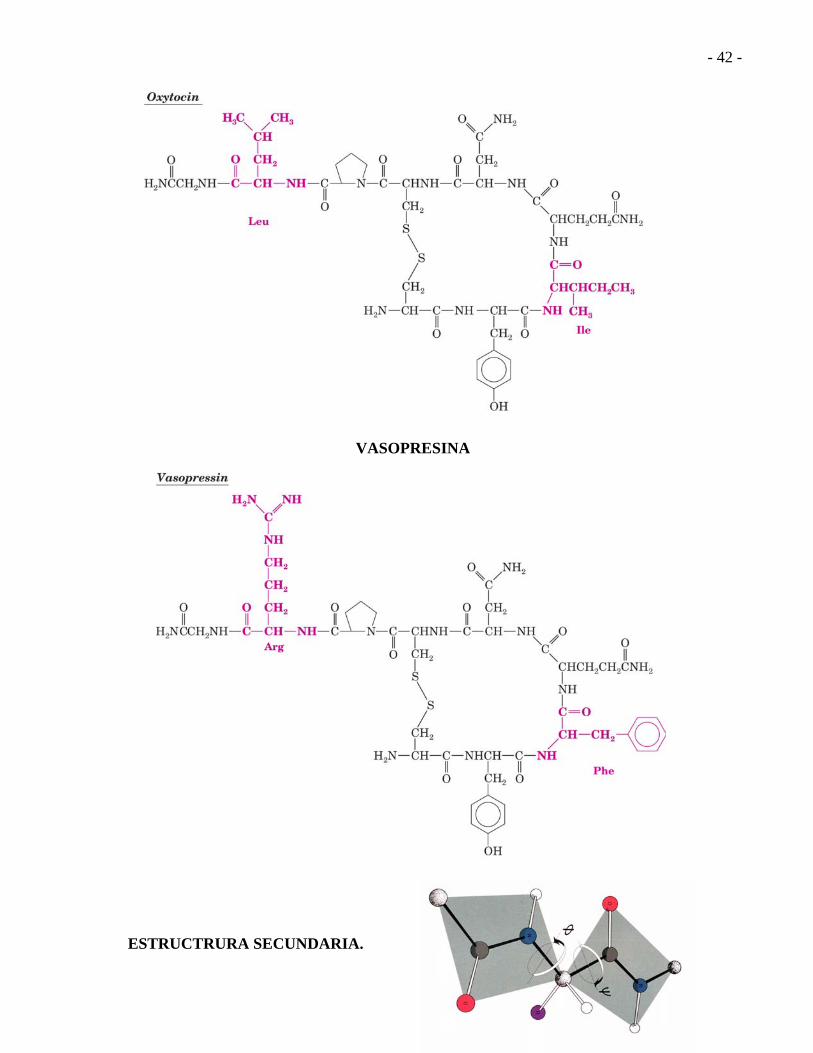
OXITOCINA
- 42 -
VASOPRESINA
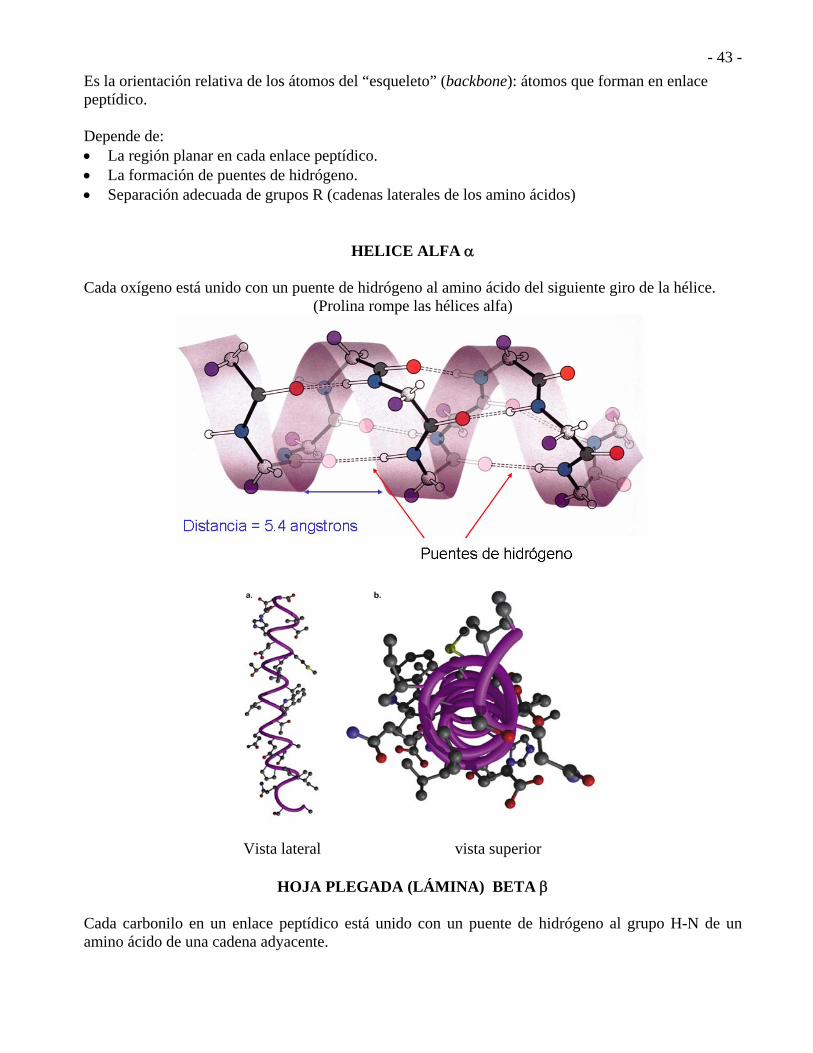
ESTRUCTRURA SECUNDARIA.
- 43 -
Es la orientación relativa de los átomos del “esqueleto” (backbone): átomos que forman en enlace peptídico. Depende de: La región planar en cada enlace peptídico. La formación de puentes de hidrógeno. Separación adecuada de grupos R (cadenas laterales de los amino ácidos)
HELICE ALFA Cada oxígeno está unido con un puente de hidrógeno al amino ácido del siguiente giro de la hélice.
(Prolina rompe las hélices alfa)
Vista lateral vista superior
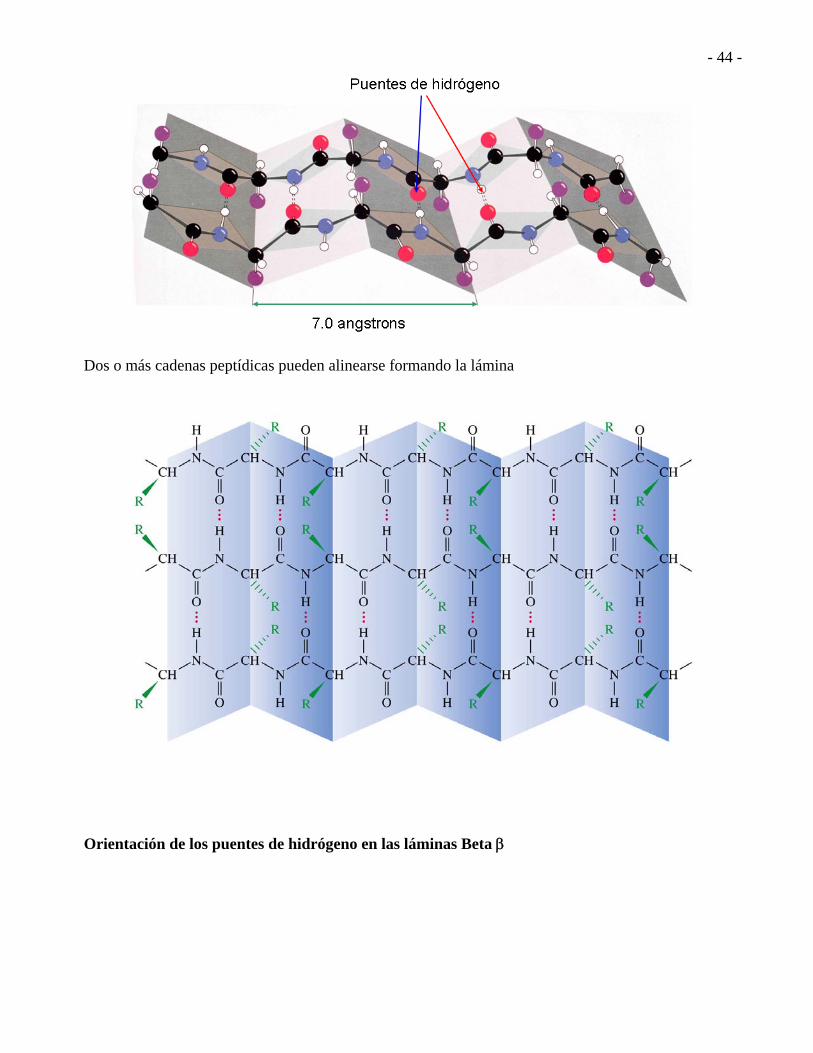
HOJA PLEGADA (LÁMINA) BETA Cada carbonilo en un enlace peptídico está unido con un puente de hidrógeno al grupo H-N de un amino ácido de una cadena adyacente.
- 44 -
Dos o más cadenas peptídicas pueden alinearse formando la lámina Orientación de los puentes de hidrógeno en las láminas Beta
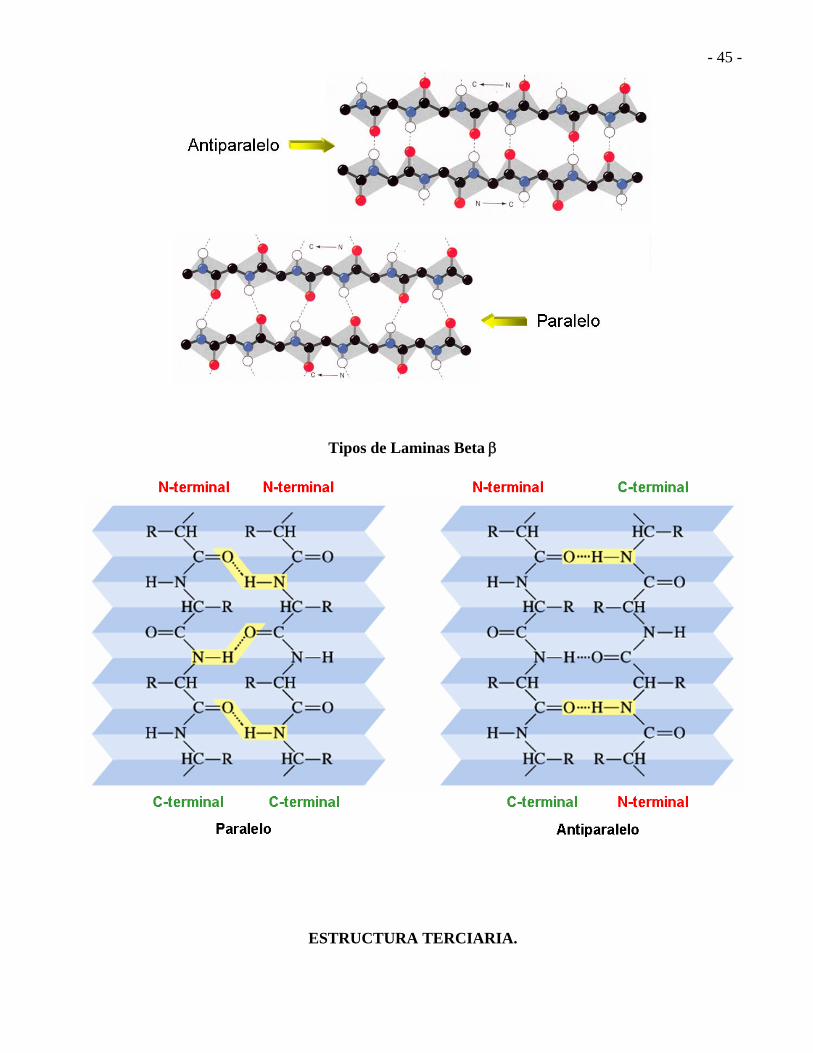
- 45 -
Tipos de Laminas Beta
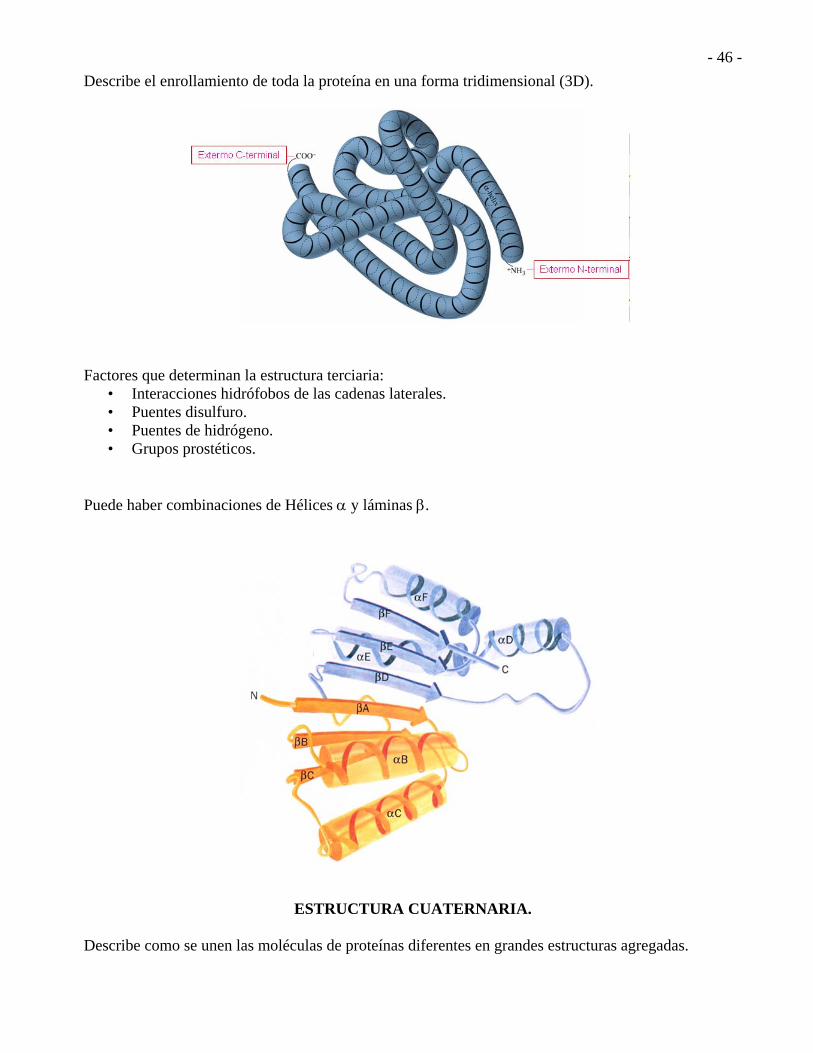
ESTRUCTURA TERCIARIA.
- 46 -
Describe el enrollamiento de toda la proteína en una forma tridimensional (3D).
Factores que determinan la estructura terciaria:
• Interacciones hidrófobos de las cadenas laterales. • Puentes disulfuro. • Puentes de hidrógeno. • Grupos prostéticos.
Puede haber combinaciones de Hélices y láminas .
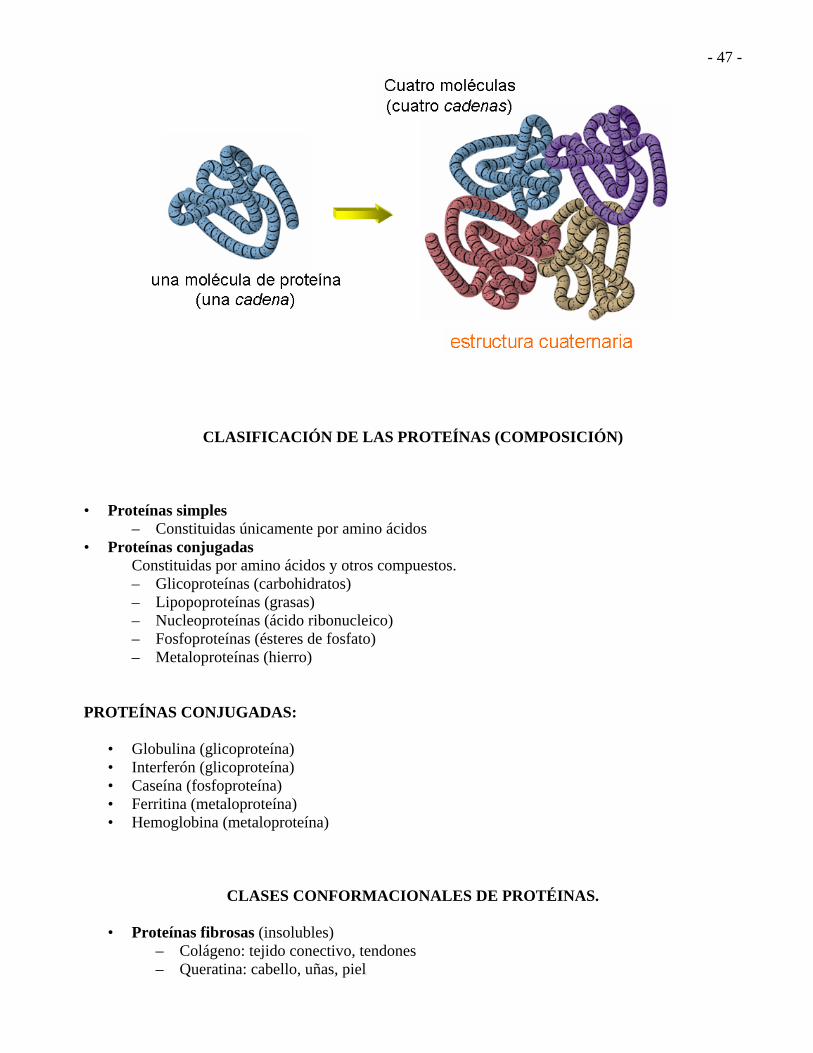
ESTRUCTURA CUATERNARIA. Describe como se unen las moléculas de proteínas diferentes en grandes estructuras agregadas.
- 47 -
CLASIFICACIÓN DE LAS PROTEÍNAS (COMPOSICIÓN) • Proteínas simples
– Constituidas únicamente por amino ácidos • Proteínas conjugadas
Constituidas por amino ácidos y otros compuestos. – Glicoproteínas (carbohidratos) – Lipopoproteínas (grasas) – Nucleoproteínas (ácido ribonucleico) – Fosfoproteínas (ésteres de fosfato) – Metaloproteínas (hierro)
PROTEÍNAS CONJUGADAS:
• Globulina (glicoproteína) • Interferón (glicoproteína) • Caseína (fosfoproteína) • Ferritina (metaloproteína) • Hemoglobina (metaloproteína)
CLASES CONFORMACIONALES DE PROTÉINAS.
• Proteínas fibrosas (insolubles) – Colágeno: tejido conectivo, tendones – Queratina: cabello, uñas, piel

- 48 -
– Elastina: tejido conectivo elástico • Proteínas globulares (solubles)
– Insulina: hormona reguladora del metabolismo de la glucosa – Mioglobina: transporte de oxígeno – Ribonucleasa: controla la síntesis del RNA
ALGUNAS FUNCIONES BIOLÓGICAS DE LAS PROTEÍNAS.
• Enzimas (catalizadores biológicos) • Hormonas (insulina) • Proteínas protectoras (anticuerpos) • Proteínas de almacenamiento (caseína) • Proteínas estructurales (queratina, elastina) • Proteínas de transporte (hemoglobina)
ENZIMAS.
• Casi todas son globulares • Algunas son conjugadas:
– La parte no proteica se llama cofactor – La parte proteica se llama apoenzima – Cofactor + apoenzima = holoenzima
• Son específicas
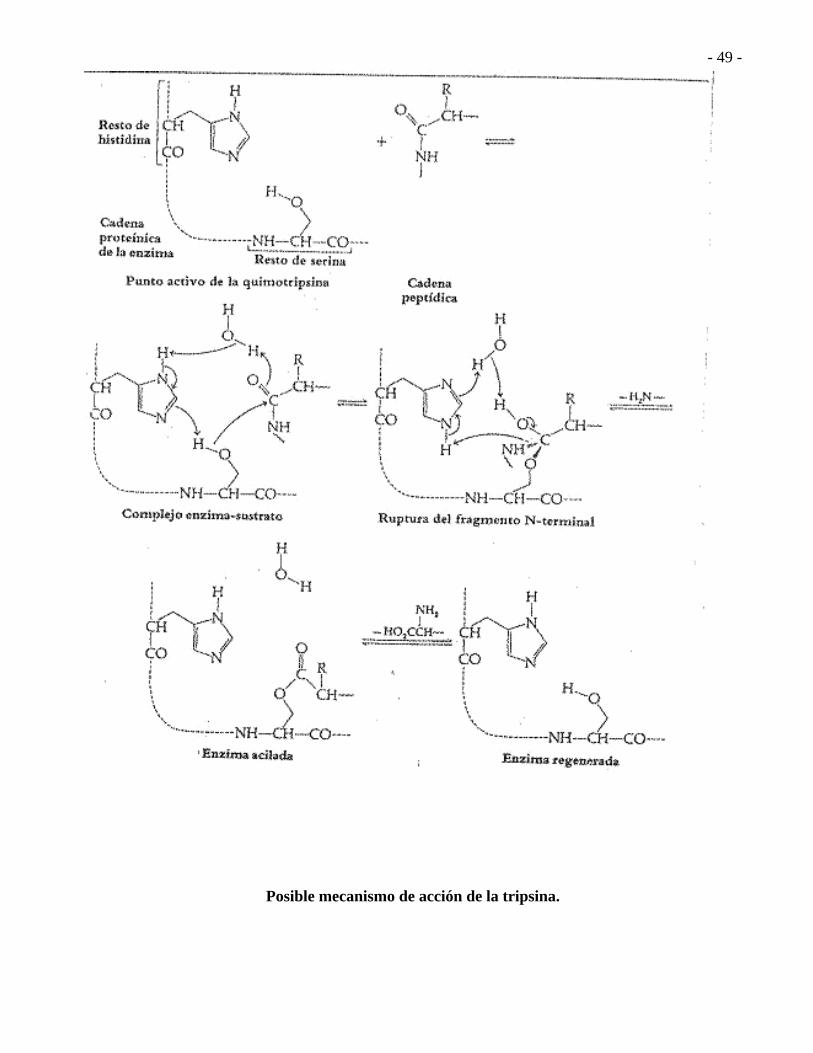
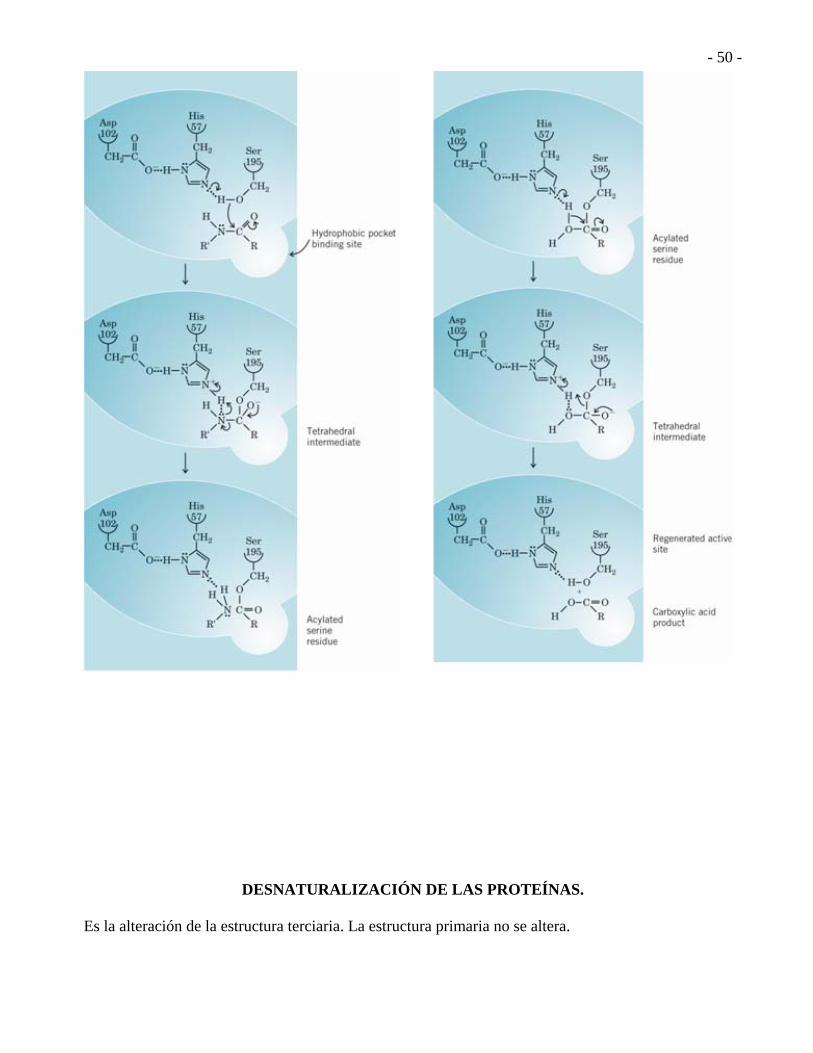
Posible mecanismo de acción de la quimiotripsina.
- 49 -
Posible mecanismo de acción de la tripsina.
- 50 -
DESNATURALIZACIÓN DE LAS PROTEÍNAS. Es la alteración de la estructura terciaria. La estructura primaria no se altera.

- 51 -
ESTRUCTURAS DE PROTEÍNAS EN INTERNET.
Protein Data Bank http://www.rcsb.org/pdb/

- 52 -
N+
S
CH3
CH3
CH3
CCH3
OHH
Problema 29. se cree que la descarboxilación de los -cetoácidos catalizada por tiamina se inicia con la eliminación de un átomo de hidrógeno ácido del anillo de tiazol de la tiamina.
N+
S
CH3
H
CH3
CH3
N+
C- S
CH3
CH3
CH3
H+
+
a. ¿Por qué es ácido este hidrógeno? b. Proponga un mecanismo para la descarboxilación del ácido pirúvico (CH3COCO2H) mediante
tiamina. Se sabe que el intermedio final es la especie A.
A = La vitamina ácido lipoico es una coenzima en la conversión de A y CoA en acetil coenzima A. indique de que manera el intermedio A puede justificar la formación de acetil coenzima A a partir del piruvato inicial.
S
S
(CH2)4 CO2H





























