Embed Size (px)
Citation preview

An Electron Microscope Study of Sarcoma I in a HomologousHost. I. The Cells of the Growing Tumor*
0
VELMAC. CHAMBERSAND RUSSELL,S. WEISER(Department of Microbiology, University of Washington, School of Medicine, Seattle, Washington)
SUMMARY
The fine structure of Sarcoma I cells from growing ascites tumors of C57BL/6Kmice (homologous strain) was compared with the fine structure of cells from the ascitestumors of A/Jax mice (isologous strain). No differences were observed in the tumorcells grown in either strain of mouse. The following ultrastructural features weredemonstrated: numerous free ribosomes generally occurring in clusters; a relativelysmall amount of rough-surfaced endoplasmic reticulum; a fine filamentous componentof the cytoplasm; large, spherical lipide bodies; occasional annulate lamellae; and occasional doughnut-shaped, virus-like bodies within cisternae of endoplasmic reticulum.Similar ultrastructural features have been described by other workers in studies ofvarious kinds of tumor cells.
Sarcoma I is a transplantable tumor that originated in1947 in a mouse of the A/Jax strain after treatment withdibenzanthracene. The ascites form of the tumor growsin the A/Jax mouse, producing death on the 12th to the14th day. After transplantation to the peritoneal cavityof the homologous C57BL/6K mouse the sarcoma cellsproliferate for about 6 days, at which time an immuneresponse begins and the tumor is rejected completelybetween the 10th and the 14th day.
The cytology of Sarcoma I cells obtained from theperitoneal cavity of the A/Jax and the C57BL/6K mousewas described by Baker, Weiser, Jutila, Evans, and Blan-dau (1). Their report contributed valuable informationconcerning the cytological features of the tumor cellsrevealed by light microscopy, phase-contrast microscopy,and phase-contrast cinematography.
In the present study the ultrastructural features of theSarcoma I cell during growth of the tumor in the homologous C57BL/6K mouse are described. These featuresare compared with those of the sarcoma cell from thesusceptible A/Jax mouse.
MATERIALS AND METHODS
Sarcoma I cells.—SarcomaI cells were obtained in 1958from the Roscoe B. Jackson Memorial Laboratory, BarHarbor, Maine. The tumor has been maintained in ourlaboratory by weekly serial transfer in the peritonealcavity of A/Jax mice, the strain of origin. Sarcoma cellswere removed from the peritoneal cavity of an A/Jaxmouse on the 7th or 8th day of tumor growth and were
* Supported in part by United States Public Health ServiceGrant CA-05698.
Received for publication December 16, 1963.
injected intraperitoneally into C57BL/6K mice for thestudies in the homologous host.
Electron microscopy.—Cells were removed from theperitoneal cavity by means of a 25-gauge needle attachedto a tuberculin syringe. The needle was removed fromthe syringe, and the cells were transferred immediatelyto cold osmium tetroxide buffered with s-collidine. Theconcentration of osmium was 2.67 per cent and that ofs-collidine was 0.067 M before the addition of cells. Fixation time was between 40 minutes and 1 hour. The cellswere rinsed briefly in distilled water or in 0.067 M s-collidine buffer, dehydrated in alcohol and in propylene oxide,and embedded in epoxy resin (14).
Sections were cut with a glass knife or with a Dupontdiamond knife mounted on an LKB Ultratome or on aPorter-Blum microtome. Thin sections were stainedwith lead hydroxide (16) or with 2 per cent uranyl acetateand were viewed with an RCA EMU-2C, -3E, or -3Gelectron microscope.
RESULTS
The ascitic fluid removed from C57BL/6K mice 6 daysafter transplantation of Sarcoma I intraperitoneally contained numerous sarcoma cells and a few host macrophages. Although the presence of host macrophagesmarked the onset of rejection, the majority of tumor cellsat this time appeared unchanged. The number of hostcells increased during the 7th and 8th days, and by the9th day the cells were predominantly macrophages. Tumor rejection was complete by the 10th to the 14th day.
The cellular component of the ascitic fluid from tumor-bearing A/Jax mice was almost entirely sarcoma cellsthroughout the course of the tumor, which terminated indeath of the mice. A comparison of sarcoma cells from
693
on April 8, 2019. © 1964 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

694 Cancer Research Vol. 24, May 1964
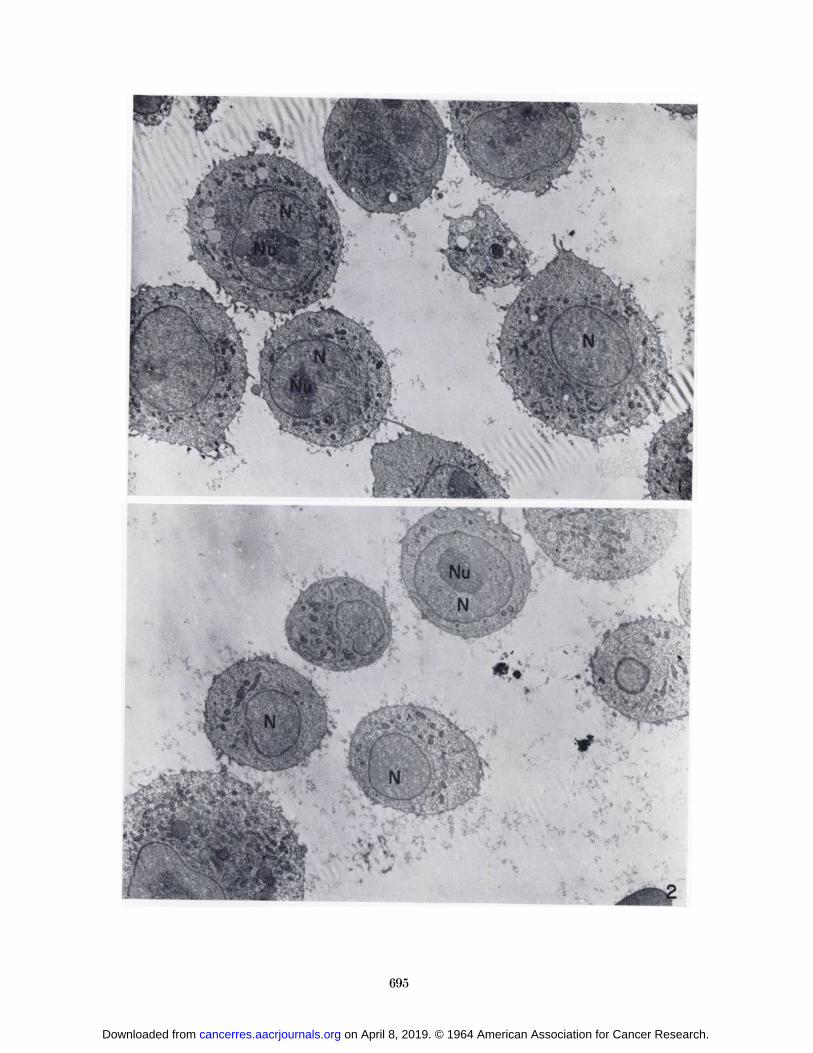
a C57BL/6K mouse on the 6th day and an A/Jax mouseon the 8th day is seen in low magnification electron micrographs in Figures 1 and 2, respectively.
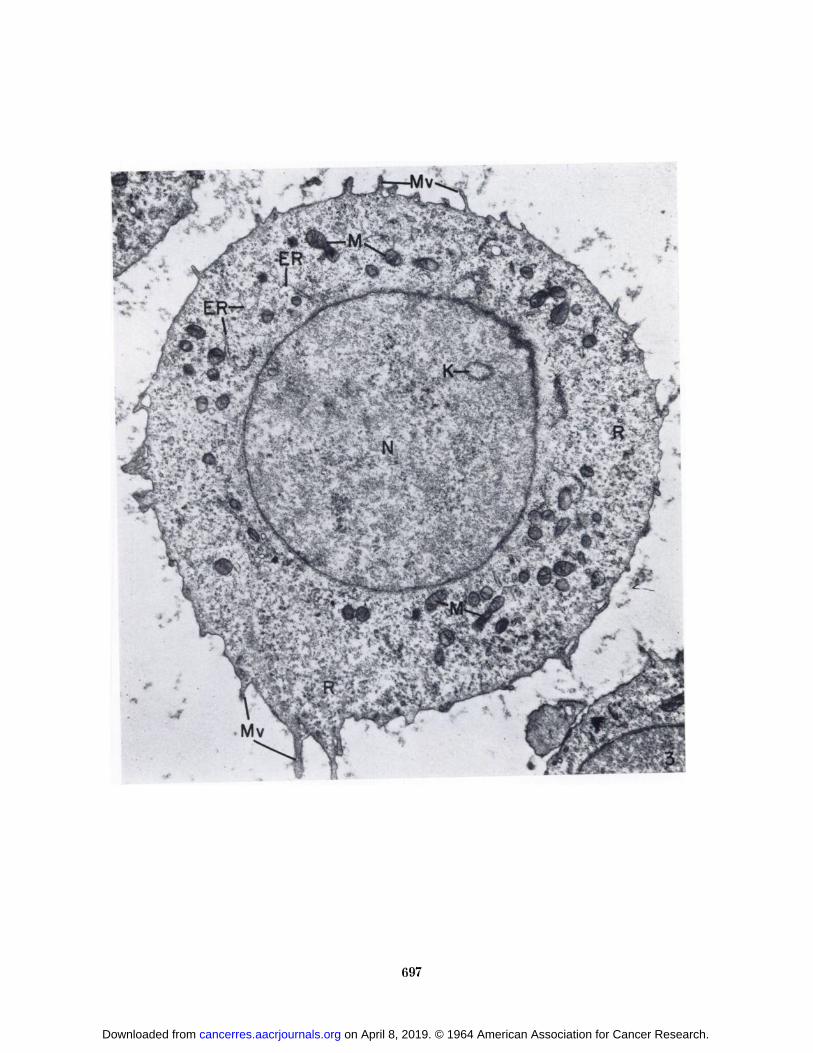
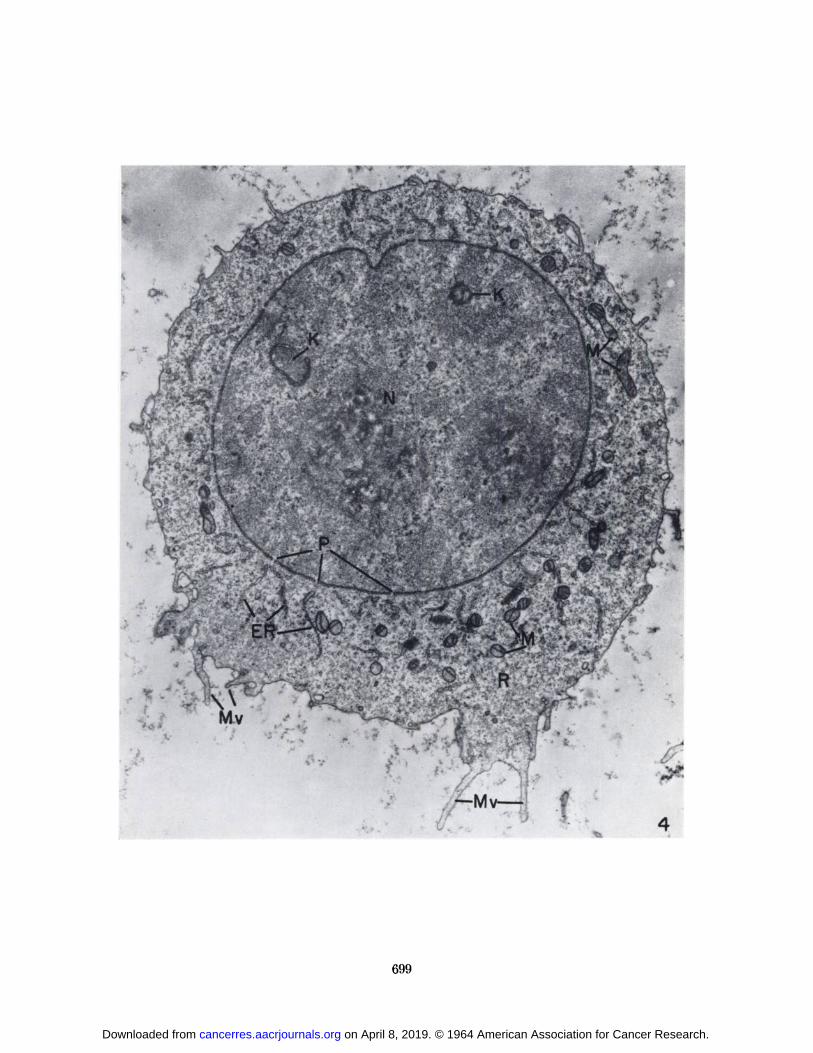
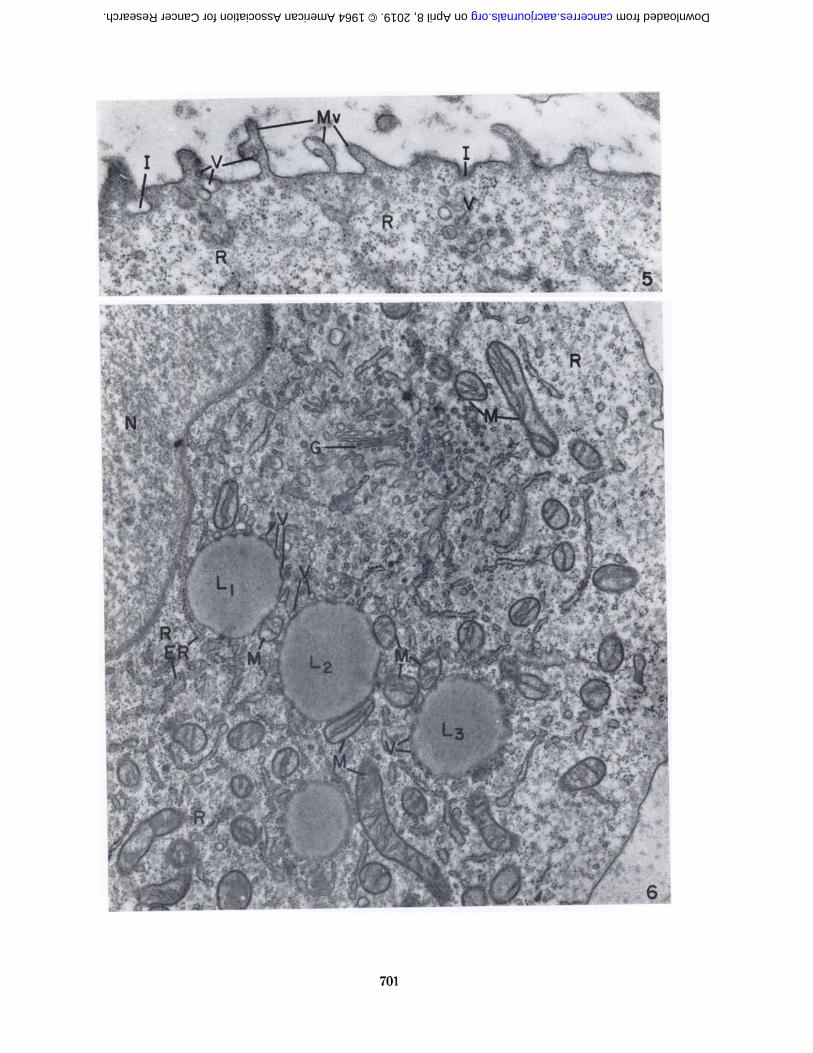
At higher magnification, the fine structure of SarcomaI cells from the C57BL/6K mice during growth of thetumor (Fig. 3) was similar to that of sarcoma cells fromthe A/Jax mice (Fig. 4). The cells were usually spherical,with microvilli of varying shape, length, and number protruding from the cell surface. Small vesicles often occurred within the microvilli or at their bases (Fig. 5).In addition to the outfolding of the cell surface to formmicrovilli, small invaginations of the cell surface were seen(Fig. 5). It is not possible to distinguish between invaginations that represent an intermediate step in theformation of pinocytotic vesicles for the uptake of material into the cell and those that are formed as the resultof vesicles discharging their contents into the extracellularmedium.
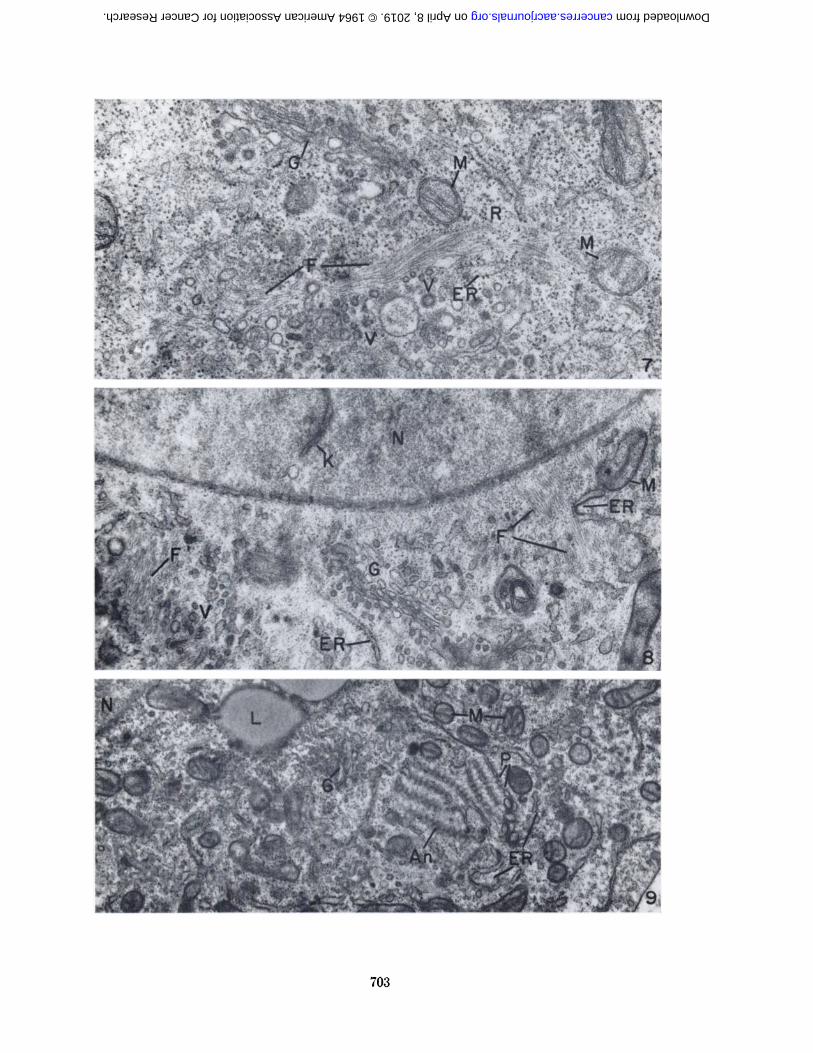
One of the outstanding features of the Sarcoma I cellswas the abundance of ribosomes in the cytoplasm. Manyof these ribonucleoprotein particles occurred free and wereusually arranged in small clusters (Fig. 6). Others wereattached to membranes of the endoplasmic reticulum.Although the section presented in Figure 6 contains considerable endoplasmic reticulum, this cytoplasmic component was often rather sparse (Fig. 3).
The Golgi complex was prominent in some sections(Figs. 6-8). It consisted of parallel cisternae bound bysmooth membranes and associated with numerous, small,smooth-surfaced vesicles.
The mitochondria were seen in spherical, oval, and longprofiles with cristae extending from one side to the other,from end to end, or diagonally (Fig. 6).
The Sarcoma I cell often contained large, sphericalbodies which, in electron micrographs, were characterizedby a homogeneous appearance and low electron density(Fig. 6). They were frequently bound by a narrow marginof increased density which appeared beaded or granular.In l-p sections of epoxy-embedded cells stained withAzure II and méthylèneblue (20) these bodies were alight green color, which is the characteristic reaction forlipide-containing structures. In smears of whole cellsfixed with formalin or osmium vapor and stained withOil Red 0. they were bright red. The fine structure ofthese bodies, together with their staining properties, established that they were lipide. They were undoubtedlyidentical to the "refractile bodies or spherules" described
by Baker et al. in earlier light microscope studies (1).Lipide bodies were observed in association with mito
chondria, smooth-surfaced vesicles, endoplasmic reticulum, and ribosomes (Fig. 6). An electron micrographpresented in an earlier publication (6) showed a lipidebody in contact with the membranes of annulate lamellae.
Annulate lamellae were observed in a number of sarcomacells from both A/Jax and C57BL/6K mouse tumors(Fig. 9). They occurred in the region of the Golgi complex more frequently than in other positions in the cellas described in an earlier publication (6).
Bundles of fine filaments were observed frequently athigh magnification (Figs. 7, 8). They were seen in thecytoplasm near the nuclear membrane, in the region of theGolgi complex, associated with mitochondria, and amongribosomes and profiles of rough-surfaced endoplasmicreticulum.
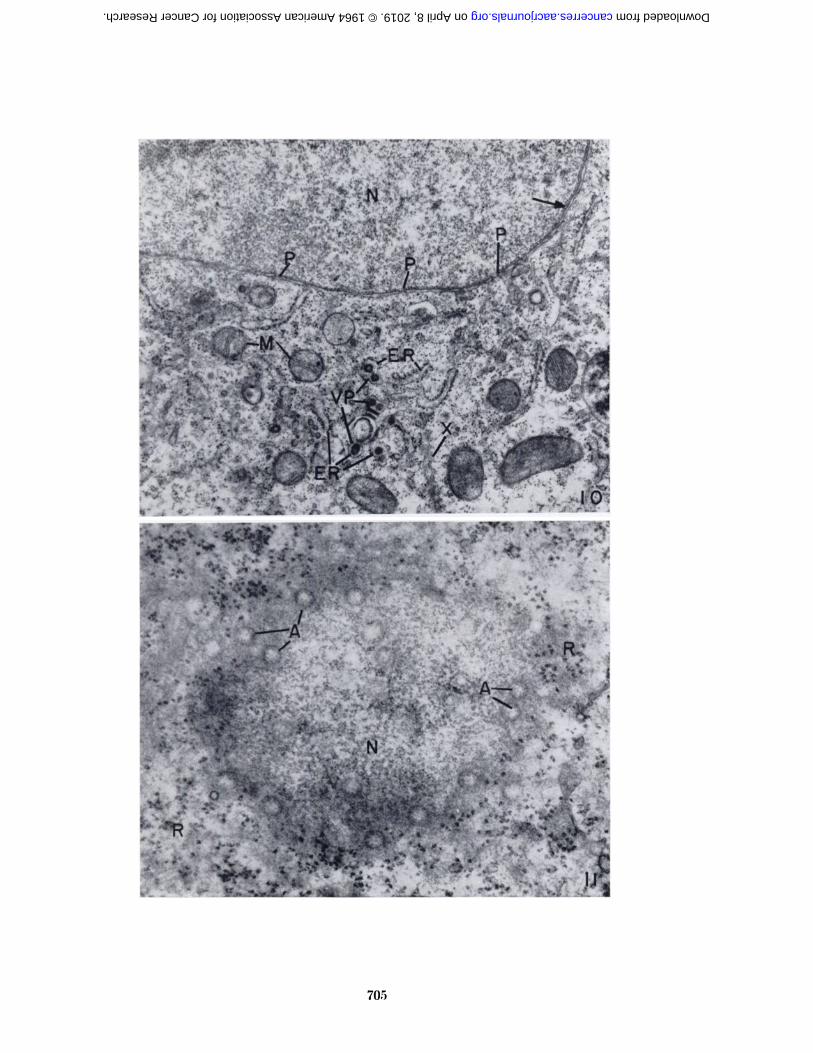
FIGS. 1-11.—Abbreviations:A = annulus of nuclear enve- M = mitochondria
lope Mv = microvilliAn = annulate lamellae N = nucleusER = endoplasmic reticulum Nu = nucleolusF = cytoplasmic filaments P = poreG = Golgi complex R = ribosomesI = invagination V = vesiclesK = membranous profile in nu- VP = virus-like particles
cleusL = lipide body
FIG. 1.—Electron micrograph of a thin section of Sarcoma Icells from a 6-day tumor in a C57BL/6K mouse. X 3000.
FIG. 2.-—Electron micrograph of sectioned Sarcoma I cellsfrom an 8-day tumor in an A/Jax mouse. X 3200.
on April 8, 2019. © 1964 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Ir
.
•>rf N
L
695
on April 8, 2019. © 1964 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

FIG. 3.—Sarcoma I cell from a 6-day tumor in a C57BL/6Kmouse. Microvilli (Mr) protrude from the cell surface. Thecytoplasm contains an abundance of free ribosomes (/Õ)and a smallamount of endoplasmic reticulum (Eli). Note the membranousprofile (A') in the nucleus (A7). X 12,000.
690
on April 8, 2019. © 1964 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
•**JVii>â„¢r"̂ *i- *y^TSA £***3. "
l•.i*- &
m
-?J
697
on April 8, 2019. © 1964 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

FIG. 4.—SarcomaI cell from an 8-day tumor in an A/Jax mouse.Note the similarity to the cell in Fig. 3. Two membranous profiles(K) are present in the nucleus (A'). Nuclear pores (P) occur at
frequent intervals along the nuclear envelope. X 11,000.
on April 8, 2019. © 1964 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
•%
v
•Õ-Mv—'
699
on April 8, 2019. © 1964 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

Fio. 5.—Sarcoma I cell from a 6-day tumor in a C57BL/GKmouse. Microvilli (Mv) protrude from the cell surface. Smooth-surfaced vesicles (V) are seen within the microvilli and at theirbase. Invaginations of the plasma membrane are also present(/). Ribosomes (R) are numerous. X 35,(KM).
FIG. 6.—Aportion of a healthy-appearing Sarcoma I cell froman 8-day tumor in a C57BL/(>K mouse. Lipide bodies (Lt , L2 ,and L3) are closely associated with mitochondria. All four lipidebodies have several small vesicles in contact with their surface.Lipide body (L¡)is in close proximity to ER. X 23,000.
700
on April 8, 2019. © 1964 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
10¿
9
on April 8, 2019. © 1964 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

'OOO'üIX'aB[|auiB|d}B[nuuBjoA")iui.>iAai(niisi(£))xa[duioai8[or)-(rf)saaodXqpd)dnjja)utaJBipiq.ttsoinu(|-mamjo.vvaiAuojiaas-ssojaB^uatojd}i{äuai{)^Batî[iaiuB[ailX"asnoui5l9/rIQ¿SOBu!A"i(i[Tîai( X[j(iojTîdcliî ui!u;(uy)
•OOO'OKX'snaianuaqjui(Y)a[y<udsnouT?j<nuaiua^osj'(Q)xa[tluio.>iSjor)amjoX)iui3iAamuipun'(Ã/^)tunjnaiÃajaitusB[dopua'(/V)"!Jpti"t|-">(uti'H!-tttioi)i!io<>ssBaso[auiado[aAua.nsapnuaq^o^sa]Sin!n)S|j}tÃp̄utìjBaujnaao(^)sjuauiB]ij'asnoiuxiÃf/vÃœBu;jtmin}Xcp-gÃœBluojj[jaa¡BUIOOJBB̂jouoi^jody—'8''Jlá
•OOO'SS X'(}¡3)»iii(n3i;aj3uusB|dopuapuB'(£))saiiBjqiuauii3¡OQ'(f\¡)Bup-uoqaojuuJBBUpuiÃ'(y)sauÃosoqu(^)sajaisaAjo^spuutu(¿f)s^uauiB|ijjoajptmqajojyj-asnomJiBuijoiunÃÃtBp-9Bmojj[¡aajBtuoajByBjouoi^jody—'¿'
on April 8, 2019. © 1964 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
/^'"'SS*:
IA1
A
/
aat-
on April 8, 2019. © 1964 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

toi
•QOO'ISX'-VV8IAUO-83BJuiusasai-esajodaqjSuipunojjns(y)i[nuuy'asnouiBuijouiri}XTjp-gUBuiojj[|9DJBiuoajB^Xi{}[Ba
auttjquiauiJiîa|anujouoi^aas[Bi^ua3uiî|y—'\\'(KK)'eSX'(,[)sajoclXqpa^dnjja^u;siadojaAua
(I)auBj<|uiauijBa[onujauniaqijoaotîjjnsaptsuiaqjimajBsaAiäaa^BjjB|HUBa3y'(.Y)T8uaass!
-uiaiu,ÎAB.\\ |iînsnunuy'(}I3)\un\nar\aiJimstîjdopuajoaBuaa|siauiqjiM(f/yl)saprjjBct35(i[-snaiA'adBqs-)nuqSnopSuiMoqsasnouiH9/ia¿SO«u!JouiniX«p-yBluojj[jaajBuioajBgy—'QI'OIJ
on April 8, 2019. © 1964 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
r-
:
on April 8, 2019. © 1964 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

CHAMBERSANDWEISER—SarcomaI in Homologous Host 707
Small, dense, doughnut-shaped bodies were occasionallyseen in electron micrographs of Sarcoma I cells from bothA/Jax and C57BL/6K mice (Fig. 10). These bodies wereapproximately 90 m/u in diameter and usually occurredwithin the cisternae of rough-surfaced endoplasmic reticu-lum. They resembled the type A virus-like bodiesdescribed by Dalton and co-workers (7, 8) and others(10, 27).
The nucleus was usually spherical and was frequentlylocated in a slightly eccentric position in the cell (Figs. 1,2). The nucleoplasm was granular and often contained amembranous profile which probably represents the invagination of the nuclear envelope. A suggestion of thisis seen in Figure 8. The nuclear envelope was composedof the usual two membranes, interrupted at irregular intervals by pores (Figs. 4 and 10). A section showing thecharacteristics of the pore wall or annulus described byWischnitzer (28) is shown in Figure 11. A layer of granular material was observed along the inner surface of thenuclear membrane (Fig. 10). The granules were approximately the same size as the cytoplasmic ribosomes, butappeared less dense. This layer gave the impression of avery thick membrane in low-magnification electron micrographs and in photomicrographs of stained sections.
Sections through nuceloli showed irregular masses ofcoarse, granular material of greater density than the surrounding nucleoplasm (Figs. 1, 2).
DISCUSSION
The electron microscope study of Sarcoma I cells duringthe growth of the tumor in isologous and homologous hostsshows certain aspects of fine structure which supplementthe observations made with the light microscope (1). Incomparing the cells from the two strains of mice duringtumor growth, no significant differences in fine structurewere noted.
Microvilli varying in size and number were commonlyfound distributed irregularly over the surface of the cells.These have been described in MCiM cells (2, 3), in Ehrlichascites cells (27), Walker tumor (15), and in normal cellsthat have grown in peritoneal fluid or in tissue culture (23).The small vesicles seen at the base or within the microvilliare apparently related to the uptake of material by pinocy-tosis or to the release of substances from the cell. Microvilli increase in number and in length during incubationin vitro, as described by Baker et al. (1).
The numerous ribosomes in Sarcoma I cells caused highelectron density of the cytoplasm in electron micrographsand a strong basophilia in smears and thick sections stainedwith basic dyes. Many of the ribonucleoprotein particlesoccurred free in the cytoplasm of Sarcoma I cells and wereusually arranged in clusters. Others were attached tomembranes of endoplasmic reticulum which was present inrelatively small amounts. Abundant free ribosomes andpoorly developed systems of endoplasmic reticulum arecommon features of many kinds of tumor cells (2, 4, 7, 11,13, 17, 18, 27). Howatson and Ham (11) suggested thatthese features may be related to cell growth, whereas organized endoplasmic reticulum may be related to special
ized cellular activities of differentiated cells. Birbeck andMercer (5) pointed out that free ribosomes are generallyassociated with protein synthesis in cells that retain theirprotein, and an abundance of ribosome-studded membranes is associated with protein synthesis in secretorycells. The occurrence of numerous free ribosomes in Sarcoma I cells is probably related to the rapid rate of growthand reproduction of these cells.
Lipide bodies were observed frequently in Sarcoma Icells. They increased in number, not only in the sarcomacells but also in the host peritoneal cells, as the diseaseprogressed in the A/Jax mouse and during tumor rejectionin the C57BL/6K mouse.
The accumulation of lipide deposits in cells is a widespread phenomenon. Among the cells in which largeamounts of lipide have been described are cells grown intissue culture (23), pancreas cells of starved guinea pigs(19), liver cells of rats during carbon tetrachloride intoxication (25), fibroblasts of skin wounds of scorbutic guineapigs (22), Sarcoma 37 cells (9), and MCiM cells (2, 3, 12).Palade (19) described a close association of lipide bodieswith mitochondria in the cells of the pancreas of starvedguinea pigs. He speculated that the lipide bodies represent fat that has been mobilized from adipose tissue andthat their association with mitochondria was for thepurpose of fatty acid oxidation by the oxidative enzymes ofthe mitochondria. Smuckler et al. (25) attributed theaccumulations of lipide in the liver cells of carbon tetra-chloride-intoxicated rats to the lack of protein with whichtriglycéridemay be coupled for transport from the cell.This theory was also invoked by Ross and Benditi (22)as the most plausible explanation for the development oflipide bodies in fibroblasts of healing wounds of scorbuticguinea pigs. Whether the development of lipide bodiesin Sarcoma I cells growing in either the susceptible A/Jaxmouse or the resistant C57BL/6K mouse is due to one ofthe above alterations in metabolism or to some other causeis not yet known.
The fine filaments in the cytoplasm of the Sarcoma Icells resemble those seen in MCiM sarcoma cells (2, 3, 12),in Sarcoma 37 cells (9), in fibroblasts (21), and the tono-filaments of epithelial cells (24, 26). The electron densityof the cytoplasmic matrix of Sarcoma I cells was partiallydue to this fine filamentous component.
REFERENCES
1. BAKER, P.; WEISER, R. S.; JUTILA, J.; EVANS, C. A.; ANDBLANDAU,R. J. Mechanisms of Tumor Homograft Rejection:The Behavior of Sarcoma I Ascites Tumor in the A/Jax andthe C57BL/6K Mouse. Ann. N. Y. Acad. Sci., 101:46-62, 1962.
2. BERGSTRAND,A., AND RINOERTZ, N. Electron MicroscopicExamination of the MCiM Tumor. I. The Tumor in AsciteaForm. J. Nati. Cancer Inst., 25:501-21, 1960.
3. . Electron Microscopic Examination of the MCiMTumor. II. A Comparison between the Sublines MAA, MASI,and MSB •Ibid., pp. 523-45.
4. BERNHARD,W. Electron Microscopy of Tumor Cells and TumorViruses: A Review. Cancer Res., 18:491-509, 1958.
5. BIHBECK, M. S. C., AND MERCER, E. H. Cytology of CellsWhich Synthesize Protein. Nature, 189:558-60, 1961.
6. CHAMBERS,V. C., ANDWEISER, R. S. Annulate Lamellae inSarcoma I Cells. J. Cell Biol. (in press).
on April 8, 2019. © 1964 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

708 Cancer Research Vol. 24, May 1964
7. DALTON,A. J., ANDFELIX, M. D. The Electron Microscopy ofNormal and Malignant Cells. Ann. N. Y. Acad. Sci., 63(6):1117-40, 1956.
8. DALTON,A. J.; POTTER, M.; ANDMERWIN, R. M. Some Ultra-structural Characteristics of a Series of Primary and Transplanted Plasma-Cell Tumors of the Mouse. J. Nati. CancerInst., 26:1221-68, 1961.
9. EPSTEIN, M. A. The Fine Structure of the Cells in Mouse Sarcoma 37 Ascitic Fluids. J. Biophys. Biochem. Cytol., 3:567-76,1957.
10. GRANBOULAN, N.; RIVIERE, M. R.; AND BERNHARD, W.Présencede particules d'aspect viral dans un sarcome creffa-
ble de la souris provoquépar le méthylcholanthrène. Bull.Assoc. Franc, étudeCancer, 47:291-307, 1960.
11. HOWATSON,A. F., ANDHAM, A. W. Electron Microscope Studyof Sections of Two Rat Liver Tumors. Cancer Res., 15: 62-69,1955.
12. JOURNEY, L. J., AND AMOS, D. B. An Electron MicroscopeStudy of Histiocyte Response to Ascites Tumor Homografts.Cancer Res., 22:998-1001,1962.
13. KUFF, E. L., ANDZEIGEL, R. F. Cytoplasmic Ribonucleopro-tein Components of the Novikoff Hepatoma. J. Biophys. Biochem. Cytol., 7:465-78,1960.
14. LUFT, J. H. Improvements in Epoxy Resin Embedding Methods. J. Biophys. Biochem. Cytol., 9:409-414, 1961.
15. MERCER, E. H., ANDEASTY, G. C. The Fine Cytology of theWalker Tumor. Cancer Res., 21:52-56, 1961.
16. MILLONIG,G. A Modified Procedure for Lead Staining of ThinSections. J. Biophys. Biochem. Cytol., 11:736-739, 1961.
17. NAGUMO, S. Electron Microscopic Observation of Méthylcholanthrène Induced Mouse Lymphomatosis. J. Electron-microscopy, 7:48-56, 1959.
18. NOVIKOFF,A. B. A Transplantable Rat Liver Tumor Inducedby 4-Dimethylaminoazobenzene. Cancer Res., 17:1010-27,1957.
19. PALADE, G. E. Functional Changes in the Structure of CellComponents, pp. 64-83. In Subcellular Particles. New York:The Ronald Press Co., 1959.
20. RICHARDSON,K. C.; JARETT, L.; ANDFINKE, E. H. Embeddingin Epoxy Resins for Ultrathin Sectioning in Electron Microscopy. Stain Technol., 35:313-23, 1960.
21. Ross, R., ANDBENDITT, E. P. Wound Healing and CollagenFormation. I. Sequential Changes in Components of GuineaPig Skin Wounds Observed in the Electron Microscope. J.Biophys. Biochem. Cytol., 11:677-700, 1961.
22. . Wound Healing and Collagen Formation. II. FineStructure in Experimental Scurvy. J. Cell Biol., 12:533-52,1962.
23. SELBY, C. C. Microscopy. II. Electron Microscopy: A Review.Cancer Res., 13:753-75, 1953.
24. . An Electron Microscope Study of Mammalian Skin inThin Sections. I. Dermo-Epidermal Junction and Basal CellLayer. J. Biophys. Biochem. Cytol., 1:429-44, 1955.
25. SMUCKLER,E. A.; ISEHI, O. A.; ANDBENDITT, E. P. Studies onCarbon Tetrachloride Intoxication. Biochem. Biophys. Res.Comm., 5:270-75,1961.
26. WEISS, P., AND FERRIS, W. Electronmicrograms of LarvalAmphibian Epidermis. Exp. Cell Res., 6:546-49, 1954.
27. WESSEL, W., AND BERNHARD,W. Vergleichende elektronenmikroskopische Untersuchung von Ehrlich- und Yoshida-Ascitestumorzellen. Z. Krebsforsch., 62:140-62, 1957.
28. WISCHNITZEH,S. An Electron Microscope Study of the Nuclear Envelope of Amphibian Oocytes. J. Ultrastruct. Res.,1:201-22, 1958.
on April 8, 2019. © 1964 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

1964;24:693-708. Cancer Res Velma C. Chambers and Russell S. Weiser Host. I. The Cells of the Growing TumorAn Electron Microscope Study of Sarcoma I in a Homologous
Updated version
http://cancerres.aacrjournals.org/content/24/4_Part_1/693
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/24/4_Part_1/693To request permission to re-use all or part of this article, use this link
on April 8, 2019. © 1964 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from





























