Embed Size (px)
Citation preview

The Plant Cell, Vol. 2, 1239-1248, December 1990 O 1990 American Society of Plant Physiologists
Analysis of Tomato Polygalacturonase Expression in Transgenic Tobacco
Katherine W. Osteryoung,’ Kurt Toenjes, Bradford Hall, Vickie Winkler, and Alan 6. Bennett2 Mann Laboratory, Department of Vegetable Crops, University of California, Davis, California 9561 6
Tomato polygalacturonase is a cell wall enzyme secreted in large amounts during tomato fruit ripening. Polygalac- turonase is synthesized as a glycoprotein precursor that undergoes numerous cotranslational and post-translational processing steps during its maturation, yielding three isozymes in tomato fruit, PG1, PGPA, and PGPB. To investigate the physiological roles of the three isozymes and the functional significance of the polygalacturonase processing domains in its intracellular transport and activity, we have examined polygalacturonase expression in transgenic tobacco plants. A full-length polygalacturonase cDNA was placed under control of the cauliflower mosaic virus 35s promoter and introduced into tobacco by way of Agrobacterium-mediated transformation. Analysis of transgenic tobacco plants indicated that (1) immunologically detectable polygalacturonase can be extracted from leaves, roots, and stems of transgenic tobacco plants; (2 ) only PGSA and PGPB were detectable in transgenic tobacco; (3) the polygalacturonase isozymes present in transgenic tobacco were electrophoretically indistinguishable from the tomato isozymes; (4) the N-terminal sequence, degree of N-linked glycosylation, and extent of oligosaccharide processing were similar in polygalacturonase from transgenic tobacco and tomato; (5) polygalacturonase was properly localized in cell walls of transgenic tissue; (6) the protein was enzymically active in vitro; however, (7) accumulation of PGPA and PGSB in cell walls of transgenic tobacco did not result in pectin degradation in vivo. These results indicated that tomato polygalacturonase was properly processed and transported to the cell wall of tobacco. However, accumulation of the two polygalacturonase isozymes expressed in this heterologous host was insufficient to promote polyuronide degradation in tobacco leaf tissue.
INTRODUCTION
Tomato fruit ripening is the result of a complex devel- opmental program affecting metabolic processes in every compartment of the cell. A major component of this de- velopmental program is the degradation of pectic polymers in the cell wall, which is brought about by the action of a ripening-induced enzyme, polygalacturonase (Themmen et al., 1982; Crookes and Grierson, 1983; Huber, 1983a; Giovannoni et al., 1989). Polygalacturonase accumulates to high levels in ripening tomato fruit, suggesting an im- portant role for the enzyme in the developing fruit. Although a correlation between polygalacturonase accumulation and tomato fruit softening has been documented (Huber, 1983b, and references therein), recent evidence indicates that, by itself, degradation of polyuronides by polygalac- turonase is insufficient to cause tomato fruit softening (Giovannoni et al., 1989). Thus, the physiological function of this enzyme remains unclear.
Tomato fruit polygalacturonase comprises three iso- zymes: PGI , PGPA, and PG2B (Pressey and Avants, 1973;
’ Current address: Department of Biochemistry, Life Sciences South, Room 303, University of Arizona, Tucson, AZ 85721. * To whom correspondence should be addressed.
Tucker et al., 1980; Ali and Brady, 1982). PG2A and PG2B appear to function as monomeric catalytic subunits of 45 kD and 46 kD, respectively, with the size difference arising from a difference in extent of glycosylation (DellaPenna and Bennett, 1988). PG1 is a more complex protein of approximately 1 O0 kD consisting of a single catalytic poly- galacturonase subunit (either PG2A or PGPB) in associa- tion with a second glycoprotein subunit of 41 kD (Moshrefi and Luh, 1983). This subunit is immunologically unrelated to PG2A and PG2B (T. Moore and A.B. Bennett, unpub- lished results) and may be the polygalacturonase converter that converts the PG2 isozymes to PGI in vitro (Tucker et al., 1981; Pressey, 1984).
Most work on polygalacturonase has focused on char- acterizing the PGPA and PG2B catalytic subunits, which function both as monomers and as subunits of PG1. The biosynthesis and maturation of these monomers are them- selves complex. Both PG2A and PG2B appear to be the products of a single gene and are synthesized as a larger molecular weight precursor that undergoes a number of processing events during conversion to the mature protein. These include cotranslational removal of the hydrophobic signal sequence and asparagine-linked glycosylation in the

1240 The Plant Cell
JPTransgenic Tobacco
2B-,2A-
1Figure 1. Immunodetection of Polygalacturonase Protein in CellWall Extracts from Leaves of Transgenic Tobacco.
Cell wall proteins were extracted as described in Methods, sepa-rated by SDS-PAGE, electroblotted to nitrocellulose, and probedwith polygalacturonase antiserum. Lane 1, 0.5 /ig of purifiedtomato PG2A and PG2B; lane 2, 50 ng of cell wall protein from acontrol plant transformed with vector alone; lanes 3 to 7, 50 ^gof cell wall protein from five different transgenic tobacco plants.The sizes of PG2A and PG2B are 45 kD and 46 kD, respectively.
similar analysis in transgenic rin tomato fruit expressing achimeric polygalacturonase gene suggests that PG1 alonecatalyzes polyuronide degradation in vivo (DellaPenna etal., 1990).
To address the functional significance of processingevents that contribute to the maturation, intracellular trans-port, and physiological function of the polygalacturonaseisozymes, we have characterized expression of the cata-lytic polygalacturonase subunits in transgenic tobacco.The results of these experiments indicated that the proteinprocessing apparatus, which functions in the maturationof tomato fruit polygalacturonase, is conserved in vegeta-tive tissue of tobacco. In addition, these results haveprovided an opportunity to assess the function of the PG2isozymes in the absence of PG1.
RESULTS
endoplasmic reticulum (DellaPenna and Bennett, 1988), aswell as extensive post-translational modification of the coreoligosaccharides in the Golgi (Moshrefi and Luh, 1983).Recent evidence suggests that the differences in glyco-sylation between PG2A and PG2B result from differencesin cotranslational core glycosylation rather than from dif-ferential processing of the glycan side chains (DellaPennaand Bennett, 1988). Comparison of the amino acid se-quence of mature PG2A and PG2B with the amino acidsequence deduced from cDNA clones further indicates atleast two additional proteolytic processing steps, one thatremoves 47 amino acids following the signal sequence(DellaPenna and Bennett, 1988), and another that removes13 amino acids from the C terminus of the precursorprotein (Sheehy et al., 1987). Both of these proteolyticevents are presumed to occur post-translationally, al-though the subcellular sites of these cleavages andwhether they occur in one or in multiple steps are notknown.
Although all three polygalacturonase isozymes are ac-tive when assayed in vitro (Pressey and Avants, 1973;Tucker et al., 1980; Ali and Brady, 1982), their distincttemporal patterns of accumulation in ripening tomato fruithave led to some controversy regarding the function ofeach isozyme in vivo. Pressey (1988) has suggested thatPG1 is an artifact of extraction, PG2A and PG2B being theonly isozymes present in vivo, whereas Knegt et al. (1988)have presented data contrary to this view and proposedthat PG1 is the physiologically functional isozyme in vivo.Recent analysis of polygalacturonase isozyme accumula-tion in transgenic tomato fruit exhibiting reduced levels ofpolygalacturonase has led to the proposal that the PG1and PG2 isozymes are responsible for distinct aspects ofpolyuronide degradation (Smith et al., 1990), whereas a
Synthesis, Processing, and Isozyme Composition ofPolygalacturonase in Transgenic Tobacco
A full-length tomato fruit polygalacturonase cDNA(DellaPenna and Bennett, 1988) was fused to the cauli-flower mosaic virus 35S promoter and NOS 3' terminatorin the binary vector pMON530 and introduced into tobaccoby way of •Agrotoacter/i/m-mediated transformation. Trans-formants selected for kanamycin resistance and nopalinesynthase activity were assayed for the presence of poly-galacturonase protein by immunoblotting with polygalac-turonase antiserum. Figure 1 shows an immunoblot ofcrude cell wall extracts from leaves of five different trans-genic tobacco plants and one control plant transformedwith vector only. Levels of immunologically detectablepolygalacturonase varied among transformants, as shownby the varying band intensities in Figure 1. Polygalacturon-ase was also detected in cell wall extracts from stems androots of transgenic tobacco plants, but at somewhat lowerlevels than in leaves (data not shown).
The immunoblot in Figure 1 also shows that two immu-noreactive polypeptides corresponding to purified tomatofruit PG2A and PG2B accumulated in transgenic tobaccoleaves. This was true for all transgenic plants that hadimmunologically detectable levels of polygalacturonase.These results clearly demonstrated that PG2A and PG2Brepresent differentially processed forms of a single geneproduct because a single cDNA was transferred into to-bacco. In addition, the electrophoretic mobilities of thesepolypeptides on SDS gels (Figure 1) and native gels (seebelow) are identical to the mobilities of authentic tomatofruit PG2A and PG2B, suggesting that the cotranslationaland post-translational processing of polygalacturonase intransgenic tobacco closely parallel the normal processingin ripening tomato fruit.
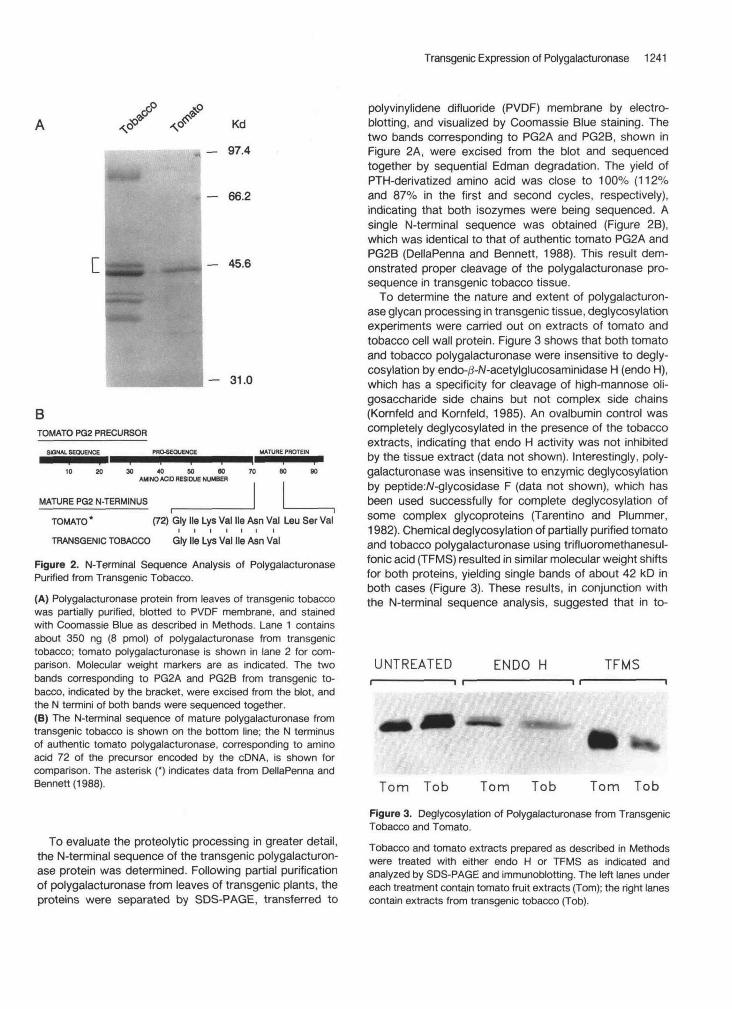
Transgenic Expression of Polygalacturonase 1241
Kd
.. — 97.4
— 66.2
_ — 45.6
— 31.0
BTOMATO PG2 PRECURSOR
SIGNAL SEQUENCE ________PUP-SEQUENCE MATURE PROTEIN
30 40 50 60 70AMINO ACID RESIDUE NUMBER
MATURE PG2 N-TERMINUS
TOMATO * (72) Gly lie Lys Val He Asn Val Leu Ser ValI I I I I I I
TRANSGENIC TOBACCO Gly He Lys Val lie Asn Val
Figure 2. N-Terminal Sequence Analysis of PolygalacturonasePurified from Transgenic Tobacco.
(A) Polygalacturonase protein from leaves of transgenic tobaccowas partially purified, blotted to PVDF membrane, and stainedwith Coomassie Blue as described in Methods. Lane 1 containsabout 350 ng (8 pmol) of polygalacturonase from transgenictobacco; tomato polygalacturonase is shown in lane 2 for com-parison. Molecular weight markers are as indicated. The twobands corresponding to PG2A and PG2B from transgenic to-bacco, indicated by the bracket, were excised from the blot, andthe N termini of both bands were sequenced together.(B) The N-terminal sequence of mature polygalacturonase fromtransgenic tobacco is shown on the bottom line; the N terminusof authentic tomato polygalacturonase, corresponding to aminoacid 72 of the precursor encoded by the cDNA, is shown forcomparison. The asterisk (*) indicates data from DellaPenna andBennett (1988).
To evaluate the proteolytic processing in greater detail,the N-terminal sequence of the transgenic polygalacturon-ase protein was determined. Following partial purificationof polygalacturonase from leaves of transgenic plants, theproteins were separated by SDS-PAGE, transferred to
polyvinylidene difluoride (PVDF) membrane by electro-blotting, and visualized by Coomassie Blue staining. Thetwo bands corresponding to PG2A and PG2B, shown inFigure 2A, were excised from the blot and sequencedtogether by sequential Edman degradation. The yield ofPTH-derivatized amino acid was close to 100% (112%and 87% in the first and second cycles, respectively),indicating that both isozymes were being sequenced. Asingle N-terminal sequence was obtained (Figure 2B),which was identical to that of authentic tomato PG2A andPG2B (DellaPenna and Bennett, 1988). This result dem-onstrated proper cleavage of the polygalacturonase pro-sequence in transgenic tobacco tissue.
To determine the nature and extent of polygalacturon-ase glycan processing in transgenic tissue, deglycosylationexperiments were carried out on extracts of tomato andtobacco cell wall protein. Figure 3 shows that both tomatoand tobacco polygalacturonase were insensitive to degly-cosylation by endo-/3-A/-acetylglucosaminidase H (endo H),which has a specificity for cleavage of high-mannose oli-gosaccharide side chains but not complex side chains(Kornfeld and Kornfeld, 1985). An ovalbumin control wascompletely deglycosylated in the presence of the tobaccoextracts, indicating that endo H activity was not inhibitedby the tissue extract (data not shown). Interestingly, poly-galacturonase was insensitive to enzymic deglycosylationby peptide:A/-glycosidase F (data not shown), which hasbeen used successfully for complete deglycosylation ofsome complex glycoproteins (Tarentino and Plummer,1982). Chemical deglycosylation of partially purified tomatoand tobacco polygalacturonase using trifluoromethanesul-fonic acid (TFMS) resulted in similar molecular weight shiftsfor both proteins, yielding single bands of about 42 kD inboth cases (Figure 3). These results, in conjunction withthe N-terminal sequence analysis, suggested that in to-
UNTREATED ENDO H TFMS
Tom Tob Tom Tob Tom Tob
Figure 3. Deglycosylation of Polygalacturonase from TransgenicTobacco and Tomato.
Tobacco and tomato extracts prepared as described in Methodswere treated with either endo H or TFMS as indicated andanalyzed by SDS-PAGE and immunoblotting. The left lanes undereach treatment contain tomato fruit extracts (Tom); the right lanescontain extracts from transgenic tobacco (Tob).
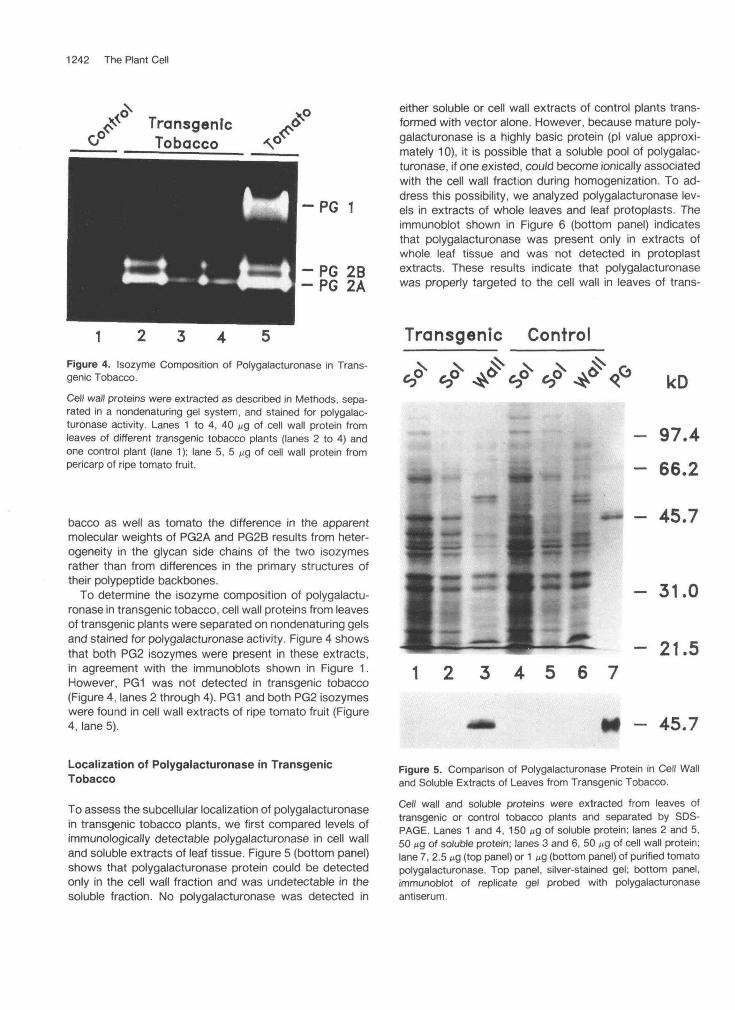
1242 The Plant Cell
TransgenlcTobacco
-PG 1
-PG 2B-PG 2A
Figure 4. Isozyme Composition of Polygalacturonase in Trans-genic Tobacco.
Cell wall proteins were extracted as described in Methods, sepa-rated in a nondenaturing gel system, and stained for polygalac-turonase activity. Lanes 1 to 4, 40 ^9 of cell wall protein fromleaves of different transgenic tobacco plants (lanes 2 to 4) andone control plant (lane 1); lane 5, 5 ng of cell wall protein frompericarp of ripe tomato fruit.
bacco as well as tomato the difference in the apparentmolecular weights of PG2A and PG2B results from heter-ogeneity in the glycan side chains of the two isozymesrather than from differences in the primary structures oftheir polypeptide backbones.
To determine the isozyme composition of polygalactu-ronase in transgenic tobacco, cell wall proteins from leavesof transgenic plants were separated on nondenaturing gelsand stained for polygalacturonase activity. Figure 4 showsthat both PG2 isozymes were present in these extracts,in agreement with the immunoblots shown in Figure 1.However, PG1 was not detected in transgenic tobacco(Figure 4, lanes 2 through 4). PG1 and both PG2 isozymeswere found in cell wall extracts of ripe tomato fruit (Figure4, lane 5).
either soluble or cell wall extracts of control plants trans-formed with vector alone. However, because mature poly-galacturonase is a highly basic protein (pi value approxi-mately 10), it is possible that a soluble pool of polygalac-turonase, if one existed, could become ionically associatedwith the cell wall fraction during homogenization. To ad-dress this possibility, we analyzed polygalacturonase lev-els in extracts of whole leaves and leaf protoplasts. Theimmunoblot shown in Figure 6 (bottom panel) indicatesthat polygalacturonase was present only in extracts ofwhole leaf tissue and was not detected in protoplastextracts. These results indicate that polygalacturonasewas properly targeted to the cell wall in leaves of trans-
Transgenic Control
?<?<? <T kD
- 97.4- 66.2
— - 45.7
- 31.0
1 2 3 4 5 6 7- 21.5
- 45.7
Localization of Polygalacturonase in TransgenicTobacco
To assess the subcellular localization of polygalacturonasein transgenic tobacco plants, we first compared levels ofimmunologically detectable polygalacturonase in cell walland soluble extracts of leaf tissue. Figure 5 (bottom panel)shows that polygalacturonase protein could be detectedonly in the cell wall fraction and was undetectable in thesoluble fraction. No polygalacturonase was detected in
Figure 5. Comparison of Polygalacturonase Protein in Cell Walland Soluble Extracts of Leaves from Transgenic Tobacco.
Cell wall and soluble proteins were extracted from leaves oftransgenic or control tobacco plants and separated by SDS-PAGE. Lanes 1 and 4, 150 M9 of soluble protein; lanes 2 and 5,50 M9 of soluble protein; lanes 3 and 6, 50 M9 of cell wall protein;lane 7, 2.5 ̂ 9 (top panel) or 1 ng (bottom panel) of purified tomatopolygalacturonase. Top panel, silver-stained gel; bottom panel,immunoblot of replicate gel probed with polygalacturonaseantiserum.
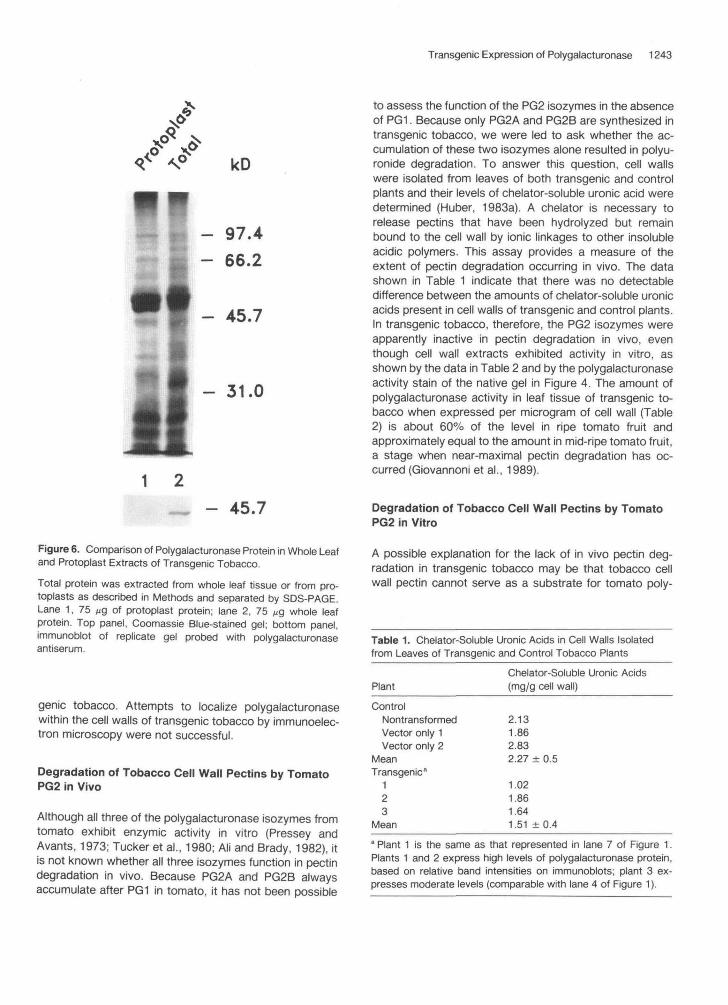
Transgenic Expression of Polygalacturonase 1243
kD
- 31.0
to assess the function of the PG2 isozymes in the absenceof PG1. Because only PG2A and PG2B are synthesized intransgenic tobacco, we were led to ask whether the ac-cumulation of these two isozymes alone resulted in polyu-ronide degradation. To answer this question, cell wallswere isolated from leaves of both transgenic and controlplants and their levels of chelator-soluble uronic acid weredetermined (Huber, 1983a). A chelator is necessary torelease pectins that have been hydrolyzed but remainbound to the cell wall by ionic linkages to other insolubleacidic polymers. This assay provides a measure of theextent of pectin degradation occurring in vivo. The datashown in Table 1 indicate that there was no detectabledifference between the amounts of chelator-soluble uronicacids present in cell walls of transgenic and control plants.In transgenic tobacco, therefore, the PG2 isozymes wereapparently inactive in pectin degradation in vivo, eventhough cell wall extracts exhibited activity in vitro, asshown by the data in Table 2 and by the polygalacturonaseactivity stain of the native gel in Figure 4. The amount ofpolygalacturonase activity in leaf tissue of transgenic to-bacco when expressed per microgram of cell wall (Table2) is about 60% of the level in ripe tomato fruit andapproximately equal to the amount in mid-ripe tomato fruit,a stage when near-maximal pectin degradation has oc-curred (Giovannoni et al., 1989).
— - 45.7
Figure 6. Comparison of Polygalacturonase Protein in Whole Leafand Protoplast Extracts of Transgenic Tobacco.
Total protein was extracted from whole leaf tissue or from pro-toplasts as described in Methods and separated by SDS-PAGE.Lane 1, 75 ^g of protoplast protein; lane 2, 75 //g whole leafprotein. Top panel, Coomassie Blue-stained gel; bottom panel,immunoblot of replicate gel probed with polygalacturonaseantiserum.
genie tobacco. Attempts to localize polygalacturonasewithin the cell walls of transgenic tobacco by immunoelec-tron microscopy were not successful.
Degradation of Tobacco Cell Wall Pectins by TomatoPG2 in Vivo
Although all three of the polygalacturonase isozymes fromtomato exhibit enzymic activity in vitro (Pressey andAvants, 1973; Tucker et al., 1980; Ali and Brady, 1982), itis not known whether all three isozymes function in pectindegradation in vivo. Because PG2A and PG2B alwaysaccumulate after PG1 in tomato, it has not been possible
Degradation of Tobacco Cell Wall Pectins by TomatoPG2 in Vitro
A possible explanation for the lack of in vivo pectin deg-radation in transgenic tobacco may be that tobacco cellwall pectin cannot serve as a substrate for tomato poly-
Table 1. Chelator-Soluble Uronic Acids in Cell Walls Isolatedfrom Leaves of Transgenic and Control Tobacco Plants
PlantChelator-Soluble Uronic Acids(mg/g cell wall)
ControlNontransformedVector only 1Vector only 2
MeanTransgenic8
123
Mean
2.131.862.832.27
1.021.861.641.51
±0.5
± 0.4
* Plant 1 is the same as that represented in lane 7 of Figure 1.Plants 1 and 2 express high levels of polygalacturonase protein,based on relative band intensities on immunoblots; plant 3 ex-presses moderate levels (comparable with lane 4 of Figure 1).
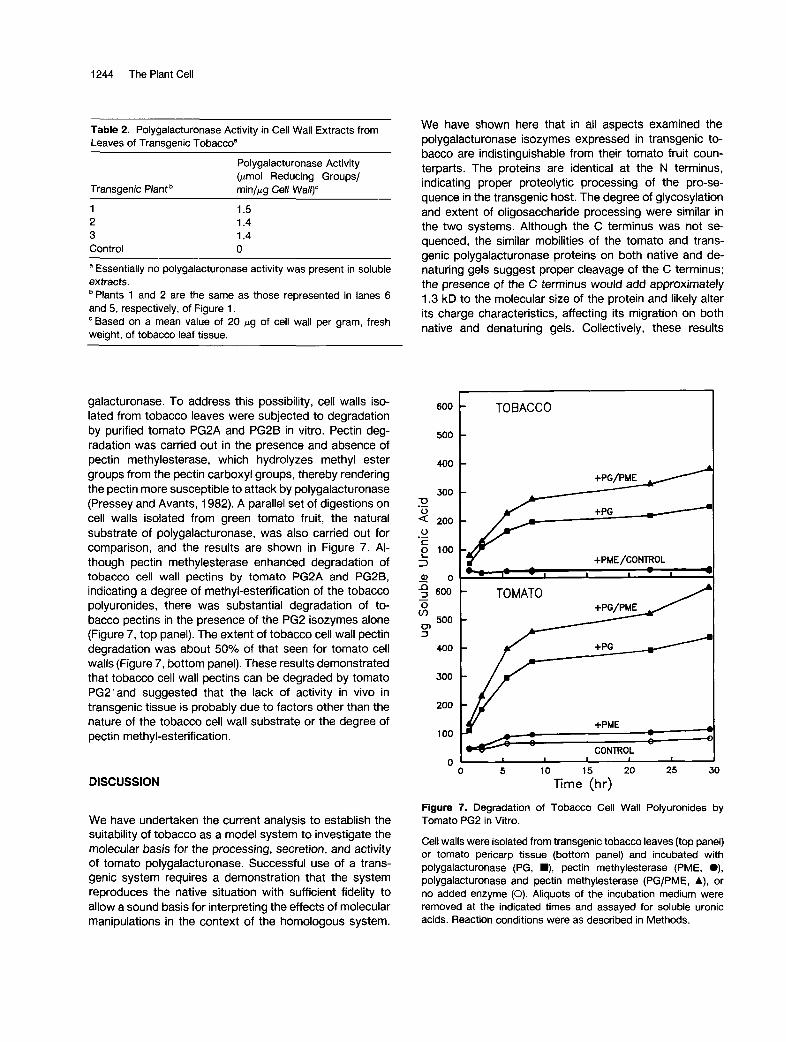
1244 The Plant Cell
Table 2. Polygalacturonase Activity in Cell Wall Extracts from Leaves of Transaenic Tobacco"
Polygalacturonase Activity (pmol Reducing Groups/
Transgenic Plantb min/pg Cell Wally
1 2 3 Control
1.5 1.4 1.4 O
a Essentially no polygalacturonase activity was present in soluble extracts.
Plants 1 and 2 are the same as those represented in lanes 6 and 5, respectively, of Figure 1. "Based on a mean value of 20 pg of cell wall per gram, fresh weight, of tobacco leaf tissue.
galacturonase. To address this possibility, cell walls iso- lated from tobacco leaves were subjected to degradation by purified tomato PG2A and PGPB in vitro. Pectin deg- radation was carried out in the presence and absence of pectin methylesterase, which hydrolyzes methyl ester groups from the pectin carboxyl groups, thereby rendering the pectin more susceptible to attack by polygalacturonase (Pressey and Avants, 1982). A parallel set of digestions on cell walls isolated from green tomato fruit, the natural substrate of polygalacturonase, was also carried out for comparison, and the results are shown in Figure 7. Al- though pectin methylesterase enhanced degradation of tobacco cell wall pectins by tomato PG2A and PG2B, indicating a degree of methyl-esterification of the tobacco polyuronides, there was substantial degradation of to- bacco pectins in the presence of the PG2 isozymes alone (Figure 7 , top panel). The extent of tobacco cell wall pectin degradation was about 50% of that seen for tomato cell walls (Figure 7, bottom panel). These results demonstrated that tobacco cell wall pectins can be degraded by tomato PG2'and suggested that the lack of activity in vivo in transgenic tissue is probably due to factors other than the nature of the tobacco cell wall substrate or the degree of pectin methyl-esterification.
DISCUSSION
We have undertaken the current analysis to establish the suitability of tobacco as a model system to investigate the molecular basis for the processing, secretion, and activity of tomato polygalacturonase. Successful use of a trans- genic system requires a demonstration that the system reproduces the native situation with sufficient fidelity to allow a sound basis for interpreting the effects of molecular manipulations in the context of the homologous system.
We have shown here that in all aspects examined the polygalacturonase isozymes expressed in transgenic to- bacco are indistinguishable from their tomato fruit coun- terparts. The proteins are identical at the N terminus, indicating proper proteolytic processing of the pro-se- quence in the transgenic host. The degree of glycosylation and extent of oligosaccharide processing were similar in the two systems. Although the C terminus was not se- quenced, the similar mobilities of the tomato and trans- genic polygalacturonase proteins on both native and de- naturing gels suggest proper cleavage of the C terminus; the presence of the C terminus would add approximately 1.3 kD to the molecular size of the protein and likely alter its charge characteristics, affecting its migration on both native and denaturing gels. Collectively, these results
600
500
400
300 73 .- 2 200 o c
3 - 0 0 5 600 O v)
500 0 3
.- 2 100
-
400
300
200
1 O0
O
TO BACCO
+PME/CONlROL - - a
Y I- - 1 I 1 - 1 I
TOMATO / +PG/PME
E Y +PME - -
" - I -
CONlROL
O 5 10 15 20 25 30
Time (hr)
Figure 7. Degradation of Tobacco Cell Wall Polyuronides by Tomato PG2 in Vitro.
Cell walls were isolated from transgenic tobacco leaves (top panel) or tomato pericarp tissue (bottom panel) and incubated with polygalacturonase (PG, m, pectin methylesterase (PME, O), polygalacturonase and pectin methylesterase (PG/PME, A), or no added enzyme (O). Aliquots of the incubation medium were removed at the indicated times and assayed for soluble uronic acids. Reaction conditions were as described in Methods.

Transgenic Expression of Polygalacturonase 1245
strongly suggest that the protein processing apparatus in tobacco faithfully reproduces all of the processing events that normally occur for PG2A and PG2B in tomato fruit. Although other plant proteins have been expressed and shown to be correctly processed in transgenic tobacco (Hoffman et al., 1987; Sturm et al., 1988; Sonnewald et al., 1989; Wilkins et al., 1990), expression of heterologous proteins in this system sometimes results in accumulation of molecular forms not normally observed in the native tissue (Higgins et al., 1988; Post-Beitenmiller et al., 1989; Altabella and Chrispeels, 1990). It is, therefore, significant that polygalacturonase, whose complex maturation proc- ess includes core glycosylation, oligosaccharide modi- fication, and at least three proteolytic processing steps, appears to undergo identical processing events in the transgenic host, including the heterogeneous reac- tions associated with maturation of the PG2A and PG2B isozymes.
Only two of the three polygalacturonase isozymes nor- mally present in tomato fruit were synthesized in trans- genic tobacco; PG1 was not detected. The most likely explanation for this is that the 41-kD subunit required for assembly of PG1 is not encoded by the tobacco genome or is not expressed in vegetative tissue. Unfortunately, this subunit has neither been purified nor its cDNA cloned, so the molecular probes necessary to examine its expression are not yet available.
Upon undertaking this study, we were concerned that constitutive expression of a cell wall hydrolase normally present only during a senescent stage of the tomato life cycle might preclude the regeneration of transformed plants. Nevertheless, transgenic plants expressing high levels of polygalacturonase in leaves, stems, and roots regenerated readily, grew normally, flowered, and set via- ble seed. In addition, ultrastructural observations revealed no obvious differences in cell wall structure between trans- genic and control plants (results not shown), as is observed as a consequence of polygalacturonase action in tomato fruit (Craokes and Grierson, 1983). These observations. in
of polygalacturonase in ripening tomato fruit, as has been proposed previously (Knegt et al., 1988). The results pre- sented here showing that accumulation of PG2A and PG2B in transgenic tobacco is insufficient to promote detectable pectin degradation in vivo are consistent with this hypoth- esis. When the clone for the 41-kD subunit of PGl be- comes available, this hypothesis might be tested by ex- pressing this subunit in these same transgenic tobacco plants and assaying for PG1 accumulation and pectin degradation.
Although the absence of PG1 is an intriguing explanation for the lack of pectin degradation in the leaves of trans- genic tobacco, there are other possible explanations. One possibility is that the biochemical milieu of the tobacco cell wall is incompatible with polygalacturonase activity. For instance, high calcium levels, if present in the cell wall, would inhibit polygalacturonase activity (Buescher and Hobson, 1982). Another possibility is that although the enzyme appears to be secreted to the cell wall, it may not be accessible to its polyuronide substrate because of improper targeting within the cell wall. It is also conceivable that the native structure of the tobacco cell wall is unsuited to degradation by the tomato enzyme in vivo, even though our results show that isolated tobacco cell walls are an acceptable substrate for purified tomato PG2 in vitro.
We have shown that the fidelity of polygalacturonase transpor? and processing is conserved in transgenic to- bacco. This heterologous expression system thus provides a means for examining the functions of the various proc- essing steps associated with maturation of this complex protein. In addition, this transgenic host may allow a critical test of the hypothesis that PG1 is required to catalyze polyuronide degradation in vivo.
METHODS
Plant Transformation and Growth
conjunction with the absence of PG1, led us to investigate the possibility that the PG2 isozymes expressed in the transgenic wereJ not causing the degradation Of pectins in vivo^ even though active enzyme could be extracted.
In ripening tomato fruit, PG1 accumulates before PG2 (Tucker et al., 1980; Brady et al., 1983). Pectin degradation commences with the appearance of PG1, and continues after the accumulation of PG2 has begun (Pressey, 1986). However, in a recent experiment in our laboratory that
ase in transgenic rin tomato fruit (Giovannoni et al., 1989),
The full-length polygalacturonase cDNA (DellaPenna and Bennett, 1988) was subcloned into the Smal site of the binary plant expression vector pMON530 (Shah et al., 1986). The resulting plasmid was used to transfect the Agrobacterium tumefaciens strain GV3111 SE harboring the disarmed Ti plasmid pTiB6S3SE (Rogers et ai., 1986). Leaf pieces of axenically grown Nicotiana tabacum cv Xanthi and A. tumefaciens were cocultivated at 28% on nurse culture plates essentially by the method of Rogers et al. (1986). Putative transfOr"nts were regenerated on 100 mg/L kanamycin sulfate (Boehringer Mannheim) and 300 mg/L carben-
medium (GIBCO) containing Nitsch's vitamins (Nitsch, 1969), 1 mg/L benzyladenine, and 0.1 mg/L naphthalene acetic acid.
shoots in 1 O0 mg/L kanamycin in the same medium but without hormones, and also by assaying rooted shoots for the presence of nopaline (Otten and Schilperoort, 1978). Kanamycin-resistant, nopaline-positive transformants were maintained axenically in 1-
resulted in a more gradual accumulation of polygalacturon- icillin (GeoPen) in a Murashige and Skoog (1962) complete agar
it was noted that pectin degradation coiricided with the Transformants were fufiher s&cted by rooting regenerated Of PG1 and that no further degradation
curred after PG2A and PG2B b%an to accumulate (DellaPenna et al., 1990). These reSultS suggested the possibility that PG1 is the only physiologically active form

1246 The Plant Cell
quart canning jars on the same medium containing 1 mg/L IAA and 0.15 mg/L kinetin without antibiotics. Except during coculti- vation, all plant material was grown at 25°C with a 16-hr day length at a light intensity of 70 pmol m-' sec-'. Transgenic plants transformed with the empty pMON530 vector were also gener- ated and are the controls referred to in the text and figures.
Protein Extraction and Polygalacturonase Assays
Protein was extracted from tobacco tissue essentially as previ- ously described for tomato fruit tissue (DellaPenna et al., 1987). Briefly, leaf tissue was homogenized on ice in 1 volume to 3 volumes of 20 mM Tris, 5 mM 2-mercaptoethanol, pH 7, or 20 mM Mes, 3 mM DTT, pH 5.5 (low-salt buffers). Homogenization at pH 5.5 yielded polygalacturonase protein of higher activity and was used for enzyme activity data shown in Figure 4 and Table 2. The homogenate was centrifuged at 15,0009 for 15 min at 4OC. The cell debris pellet was resuspended for 1 hr at O°C in 1.7 M NaCI, 15 mM EDTA, 50 mM sodium citrate, pH 5.5 (high-salt buffer), with 3 mM DTT added for data shown in Figure 4 and Table 2. The slurry was centrifuged at 15,0009 for 20 min at 4OC. The supernatants from both centrifugations were brought to 80% saturation with ammonium sulfate. The precipitates were collected by centrifugation as above, dissolved in 100 mM NaCI, 10 mM sodium acetate, 1 mM DTT, pH 6.0 (pH 5.0 for Table 2 and Figure 4), and dialyzed against the same buffer. The ammonium sulfate precipitates from the low-salt and high-salt supernatants are the soluble and cell wall fractions, respectively, referred to in the text and figure legends.
For the data shown in Figure 6, total (soluble and cell wall) protein was extracted by homogenizing protoplasts or whole leaves in high-salt buffer to release soluble and cell wall proteins into the supernatant. The cell debris was pelleted and discarded and the supernatants were treated as above.
Protein was determined by the method of Bradford (1976) or Smith et al. (1985). Polygalacturonase activity was measured as in DellaPenna et al. (1987).
N-Terminal Sequence Analysis
Polygalacturonase was partially purified from leaves of a single transgenic plant essentially as described previously for tomato (DellaPenna et al., 1986). The ammonium sulfate precipitate ob- tained as described above was resuspended in 0.1 M NaCI, 20 mM Mes, pH 6.0, dialyzed against the same buffer, concentrated by ultrafiltration (Amicon Corp.), filtered on a 0.22-pm filter (Milli- pore Corp.), and chromatographed on a 1-mL Mono S cation- exchange column (Pharmacia LKB Biotechnology Inc.). Proteins were eluted over 20 min using a linear gradient of 0.1 M to 0.8 M NaCl in 20 mM Mes, pH 6.0, at a flow rate of 1 mL/min. Fractions of 0.5 mL were collected and those containing immunologically reactive polygalacturonase protein were pooled, dialyzed against a buffer containing 0.1 M sodium acetate, 1 M NaCI, 1 mM MgCI,, 1 mM CaCI2, 1 mM MnCI,, and subjected to concanavalin A- Sepharose chromatography (Sigma) as described previously (DellaPenna et al., 1986). The eluant was dialyzed against 20 mM Tris, pH 7.8,l O0 mM NaCI, concentrated by ultrafiltration, applied to the Mono S column, and eluted over a 30-min period using a
linear gradient of 0.1 M to 0.8 M NaCl in 20 mM Tris, pH 7.8, at a flow rate of 1 mL/min. Fractions containing polygalacturonase were pooled and concentrated to about 50 pL using a Centricon filter.
A portion of the purified sample containing an estimated 8 pmol of polygalacturonase protein was separated on a 0.5-mm 10% SDS-polyacrylamide gel (Laemmli, 1970) allowed to age overnight before loading. Sodium thioglycolic acid (0.1 mM, Sigma) was included in the upper buffer reservoir and sample loading buffer (Hunkapiller et al., 1981). The proteins were transferred to Im- mobilon-P PVDF membrane (Millipore) by electroblotting in 1 O mM 3-(cyclohexylamino)-l -propanesulfonic acid, pH 1 1, 10% metha- nol. The blot was washed in water, stained with 0.1% Coomassie Blue R-250 in 50% methanol, destained with severa1 changes of 50% methanol, 10% acetic acid, and washed with water (Matsu- daira, 1987). All solutions were freshly prepared and methanol and acetic acid were HPLC grade. The two bands corresponding to PG2A and PG2B were excised from the blot and sequenced together by automated sequential Edman degradation on a gas phase sequenator. The yield of PTH-derivatized amino acid in the initial cycle was near 100%.
Polygalacturonase Deglycosylation
Enzymic deglycosylation of polygalacturonase from transgenic tobacco and tomato was carried out on crude cell wall extracts prepared as above except that the ammonium sulfate pellet was dialyzed against 10 mM sodium acetate pH 5.5. One hundred micrograms of tobacco protein or 5 pg of tomato protein were denatured by boiling for 5 min in the same buffer containing 0.14% SDS and 5 mM 2-mercaptoethanol. The solutions were diluted 1 .5-fold with 1 O mM sodium acetate, pH 5.5, phenylmethylsulfonyl fluotide was added to a concentration of 0.5 mM, and the solutions were incubated overnight at 37°C in the presence of 5 milliunits ofendo H (Boehringer Mannheim).
For chemical deglycosylation, tomato and tobacco cell wall extracts were chromatographed on concanavalin A-Sepharose (Sigma) as described previously (DellaPenna et al., 1986), dialyzed against 0.1% ammonium bicarbonate adjusted to pH 7.2 by bubbling in CO,, lyophilized, and held over phosphorus pentoxide until treated. Deglycosylation with TFMS (Sigma) was carried out in the presence of anisole (Aldrich) as described (Karp et al., 1982). The deglycosylated proteins were dialyzed extensively against 0.1 "/O ammonium bicarbonate and lyophilized.
Both enzymically and chemically deglycosylated proteins were analyzed by SDS-PAGE and immunoblotting.
Gel Electrophoresis and lmmunoblotting
Conditions used for protein separation on native 7.5% polyacryl- amide gels were as described (Reisfeld et al., 1962). Native gels were stained for polygalacturonase activity by the method of Lisker and Retig (1 974). Proteins separated on denaturing lO0/o
polyacrylamide gels (Laemmli, 1970) were either Coomassie Blue stained or silver stained, or electroblotted to nitrocellulose for immunodetection using a 1000-fold dilution of polygalacturonase antiserum (DellaPenna et al., 1986).

Transgenic Expression of Polygalacturonase 1247
Cell Wall Isolation, Digestion, and Polyuronide Determination
Acetone-insoluble, phenol-extracted cell walls were isolated from tobacco leaves and from green tomato fruit pericarp as described previously (Huber, 1983a). EDTA-soluble polyuronides were ex- tracted from the isolated cell walls (Huber, 1983a) and quantitated by the method of Blumenkrantz and Asboe-Hansen (1973).
For enzymic digestion of tobacco and tomato cell walls by tomato polygalacturonase, 5 mg of cell wall isolated as indicated above were digested with 10 pg (approximately 600 nmol of reducing groups min-') of a mixture of purified tomato PG2A and PG2B (DellaPenna et al., 1986) in the presence or absence of 50 pg (7.5 units) of tomato pectin methylesterase (Sigma). The digest was carried out at 25OC in 20 mM sodium citrate, 1 O0 mM NaCI, pH 4.5, with gentle shaking. At various times, aliquots were removed and assayed for soluble uronic acids as above.
Protoplast lsolation
Tobacco leaf tissue was cut into small pieces in a sterile Petri dish in 0.5 M mannitol, 0.1% BSA, 3 mM Mes, pH 5.7 (incubation medium), and washed twice with the same medium. The leaf pieces were incubated at 25OC overnight in incubation medium containing 0.5% Cellulase "Onazuka" R-1 O (Yakult Honsha, To- kyo, Japan) and 0.5% Macerase (Behring Diagnostics), and then shaken gently for 1 hr at 25°C to release protoplasts. Protoplasts were filtered through four layers of cheesecloth, centrifuged at 509 for 15 min (all centrifugations were done in a swinging bucket rotor at 4OC), resuspended in a small volume of incubation me- dium, and layered onto a 10%/17°/o discontinuous gradient of Ficoll 400 (Pharmacia) in incubation medium. After centrifugation at 150g for 30 min, protoplasts collecting at the top of both the 10% and 17% Ficoll layers were pooled, diluted with incubation medium, and centrifuged for 15 min at 509. The pellet was resuspended in a small volume of incubation medium and the protoplasts were counted on a hemacytometer. Yields were 1.4 x 106 to 4.7 x 106 protoplasts per gram of leaf tissue. Protein was extracted from protoplasts as described above.
ACKNOWLEDGMENTS
We thank Monsanto Corporation for providing the plasmid pMON530. This work was supported by the University of Califor- nia Biotechnology Research and Education Program, U.S. De- partment of Agriculture-Competitive Research Grants Office Grant 89-37261 -4641, and Enimont America, Inc.
Received August 7, 1990; accepted October 5, 1990.
REFERENCES
Ali, Z.M., and Brady, C.J. (1 982). Purification and characterization of polygalacturonase of tomato fruits. Aust. J. Plant Physiol. 9, 155-1 69.
Altabella, T., and Chrispeels, M.J. (1 990). Tobacco plants trans- formed with the bean aai gene express an inhibitor of insect 01-
amylase in their seeds. Plant Physiol 93, 805-810. Blumenkrantz, N., and Asboe-Hansen, G. (1973). New method
for quantitative determination of uronic acids. Anal. Biochem. 54,484-489.
Bradford, M.M. (1976). A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72, 248-254.
Brady, C.J., Meldrum, S.K., and McGlasson, W.B. (1983). Dif- ferential accumulation of the molecular forms of polygalacturon- ase in tomato mutants. J. Food Biochem. 7, 7-14.
Buescher, R.W., and Hobson, G.E. (1982). Role of calcium and chelating agents in regulating the degradation of tomato fruit tissue by polygalacturonase. J. Food Biochem. 6, 147-1 60.
Crookes, P.R., and Grierson, D. (1983). Ultrastructure of tomato fruit ripening and the role of polygalacturonase isozymes in cell wall degradation. Plant Physiol. 72, 1088-1 093.
DellaPenna, D., and Bennett, A.B. (1988). In vitro synthesis and processing of tomato fruit polygalacturonase. Plant Physiol. 86,
DellaPenna, D., Alexander, D.C., and Bennett, A.B. (1986). Molecular cloning of tomato fruit polygalacturonase: Analysis of polygalacturonase mRNA levels during ripening. Proc. Natl. Acad. Sci. USA 83,6420-6424.
DellaPenna, D., Kates, D.S., and Bennett, A.B. (1987). Polyga- lacturonase gene expression in Rutgers, rin, nor, and Nr tomato fruit. Plant Physiol. 85, 502-507.
DellaPenna, D., Lashbrook, C.C., Toenjes, K., Giovannoni, J.J., Fischer, R.L., and Bennett, A.B. (1 990). Polygalacturonase isozymes and pectin degradation in transgenic rin tomato fruit. Plant Physiol. 94, 1882-1 886.
Giovannoni, J.J., DellaPenna, D., Bennett, A.B., and Fischer, R.L. (1 989). Expression of a chimeric polygalacturonase gene in transgenic rin (ripening inhibitor) tomato fruit results in polyu- ronide degradation but not fruit softening. Plant Cell 1, 53-63.
Higgins, J.V., Newbigin, E.J., Spencer, D., Llewellyn, D.J., and Craig, C. (1988). The sequence of a pea vicilin gene and its expression in transgenic tobacco plants. Plant MOI. Biol. 11,
Hoffman, L.M., Donaldson, D.D., Bookland, R., Rashka, K., and Herman, E.M. (1 987). Synthesis and protein body deposition of maize 15-kD zein in transgenic tobacco seeds. EMBO J. 6,
Huber, D.J. (1 983a). Polyuronide degradation and hemicellulose modifications in ripening tomato fruit. J. Am. SOC. Hort. Sci.
Huber, D.J. (1983b). The role of cell wall hydrolases in fruit softening. Hort. Rev. 5, 169-219.
Hunkapiller, M.W., Lujan, E., Ostrander, F., and Hood, L. (1981). lsolation of microgram quantities of protein from polyacrylamide gels for amino acid sequence analysis. Methods Enzymol. 91,
Karp, D.R., Atkinson, J.P., and Shreffler, D.C. (1982). Genetic variation in glycosylation of the fourth component of murine complement. J. Biol. Chem. 257, 7330-7335.
1057-1 063.
683-695.
3213-3221.
108,405-409.
227-236.

1248 The Plant Cell
Knegt, E., Vermeer, E., and Bruinsma, J. (1988). Conversion of the polygalacturonase isozymes from ripening tomato fruits. Physiol. Plant. 72, 108-1 14.
Kornfeld, R., and Kornfeld, S. (1985). Assembly of asparagine- linked oligosaccharides. Annu. Rev. Biochem. 54, 631 -664.
Laemmli, U.K. (1 970). Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227,
Lisker, N., and Retig, N. (1974). Detection of polygalacturonase and pectinylase isozymes in polyacrylamide gels. J. Chroma- togr. 96,245-249.
Matsudaira, P. (1 987). Sequence from picomole quantities of proteins electroblotted onto polyvinylidene difluoride mem- branes. J. Biol. Chem. 262, 10035-10038.
Moshrefi, M., and Luh, B.S. (1 983). Carbohydrate composition and electrophoretic properties of tomato polygalacturonase isozymes. Eur. J. Biochem. 135, 511-514.
Murashige, T., and Skoog, F. (1 962). A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 15, 473-497.
Nitsch, J.P. (1 969). Experimental androgenesis in Nicotiana. Py- tomorphology 19, 389-404.
Otten, L.B., and Schilperoort, R.A. (1978). A rapid micro scale method for the detection of lysopine and nopaline dehydroge- nase activities. Biochim. Biophys. Acta 527, 497-500.
Post-Beittenmiller, M.A., Schmid, K.M., and Ohlrogge, J.B. (1 989). Expression of holo and apo forms of spinach acyl carrier protein-l in leaves of transgenic tobacco plants. Plant Cell 1, 889-899.
Pressey, R. (1 984). Purification and characterization of tomato polygalacturonase converter. Eur. J. Biochem. 144,217-221.
Pressey, R. (1 986). Changes in polygalacturonase isozymes and converter in tomatoes during ripening. HortScience 21,
Pressey, R. (1 988). Reevaluation of the changes in polygalactu- ronases in tomatoes during ripening. Planta 174, 39-43.
Pressey, R., and Avants, J.K. (1973). Two forms of polygalac- turonase in tomatoes. Biochim. Biophys. Acta 309, 363-369.
Pressey, R., and Avants, J.K. (1982). Solubilization of cell walls by tomato polygalacturonases: Effects of pectinesterases. J. Food Biochem. 6,57-74.
Reisfeld, R.A., Lewis, U.J., and Williams, D.E. (1962). Disc electrophoresis of basic proteins and peptides in polyacrylamide gels. Nature 195, 281-283.
680-685.
11 83-1 185.
Rogers, S.G., Horsch, R.B., and Fraley, R.T. (1986). Gene tranS- fer in plants: Production of transformed plants using Ti plasmid vectors. Methods Enzymol. 118, 627-641.
Shah, D.M., Horsch, R.B., Klee, H.J., Kishore, G.M., Winter, J.A., Tumer, N.E., Hironaka, C.M., Sanders, D.R., Gasser, C.S., Aykent, S., Siegel, N.R., Rogers, S.G., and Fraley, R.T. (1 986). Engineering herbicide tolerance in transgenic plants. Science 233,478-481.
Sheehy, R.E., Pearson, J., Brady, C.J., and Hiatt, W.R. (1987). Molecular characterization of tomato fruit polygalacturonase. MOI. Gen. Genet. 208, 30-36.
Smith, C.J.S., Watson, C.F., Morris, P.C., Bird, C.R., Seymour, G.B., Gray, J.E., Arnold, C., Tucker, G.A., Schuch, W., Hard- ing, S., and Grierson, D. (1990). lnheritance and effect on ripening of antisense polygalacturonase genes in transgenic tomatoes. Plant MOI. Biol. 14, 369-379.
Smith, P.K., Krohn, R.I., Hermanson, G.T., Molha, A.K., Gartner, F.H., Provenrano, M.D., Fujimoto, E.K., Goeke, N.M., Olson, B.J., and Klenk, D.C. (1985). Measurement of protein using bicinchoninic acid. Anal. Biochem. 150, 76-85.
Sonnewald, U., Sturm, A., Chrispeels, M.J., and Willmitrer, L. (1 989). Targeting and glycosylation of patatin, the major potato tuber protein, in leaves of transgenic tobacco. Planta 179,
Sturm, A., Voelker, T.A., Herman, E.M., and Chrispeels, M.J. (1 988). Correct glycosylation, Golgi processing, and targeting to protein bodies of the vacuolar protein phytohemagglutinin in transgenic tobacco. Planta 175, 170-183.
Tarentino, A.L., and Plummer, T.H., Jr. (1 982). Oligosaccharide accessibility to peptide:N-glycosidase as promoted by protein- unfolding reagents. J. Biol. Chem. 257, 10776-10780.
Themmen, A.P., Tucker, G.A., and Grierson, D. (1982). Degra- dation of isolated tomato cell walls by purified polygalacturon- ase in vitro. Plant Physiol. 69, 122-124.
Tucker, G.A., Robertson, N.G., and Grierson, D. (1980). Changes in polygalacturonase isozymes during "ripening" of normal and mutant tomato fruit. Eur. J. Biochem. 112,
Tucker, G.A., Robertson, N.G., and Grierson, D. (1981). The conversion of tomato-fruit polygalacturonase isozyme 2 into isozyme 1 in vitro. Eur. J. Biochem. 115, 87-90.
Wilkins, T.A., Bednarek, S.Y., and Raikhel, N.V. (1990). Role of propeptide glycan in post-translational processing and transport of barley lectin to vacuoles in transgenic tobacco. Plant Cell 2,
171-180.
1 19-1 24.
301 -31 3.

DOI 10.1105/tpc.2.12.1239 1990;2;1239-1248Plant Cell
K W Osteryoung, K Toenjes, B Hall, V Winkler and A B BennettAnalysis of tomato polygalacturonase expression in transgenic tobacco.
This information is current as of November 23, 2018
Permissions https://www.copyright.com/ccc/openurl.do?sid=pd_hw1532298X&issn=1532298X&WT.mc_id=pd_hw1532298X
eTOCs http://www.plantcell.org/cgi/alerts/ctmain
Sign up for eTOCs at:
CiteTrack Alerts http://www.plantcell.org/cgi/alerts/ctmain
Sign up for CiteTrack Alerts at:
Subscription Information http://www.aspb.org/publications/subscriptions.cfm
is available at:Plant Physiology and The Plant CellSubscription Information for
ADVANCING THE SCIENCE OF PLANT BIOLOGY © American Society of Plant Biologists






![Transgenic Expression ofaFungalendo-Polygalacturonase ... · Transgenic Expression ofaFungalendo-Polygalacturonase Increases Plant Resistance to Pathogens and Reduces Auxin Sensitivity1[W]](https://img.pdfslide.net/doc/110x75/5f5599280b86d949f711ae38/transgenic-expression-ofafungalendo-polygalacturonase-transgenic-expression.jpg)






















