Embed Size (px)
Citation preview

Cancer Therapy: Clinical
Antibiotic Treatment Decreases Microbial BurdenAssociated with Pseudomyxoma Peritonei and Affectsb-Catenin Distribution
Cristina Semino-Mora1, Traci L. Testerman4, Hui Liu1, Jeannette M. Whitmire1, Kimberley Studeman2,Yali Jia4, Thomas J. McAvoy3, Jennifer Francis4, Carol Nieroda4, Armando Sardi4, D. Scott Merrell1, andAndre Dubois1
AbstractPurpose: Pseudomyxoma peritonei is an understudied cancer in which an appendiceal neoplasm
invades the peritoneum and forms tumor foci on abdominal organs. Previous studies have shown that
bacteria reside within pseudomyxoma peritonei tumors andmucin. Thus, we sought to analyze the effect of
antibiotics on bacterial density and b-catenin expression within pseudomyxoma peritonei samples.
ExperimentalDesign: The study included 48patients: 19with disseminated peritoneal adenomucinosis
(DPAM)and29withperitonealmucinous carcinomatosis (PMCA). Fourteenpatientswere given antibiotics
(30 mg lansoprazole, 1 g amoxicillin, and 500 mg clarithromycin) twice a day for 14 days. One week after
completion of therapy, surgery was conducted and specimens were harvested for pathology, bacterial
culture, ISH, and immunohistochemistry.
Results: ISH showed the presence of bacteria in 83% of the patient samples, with a higher Helicobacter
pylori density observed in PMCA versus DPAM. PMCA patients treated with antibiotics had a significantly
lower bacterial density anddecreasedb-catenin levels in the cytoplasm, the cell nuclei, andmucin-associated
cells. Although not significant, similar trends were observed in DPAM patients. Cell membrane b-cateninwas significantly increased in both DPAM and PMCA patients receiving antibiotics.
Conclusions: Bacteria play an important role in pseudomyxoma peritonei. Antibiotic treatment
improved the histopathology of tissue, particularly in PMCA patients. In PMCA, antibiotics decreased
bacterial density and were associated with a significant b-catenin decrease in the cytoplasm, cell nuclei,
and mucin along with a small membrane increase. These results suggest that antibiotics offer potential
protection against cell detachment, cellular invasion, and metastasis. Clin Cancer Res; 19(14); 3966–76.
�2013 AACR.
IntroductionPseudomyxoma peritonei is characterized by the pres-
ence of multifocal peritoneal and omental implants ofmucus-secreting epithelial cells and dissecting gelatinousascites (1). This complex disease is found in 0.02% oflaparotomies, and its incidence is threefold higher in
females than in males (2, 3). Pseudomyxoma peritoneiusually originates from a perforated low-grade appendi-ceal mucinous neoplasm or appendiceal adenocarcino-ma that protrudes into the peritoneum, and the initialpresentation of the disease often includes increasedabdominal girth and/or appendicitis-like symptoms(4). Pseudomyxoma peritonei can take on a protractedcourse if the histopathology is that of disseminatedperitoneal adenomucinosis (DPAM), or rapidly evolveas a highly malignant tumor with peritoneal mucinouscarcinomatosis (PMCA) histopathology. Current treat-ment combines cytoreductive surgery (resection of thetumor and peritoneal implants) and intraperitonealhyperthermic chemotherapy (5–8). Despite extensiveresections, pseudomyxoma peritonei frequently relapsesand 5-year survival is only 53% (9), although survival issignificantly better if lymph nodes are negative (5-yearsurvival is 76% for lymph node-negative patients and11% for lymph node-positive patients; P < 0.001; ref. 10).Even in patients with low-grade appendiceal mucinousneoplasms, most patients succumb to the disease due
Authors' Affiliations: 1Uniformed Services University and United StatesMilitary Cancer Institute, Bethesda; 2Mercy Medical Center, Baltimore;3University of Maryland, College Park, Maryland; and 4Louisiana StateUniversity Health Sciences Center-Shreveport, Shreveport, Louisiana
Note: Supplementary data for this article are available at Clinical CancerResearch Online (http://clincancerres.aacrjournals.org/).
Corresponding Authors: Thomas J. McAvoy, University of Maryland,College Park, MD 20742. Phone: 301-405-1935; Fax: 301-314-9126;E-mail: [email protected]; and D. Scott Merrell, Department of Microbi-ology, UniformedServicesUniversity of theHealth Sciences, RoomB4140,Bethesda, MD 20814. Phone: 301-295-1584; Fax: 301-295-3773; E-mail:[email protected]
doi: 10.1158/1078-0432.CCR-13-0616
�2013 American Association for Cancer Research.
ClinicalCancer
Research
Clin Cancer Res; 19(14) July 15, 20133966

to recurrent mucoid ascites, proliferation of gelatinousmucin, and mechanical compression and obstruction ofabdominal organs, heart, and lungs (11).A key aspect of colon cancer and several other cancers is
deregulation of theWnt/b-catenin pathway (12). The trans-membrane protein b-catenin is associated with the cyto-plasmic region of E-cadherin within adherens junctions. Innormal, polarized gut epithelial cells, b-catenin provides alinkage mechanism between cytoskeletal proteins and cell-to-cell junctional proteins (e.g., E-cadherin), allowing cellsto tightly bind to each other (13). These interactions arecrucial formaintenanceof epithelial cell polarity, regulationof cell growth, and cell-to-cell focal adhesion. During car-cinogenesis, b-catenin relocates into the cytoplasm andnucleus. Loss of membrane b-catenin leads to cell separa-tion andmigration,movement of abnormal neoplastic cellsto the stroma, entry into blood vessels, and metastaticspread to different tissues through the bloodstream (14).Furthermore, following nuclear translocation, b-catenininteracts with T-cell factors (TCF) or lymphocyte-enhancerfactors that can then act as transcription factors. Thus,b-catenin nuclear localization triggers expression of variousgenes, including genes required for cell proliferation (15).Several infectious agents have been found to stimulate
cell proliferation via theWnt/b-catenin pathway. In fact, thepotential for certain viruses to cause cancer has been rec-ognized for some time. Recent studies suggest that Epstein–Barr virus, Kaposi’s sarcoma-associated virus, and hepatitisC virus promote carcinogenesis by activating the Wnt/b-catenin pathway (16, 17). Some bacteria also influencethe Wnt/b-catenin pathway. Helicobacter pylori expressesnumerous virulence factors that promote carcinogenesis,including the cytotoxin-associated gene A (CagA). Translo-cation of CagA into epithelial cells results in changes inexpression of b-catenin (18) as well as causes nuclear
accumulation of b-catenin (19), which is associated withaggressive and invasive tumors (20). Salmonella and Chla-mydia trachomatis also stimulate epithelial proliferation viathe Wnt/b-catenin pathway (21, 22).
Although pseudomyxoma peritonei patients have nosymptoms of peritonitis, we previously hypothesized thatintestinal bacteria spread to the peritoneum at the time ofappendiceal perforation. In keeping with this idea, werecently showed that H. pylori and other bacteria can bedetected in pseudomyxoma peritonei tissues (23). Giventhis finding, herein we studied the effect of preoperativeantibiotic therapy on bacterial density, and on the concur-rent expression of b-cateninwithin the neoplastic cancerouscells of DPAM and PMCA patients. We observed that afterantibiotic treatment, PMCA patients showed a significantdecrease in bacterial density along with significantlydecreased b-catenin expression in the cytoplasm, nuclei,and mucin. We found that in neoplastic cancerous cells,b-catenin is redistributed into the cytoplasm and accumu-lates in different areas of the cytosol and ground substance(connective tissue in the stroma that supports fibers). Enmasse, our data suggest that antibiotic treatment of PMCAmay affect the carcinogen pathway elicited by b-catenin andmay serve as a novel treatment of this understudied cancer.
Materials and MethodsPatients, histopathology, and treatment
Forty-eight patients with the diagnosis of peritonealdissemination of appendiceal mucinous neoplasms thathad been scheduled to undergo laparotomy for staging,extensive cytoreductive surgery, and hyperthermic intraper-itoneal chemotherapywere studied. Patients’ agewas 53�2years and weight 73 � 2 kg (means and SEM).
Observation and analysis of biopsies stained with hema-toxylin and eosin (H&E) from cytoreductive surgical speci-mens by a board certified and experienced pathologistallowed tumor classification as either DPAM or as the moremalignant PMCA (24). Tissue from a patient with a non-perforated, nonneoplastic appendix (NNA) was used as acontrol. Analysis of the grade of inflammation was con-ducted according to conditions previously described (25).
Three weeks before surgery, an open label anti-H. pyloritriple therapy (Prevpac, i.e., 30 mg lansoprazole, 1,000 mgamoxicillin, and 500 mg clarithromycin given twice a dayfor 14 days) was given to a total of 14 of the pseudomyxomaperitonei patients: 6 DPAM and 8 PMCA. A total of 34patients received no antibiotics and were maintained asuntreated controls: 13DPAMand 21PMCA.Oneweek aftercompletion of the antibiotic therapy, patients and controlsunderwent cytoreductive surgery (resection of the tumorand peritoneal implants) and hyperthermic intraperitonealchemotherapy to achieve complete or near-complete resec-tion of pseudomyxoma peritonei cancerous tissues (8).
ISH studiesProbes. Two previously described 16S rDNA bacterial
probes (23) labeled with biotin were as follows: (i) a probe
Translational RelevancePseudomyxoma peritonei is an underresearched
abdominal cancer that originates from an appendicealneoplasm. Patients frequently relapse and succumb tothe disease, necessitating the development of noveltherapeutics to treat patients. Recent studies have indi-cated the presence of bacteria within the tumors andmucin of pseudomyxoma peritonei patients. The studyherein indicates that antibiotic treatment significantlyreduces bacterial density along with b-catenin levels inthe cytoplasm, the cell nuclei, andmucin-associated cellsof peritoneal mucinous carcinomatosis (PMCA)patients. In addition, the b-catenin levels within mem-branes significantly increase in both disseminated peri-toneal adenomucinosis and PMCA patients. The renor-malization of b-catenin distribution combined with thereduction in bacterial density make antibiotic treatmenta potential therapeutic for pseudomyxoma peritoneidisease.
Pseudomyxoma Peritonei, Bacteria, and Antibiotics
www.aacrjournals.org Clin Cancer Res; 19(14) July 15, 2013 3967

that can detect 19,973 typed and nonculturable bacteria(TNCB; including Campylobacter jejuni, Escherichia coli, Sal-monella enterica, H. pylori, and Enterococcus faecalis; 5’-AGCAAA CAG GAT TAG ATA CCC TGG TAG TCC AC-3’); and (ii) a probe specific for H. pylori (5’-ATT TCA CACCTG ACT GAC TAT CCC GCC TAC GCG-3’; ref. 26). Ininitial experiments, we used both cRNA and cDNA probesconcurrently, and confirmed earlier observations that these2 probes can detect the same bacteria (27).
Paraffin blocks of formalin-fixed tissue were sectioned(5 mm) and analyzed as previously described (27). Briefly,each unstained section was deparaffinized, prehybridized,hybridized with denatured probe solution, and then incu-bated for 18 hours with a probe labeled with biotin fol-lowed by the removal of unbound probe using salinecitrate solution. DNA was detected by incubating theslide for 2 hours with streptavidin-conjugated alkalinephosphatase (Roche Diagnostic; 1:500 dilution in blockingimmuno-Tris buffer), washing, and applying, the chromo-genic substrate BCIP/NBT/kit (5-bromop-4-chloro-3-indo-lyl phosphate/nitro-blue-tetrazolium), yielding a blue colorreaction (Vector Labs). The slide was then counterstainedwithNuclear Fast Red, washed inwater, dehydrated, clearedin xylene, air dried, and mounted with permount. TNCBand H. pylori 16S rDNA were detected in serial sectionsunder bright light as follows: first section: detection ofTNCB 16S rDNA, second section: detection of H. pylori16S rDNA. Parallel dual FISH studies (27) were also con-ductedusing the16S rDNA-TNCBprobe labeledwithbiotinand then detected with avidin labeled with fluorescein–fluorescein isothiocyanate (FITC; green reaction), and the16S rDNA-H. pylori probe labeled with digoxigenin andthen detected with anti-digoxigenin mouse monoclonalantibody conjugated to biotin (Abcam) and then detectedusing avidin-Tx red (red reaction). Colocalization of bothTNCB and H. pylori was visualized as yellow.
As previously described (27), control for nonspecific bind-ing included: (i) sense probe instead of antisense probe, (ii)hybridization buffer instead of the antisense probe, (iii)unlabeledantisenseprobe, (iv)digoxigeninorbiotin-labeledprobe for a sequence completely unrelated to man andprokaryotes—the scorpion Buthus martensi Karsch neurotox-insequence(5’-GGCCACGCGTCGACTAGTAC-3’; ref. 28),(v) RNaseApretreatment (Roche), (vi)DNase I pretreatment(Roche), and (vii) RNase þ DNase I pretreatment.
For fluorescence, a Nikon Eclipse 80i microscope DSblack-white camera with a DS-L2 control unit was usedand software from NIS-Elements Advanced Research wasused for analysis and reproduction of b-catenin images. Forbrightfield microscopy, an Eclipse 800 Nikon microscopewith digital camera (QCapture Pro, Micropublisher 6.0;Burnaby) was used for analysis and reproduction of bacte-rial images (TNCB and H. pylori). Fluorescence immuno-histochemistry (FIHC) was used for studies of b-catenin.Two different studies were conducted:
Laser confocal microscopy was used to quantify cell mem-brane andnuclear localizationofb-catenin. Antigen retrievalwas first achieved by sequentially treating deparaffinized
sections with saline citrate solution in a pressure cookersystem for 9 minutes (27). b-Catenin expression was ana-lyzed using a mouse anti-b-catenin monoclonal antibody(Santa Cruz Biotechnology, Inc.) at 1:150 dilution followedby biotinylated anti-mouse immunoglobulin G and avidin,labeled with either FITC for b-catenin cell membrane local-ization or Texas red for b-catenin nuclear localization. Fornuclear localization, DNA was also stained using 4’,6-dia-mino-2-phenylindole (DAPI) because it specifically bindsDNA without overlap with fluorochromes. Importantly,DAPI binds DNA (heterochromatin and euchromatin) ofan interphase nucleus. In contrast to usual dual-fluorescencestains, DAPI stain cannot be easily merged with the expres-sion of any fluorochrome (in this case with b-catenin-Texasred) to produce a third color because nuclear DNA is boundto b-catenin (first layer of procedure) and, as a result, DAPIbinding to DNA is not as efficient at the same site. However,DAPI can bind to DNA that is not bound to b-catenin.
b-Catenin compartment localization was determined fromstandard fluorescence (FIHC) of paraffin sections mountedwith Vectashield alone (w/o DAPI) and observed using aNikon Eclipse 80i microscope as described earlier. Thefollowing tissue types were analyzed: (i) epithelia; (ii)lymphocytes, monocytes, and granulocytes; (iii) stroma(connective tissue, vessels, fibroblasts, and fibers); and (iv)mucin pools. In addition, specific analyses of (i) lateral cell–cell contact and (ii) cytoplasm localizationwere conducted.
Fluorescence IHC images were observed and reproducedusing the same microscope and digital camera describedearlier for the ISH studies.
For b-catenin nuclear localization, sections weremounted using Vectashield containing DAPI and confocalimages were collected on a Zeiss S710 NLO laser scanningconfocal microscope (Carl Zeiss) equipped with a 63� oilimmersion lens (1.4NA). DAPI-stained samples were excit-edwith a 405-nm laser diode and emitted light in the 410 to556 nm range was collected. For Texas red–stained samples,a laser line of 561 nm was used for excitation and lightbetween 566 and 690 nm was used for imaging (BIC,USUHS). Phosphate buffer solution instead of the firstb-catenin antibody was used as a negative control.
Morphometric analysisQuantitation of bacteria and b-catenin. Determination
of bacterial and b-catenin density in cellular compartmentswas conducted as previously described (27). Briefly, anintraocular grid-based method was used to examine sam-ples by a microscopist who was blinded to the histopath-ologic diagnoses (i.e., DPAMor PMCA) as well as antibiotictreatment category. The number of bacterial clusters expres-sing 16S rDNA of TNCB and/or H. pylori was quantified at400� magnification in 3 randomly selected fields of viewaccording to a modification of the point-counting stereo-logical method, and using an intraocular reticle of 27-mmdiameter, covering 3578 mm2 (i.e., 17,892 mm3 for 5-mm-thick sections; Kr409, Klarman Rulings, Inc.; ref. 23).Counting of the number of intersections of vertical andhorizontal lines that overlapped a bacterium in the area
Semino-Mora et al.
Clin Cancer Res; 19(14) July 15, 2013 Clinical Cancer Research3968
delimited by a projected grid on the tissue was conducted.All data were expressed asmeans� SEMnumber of bacteriaper 106 mm3 (representing an imaginary cube with sides of100 mm or 0.1 mm).Total b-catenin density in biopsies was determined in a
similar fashion and expressed as Vvi (volumetric density orvolume occupied for the b-catenin reaction) in 106 mm3 oftissue.Computerized quantification of b-catenin in the nuclei.
Nuclear localization of b-catenin and analysis were con-ducted using a Zeiss 710 NLO laser scanning confocalmicroscope (Carl Zeiss) as described earlier (BIC, USUHS).Software developed by TMusing aMATLAB platform and
associated toolboxes (http://www.mathworks.com/) wasused to quantify the fraction of the nucleus occupied byb-catenin in confocal microscopic pictures of IHC sectionsthat had been double stained for both b-catenin (Texas red)and DNA (DAPI; Supplementary Methods). In brief, thepercent of nuclei volume occupied by b-catenin was calcu-lated as the number of bright red pixels that overlapped thedark blueDAPI stain divided by total nuclei pixels (nuclei inblue) multiplied by 100.
Statistical analysisStatistical significance was determined using t tests to
compare results in patients receiving antibiotics versus no
antibiotics in each cell of the tables. Values were expressedas means � SEM.
ResultsPathological evaluation of DPAM and PMCA
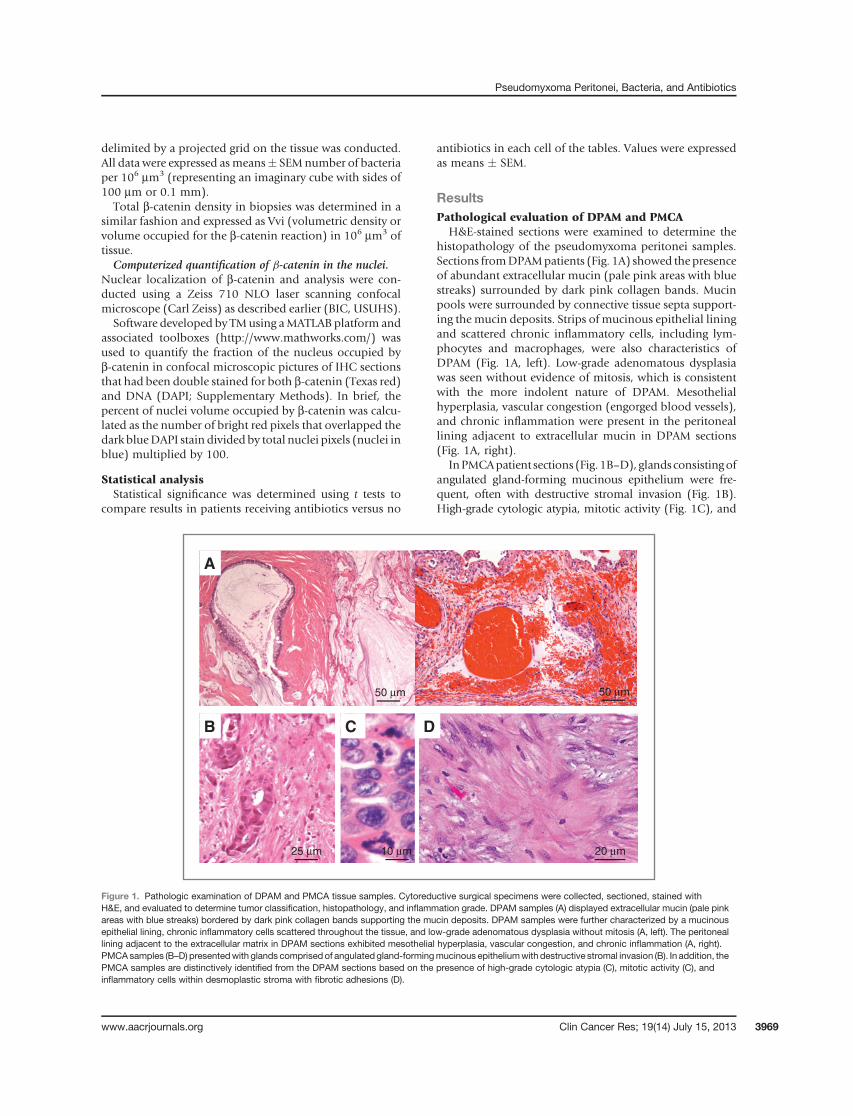
H&E-stained sections were examined to determine thehistopathology of the pseudomyxoma peritonei samples.Sections fromDPAMpatients (Fig. 1A) showed the presenceof abundant extracellular mucin (pale pink areas with bluestreaks) surrounded by dark pink collagen bands. Mucinpools were surrounded by connective tissue septa support-ing the mucin deposits. Strips of mucinous epithelial liningand scattered chronic inflammatory cells, including lym-phocytes and macrophages, were also characteristics ofDPAM (Fig. 1A, left). Low-grade adenomatous dysplasiawas seen without evidence of mitosis, which is consistentwith the more indolent nature of DPAM. Mesothelialhyperplasia, vascular congestion (engorged blood vessels),and chronic inflammation were present in the peritoneallining adjacent to extracellular mucin in DPAM sections(Fig. 1A, right).
InPMCApatient sections (Fig.1B–D), glandsconsistingofangulated gland-forming mucinous epithelium were fre-quent, often with destructive stromal invasion (Fig. 1B).High-grade cytologic atypia, mitotic activity (Fig. 1C), and
A
B C D
25 μm 10 μm 20 μm
50 μm 50 μm
Figure 1. Pathologic examination of DPAM and PMCA tissue samples. Cytoreductive surgical specimens were collected, sectioned, stained withH&E, and evaluated to determine tumor classification, histopathology, and inflammation grade. DPAM samples (A) displayed extracellular mucin (pale pinkareas with blue streaks) bordered by dark pink collagen bands supporting the mucin deposits. DPAM samples were further characterized by a mucinousepithelial lining, chronic inflammatory cells scattered throughout the tissue, and low-grade adenomatous dysplasia without mitosis (A, left). The peritoneallining adjacent to the extracellular matrix in DPAM sections exhibited mesothelial hyperplasia, vascular congestion, and chronic inflammation (A, right).PMCAsamples (B–D) presentedwith glands comprised of angulated gland-formingmucinous epitheliumwith destructive stromal invasion (B). In addition, thePMCA samples are distinctively identified from the DPAM sections based on the presence of high-grade cytologic atypia (C), mitotic activity (C), andinflammatory cells within desmoplastic stroma with fibrotic adhesions (D).
Pseudomyxoma Peritonei, Bacteria, and Antibiotics
www.aacrjournals.org Clin Cancer Res; 19(14) July 15, 2013 3969
inflammatory cells within a desmoplastic stroma with fibro-tic adhesions differentiate PMCA from DPAM (Fig. 1D);desmoplasia is a fibrotic reaction tomalignant cells invadingnormal tissue. No significant differences in inflammatorycell densitieswere observed across the 2 disease types inH&Eslides (scores of 0–3; ref. 29).
Presence of bacteria in DPAM and PMCA as determinedby ISH
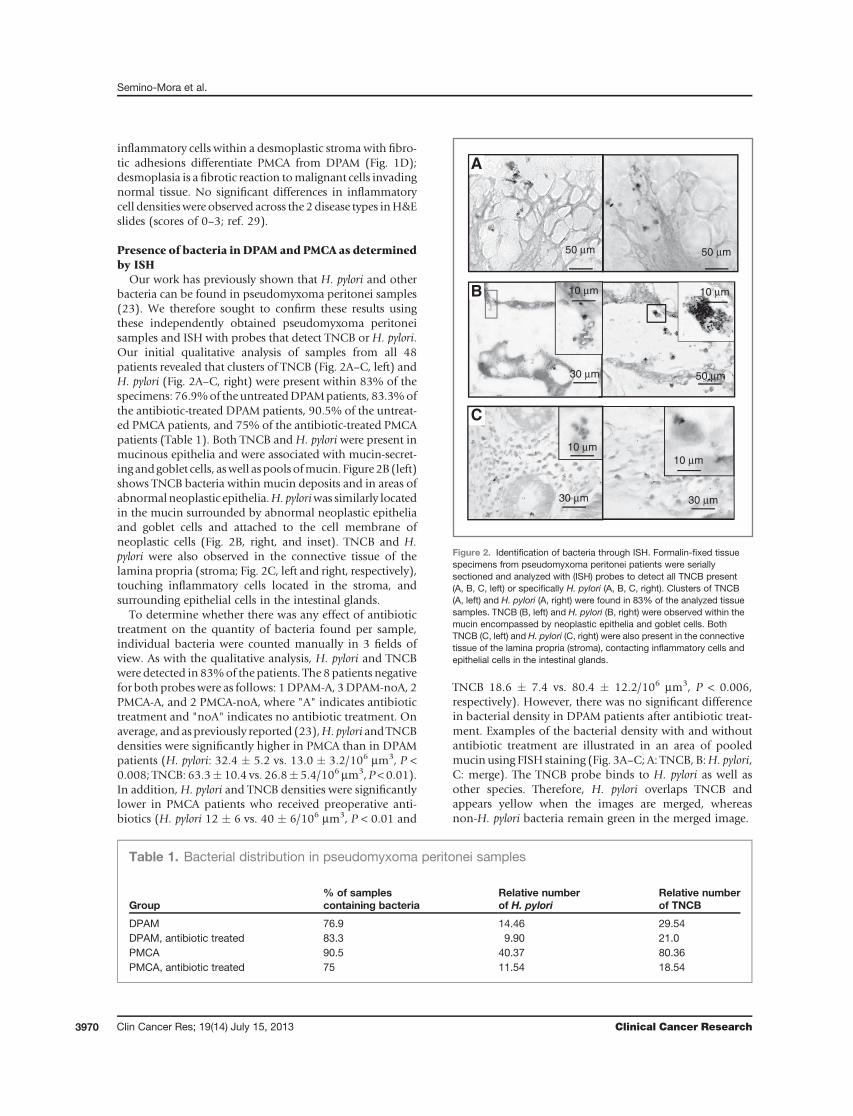
Our work has previously shown that H. pylori and otherbacteria can be found in pseudomyxoma peritonei samples(23). We therefore sought to confirm these results usingthese independently obtained pseudomyxoma peritoneisamples and ISH with probes that detect TNCB or H. pylori.Our initial qualitative analysis of samples from all 48patients revealed that clusters of TNCB (Fig. 2A–C, left) andH. pylori (Fig. 2A–C, right) were present within 83% of thespecimens: 76.9%of the untreatedDPAMpatients, 83.3%ofthe antibiotic-treated DPAM patients, 90.5% of the untreat-ed PMCA patients, and 75% of the antibiotic-treated PMCApatients (Table 1). Both TNCB and H. pylori were present inmucinous epithelia and were associated with mucin-secret-ing andgoblet cells, aswell aspools ofmucin. Figure2B(left)shows TNCB bacteria within mucin deposits and in areas ofabnormalneoplastic epithelia.H.pyloriwas similarly locatedin the mucin surrounded by abnormal neoplastic epitheliaand goblet cells and attached to the cell membrane ofneoplastic cells (Fig. 2B, right, and inset). TNCB and H.pylori were also observed in the connective tissue of thelamina propria (stroma; Fig. 2C, left and right, respectively),touching inflammatory cells located in the stroma, andsurrounding epithelial cells in the intestinal glands.
To determine whether there was any effect of antibiotictreatment on the quantity of bacteria found per sample,individual bacteria were counted manually in 3 fields ofview. As with the qualitative analysis, H. pylori and TNCBwere detected in 83%of the patients. The 8patients negativefor both probeswere as follows: 1DPAM-A, 3DPAM-noA, 2PMCA-A, and 2 PMCA-noA, where "A" indicates antibiotictreatment and "noA" indicates no antibiotic treatment. Onaverage, andaspreviously reported (23),H. pylori andTNCBdensities were significantly higher in PMCA than in DPAMpatients (H. pylori: 32.4 � 5.2 vs. 13.0 � 3.2/106 mm3, P <0.008; TNCB: 63.3� 10.4 vs. 26.8� 5.4/106 mm3, P < 0.01).In addition,H. pylori and TNCB densities were significantlylower in PMCA patients who received preoperative anti-biotics (H. pylori 12 � 6 vs. 40 � 6/106 mm3, P < 0.01 and
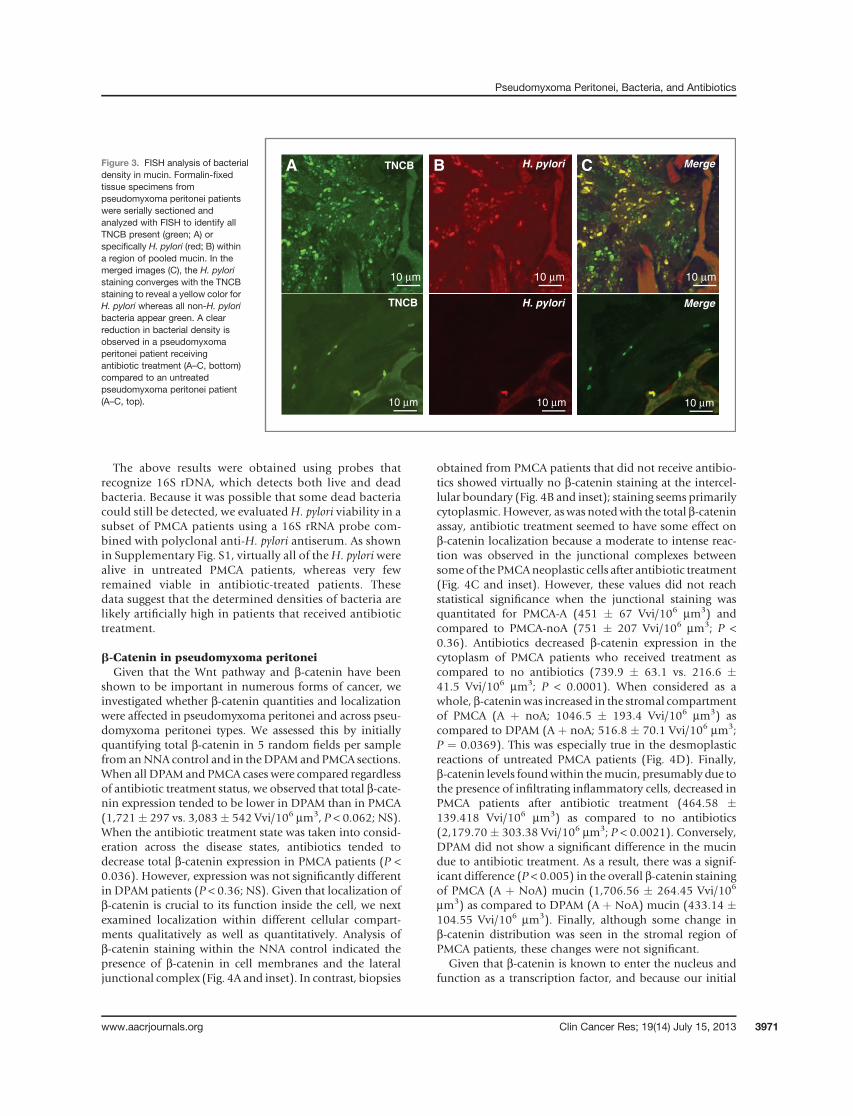
TNCB 18.6 � 7.4 vs. 80.4 � 12.2/106 mm3, P < 0.006,respectively). However, there was no significant differencein bacterial density in DPAM patients after antibiotic treat-ment. Examples of the bacterial density with and withoutantibiotic treatment are illustrated in an area of pooledmucin using FISH staining (Fig. 3A–C; A: TNCB, B:H. pylori,C: merge). The TNCB probe binds to H. pylori as well asother species. Therefore, H. pylori overlaps TNCB andappears yellow when the images are merged, whereasnon-H. pylori bacteria remain green in the merged image.
B
50umA
50 μm 50 μm
B
30 μm
10 μm
50 μm
10 μm
E
C
30 μm30 μm
10 μm10 μm
Figure 2. Identification of bacteria through ISH. Formalin-fixed tissuespecimens from pseudomyxoma peritonei patients were seriallysectioned and analyzed with (ISH) probes to detect all TNCB present(A, B, C, left) or specifically H. pylori (A, B, C, right). Clusters of TNCB(A, left) and H. pylori (A, right) were found in 83% of the analyzed tissuesamples. TNCB (B, left) and H. pylori (B, right) were observed within themucin encompassed by neoplastic epithelia and goblet cells. BothTNCB (C, left) and H. pylori (C, right) were also present in the connectivetissue of the lamina propria (stroma), contacting inflammatory cells andepithelial cells in the intestinal glands.
Table 1. Bacterial distribution in pseudomyxoma peritonei samples
Group% of samplescontaining bacteria
Relative numberof H. pylori
Relative numberof TNCB
DPAM 76.9 14.46 29.54DPAM, antibiotic treated 83.3 9.90 21.0PMCA 90.5 40.37 80.36PMCA, antibiotic treated 75 11.54 18.54
Semino-Mora et al.
Clin Cancer Res; 19(14) July 15, 2013 Clinical Cancer Research3970
The above results were obtained using probes thatrecognize 16S rDNA, which detects both live and deadbacteria. Because it was possible that some dead bacteriacould still be detected, we evaluatedH. pylori viability in asubset of PMCA patients using a 16S rRNA probe com-bined with polyclonal anti-H. pylori antiserum. As shownin Supplementary Fig. S1, virtually all of theH. pyloriwerealive in untreated PMCA patients, whereas very fewremained viable in antibiotic-treated patients. Thesedata suggest that the determined densities of bacteria arelikely artificially high in patients that received antibiotictreatment.
b-Catenin in pseudomyxoma peritoneiGiven that the Wnt pathway and b-catenin have been
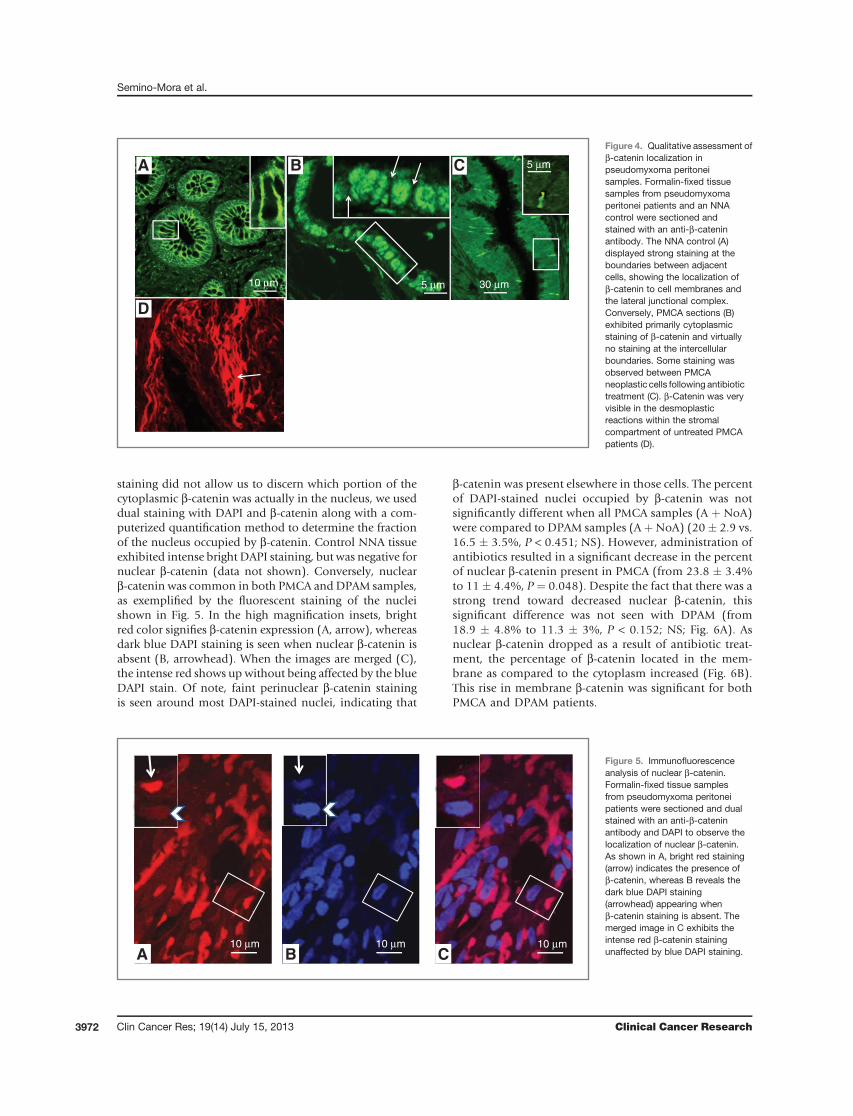
shown to be important in numerous forms of cancer, weinvestigated whether b-catenin quantities and localizationwere affected in pseudomyxoma peritonei and across pseu-domyxoma peritonei types. We assessed this by initiallyquantifying total b-catenin in 5 random fields per samplefromanNNA control and in theDPAMand PMCA sections.When all DPAM and PMCA cases were compared regardlessof antibiotic treatment status, we observed that total b-cate-nin expression tended to be lower in DPAM than in PMCA(1,721� 297 vs. 3,083� 542 Vvi/106 mm3, P < 0.062; NS).When the antibiotic treatment state was taken into consid-eration across the disease states, antibiotics tended todecrease total b-catenin expression in PMCA patients (P <0.036). However, expression was not significantly differentin DPAM patients (P < 0.36; NS). Given that localization ofb-catenin is crucial to its function inside the cell, we nextexamined localization within different cellular compart-ments qualitatively as well as quantitatively. Analysis ofb-catenin staining within the NNA control indicated thepresence of b-catenin in cell membranes and the lateraljunctional complex (Fig. 4A and inset). In contrast, biopsies
obtained from PMCA patients that did not receive antibio-tics showed virtually no b-catenin staining at the intercel-lular boundary (Fig. 4B and inset); staining seems primarilycytoplasmic. However, aswas notedwith the total b-cateninassay, antibiotic treatment seemed to have some effect onb-catenin localization because a moderate to intense reac-tion was observed in the junctional complexes betweensomeof the PMCAneoplastic cells after antibiotic treatment(Fig. 4C and inset). However, these values did not reachstatistical significance when the junctional staining wasquantitated for PMCA-A (451 � 67 Vvi/106 mm3) andcompared to PMCA-noA (751 � 207 Vvi/106 mm3; P <0.36). Antibiotics decreased b-catenin expression in thecytoplasm of PMCA patients who received treatment ascompared to no antibiotics (739.9 � 63.1 vs. 216.6 �41.5 Vvi/106 mm3; P < 0.0001). When considered as awhole, b-cateninwas increased in the stromal compartmentof PMCA (A þ noA; 1046.5 � 193.4 Vvi/106 mm3) ascompared to DPAM (A þ noA; 516.8 � 70.1 Vvi/106 mm3;P ¼ 0.0369). This was especially true in the desmoplasticreactions of untreated PMCA patients (Fig. 4D). Finally,b-catenin levels foundwithin themucin, presumably due tothe presence of infiltrating inflammatory cells, decreased inPMCA patients after antibiotic treatment (464.58 �139.418 Vvi/106 mm3) as compared to no antibiotics(2,179.70 � 303.38 Vvi/106 mm3; P < 0.0021). Conversely,DPAM did not show a significant difference in the mucindue to antibiotic treatment. As a result, there was a signif-icant difference (P < 0.005) in the overall b-catenin stainingof PMCA (A þ NoA) mucin (1,706.56 � 264.45 Vvi/106
mm3) as compared to DPAM (A þ NoA) mucin (433.14 �104.55 Vvi/106 mm3). Finally, although some change inb-catenin distribution was seen in the stromal region ofPMCA patients, these changes were not significant.
Given that b-catenin is known to enter the nucleus andfunction as a transcription factor, and because our initial
B
CA BTNCB
TNCB H. pylori
H. pylori
Merge
Merge
10 μm
10 μm10 μm 10 μm
10 μm10 μm
Figure 3. FISH analysis of bacterialdensity in mucin. Formalin-fixedtissue specimens frompseudomyxoma peritonei patientswere serially sectioned andanalyzed with FISH to identify allTNCB present (green; A) orspecifically H. pylori (red; B) withina region of pooled mucin. In themerged images (C), the H. pyloristaining converges with the TNCBstaining to reveal a yellow color forH. pylori whereas all non-H. pyloribacteria appear green. A clearreduction in bacterial density isobserved in a pseudomyxomaperitonei patient receivingantibiotic treatment (A–C, bottom)compared to an untreatedpseudomyxoma peritonei patient(A–C, top).
Pseudomyxoma Peritonei, Bacteria, and Antibiotics
www.aacrjournals.org Clin Cancer Res; 19(14) July 15, 2013 3971
staining did not allow us to discern which portion of thecytoplasmic b-catenin was actually in the nucleus, we useddual staining with DAPI and b-catenin along with a com-puterized quantification method to determine the fractionof the nucleus occupied by b-catenin. Control NNA tissueexhibited intense bright DAPI staining, but was negative fornuclear b-catenin (data not shown). Conversely, nuclearb-catenin was common in both PMCA and DPAM samples,as exemplified by the fluorescent staining of the nucleishown in Fig. 5. In the high magnification insets, brightred color signifies b-catenin expression (A, arrow), whereasdark blue DAPI staining is seen when nuclear b-catenin isabsent (B, arrowhead). When the images are merged (C),the intense red shows up without being affected by the blueDAPI stain. Of note, faint perinuclear b-catenin stainingis seen around most DAPI-stained nuclei, indicating that
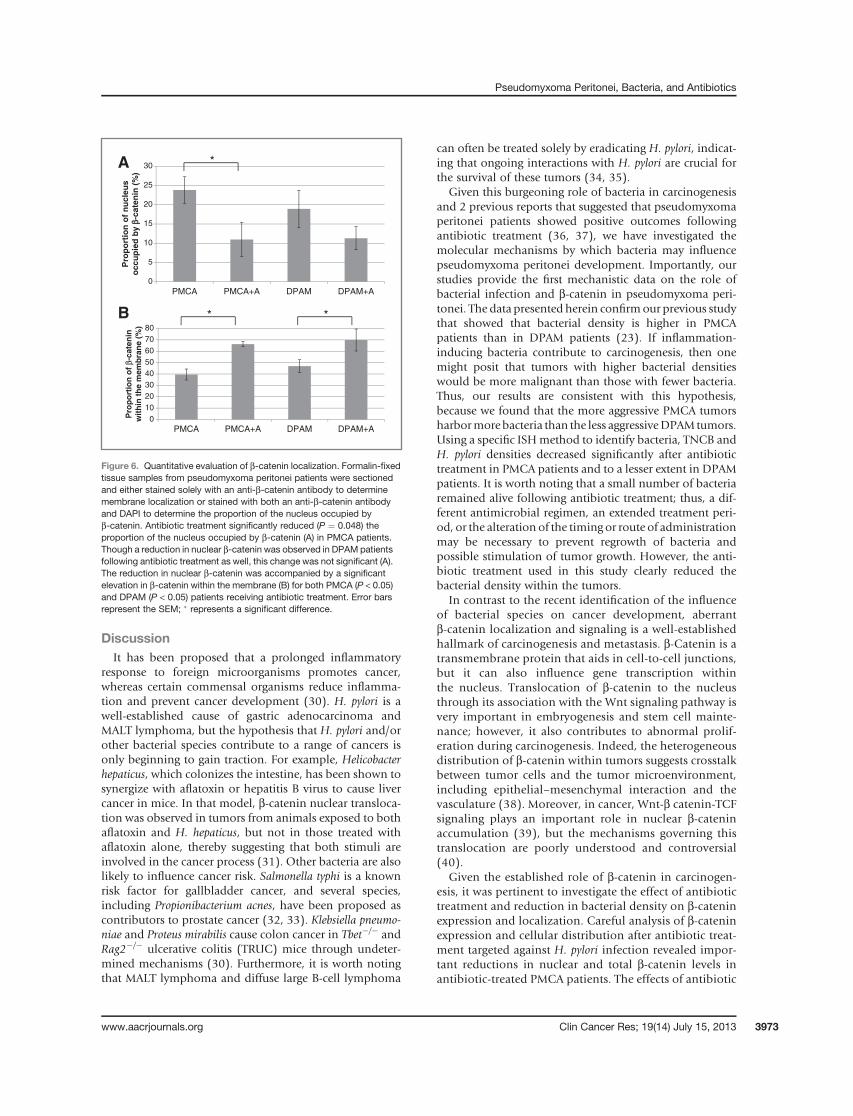
b-catenin was present elsewhere in those cells. The percentof DAPI-stained nuclei occupied by b-catenin was notsignificantly different when all PMCA samples (A þ NoA)were compared to DPAM samples (AþNoA) (20� 2.9 vs.16.5 � 3.5%, P < 0.451; NS). However, administration ofantibiotics resulted in a significant decrease in the percentof nuclear b-catenin present in PMCA (from 23.8 � 3.4%to 11 � 4.4%, P ¼ 0.048). Despite the fact that there was astrong trend toward decreased nuclear b-catenin, thissignificant difference was not seen with DPAM (from18.9 � 4.8% to 11.3 � 3%, P < 0.152; NS; Fig. 6A). Asnuclear b-catenin dropped as a result of antibiotic treat-ment, the percentage of b-catenin located in the mem-brane as compared to the cytoplasm increased (Fig. 6B).This rise in membrane b-catenin was significant for bothPMCA and DPAM patients.
A CB10 μm10 μm10 μm
Figure 5. Immunofluorescenceanalysis of nuclear b-catenin.Formalin-fixed tissue samplesfrom pseudomyxoma peritoneipatients were sectioned and dualstained with an anti-b-cateninantibody and DAPI to observe thelocalization of nuclear b-catenin.As shown in A, bright red staining(arrow) indicates the presence ofb-catenin, whereas B reveals thedark blue DAPI staining(arrowhead) appearing whenb-catenin staining is absent. Themerged image in C exhibits theintense red b-catenin stainingunaffected by blue DAPI staining.
A B C
10 μm
30um
30 μm
5 μm
5 μm
D
Figure 4. Qualitative assessment ofb-catenin localization inpseudomyxoma peritoneisamples. Formalin-fixed tissuesamples from pseudomyxomaperitonei patients and an NNAcontrol were sectioned andstained with an anti-b-cateninantibody. The NNA control (A)displayed strong staining at theboundaries between adjacentcells, showing the localization ofb-catenin to cell membranes andthe lateral junctional complex.Conversely, PMCA sections (B)exhibited primarily cytoplasmicstaining of b-catenin and virtuallyno staining at the intercellularboundaries. Some staining wasobserved between PMCAneoplastic cells following antibiotictreatment (C). b-Catenin was veryvisible in the desmoplasticreactions within the stromalcompartment of untreated PMCApatients (D).
Semino-Mora et al.
Clin Cancer Res; 19(14) July 15, 2013 Clinical Cancer Research3972
DiscussionIt has been proposed that a prolonged inflammatory
response to foreign microorganisms promotes cancer,whereas certain commensal organisms reduce inflamma-tion and prevent cancer development (30). H. pylori is awell-established cause of gastric adenocarcinoma andMALT lymphoma, but the hypothesis that H. pylori and/orother bacterial species contribute to a range of cancers isonly beginning to gain traction. For example, Helicobacterhepaticus, which colonizes the intestine, has been shown tosynergize with aflatoxin or hepatitis B virus to cause livercancer in mice. In that model, b-catenin nuclear transloca-tion was observed in tumors from animals exposed to bothaflatoxin and H. hepaticus, but not in those treated withaflatoxin alone, thereby suggesting that both stimuli areinvolved in the cancer process (31). Other bacteria are alsolikely to influence cancer risk. Salmonella typhi is a knownrisk factor for gallbladder cancer, and several species,including Propionibacterium acnes, have been proposed ascontributors to prostate cancer (32, 33). Klebsiella pneumo-niae and Proteus mirabilis cause colon cancer in Tbet�/� andRag2�/� ulcerative colitis (TRUC) mice through undeter-mined mechanisms (30). Furthermore, it is worth notingthat MALT lymphoma and diffuse large B-cell lymphoma
can often be treated solely by eradicating H. pylori, indicat-ing that ongoing interactions with H. pylori are crucial forthe survival of these tumors (34, 35).
Given this burgeoning role of bacteria in carcinogenesisand 2 previous reports that suggested that pseudomyxomaperitonei patients showed positive outcomes followingantibiotic treatment (36, 37), we have investigated themolecular mechanisms by which bacteria may influencepseudomyxoma peritonei development. Importantly, ourstudies provide the first mechanistic data on the role ofbacterial infection and b-catenin in pseudomyxoma peri-tonei. The data presented herein confirmour previous studythat showed that bacterial density is higher in PMCApatients than in DPAM patients (23). If inflammation-inducing bacteria contribute to carcinogenesis, then onemight posit that tumors with higher bacterial densitieswould be more malignant than those with fewer bacteria.Thus, our results are consistent with this hypothesis,because we found that the more aggressive PMCA tumorsharbormorebacteria than the less aggressiveDPAMtumors.Using a specific ISHmethod to identify bacteria, TNCB andH. pylori densities decreased significantly after antibiotictreatment in PMCA patients and to a lesser extent in DPAMpatients. It is worth noting that a small number of bacteriaremained alive following antibiotic treatment; thus, a dif-ferent antimicrobial regimen, an extended treatment peri-od, or the alteration of the timing or route of administrationmay be necessary to prevent regrowth of bacteria andpossible stimulation of tumor growth. However, the anti-biotic treatment used in this study clearly reduced thebacterial density within the tumors.
In contrast to the recent identification of the influenceof bacterial species on cancer development, aberrantb-catenin localization and signaling is a well-establishedhallmark of carcinogenesis and metastasis. b-Catenin is atransmembrane protein that aids in cell-to-cell junctions,but it can also influence gene transcription withinthe nucleus. Translocation of b-catenin to the nucleusthrough its association with the Wnt signaling pathway isvery important in embryogenesis and stem cell mainte-nance; however, it also contributes to abnormal prolif-eration during carcinogenesis. Indeed, the heterogeneousdistribution of b-catenin within tumors suggests crosstalkbetween tumor cells and the tumor microenvironment,including epithelial–mesenchymal interaction and thevasculature (38). Moreover, in cancer, Wnt-b catenin-TCFsignaling plays an important role in nuclear b-cateninaccumulation (39), but the mechanisms governing thistranslocation are poorly understood and controversial(40).
Given the established role of b-catenin in carcinogen-esis, it was pertinent to investigate the effect of antibiotictreatment and reduction in bacterial density on b-cateninexpression and localization. Careful analysis of b-cateninexpression and cellular distribution after antibiotic treat-ment targeted against H. pylori infection revealed impor-tant reductions in nuclear and total b-catenin levels inantibiotic-treated PMCA patients. The effects of antibiotic
0
5
10
15
20
25
30
DPAM+ADPAMPMCA+APMCA
01020304050607080
DPAM+ADPAMPMCA+APMCA
A
B
Pro
po
rtio
n o
f n
ucl
eus
occ
up
ied
by
ββ-ca
ten
in (
%)
Pro
po
rtio
n o
f β-
cate
nin
wit
hin
th
e m
emb
ran
e (%
)
*
**
Figure 6. Quantitative evaluation of b-catenin localization. Formalin-fixedtissue samples from pseudomyxoma peritonei patients were sectionedand either stained solely with an anti-b-catenin antibody to determinemembrane localization or stained with both an anti-b-catenin antibodyand DAPI to determine the proportion of the nucleus occupied byb-catenin. Antibiotic treatment significantly reduced (P ¼ 0.048) theproportion of the nucleus occupied by b-catenin (A) in PMCA patients.Though a reduction in nuclear b-catenin was observed in DPAM patientsfollowing antibiotic treatment as well, this change was not significant (A).The reduction in nuclear b-catenin was accompanied by a significantelevation in b-catenin within the membrane (B) for both PMCA (P < 0.05)and DPAM (P < 0.05) patients receiving antibiotic treatment. Error barsrepresent the SEM; � represents a significant difference.
Pseudomyxoma Peritonei, Bacteria, and Antibiotics
www.aacrjournals.org Clin Cancer Res; 19(14) July 15, 2013 3973

treatment reached statistical significance primarily inPMCA samples and not in DPAM samples, which couldbe the result of the small sample size (n ¼ 6) of antibiotictreated DPAM patients, making it more difficult to reachstatistical significance; similar trends were seen in bothgroups of antibiotic-treated patients. The levels and local-ization of b-catenin were also altered throughout thevarious cell types within the analyzed peritoneal tissue,where antibiotic treatment significantly decreased b-cate-nin expression in the cytoplasm of epithelial and inflam-matory cells, in the nuclei, and in the mucin of PMCApatients. Although not statistically significant, stromalb-catenin localization was elevated in PMCA patients nottreated with antibiotics. Increased levels of b-cateninwithin the stroma is associated with desmoplastic reac-tion in malignancy (Fig. 4D), where b-catenin is attachedto the fibrous elements of connective tissue of the stromalcompartment (41). The stromal compartment plays animportant role during neoplastic cell invasion, detachingfrom neoplastic intestinal glands, invading the stromalblood vessels and lymphatics, and initiating metastasis(42). b-Catenin within mucin was higher in PMCApatients as compared to DPAM patients. Unlike the stro-ma, antibiotic treatment had a significant effect ondecreasing b-catenin in the mucin. Because mucin is themost important histopathology hallmark of pseudomyx-oma peritonei, decreased levels of b-catenin within mucinafter antibiotic treatment suggests that antimicrobialsmight be useful in the treatment of this disease. Theeffectiveness of antibiotic treatment is further evidencedby the significant elevation of b-catenin levels within themembrane in both PMCA and DPAM patients accompa-nied by a reduction in nuclear b-catenin levels followingadministration of antibiotics. In normal cells, the b-cate-nin complex is present within the adherens junctions, asconfirmed by immunogold and transmission electronmicroscopy (43, 44). b-Catenin within the membranepromotes cell-to-cell communication and is likely toreduce neoplastic cell migration toward the stroma andmetastasis; thus, increased levels of b-catenin within themembrane could be helpful in improving the prognosisof pseudomyxoma peritonei disease. Moreover, the renor-malization of b-catenin distribution in PMCA patientsafter antibiotic treatment confirms the pivotal contribu-tion of bacteria to the pseudomyxoma peritonei diseaseprocess.
Targeting nuclear b-catenin is a potential strategy forcancer therapy. Other reports have discussed the benefit ofusing antimicrobial agents to treat cancer; however, mostattribute normalization of b-catenin with specific effects ofantimicrobial agents on the Wnt/b-catenin pathway unre-lated to thekilling ofmicroorganisms. For example, patientswith ovarian endometric adenocarcinoma receiving rapa-mycin had a demonstrated inhibition of the Wnt/b-cateninpathway along with a decrease in tumor burden (45).Rapamycin blocksWnt-mediated cell growth by interactingwithmTOR, which is a central regulator of cell growth (46).Similarly, the antibiotic Streptonigrin was reported to block
the complex formation of b-catenin/TCF (T-cell factor sig-naling)withDNA inhuman cell lines (47). In our study, thereduction of nuclear b-catenin following treatment withantimicrobial agents is likely due to bacterial killing,because, to our knowledge, no evidence indicates thatamoxicillin or clarithromycin influence the Wnt/b-cateninsignaling pathway. Of note, companion studies conductedby our group (J.J. Gilbreath et al., submitted for publica-tion) have defined the microbiome of pseudomyxomaperitonei tumors from 11 patients and shown the abilityto culture bacteria directly from pseudomyxoma peritoneitumor tissue. That study further indicated that antibioticadministration improves the survival of PMCA patientswithout pseudomyxoma peritonei observed in the lymphnodes when compared with untreated patients and histor-ical data. Antibiotic treatment did not improve the survivalof patients with lymph nodes positive for pseudomyxomaperitonei. Thus, treatment with antibiotics may be a ben-eficial therapy for pseudomyxoma peritonei patients withnonmetastatic disease.
We have not determined which mechanism is responsi-ble for the reduction of nuclear b-catenin following anti-biotic treatment or whether bacteria other than H. pyloricontribute to abnormal b-catenin localization. It is plausi-ble that the diminished b-catenin levels could be due to theloss of direct contact between tumor cells and bacteria orcaused by a reduced inflammatory response to bacteria. Infact, DeNardo and colleagues propose that chronic inflam-mation could lead to Wnt/b-catenin activation withoutspecific interactions between bacteria and epithelial cells(48). In this model, TNF-a produced by activated macro-phages binds to TNF receptors on nearby epithelial cells,leading to AKT phosphorylation and the subsequent stabi-lization of b-catenin.
Recent studies have revealed the influence of severalbacterial species on b-catenin expression and localization.H. pylori is present in the intercellular spaces, where thebacteria disrupt apical tight junctions as well as perturbmolecular expression of b-catenin and other proteins ofthe lateral cell-to-cell membrane (49, 50). Salmonellaactivates the Wnt/b-catenin pathway via the secretedeffector protein AvrA (21). Campylobacter rodentium, Bac-teroides fragilis, and Chlamydia trachomatis also influenceWnt/b-catenin signaling (15, 22). Given the effect ofbacteria on b-catenin signaling, which is known to con-tribute to the carcinogenic process, it is interesting to notethat many common cancers occur in anatomical regionsfrequently exposed to bacteria, including cancers of thethroat, lung, gastrointestinal tract, reproductive tract, andskin. Other locations, such as the breast, may seem toexist as sterile sites; however, bacteria can often gainentry, raising the possibility that individual bacterialspecies or a combination of different species could con-tribute to the carcinogenic process in multiple cancers.Thus, determining which bacterial species are involved inthese processes could lead to targeted antimicrobial ther-apy and help to improve current methods for preventingand treating cancer.
Semino-Mora et al.
Clin Cancer Res; 19(14) July 15, 2013 Clinical Cancer Research3974

Disclosure of Potential Conflicts of InterestNo potential conflicts of interest were disclosed.
Authors' ContributionsConception and design: C. Semino-Mora, T.L. Testerman, C. Nieroda, D.S.Merrell, A. DuboisDevelopment of methodology: C. Semino-Mora, T.J. McAvoy, C. NierodaAcquisitionofdata (provided animals, acquired andmanagedpatients,provided facilities, etc.): C. Semino-Mora, H. Liu, K. Studeman, Y. Jia, C.NierodaAnalysis and interpretation of data (e.g., statistical analysis, biosta-tistics, computational analysis): C. Semino-Mora, T.L. Testerman, H. Liu,T.J. McAvoy, D.S. Merrell, A. DuboisWriting, review, and/or revisionof themanuscript:C. Semino-Mora, T.L.Testerman, J.M. Whitmire, T.J. McAvoy, C. Nieroda, A. Sardi, D.S. Merrell, A.DuboisAdministrative, technical, or material support (i.e., reporting or orga-nizing data, constructing databases): H. Liu, T.J. McAvoy, J. FrancisStudy supervision: A. Sardi, D.S. Merrell, A. Dubois
AcknowledgmentsDuring the writing of this article, Dr. A. Dubois passed away unex-
pectedly. Although ill for a length of time, he did not want others to beconcerned and was silent about this fact. Throughout his illness, he
worked diligently and passionately on the pseudomyxoma peritoneiresearch. His wisdom, generosity, and expertise will be sorely missed byhis colleagues and the research community. In addition, Dr. C. Semino-Mora passed away after the submission of the original manuscript.Despite a lengthy illness, she continued to contribute her abilities toseveral research projects. Her extensive skills and expertise were a tre-mendous asset that cannot be replaced. The authors thank Dr. D.McDaniel for his assistance with laser confocal imaging along with M.Sittig and M. Putman for their contributions to this project.
Grant SupportThis work was supported by R0832L, which was funded by Uniformed
Services University, as well as funding from the US Military Cancer Institute.The study was approved by the Institutional Review Boards of the MercyMedical Center and the Uniformed Services University of the HealthSciences, and written informed consent was obtained from all patientsbefore study entry.
The costs of publication of this article were defrayed in part by thepayment of page charges. This article must therefore be hereby markedadvertisement in accordance with 18 U.S.C. Section 1734 solely to indicatethis fact.
Received March 7, 2013; revised May 10, 2013; accepted May 27, 2013;published OnlineFirst June 6, 2013.
References1. Bradley RF, Stewart JHt, Russell GB, Levine EA, Geisinger KR. Pseu-
domyxoma peritonei of appendiceal origin: a clinicopathologic anal-ysis of 101 patients uniformly treated at a single institution, withliterature review. Am J Surg Pathol 2006;30:551–9.
2. Mann WJ Jr, Wagner J, Chumas J, Chalas E. The management ofpseudomyxoma peritonei. Cancer 1990;66:1636–40.
3. Aho AJ, Heinonen R, Lauren P. Benign and malignant mucocele of theappendix. Histological types and prognosis. Acta Chir Scand 1973;139:392–400.
4. Smeenk RM, Bruin SC, van Velthuysen ML, Verwaal VJ. Pseudomyx-oma peritonei. Curr Probl Surg 2008;45:527–75.
5. WitkampAJ, deBreeE, VanGoethemR, Zoetmulder FA. Rationale andtechniques of intra-operative hyperthermic intraperitoneal chemother-apy. Cancer Treat Rev 2001;27:365–74.
6. Galani E, Marx GM, Steer CB, Culora G, Harper PG. Pseudomyxomaperitonei: the 'controversial' disease. Int J Gynecol Cancer 2003;13:413–8.
7. Loungnarath R, Causeret S, Brigand C, Gilly FN, Glehen O.[Pseudomyxoma peritonei: new concept and new therapeuticapproach]. Ann Chir 2005;130:63–9.
8. Levine EA, Stewart JHt, Russell GB, Geisinger KR, Loggie BL, Shen P.Cytoreductive surgery and intraperitoneal hyperthermic chemothera-py for peritoneal surfacemalignancy: experience with 501 procedures.J Am Coll Surg 2007;204:943–53; discussion 53-5.
9. GoughDB, Donohue JH, Schutt AJ,Gonchoroff N,Goellner JR,WilsonTO, et al. Pseudomyxomaperitonei. Long-term patient survival with anaggressive regional approach. Ann Surg 1994;219:112–9.
10. Halabi HE, Gushchin V, Francis J, Athas N, Macdonald R, Nieroda C,et al. Prognostic significance of lymph node metastases in patientswith high-grade appendiceal cancer. Ann Surg Oncol 2012;19:122–5.
11. Yan H, Pestieau SR, Shmookler BM, Sugarbaker PH. Histopathologicanalysis in 46patientswithpseudomyxomaperitonei syndrome: failureversus success with a second-look operation. Mod Pathol 2001;14:164–71.
12. Miller JR. The Wnts. Genome Biol 2001;3:REVIEWS3001-REVIEWS3001.15.
13. Hayashida Y, Honda K, IdogawaM, Ino Y, OnoM, Tsuchida A, et al. E-cadherin regulates the association between beta-catenin and actinin-4. Cancer Res 2005;65:8836–45.
14. Takayama T, Shiozaki H, Shibamoto S, Oka H, Kimura Y, Tamura S,et al. Beta-catenin expression in human cancers. Am J Pathol1996;148:39–46.
15. Sun J. Enteric bacteria and cancer stem cells. Cancers (Basel) 2010;3:285–97.
16. Hayward SD, Liu J, FujimuroM. Notch andWnt signaling: mimicry andmanipulation by gamma herpesviruses. Sci STKE 2006;2006:re4.
17. Zhang Y, Wei W, Cheng N, Wang K, Li B, Jiang X, et al. Hepatitis CVirus-induced upregulation of miR-155 promotes hepatocarcino-genesis by activating Wnt signaling. Hepatology 2012;56:1631–40.
18. El-Etr SH, Mueller A, Tompkins LS, Falkow S, Merrell DS. Phosphor-ylation-independent effects of CagA during interaction between Heli-cobacter pylori and T84 polarized monolayers. J Infect Dis 2004;190:1516–23.
19. Franco AT, Israel DA, Washington MK, Krishna U, Fox JG, Rogers AB,et al. Activation of beta-catenin by carcinogenic Helicobacter pylori.Proc Natl Acad Sci U S A 2005;102:10646–51.
20. Nakopoulou L, Mylona E, Papadaki I, Kavantzas N, Giannopoulou I,Markaki S, et al. Study of phospho-beta-catenin subcellular distribu-tion in invasive breast carcinomas in relation to their phenotype and theclinical outcome. Mod Pathol 2006;19:556–63.
21. Liu X, Lu R, Wu S, Sun J. Salmonella regulation of intestinal stemcells through the Wnt/beta-catenin pathway. FEBS Lett 2010;584:911–6.
22. Kessler M, Zielecki J, Thieck O, Mollenkopf HJ, Fotopoulou C, MeyerTF. Chlamydia trachomatis disturbs epithelial tissue homeostasis infallopian tubes via paracrine Wnt signaling. Am J Pathol 2012;180:186–98.
23. Semino-Mora C, Liu H, McAvoy T, Nieroda C, Studeman K, Sardi A,et al. Pseudomyxoma peritonei: is disease progression related tomicrobial agents? A study of bacteria, MUC2 AND MUC5AC expres-sion in disseminated peritoneal adenomucinosis and peritonealmucinous carcinomatosis. Ann Surg Oncol 2008;15:1414–23.
24. Ronnett BM, Zahn CM, Kurman RJ, Kass ME, Sugarbaker PH,Shmookler BM. Disseminated peritoneal adenomucinosis and perito-neal mucinous carcinomatosis. A clinicopathologic analysis of 109cases with emphasis on distinguishing pathologic features, site oforigin, prognosis, and relationship to "pseudomyxoma peritonei." AmJ Surg Pathol 1995;19:1390–408.
25. Liu H, Merrell DS, Semino-Mora C, Goldman M, Rahman A, Mog S,et al. Diet synergistically affects Helicobacter pylori-induced gastriccarcinogenesis in nonhuman primates. Gastroenterology 2009;137:1367–79 e1–6.
26. Liu H, Rahman A, Semino-Mora C, Doi SQ, Dubois A. Specific andsensitive detection ofH. pylori in biological specimens by real-timeRT-PCR and in situ hybridization. PLoS One 2008;3:e2689.
27. Semino-Mora C, Doi SQ, Marty A, Simko V, Carlstedt I, Dubois A.Intracellular and interstitial expression of Helicobacter pylori virulence
Pseudomyxoma Peritonei, Bacteria, and Antibiotics
www.aacrjournals.org Clin Cancer Res; 19(14) July 15, 2013 3975

genes in gastric precancerous intestinal metaplasia and adenocarci-noma. J Infect Dis 2003;187:1165–77.
28. Lan ZD, Dai L, Zhuo XL, Feng JC, Xu K, Chi CW. Gene cloning andsequencing of BmKASandBmKAS-1, two novel neurotoxins from thescorpion Buthus martensi Karsch. Toxicon 1999;37:815–23.
29. Dixon MF, Genta RM, Yardley JH, Correa P. Histological classificationof gastritis andHelicobacter pylori infection: an agreement at last? TheInternational Workshop on the Histopathology of Gastritis. Helicobac-ter 1997;2 Suppl 1: S17–24.
30. Trinchieri G. Cancer and inflammation: an old intuition with rapidlyevolving new concepts. Annu Rev Immunol 2012;30:677–706.
31. Martin HR, Shakya KP, Muthupalani S, Ge Z, Klei TR, Whary MT, et al.Brugia filariasis differentially modulates persistent Helicobacter pylorigastritis in the gerbil model. Microbes Infect 2010;12:748–58.
32. Samaras V, Rafailidis PI, Mourtzoukou EG, Peppas G, Falagas ME.Chronic bacterial and parasitic infections and cancer: a review. J InfectDev Ctries 2010;4:267–81.
33. Fassi Fehri L, Mak TN, Laube B, Brinkmann V, Ogilvie LA, MollenkopfH, et al. Prevalence of Propionibacterium acnes in diseased prostatesand its inflammatory and transforming activity on prostate epithelialcells. Int J Med Microbiol 2011;301:69–78.
34. Kuo SH, Yeh KH, Wu MS, Lin CW, Hsu PN, Wang HP, et al. Helico-bacter pylori eradication therapy is effective in the treatment of early-stage H. pylori-positive gastric diffuse large B-cell lymphomas. Blood2012;119:4838–44; quiz 5057.
35. Ferreri AJ, Govi S, Raderer M, Mule A, Andriani A, Caracciolo D, et al.Helicobacter pylorieradication as exclusive treatment for limited-stagegastric diffuse large B-cell lymphoma: results of a multicenter phase 2trial. Blood 2012;120:3858–60.
36. Creel NJ, Dart BWt. Goblet cell carcinoid and mucinous cystadenomain an interval appendectomy specimen. Am Surg 2012;78:E49–50.
37. Thompson MA, Ashton RW, Pitot HC. Mucinous appendiceal adeno-carcinoma presenting 5 years after appendectomy. Ann Intern Med2004;140:W33.
38. FisherM,HuangYS, Li X,McIverKS, ToukokiC, EichenbaumZ.Shr is abroad-spectrum surface receptor that contributes to adherence andvirulence in group A streptococcus. Infect Immun 2008;76:5006–15.
39. van de Wetering M, Sancho E, Verweij C, de Lau W, Oving I,Hurlstone A, et al. The beta-catenin/TCF-4 complex imposes a
crypt progenitor phenotype on colorectal cancer cells. Cell 2002;111:241–50.
40. WuX, Tu X, Joeng KS, HiltonMJ,Williams DA, Long F. Rac1 activationcontrols nuclear localization of beta-catenin during canonical Wntsignaling. Cell 2008;133:340–53.
41. Osunkoya AO, Netto GJ, Epstein JI. Colorectal adenocarcinomainvolving the prostate: report of 9 cases. Hum Pathol 2007;38:1836–41.
42. Karagiannis GS, Petraki C, Prassas I, Saraon P, Musrap N, Dimitro-manolakis A, et al. Proteomic signatures of the desmoplastic invasionfront reveal collagen type XII as a marker of myofibroblastic differen-tiation during colorectal cancer metastasis. Oncotarget 2012;3:267–85.
43. Aberle H, Schwartz H, Kemler R. Cadherin-catenin complex: proteininteractions and their implications for cadherin function. J Cell Bio-chem 1996;61:514–23.
44. Bierkamp C, Schwarz H, Huber O, Kemler R. Desmosomal localizationof beta-catenin in the skin of plakoglobin null-mutant mice. Develop-ment 1999;126:371–81.
45. Tanwar PS, Zhang L, Kaneko-Tarui T, Curley MD, Taketo MM, Rani P,et al. Mammalian target of rapamycin is a therapeutic target for murineovarian endometrioid adenocarcinomas with dysregulated Wnt/beta-catenin and PTEN. PLoS One 2011;6:e20715.
46. Inoki K, Ouyang H, Zhu T, Lindvall C, Wang Y, Zhang X, et al. TSC2integrates Wnt and energy signals via a coordinated phosphorylationby AMPK and GSK3 to regulate cell growth. Cell 2006;126:955–68.
47. Park S, Chun S. Streptonigrin inhibits beta-catenin/Tcf signaling andshows cytotoxicity in beta-catenin-activated cells. Biochim BiophysActa 2011;1810:1340–5.
48. DeNardo DG, Johansson M, Coussens LM. Inflaming gastrointestinaloncogenic programming. Cancer Cell 2008;14:7–9.
49. Necchi V, Candusso ME, Tava F, Luinetti O, Ventura U, Fiocca R, et al.Intracellular, intercellular, and stromal invasion of gastric mucosa,preneoplastic lesions, and cancer by Helicobacter pylori. Gastroen-terology 2007;132:1009–23.
50. Suzuki M, Mimuro H, Suzuki T, Park M, Yamamoto T, Sasakawa C.Interaction of CagA with Crk plays an important role in Helicobacterpylori-induced loss of gastric epithelial cell adhesion. J ExpMed 2005;202:1235–47.
Semino-Mora et al.
Clin Cancer Res; 19(14) July 15, 2013 Clinical Cancer Research3976








![€¦ · Web view2009. 4. 23. · [Cr2O72-] Reverse Rate. A. increases increases. B. increases decreases. C. decreases decreases. D. decreases increases. 31. A small amount of H2SO4](https://img.pdfslide.net/doc/110x75/608f2c47b9e3f5096f2e5efc/web-view-2009-4-23-cr2o72-reverse-rate-a-increases-increases-b-increases.jpg)




















