Embed Size (px)
Citation preview

of April 13, 2018.This information is current as Antilymphocyte Autoimmunity
Associated Hemolytic Anemia and Immunodeficiency-Like Condition withSpontaneous Common Variable Myeloid Glycosylation Defects Lead to a
Sean O. Ryan, Derek W. Abbott and Brian A. Cobb
ol.1400385http://www.jimmunol.org/content/early/2014/05/02/jimmun
published online 2 May 2014J Immunol
MaterialSupplementary
5.DCSupplementalhttp://www.jimmunol.org/content/suppl/2014/05/02/jimmunol.140038
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved.Copyright © 2014 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on April 13, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on April 13, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

The Journal of Immunology
Myeloid Glycosylation Defects Lead to a SpontaneousCommon Variable Immunodeficiency-Like Condition withAssociated Hemolytic Anemia and AntilymphocyteAutoimmunity
Sean O. Ryan, Derek W. Abbott, and Brian A. Cobb
Common variable immunodeficiency (CVID), the most frequent symptomatic primary immune deficiency in humans, is a hetero-
geneous group of immunologic disorders estimated to affect 1:10,000–1:50,000. Although a clear disease etiology remains elusive,
a common characteristic of CVID is deficient IgG Ab production in response to infection or vaccination. Patients often also exhibit
autoimmune cytopenias with symptoms of abnormal T cell function, including reductions in naive T cells, which correlate with
clinical severity. In this study, we discovered that targeted alterations in the glycome of the myeloid lineage lead to spontaneous
immunodeficiency characteristic of both humoral and T cell dysfunction regularly found in human CVID. Mice carrying a my-
eloid-specific knockout of the Mgat2 gene encoding UDP-GlcNAc:a-6-D-mannoside b-1,2-N-acetylglucosaminyltransferase II
enzyme exhibit deficiencies in IgG responses to both protein and polysaccharide conjugate vaccines. Interestingly, the immuno-
deficiency is associated with decreased T cell activity because of a persistent autoimmune-mediated depletion of naive T cells,
which is induced by changes in erythrocyte surface glycosylation. The N-glycosylation dependent autoepitopes that emerge on
erythrocytes lead to autoimmune hemolytic anemia, and the causative auto-IgM cross-reacts with naive T cells despite the lack of
glycan change on T cells. These findings demonstrate that alterations in erythrocyte glycosylation trigger the development of
autoantibodies directed at both erythrocytes and naive T cells, revealing a possible mechanistic link between the induction
of autoimmune hemolytic anemia, the reduction in naive T cells, and poor Ab responses to vaccine in severe CVID patients.
The Journal of Immunology, 2014, 192: 000–000.
Common variable immunodeficiency (CVID) is the mostcommonly diagnosed primary immunodeficiency (1, 2),likely because of the variety of clinical presentations,
although .90% of analyzed CVID patients lack a definitive ge-netic defect (2). A key characteristic of CVID is poor Ab pro-duction, especially IgG, in response to infection or immunization;however, many patients also exhibit symptoms of abnormal T cellfunction and immune dysregulation (3). The multifaceted natureof this disease coupled with the lack of solid genetics has led toa dearth of available mouse models to understand the manifes-tations and mechanisms underlying CVID (4). Moreover, the useof Ig replacement therapy (i.e., i.v. Ig [IVIg]) and antibiotics hasreduced the number of infections, yet the major inflamma-tory complications such as autoimmunity still develop in CVID
patients (5). As a result, the deficient Ig production common toCVID may not encompass the full mechanistic extent of the dis-order.A broader explanation may be found in multiple observations in
which T cell defects were found in a still undefined proportion ofhuman CVID patients. Some CVID patients have significantlydecreased T cell numbers, in particular naive CD4+ T cells (6, 7).In a recent clinical study of 313 CVID subjects, patients whodeveloped recurring infections but not autoimmunity showedmodest decreases in both class switched memory B cells and naiveCD4+ T cells, whereas patients with more severe sequelae such aschronic enteropathy showed a greater loss of these cells (8). Thesestudies reveal that the degree of reduction in naive CD4+ T cells iscorrelated with clinical disease severity and that this is indepen-dent of age, gender, and even Ig titer (7, 8). To date, the mecha-nisms underlying naive T cell deficiency have not been identified,although some have postulated that decreased thymic output couldaccount for the changes (7).In the CVID patient population, autoimmune diseases affect
∼20% of patients and are commonly the first clinical sign of im-munodeficiency (9). Among the various cytopenias, autoimmunehemolytic anemia (AIHA) and autoimmune thrombocytopenicpurpura are the most prevalent in CVID patients (10). Although thelinkage between CVID and these Ab-mediated disorders remainsa matter of investigation, it is particularly interesting to note thatCVID-associated cytopenia is inversely proportional to the numberof naive CD4+ T cells in circulation, raising the possibility that thespecific loss of naive T cells is mechanistically tied to the de-struction of platelets and/or erythrocytes.We recently described the creation of a novel mouse strain in
which we used the cre-lox recombination system to delete the
Department of Pathology, Case Western Reserve University School of Medicine,Cleveland, OH 44106
Received for publication February 10, 2014. Accepted for publication April 7, 2014.
This work was supported by National Institutes of Health Grants GM082916 andOD004225 (to B.A.C.). Tissue histology was performed by the Tissue ResourcesCore Facility of the Case Comprehensive Cancer Center, which is supported byNational Institutes of Health Grant P30 CA43703.
Address correspondence and reprint requests to Dr. Brian A. Cobb, Departmentof Pathology, Case Western Reserve University School of Medicine, 10900 EuclidAvenue, WRB 6532, Cleveland, OH 44106-7288. E-mail address: [email protected]
The online version of this article contains supplemental material.
Abbreviations used in this article: AIHA, autoimmune hemolytic anemia; cN-glycan,complex-type N-glycan; CVID, common variable immunodeficiency; GlcNAcT-II,b-1,2-N-acetylglucosaminyltransferase II; IVIg, i.v. Ig; PHA-L, Phaseolus vulgarisleucoagglutinin.
Copyright� 2014 by The American Association of Immunologists, Inc. 0022-1767/14/$16.00
www.jimmunol.org/cgi/doi/10.4049/jimmunol.1400385
Published May 2, 2014, doi:10.4049/jimmunol.1400385 by guest on A
pril 13, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

Mgat2 gene encoding UDP-GlcNAc:a-6-D-mannoside b-1,2-N-acetylglucosaminyltransferase II (GlcNAcT-II) within the myeloidlineage (11). These Mgat2DM/DM mice lack the ability to synthe-size complex-type N-glycans (cN-glycans) in myeloid cells andare hyporesponsive to T cell–dependent bacterial polysaccharideAgs because of defects in MHC class II–dependent presentation,although peptide responses remain normal and most of these micedo not show overt developmental or systemic defects. However,∼25% of these mice show a spontaneous and specific loss of CD4+
naive T cells despite the lack of change in T cell glycosylation.In this study, we describe the characterization of these sponta-
neous T cell–depleted Mgat2DM/DM mice to reveal a CVID-likecondition that incorporates not only the associated humoral re-sponse deficiencies seen in human patients but also the presence ofAIHA coupled with the selective loss of naive T cells. Our datashow that loss of naive T cells is independent of thymic devel-opment or cell-intrinsic Mgat2 activity, but is associated withsignificant autoantibody (IgM) deposition on T cells in circulation,indicating an Ab-mediated mechanism of cell depletion. Re-markably, we found that auto-IgM Abs arise in response toerythrocytes with altered glycosylation, yet cross-react with naiveT cells from both Mgat2DM/DM and wild type mice. The CVID-like phenotype could also be induced within the “normal”Mgat2DM/DM mice upon the adoptive transfer of glycan-alterederythrocytes. These findings demonstrate that changes in eryth-rocyte glycosylation can trigger IgM-mediated autoimmunity thatnot only leads to hemolytic anemia but also cross-reacts withnaive T cells, thereby depleting the helper capacity of the adaptiveimmune response and preventing robust IgG responses and classswitching upon vaccination. Through incorporation of hypogam-maglobulinemia, loss of naive T cells, and AIHA, our findingsraise the possibility that altered erythrocyte and/or platelet gly-cosylation may play an unexpected role in human CVID severity.
Materials and MethodsAnimals and common reagents
Animal colonies were maintained in a specific pathogen-free environmentat Case Western Reserve University and were treated under InstitutionalAnimal Care and Use Committee–approved guidelines in accordance withapproved protocols. Mgat2DM/DM mice were generated by crossing theMgat2 (B6.129-Mgat2tm1Jxm/J; stock 006892) and LyzM-Cre (B6.129P2-Lyz2tm1(cre)Ifo/J; stock 004781) parental strains. As reported by The JacksonLaboratory, the parental strains were backcrossed to C57BL/6 before in-breeding for nine generations (Mgat2) and at least six generations (LyzM-Cre), respectively. The Mgat2wt/wt mice used were either B6.129-Mgat2tm1Jxm/J mice or C57BL/6J mice (stock 000664), both of which havewild type Mgat2 activity. All mice were originally purchased from TheJackson Laboratory. Mouse genotypes were confirmed using The JacksonLaboratory PCR protocols. Cell culture was performed using RPMI 1640medium supplemented with 10% FBS, penicillin, streptomycin, L-gluta-mine, and 2-ME (Life Technologies).
Vaccinations
For protein vaccinations each mouse received an i.p. injection of 2 mgOVA (Sigma-Aldrich) adsorbed to 25 ml alum adjuvant (Alhydrogel 2%;InvivoGen) in a final volume of 100 ml, diluted in PBS. For polysaccharidevaccinations each mouse received an i.p. injection of 40 ml Prevnar-13(supplied by J. Schreiber, Tufts University, Boston, MA) diluted to 200 mlin PBS. Mice received a second dose at two weeks. Prevnar-13 contains4.4 mg/ml polysaccharide each from Streptococcus pneumoniae serotypes1, 3, 4, 5, 6A, 7F, 9V, 14, 18C, 19A, 19F, 23F, and 8.8 mg/ml from serotype6B. Mice were analyzed 10 d after the final dose.
Serum Ab detection
Anti-protein and anti-polysaccharide serum Abs were detected by indirectELISA as described previously (12, 13). Briefly, Microlon high bindingplates (Greiner Bio-One) were coated overnight at 4˚C with either10 mg/ml OVA (Sigma-Aldrich) or S. pneumoniae serotype 14 polysaccharide
(supplied by J. Schreiber) diluted in PBS. Serial dilutions of serum wereused to probe Ag-bound plates. Detection was performed using bio-tinylated anti-mouse IgG polyclonal Ab (Jackson ImmunoResearch Lab-oratories) and europium-conjugated streptavidin (PerkinElmer), followedby quantification by time-resolved fluorescence on a Victor3V MultilabelCounter using DELFIA Enhancement Solution, according to the manu-facturer’s protocol (PerkinElmer).
Flow cytometry
Flow cytometry was performed as described previously (11). Briefly, cellswere stained with Alexa Fluor 647–conjugated Phaseolus vulgaris leu-coagglutinin (PHA-L) lectin (Life Technologies) and/or the indicated Abs(BioLegend) for 30 min at 4˚C. FACS data were collected using an AccuriC6 flow cytometer (BD Biosciences). Analyses of FACS data were per-formed using FCS Express (De Novo Software).
In vitro T cell Ag recall assay
T cell recall assays were performed as described previously (14). CD4+
T cells were isolated from the spleen by CD4+ magnetic bead positiveselection (Miltenyi Biotec) and labeled with 2.5 mM CFSE (Life Tech-nologies). CD4+ T cells (1 3 105) were cocultured with T cell–depletedsplenocytes (1 3 105) and 50 mg/ml OVA (Sigma-Aldrich) or media alone.On day 3, culture supernatants were analyzed for IL-5 by sandwich ELISA,according to the manufacturer’s protocol (BioLegend). To measure prolif-eration, CFSE-labeled cells were collected and analyzed by flow cytometry.
Coombs test for autoantibody detection
For direct Coombs tests, cells were collected from blood or spleen andprobed directly with biotinylated anti-mouse IgM (Jackson Immuno-Research) and the indicated phenotype Abs for 30 min at 4˚C. For indirectCoombs tests, cells were first incubated at 4˚C for 2 h with the indicatedmouse serum diluted 1:30 in PBS before being probed with biotinylatedanti-mouse IgM and the indicated phenotype Abs. Cells were then washedand probed with AlexaFluor-488 conjugated streptavidin (JacksonImmunoResearch Laboratories) followed by analysis by flow cytometry.
Tissue histology and blood differentials
Whole spleens were collected from indicated mice and preserved in 10%buffered formalin (Fisher Scientific). Paraffin embedding, tissue sectioning,and Perls’ Prussian blue iron staining were performed by the TissueResources Core Facility of the Case Comprehensive Cancer Center(Cleveland, OH). Microscopy images were acquired using a Leica DM ILLED microscope and LAS V4.1 software (Leica Microsystems). Hema-tological differentials were performed using a HEMAVET analyzer (DrewScientific).
Complement-mediated lysis assay
Complement lysis assays used Low-Tox-M rabbit complement (CedarlaneLabs) following the manufacturer’s instructions. Blood containing PHA-L2
erythrocytes was collected from naive Mgat2DM/DM mice into an equalvolume of 2 mg/ml EDTA in PBS (pH 7.2). Following three washes withPBS at 1500 3 g erythrocyte numbers were determined. A total of 10 3106 erythrocytes were incubated at 4˚C for 2 h in 50 ml of the indicatedmouse serum diluted 1:30 in PBS. After two washes with PBS, erythro-cytes were suspended in 50 ml assay buffer (RPMI 1640 with 25 mMHEPES and 0.3% BSA). To each well an additional 50 ml was added ofeither assay buffer, 1:10 complement diluted in assay buffer, or water.After 1 h incubation at 37˚C, intact erythrocytes were removed by cen-trifugation. Hemolysis was quantified by measuring supernatant absor-bance at 405 nm to determine levels of hemoglobin release. Percent lysiswas calculated relative to the water and buffer alone controls.
Adoptive transfer
Cell labeling and adoptive cell transfer experiments were done as describedpreviously (15). Briefly, splenocytes collected from donor Mgat2wt/wt micewere labeled with 6 mM CFSE (Life Technologies) and transferred i.v. at20 3 106 per recipient Mgat2wt/wt mice or CVID-Mgat2DM/DM mice. Onday 4, spleens from recipient mice were analyzed by flow cytometry todetermine absolute numbers of donor cells based on CFSE fluorescence.
Serum Ab adsorption assay
Serum collected from the indicated mice was incubated with erythrocytesfrom naive Mgat2DM/DM mice that lack the CVID-like phenotype for 2 h onice to adsorb auto-IgM Abs. This preadsorbed serum was collected and
2 MYELOID GLYCAN–INDUCED CVID
by guest on April 13, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
incubated with freshly isolated Mgat2wt/wt T cells for 2 h on ice or freshMgat2DM/DM erythrocytes to confirm complete auto-IgM removal. Cellswere then collected and surface-bound IgM was detected using bio-tinylated anti-mouse IgM and Alexa Fluor 488–conjugated streptavidin.T cells were labeled with anti-CD4 and anti-CD44 Abs. Cells were thenanalyzed by flow cytometry.
Erythrocyte immunizations
Blood containing PHA-L2 erythrocytes was collected from naiveMgat2DM/DM mice into an equal volume of 2 mg/ml EDTA in PBS (pH7.2). Following three washes with PBS at 1500 3 g erythrocyte numberswere determined and then subjected to hypotonic lysis using cold buffercontaining 5 mM Na2PO4 and 1 mM EDTA (pH 8). Membranes werewashed approximately five times with lysis buffer at 14,000 3 g or untilresidual hemoglobin was not detected in the supernatant as determined byabsorbance at 405 nm. Membrane preparations were suspended at 2.5 3108 input erythrocytes per ml PBS. Each mouse received 200 ml membranepreparation adsorbed to 100 ml alum adjuvant (Alhydrogel 2%; InvivoGen)by i.p. injection followed by a second injection at 2 wk. Mice were ana-lyzed five days after the final injection.
Statistics
Statistical comparisons were performed using a Student t test or ANOVAfor significance with a 95% confidence interval using GraphPad Instatsoftware. A p value , 0.05 was considered significant.
ResultsMyeloid-associated deletion of Mgat2 inducesa hypogammaglobulin state
During our routine characterization of the novel Mgat2DM/DM
mouse line, described recently (11), we examined humoral and
cellular immune responses induced by vaccination. Mgat2DM/DM
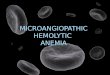
mice or Mgat2wt/wt mice were primed with alum-adsorbed OVA.Although a majority of the Mgat2DM/DM responded normally, a pop-ulation of these mice was severely IgG hyporesponsive as comparedwith both Mgat2wt/wt and most Mgat2DM/DM littermates (Fig. 1A).Using the Ab response to segregate Mgat2DM/DM mice into normaland low IgG groups, we could not detect induction of Ag-specificCD4+ T cell recall responses, measured by cell proliferation (Fig. 1B)and cytokine production (Fig. 1C), in low IgG Mgat2DM/DM mice ascompared with normal IgG Mgat2DM/DM and Mgat2wt/wt mice.A retrospective analysis of data collected prevaccination showed
a clear correlation between abnormal CD4+ T cell distribution inthe blood and vaccine failure. Both Mgat2wt/wt and normal IgGMgat2DM/DM mice exhibited a “naive” phenotype prevaccination,where CD4+ T cells expressing the naive CD44low surface phe-notype were on average at a .10:1 ratio to T cells expressing theeffector/memory CD44hi surface phenotype (Fig. 1D). However,low IgG Mgat2DM/DM mice exhibited a significantly skewed , 1:1ratio (Fig. 1D), indicating that a spontaneous pre-existing condi-tion had developed in these mice prior to immunization.The combination of IgG hyporesponsiveness and T cell dys-
function are characteristics of CVID in humans, with the lattershowing a strong association with disease severity (7). Becausea common diagnostic of CVID patients is hyporesponsiveness topolysaccharide-conjugate vaccines, we further tested our hypoth-esis that Mgat2DM/DM mice develop a spontaneous CVID-likecondition by examining responses to the clinically used Prevnar-13 vaccine. Prevnar-13 is a 13-valent capsular polysaccharide
FIGURE 1. Myeloid-associated deletion of Mgat2 induces a spontaneous CVID-like condition. Mgat2wt/wt and Mgat2DM/DM mice were immunized with
OVA and alum. (A) Serum anti-ova IgG end-point titers identified a subset of Mgat2DM/DM mice as low IgG responders (N). These low IgG responders did
not induce detectable T cell responses to OVA as measured by cell proliferation (B) and IL-5 production (C). (D) Flow cytometry analysis of blood CD4+
T cells prior to vaccination showing the ratio of naive (CD44low) to effector/memory (CD44hi) cells. (E) The CD4+ T cell ratio phenotype was used to
predict low IgG responders to Prevnar-13 vaccination and shown as induced antipolysaccharide IgG levels by ELISA at 1:50 serum dilution. (Eu, euro-
pium). *p , 0.05, **p , 0.01.
The Journal of Immunology 3
by guest on April 13, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

conjugate vaccine designed to induce protective Ab-mediatedimmunity against pneumococcal infection (16). Mgat2DM/DM
mice predicted to have the potential CVID-like condition (CVID-Mgat2DM/DM) were identified prevaccination based on a skewedT cell phenotype in the blood as shown in Fig. 1D. As in non-vaccinated mice, humoral IgG responses were undetectable afterPrevnar-13 vaccination in CVID-Mgat2DM/DM mice, whereasresponses in “naıve” Mgat2DM/DM mice were induced to a similarextent as control Mgat2wt/wt mice (Fig. 1E).
The abnormal T cell phenotype in CVID-Mgat2DM/DM mice ismaintained with age and independent of changes in T cellsurface glycosylation
Using the distinct T cell distribution in the blood to identifyCVID-Mgat2DM/DM mice, we determined disease penetrance tobe ∼25% of all Mgat2DM/DM mice (Table I). In addition, we foundthat the phenotype was maintained with age without any detectableconversion between the naive and CVID-like phenotype (Fig. 2A).We next examined whether the Mgat2 conditional deletion was
unexpectedly modifying T cells from Mgat2DM/DM mice and thuscontributing to the spontaneous immune phenotype. Cell surfacelabeling with the lectin PHA-L, which preferentially binds cN-glycans and thus can differentiate cells lacking Mgat2 (11, 14),showed no difference in T cell (CD3+) surface glycosylation(Fig. 2B). Alternatively, neutrophils (Gr1+CD11b+), which are knownto actively express lysozyme M and thus the focus of most studiesusing the LyzM-Cre system, showed the expected decrease in cellsurface cN-glycans indicated by decreased PHA-L binding (Fig. 2C).
CVID-Mgat2DM/DM mice exhibit autoimmune hemolytic anemia
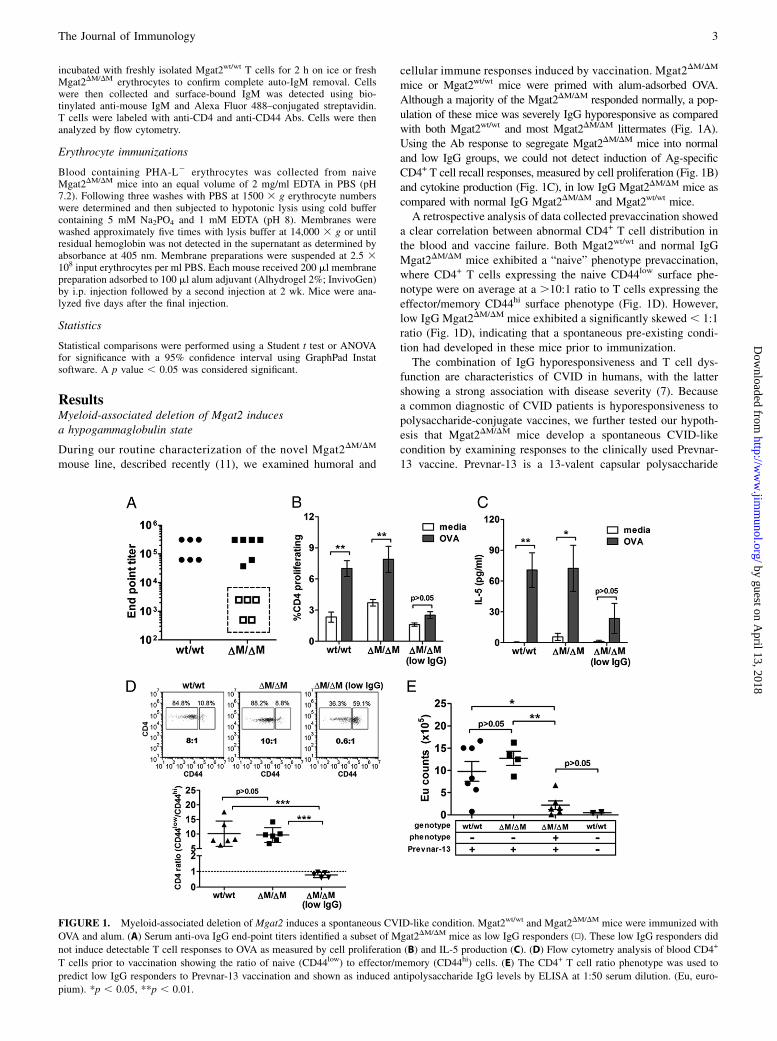
In addition to poor IgG responses and a loss of naive T cells, humanCVID is also commonly linked to autoimmune cytopenias. Toevaluate this in our model, we performed standard hematolog-ical analyses and found that the CVID-Mgat2DM/DM mice weresomewhat anemic (Fig. 3A). We had shown previously that thehematological profile of naive Mgat2DM/DM mice was similar toMgat2wt/wt mice (11). Upon further analysis of the blood, wefound a clear population of circulating erythrocytes that failedto bind PHA-L, indicating the lack of cN-glycans, as comparedwith the uniformly PHA-L+ erythrocytes found in Mgat2wt/wt mice(Fig. 3B). Although lysozyme M expression is not commonlyassociated with the erythroid lineage, the PHA-L binding patternindicated the loss ofMgat2 at some point during cell development.We found that while PHA-L- erythrocytes are a significant
proportion of circulating erythrocytes in naive Mgat2DM/DM mice,the number of PHA-L2 erythrocytes in CVID-Mgat2DM/DM micewas much lower (Fig. 3B). Direct Coombs tests showed thatCVID-Mgat2DM/DM mice, but not naive Mgat2DM/DM littermates,had significantly elevated levels of auto-IgM Ab coating eryth-rocytes, suggesting an antierythrocyte response (Fig. 3C). PHA-L+
erythrocytes did not exhibit significant auto-IgM Ab depositionbetween Mgat2DM/DM groups (p . 0.05), which were similar tocontrol Mgat2wt/wt erythrocytes (Fig. 3D), but rather it was thePHA-L2 erythrocytes in CVID-Mgat2DM/DM mice that were the
targets of the autoreactive Abs (Fig. 3E). Further analysis foundthat the autoantibody was functional and could mediate comple-ment lysis (Fig. 3F). Finally, histological analysis showed in-creased iron deposition in the splenic red pulp of only CVID-Mgat2DM/DM mice (Fig. 3G), indicating heightened clearance ofopsonized erythrocytes by resident macrophages. Taken together,these data demonstrate that the spontaneous CVID-like phenotypeis also associated with autoimmune hemolytic anemia, similar tomany human patients (17), and that this autoimmunity was di-rected at erythrocytes carrying altered glycans at the cell surface.
CVID-Mgat2DM/DM mice show selective naive T celllymphopenia
On the basis of the active depletion of erythrocytes from CVID-Mgat2DM/DM mice, we postulated that the skewed T cell pheno-type (Fig. 1D) may also be due to cell depletion. Indeed, uponcloser examination, we found that in CVID-Mgat2DM/DM mice,overall CD4+ and CD8+ T cell numbers in the blood (Fig. 4A) andspleen (Fig. 4B) were significantly decreased. Like human CVIDpatients, the observed lymphopenia was focused selectively on thenaive T cell population (CD44low; Fig. 4C, 4D), whereas effector/memory T cell numbers were similar if not slightly increased(CD44hi; Fig. 4E, 4F). On the basis of these data, the skewedT cell phenotype ratio (Fig. 1D) appears to be due at least in partto decreased numbers of naive T cells.A possible rationale for decreased naive T cell numbers in the
periphery could be related to decreases in thymic output of newnaive T cells. However, overall thymocyte numbers (Fig. 4G) andthe relative distribution of cells in the CD4/CD8 double-negative,double-positive, and single-positive stages of development werenormal (Fig. 4H). This indicated that the loss of naive T cells wasindependent of thymic development and suggests that the processis actively occurring in the periphery.
Naive T cells from CVID-Mgat2DM/DM mice are labeled withauto-IgM
On the basis of the presentation of autoimmune hemolytic anemia,we next sought to determine whether the selective absence of naive
Table I. Incidence of spontaneous CVID-like phenotype
Phenotype Distribution
No. of Naive No. of CVID Percentage of CVID
Mgat2wt/wt 19 0 0Mgat2DM/DM 218 74 25.3
All Mgat2wt/wt mice (n = 19) and Mgat2DM/DM mice (n = 292) were phenotypedbased on CD4 T cell distribution in the blood as shown in Fig. 1D.
FIGURE 2. The abnormal T cell distribution in CVID-Mgat2DM/DM
mice is maintained with age and independent of changes in T cell surface
glycosylation. (A) Representative analysis of blood CD4+ T cell phenotype
from the same naive Mgat2DM/DM or CVID-Mgat2DM/DM mouse at 12 and
27 wk of age. Comparison between T cell (CD3+) (B) and neutrophil (Gr1+
CD11b+) cell (C) glycosylation profiles from Mgat2wt/wt, naive Mgat2DM/DM,
and CVID-Mgat2DM/DM mice using flow cytometry and fluorescently labeled
PHA-L lectin. *p , 0.05.
4 MYELOID GLYCAN–INDUCED CVID
by guest on April 13, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
T cells in CVID-Mgat2DM/DM mice was due to an ongoing anti-lymphocyte autoimmune response. We found significant levels ofauto-IgM deposition on total CD4+ T cells but not CD8+ T cells(Fig. 5A). Closer examination of the naive (CD44low) and effector/memory (CD44hi) T cell subsets show that, as predicted, naiveCD4+ T cells (Fig. 5B), and to a lesser extent naive CD8+ T cells(Fig. 5B) from CVID-Mgat2DM/DM mice were the targets of theauto-IgM Ab. There was no significant Ab deposition on effector/memory T cells as compared with Mgat2wt/wt mice (Fig. 5C).
Naive T cell lymphopenia in CVID-Mgat2DM/DM mice is notdependent on T cell genotype
Although there was no indication of Mgat2 deletion in Mgat2DM/DM
T cells (Fig. 2), we wanted to confirm that the loss of naiveT cells was independent of cell-intrinsic changes because of theMgat2DM/DM genotype and thymic development. To this end, wetransferred bulk splenocytes from donor Mgat2wt/wt mice to re-cipient CVID-Mgat2DM/DM mice or control Mgat2wt/wt mice.Three days after transfer, the total number of donor CD4+ andCD8+ T cells remaining in recipient CVID-Mgat2DM/DM micewere significantly decreased compared with recipient Mgat2wt/wt
mice (Fig. 6A). As an internal control, the number of donorCD11b+ splenocytes was also determined (Fig. 6A). These mye-loid cells were found at equal levels in both recipient groups in-dicating that the wild-type T cells were being selectively depleted.
Interestingly, we found that decreased T cell numbers was againbecause of a selective depletion of naive CD4+ and CD8+ T cellsbut not the effector/memory T cell populations (Fig. 6B). Thesefindings confirm that the mechanism creating T cell lymphopeniain CVID-Mgat2DM/DM mice is an ongoing autoimmune responseagainst naive T cells.
The autoantibody response to naive T cells shares commonepitope(s) with erythrocytes expressing the alteredglycophenotype
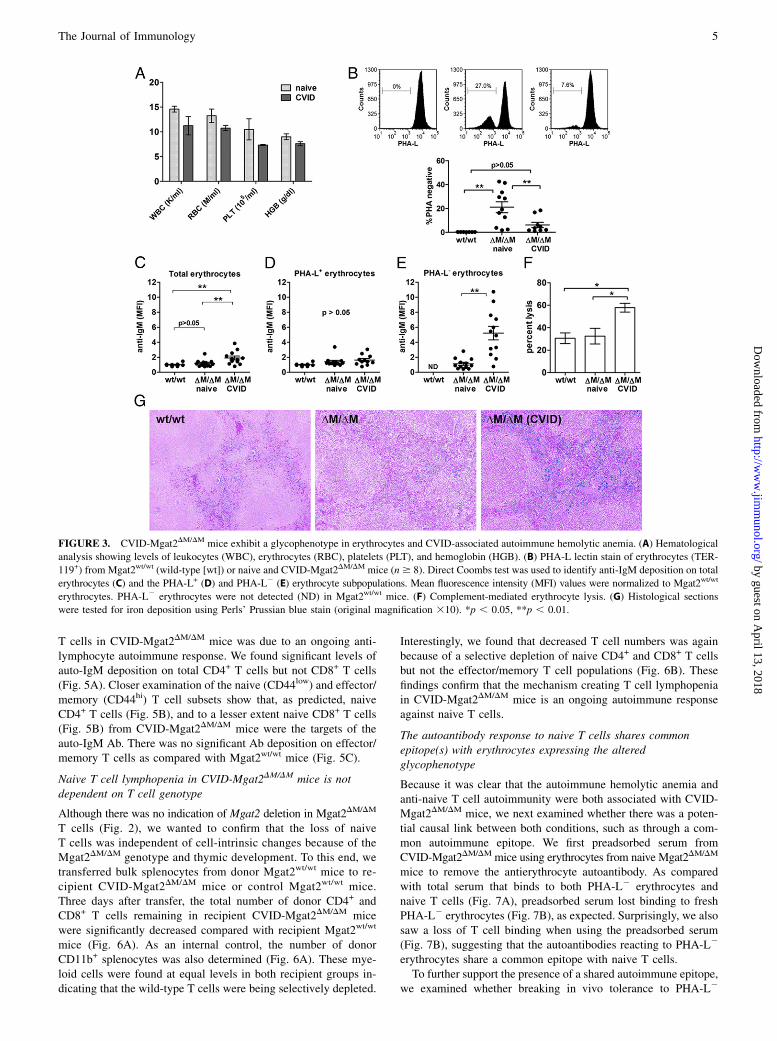
Because it was clear that the autoimmune hemolytic anemia andanti-naive T cell autoimmunity were both associated with CVID-Mgat2DM/DM mice, we next examined whether there was a poten-tial causal link between both conditions, such as through a com-mon autoimmune epitope. We first preadsorbed serum fromCVID-Mgat2DM/DMmice using erythrocytes from naive Mgat2DM/DM
mice to remove the antierythrocyte autoantibody. As comparedwith total serum that binds to both PHA-L2 erythrocytes andnaive T cells (Fig. 7A), preadsorbed serum lost binding to freshPHA-L2 erythrocytes (Fig. 7B), as expected. Surprisingly, we alsosaw a loss of T cell binding when using the preadsorbed serum(Fig. 7B), suggesting that the autoantibodies reacting to PHA-L2
erythrocytes share a common epitope with naive T cells.To further support the presence of a shared autoimmune epitope,
we examined whether breaking in vivo tolerance to PHA-L2
FIGURE 3. CVID-Mgat2DM/DM mice exhibit a glycophenotype in erythrocytes and CVID-associated autoimmune hemolytic anemia. (A) Hematological
analysis showing levels of leukocytes (WBC), erythrocytes (RBC), platelets (PLT), and hemoglobin (HGB). (B) PHA-L lectin stain of erythrocytes (TER-
119+) from Mgat2wt/wt (wild-type [wt]) or naive and CVID-Mgat2DM/DM mice (n$ 8). Direct Coombs test was used to identify anti-IgM deposition on total
erythrocytes (C) and the PHA-L+ (D) and PHA-L2 (E) erythrocyte subpopulations. Mean fluorescence intensity (MFI) values were normalized to Mgat2wt/wt
erythrocytes. PHA-L2 erythrocytes were not detected (ND) in Mgat2wt/wt mice. (F) Complement-mediated erythrocyte lysis. (G) Histological sections
were tested for iron deposition using Perls’ Prussian blue stain (original magnification 310). *p , 0.05, **p , 0.01.
The Journal of Immunology 5
by guest on April 13, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

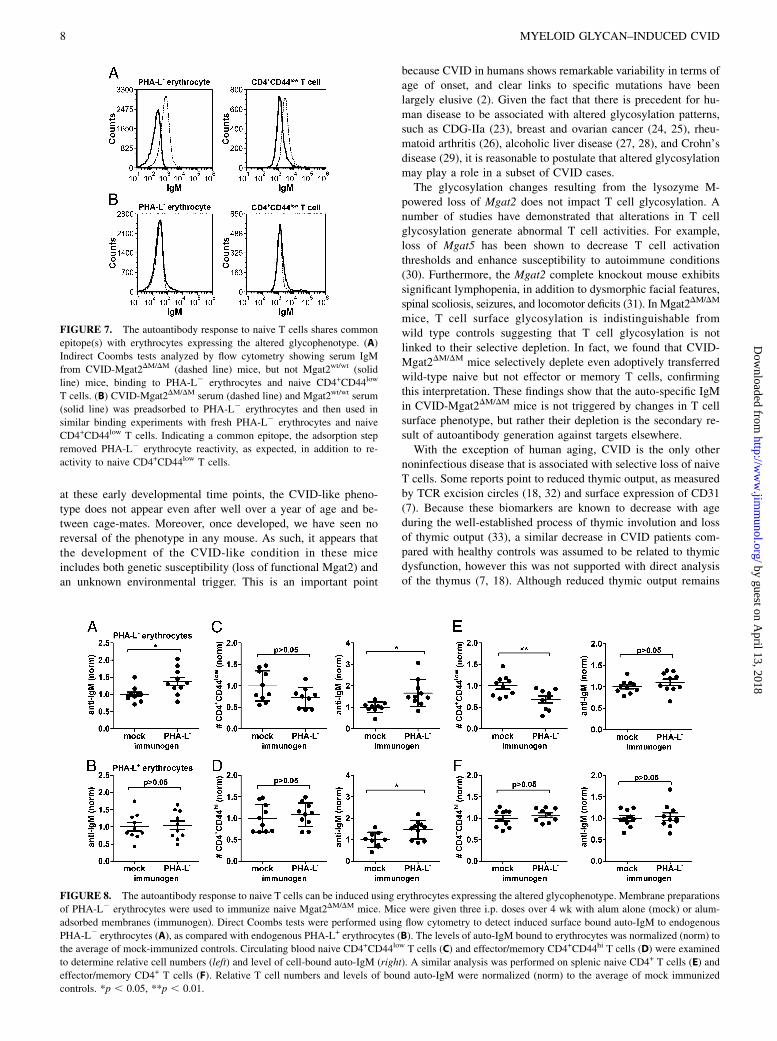
erythrocytes in naive Mgat2DM/DM mice could induce the CVID-like phenotype. Mice were immunized with membrane prepara-tions of PHA-L2 erythrocytes adsorbed to alum adjuvant or givenalum alone. Although fold reactivity was lower than that seen inspontaneous CVID-Mgat2DM/DM mice, PHA-L2 membrane prep-arations did induce a significant autoantibody response againstendogenous PHA-L2 erythrocytes (Fig. 8A) yet did not cross-react with the circulating PHA-L+ erythrocytes (Fig. 8B). Fur-ther indicating a common epitope shared by PHA-L2 erythrocytesand naive T cells, we found significant induction of autoantibodydeposition on circulating naive CD4+ T cells that correlated witha trend of decreased cell numbers (Fig. 8C). As predicted thenumbers of effector/memory CD4+ T cells were unchanged afterimmunization, however we did detect some autoantibody on thesecells (Fig. 8D). Unlike the blood, where naive T cell numbers onlytrended lower, we did see a significant reduction in their numbersin the spleen of PHA-L2 erythrocyte immunized mice (Fig. 8E).Similar to CVID-Mgat2DM/DM mice, there was no significantchange in effector/memory T cell numbers (Fig. 8F). Interestingly,there was an inverse relationship between detection of significant
autoantibody deposition in the blood and significant decreases innaive T cell numbers found in the spleen, suggesting that theopsonized cells in circulation had yet to be cleared from thesystem. Taken together, our findings indicate that the spontaneousCVID-like condition is directly linked to an autoantibody responseto naive T cells that shares common epitope(s) with erythrocytesexpressing the altered glycophenotype.
DiscussionCVID is a highly heterogeneous disease that is historically cate-gorized by defects in humoral immunity and B cells; however, it isestimated that .30% of the patients also have decreases in ab-solute T cell numbers, which is due to a selective loss of naiveT cells (6, 18). This phenotype has been suggested to indicatea distinct category of CVID, and it perhaps represents the mostclinically relevant because of the strong correlation to diseaseseverity (7). Moreover, it is not uncommon for severe CVID toalso correlate with autoimmune cytopenias, such as autoimmunethrombocytopenic purpura and AIHA (10). In this study, we foundthat changes in erythrocyte glycosylation can lead to the unmasking
FIGURE 4. The skewed T cell distribution identifying CVID-Mgat2DM/DM mice is due to selective naive T cell lymphopenia. Mgat2wt/wt or naive and
CVID-Mgat2DM/DM mice were used to determine total CD4+ and CD8+ T cell counts in the blood (A) and the spleen (B). Naive (CD44low) CD4+ and CD8+
T cell counts in the blood (C) and the spleen (D). Effector/memory (CD44hi) CD4+ and CD8+ T cell counts in the blood (E) and the spleen (F). Total
thymocyte counts (G) and the stages (H) of thymocyte development based on CD4 and CD8 expression were similar suggesting that loss of naive T cells is
not due to thymic dysfunction in CVID-Mgat2DM/DM mice. *p , 0.05, **p , 0.01, ***p , 0.001.
6 MYELOID GLYCAN–INDUCED CVID
by guest on April 13, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

of an autoepitope shared specifically with naive but not memory oreffector T cells. The auto-specific IgM produced leads to the se-lective depletion of both erythrocytes (AIHA) and naive T cells. Asa result of the loss of naive T cells, IgG responses to traditionalprotein and polysaccharide-conjugate vaccines are highly muted.These findings closely mirror a common cluster of phenotypespresent in some patients, suggesting that abnormal erythrocyteglycosylation arising from environmental and/or genetic circum-stances may contribute to human CVID onset and severity.We recently described the creation of the Mgat2DM/DM mouse
used in these studies (11). The goal underlying that study was to
understand the in vivo impact of N-linked glycosylation changesupon the MHC class II–mediated presentation of zwitterioniccapsular polysaccharide Ags (glycoantigens) to CD4+ T cells byaltering glycosylation in professional APCs (excluding B cells).As we reported, the mice used in that study were from the ∼75%of mice from the colony that did not show the autoimmune phe-notype. These mice had distinct defects in MHC class II–depen-dent presentation of glycoantigen but not peptide Ag (11). Noother defects were seen in any tissue or blood chemistry measures.In this study, we have focused on the ∼25% of the colony thatdevelops spontaneous loss of naive T cells and anemia.Although initially unexpected, expression of lysozyme M within
myeloid stem cell progenitors has been reported previously (19),providing a rationale for why Mgat2DM/DM mice carry as many as50% of the circulating erythrocytes with a PHA-L-low phenotype.The change in erythrocyte glycophenotype was not independentlysufficient to induce anemia or loss of naive T cells in everyMgat2DM/DM mouse. This incomplete disease penetrance inMgat2DM/DM mice suggests that the etiology likely has epigeneticorigins driven by the microflora. For example, IL-10 knockout (IL-102/2) mice are a common model for spontaneous inflammatorybowel disease; however, disease penetrance is highly sensitive tothe local environment. IL-102/2 mice housed in full barrier spe-cific pathogen-free facilities tend to show incomplete penetranceand mild disease compared with mice in vivariums without afull barrier (20). Furthermore, we detect very little switched(i.e., induced) IgG in nonmanipulated mice (wild-type and oth-erwise). Others have reported serum concentrations of IgG to be∼10 times higher (∼2–3 mg/ml) (21, 22) than we detect (∼0.25mg/ml) (Supplemental Fig. 1). However, when challenged witha generic microbial stimulus such as heat-killed cecal contents,total IgG levels in wild-type control mice were significantly in-creased over that which was induced in CVID-Mgat2DM/DM mice(Supplemental Fig. 1), and thus like CVID patients, we wouldexpect the CVID-Mgat2DM/DM mice to be more susceptible toinfection in a nonspecific pathogen-free environment. Along withthe lack of switched IgG induced by vaccination, we also foundlow B cell numbers in the spleen (Supplemental Fig. 1). In con-trast, the number of B cells in the blood was similar to controlanimals (Supplemental Fig. 1), which is a phenomenon commonlyseen in CVID patients. This is intriguing because it presents aninteresting dichotomy in the mouse that may be missed in humanswhere measurements are most often taken from the blood ratherthan a biopsy from the spleen.The trigger to become CVID-like or not in Mgat2DM/DM mice
appears to occur sometime before 5 wk of age. If it does not occur
FIGURE 5. Naive T cells from CVID-Mgat2DM/DM mice are labeled
with auto-IgM Ab. Direct Coombs tests were done to detect auto-IgM
adsorbed to total CD4+ and CD8+ T cells (A), naive (CD44low) CD4+ and
CD8+ T cells (B), and effector/memory (CD44hi) CD4+ and CD8+ T cells
(C) in the blood. *p , 0.05, **p , 0.01.
FIGURE 6. Naive T cell lymphopenia
in CVID-Mgat2DM/DM mice is not de-
pendent on T cell genotype. Donor
Mgat2wt/wt splenocytes were labeled with
CFSE and transferred into either recipi-
ent Mgat2wt/wt mice (wild-type [wt]) or
CVID-Mgat2DM/DM mice. After 3 d, re-
cipient mice were analyzed to determine
the number of remaining donor (CFSE+)
CD4+ and CD8+ T cells (A) and CD11b+
myeloid cells and naive T cells (top,
CD44low) versus effector/memory T
cells (bottom, CD44hi) (B). **p , 0.01,
***p , 0.001.
The Journal of Immunology 7
by guest on April 13, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
at these early developmental time points, the CVID-like pheno-type does not appear even after well over a year of age and be-tween cage-mates. Moreover, once developed, we have seen noreversal of the phenotype in any mouse. As such, it appears thatthe development of the CVID-like condition in these miceincludes both genetic susceptibility (loss of functional Mgat2) andan unknown environmental trigger. This is an important point
because CVID in humans shows remarkable variability in terms ofage of onset, and clear links to specific mutations have beenlargely elusive (2). Given the fact that there is precedent for hu-man disease to be associated with altered glycosylation patterns,such as CDG-IIa (23), breast and ovarian cancer (24, 25), rheu-matoid arthritis (26), alcoholic liver disease (27, 28), and Crohn’sdisease (29), it is reasonable to postulate that altered glycosylationmay play a role in a subset of CVID cases.The glycosylation changes resulting from the lysozyme M-
powered loss of Mgat2 does not impact T cell glycosylation. Anumber of studies have demonstrated that alterations in T cellglycosylation generate abnormal T cell activities. For example,loss of Mgat5 has been shown to decrease T cell activationthresholds and enhance susceptibility to autoimmune conditions(30). Furthermore, the Mgat2 complete knockout mouse exhibitssignificant lymphopenia, in addition to dysmorphic facial features,spinal scoliosis, seizures, and locomotor deficits (31). In Mgat2DM/DM
mice, T cell surface glycosylation is indistinguishable fromwild type controls suggesting that T cell glycosylation is notlinked to their selective depletion. In fact, we found that CVID-Mgat2DM/DM mice selectively deplete even adoptively transferredwild-type naive but not effector or memory T cells, confirmingthis interpretation. These findings show that the auto-specific IgMin CVID-Mgat2DM/DM mice is not triggered by changes in T cellsurface phenotype, but rather their depletion is the secondary re-sult of autoantibody generation against targets elsewhere.With the exception of human aging, CVID is the only other
noninfectious disease that is associated with selective loss of naiveT cells. Some reports point to reduced thymic output, as measuredby TCR excision circles (18, 32) and surface expression of CD31(7). Because these biomarkers are known to decrease with ageduring the well-established process of thymic involution and lossof thymic output (33), a similar decrease in CVID patients com-pared with healthy controls was assumed to be related to thymicdysfunction, however this was not supported with direct analysisof the thymus (7, 18). Although reduced thymic output remains
FIGURE 7. The autoantibody response to naive T cells shares common
epitope(s) with erythrocytes expressing the altered glycophenotype. (A)
Indirect Coombs tests analyzed by flow cytometry showing serum IgM
from CVID-Mgat2DM/DM (dashed line) mice, but not Mgat2wt/wt (solid
line) mice, binding to PHA-L2 erythrocytes and naive CD4+CD44low
T cells. (B) CVID-Mgat2DM/DM serum (dashed line) and Mgat2wt/wt serum
(solid line) was preadsorbed to PHA-L2 erythrocytes and then used in
similar binding experiments with fresh PHA-L2 erythrocytes and naive
CD4+CD44low T cells. Indicating a common epitope, the adsorption step
removed PHA-L2 erythrocyte reactivity, as expected, in addition to re-
activity to naive CD4+CD44low T cells.
FIGURE 8. The autoantibody response to naive T cells can be induced using erythrocytes expressing the altered glycophenotype. Membrane preparations
of PHA-L2 erythrocytes were used to immunize naive Mgat2DM/DM mice. Mice were given three i.p. doses over 4 wk with alum alone (mock) or alum-
adsorbed membranes (immunogen). Direct Coombs tests were performed using flow cytometry to detect induced surface bound auto-IgM to endogenous
PHA-L2 erythrocytes (A), as compared with endogenous PHA-L+ erythrocytes (B). The levels of auto-IgM bound to erythrocytes was normalized (norm) to
the average of mock-immunized controls. Circulating blood naive CD4+CD44low T cells (C) and effector/memory CD4+CD44hi T cells (D) were examined
to determine relative cell numbers (left) and level of cell-bound auto-IgM (right). A similar analysis was performed on splenic naive CD4+ T cells (E) and
effector/memory CD4+ T cells (F). Relative T cell numbers and levels of bound auto-IgM were normalized (norm) to the average of mock immunized
controls. *p , 0.05, **p , 0.01.
8 MYELOID GLYCAN–INDUCED CVID
by guest on April 13, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

a possibility, our findings revealed indistinguishable numbers ofsingle and double positive T cells in the thymus in these mice,leading to an alternative hypothesis whereby the mechanism for lossof naive T cells is due to a targeted autoimmune response. Inhumans, this loss of naive T cells would include recent thymicemigrants and thus when compared with healthy controls, theaforementioned biomarkers would also be expected to be decreased.Interestingly, we found that the occurrence of AIHA and naive
T cell autoimmunity was related through an apparent commonepitope. We have not seen mice with AIHA that did not also havethe selective depletion of naive T cells, nor did we findmice lackingnaive T cells that did not show AIHA. More importantly, we foundthat “immunization” of mice with PHA-L-negative erythrocyteghosts could not only induce autoreactive IgM, but that this IgMalso selectively depleted naive T cells even if those cells werefrom wild-type mice. Finally, “absorbed IgM” from CVID-Mgat2DM/DM serum using PHA-L-negative erythrocyte ghosts alsoeliminated IgM deposition on native T cells in vitro (indirectCoombs test). These data show that alterations in the surface gly-cosylation of erythrocytes reveal a cryptic epitope shared betweenerythrocytes and naive T cells that lead directly to their collec-tive and selective depletion. Therefore, in addition to CVID, theMgat2DM/DM mice may also be informative for studies of AIHA.Finally, once naive T cells are depleted in CVID-Mgat2DM/DM
mice, IgG responses to vaccine are severely attenuated. The over-all titer of IgG in these mice prior to immunization is not sig-nificantly different, further supporting the notion that the naiveT cells are not lost at birth, but rather requires a potent immuneresponse triggered by the presence of PHA-L-negative erythro-cytes. However, once those T cells are lost, IgG responses againsta model Ag (OVA) and the human conjugate vaccine Prevnar13mirror the poor response reported for many human CVID patients,thereby completing the circle between loss of erythrocyte complexN-glycans and the defining characteristic of CVID.In total, we have found that glycosylation changes in the
erythrocyte population could explain the incidence of AIHA, lossof naive T cells, and failure to mount potent IgG responses in someCVID patients. Although we cannot yet say whether this modelrecapitulates the mechanism(s) underlying human disease, ourfindings open exciting new avenues for the study of human CVIDwith particular attention paid to changes in glycosylation of keyhematopoietic cells as a triggering event and possibly as a bio-marker for severity.
AcknowledgmentsWe thank Dr. John Schreiber for supplying conjugate vaccine components
and Dr. Alex Huang for assistance with microscopy image acquisition.
DisclosuresThe authors have no financial conflicts of interest.
References1. International Union of Immunological Societies. 1999. Primary immunodefi-
ciency diseases. Report of an IUIS Scientific Committee. Clin. Exp. Immunol.118(Suppl 1): 1–28.
2. Salzer, U., K. Warnatz, and H. H. Peter. 2012. Common variable immunodefi-ciency - an update. Arthritis Res. Ther. 14: 223.
3. Park, J. H., E. S. Resnick, and C. Cunningham-Rundles. 2011. Perspectives oncommon variable immune deficiency. Ann. N. Y. Acad. Sci. 1246: 41–49.
4. Lee, J. J., I. Rauter, L. Garibyan, E. Ozcan, T. Sannikova, S. R. Dillon,A. C. Cruz, R. M. Siegel, R. Bram, H. Jabara, and R. S. Geha. 2009. The murineequivalent of the A181E TACI mutation associated with common variable im-munodeficiency severely impairs B-cell function. Blood 114: 2254–2262.
5. Resnick, E. S., and C. Cunningham-Rundles. 2012. The many faces of theclinical picture of common variable immune deficiency. Curr. Opin. Allergy Clin.Immunol. 12: 595–601.
6. Bateman, E. A., L. Ayers, R. Sadler, M. Lucas, C. Roberts, A. Woods,K. Packwood, J. Burden, D. Harrison, N. Kaenzig, et al. 2012. T cell phenotypesin patients with common variable immunodeficiency disorders: associations withclinical phenotypes in comparison with other groups with recurrent infections.Clin. Exp. Immunol. 170: 202–211.
7. Giovannetti, A., M. Pierdominici, F. Mazzetta, M. Marziali, C. Renzi,A. M. Mileo, M. De Felice, B. Mora, A. Esposito, R. Carello, et al. 2007.Unravelling the complexity of T cell abnormalities in common variable immu-nodeficiency. J. Immunol. 178: 3932–3943.
8. Mouillot, G., M. Carmagnat, L. Gerard, J. L. Garnier, C. Fieschi, N. Vince,L. Karlin, J. F. Viallard, R. Jaussaud, J. Boileau, et al. 2010. B-cell and T-cellphenotypes in CVID patients correlate with the clinical phenotype of the disease.J. Clin. Immunol. 30: 746–755.
9. Agarwal, S., and C. Cunningham-Rundles. 2009. Autoimmunity in commonvariable immunodeficiency. Curr. Allergy Asthma Rep. 9: 347–352.
10. Cunningham-Rundles, C. 2008. Autoimmune manifestations in common vari-able immunodeficiency. J. Clin. Immunol. 28(Suppl. 1): S42–S45.
11. Ryan, S. O., S. M. Leal, Jr., D. W. Abbott, E. Pearlman, and B. A. Cobb. 2014.Mgat2 ablation in the myeloid lineage leads to defective glycoantigen T cellresponses. Glycobiology 24: 262–271.
12. Lai, Z., and J. R. Schreiber. 2011. Outer membrane protein complex of Me-ningococcus enhances the antipolysaccharide antibody response to pneumo-coccal polysaccharide-CRM197 conjugate vaccine. Clin. Vaccine Immunol. 18:724–729.
13. Ryan, S. O., A. M. Vlad, K. Islam, J. Gariepy, and O. J. Finn. 2009. Tumor-associated MUC1 glycopeptide epitopes are not subject to self-tolerance andimprove responses to MUC1 peptide epitopes in MUC1 transgenic mice. Biol.Chem. 390: 611–618.
14. Ryan, S. O., J. A. Bonomo, F. Zhao, and B. A. Cobb. 2011. MHCII glycosylationmodulates Bacteroides fragilis carbohydrate antigen presentation. J. Exp. Med.208: 1041–1053.
15. Ryan, S. O., J. L. Johnson, and B. A. Cobb. 2013. Neutrophils confer T cellresistance to myeloid-derived suppressor cell-mediated suppression to promotechronic inflammation. J. Immunol. 190: 5037–5047.
16. Gruber, W. C., D. A. Scott, and E. A. Emini. 2012. Development and clinicalevaluation of Prevnar 13, a 13-valent pneumocococcal CRM197 conjugatevaccine. Ann. N. Y. Acad. Sci. 1263: 15–26.
17. Podjasek, J. C., and R. S. Abraham. 2012. Autoimmune cytopenias in commonvariable immunodeficiency. Front. Immunol. 3: 189.
18. Guazzi, V., F. Aiuti, I. Mezzaroma, F. Mazzetta, G. Andolfi, A. Mortellaro,M. Pierdominici, R. Fantini, M. Marziali, and A. Aiuti. 2002. Assessment ofthymic output in common variable immunodeficiency patients by evaluation ofT cell receptor excision circles. Clin. Exp. Immunol. 129: 346–353.
19. Ye, M., H. Iwasaki, C. V. Laiosa, M. Stadtfeld, H. Xie, S. Heck, B. Clausen,K. Akashi, and T. Graf. 2003. Hematopoietic stem cells expressing the myeloidlysozyme gene retain long-term, multilineage repopulation potential. Immunity19: 689–699.
20. Sellon, R. K., S. Tonkonogy, M. Schultz, L. A. Dieleman, W. Grenther,E. Balish, D. M. Rennick, and R. B. Sartor. 1998. Resident enteric bacteria arenecessary for development of spontaneous colitis and immune system activationin interleukin-10‑deficient mice. Infect. Immun. 66: 5224–5231.
21. Haury, M., A. Sundblad, A. Grandien, C. Barreau, A. Coutinho, and A. Nobrega.1997. The repertoire of serum IgM in normal mice is largely independent ofexternal antigenic contact. Eur. J. Immunol. 27: 1557–1563.
22. Amaral, J. F., D. A. Foschetti, F. A. Assis, J. S. Menezes, N. M. Vaz, andA. M. Faria. 2006. Immunoglobulin production is impaired in protein-deprivedmice and can be restored by dietary protein supplementation. Braz. J. Med. Biol.Res. 39: 1581–1586.
23. Jaeken, J. 2010. Congenital disorders of glycosylation. Ann. N. Y. Acad. Sci.1214: 190–198.
24. Saldova, R., L. Royle, C. M. Radcliffe, U. M. Abd Hamid, R. Evans,J. N. Arnold, R. E. Banks, R. Hutson, D. J. Harvey, R. Antrobus, et al. 2007.Ovarian cancer is associated with changes in glycosylation in both acute-phaseproteins and IgG. Glycobiology 17: 1344–1356.
25. Turner, G. A., M. T. Goodarzi, and S. Thompson. 1995. Glycosylation of a-1-proteinase inhibitor and haptoglobin in ovarian cancer: evidence for two dif-ferent mechanisms. Glycoconj. J. 12: 211–218.
26. Thompson, S., E. Dargan, I. D. Griffiths, C. A. Kelly, and G. A. Turner. 1993.The glycosylation of haptoglobin in rheumatoid arthritis. Clin. Chim. Acta 220:107–114.
27. Gravel, P., C. Walzer, C. Aubry, L. P. Balant, B. Yersin, D. F. Hochstrasser, andJ. Guimon. 1996. New alterations of serum glycoproteins in alcoholic and cir-rhotic patients revealed by high resolution two-dimensional gel electrophoresis.Biochem. Biophys. Res. Commun. 220: 78–85.
28. Mann, A. C., C. O. Record, C. H. Self, and G. A. Turner. 1994. Monosaccharidecomposition of haptoglobin in liver diseases and alcohol abuse: large changes inglycosylation associated with alcoholic liver disease. Clin. Chim. Acta 227: 69–78.
29. Goodarzi, M. T., and G. A. Turner. 1998. Reproducible and sensitive determi-nation of charged oligosaccharides from haptoglobin by PNGase F digestion andHPAEC/PAD analysis: glycan composition varies with disease. Glycoconj. J. 15:469–475.
30. Demetriou, M., M. Granovsky, S. Quaggin, and J. W. Dennis. 2001. Negativeregulation of T-cell activation and autoimmunity by Mgat5 N-glycosylation.Nature 409: 733–739.
31. Wang, Y., J. Tan, M. Sutton-Smith, D. Ditto, M. Panico, R. M. Campbell,N. M. Varki, J. M. Long, J. Jaeken, S. R. Levinson, et al. 2001. Modeling
The Journal of Immunology 9
by guest on April 13, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

human congenital disorder of glycosylation type IIa in the mouse: conser-vation of asparagine-linked glycan-dependent functions in mammalianphysiology and insights into disease pathogenesis. Glycobiology 11: 1051–1070.
32. De Vera, M. J., L. Al-Harthi, and A. T. Gewurz. 2004. Assessing thymo-poiesis in patients with common variable immunodeficiency as measured
by T-cell receptor excision circles. Ann. Allergy Asthma Immunol. 93: 478–484.
33. Kimmig, S., G. K. Przybylski, C. A. Schmidt, K. Laurisch, B. Mowes,A. Radbruch, and A. Thiel. 2002. Two subsets of naive T helper cells withdistinct T cell receptor excision circle content in human adult peripheral blood. J.Exp. Med. 195: 789–794.
10 MYELOID GLYCAN–INDUCED CVID
by guest on April 13, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from