Embed Size (px)
Citation preview

Antropologia Molecolare
6 CFU (4+2)
Prof.ssa Carla M. Calò
Tel.: 070 6754154
E-mail: [email protected]

Cos’è l’Antropologia Antropos: uomo
Logos: discorso
Antropologia
Paleontologia
Antropometria
Auxologia
Demografia
Antropologia
molecolare

Antropologia:
Lo studio dell’uomo in rapporto al suo contesto biologico, demografico e storico-culturale.
Antropologia molecolare:
L’introduzione di nuovi strumenti basati sulla variabilità del DNA ha permesso di affrontare queste problematiche da un ulteriore punto di vista e creando una nuova branca dell’antropologia “L’ Antropologia molecolare”.

Programma
Introduzione all’antropologia molecolare: nascita, sviluppi e finalità. Dall'analisi dei marcatori
classici alla NGS.
Variabilità inter e intra-popolazionistica: significato e come verificarla.
Le forze evolutive. I processi microevolutivi nelle popolazioni umane.
I marcatori molecolari nello studio delle popolazioni umane (marcatori biallelici, VNTRs,
STRs, SNPs): origine, tassi di mutazione e applicazioni. Vantaggi nell’utilizzo dei marcatori a
trasmissione uniparentale (mtDNA e porzione non ricombinante del cromosoma Y).
Richiami sulle teorie dell’evoluzione umana. Origine e diffusione dell’uomo anatomicamente
moderno: le diverse teorie analizzate da un punto di vista molecolare. La posizione
dell’uomo di Neanderthal. La posizione dei nuragici. Il popolamento dei vari continenti da
parte di Homo sapiens ricostruito attraverso i dati molecolari.
Caratteristiche del DNA antico. Le Metodologie di analisi. Il problema delle contaminazioni:
precauzioni e protocolli di lavoro. Marcatori utilizzati nello studio del DNA antico. Analisi
bioinformatiche per la ricostruzione e l’analisi delle sequenze di DNA.
Esempi di evoluzione recente: la persistenza della lattasi.
Variabilità genetica e suscettibilità individuale verso patologie complesse o fenotipi
multifattoriali.
La fallacità del concetto di “razza umana” secondo gli studi sul DNA.
Cenni su DNA e analisi forensi. I marcatori utilizzati nell’analisi forense e negli studi di
associazione.
Metodi di analisi della variabilità genetica: allineamento e confronto di sequenze, distanze
genetiche, alberi genetici, metodi per la ricostruzione degli alberi. L'orologio molecolare e la
datazione dell'ancestore comune.

Esercitazioni
Prelievo campione buccale per estrazione: confronto tra diverse
metodiche (spazzolino tradizionale, spazzolino in etanolo, raccolta di
saliva e sciacqui con colluttorio).
Estrazione DNA, quantificazione con fluorimetro e spettrofotometro.
PCR
Digestione enzimatica per il riconoscimento di polimorfismi SNPs
Vari tipi di elettroforesi (in agarosio e in poliacrilamide) per il
riconoscimento e l'identificazione di polimorfismi.
Esercitazioni al computer per l’utilizzo di banche dati e allineamento di
sequenze
Testi consigliati:
•Jobling, Hollox, Hurles, Kivisild, Tyler-Smith. Human Evolutionary
Genetics. Garland Science, 2014
Antropologia Molecolare – D. Caramelli Ed. Firenze University Press
•Genetica delle popolazioni umane . J.H. Relethford. Casa Ed.
Ambrosiana (no i primi 3 capitoli).
•Materiale e Articoli scientifici forniti dalla docente

Di cosa ci occupiamo oggi?
Introduzione all’antropologia molecolare: nascita,
sviluppi e finalità. Dall'analisi dei marcatori classici alla
NGS.
Variabilità inter e intra-popolazionistica: significato e
come verificarla.
Le forze evolutive. I processi microevolutivi nelle
popolazioni umane.

Prima della sua definizione formale (1962)
1901 Landsteiner scopre i gruppi sanguigni ABO, Vienna (Nobel nel 1930)
gli studi sulle reazioni tra diversi tipi di sangue sono i primi a mettere in luce l’esistenza di una variabilità molecolare tra individui
1919 primo studio di popolazione sul sitema ABO
successivamente Rh, MNS, etc... sempre con tecniche immunologiche
1949 introduzione della tecnica dell’elettroforesi per separare diverse varianti dell’emoglobina
questo permette di allargare il numero di varianti, si iniziano a descrivere le popolazioni in termini di frequenze alleliche (1954 primo libro ad opera di Mourant)
contemporaneamente nasce la genetica di popolazioni e si vanno delineando le linee guida della teoria sintetica dell’evoluzione (la selezione come elemento chiave nella distribuzione della variabilità – Fisher, Haldane, Wright)
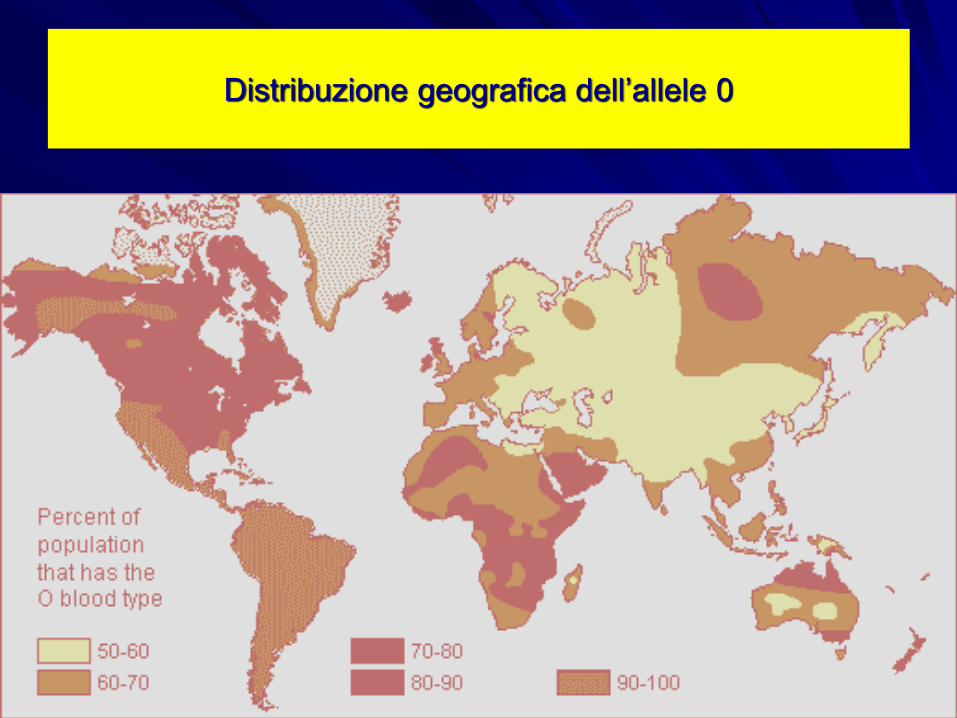
Distribuzione geografica dell’allele 0
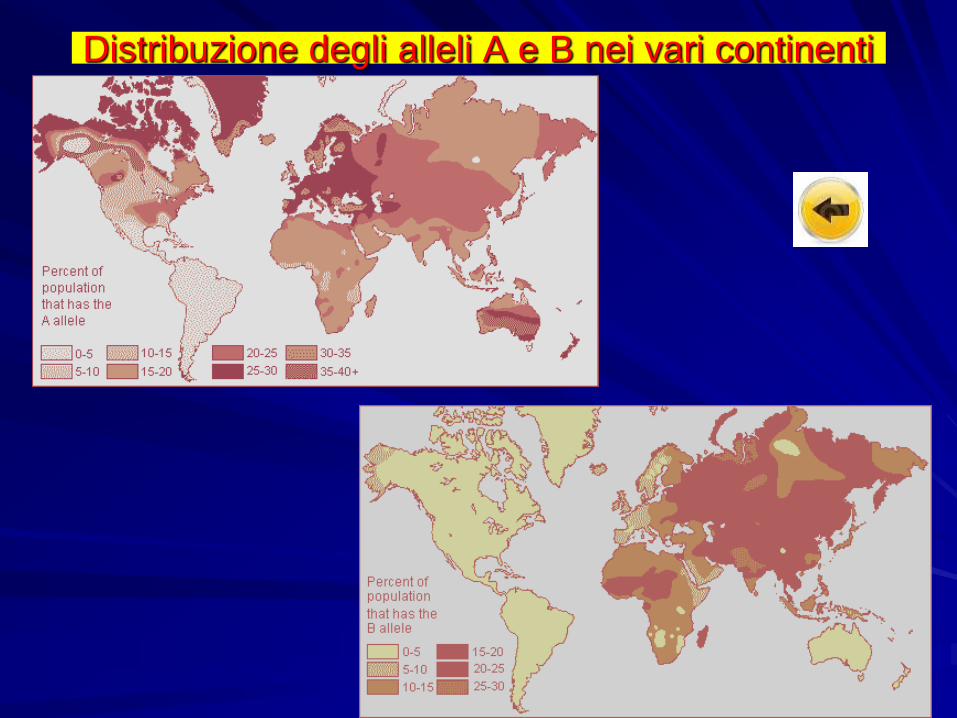
Distribuzione degli alleli A e B nei vari continenti
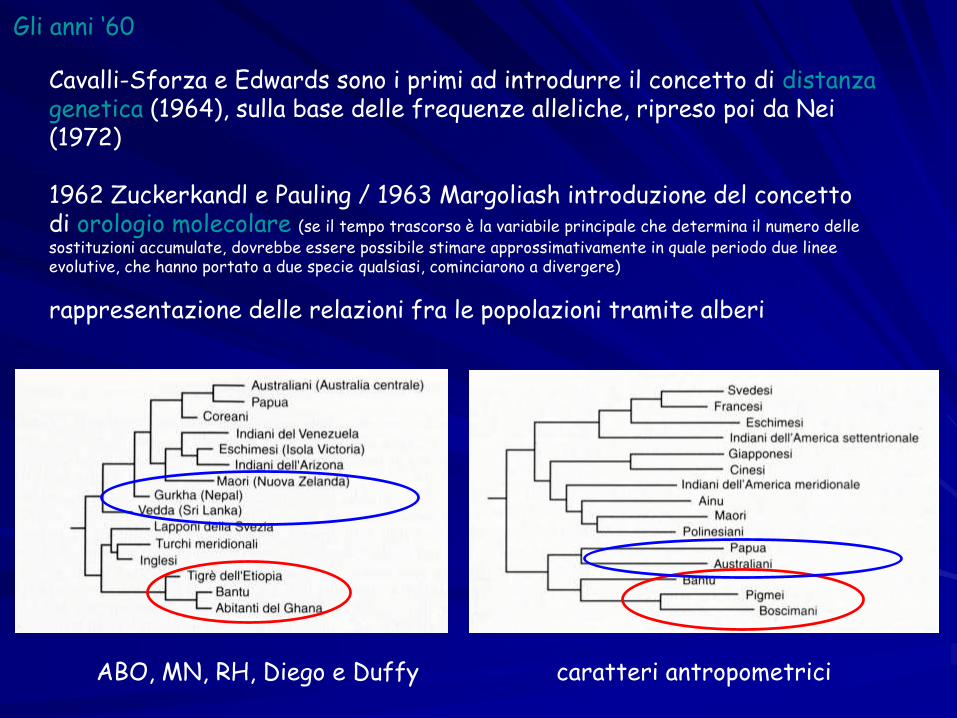
Gli anni ‘60
Cavalli-Sforza e Edwards sono i primi ad introdurre il concetto di distanza genetica (1964), sulla base delle frequenze alleliche, ripreso poi da Nei (1972)
1962 Zuckerkandl e Pauling / 1963 Margoliash introduzione del concetto di orologio molecolare (se il tempo trascorso è la variabile principale che determina il numero delle sostituzioni accumulate, dovrebbe essere possibile stimare approssimativamente in quale periodo due linee evolutive, che hanno portato a due specie qualsiasi, cominciarono a divergere)
rappresentazione delle relazioni fra le popolazioni tramite alberi
ABO, MN, RH, Diego e Duffy caratteri antropometrici
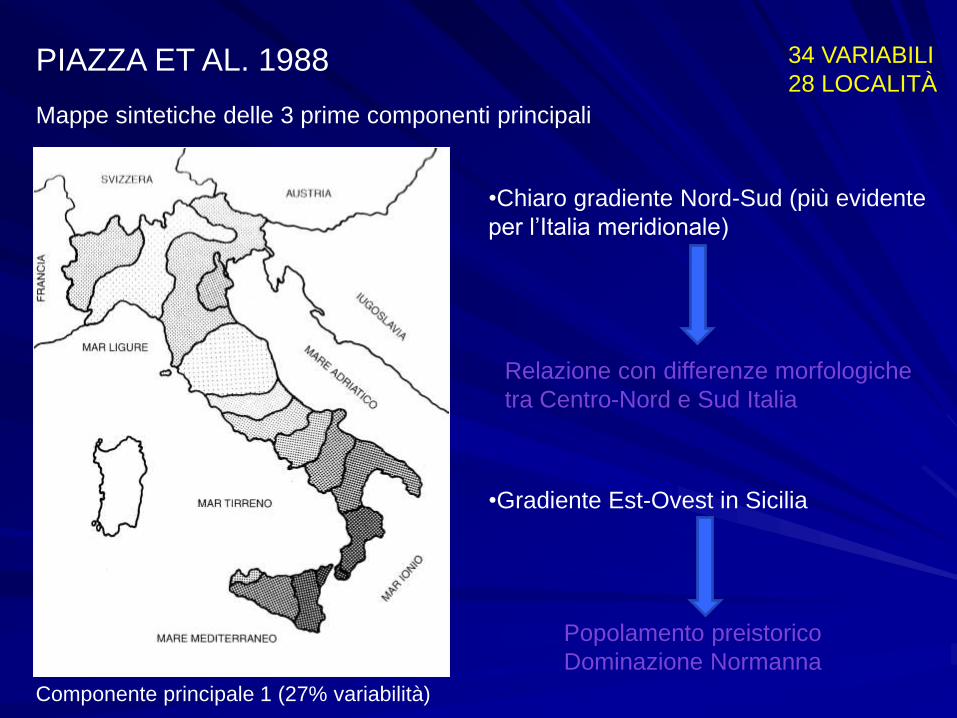
PIAZZA ET AL. 1988 34 VARIABILI
28 LOCALITÀ
Mappe sintetiche delle 3 prime componenti principali
Componente principale 1 (27% variabilità)
•Chiaro gradiente Nord-Sud (più evidente
per l’Italia meridionale)
Relazione con differenze morfologiche
tra Centro-Nord e Sud Italia
•Gradiente Est-Ovest in Sicilia
Popolamento preistorico
Dominazione Normanna

Gli anni ‘70
inizio ’70 scoperta degli enzimi di restrizione di E.coli – nel 1978 Nathans, Arber e Smith ricevono il Nobel per la Medicina
1974 Cohen e Boyer - tecnica del DNA ricombinante
1977 Maxam e Gilbert – sequenziamento del DNA tramite il metodo della degradazione chimica
1977 Sanger – sequenziamento del DNA tramite metodo della terminazione della catena (che si usa ancora oggi)
contemporaneamente Kimura propone la teoria neutrale dell’evoluzione molecolare (la deriva come elemento chiave nella distribuzione della variabilità)

quindi siamo passati da…
antigeni
proteine
DNA bassa risoluzione (enzimi di restrizione)
DNA alta risoluzione(sequenze)
Next generation sequences
aumenta la risoluzione molecolare, la precisione e l’informazione ottenuta
diminuiscono le costrizioni funzionali sul polimorfismo analizzato

tutte le cellule di un individuo recano il medesimo
patrimonio genetico (DNA nucleare, DNA mitocondriale
citoplasmatico)
il 99,9% della sequenza di DNA è identica in tutti gli esseri
umani
nel restante 0,1% sono possibili delle differenze inter-
individuali (polimorfismi)
Ricordiamo che:

Le popolazioni naturali possiedono unanotevole variabilità genetica
(fattore determinante per l’evoluzione della specie e l’adattamento all’ambiente)

Variabilità: il punto di partenza
per ogni analisi genetica o
evoluzionistica è l’osservazione
che un certo carattere, o
complesso di caratteri, è
variabile o polimorfico nei
diversi individui

Variabilità Discontinua: due o più forme (fenotipi distinti) il mutante e il wild type, es. albinismo.
Variabilità continua: gamma ininterrotta di fenotipi (es. altezza, colore della pelle). Forte influenza ambientale.
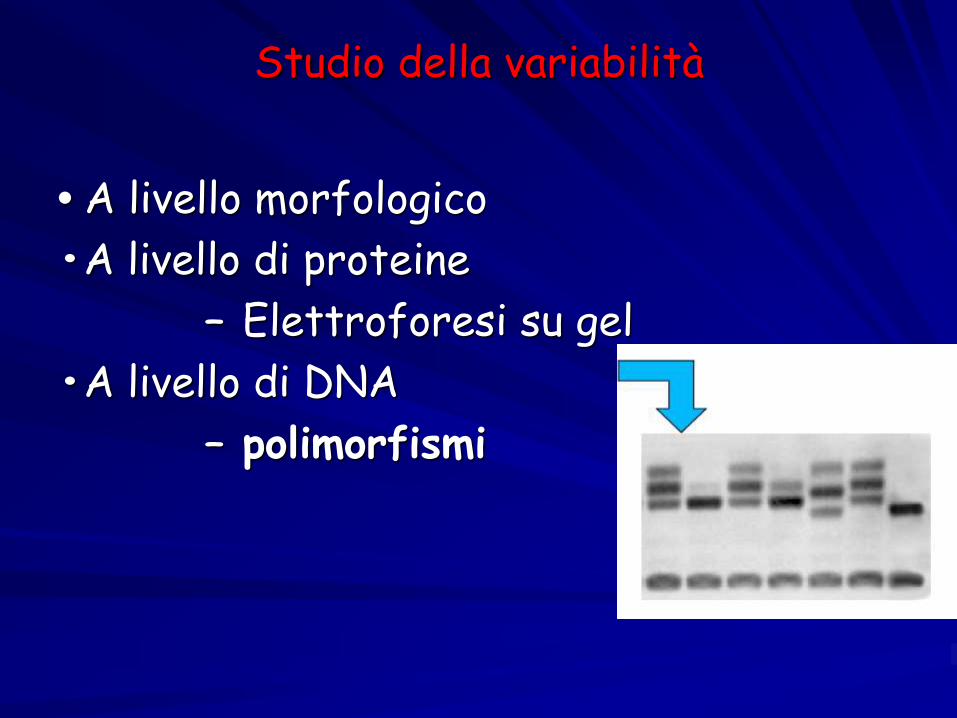
Studio della variabilità
• A livello morfologico
•A livello di proteine
– Elettroforesi su gel
•A livello di DNA
– polimorfismi

Fattori che influiscono sulla variazione della
struttura genetica di una popolazione
• Mutazione
• Migrazione (flusso genico)
• Dimensioni della popolazione e deriva genetica casuale
• Selezione
• Sistema di accoppiamento

la mutazioneUna differenza fra 2 sequenze umane può
essere considerata un
polimorfismo se l’allele a minore frequenza ha nella popolazione una frequenze >1%.
Mutazione se patogena
Variante
Cambia la costituzione genetica delle popolazioni con un tasso molto basso

Tasso di mutazione
La mutazione è un evento raro: 2.3x10-8
Ma varia a seconda della regione genomica
del tipo di mutazione.
Siamo tutti
portatori di alleli
mutanti, ogni
persona porta
circa 147
mutazioni
(22-529 miliardi di
nuove mutazioni
per generazione)

Tipi di mutazioni:
•Mutazioni puntiformi (sostituzioni singoli
nucleotidi)
•Inserzioni o delezioni
•Cambiamenti cromosomici

Mutazioni in cellule somaticheMutazioni in cellule germinali : ereditarie. Su di esse agisce la selezione
A seconda dell’effetto fenotipico:Silenti: la mutazione cambia il codone per un aa in un altro
codone per lo stesso aa (es CUA e UUA specificano entrambi per
Leucina)
Nonsenso: la mutazione cambia il codone per un aa in un
codone di stop
Missenso: la mutazione cambia il codone per un aa in un
codone per un altro aa

Possono essere classificate in sottogruppi:
- Sostituzione conservativa: l’aa sostituito ha caratteristiche
chimiche simili al residuo fisiologico, con conseguenze spesso
non gravi sulla funzione proteica.
- Sostituzione NON-conservativa: l’aa sostituito ha
caratteristiche chimiche diverse dal residuo fisiologico, con
conseguenze spesso gravi sulla funzione proteica.
- Sostituzione Read-through: un codone di stop viene sostituto
da un codone che specifica per un aa con conseguente
allungamento del prodotto proteico

Frameshift: Inserzioni o delezioni di 1, 2, 4, 5… nucleotidi provocano la lettura errata di tutto il tratto di DNA a valle.
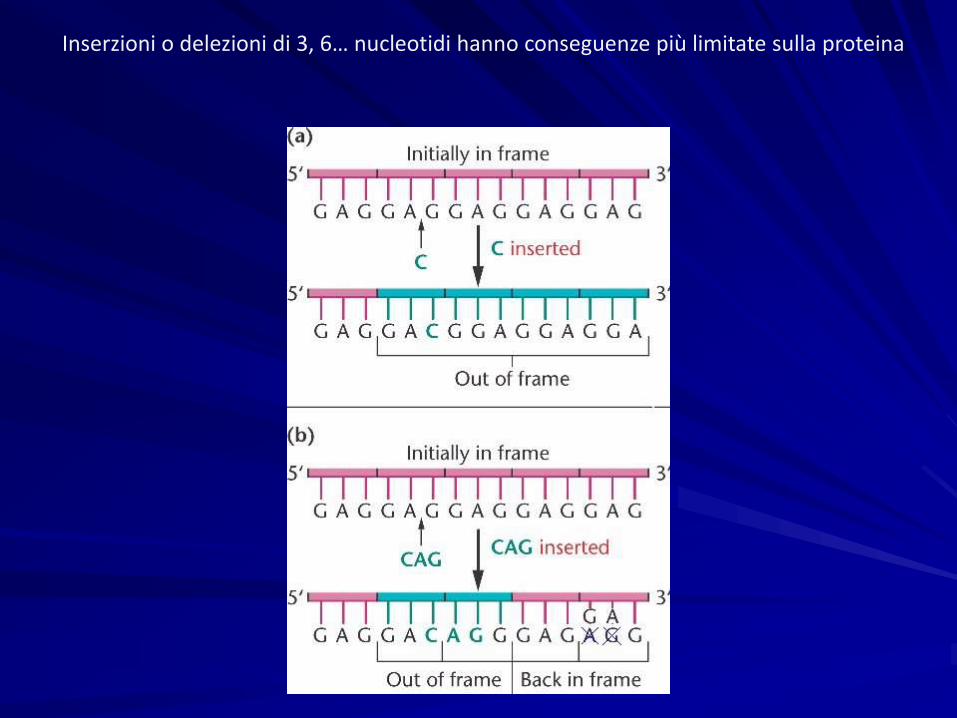
Inserzioni o delezioni di 3, 6… nucleotidi hanno conseguenze più limitate sulla proteina

L’impatto evolutivo della mutazione
1) La mutazione è una causa di evoluzione
necessaria (non sufficiente)
2) La mutazione è un evento casuale
3) La mutazione deve manifestarsi nelle cellule
sessuali

Con le mutazioni si possono calcolare delle date
Più tempo passa, più mutazioni si accumulano
Il numero di mutazioni che separa due specie è proporzionale al tempo intercorso dall’antenato comune

Conoscendo (p.e.s., sulla base di dati fossili) il tempo di separazione fra la specie A e la specie B, possiamo calcolare un tasso di differenziazione molecolare, che poi ci permetterà di stimare i tempi di separazione fra A e C, D, E, ecc.
L’orologio molecolare
Uomo e scimpanzè si sono separati fra 8 e 6 milioni di anni fa (diciamo 6).Se per un certo tratto di DNA troviamo fra loro 15 differenzeTasso di divergenza = TD = 15/6milioni = 2,5 per milione di anni
Se fra uomo e gorilla ci sono 17 differenze, l’antenato comune fra uomo e gorilla sarà vissuto 17 / 2,5 7 milioni di anni fa


Tre modelli di mutazione
Alleli infiniti: ogni evento mutazionale genera un allele diverso
Siti infiniti: ogni evento mutazionale colpisce un sito diverso
Stepwise: ogni evento mutazionale allunga o accorcia di un repeat un locus STR o VNTR

Associare a ciascuna definizione il termine corrispondente.
1. Sostituzione nucleotidica che genera un codone di stop2. Perdita o acquisto di un tratto di DNA3. Sostituzione nucleotidica che provoca il cambio di un codone in un altro
codone per lo stesso amminoacido4. Sostituzione nucleotidica che provoca il cambio di un codone in un codone per
un altro amminoacido5. Perdita o acquisto di pochi nucleotidi, che alterano la lettura della sequenza in
tutto il tratto a valle6. Sostituzione, perdita o acquisto di un singolo nucleotide
a. Indel c. Puntiforme d. Silente
e. Nonsenso f. Missenso g. Frameshift
Vediamo se ci siamo capiti

La migrazione e flusso genico
Si ha quando individui si spostano da una
popolazione ad un’altra, con la quale si
accoppiano
Determina un cambiamento locale delle
frequenze alleliche
Dipende dal tasso della migrazione

migrazione
La migrazione tende a rendere omogenee le frequenze alleliche tra
diverse popolazioni e determina quindi una diminuzione della
variabilità interpopolazioni (FST tende a zero).
Allo stesso tempo introduce nuova variazione nelle popolazioni,
determinando un aumento medio della variabilità intrapopolazioni (H
aumenta)
1. Migrazione unidirezionale
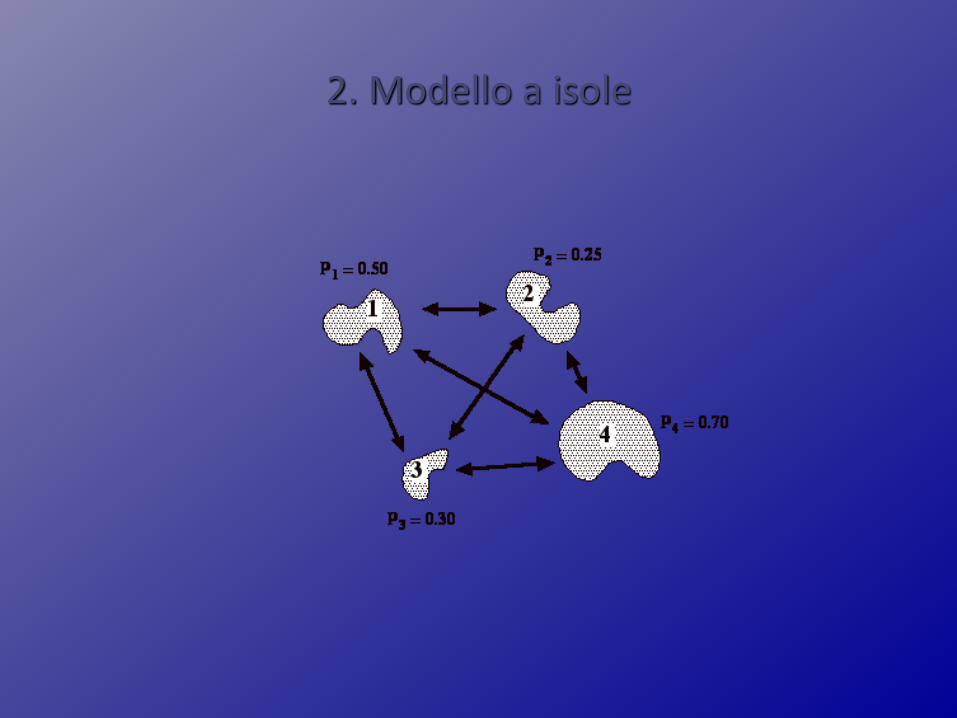
2. Modello a isole
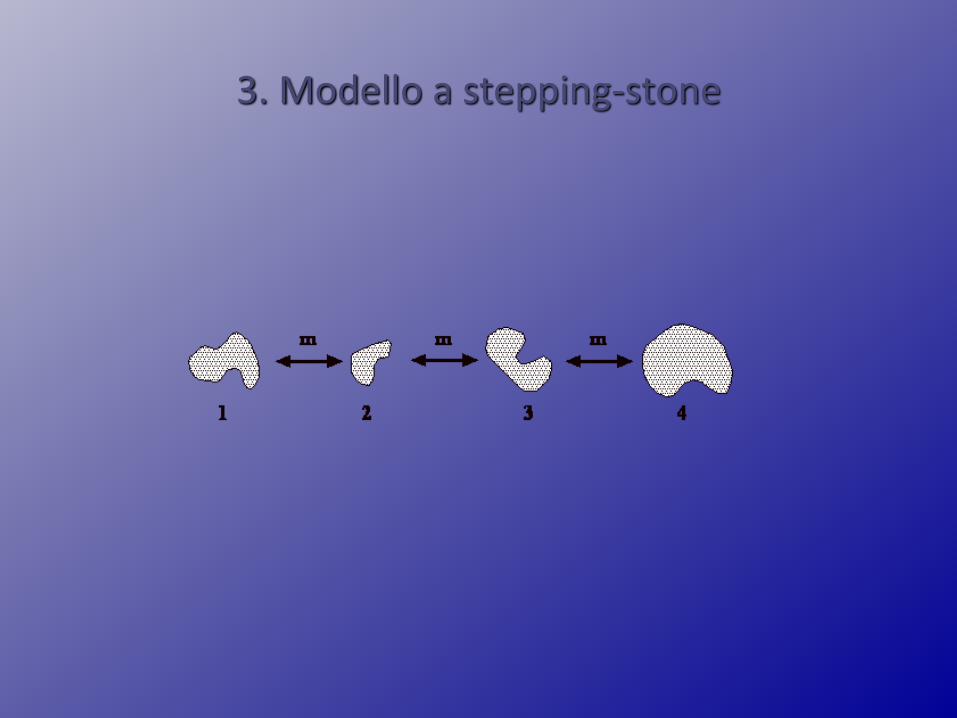
3. Modello a stepping-stone
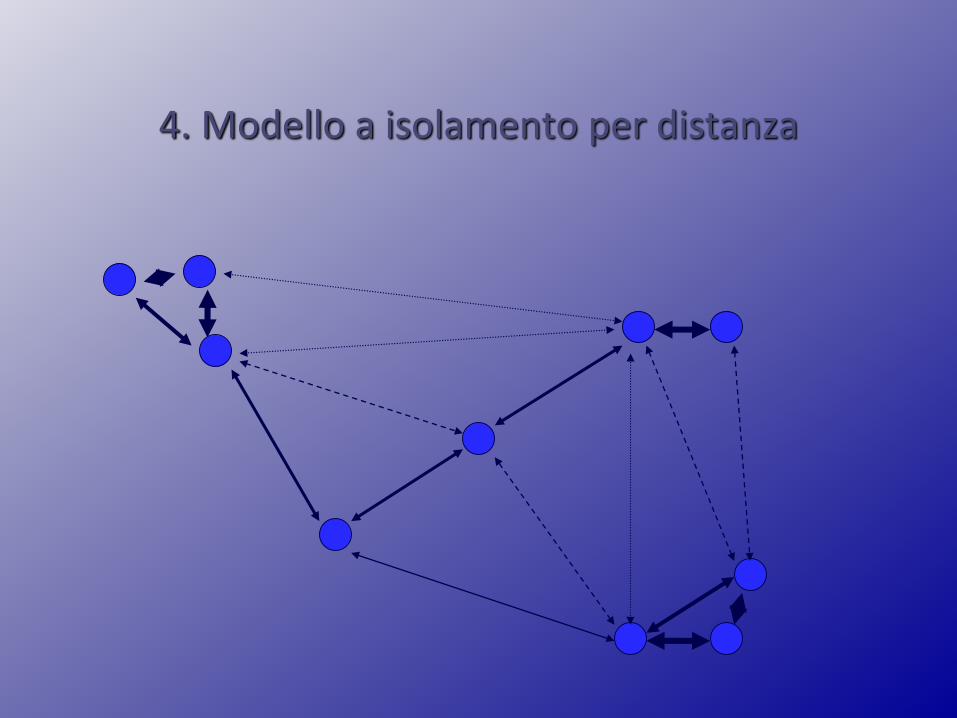
4. Modello a isolamento per distanza

Modello: migrazione unidirezionale
Popolazione 1
N grande
Freq. A = p
Popolazione 2
N piccolomigrazione
Xtrascurabile
Freq. A = r
Dopo una generazione di migrazione:
r1 = r (1-m) + p (m)
Dopo t generazioni di migrazione:
rt= p + (r – p) (1-m)t
m = tasso di migrazione (proporzione di migranti
da pop1 in pop2 ad ogni generazione)
da cui, per t= ∞ r = p
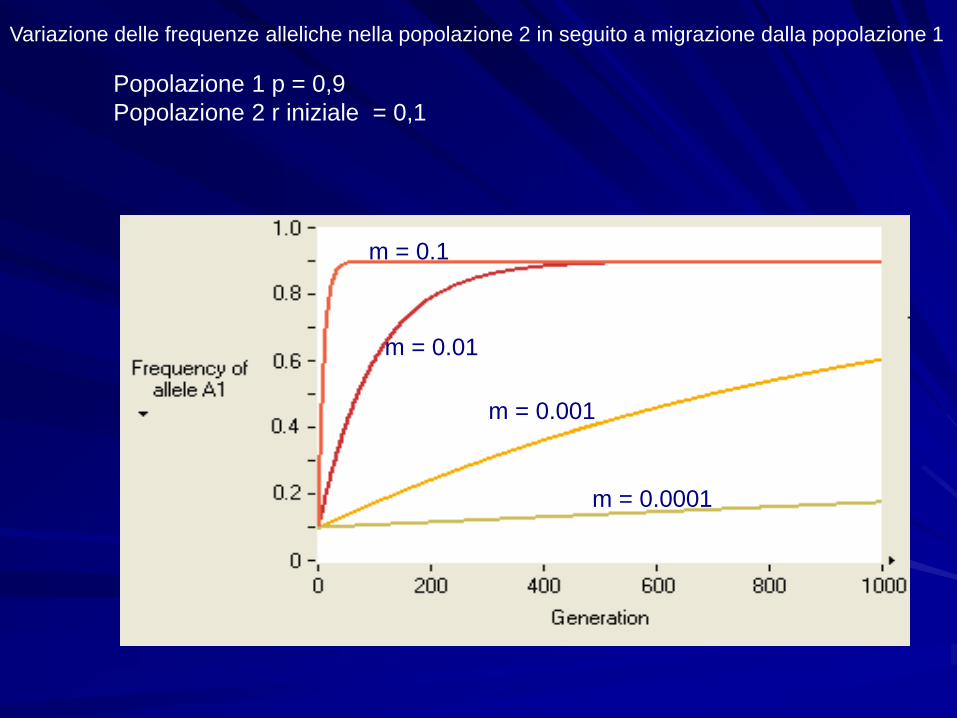
Variazione delle frequenze alleliche nella popolazione 2 in seguito a migrazione dalla popolazione 1
Popolazione 1 p = 0,9
Popolazione 2 r iniziale = 0,1
m = 0.0001
m = 0.1
m = 0.01
m = 0.001
150/200 = 0.7550/200 = 0.25
3/12 = 0.25
9/12 = 0.75
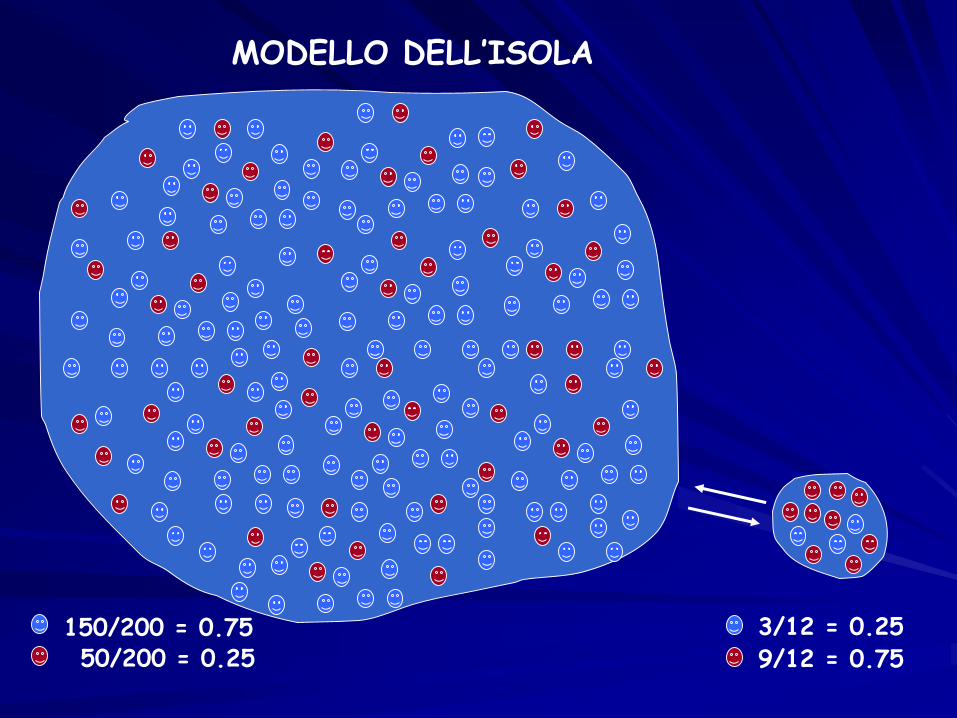
MODELLO DELL’ISOLA
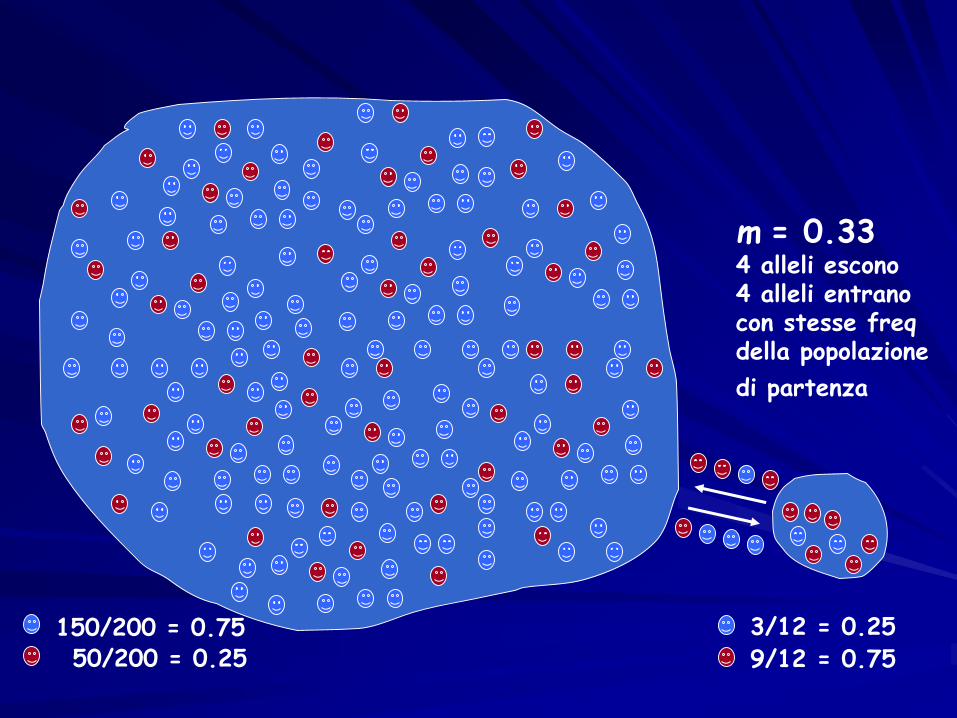
150/200 = 0.7550/200 = 0.25
3/12 = 0.25
9/12 = 0.75
m = 0.334 alleli escono4 alleli entranocon stesse freqdella popolazione
di partenza
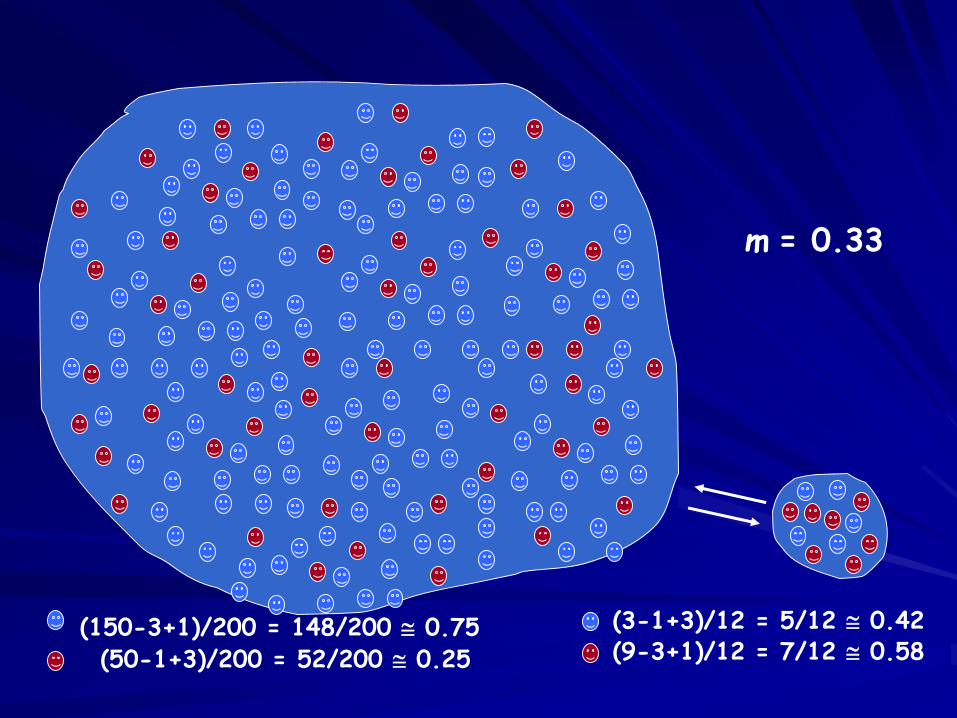
(150-3+1)/200 = 148/200 0.75(50-1+3)/200 = 52/200 0.25
m = 0.33
(3-1+3)/12 = 5/12 0.42(9-3+1)/12 = 7/12 0.58
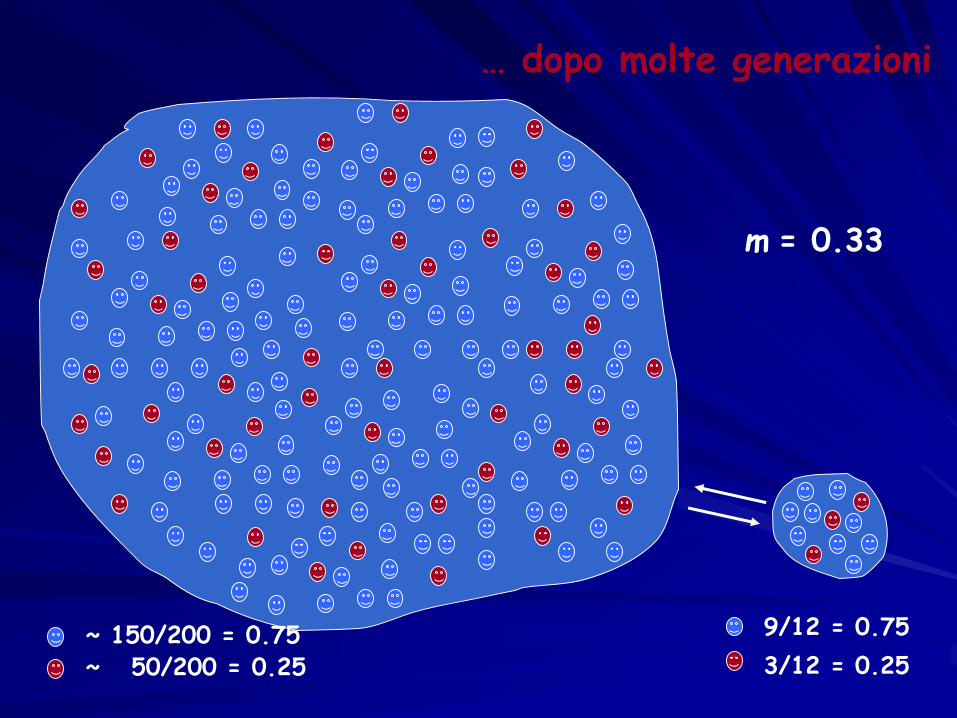
m = 0.33
… dopo molte generazioni
~ 150/200 = 0.75~ 50/200 = 0.25
9/12 = 0.75
3/12 = 0.25

Esempio:
l’aplotipo R0 del sistema Rh ha una frequenza molto bassa negli europei (3%), del 63% negli africani e del 45% negli afroamericani degli USAIsola: africani che migrano verso il continente (USA)
MIGRAZIONE: modello dell’isola
Dopo n generazioni quale è stato l’effetto nella popolazione europea degli USA (continente) della migrazione dei geni africani?E l’effetto negli africani dei geni europei?
Qual è il tasso di migrazione genica (allelica) media dagli europei (E) agli africani (A) nell’attuale popolazione afroamericana (AA) degli USA?

rAA - pE = (1 m)t (pA - pE)
0,45 - 0,03 = (1 m)t (0,63 - 0,03)
0,42----- = (1 m)t
0,60
pA= 0.63 (frequenza di R0 negli africani occidentali attuali, cioè freq. allelica iniziale nell’isola)
pE = 0.03 (frequenza di R0 negli europei attuali, cioèfreq. allelica nel continente)
rAA = 0.45 (frequenza di R0 negli afroamericani attuali, cioè freq. allelica dopo n generazioni)
rt= p + (r – p) (1-m)t

E’ il tasso di migrazione genica dagli europei agli africani
nella popolazione afroamericana degli USA, cioè a ogni
generazione il 3,5% di geni europei è entrato a far parte
del pool di geni della popolazione degli afroamericani degli
USA.
n = 10 (n. generazioni di immigrazione di africani negli USA a partire da circa 250-300 anni fa con la schiavitù)
0,42----- = (1 m)10
0,60
-0,3567 = 10 ln (1-m)
ln (1-m) = -0,03567
m = 0.035

Stima dell’admixture
Sistema sanguigno Duffy negli Afroamericani
Fya nei bianchi: 0.43 (px)
Fya negli Africani: 0 (py)
Nella popolazione nera dell’Oklaoma Fya: 0.0941 (ry)
M= (ry-py)/(px-py)= (0.0941-0)/(0.43-0)=0.21
Il contributo dei geni bianchi al pool genico della popolazione nera
dell’Oklaoma (dovuto agli schiavi) risulta circa il 21%.
Tenendo conto che dall’arrivo dei neri negli USA sono passati circa
300 anni, cioè 10 generazioni, il flusso genico è di circa il 2% (21/10)
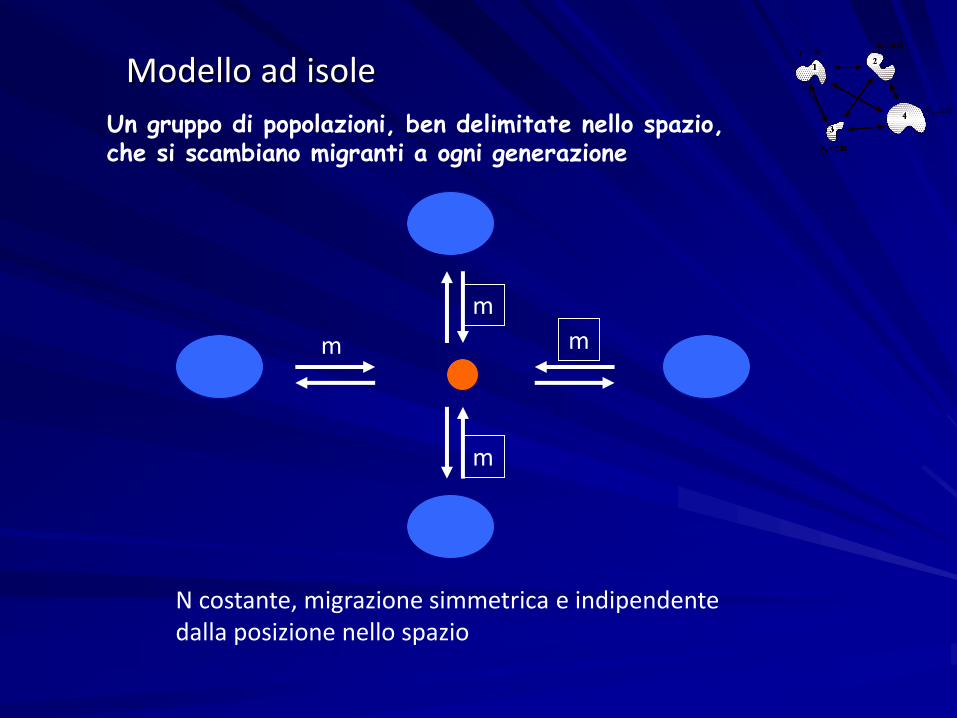
Modello ad isole
mm
m
m
N costante, migrazione simmetrica e indipendentedalla posizione nello spazio
Un gruppo di popolazioni, ben delimitate nello spazio, che si scambiano migranti a ogni generazione
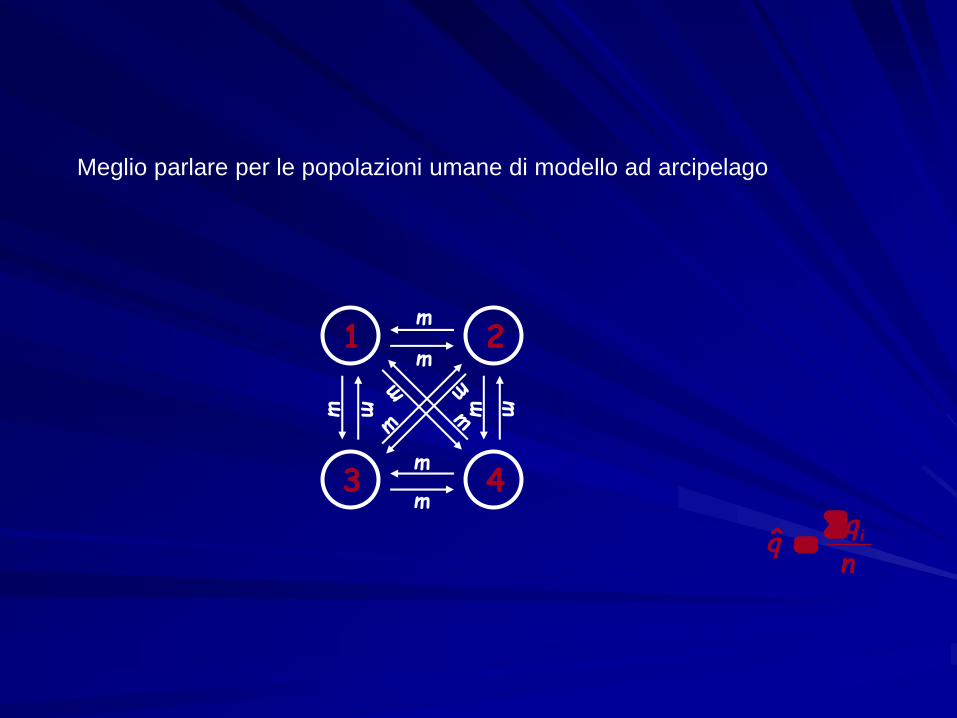
Meglio parlare per le popolazioni umane di modello ad arcipelago
1
43
2
m
mm
m
m
mm
mn
qq̂ i
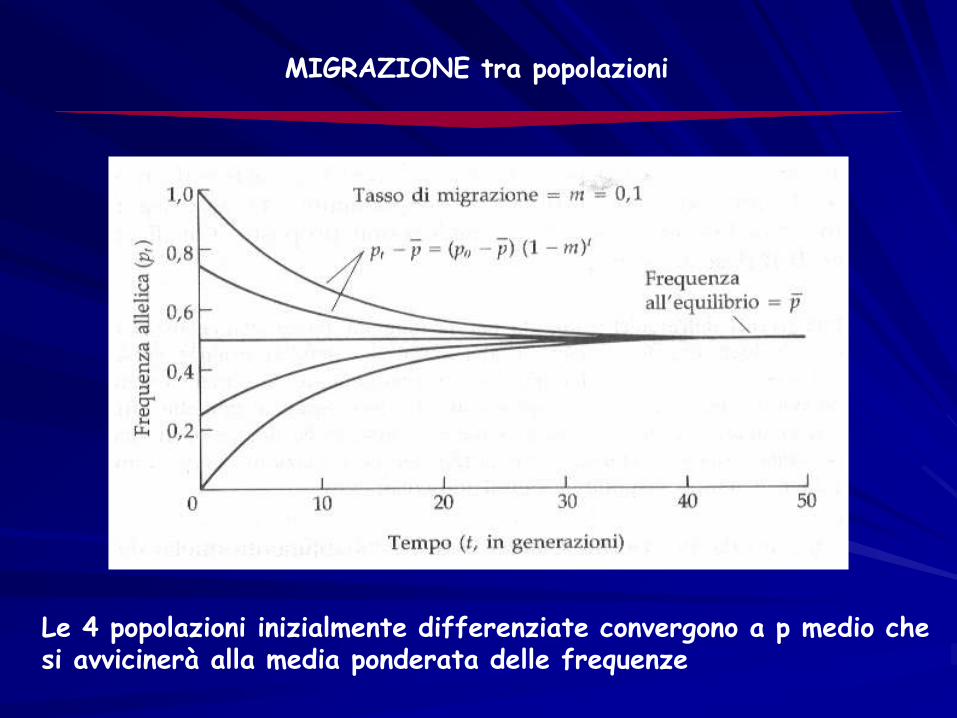
MIGRAZIONE tra popolazioni
Le 4 popolazioni inizialmente differenziate convergono a p medio chesi avvicinerà alla media ponderata delle frequenze
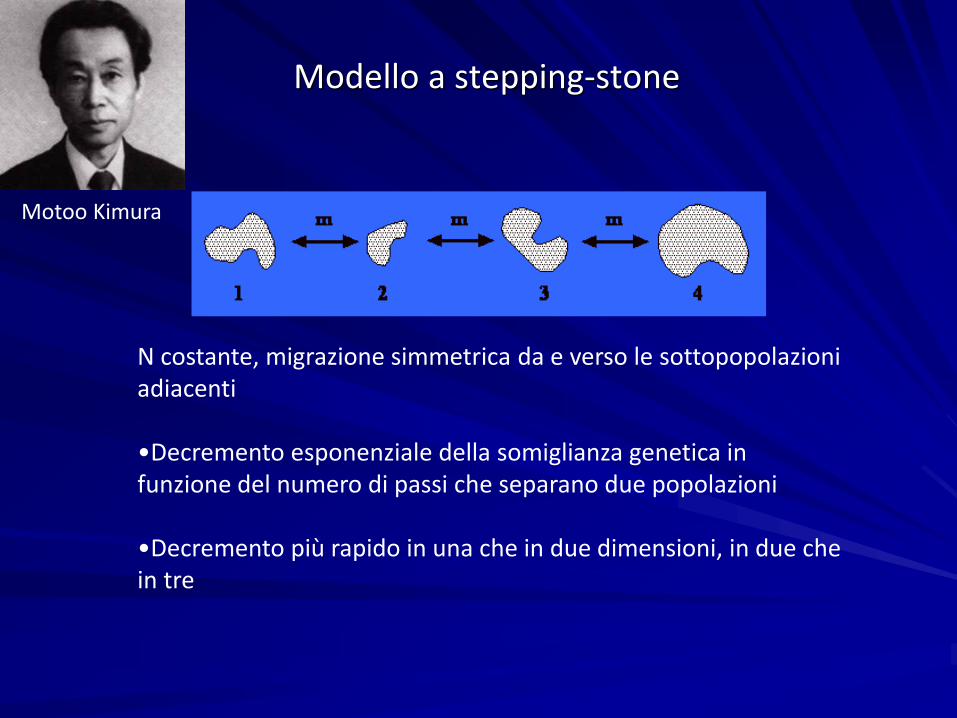
Modello a stepping-stone
N costante, migrazione simmetrica da e verso le sottopopolazioniadiacenti
•Decremento esponenziale della somiglianza genetica in funzione del numero di passi che separano due popolazioni
•Decremento più rapido in una che in due dimensioni, in due chein tre
Motoo Kimura

Modello a isolamento per distanza
N costante, migrazione simmetrica in funzione della distanza geografica frapopolazioni, deriva
Decremento esponenziale della somiglianza genetica (kinship) in funzionedella distanza geografica
Kinship = φij = (pi-pmed) (pj-pmed)
φ(d) = a + e-bd + L (Malécot-Morton)
d
ln φ
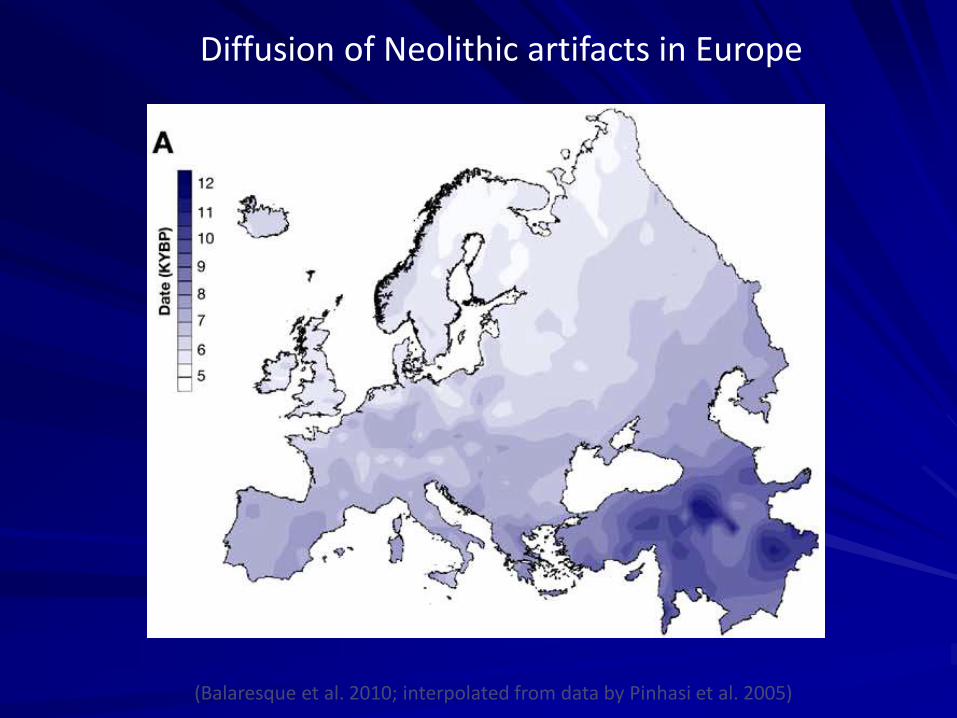
Diffusion of Neolithic artifacts in Europe
(Balaresque et al. 2010; interpolated from data by Pinhasi et al. 2005)
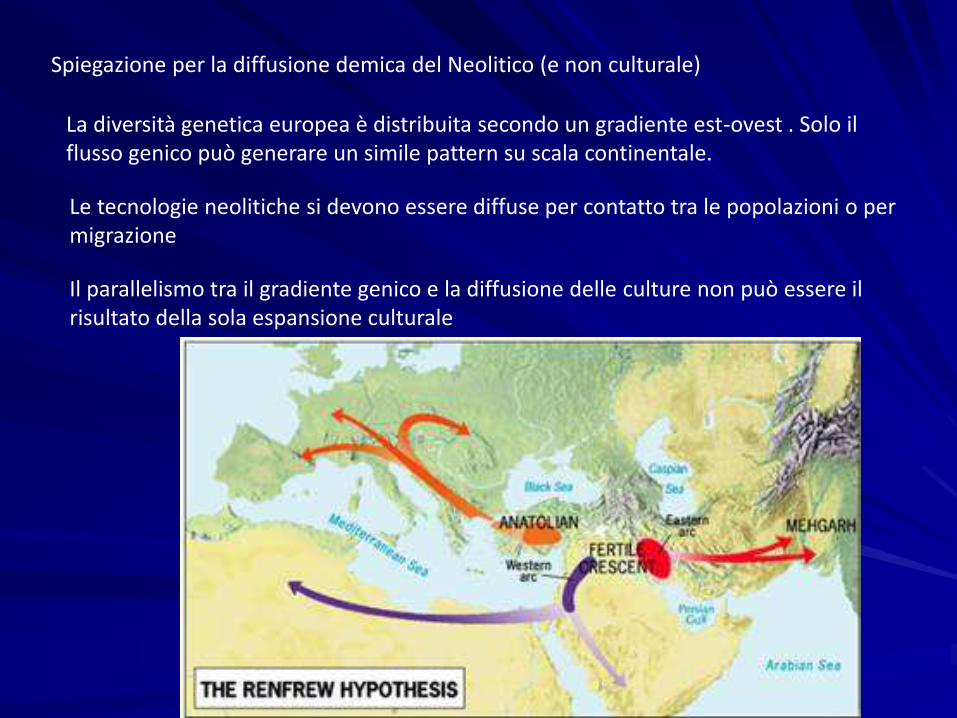
Spiegazione per la diffusione demica del Neolitico (e non culturale)
La diversità genetica europea è distribuita secondo un gradiente est-ovest . Solo il flusso genico può generare un simile pattern su scala continentale.
Le tecnologie neolitiche si devono essere diffuse per contatto tra le popolazioni o per migrazione
Il parallelismo tra il gradiente genico e la diffusione delle culture non può essere il risultato della sola espansione culturale

There can be sex-specific differences.
Marital residence patternes are critical to
investigating local migration patterns in
migration.
Patrilocality
Matrilocality
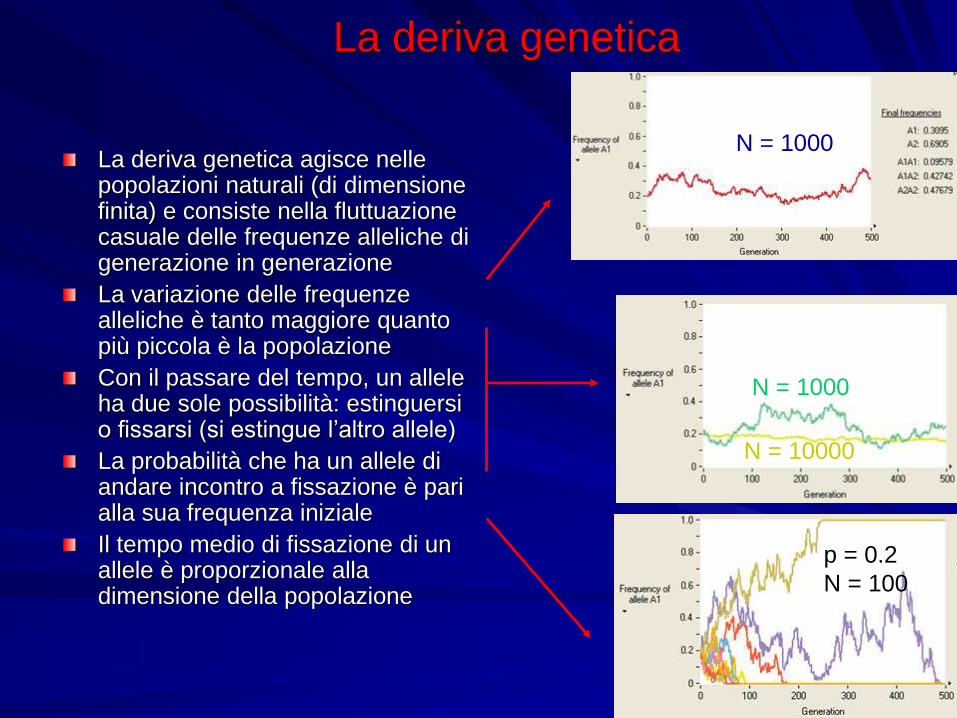
La deriva genetica
La deriva genetica agisce nelle popolazioni naturali (di dimensione finita) e consiste nella fluttuazione casuale delle frequenze alleliche di generazione in generazione
La variazione delle frequenze alleliche è tanto maggiore quanto più piccola è la popolazione
Con il passare del tempo, un allele ha due sole possibilità: estinguersi o fissarsi (si estingue l’altro allele)
La probabilità che ha un allele di andare incontro a fissazione è pari alla sua frequenza iniziale
Il tempo medio di fissazione di un allele è proporzionale alla dimensione della popolazione
N = 1000
N = 1000
N = 10000
p = 0.2
N = 100
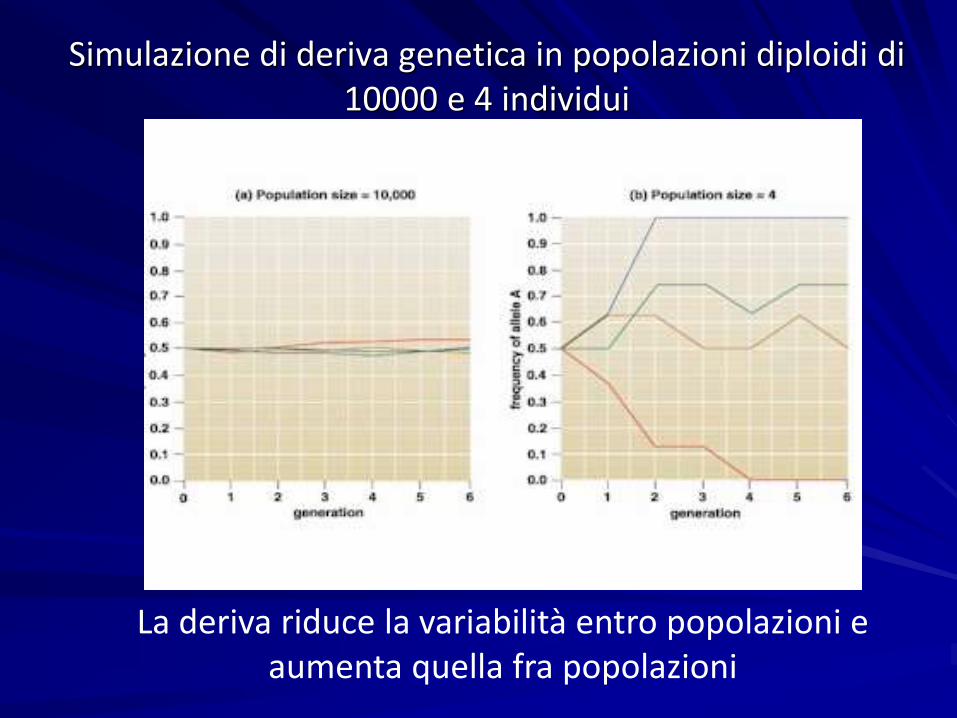
Simulazione di deriva genetica in popolazioni diploidi di10000 e 4 individui
La deriva riduce la variabilità entro popolazioni e aumenta quella fra popolazioni
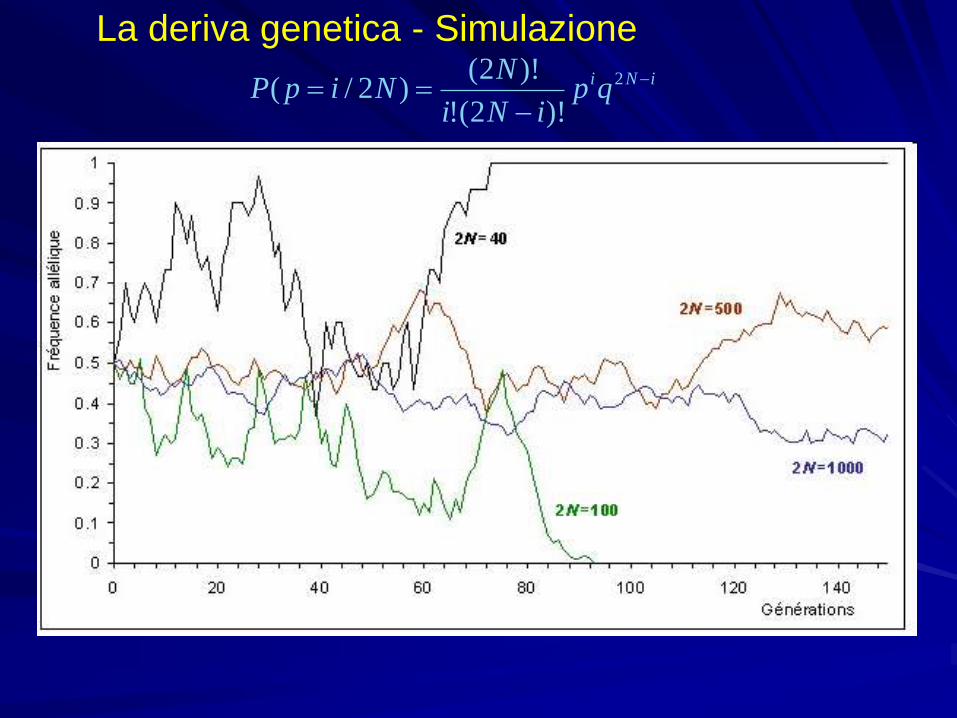
La deriva genetica - SimulazioneiNiqp
iNi
NNipP
2
)!2(!
)!2()2/(

Le conseguenze della deriva genetica sul grado di
variabilità intra- ed interpopolazioni (1)
La deriva genetica determina una diminuzione MEDIA del grado di var.
intrapopolazione (diminuzione della eterozigosità), ad esempio
determinando la fissazione di alleli (loci monomorfici).
Può anche determinare un aumento della variabilità stessa (es. una
freq. allelica passa da 0.6 a 0.5 per effetto della deriva maggiore
eterozigosità)
agisce in modo casuale, quindi determinerà un aumento MEDIO del
grado di diversità interpopolazioni (FST), portando le popolazioni a
divergere in relazione alle frequenze alleliche di un determinato locus.
Ciò non toglie che (come sopra) possa verificarsi una diminuzione di
FST, con popolazioni inizialmente differenti che vengono, per caso (per
effetto della deriva), ad avere le stesse frequenze alleliche

Considerando tuttavia più loci contemporaneamente
avremo sempre:
Diminuzione della eterozigosità (var. intra)
Aumento di FST (var. inter)
Le conseguenze della deriva genetica sul grado di
variabilità intra- ed interpopolazioni (2)
Entrambi gli effetti si accentuano con il passare delle generazioni
conseguenze

Gli effetti della deriva genetica sulla variabilità
interpopolazioni
+
+++
++++
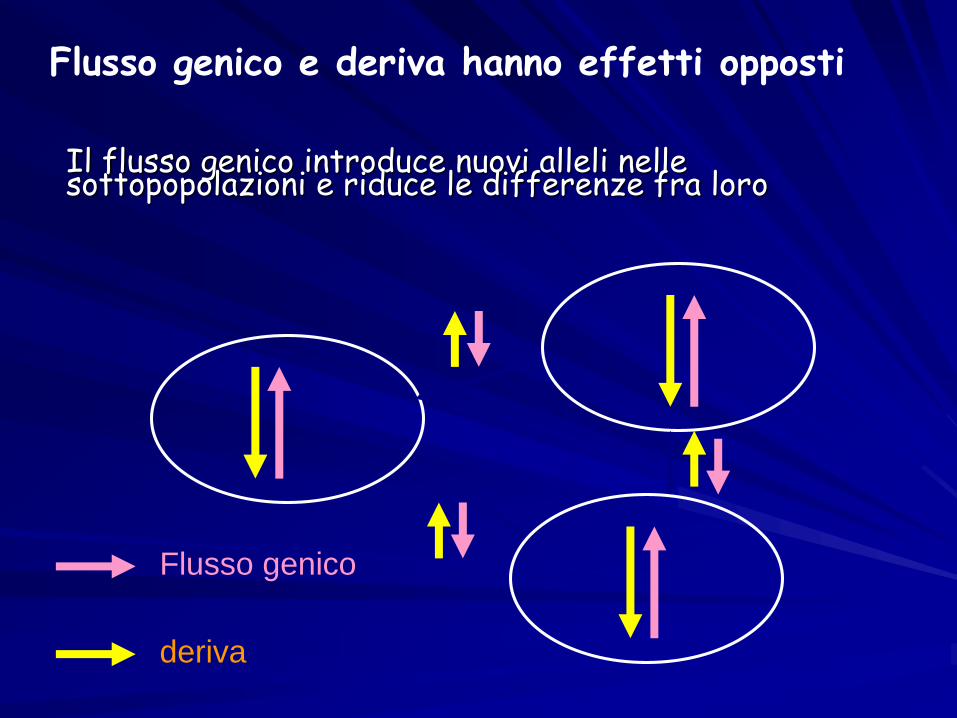
Il flusso genico introduce nuovi alleli nelle sottopopolazioni e riduce le differenze fra loro
Flusso genico
deriva
Flusso genico e deriva hanno effetti opposti
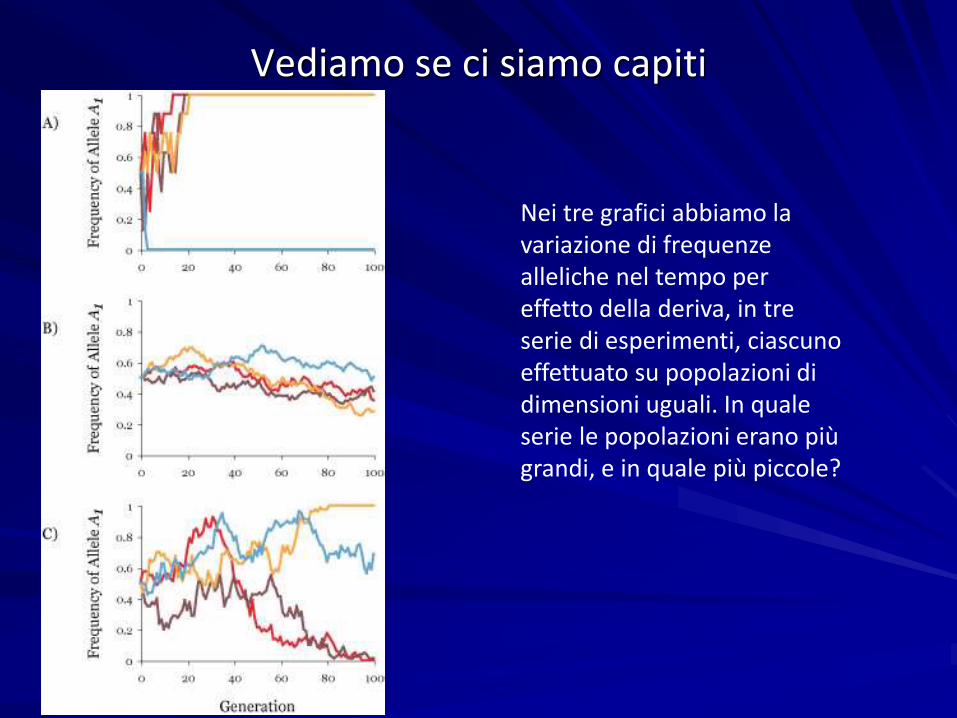
Vediamo se ci siamo capiti
Nei tre grafici abbiamo la variazione di frequenze alleliche nel tempo per effetto della deriva, in tre serie di esperimenti, ciascuno effettuato su popolazioni di dimensioni uguali. In quale serie le popolazioni erano più grandi, e in quale più piccole?

Perché è importante la derivagenetica?
Importanza evolutiva: cambiamento non adattativo, specie in piccole popolazioni
Importanza per la conservazione: perdita di diversitàgenetica, specie in piccole popolazioni
Importanza biomedica: alleli patologici altrove raripossono essere comuni in piccole popolazioni
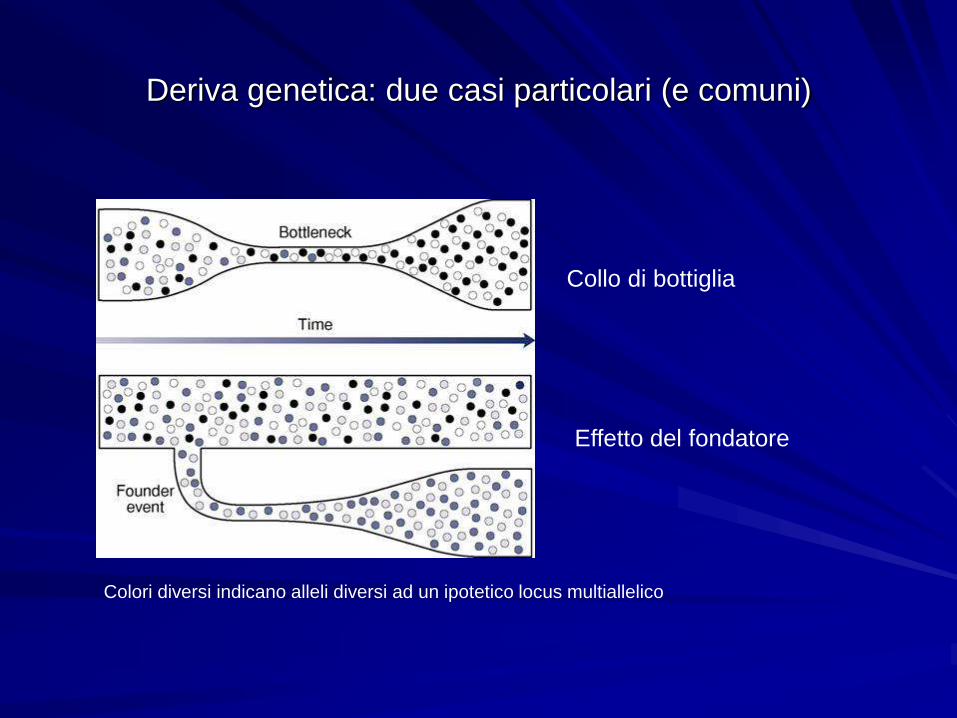
Deriva genetica: due casi particolari (e comuni)
Collo di bottiglia
Effetto del fondatore
Colori diversi indicano alleli diversi ad un ipotetico locus multiallelico
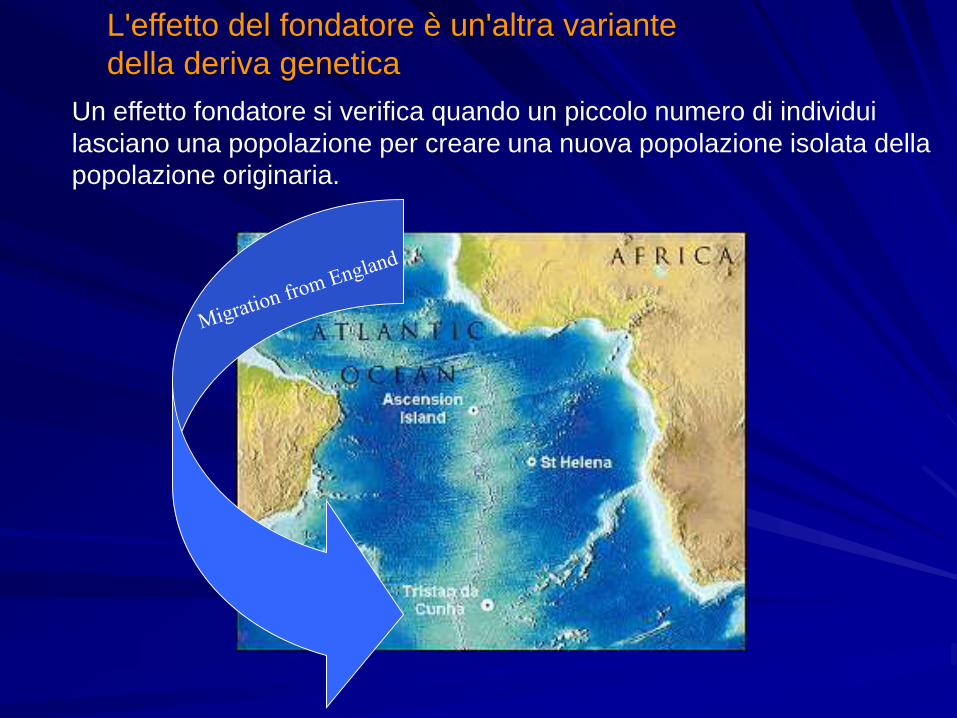
L'effetto del fondatore è un'altra variante
della deriva genetica
Un effetto fondatore si verifica quando un piccolo numero di individui
lasciano una popolazione per creare una nuova popolazione isolata della
popolazione originaria.

The Founder Effect is Another Variation of Genetic Drift
L’isola di Tristan da Cunha in
Atlantico del sud, è stata
colonizzata da 15 britannici nel
1814, uno di loro ha portato un
allele per pigmentosum retinite.
Tra i loro 240 discendenti che
vivono sull'isola oggi, quattro
sono ciechi dalla malattia e 9
altri sono portatori.

The Founder Effect
Popolazione Amish derivano da qualche decina coloni sfuggiti delle
persecuzione religiose in Germania nel 1719, per stabilirsi in
Pennsylvania.
La comunità è chiusa.
Le frequenze alleliche delle malattie genetiche in Amish sono
significativamente diversi dal ancestrale populazione tedesca e dalle
popolazioni locali nei dintorni.

Donna Amish con il figlioportatore della sindromeEllis–van Creveld: arti corti, dita sopranumerarie e difetticardiaci. L’ effetto fondatorespiega la prevalenza dellasindrome tra gli Amish residenti di Lancaster County, Pennsylvania.
Negli ultimi 40 anni del 20° secolo, 61
bambini con microcefalia sono nati in 23
famiglie Amish (tutte discendenti di una
singola coppia datata 9 generazioni
indietro).
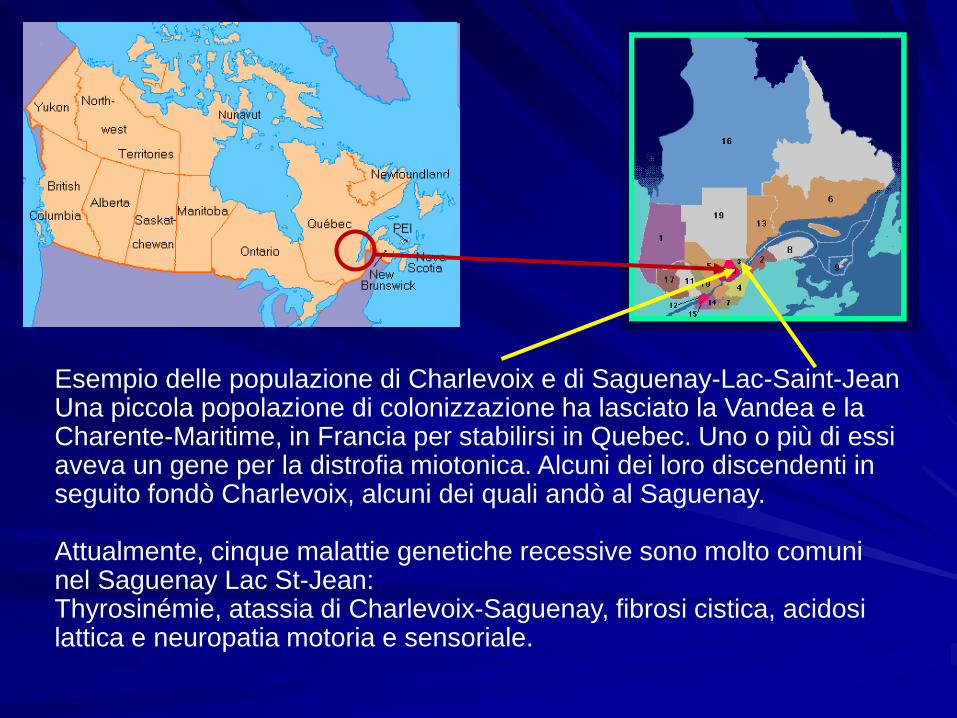
Esempio delle populazione di Charlevoix e di Saguenay-Lac-Saint-JeanUna piccola popolazione di colonizzazione ha lasciato la Vandea e la Charente-Maritime, in Francia per stabilirsi in Quebec. Uno o più di essi aveva un gene per la distrofia miotonica. Alcuni dei loro discendenti in seguito fondò Charlevoix, alcuni dei quali andò al Saguenay.
Attualmente, cinque malattie genetiche recessive sono molto comuni nel Saguenay Lac St-Jean:Thyrosinémie, atassia di Charlevoix-Saguenay, fibrosi cistica, acidosi lattica e neuropatia motoria e sensoriale.
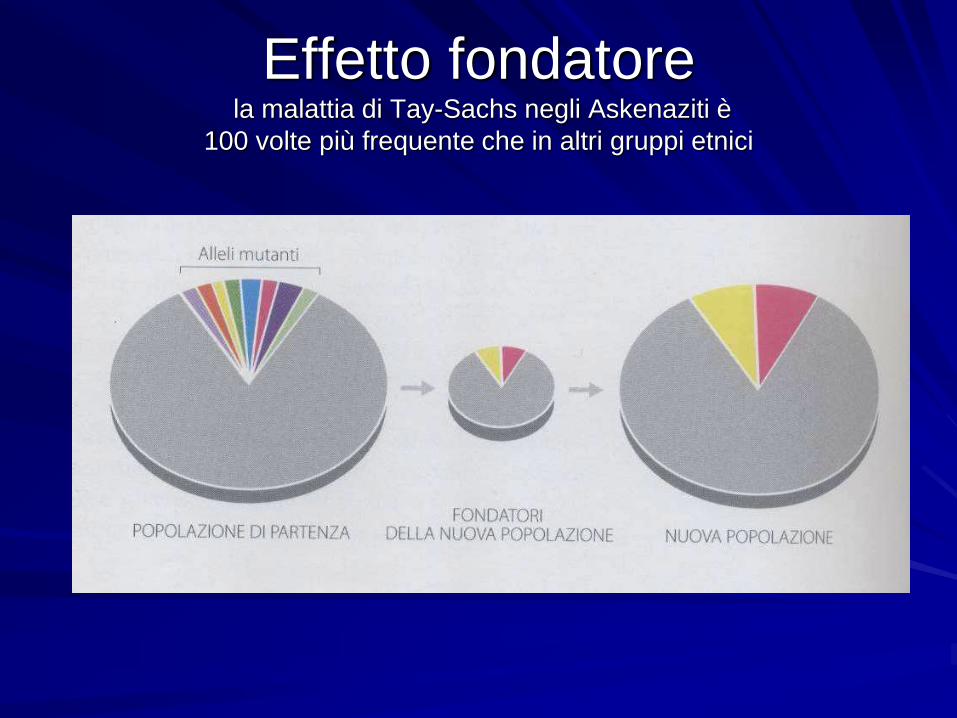
Effetto fondatorela malattia di Tay-Sachs negli Askenaziti è
100 volte più frequente che in altri gruppi etnici

Effetto fondatore
Nel Maracaibo (Venezuela): alta frequenza della Corea di
Huntington's.
L’origne della malattia si fa risalire ai primi del 19° secolo, quando
una donna, Maria Concepción Soto, si trasferì nella zona. Lei
(affetta da tale patologie) ha avuto numerosi figli ("founder“).
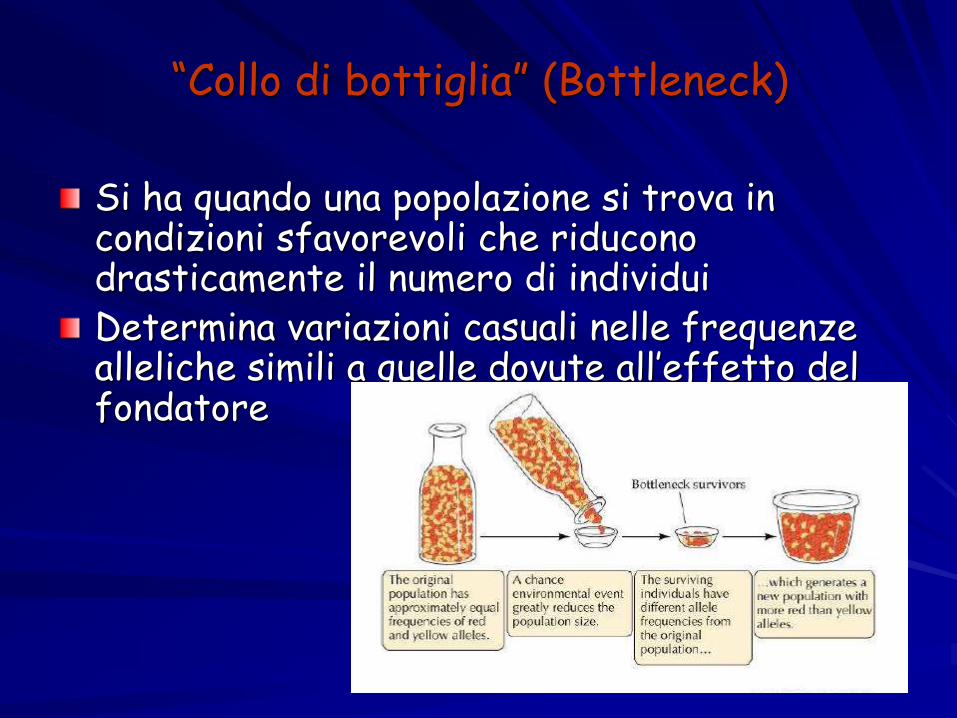
“Collo di bottiglia” (Bottleneck)
Si ha quando una popolazione si trova in condizioni sfavorevoli che riducono drasticamente il numero di individuiDetermina variazioni casuali nelle frequenze alleliche simili a quelle dovute all’effetto del fondatore

La selezione naturale
E’ il solo fattore evolutivo che permette alle popolazioni di adattarsi al loro ambiente
Può mantenere inalterate le frequenze alleliche o causare il loro cambiamento nel tempo

La selezione naturale
Insieme dei fattori che tendono a favorire, o a sfavorire, la tendenza a riprodursi di un dato genotipo.L'effetto della selezione è l'adattamento; in termini evolutivi indica una variazione delle caratteristiche di un organismo in relazione all'ambiente.In senso darwiniano la selezione naturale può essere definita come la riproduzione differenziale di varianti genetiche alternative.

Gli individui sono incapaci di evolvere "geneticamente parlando".
Si limitano passare i loro geni quando possono.
Di generazione in generazione, i tratti determinati da questi geni aumento della popolazione che "evolve".
La selezione naturale agisce sugli individui, ma solo le popolazioni evolvono

SELEZIONE NATURALE
Teoria di Lamarck : Lamarckismo 1809
Motore dell’ evoluzione : ambiente
Teoria di Darwin : Darwinismo 1859
Motore dell’ evoluzione : selezione naturale

FITNESS (w)
Misura l’efficienza riproduttiva di un dato genotipoComponenti della fitness:sopravvivenzatasso di svilupposuccesso nell’accoppiamentofertilità (n. gameti)

Coefficiente di selezione (s)
La fitness viene espressa anche in termini di coefficiente di selezione: s = 1-wMisura la riduzione di fitness di un datogenotipo rispetto a quello migliore.Ad es. s = 0,01 indica che un dato genotipo ha una probabilità di sopravvivere inferiore dell’1% rispetto al genotipo migliore.

Effetto della selezione naturale
• Selezione contro l’allele recessivo
• Selezione contro l’allele dominante
• Selezione in favore dell’eterozigote
• Selezione contro l’eterozigote

La selezione naturale
Stabilizzante
Divergente
Direzionale
frequenza dipendente
sessuale
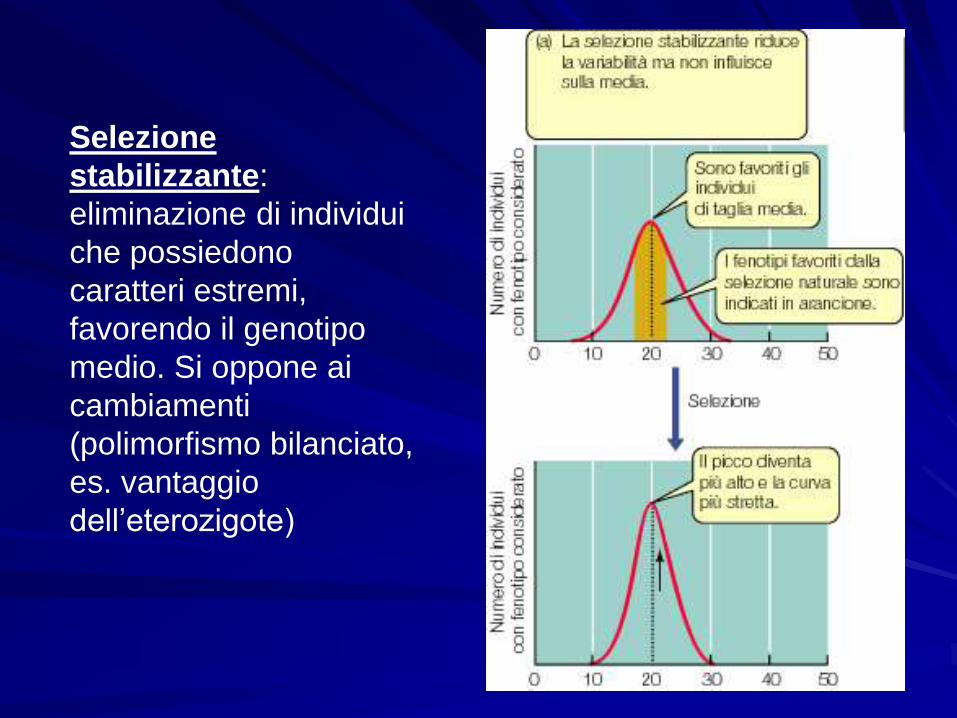
Selezione
stabilizzante:
eliminazione di individui
che possiedono
caratteri estremi,
favorendo il genotipo
medio. Si oppone ai
cambiamenti
(polimorfismo bilanciato,
es. vantaggio
dell’eterozigote)

Selezione in favore dell’eterozigote
Detta anche sovradominanza
Porta ad un equilibrio polimorfico stabile
(polimorfismo bilanciato)
Es. l’anemia falciforme
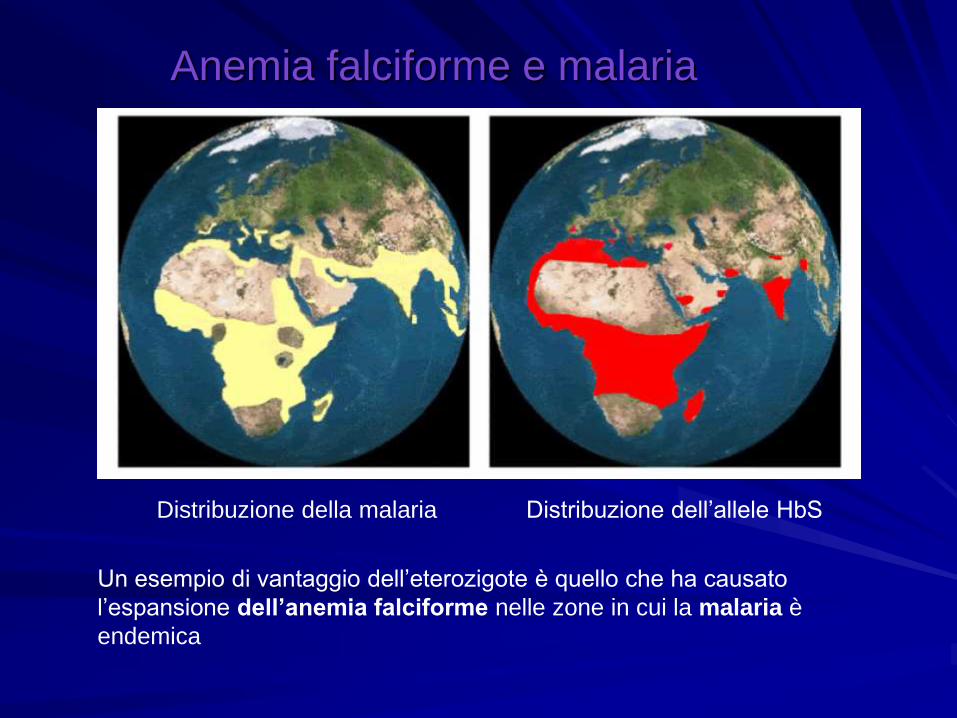
Anemia falciforme e malaria
Distribuzione della malaria Distribuzione dell’allele HbS
Un esempio di vantaggio dell’eterozigote è quello che ha causato
l’espansione dell’anemia falciforme nelle zone in cui la malaria è
endemica
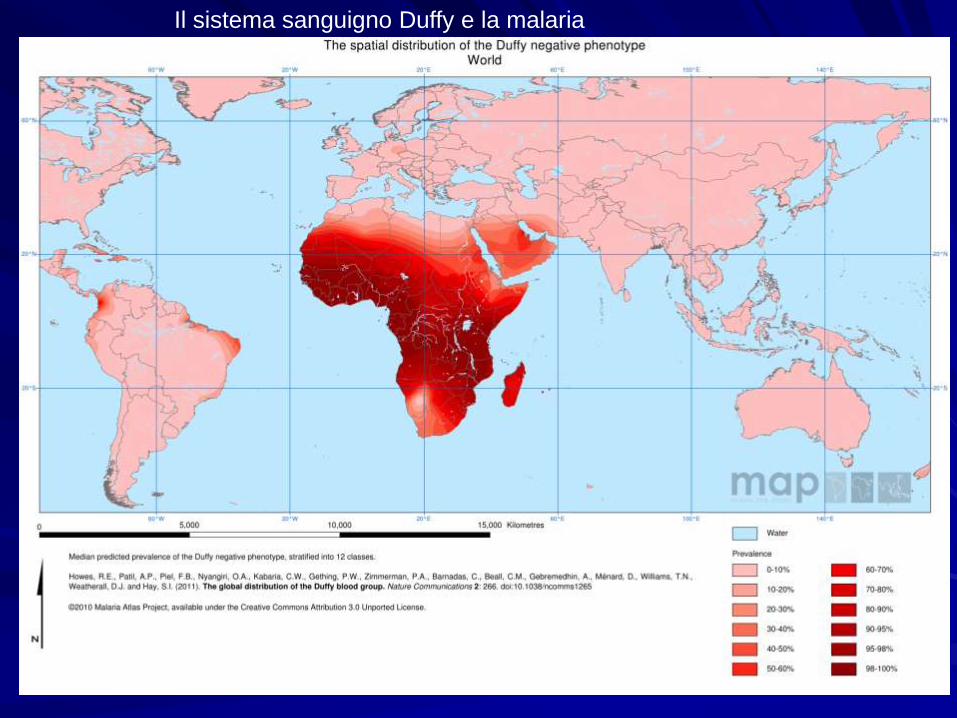
Il sistema sanguigno Duffy e la malaria
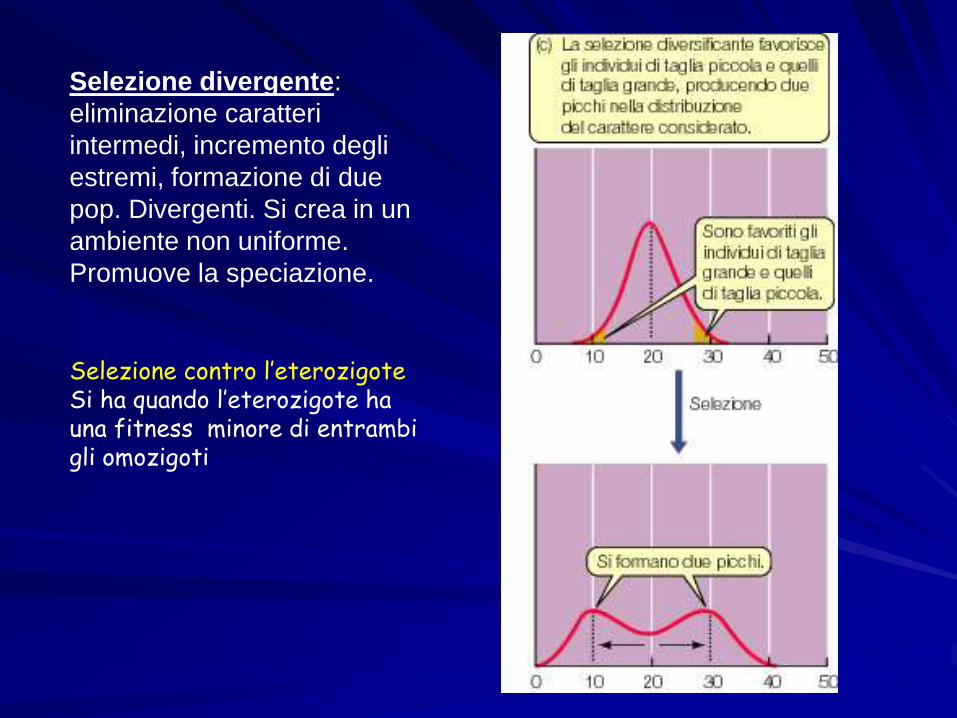
Selezione divergente:
eliminazione caratteri
intermedi, incremento degli
estremi, formazione di due
pop. Divergenti. Si crea in un
ambiente non uniforme.
Promuove la speciazione.
Selezione contro l’eterozigoteSi ha quando l’eterozigote ha una fitness minore di entrambi gli omozigoti
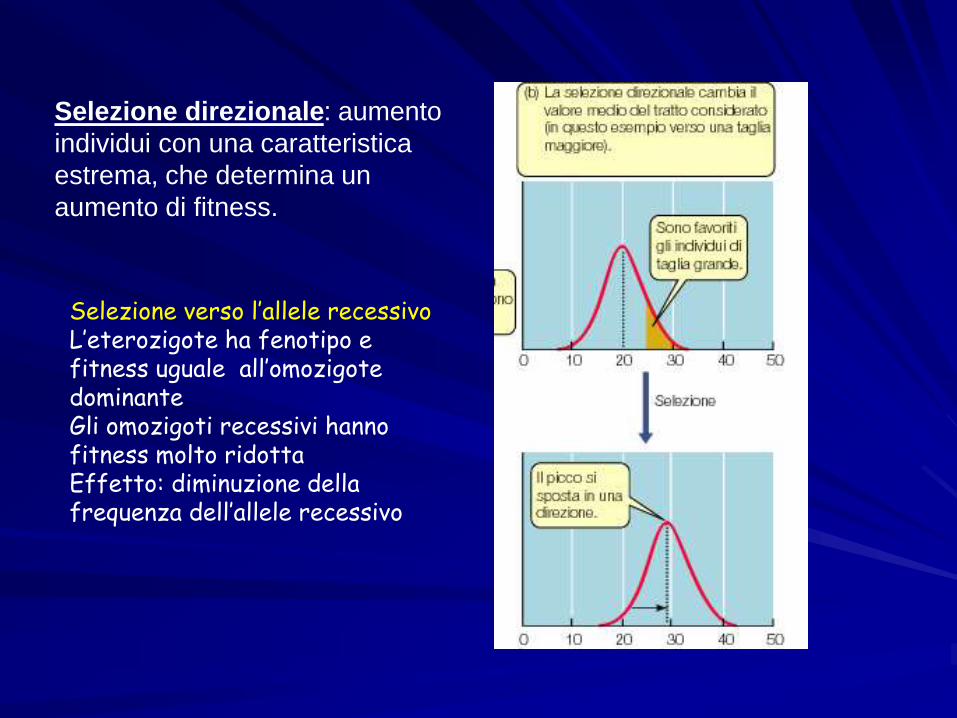
Selezione direzionale: aumento
individui con una caratteristica
estrema, che determina un
aumento di fitness.
Selezione verso l’allele recessivoL’eterozigote ha fenotipo e fitness uguale all’omozigote dominanteGli omozigoti recessivi hanno fitness molto ridottaEffetto: diminuzione della frequenza dell’allele recessivo
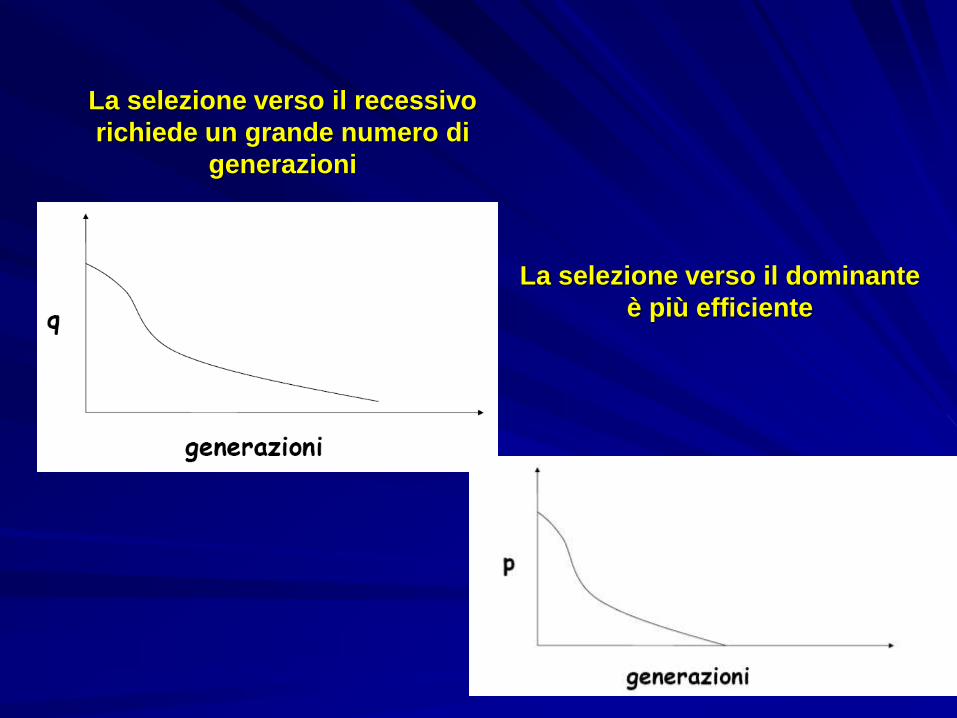
La selezione verso il recessivo
richiede un grande numero di
generazioni
La selezione verso il dominante
è più efficiente

Selezione frequenza-dipendente: riduzione
frequenza fenotipi più diffusi, aumento di quelli meno
frequenti, poiché la competizione tra i fenotipi più
frequenti ridurrà le possibilità di sopravvivenza di quei
fenotipi aumentando quella dei fenotipi rari, soggetti a
minore competizione.
Un esempio analogo è il mancinismo negli umani:
dato che la maggioranza delle persone è destrimane, i
mancini godono di notevoli vantaggi nella lotta e negli
sport come il tennis. Questo vantaggio diminuisce con
la maggiore diffusione del mancinismo; di
conseguenza, la percentuale di mancini nella
popolazione umana si è attestata ad un livello ottimale
pari a circa il 5%

Selezione sessuale: (è una selezione direzionale)
opera sulle caratteristiche che hanno come
conseguenza la conquista del partner: la frequenza
di alcuni fenotipi può aumentare o diminuire in base
all’attrattiva esercitata dall’individuo che possiede
una determinata caratteristica. Avviene in due forme:
intersessuale (es. combattimento maschio contro
maschio) e intersessuale (es. scelta preferenziale
del partner)
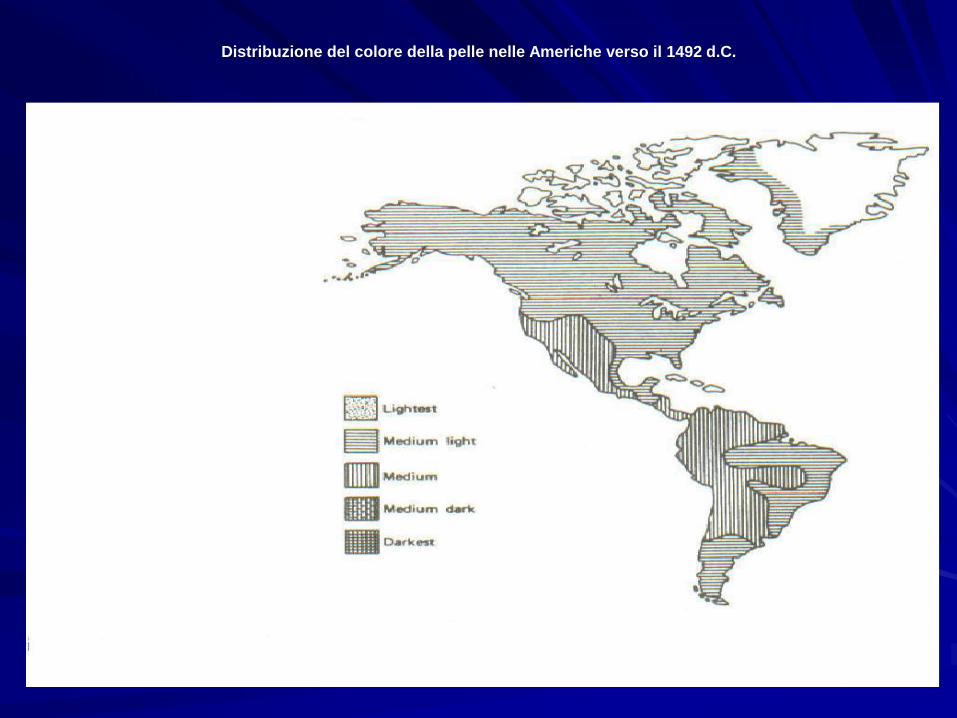
Distribuzione del colore della pelle nelle Americhe verso il 1492 d.C.
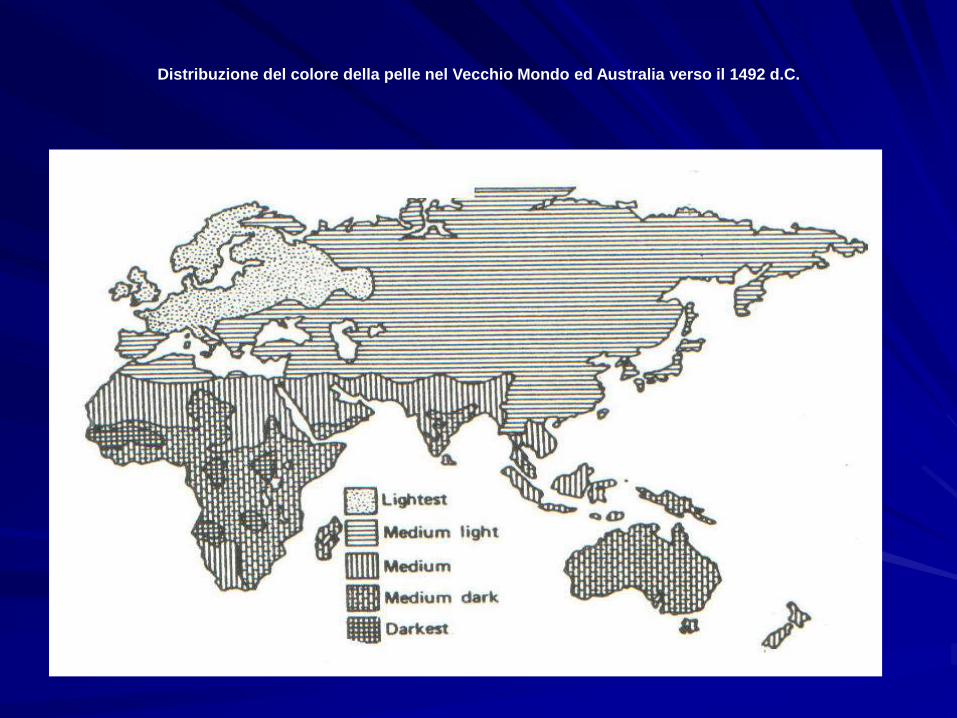
Distribuzione del colore della pelle nel Vecchio Mondo ed Australia verso il 1492 d.C.

La non perfetta corrispondenza tra latitudine e pigmentazione
ha fatto ipotizzare che la variazione geografica del colore della
pelle nelle popolazioni umane sia frutto di un compromesso
selettivo tra due forze controbilanciantesi:
•selezione naturale, che ha favorito gli individui meno
suscettibili al cancro della pelle, alle scottature solari, al deficit
di vitamina D, ed al surriscaldamento. Riguardo a quest’ultimo
punto ricordiamo che i melanodermi assorbono circa un 30% in
più di irradiazione solare rispetto ai leucodermi, e corrono
pertanto soprattutto nei tropici dei rischi di surriscaldamento.
•selezione sessuale, che avrebbe favorito nelle latitudine
elevate una selezione sessuale in favore delle femmine più
chiare, mentre nell’Africa sub-Sahariana la selezione sessuale
sarebbe stata positiva per i maschi più scuri.

La teoria della selezione sessuale si basa sull’ipotesi:
•che nelle zone artiche ci sia stato un surplus di femmine in
seguito ad un’elevata mortalità maschile attribuita ai rischi
conseguenti alle modalità dell’attività di caccia (Frost, 1994).
Inoltre, date le particolari condizioni climatiche, ci sarebbe stato
uno scarso contributo alla sussistenza da parte della raccolta
effettuata dalla componente femminile della comunità, il che
avrebbe scoraggiato la poligamia;
•che nell’Africa sub-Sahariana in un regime matrimoniale di tipo
poligamico ci sia stato un surplus di maschi, per una ridotta
mortalità maschile, pertanto i maschi con aspetto maggiormente
maschile sarebbero stati favoriti nella scelta sessuale. Ciò
giustificherebbe il fatto che le popolazioni africane sub-Sahariani
siano più scure delle altre popolazioni umane situate nei tropici
(Frost, 1994).

Accoppiamento assortativo
Individui con un dato genotipo hanno più probabilità diaccoppiarsi con individui di un altro genotipo di quanto atteso inbase alle loro frequenze
L’accoppiamento assortativo cambia le frequenze genotipiche
• Se la probabilità di accoppiamento fra genotipi simili è maggiorerispetto a quanto atteso in base al caso, la frequenza degliomozigoti tenderà ad aumentare
• Se tale probabilità è minore, la frequenza degli omozigotitenderà a diminuire

Selezione e mutazione
• Il risultato finale della selezione è
l’eliminazione dell’allele
• La mutazione fa sì che questi alleli
rimangano nella popolazione
• Selezione e mutazione hanno effetti
opposti

Effetti della selezione sul genoma
L’azione della selezione naturale si manifesta solo in
regioni limitate del genoma. L’azione esercitata su un sito
genomico ha degli effetti anche sui siti adiacenti (in
relazione al tasso di ricombinazione). Possiamo avere un
aumento di frequenza di una mutazione e quindi anche
delle mutazioni neutrali legate a quel sito (genetic hitch-
hiking). Al contrario le varianti non associate all’allele
vantaggioso saranno perse, determinando una diminuzione
della variabilità (selective sweep).
Invece si parla di background selection in caso di
pressione selettiva negativa: viene eliminato l’allele
svantaggioso e tutta la variabilità ad esso associata.

Esempi nell’uomo
Persistenza della lattasi. A selective sweep occurred for the Europeans
and the ability to get nutrition from milk and milk products was highly
positively selected. Therefore, the majority of Europeans possessed the
ability to make lactase. Other genes hitchhiked along with this selection. In
fact, researchers estimate that about a million base pairs of DNA
hitchhiked along with the sequence that coded for the lactase enzyme.
Colore della pelle. As human ancestors moved from Africa (where dark
skin is a necessary protection against the direct ultraviolet rays of the sun),
to Europe and Asia, gradually lost the dark pigmentation in favor of a
lighter coloring for the skin. Nearby alleles that controlled the rate of
metabolism hitchhiked along. It is proposed that the skin pigmentation
gene and the metabolic rate gene were involved in the same selective
sweep in the early human ancestors.





























