Embed Size (px)
Citation preview

Arabidopsis Microtubule-Associated Protein MAP65-3Cross-Links Antiparallel Microtubules toward Their Plus Endsin the Phragmoplast via Its Distinct C-Terminal MicrotubuleBinding Domain W
Chin-Min Kimmy Ho, Yuh-Ru Julie Lee, Lindsay D. Kiyama, Savithramma P. Dinesh-Kumar, and Bo Liu1
Department of Plant Biology, University of California, Davis, California 95616
Plant cytokinesis is brought about by the phragmoplast, which contains an antiparallel microtubule (MT) array. The MT-associated protein MAP65-3 acts as an MT-bundling factor that specifically cross-links antiparallel MTs near their plus ends.MAP65 family proteins contain an N-terminal dimerization domain and C-terminal MT interaction domain. Compared withother MAP65 isoforms, MAP65-3 contains an extended C terminus. A MT binding site was discovered in the region betweenamino acids 496 and 588 and found to be essential for the organization of phragmoplast MTs. The frequent cytokinetic failurecaused by loss of MAP65-3 was not rescued by ectopic expression of MAP65-1 under the control of the MAP65-3 promoter,indicating nonoverlapping functions between the two isoforms. In the presence of MAP65-3, however, ectopic MAP65-1appeared in the phragmoplast midline. We show that MAP65-1 could acquire the function of MAP65-3 when the C terminus ofMAP65-3, which contains the MT binding site, was grafted to it. Our results also show that MAP65-1 and MAP65-3 may shareredundant functions in MT stabilization. Such a stabilization effect was likely brought about by MT binding and bundling. Weconclude that MAP65-3 contains a distinct C-terminal MT binding site with a specific role in cross-linking antiparallel MTstoward their plus ends in the phragmoplast.
INTRODUCTION
In plants, cytokinesis is executed by the phragmoplast, whichcontains a framework of two opposing sets of microtubules(MTs) with their plus ends facing each other at or near the celldivision site (Staehelin and Hepler, 1996). Golgi-derived vesiclesare transported by motors along MTs toward the plus end inorder for the cell plate materials they carry to be deposited in thephragmoplast midzone (Jürgens, 2005). The phragmoplast MTarray, which is derived from the spindle midzone MTs during lateanaphase and telophase, exhibits high dynamicity (Zhang et al.,1990). Concomitant with cell plate formation, MT polymeriza-tion takes place at the phragmoplast periphery, while depoly-merization happens in the central region where vesicle transportand fusion have already been completed. In other words, pro-gressive MT turnover is accompanied by the expansion of thephragmoplast and cell plate toward to the parental cell wall.
Phragmoplast MTs exhibit a high degree of dynamic instabilitythat is required for the expansion of the array (Lloyd, 2011;Smertenko et al., 2011). The organization of the phragmoplastMT array is dependent on factors that regulate specific activi-ties, such as MT nucleation, polymerization, and bundling. Forexample, mutations in genes encoding g-tubulin and augmin
cause serious defects in MT nucleation and consequently resultin collapse of the array in Arabidopsis thaliana (Pastugliaet al., 2006; Zeng et al., 2009; Ho et al., 2011b). Mutations ina gene encoding MICROTUBULE ORGANIZATION1/GEMINIPOLLEN1 of the Xenopus laevis MT-ASSOCIATED PROTEIN215/DEFECTIVE IN SISTER CHROMATID SEPARATION1 familycause fragmentation and branching of the phragmoplast MTarray, suggesting that its role in promoting MT polymerization iscritical for the dynamics of the array (Eleftheriou et al., 2005;Kawamura et al., 2006; Oh et al., 2010). The loss of the MTplus end tracking protein END BINDING1c also causes a similarphenotype upon challenge with low doses of the herbicideoryzalin (Liu et al., 2011). In addition, MT bundling activitiesconferred by the MT-associated proteins (MAPs) in the MAP65/ANAPHASE SPINDLE ELONGATION1 (Ase1)/PROTEIN REGU-LATING CYTOKINESIS1 (PRC1) family are also critical for theorganization of the bipolar MT array (Liu et al., 2011). Besidesthe aforementioned proteins, the Kinesin-12 MT motors playcritical roles in maintaining the MT plus ends in the phragmo-plast midzone, likely by pushing newly polymerized MT seg-ments apart (Lee et al., 2007).It is believed that concerted roles of these regulatory proteins
allow the phragmoplast MT array to sustain its bipolar configu-ration while expanding toward the cell periphery (Liu et al.,2011). However, it is intriguing how the two sets of antiparallelMTs are engaged dynamically through the rapid progressionof cytokinesis. Electron microscopy tomography indicates thatthe majority of the antiparallel MTs do not interdigitate in thephragmoplast midzone (Austin et al., 2005). This study led tothe hypothesis that MTs are anchored and stabilized by the
1Address correspondence to [email protected] author responsible for distribution of materials integral to the findingspresented in this article in accordance with the policy described in theInstructions for Authors (www.plantcell.org) is: Bo Liu ([email protected]).WOnline version contains Web-only data.www.plantcell.org/cgi/doi/10.1105/tpc.111.092569
The Plant Cell, Vol. 24: 2071–2085, May 2012, www.plantcell.org ã 2012 American Society of Plant Biologists. All rights reserved.

amorphous cell plate assembly matrix in the phragmoplast.However, this hypothesis contradicts observations that a numberof proteins, including MAP65 members and Kinesin-12, con-spicuously appear in the midzone in the phragmoplast (Lee andLiu, 2000; Smertenko et al., 2000, 2008; Pan et al., 2004). Amongthem, MAP65-3 plays a critical role in engaging the antiparallelMTs because mutations in the corresponding gene lead toa wider midzone and consequently result in frequent cytokinesisfailures (Müller et al., 2004; Caillaud et al., 2008; Ho et al., 2011a).
Following the initial discovery of the;65-kD MAP65 protein inthe tobacco (Nicotiana tabacum) (Chang-Jie and Sonobe, 1993),nine homologous proteins were predicted to be encoded bythe Arabidopsis genome and shared 28 to 79% amino acididentity with the tobacco protein (Hussey et al., 2002). Proteinsin this family form dimers that cross-link MTs (Smertenko et al.,2004; Li et al., 2007; Gaillard et al., 2008; Kapitein et al., 2008).Besides MAP65-3, other MAP65 isoforms, such as MAP65-1and MAP65-5, also decorate the phragmoplast midzone asvisualized by immunostaining using antibodies raised againstantigens derived from the corresponding proteins (Smertenkoet al., 2008). However, unlike MAP65-3, which is restricted inthe midline where it is concentrated with MT plus ends, MAP65-1shows a broader localization pattern along MTs in the phrag-moplast midzone (Smertenko et al., 2008). On the other hand,a MAP65-4-green fluorescent protein (GFP) fusion associateswith kinetochore fibers in mitotic spindles but not the phrag-moplast midzone, despite the fact that the isoform is moreclosely related to MAP65-3 than to MAP65-1 or MAP65-5 (Husseyet al., 2002; Guo et al., 2009; Fache et al., 2010). Therefore, itremains unclear how the interaction between MAP65 isoformsand MTs of different arrays is established. Different patterns ofMT association suggest Arabidopsis MAP65 isoforms may as-sume divergent functions. However, a recent study revealedpossible functional redundancy among MAP65-1, MAP65-2, andMAP65-3 in Arabidopsis, and knocking out the genes encodingMAP65-1 or MAP65-2 did not seem to cause obvious cell di-vision defects (Lucas et al., 2011; Sasabe et al., 2011).
Because the loss of MAP65-3 alone causes severe defects incytokinesis, we were prompted to further analyze the structuralfeature that enables the protein to play its critical role in thephragmoplast. Sequence alignment of Arabidopsis MAP65family proteins shows relatively conserved N-terminal halvesand variable C-terminal halves (Smertenko et al., 2008). TheN-terminal part is required for protein dimerization (Smertenkoet al., 2004; Kapitein et al., 2008). When the C-terminal halvesof MAP65 proteins were further divided into the MTB1 (for MTbinding domain 1) and MTB2 domains, the C-terminal-mostMTB2 domains exhibited the most divergence (Smertenko et al.,2008). For simplicity, MTB1 and MTB2 domains of the C ter-minus are referred in this article as C1 and C2, respectively.Biochemical studies showed that the C1 domains of MAP65-1,-4, and -5 possess MT binding activity (Smertenko et al., 2004;Gaillard et al., 2008; Fache et al., 2010). A recent structuralanalysis of the human counterpart of MAP65, PRC1, alsoshowed that its most C-terminal region facilitated MT binding(Subramanian et al., 2010). Compared with other isoforms,MAP65-3 contains an extended C2 domain and is responsiblefor the slow turnover rate (half-time = 143 s) of the protein in the
phragmoplast (Smertenko et al., 2008). However, it is unclearwhether this domain is responsible for the distinct localizationpattern of the protein and critical for its role in cytokinesis.To examine whether the divergent C2 domain of MAP65-3
might contribute to its unique role, we expressed full-length andseveral truncated MAP65-3 proteins in the dyc283 map65-3 nullmutant background. When compared with a functional full-length MAP65-3 fusion protein, the deletion of the C2 domainabolished the localization and function of MAP65-3. MAP65-1was also tested as a substitute for the role of MAP65-3 in cy-tokinesis. Only when the C2 domain of MAP65-3 was fused toMAP65-1 did the fusion protein decorate the phragmoplastmidline and largely restore successful cytokinesis. We reporthere that the C2 domain of MAP65-3 contains a MT binding sitethat enables specific activity in the phragmoplast midzone.
RESULTS
The Function of MAP65-3 Is Dependent on ItsC-Terminal Domain
Among members of the MAP65 family, the C-terminal halves arehighly variable compared with the more or less conservedN-terminal halves (Smertenko et al., 2008). As reported by oth-ers (Müller et al., 2004; Caillaud et al., 2008), the loss of MAP65-3 caused obvious defects in cell division and seedling growththat can be reflected by dwarf shoots and shortened roots. Thedefects were completely suppressed by the expression of full-length MAP65-3 fused with the FLAG epitope and expressedunder the control of its native promoter (Figures 1A to 1D). Be-cause the C2 domain in the C-terminal half has a 119–aminoacid region that is not present in MAP65-1, we asked whether theextended sequence was responsible for the functional differencebetween MAP65-3 and MAP65-1. MAP651-588, resulting from thedeletion of this 119–amino acid region, was expressed in fusionwith FLAG under the control of the MAP65-3 promoter. The fusionprotein almost completely rescued the defects in root growth atthe seedling stage and shoot growth at the mature stage causedby the dyc283 mutation (Figures 1A to 1D). The deletion of theentire C2 domain, however, failed to rescue the dyc283 mutation,indicating that this MAP65-31-495-FLAG fusion protein did notretain the function of MAP65-3 (Figures 1A to 1D).A published study showed that MAP65-1 appeared in the
phragmoplast midzone in cultured Arabidopsis cells (Smertenkoet al., 2008). A more recent genetic study suggested thatMAP65-1 and MAP65-3 exhibited functional redundancy (Sasabeet al., 2011). Therefore, we tested whether expression of MAP65-1could substitute MAP65-3 in performing its antiparallel MTbinding function. Overall growth defects caused by the dyc283mutation were partially suppressed, albeit not to a high degree,when MAP65-1 was expressed under the control of the MAP65-3 promoter (Figures 1A to 1D). Because the C2 domain is re-quired for the function of MAP65-3, we swapped the C-terminalfragment of MAP65-1 for the C2 domain of MAP65-3 and ex-pressed a FLAG-fusion of this chimeric protein under the controlof the MAP65-3 promoter. This MAP65-1swap fusion protein wasable to significantly rescue, although incompletely, the growthdefects of the dyc283 mutant (Figures 1A to 1D). Collectively,
2072 The Plant Cell

the dyc283 seedlings formed stunted roots. While MAP65-31-495
could not enhance root growth, MAP65-1 slightly restored rootlength to 25.0% of the wild-type control from 14.8% in dyc283mutant (Figures 1B and 1C). MAP65-1SWAP restored the rootgrowth to 44.8% and MAP65-31-588 restored it to 66.6% (Fig-ures 1B and 1C). Consistent with the root growth phenotypes,mature plants also showed corresponding suppression effectsby expressing these versions of proteins. While MAP65-31-495
and MAP65-1 hardly made any difference when expressed inthe dyc283 mutant, MAP65-1SWAP and MAP65-31-588 enabledthe production of mature plants that were significantly healthierthan dyc283. These results suggest that the C2 domain is criticalfor the function of MAP65-3. This domain could enable MAP65-1to acquire the function of MAP65-3 in the phragmoplast.
Remediation of Defects in Phragmoplast MT Organizationand Cytokinesis by MAP65 Proteins and Derivatives
Published studies showed that the loss of MAP65-3 led toa disorganized MT array in the phragmoplast that showeda wider gap in the phragmoplast midzone when compared withwild-type cells (Müller et al., 2004; Ho et al., 2011a). Whenphragmoplast MTs were revealed by immunofluorescence, cleardifferences were detected in the dyc283 mutant cells comparedwith the wild-type control (Figure 2A, panels a and b). While MT
bundles are tightly packed and organized in the wild-typephragmoplasts, the mutant phragmoplasts possessed bundlesthat were loosely packaged and disorganized. Besides the widegap in the phragmoplast midzone, the overall length of thephragmoplast MT array appeared to be longer (left bracket,Figure 2A, panel b). The overall length of the phragmoplast MTarray increased to 4.66 µm in dyc283 cells (n = 26) comparedwith 2.52 µm in wild-type cells (n = 23) (Figure 2B). The width ofthe antitubulin dark gap (right bracket, Figure 2A, panel b) in-creased to 1.15 µm from 0.42 µm (Figure 2C). In addition, emi-nent MTs emanated from the surface of the reforming daughternuclei (arrow, Figure 2A, panel b). Both the full-length MAP65-3and truncated MAP65-31-588 fusions restored phragmoplast MTarrays to the normal organization patterns seen in the controlcells (Figure 2A), and their phragmoplast length and gap werecomparable to those of the wild-type control cells (Figures 2Band 2C). However, the expression of MAP65-31-495 or MAP65-1did not significantly alter the loose and disorganized phragmo-plast MTs when compared with the dyc283 root cells (Figure2A). The widths of the dark gaps were similar to those in dyc283cells (Figure 2C). However, the length of the phragmoplast MTarray was shorter in cells expressing MAP65-31-495 than indyc283 cells (Figure 2B). The expression of either MAP65-31-495
or MAP65-1 also did not alter the appearance of abundant MTs
Figure 1. Suppression of dyc283 by MAP65 Variants.
(A) Bar graphs represent MAP65-3, MAP65-1, and their derivatives. Under the control of the endogenous MAP65-3 promoter, they were expressed infusions with the FLAG tag at the C terminus in the dyc283 mutant background. The numbers shown are the amino acid positions in the polypeptides.The C-terminal half is divided into the C1 and C2 domains.(B) Growth phenotypes of 5-d-old seedlings of the control wild-type (WT), dyc283, and transformants expressing the designated FLAG fusion proteins.Note the difference in root lengths. Bar = 1 cm.(C) Quantitative analysis of root lengths of 5-d-old seedlings of the control and transformants (n = 57, 50, 72, 63, 66, 77, and 51 for wild-type, dyc283,and mutant plants expressing MAP65-3, MAP65-31-588, MAP65-31-495, MAP65-1, and MAP65-3swap, respectively). Error bars represent SD.(D) Growth phenotypes of mature plants of wild-type, dyc283, and mutant plants expressing MAP65-3, MAP65-31-588, MAP65-31-495, MAP65-1, andMAP65-3swap. Note that MAP65-1SWAP suppresses the dyc283 mutation.
MAP65-3 Contains a Unique C Terminus 2073

Figure 2. Assays of Phragmoplast MT Organization and Cytokinesis in Root Cells in the Wild Type, dyc283, and Various MAP65 Transformants.
(A) The C2 domain of MAP65-3 is important for the integrity of the phragmoplast MT array. At similar stages of cytokinesis, cells of the wild type (WT)(a), dyc283 (b), and dyc283 plants expressing MAP65-3 (c), MAP65-31-588 (d), MAP65-31-495 (e), MAP65-1 (f), and MAP65-1swap (g) exhibit differentpatterns of the phragmoplast array. In (b), the bracket on the left represents the phragmoplast length for the measurement presented in (B) and that onthe right for the gap width for the measurement presented in (C). Eminent MTs radiating from the two reforming daughter nuclei (arrows in [b], [e], and[f]) are observed in cells of dyc283 plants and dyc283 plants expressing MAP65-31-495 and MAP65-1proteins.(B) Quantitative assessment of the phragmoplast length in the cells of the aforementioned lines. The phragmoplasts in MAP65-31-495 (n = 29) andMAP65-1SWAP (n = 30) are significantly reduced compared with that in the dyc283 mutant. Error bars represent SD.(C) Comparison of the width of the phragmoplast midline in wild-type (n = 23), dyc283 (n = 26), and dyc283 plants expressing MAP65-3 (n = 34),MAP65-31-588 (n = 29), MAP65-31-495 (n = 29), MAP65-1 (n = 22), and MAP65-1SWAP (n = 30). The midline is much narrower in cells expressing MAP65-1SWAP than in those of the dyc283 mutant expressing MAP65-1. Error bars represent SD.(D) The root cell profiles of 5-d-old seedlings of wild-type (n = 34), dyc283 (n = 20), and dyc283 plants expressing MAP65-3 (n = 36), MAP65-31-588 (n =35), MAP65-31-495 (n = 21), MAP65-1 (n = 28), and MAP65-1swap (n = 25). Cell wall stubs can be observed in dyc283 and dyc283 cells expressingMAP65-31-495, MAP65-1, and MAP65-1SWAP (asterisks) but not those expressing MAP65-3 or MAP65-31-588.
2074 The Plant Cell

radiating from the surface of the nuclear envelope (arrows,Figure 2A, panels e and f). The expression of MAP65-1SWAP-FLAG further reduced the phragmoplast length and caused thedark gap in the phragmoplast midzone to be significantly re-duced but not to the level seen in wild-type cells (Figures 2B and2C). These data suggest that redirection of MAP65-1 by theaddition of the C2 domain of MAP65-3 significantly restored MTorganization in the phragmoplast.
Furthermore, we investigated how the aforementionedchanges in phragmoplast MT organization might correlate tohow well cytokinesis was brought about. The wild-type rootscontained organized cell profiles, as indicated by intact cell wallsseparating individual cells (Figure 2D). In the roots of the dyc283mutant, however, cell wall stubs were frequently found, in-dicating failures in cytokinesis (asterisk, Figure 2D). In fact,;38.1% of the dyc283 root cells contained cell wall stubs(Figure 2E). We found that the seed germination rate dropped to66.8% (n = 244) in the dyc283mutant compared with 99.1% (n =222) in the wild-type control. Frequent cytokinetic failures mighthave caused such defects. The expression of either MAP65-3 orMAP651-588 resulted in root cell profiles similar to those in wild-type roots and hardly had any cells show cytokinesis defects(Figures 2D and 2E). Surprisingly, although MAP65-31-495 andMAP65-1 did not appear at the phragmoplast midzone, ex-pression of both of these proteins significantly suppressed thecytokinetic defect caused by the dyc283 mutation (Figures 2Dand 2E). MAP65-1 seemed to exert a greater rescuing effectthan MAP65-31-495 (Figure 2E). The expression of MAP65-1SWAP
even restored cytokinesis to a greater extent, with <5% of rootcells showing cell wall stubs (Figures 2D and 2E). These resultssuggest that the function of MAP65-3 at the phragmoplastmidzone is essential for MT arrangement but that that ofMAP65-1 elsewhere also contributes to MT organization in thephragmoplast, although to a lesser extent.
Localization of MAP65-3-FLAG and Its Derivatives inthe Phragmoplast
We first examined the localization of the MAP65-3-FLAG fusionprotein when expressed in dyc283 mutant cells. Consistent withthe complete rescue of the mutant, the localization of full-lengthMAP65-3 fusion protein recapitulated that of the native protein(Ho et al., 2011a). In cells bearing an early phragmoplast, thefusion protein appeared intensively in the phragmoplast midline,where the antitubulin signal was seen weakly as a dark line influorescent images (Figure 3A, panels a to c). This localizationpattern persisted throughout cytokinesis (Figure 3A, panels d tof). We then examined the localization of the MAP65-31-588-FLAGfusion in cells undergoing cytokinesis. In a cell at the early stage
of cytokinesis, the fusion protein was detected in the midline ofthe phragmoplast, highlighting the dark line of the antitubulinimmunofluorescence (Figure 3B, a to c) and similar to what wasobserved for the full-length fusion. Compared with the full-length protein, this truncated protein showed an extended lo-calization pattern, decorating MTs beyond the phragmoplastmidline (arrows, Figure 3B, panel d), suggesting that it wasprobably associated with parallel MTs as well. Unlike the full-length and MAP651-588 fusions, the complete removal of the C2domain resulted in the MAP65-31-495-FLAG fusion protein nolonger being concentrated at the phragmoplast midline (Figure3C, panels a to f). Instead, the anti-FLAG signal became dis-persed across the cytoplasm at both the early (Figure 3C, panelsa to c) and late (Figure 3C, panels d to f) stages of cytokinesis.The phragmoplast MT array consisted of two fan-shaped halves(Figure 3C, panel b). The abnormal MT organization pattern inmutant plants expressing MAP65-31-495-FLAG (Figure 1B) wasconsistent with the result that expression of this truncatedprotein did not rescue the dyc283 mutant. Thus, the resultssuggest that the localization of MAP65-3 at the phragmoplastmidline is essential for its function. Upon the loss of its locali-zation by the deletion of the C2 domain, its function in thephragmoplast is abolished.Because MAP65-3 is required for the localization of Kinesin-
12 (Ho et al., 2011a), we examined whether the new localizationpattern of MAP651-588 would change the localization of themotor. We also wondered whether the extending MAP651-588
localization zone represented a broadened overlapping region ofantiparallel MTs in the phragmoplast. Unlike MAP65-31-588-FLAG, which showed a wide localization pattern, especially to-ward the phragmoplast periphery (arrows, Figures 4A and 4D),Kinesin-12 remained at the phragmoplast midline (Figures 4Band 4E). Moreover, MAP651-588, but not Kinesin-12, appearedconspicuously at the expanding front of the phragmoplast array.This result suggests that MAP65-31-588-FLAG retained the na-tive protein’s function in allowing proteins like Kinesin-12 tospecifically interact with MT plus ends in the phragmoplastmidline, albeit losing its highly restricted localization in thephragmoplast midline. Again, this is consistent with the com-plementation of the dyc283 mutation by expressing this trun-cated version of MAP65-3. The result also indicated that theregion of amino acids 589 to 707 is required for MAP65-3 todefine its localization specifically at or near MT plus ends.
MAP65-1 Localization in the Phragmoplast Can Be Alteredby MAP65-3
The phenotypic differences between the transgenic plants ex-pressing MAP65-1 and MAP65-1SWAP prompted us to investigate
Figure 2. (continued).
(E) Quantitative analysis of cells with cell wall stubs caused by defective cytokinesis. Compared with the dyc283 mutant, which has the most severecytokinesis defects, the transformant expressing MAP65-31-495 showed significant improvement. Greater improvement can be seen with MAP65-1expression. The expression of MAP65-1SWAP reduces cytokinesis defects to an even greater extent. Less than 1% of wild-type, MAP65-3, and MAP65-31-588 cells exhibited cell wall stubs caused by incomplete cytokinesis. Error bars represent SD.Bars = 5 µm in (A) and 50 µm in (D).
MAP65-3 Contains a Unique C Terminus 2075

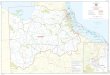
whether these differences were related to different localizationpatterns of the two proteins. Immunostaining of the MAP65-1fusion protein by the anti-FLAG antibody showed punctatesignals along MTs but flanking the phragmoplast midline (Figure5A, panel a). While the conspicuous MAP65-1-FLAG signal wasdetected in the phragmoplast, these dyc283-derived cells didnot possess obvious MAP65-3 signals when probed with anti-MAP65-3 specific antibodies (Figure 5B, panel b). This resultwas surprising because previously published data suggestedMAP65-1 locates in the spindle and phragmoplast midzones(Smertenko et al., 2004, 2008; Mao et al., 2005a). We then testedwhether the localization of MAP65-1 to the phragmoplast mid-zone was dependent on MAP65-3. When the MAP65-1-FLAGfusion was expressed in the wild-type background, in otherwords in the presence of MAP65-3, MAP65-1 was detected inthe phragmoplast midline at all stages of cytokinesis (Figure 5C).These results suggested MAP65-1 localization to the phrag-moplast midline requires MAP65-3 under this artificial condition.A likely scenario was that the ectopically expressed MAP65-1
protein directly interacted with the endogenous MAP65-3. Totest this possibility, we employed the bimolecular fluorescencecomplementation (BiFC) technique using vectors that expresstarget proteins at low levels in Nicotiana benthamiana leaf cells.Because the dimerization domain is located in the N-terminalpart of MAP65 proteins (Smertenko et al., 2004), we expressedthe nCitrine fusion proteins with truncated MAP65-11-339 andMAP65-31-340. BiFC signal highlighting parallel MTs was ob-served due to the self-interactions between MAP65-11-339-nCi-trine and MAP65-1-cCitrine and between MAP65-31-340-nCitrineand MAP65-3-cCitrine (Figures 6A and 6D). Fluorescencewas also established along cortical MTs when MAP65-31-340-nCitrine and MAP65-1-cCitrine were expressed (Figure 6B).Reciprocally, expression of MAP65-11-339-nCitrine and MAP65-3-cCitrine also rendered a positive BiFC (Figure 6C). To confirmthe specificity of the interactions and ascertain that the ob-served fluorescence was not due to accidental association be-tween nCitrine and cCitrine, the motor domain of Kinesin-12Athat harbors a MT binding site was used in place of full-lengthMAP65-1 or MAP65-3. No fluorescence was observed whenKinesin-12A61-425-cCitrine was coexpressed together with eitherMAP65-11-339-nCitrine or MAP65-31-340-nCitrine (Figures 6Eand 6F). These results further suggest that MAP65-1 andMAP65-3 would likely interact with each other when their ex-pressions were temporally coordinated. Because of such aninteraction, MAP65-3 could bring MAP65-1 to the phragmoplastmidline when both were present during cytokinesis.
Figure 3. The C2 Domain Is Required for MAP65-3 Localization to thePhragmoplast Midline.
The merged images have fusion proteins of MAP65-3 and its derivativesin green, MTs in red, and DNA in blue. Bar = 5 µm.(A) Localization of MAP65-3-FLAG. During early ([a] to [c]) and late ([d]to [f]) stages of cytokinesis. MAP65-3 consistently associates with thephragmoplast midline.(B) Localization of MAP65-31-588-FLAG. (a) to (c) The fusion protein lo-calizes to the phragmoplast midline during the early stage of cytokinesis.(d) to (f) In a mature phragmoplast, MAP65-31-588-FLAG exhibits
a broader localization pattern (arrows in [d]) along phragmoplast MTsand across the midline.(C) Localization of MAP65-31-495-FLAG. The fusion protein is detected asdiffuse signals across the cytoplasm in both the early ([a] to [c]) and late([d] to [f]) stages of cytokinesis.
2076 The Plant Cell

The C2 Domain of MAP65-3 Can Redirect MAP65-1 toBundled Antiparallel MTs in the Phragmoplast Midzone
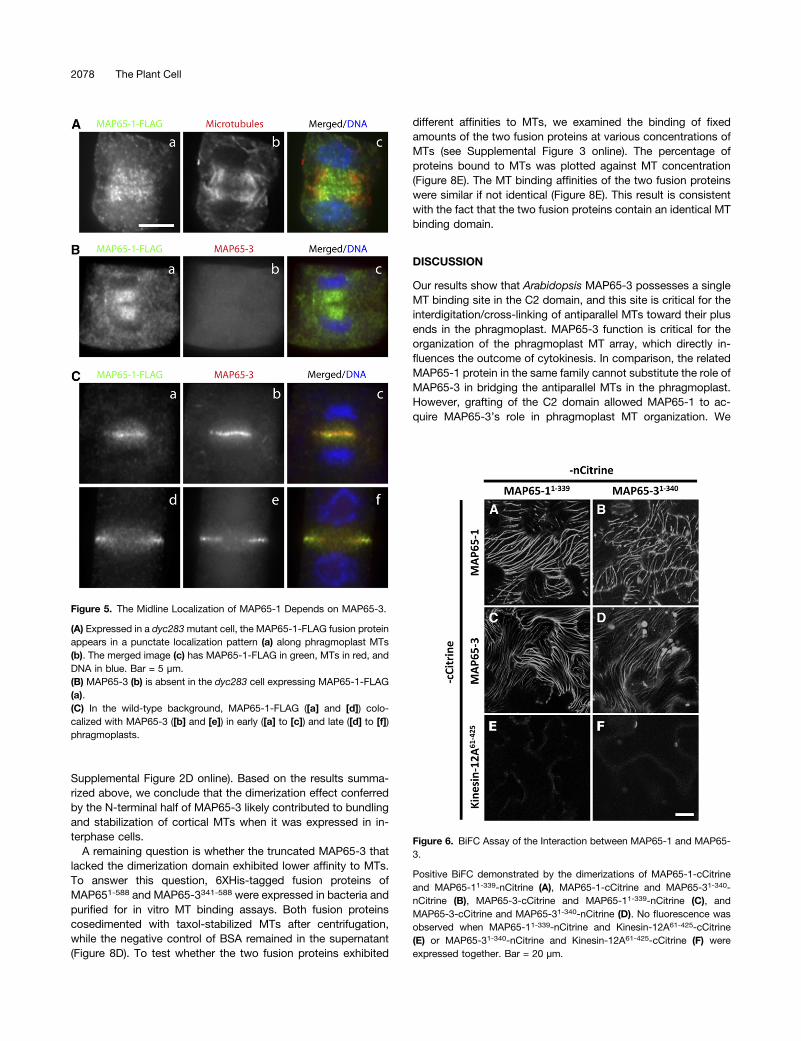
Because MAP65-1SWAP but not MAP65-1 was able to suppressthe dyc283 mutation, we wondered whether MAP65-1SWAP hada different localization pattern than MAP65-1. Indeed, theMAP65-1SWAP-FLAG fusion protein conspicuously appeared atthe midzone of the early phragmoplast (Figure 7A, panels a to c).At later stages of cytokinesis, the protein continued to decoratethe areas that bridged MT bundles from the opposite sides ofthe phragmoplast (arrowheads, Figure 7A, panels d to i). Thisresult suggested that the fusion protein most likely bundled anti-parallel MTs in the phragmoplast midzone and suppressed thedefects in cytokinesis caused by the dyc283 mutation. Becausethese cells lacked MAP65-3, no obvious signals were detectedby the anti-MAP65-3 immunofluorescence (Figure 7B, panelsa to c). These results suggested that, in the absence of MAP65-3, the addition of the MAP65-3 C2 domain was sufficient toredirect MAP65-1 to engage antiparallel MTs in the phragmo-plast midzone.
A MT Binding Domain Resides in the C2 Domainof MAP65-3
To further understand how MAP65-3 interacts with MTs, GFP fu-sions of MAP65-3 and its derivatives (Figure 8A) were expressed intobacco leaf cells under the control of a 35S promoter. The ec-topically expressed MAP65-3-GFP fusion appeared in linear thickbundles, resembling MT bundles (Figure 8B, panel a). Similar re-sults were obtained when the truncated MAP65-31-588-GFP wasexpressed (Figure 8B, panel b). MAP651-495-GFP, however, ex-hibited a diffuse localization pattern in the cytoplasm (Figure 8B,panel c). Because a published study showed that the MT bindingsite in MAP65 family proteins is located at the C-terminal half of
the protein (Smertenko et al., 2004), we first tested the entireC-terminal half consisting of both C1 and C2 domains. The re-sulting MAP65-3C-GFP fusion decorated a filamentous meshworkat the cell cortex (Figure 8B, panel d). Such a distribution patternwas also detected by the MAP65-3C2-GFP containing the C2domain alone, but not MAP65-3C1-GFP with the C1 domain(Figure 8B, panels e and f). This result further suggested that theMT binding site resides in the C2 domain. Because of the dif-ference in suppression of the dyc283 mutation by MAP65-31-588
and MAP65-31-495, we asked whether the region of amino acids496 to 588 in MAP65-3 constituted a MT-interacting site. Usingthis region, the MAP65-3C2N-GFP fusion also decorated a MT-like meshwork at the cell cortex (Figure 8B, panel g). These re-sults support the notion that the fusion proteins can directly bindto cortical MTs. The peptides of MAP65-3C1, MAP65-3C2, andMAP65-3C2N have calculated isoelectric points of 5.15, 10.24,and 11.38, respectively. This result indicates that the interactionbetween MAP65-3C2N and MTs is likely charge dependent andthe acidic nature of the MAP65-3C1 domain prevents it fromdecorating MTs. However, such a mechanism would not causeMAP65-3 to selectively bundle antiparallel MTs only near theirplus ends in the phragmoplast midzone.To test whether the filamentous networks decorated by the
MAP65-3-GFP fusions truly represented cortical MTs, they werecoexpressed with the MT binding domain derived from CKL6(for casein kinase 1-like 6) in an mCherry fusion (Ben-Nissanet al., 2008). The results indicate that all filamentous networkshighlighted by the GFP fusions colocalize perfectly with mCherry-marked cortical MTs (see Supplemental Figure 1 online). In cellsshowing diffuse signals of GFP alone, MAP65-31-495-GFP, andMAP65-3C1-GFP, cortical MTs were still detected by the CKL6-mCherry fusion (see Supplemental Figure 1 online).Published work shows that ectopic expression of MAP65-1 in
interphase cells increases the resistance of cortical MTs tochallenges of MT-depolymerizing agents (Van Damme et al.,2004b). We asked whether a similar effect was conferred byMAP65-3 by examining the turnover of the MAP65-3-GFP fusionproteins. Fluorescence recovery after photobleaching (FRAP)analysis was conducted on the GFP signals decorating corticalMTs. Both MAP65-3-GFP and MAP65-31-588-GFP had a stableappearance with low turnover rates of half-time values >90 s(Figures 8A and 8C). However, the complex network of finecortical MTs decorated by MAP65-3C, MAP65-3C2, or MAP65-3C2N all had greater turnover rates with half-time values <2 s(Figures 8A and 8C). To further test whether MTs became sta-bilized upon MAP65-3 expression, we challenged the tobaccoleaf cells with oryzalin for 30 min before they were observed byconfocal microscopy. Compared with cells expressing full-length MAP65-3 that showed bundled MTs decorated byMAP65-3-GFP (see Supplemental Figure 2A online), cells ex-pressing MAP65-3C that lacked the N-terminal dimerizationdomain had cortical MTs organized into a fine and complexnetwork decorated by MAP65-3C-GFP (see SupplementalFigure 2B online). Upon oryzalin treatment, cortical MTs re-mained highly bundled in cells expressing MAP65-3-GFP (seeSupplemental Figure 2C online). However, the fine and densecortical MT network was replaced with scarce and wavy fila-ments in oryzalin-treated cells expressing MAP65-3C-GFP (see
Figure 4. Dual Localization of MAP65-31-588-FLAG and Kinesin-12 in thePhragmoplast.
The merged images have MAP65-31-588 in green, Kinesin-12 in red, andDNA in blue. At both early ([A] to [C]) and late ([D] to [F]) stages ofcytokinesis, Kinesin-12 exclusively appears in the phragmoplast midline.By contrast, MAP65-31-588-FLAG has an extended localization pattern(arrows). Bar = 5 µm.
MAP65-3 Contains a Unique C Terminus 2077
Supplemental Figure 2D online). Based on the results summa-rized above, we conclude that the dimerization effect conferredby the N-terminal half of MAP65-3 likely contributed to bundlingand stabilization of cortical MTs when it was expressed in in-terphase cells.
A remaining question is whether the truncated MAP65-3 thatlacked the dimerization domain exhibited lower affinity to MTs.To answer this question, 6XHis-tagged fusion proteins ofMAP651-588 and MAP65-3341-588 were expressed in bacteria andpurified for in vitro MT binding assays. Both fusion proteinscosedimented with taxol-stabilized MTs after centrifugation,while the negative control of BSA remained in the supernatant(Figure 8D). To test whether the two fusion proteins exhibited
different affinities to MTs, we examined the binding of fixedamounts of the two fusion proteins at various concentrations ofMTs (see Supplemental Figure 3 online). The percentage ofproteins bound to MTs was plotted against MT concentration(Figure 8E). The MT binding affinities of the two fusion proteinswere similar if not identical (Figure 8E). This result is consistentwith the fact that the two fusion proteins contain an identical MTbinding domain.
DISCUSSION
Our results show that Arabidopsis MAP65-3 possesses a singleMT binding site in the C2 domain, and this site is critical for theinterdigitation/cross-linking of antiparallel MTs toward their plusends in the phragmoplast. MAP65-3 function is critical for theorganization of the phragmoplast MT array, which directly in-fluences the outcome of cytokinesis. In comparison, the relatedMAP65-1 protein in the same family cannot substitute the role ofMAP65-3 in bridging the antiparallel MTs in the phragmoplast.However, grafting of the C2 domain allowed MAP65-1 to ac-quire MAP65-3’s role in phragmoplast MT organization. We
Figure 5. The Midline Localization of MAP65-1 Depends on MAP65-3.
(A) Expressed in a dyc283mutant cell, the MAP65-1-FLAG fusion proteinappears in a punctate localization pattern (a) along phragmoplast MTs(b). The merged image (c) has MAP65-1-FLAG in green, MTs in red, andDNA in blue. Bar = 5 µm.(B) MAP65-3 (b) is absent in the dyc283 cell expressing MAP65-1-FLAG(a).(C) In the wild-type background, MAP65-1-FLAG ([a] and [d]) colo-calized with MAP65-3 ([b] and [e]) in early ([a] to [c]) and late ([d] to [f])phragmoplasts.
Figure 6. BiFC Assay of the Interaction between MAP65-1 and MAP65-3.
Positive BiFC demonstrated by the dimerizations of MAP65-1-cCitrineand MAP65-11-339-nCitrine (A), MAP65-1-cCitrine and MAP65-31-340-nCitrine (B), MAP65-3-cCitrine and MAP65-11-339-nCitrine (C), andMAP65-3-cCitrine and MAP65-31-340-nCitrine (D). No fluorescence wasobserved when MAP65-11-339-nCitrine and Kinesin-12A61-425-cCitrine(E) or MAP65-31-340-nCitrine and Kinesin-12A61-425-cCitrine (F) wereexpressed together. Bar = 20 µm.
2078 The Plant Cell
conclude that the C2 domain specifies the function of MAP65-3in maintaining the integrity of the phragmoplast MT array duringcytokinesis.
The C2 Domain Distinguishes MAP65-3 from Other MAP65Family Proteins
The MAP65/Ase1/PRC1 family of proteins act in dimers tocross-link and bundle MTs (Walczak and Shaw, 2010). Theypossess a dimerization domain toward the N terminus and MTbinding domain(s) toward the C terminus (Smertenko et al.,2004). Both monocot and dicot plants produce a large numberof proteins in this family (Smertenko et al., 2008; Guo et al.,
2009). These MAP65 isoforms can be nicely aligned accordingto their sequence homology except for the very C-terminal re-gions (Smertenko et al., 2008). The divergence of their C terminimay be responsible for the diversity of their intracellular locali-zation patterns (Van Damme et al., 2004b; Mao et al., 2005b)and consequently for their varied functions. A recent structuralanalysis presented a model showing that the human MAP65homolog PRC1 uses its spectrin-featured domain to bind to MTsand decode their polarity so that it can specifically mark theoverlapping region of the anaphase spindle (Subramanian et al.,2010). Earlier studies delineated two MT binding sites, namely,MTB1 and MTB2, in MAP65-1 (Smertenko et al., 2004). How-ever, our results showed that only the MTB2/C2 region ofMAP65-3 and not the MTB1 region binds directly to MTs. Thus,MAP65-3 may have adopted a MT binding mechanism differentfrom that of MAP65-1 or other isoforms.We demonstrated that the presence of the C2 domain was
most critical for the interaction of MAP65-3 with antiparallel MTsin the phragmoplast. Compared with MAP65-3, MAP65-1 doesnot contain a similar homologous region, but mixed results havebeen published on MAP65-1 localization in the phragmoplast.Antibodies raised against a MAP65-1 region decorate thephragmoplast midline in dividing suspension cultured cells(Smertenko et al., 2008). When expressed in GFP fusions underthe control of the 35S promoter in tobacco cells, MAP65-1 doesnot appear in the midline of mature phragmoplasts, except for inthe spindle midzone at anaphase (Van Damme et al., 2004a;Mao et al., 2005a; Gaillard et al., 2008). This discrepancy couldhave arisen from the differences in cell types and/or expressionlevels of the protein under different conditions. A recent studyhad MAP65-1-GFP expressed under its native promoter (Lucaset al., 2011). In root meristematic cells, this fusion protein uni-formly decorates the entire phragmoplast MT array and does notappear predominantly in the phragmoplast midzone (Lucas andShaw, 2012). In this study, we showed that MAP65-1 alone wasinsufficient to cross-bridge antiparallel MTs in the phragmoplast.It would appear at the phragmoplast midline when expressedunder the control of the MAP65-3 promoter and in the presenceof MAP65-3. This result may arise from two scenarios. First, theexpressed MAP65-1 fusion protein might have interacted withthe endogenous MAP65-3 protein to be localized at the midline.This notion is supported by our BiFC data that showed that thetwo proteins interact directly when their expression is temporallycoordinated. The other possibility is that the elevated expressionlevel of MAP65-1 at the mitotic stages would allow it to appearin the phragmoplast midzone in the presence of interdigitatedantiparallel MTs cross-linked by MAP65-3.Compared with MAP65-1 and MAP65-3, other MAP65 iso-
forms also exhibit distinct localization patterns. For example,MAP65-6 decorates mitochondria (Mao et al., 2005b). At pro-phase, MAP65-5 decorates both the prophase spindle and thepreprophase band, while MAP65-1 only appears at the pre-prophase band (Gaillard et al., 2008). Unlike MAP65-1, whichappears at the spindle midzone, MAP65-4 is specifically asso-ciated with the kinetochore fibers but not the preprophase band(Fache et al., 2010). It is intriguing how these related proteinsachieve such dynamic localization patterns at particular stagesof cell division. Detailed structural and functional dissections are
Figure 7. The MAP65-3 C2 Domain Redirects MAP65-1 to the Phrag-moplast Midline.
(A) The localization of MAP65-1SWAP-FLAG (green) at different stages ofcytokinesis. The merged image has MAP65-1SWAP-FLAG in green, MTsin red, and DNA in blue. The MAP65-1SWAP-FLAG signal is concentratedbetween two sets of antiparallel MTs during late anaphase/telophase ([a]to [c]). The signal is pronounced in the midline (arrowheads in [a], [d],and [g]) of cells bearing mature phragmoplasts. Bar = 5 µm.(B) In the dyc283 cell expressing MAP65-1SWAP-FLAG, which was de-tected at the phragmoplast midline (arrowhead in [a]), MAP65-3 is ab-sent (b). The merged image has MAP65-1SWAP-FLAG in green, MAP65-3in red, and DNA in blue (c).
MAP65-3 Contains a Unique C Terminus 2079
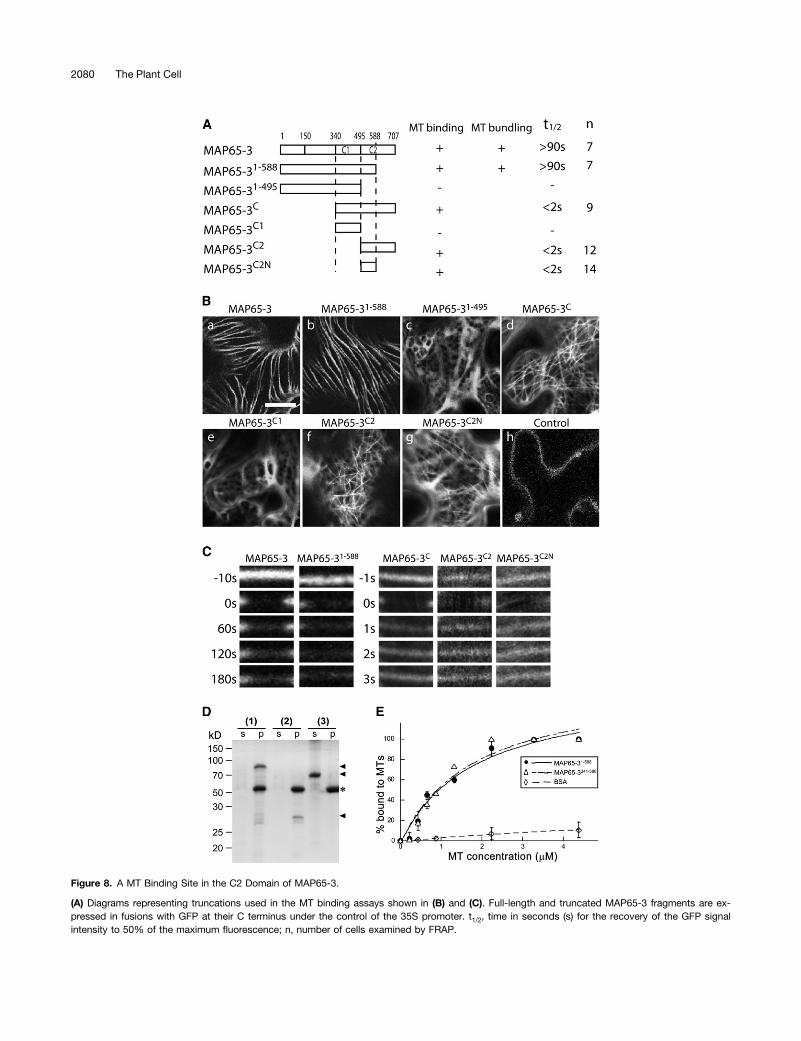
Figure 8. A MT Binding Site in the C2 Domain of MAP65-3.
(A) Diagrams representing truncations used in the MT binding assays shown in (B) and (C). Full-length and truncated MAP65-3 fragments are ex-pressed in fusions with GFP at their C terminus under the control of the 35S promoter. t1/2, time in seconds (s) for the recovery of the GFP signalintensity to 50% of the maximum fluorescence; n, number of cells examined by FRAP.
2080 The Plant Cell

required to further characterize their activities in rapidly growingplant cells.
Functional Redundancy among the MAP65 Isoforms
The sequence similarities among the MAP65 family proteinswould imply functional redundancies. Indeed, a recent workshowed that MAP65-1, MAP65-2, and MAP65-3 were sub-strates of an important MAP kinase in cytokinesis (Sasabe et al.,2011). It has been shown that mutations in genes encoding ei-ther MAP65-1 or MAP65-2 would enhance the phenotype byadding a mutation in the MAP65-3-encoding gene in Arabidopsis(Sasabe et al., 2011). Our results support such a genetic in-teraction. The expression of MAP65-1 significantly suppressed thecytokinetic defects and root growth problems caused by thedyc283 mutation. This suppression phenomenon may be beyondthe localization in the phragmoplast midline because the ectopi-cally expressed MAP65-1 did not appear at the site normally oc-cupied by MAP65-3. It implied that MAP65-3 might play a role inaspects other than cross-linking antiparallel MTs, and such a rolecould be shared with MAP65-1. If this were true, the suppressionwould be brought about by the presence of an increased amountof MAP65-1.
Due to the high sequence similarity between MAP65-3 andMAP65-4, one would expect that they may share similar func-tions. However, the ectopically expressed MAP65-4 in tobaccoBright Yellow-2 cells that contain a putative ortholog(s) of MAP65-3does not appear in the phragmoplast (Fache et al., 2010). In otherwords, it might not be able to interact with MAP65-3. Some in-trinsic features embedded in the sequence differences betweenMAP65-3 and MAP65-4 must demark the drastically differentlocalization patterns of these two highly similar proteins.
A Generalized Role in MT Stabilization by MAP65Family Proteins
Published data show that non-MAP65-3 isoforms of MAP65stabilize cortical MTs when they are ectopically expressed (VanDamme et al., 2004b; Li et al., 2009). Here, we showed thatectopically expressed MAP65-3 cause hyperbundling of corticalMTs with low turnover rates. The result suggests that the un-derlying cortical MTs became stabilized after being cross-linkedtogether into bundles. There is no evidence showing that theendogenous MAP65-3 protein interacts with cortical MTs. Thismay be a result of its cell cycle–dependent expression pattern.
The ectopic expression of all tested MAP65 family proteins ininterphase cells all leads to hyperbundling of cortical MTs thatare aligned largely in parallel to each other. Such an effect wasnot observed when MAP65-1- and MAP65-2-GFP fusions wereexpressed under the control of their endogenous promoter(Lucas et al., 2011). In vitro biochemical studies showed that twoMAP65 isoforms and the human PRC1 bind to individual MTs inmonomeric forms followed by a zippering effect to allow anti-parallel MTs to be bundled together (Gaillard et al., 2008; Lloyd,2011). Taking the FRAP data into account, we believe that thestabilization of cortical MTs was likely brought about by thebundling effect of overexpressed MAP65 in interphase cells. Inthe phragmoplast, ectopically overexpressed MAP65-1 deco-rates MTs flanking the midline (Gaillard et al., 2008). In a halvedphragmoplast, MTs are arranged in a parallel fashion (Euteneuerand McIntosh, 1980). Such a pattern is essential for unidirec-tional transport of Golgi-derived vesicles toward MT plus endsto form the cell plate. If MAP65-1 only bundled antiparallel MTs,the phragmoplast of cells overproducing MAP65-1 would haveantiparallel MTs present in both phragmoplast halves. Therefore,it would be interesting to examine MT polarity in the phragmo-plasts of cells containing high concentrations of MAP65-1.Unlike MAP65-1, ectopically overexpressed MAP65-3 only
appears in the phragmoplast midline (Van Damme et al., 2004a).However, it decorated cortical MTs evenly and without anypreference in interphase cells in our study. MAP65-3, whenexpressed under the control of its native promoter, transientlyinteracted with cortical MTs when cells were ready to enter theM phase (Caillaud et al., 2008). Collectively, these results sug-gest that the interaction of MAP65-3 with phragmoplast MTs isdifferent from that with cortical MTs in interphase cells. Its ac-tivity in bundling antiparallel MTs near their plus ends could becell cycle dependent. Such an activity could be activated upona posttranslational modification of the protein.
Regulation of MAP65 Functions in the Phragmoplast
A number of protein kinases appear in the phragmoplast and playcritical roles in regulating the progression of cytokinesis (Oh et al.,2005; Sasabe and Machida, 2006; Krupnova et al., 2009). Amongthem, the TWO-IN-ONE kinase and kinases of the NACK-PQRpathway specifically appear at the phragmoplast midline (Nishihamaet al., 2002; Kosetsu et al., 2010). Currently, the substrate of TWO-IN-ONE during cytokinesis is unknown. However, the MAP kinaseof the NACK-PQR pathway NRK1 phosphorylates MAP65-1 and
Figure 8. (continued).
(B) A MT binding site resides in the region of amino acids 496 to 588 of MAP65-3. MAP65-3- and MAP65-31-588-GFP fusions decorate thick and parallelMT bundles. GFP fusions of MAP65-3C, MAP65-3C2, and MAP65-3C2N highlight a fine filamentous network of cortical MTs. However, the GFP fusionsof MAP65-31-495 and MAP65-3C1 give diffuse signals in the cytoplasm. Cells expressing GFP alone are used as the control. Bar = 20 µm.(C) The N-terminal region of MAP65-3 contributes to the stable association with bundled cortical MTs as revealed by the FRAP experiment. GFP fusionsof MAP65-3 and MAP65-31-588 have half-life times >90 s. In comparison, MAP65-3C, MAP65-3C2, and MAP-3C2N GFP fusions have half-life times of <2 s.(D) MT cosedimentation of MAP65-3 derivatives. After incubation of MAP65-31-588 (1), MAP65-3341-588 (2), and BSA (3) with taxol-stabilized MTs,supernatant (s) and pellet (p) fractions were analyzed by SDS-PAGE. Positions of MAP65-31-588, MAP65-3341-588, and BSA are indicated by arrowheadsand that of tubulins by an asterisk on the right. Molecular mass markers in kilodaltons are shown on the left.(E) Equilibrium binding assays of MAP65-31-588 (circles, solid line) and MAP65-3341-588 (triangles, broken line) with MTs. BSA (diamonds, dashed line)was used as a negative control. Error bars represent SD of three replicates.
MAP65-3 Contains a Unique C Terminus 2081

negatively regulates its MT-bundling activity in the phragmoplast intobacco cells (Sasabe et al., 2006; Komis et al., 2011). Recently, itwas found that in Arabidopsis not only MAP65-1 but also MAP65-2and MAP65-3 are substrates of the MAP kinase MPK4 (Kosetsuet al., 2010; Sasabe et al., 2011). Redundant roles were also sug-gested for these three MAP65 isoforms. Thus, the MT-stabilizingfunction of these MAPs has to be downregulated upon the com-pletion of cell plate assembly so that phragmoplast MTs would beready to undergo depolymerization. It was shown that Ser/Thr-to-Alamutations in the NRK1 phosphorylation sites led to the formation ofmultinucleate cells caused by failures of cytokinesis (Sasabe et al.,2006).
Conversely, in human cells, the MAP65-3 counterpart, PRC1,was found to be phosphorylated at prophase to metaphase anddephosphorylated at anaphase (Zhu et al., 2006). The dephos-phorylation event is critical for PRC1 to be localized to the spindlemidzone and to oligomerize there. Thus, different mechanismsmay regulate MAP65/PRC1 function in different organisms.
The Requirement of MAP65-3 for Other Proteins to Interactwith MT Plus Ends in the Phragmoplast
It is intriguing that the MT motors Kinesin-12 and PAKRP2require MAP65-3 to interact with MT plus ends in the phrag-moplast (Ho et al., 2011a). A similar function can be found for itshuman counterpart, PRC1, for the localization of the kinesinfamily protein KIF14 in the spindle midzone (Gruneberg et al.,2006). In both fission yeast and animal cells, Ase1p and PRC1interact with a number of kinesin motors (Kurasawa et al., 2004;Zhu and Jiang, 2005; Gruneberg et al., 2006). In human cells, thekinesin Kif4 plays a role in translocating PRC1 to the spindlemidzone (Zhu and Jiang, 2005). In our experiment, the loss oftwo Kinesin-12 motors that led to phragmoplast MT disorgani-zation and cytokinesis failures abolished the midline formationand MAP65-3 distribution along the phragmoplast MTs, sug-gesting that MAP65-3 and Kinesin-12 are interdependent.
The localized activity of MAP65-3 in the phragmoplast mayalso have a restricted MT bundling effect in the phragmoplastproper. During cytokinesis, newly polymerized MTs would bebrought together by the fortified phragmoplast MT array. In theabsence of MAP65-3, however, MTs nucleated elsewhere, suchas on the envelope of the reforming daughter nuclei, would re-main discrete. This is probably why MTs polymerized in placesother than the phragmoplast proper in cells lacking a functionalMAP65-3.
METHODS
Plant Materials, Growth Conditions, and Transformation
The Arabidopsis thaliana materials used in this study include the controlWassilewskija plants and the dyc283 mutant (Caillaud et al., 2008). Plantgrowth conditions and Agrobacterium tumefaciens–mediated trans-formation were performed as described previously (Kong et al., 2010; Hoet al., 2011a).
For observation of various GFP fusions, Agrobacterium with the p19and testing plasmids (see below) were grown overnight in lysogeny brothmedium at 30°C. The culture was harvested by centrifugation and re-suspended in the infiltration medium (10 mMMgCl2, 10 mMMES, pH 5.6,and 200 µM acetosyringone) to a final OD600 of 0.6 to ;1.0. The
agrobacterial suspension was incubated at 22°C for at least 3 h and thenused for filtration of Nicotiana benthamiana leaves according to a pub-lished protocol (Popescu et al., 2007).
Construction of Expression Vectors
A previously constructed plasmid containing full-length MAP65-3 (i.e.,ProMAP65-3:MAP65-3-FLAG) (Ho et al., 2011a) was used as template forthe following constructs. To make the ProMAP65-3:MAP65-31-588-FLAGand ProMAP65-3:MAP65-31-495-FLAG constructs, promoter and codingsequences were amplified by PCR using the forward primer AtMAP65 1(59-CACCACACTCTTCCCTACACAAAACCGC-39) and the reverse pri-mers AtMAP65 588r (59-GGGGGAAAACGGTTTGCGAACTAATG-39), andAtMAP65 1 and AtMAP65 495r (59-TTGTTCTGCTATGAGCTGTCCTT-GAAGC-39). To make the ProMAP65-3:MAP65-1 and ProMAP65-3:MAP65-1SWAP constructs, the MAP65-3 promoter was first amplifiedby PCR using the AtMAP65 1 and AtMAP65 22 (59-TTCGAAATGCT-TAAGCCTGTAACAGGG-39) primers. The full-length MAP65-1 cDNAsequence was amplified by PCR from the cDNA clone M78N02STM usingthe forward primer AtMAP-1a (59-CCCTGTTACAGGCTTAAGCATTTCGA-AATGGCAGTTACAGATACTGAAAGTCCTC-39) and the reverse primerAtMAP-1b (59-TGGTGAAGCTGGAACTTGATGATCTGC-39). The MAP65-3promoter and MAP65-1 fragments were fused together by PCR using theprimers AtMAP651 andAtMAP-1b. Tomake theProMAP65-3:MAP65-1SWAP
construct, two fragments in addition to the MAP65-3 promoter wereamplified individually. The MAP65-1 truncated fragment was amplifiedusing the primers AtMAP-1a and AtMAP-1c (59-GGGTTTGGACCCGTA-AAGTGCCTCTTGCTCTACGTGTGGCTGTTCTTGAAC-39). The fragmentencoding the MAP65-3 C2 domain was amplified with the primers At-MAP65 21 (59-GAGGCACTTTACGGGTCCAAACCC-39) and AtMAP65 8(59-AACCAAACGACATTCAGACTGTAGCATG-39). The entire fused Pro-MAP65-3:MAP65-1SWAP fragment was amplified by an additional PCRreaction using the primers AtMAP65 1 and AtMAP65 8 and the threefragments as templates. The aforementioned PCR products were clonedinto the Gateway pENTR/D-TOPO vector (Invitrogen), followed by LRrecombination reactions with the destination vector pGWB10 (Nakagawaet al., 2007) to render the final vectors that would allow the expression ofC-terminal FLAG fusion proteins. These plasmids were transformed intothe dyc283 mutant.
Constructs for tobacco (Nicotiana tabacum) leaf filtration experimentsweremade as following. The cDNA fragments encoding full-length MAP65-3 andtruncated MAP65-31-588 and MAP65-31-495 proteins were amplified by theforward primer AtMAP65-3 33 (59-CACCATGGCAAGTGTTCAAAAA-GATCCGATTCTTC-39) and reverse primers AtMAP 8, AtMAP 588r, andAtMAP 495r, respectively. The fragments encoding the MAP65-3C andMAP65-3C2 peptides were amplified by PCR using the primers MAP65MTB5KpnI (59-CACCGGTACCATGAGTGCTATAGATCAGACTATCGTGGCG-39)and AtMAP 8, and MTB2 5KpnI (59-CACCGGTACCATGGAGGCACTT-TACGGGTCCAAACCC-39) and AtMAP 8, respectively. The fragment en-coding MAP65-3C1 was amplified using the primers MTB65MTB 5KpnIand AtMAP 495r and that encoding MAP65-3C2N using the primers MTB25KpnI and AtMAP 588r. These PCR products were cloned into pENTR/TOPObefore being delivered into the destination vector pGWB5 (Nakagawa et al.,2007). The resulting constructs would allow the expression of these proteins/peptides in fusions with GFP at their C termini under the control of the 35Spromoter. For the MT colocalization experiment, an mCherry fusion proteinwith the C-terminal MT binding site of CKL6 (Ben-Nissan et al., 2008) wascoexpressed with the aforementioned fusion proteins in leaf cells of N.benthamiana.
Fluorescence and Confocal Microscopy
Root tip cells were prepared for indirect immunofluorescence staining asdescribed previously (Lee and Liu, 2000). Primary antibodies used in this
2082 The Plant Cell

study included anti-FLAG C2 (Shanghai Genomics), polyclonal antitubulin(Cytoskeleton), anti-MAP65-3 (Ho et al., 2011a), and anti-PAKRP1-Cfor Kinesin-12 (Lee and Liu, 2000). Secondary antibodies were fluo-rescein isothiocyanate–conjugated donkey anti-mouse IgG, fluoresceinisothiocyanate–conjugated donkey anti-rabbit IgG, and Texas Red–conjugated donkey anti-goat IgG (Rockland Immunochemicals).
Slides were observed under an Eclipse E600 microscope equippedwith epifluorescence optics (Nikon) and ET filter sets (Chroma Technol-ogy). Images were captured by an Orca charge-coupled device camera(Hamamatsu Photonics) driven by the MetaMorph software package(Molecular Devices) before being assembled by Photoshop software(Adobe).
The signals of GFP fusions with the full-length MAP65-3 protein andvarious truncations were visualized in N. benthamiana leaves 3 to 4 d afteragrobacterial filtration using an FV1000 laser scanning confocal micro-scope with a PLAPON 360 oil objective lens (Olympus USA). The GFPsignal was collected at 500 to 550 nm. The FRAP experiments wereperformed using the built-in function in FV1000. The GFP and mCherrysignals were excited by the 488- and 543-nm lasers and collected at the500- to 550-nm and 550- to 600-nm ranges, respectively. Acquiredimaged were processed by MetaMorph and Photoshop (Adobe).
The cell profiles in the root tip regions were observed according to thefluorescent signal of propidium iodide infiltrated into the cell wall. Briefly,5-d-old seedlings were incubated in 20 mg/mL propidium iodide solutionand observed under a TCS SP2 (Leica) confocal microscope with an HCXPL APO 363 water or an HC PL APO 320 objective lens. Two cell files inregions of 80 to 240 mm from the root tip were used to calculate thedegree of cytokinetic defects.
BiFC Experiments
The BiFC experiments were performed using the yellow fluorescent proteinvariant Citrine as reported (Caplan et al., 2008). Instead of the cauliflowermosaic virus 35S promoter, the much weaker NOS promoter was used forall constructs used in BiFC. Briefly, the full-length MAP65-1–coding se-quence was cloned into SPDK919 (pNOS-MCS-cCitrine) at the MluI andXbaI sites after being amplified by PCR using the primers of AtMAP65-1MluI5 (59-CACCACGCGTATGGCAGTTACAGATACTGAAAGTCCTC-39)and AtMAP65-1XbaI3 (59-CATTCTAGACGTGGTGAAGCTGGAACTTGATG-ATCTGC-39). Similarly, the construct for MAP65-3 was made using theprimers MAP65-3KM5 (59-CACCATGGCAAGTGTTCAAAAAGATC-39)and MAP65-3KM3 (59-ACTAGTACCAAACGACATTCAGACTG-39). Cod-ing sequences of the truncated MAP65-11-339 and MAP65-31-340 fusionswere amplified using primer pairs MAP65-1KM5 (59-CACCATGGCAG-TTACAGATACTGAAAG-39) and MAP65-1KM3 (59-ACTAGTTCCG-GGTTAACTTCTACATG-39), and MAP65-3KM5 and MAP65-3KM3b(59-ACTAGTCTGTCTGACACTGGCAACAAG-39). They were then clonedinto SPDK920 (pNOS-MCS-nCitrine) at the NcoI and SpeI sites. Asa negative control, the coding sequence of the motor domain of Kinesin-12A (Lee and Liu, 2000; Pan et al., 2004) was amplified using the primersof PAKRP1KM5b (59-CACCATGGAGAATCCGTTGCCTCCG-39) andPAKRP1KM3 (59-ACTAGTGCACGCTGAGCAAATCTCAG-39) and clonedinto SPDK919 between the MluI and XbaI sites. Agrobacterial infiltrationand confocal microscopy examination experiments were as describedabove.
MT Cosedimentation Assay
To express 6XHis-taggedMAP651-588 andMAP65-3341-588, the correspondingcoding sequences were amplified by PCR using the following primer pairs:AtMAP65_17 (59-CACCGGATCCGCAAGTGTTCAAAAAGATCCGATTCTTC-39) and AtMAP65_22 (59-CATGAATTCTCAGGGGGAAAACGGTTTGCGAAC-TAATG-39), and AtMAP65_19 (59-CACCGGATCCAGTGCTATAGATCA-GACTATCGTGGCG-39) and AtMAP65_22, respectively. The PCR products
were digested with BamHI and EcoRI and cloned into pET30a (EMD Millipore)between these sites. The fusion proteins were expressed and purified asdescribed previously (Ho et al., 2011a). Purified proteins at the concentration of2 µM were subjected to a MT cosedimentation assay as described previously(Lee et al., 2001). Proteins were analyzed by 12.5% SDS-PAGE and stainedwith Coomassie Brilliant Blue G 250 prior to being imaged for density analysis.
Accession Numbers
The Arabidopsis Information Resource locus identifier for the genesmentioned in this article are At5g51600 for MAP65-3 and At5g55230 forMAP65-1. The cDNA clone M78N02STM (GenBank accession numberBE530728) contains the full-length coding sequence of MAP65-1.
Supplemental Data
The following materials are available in the online version of this article.
Supplemental Figure 1. Colocalization of MAP65-3 and Its Deriva-tives with Cortical MTs.
Supplemental Figure 2. Cortical MTs upon Expression of MAP65-3 orMAP65-3341-707 with and without Oryzalin Treatment.
Supplemental Figure 3. MT Binding Assays of MAP65-31-588 andMAP65-3341-588.
ACKNOWLEDGMENTS
We thank Jung-Youn Lee at the University of Delaware for generouslyproviding the CKL6-mCherry construct, Tsuyoshi Nakagawa at ShimaneUniversity in Japan for the pGWB vectors, and Bruno Favery at InstitutNational de la Recherche Agronomique in France for the dyc283 mutant.We also thank William Lucas for sharing the TCS SP2 confocalmicroscope acquired through National Science Foundation (NSF) GrantDBI-0354790. We are especially grateful to the members of our groupand Hsou-min Li for their critical input. This report is based on worksupported by the NSF under Grant MCB-0920454 (to B.L.). BiFCconstructs used here were developed under NSF Grant DBI-1042344(to S.P.D.-K.). Any opinions, findings, conclusions, or recommendationsexpressed in this article are those of the authors and do not necessarilyreflect the views of the NSF.
AUTHOR CONTRIBUTIONS
C.-M.K.H. and B.L. designed the project. C.-M.K.H., Y.-R.J.L., L.D.K.,and S.P.D.-K. performed specific experiments and analyzed data.C.-M.K.H. wrote the article. B.L. revised and edited the article.
Received October 10, 2011; revised April 11, 2012; accepted April 23,2012; published May 8, 2012.
REFERENCES
Austin, J.R., IISeguí-Simarro, J.M., and Staehelin, L.A. (2005).Quantitative analysis of changes in spatial distribution and plus-endgeometry of microtubules involved in plant-cell cytokinesis. J. CellSci. 118: 3895–3903.
Ben-Nissan, G., Cui, W., Kim, D.J., Yang, Y., Yoo, B.C., and Lee, J.Y. (2008). Arabidopsis casein kinase 1-like 6 contains a microtubule-binding domain and affects the organization of cortical microtubules.Plant Physiol. 148: 1897–1907.
Caillaud, M.-C., Lecomte, P., Jammes, F., Quentin, M., Pagnotta,S., Andrio, E., de Almeida Engler, J., Marfaing, N., Gounon, P.,Abad, P., and Favery, B. (2008). MAP65-3 microtubule-associated
MAP65-3 Contains a Unique C Terminus 2083

protein is essential for nematode-induced giant cell ontogenesis inArabidopsis. Plant Cell 19: 423–437.
Caplan, J.L., Mamillapalli, P., Burch-Smith, T.M., Czymmek, K.,and Dinesh-Kumar, S.P. (2008). Chloroplastic protein NRIP1 me-diates innate immune receptor recognition of a viral effector. Cell132: 449–462.
Chang-Jie, J., and Sonobe, S. (1993). Identification and preliminarycharacterization of a 65 kDa higher-plant microtubule-associatedprotein. J. Cell Sci. 105: 891–901.
Eleftheriou, E.P., Baskin, T.I., and Hepler, P.K. (2005). Aberrant cellplate formation in the Arabidopsis thaliana microtubule organization1 mutant. Plant Cell Physiol. 46: 671–675.
Euteneuer, U., and McIntosh, J.R. (1980). Polarity of midbody andphragmoplast microtubules. J. Cell Biol. 87: 509–515.
Fache, V., Gaillard, J., Van Damme, D., Geelen, D., Neumann, E.,Stoppin-Mellet, V., and Vantard, M. (2010). Arabidopsis kineto-chore fiber-associated MAP65-4 cross-links microtubules andpromotes microtubule bundle elongation. Plant Cell 22: 3804–3815.
Gaillard, J., Neumann, E., Van Damme, D., Stoppin-Mellet, V., Ebel,C., Barbier, E., Geelen, D., and Vantard, M. (2008). Two micro-tubule-associated proteins of Arabidopsis MAP65s promote anti-parallel microtubule bundling. Mol. Biol. Cell 19: 4534–4544.
Gruneberg, U., Neef, R., Li, X.L., Chan, E.H.Y., Chalamalasetty,R.B., Nigg, E.A., and Barr, F.A. (2006). KIF14 and citron kinase acttogether to promote efficient cytokinesis. J. Cell Biol. 172: 363–372.
Guo, L., Ho, C.-M.K., Kong, Z., Lee, Y.-R.J., Qian, Q., and Liu, B.(2009). Evaluating the microtubule cytoskeleton and its interactingproteins in monocots by mining the rice genome. Ann. Bot. (Lond.)103: 387–402.
Ho, C.M., Hotta, T., Guo, F., Roberson, R.W., Lee, Y.R., and Liu, B.(2011a). Interaction of antiparallel microtubules in the phragmoplastis mediated by the microtubule-associated protein MAP65-3 inArabidopsis. Plant Cell 23: 2909–2923.
Ho, C.M., Hotta, T., Kong, Z., Zeng, C.J., Sun, J., Lee, Y.R., and Liu,B. (2011b). Augmin plays a critical role in organizing the spindle andphragmoplast microtubule arrays in Arabidopsis. Plant Cell 23:2606–2618.
Hussey, P.J., Hawkins, T.J., Igarashi, H., Kaloriti, D., andSmertenko, A. (2002). The plant cytoskeleton: Recent advances inthe study of the plant microtubule-associated proteins MAP-65,MAP-190 and the Xenopus MAP215-like protein, MOR1. Plant Mol.Biol. 50: 915–924.
Jürgens, G. (2005). Cytokinesis in higher plants. Annu. Rev. PlantBiol. 56: 281–299.
Kapitein, L.C., Janson, M.E., van den Wildenberg, S.M.,Hoogenraad, C.C., Schmidt, C.F., and Peterman, E.J.G. (2008).Microtubule-driven multimerization recruits ase1p onto overlappingmicrotubules. Curr. Biol. 18: 1713–1717.
Kawamura, E., Himmelspach, R., Rashbrooke, M.C., Whittington,A.T., Gale, K.R., Collings, D.A., and Wasteneys, G.O. (2006). MI-CROTUBULE ORGANIZATION 1 regulates structure and function ofmicrotubule arrays during mitosis and cytokinesis in the Arabi-dopsis root. Plant Physiol. 140: 102–114.
Komis, G., Illés, P., Beck, M., and �Samaj, J. (2011). Microtubules andmitogen-activated protein kinase signalling. Curr. Opin. Plant Biol.14: 650–657.
Kong, Z., Hotta, T., Lee, Y.R., Horio, T., and Liu, B. (2010). The g-tubulincomplex protein GCP4 is required for organizing functional microtubulearrays in Arabidopsis thaliana. Plant Cell 22: 191–204.
Kosetsu, K., Matsunaga, S., Nakagami, H., Colcombet, J., Sasabe,M., Soyano, T., Takahashi, Y., Hirt, H., and Machida, Y. (2010).The MAP kinase MPK4 is required for cytokinesis in Arabidopsisthaliana. Plant Cell 22: 3778–3790.
Krupnova, T. et al. (2009). Microtubule-associated kinase-like proteinRUNKEL needed [corrected] for cell plate expansion in Arabidopsiscytokinesis. Curr. Biol. 19: 518–523. Erratum. Curr. Biol. 19: 536.
Kurasawa, Y., Earnshaw, W.C., Mochizuki, Y., Dohmae, N., andTodokoro, K. (2004). Essential roles of KIF4 and its binding partnerPRC1 in organized central spindle midzone formation. EMBO J. 23:3237–3248.
Lee, Y.-R.J., Giang, H.M., and Liu, B. (2001). A novel plant kinesin-related protein specifically associates with the phragmoplast organelles.Plant Cell 13: 2427–2439.
Lee, Y.R.J., Li, Y., and Liu, B. (2007). Two Arabidopsis phragmoplast-associated kinesins play a critical role in cytokinesis during malegametogenesis. Plant Cell 19: 2595–2605.
Lee, Y.R.J., and Liu, B. (2000). Identification of a phragmoplast-associ-ated kinesin-related protein in higher plants. Curr. Biol. 10: 797–800.
Li, H., Mao, T.L., Zhang, Z.D., and Yuan, M. (2007). The AtMAP65-1cross-bridge between microtubules is formed by one dimer. PlantCell Physiol. 48: 866–874.
Li, H., Zeng, X., Liu, Z.Q., Meng, Q.T., Yuan, M., and Mao, T.L.(2009). Arabidopsis microtubule-associated protein AtMAP65-2acts as a microtubule stabilizer. Plant Mol. Biol. 69: 313–324.
Liu, B., Hotta, T., Ho, C.-M.K., and Lee, Y.-R.J. (2011). Microtubuleorganization in the phragmoplast. In The Plant Cytoskeleton, B. Liu,ed (New York: Springer), pp. 207–225.
Lloyd, C. (2011). Plant cytokinesis: Circles within circles. Curr. Biol.21: R926–R927.
Lucas, J.R., Courtney, S., Hassfurder, M., Dhingra, S., Bryant, A.,and Shaw, S.L. (2011). Microtubule-associated proteins MAP65-1and MAP65-2 positively regulate axial cell growth in etiolated Arabi-dopsis hypocotyls. Plant Cell 23: 1889–1903.
Lucas, J.R., and Shaw, S.L. (March 24, 2012). MAP65-1 and MAP65-2 promote cell proliferation and axial growth in Arabidopsis roots.Plant J. http://dx.doi.org/10.1111/j.1365-313X.2012.05002.x.
Mao, G.J., Chan, J., Calder, G., Doonan, J.H., and Lloyd, C.W.(2005a). Modulated targeting of GFP-AtMAP65-1 to central spindlemicrotubules during division. Plant J. 43: 469–478.
Mao, T., Jin, L., Li, H., Liu, B., and Yuan, M. (2005b). Two microtu-bule-associated proteins of the Arabidopsis MAP65 family functiondifferently on microtubules. Plant Physiol. 138: 654–662.
Müller, S., Smertenko, A., Wagner, V., Heinrich, M., Hussey, P.J.,and Hauser, M.T. (2004). The plant microtubule-associated proteinAtMAP65-3/PLE is essential for cytokinetic phragmoplast function.Curr. Biol. 14: 412–417.
Nakagawa, T., Kurose, T., Hino, T., Tanaka, K., Kawamukai, M.,Niwa, Y., Toyooka, K., Matsuoka, K., Jinbo, T., and Kimura, T.(2007). Development of series of gateway binary vectors, pGWBs,for realizing efficient construction of fusion genes for plant trans-formation. J. Biosci. Bioeng. 104: 34–41.
Nishihama, R., Soyano, T., Ishikawa, M., Araki, S., Tanaka, H., Asada,T., Irie, K., Ito, M., Terada, M., Banno, H., Yamazaki, Y., andMachida, Y. (2002). Expansion of the cell plate in plant cytokinesisrequires a kinesin-like protein/MAPKKK complex. Cell 109: 87–99.
Oh, S.A., Johnson, A., Smertenko, A., Rahman, D., Park, S.K.,Hussey, P.J., and Twell, D. (2005). A divergent cellular role for theFUSED kinase family in the plant-specific cytokinetic phragmoplast.Curr. Biol. 15: 2107–2111.
Oh, S.A., Pal, M.D., Park, S.K., Johnson, J.A., and Twell, D. (2010).The tobacco MAP215/Dis1-family protein TMBP200 is required forthe functional organization of microtubule arrays during malegermline establishment. J. Exp. Bot. 61: 969–981.
Pan, R., Lee, Y.R.J., and Liu, B. (2004). Localization of two homol-ogous Arabidopsis kinesin-related proteins in the phragmoplast.Planta 220: 156–164.
2084 The Plant Cell

Pastuglia, M., Azimzadeh, J., Goussot, M., Camilleri, C., Belcram,K., Evrard, J.L., Schmit, A.C., Guerche, P., and Bouchez, D.(2006). g-Tubulin is essential for microtubule organization and de-velopment in Arabidopsis. Plant Cell 18: 1412–1425.
Popescu, S.C., Popescu, G.V., Bachan, S., Zhang, Z., Seay, M.,Gerstein, M., Snyder, M., and Dinesh-Kumar, S.P. (2007). Dif-ferential binding of calmodulin-related proteins to their targets re-vealed through high-density Arabidopsis protein microarrays. Proc.Natl. Acad. Sci. USA 104: 4730–4735.
Sasabe, M., and Machida, Y. (2006). MAP65: A bridge linking a MAPkinase to microtubule turnover. Curr. Opin. Plant Biol. 9: 563–570.
Sasabe, M., Kosetsu, K., Hidaka, M., Murase, A., andMachida, Y. (2011).Arabidopsis thaliana MAP65-1 and MAP65-2 function redundantly withMAP65-3/PLEIADE in cytokinesis downstream of MPK4. Plant Signal.Behav. 6: 743–747.
Sasabe, M., Soyano, T., Takahashi, Y., Sonobe, S., Igarashi, H.,Itoh, T.J., Hidaka, M., and Machida, Y. (2006). Phosphorylation ofNtMAP65-1 by a MAP kinase down-regulates its activity of micro-tubule bundling and stimulates progression of cytokinesis of to-bacco cells. Genes Dev. 20: 1004–1014.
Smertenko, A., Saleh, N., Igarashi, H., Mori, H., Hauser-Hahn, I.,Jiang, C.-J., Sonobe, S., Lloyd, C.W., and Hussey, P.J. (2000). Anew class of microtubule-associated proteins in plants. Nat. CellBiol. 2: 750–753.
Smertenko, A.P., Chang, H.Y., Wagner, V., Kaloriti, D., Fenyk, S.,Sonobe, S., Lloyd, C., Hauser, M.T., and Hussey, P.J. (2004). TheArabidopsis microtubule-associated protein AtMAP65-1: Molecularanalysis of its microtubule bundling activity. Plant Cell 16: 2035–2047.
Smertenko, A.P., Kaloriti, D., Chang, H.Y., Fiserova, J., Opatrny,Z., and Hussey, P.J. (2008). The C-terminal variable region speci-fies the dynamic properties of Arabidopsis microtubule-associatedprotein MAP65 isotypes. Plant Cell 20: 3346–3358.
Smertenko, A.P., Piette, B., and Hussey, P.J. (2011). The origin ofphragmoplast asymmetry. Curr. Biol. 21: 1924–1930.
Staehelin, L.A., and Hepler, P.K. (1996). Cytokinesis in higher plants.Cell 84: 821–824.
Subramanian, R., Wilson-Kubalek, E.M., Arthur, C.P., Bick, M.J.,Campbell, E.A, Darst, S.A., Milligan, R.A., and Kapoor, T.M.(2010). Insights into antiparallel microtubule crosslinking by PRC1,a conserved nonmotor microtubule binding protein. Cell 142: 433–443.
Van Damme, D., Bouget, F.Y., Van Poucke, K., Inzé, D., andGeelen, D. (2004a). Molecular dissection of plant cytokinesis andphragmoplast structure: A survey of GFP-tagged proteins. Plant J.40: 386–398.
Van Damme, D., Van Poucke, K., Boutant, E., Ritzenthaler, C.,Inzé, D., and Geelen, D. (2004b). In vivo dynamics and differentialmicrotubule-binding activities of MAP65 proteins. Plant Physiol.136: 3956–3967.
Walczak, C.E., and Shaw, S.L. (2010). A MAP for bundling micro-tubules. Cell 142: 364–367.
Zeng, C.J., Lee, Y.R., and Liu, B. (2009). The WD40 repeat proteinNEDD1 functions in microtubule organization during cell division inArabidopsis thaliana. Plant Cell 21: 1129–1140.
Zhang, D.H., Wadsworth, P., and Hepler, P.K. (1990). Microtubuledynamics in living dividing plant cells: Confocal imaging of micro-injected fluorescent brain tubulin. Proc. Natl. Acad. Sci. USA 87:8820–8824.
Zhu, C.J., and Jiang, W. (2005). Cell cycle-dependent translocationof PRC1 on the spindle by Kif4 is essential for midzone formationand cytokinesis. Proc. Natl. Acad. Sci. USA 102: 343–348.
Zhu, C.J., Lau, E., Schwarzenbacher, R., Bossy-Wetzel, E., andJiang, W. (2006). Spatiotemporal control of spindle midzone for-mation by PRC1 in human cells. Proc. Natl. Acad. Sci. USA 103:6196–6201.
MAP65-3 Contains a Unique C Terminus 2085

DOI 10.1105/tpc.111.092569; originally published online May 8, 2012; 2012;24;2071-2085Plant Cell
LiuChin-Min Kimmy Ho, Yuh-Ru Julie Lee, Lindsay D. Kiyama, Savithramma P. Dinesh-Kumar and Bo
Domaintoward Their Plus Ends in the Phragmoplast via Its Distinct C-Terminal Microtubule Binding
Microtubule-Associated Protein MAP65-3 Cross-Links Antiparallel MicrotubulesArabidopsis
This information is current as of June 12, 2018
Supplemental Data /content/suppl/2012/05/02/tpc.111.092569.DC1.html
References /content/24/5/2071.full.html#ref-list-1
This article cites 55 articles, 27 of which can be accessed free at:
Permissions https://www.copyright.com/ccc/openurl.do?sid=pd_hw1532298X&issn=1532298X&WT.mc_id=pd_hw1532298X
eTOCs http://www.plantcell.org/cgi/alerts/ctmain
Sign up for eTOCs at:
CiteTrack Alerts http://www.plantcell.org/cgi/alerts/ctmain
Sign up for CiteTrack Alerts at:
Subscription Information http://www.aspb.org/publications/subscriptions.cfm
is available at:Plant Physiology and The Plant CellSubscription Information for
ADVANCING THE SCIENCE OF PLANT BIOLOGY © American Society of Plant Biologists