Embed Size (px)
Citation preview

Biotechnology Advances 18 (2000) 267–288
0734-9750/00/$ – see front matter © 2000 Elsevier Science Inc. All rights reserved.
PII: S0734-9750(00)00037-9
Research review paper
Are microbes intelligent beings?:An assessment of cybernetic modeling
P.R. Patnaik*
Institute of Microbial Technology, Sector 39-A, Chandigarh-160 036, India
Abstract
Microorganisms growing in a multi-substrate medium have different and varying preferences forthe various components of the medium. The preferences depend on the operating conditions and thesubstrates may be utilized sequentially or simultaneously. Sometimes an organism may change itspreferences among substrates and/or switch between sequential and simultaneous utilization. Theseaspects are difficult to describe through models based on chemical and physical laws alone. Cyber-netic modeling ascribes to microorganisms the ability to perceive their environment (i.e. the growthmedium) and make ‘intelligent’ choices regarding substrate utilization to maximize an objective,which is usually the growth rate. This article reviews the development of cybernetic modeling since itbegan in 1982. Different workers have suggested different perspectives of how microbes make opti-mal use of their resources. These are discussed and future directions for improvement areindicated. © 2000 Elsevier Science Inc. All rights reserved.
Keywords:
Microbial growth; Metabolic modeling; Cybernetic modeling
1. Introduction
Quantitative prediction of cellular growth on a macroscopic scale is at the heart of mostmicrobial processes. Growth consumes nutrients, may generate products, changes the rheo-logical and the thermodynamic properties of the medium, influences the stability of the reac-tor, and has a bearing on the subsequent (downstream) processing steps.
The classical approach to modeling microbial growth is derived from the seminal work ofMonod (1942), who used either one nutrient only or a mixture of nutrients in which all ex-cept one were in excess. In the latter case, growth was controlled by the availability of thenutrient that was not in excess. This approach leads to mechanistic models based on this sin-
* Tel.:
1
91-172-690223; fax:
1
91-172-690585.
E-mail address
: [email protected] (P.R. Patnaik)

268
P.R. Patnaik / Biotechnology Advances 18 (2000) 267–288
gle nutrient. While being attractively simple, the Monod approach has limited applicability toreal situations. Real fermentation media are rarely of the ‘minimal type’ (i.e. with only onelimiting component). Most media are complex in nature, and sometimes contain additionalcomponents. There are at least two reasons for using multi-component media. One is cost;naturally occurring media are cheaper than synthetic or purified substances. The second rea-son is that good growth depends on the optimal availability of a number of nutrients whichparticipate in the cellular reactions. The classical approach cannot satisfactorily describegrowth in multi-component environments without considerable prior information.
A conspicuous feature of microbial growth that illustrates the difficulties of the Monodmethodology is evident even with just two or three components (substrates). Under someconditions the substrates are utilized one at a time (e.g. diauxie or triauxie) while under dif-ferent conditions they are consumed simultaneously, although not necessarily at equal rates(Kompala, 1984). Moreover, the relative preferences among substrates depends both on theorganism and on the operating conditions. It was initially believed that glucose was the sub-strate most preferred by most organisms and this led to diauxie being termed the ‘glucose ef-fect’. Many cases are now known of microorganisms that do not utilize glucose preferen-tially.
Arthrobacter crystallopoietes
prefers succinate over glucose (Krulwich and Ensign,1969),
Propionibacterium shermanii
utilizes lactate in preference to glucose (Lee et al.,1974), and, depending on the culture conditions,
Escherichia coli
may have lower prefer-ences for glucose than for galactose, maltose, and arabinose (Lendenmann et al., 1996).
Narang’s (1994) studies of the growth of
E. coli
on mixtures of glucose and organic acidsrevealed shifts between sequential and simultaneous consumption. With the exception ofglucose–pyruvate mixtures, diauxic growth was observed in all cases where an organic acidwas paired with glucose. Simultaneous utilization with a growth rate between those on pureglucose and on pyruvate was observed when the inoculum was precultured on pyruvatealone, but diauxic growth (sequential utilization) occurred if preculturing was on glucose.Similar observations have also been reported for
Pseudomonas oxaliticus
in a medium ofoxalate and formate (Dijkhuizen et al., 1980), and for
Hansuela polymorpha
grown onmethanol and glucose (Egli et al., 1986), indicating that metabolic switching between dif-ferent utilization (and growth) patterns is not restricted to any particular kind of microor-ganism.
In a minimal medium, microbial growth rate is controlled by the one nutrient in limitedsupply and the organism has to decide how best to utilize this for different functions such asmaintenance of biomass composition, homeostasis, synthesis of proteins and other material.In a nonminimal (multi-substrate or complex) medium, there are additional choices in termsof the utilization patterns of the components of the medium (Pavlou and Fredrickson, 1989).As shown by the studies just cited, the utilization patterns may change with the prevailingconditions. Thus, while a constant utilization pattern may be possible at steady state in con-tinuous fermentation, this is unlikely in batch or fed-batch operation. For Monodic growthrate models to be able to express these features, such ‘informed choices’ have to be includeda priori in the modeling framework. However, this is not easy because: (1) not always are thechoices known in advance; (2) mutations and genetic changes can alter the preferential pat-tern; (3) perturbations in the operating condition influence a prevailing utilization pattern;and (4) the choices change with time in dynamic processes.

P.R. Patnaik / Biotechnology Advances 18 (2000) 267–288
269
To overcome these limitations, Ramkrishna (1982) introduced a different perspective ofmicrobial growth. His
cybernetic method
views microorganisms as ‘optimal strategists’. Thismeans that, given a set of conditions, they have the ability to ‘think’ and ‘decide’ how best toutilize the resources so as to maximize a particular objective. This may be distinguished frommechanistic modeling, which is based on chemical and physical laws alone. While Ramkrishnadeveloped a systematic and quantitative method to describe microbial growth in a laboratoryenvironment (a bioreactor), the cybernetic perspective has long been a fundamental tenet ofevolutionary biology. As Demain (1971) observed, “Microorganisms have evolved over theyears, developing better and better mechanisms to prevent overproduction of their metabo-lites. . . . All microorganisms must possess regulatory (control) mechanisms. . . . In fermenta-tion organisms, controls are less rigid but nevertheless present.”
In extending the ‘optimal strategy’ approach from adaptation in a natural environment to adesired mode of function in a controlled environment, cybernetic modeling translates theidea that cells regulate their activities by exerting control over the activities and the rates ofsynthesis of enzymes (Pavlou and Fredrickson, 1989). Indeed, the idea of microorganismsorganizing their metabolic activities to achieve an objective optimally was conceived longago when Bremmermann (1967) stated that “most biological systems are goal oriented. . . .A goal seeking system is called cybernetic if it pursues its goal while interacting with an en-vironment.” However, in order to keep the resulting equations sufficiently simple, details ofthe mechanisms of regulation are absorbed into the optimality criteria. This idea will becomeclearer as the models are discussed.
2. The cybernetic framework
An early version of Ramkrishna’s (1982) approach was applied by Dhurjati et al. (1985) tothe growth of
Klebsiella pneumoniae
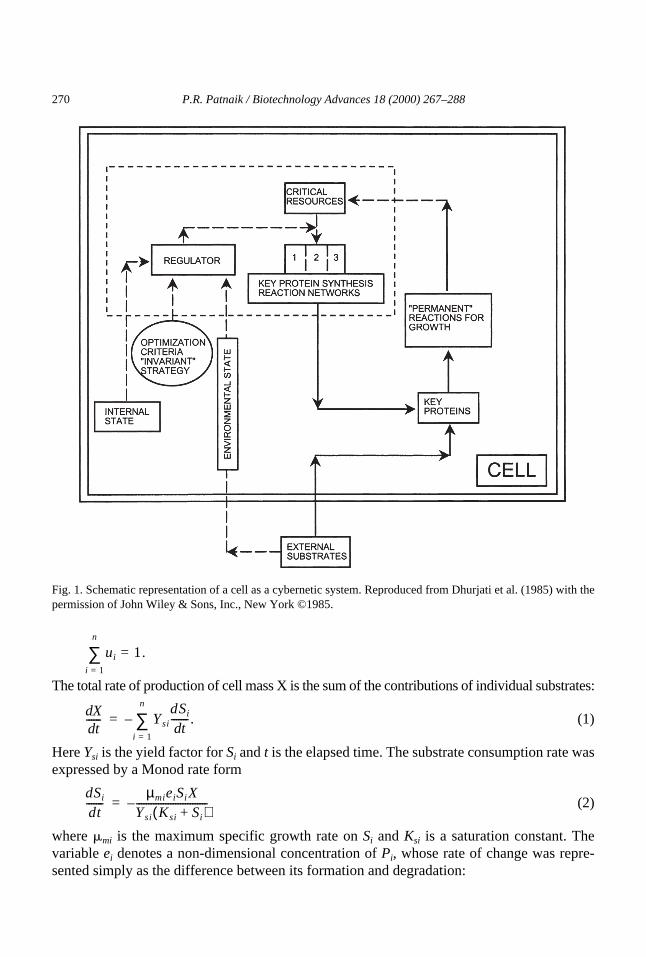
in a batch culture containing two limiting carbonsources, D-glucose and D-xylose. The ‘adaptive machinery’ controls metabolic changes inresponse to variations in the extracellular environment; it controls the synthesis of key pro-teins and includes a resource pool whose optimal allocation is the basis of cybernetic model-ing. Once the key proteins are synthesized, a ‘permanent machinery’ carries out the reactionsnecessary for the replication of cellular material. The third component is a ‘regulator’ (possi-bly ‘regulating machinery’ would be more consistent with the names of the other two), whichperforms the cybernetic functions; it continually allocates resources to the protein synthesissystems of the adaptive machinery. This would imply inter-connections between the threecomponents, as shown in Fig. 1. Although the components are conceptual entities for themodel, they have physiological counterparts in living cells (Apter, 1966); for instance, theregulator functions are carried out by certain promoter and operator sites of the operon.
Dhurjati et al. (1985) consider
n
substrates,
S
1
,
S
2
, . . .,
S
n
, to be involved in the synthesis ofan equal number of proteins,
P
1
,
P
2
, . . .,
P
n
, in a batch fermentation. This implies a one-to-one correspondence between a resource and a product, a restriction which was later removed.Let
R
i
be the allocation rate of
S
i
to
P
i
, and
R
the total allocation rate; then
u
i
5
R
i
/R is thefractional allocation rate of
S
i
to
P
i
. Based on Mandelstam and McQuillen’s (1968) work,Dhurjati and coworkers considered R to be constant, which results in the constraint
270
P.R. Patnaik / Biotechnology Advances 18 (2000) 267–288
The total rate of production of cell mass X is the sum of the contributions of individual substrates:
(1)
Here
Y
si
is the yield factor for
S
i
and
t
is the elapsed time. The substrate consumption rate wasexpressed by a Monod rate form
(2)
where
m
mi
is the maximum specific growth rate on
S
i
and
K
si
is a saturation constant. Thevariable
e
i
denotes a non-dimensional concentration of
P
i
, whose rate of change was repre-sented simply as the difference between its formation and degradation:
ui 1=i 1=
n
∑ .
dXdt------ Ysi
dSi
dt-------.
i 1=
n
∑–=
dSi
dt-------
µmieiSi XYsi Ksi Si+( )-----------------------------–=
Fig. 1. Schematic representation of a cell as a cybernetic system. Reproduced from Dhurjati et al. (1985) with thepermission of John Wiley & Sons, Inc., New York ©1985.

P.R. Patnaik / Biotechnology Advances 18 (2000) 267–288
271
(3)
Now, the cell’s regulator may apportion resources to maximize growth in either of two ways,as discussed next.
2.1. The long-term perspective
This view of cellular optimality postulates that resource allocation is done in such a waythat the total growth of cell mass over a specified duration,
t
f
, is maximized. Mathematicallythis may be stated as:
(4)
where
5
dX
/
dt.
The maximization is subject to the physical limit that growth ceases whenthe substrates are exhausted; this is expressed by a constraint on Eq. (4)
(5)
where
S
i
0
is the value of
S
i
at
t
5
0.Dhurjati et al. (1985) adopted this perspective for the batch cultivation of
K. pneumoniae
and derived a bang-bang policy for allocation of either glucose or xylose. This policy statesthat initially only one substrate (glucose) is utilized, resulting in the production of the corre-sponding key enzyme
E
1
, and then the other substrate is consumed after the concentration ofthe first has become very low. Although the long-term perspective portrayed the growth of
K.pneumoniae
, it had some inherent limitations, two of which led to its modification. One is thebang-bang utilization policy itself. This is suitable for diauxic growth but not if both (or all)components of the growth medium are utilized simultaneously. The limitation is a conse-quence of the assumption in Eq. (3) that enzyme synthesis rates depend linearly on the frac-tional allocations of resources,
u
i
. The second limitation is the inability of the model to adaptthe optimal policy to perturbations in the operating conditions. Such perturbations are com-monly expected in fed-batch and continuous fermentations but they also occur in batch oper-ations (Patnaik, 1996).
2.2. The short-term perspective
To overcome the difficulties posed above, Kompala and associates (Kompala et al., 1984)postulated an alternate short-term perspective, in which the microorganism allocates theavailable resources such that its growth rate is maximized at every instant of time. Here againthere are two strategies. One strategy is that, at any instant, the organism chooses to synthe-size the enzyme
Ej for the substrate Sj which maximizes the instantaneous acceleration of thebiomass growth rate. This means the fractional allocation uj is calculated from time to timeby maximizing d2X/dt2. Kompala et al. (1984) showed that this method also yields a bang-bang policy. Another problem with this strategy is that it ignores the effect of enzymespresent at the start of a process. To illustrate, suppose the cells are pre-cultured on a substrateS2 and then grown on a mixture of S1 and S2. Initially there is some concentration of E2, so
dei
dt------- aiui biei.–=
maxu
Xdt0
t f
∫
X·
Si0 Sidt ε ≈ 0=0
t f
∫+i 1=
n
∑

272 P.R. Patnaik / Biotechnology Advances 18 (2000) 267–288
the cells will consume S2 in the presence of S1 even if u2 5 0, but the instantaneous biomassmaximization strategy does not take account of such a feature. However, it is able to accom-modate perturbations because the policy is updated from point to point in time.
The second strategy, which is recommended by Kompala et al. (1984), is based on Herrn-stein’s (1974) matching law. Suppose a resource ri is allocated to the i-th choice and pi is theresulting return. Then the law of equal marginal utilities, which is analogous to the matchinglaw, implies that for each i, dpi/dri is non-negative and d2pi/dri
2 is nonpositive. The totalreturn
for n resources is maximized under the constraint
where r is the total amount of resource, when
(6)
which leads to
(7)
Eq. (7) is the matching law statement that the fractional allocation must match the fractionalreturn. Note also that the constraint
degenerates to
if ui is defined as in Dhurjati et al. (1985) and r is constant.Kompala et al. (1984) pointed out that the key mechanism for the control of resource allo-
cation is at the level of transcription of the DNA by the RNA polymerase to generate the mR-NAs required for the synthesis of proteins. This control is exercised through the distributionof the times of transcription for the catabolic key enzymes, the total time being limited byother required activities of the RNA polymerase. If rBi is the rate of growth of biomass on Si,then the matching law provides that
(8)
If ui 5 0, the polymerase does not transcribe the gene for Ei; if ui 5 1 then obviously only Ei
is induced.
pi
i 1=
n
∑ ri( )
ri r=i 1=
n
∑ .
dp1
dr1--------
dp2
dr2-------- . . .
dpn 1–
drn 1–--------------
dpn
drn
--------=== =
ui
dri
dr j∑-------------
dpi
dp j∑--------------.= =
ri r=i 1=
n
∑
ui 1=i 1=
n
∑
ui rBi rBj.j 1=
n
∑⁄=

P.R. Patnaik / Biotechnology Advances 18 (2000) 267–288 273
To incorporate the effect of any enzymes present initially, let vi denote the fractional allo-cation of another critical resource required for the utilization of the i-th substrate. Then theactual rate of biomass production is:
(9)
Obviously,
The vi are called cybernetic variables. If vi 5 0, growth does not take place even if the sub-strate Si is present in the medium and its key enzyme Ei exists in the protoplasm. Likewise, avi-value of close to unity signifies strong activation.
The matching law method was able to portray sequential and simultaneous consumptionof glucose and lactose by K. aerogenes, arabinose and lactose by K. oxytoca, and a mixtureof glucose, sorbitol, and glycerol by E. coli H1, all in batch cultivation. It could also depictthe transient behavior of continuous fermentations upon abrupt changes in the feed stream,without a priori specification of the order of preference among the substrates.
3. Further developments
Despite its ability to predict diauxic and triauxic growth, as well as simultaneous and shift-ing rates of utilization from a group of substrates, the model proposed by Kompala and co-workers (Kompala et al., 1984) still had limitations. First, linear dependence of the enzymesynthesis rates on the cybernetic variables necessitated frequent changes in the bang-bangpolicy to account for parallel consumptions and changing patterns of substrate consumption(Pavlou and Fredrickson, 1989). Second, by assuming a constant genotype, the model did nottake account of mutations occurring during the cultivation period. Third, the matching lawformalism produced an unrealistic situation when the organism had equal preference for twosubstrates in a mixture. According to the model predictions, the growth rate on the mixturewill be the same as that on either substrate, whereas it should be their sum (i.e. double therate on one substrate).
A subsequent study (Kompala et al., 1986) addressed the first and third problems by defin-ing vi in Eq. (9) as:
(10)
Now the constraint
no longer applied. As before, the model was tested with K. oxytoca grown on single sub-strates (glucose, lactose, and arabinose) and with combinations of two or three of these sub-
vi
i 1=
n
∑ rBi 0 vi 1.< <;
vi 1=i 1=
n
∑ .
vi ri maxj
r j( ).⁄=
vi 1=i 1=
n
∑

274 P.R. Patnaik / Biotechnology Advances 18 (2000) 267–288
strates. While diauxic and triauxic growth were faithfully portrayed for preculturing on anyof the components, the model was not satisfactory for biphasic growth, as with a combinationof glucose and fructose. This kind of growth occurs when the two substrates are close iso-mers and thus compete for the same active sites on a common enzyme E (rather than E1 andE2). Sharing of the same binding sites translates to the original conservation condition on thevi, which had been dispensed with in Eq. (10). So, not surprisingly, the optimal resource allo-cation strategy was again of the bang-bang type:
(11)
Turner and Ramkrishna (1988) further modified Kompala et al.’s (1986) work to accommo-date an important observation by Standing et al. (1972). The latter authors grew E. coli incontinuous culture on glucose until steady-state was attained. The glucose in the feed streamwas then replaced by xylose. The model would predict a very slow rate of growth because ofthe absence of the key enzyme for xylose. However, the experimentally measured rates wereappreciably larger, suggesting that a certain amount of constitutive synthesis of the enzymealso occurred. The constitutive synthesis is not influenced by environment factors and wastherefore considered to receive a fixed fraction of the critical resource.
3.1. Maintenance metabolism
For the growth of heterotrophic bacteria, the mass of bacteria formed per unit mass of sub-strate consumed (i.e. the yield coefficient Yxs) is usually constant when there is sufficient sub-strate and the growth rate is high. At the lower growth rates that occur during high-conversioncontinuous culture, the apparent Yxs is lower and does not stay constant (O’Connor et al., 1992).This variation is normally taken care of by introducing a term for the cell’s energy requirementsfor its own maintenance (Pirt, 1975). The substrate consumption rate for this purpose is given by:
(12)
where m is the specific growth rate (g g21 h21), m is the maintenance coefficient (g g21 h21),and Yxs is the yield that would be observed if m were zero. Although the assumption of a con-stant value for m is sometimes a reasonable approximation, physiological arguments (Pirt,1975; Esener et al., 1983) favor a variable m. So, Turner et al. (1989) used a Monodic typeequation suggested earlier (Ramkrishna et al., 1966), to describe the rate of maintenance me-tabolism; thus:
(13)
In Eq. (13), mM is the specific growth rate during the synthesis of a key enzyme EM that isused in non-growth-associated processes, eM is the concentration of EM and s is the (single)substrate concentration.
Their approach (Turner et al., 1989) also considered a growth-associated enzyme EG,whose kinetics are similar to Eq. (13):
vi
1 if ri r j>0 if ri r j.<
=
qs m Yxs m+⁄=
rM
mMeMsKM s+----------------.=

P.R. Patnaik / Biotechnology Advances 18 (2000) 267–288 275
(14)
Because both EG and EM may be present at the same time, the actual specific growth andmaintenance rates are cybernetically moderated:
(15)
and
(16)
The cybernetic variables vG and vM control the activities of EG and EM. The similarity be-tween these equations and Eq. (9) is not surprising since they all refer to fractional alloca-tions of resources for different purposes.
Inclusion of maintenance metabolism enabled the model to display accurately the temporalvariations of glucose and biomass concentrations and the yield Yxs during perturbed fed-batchfermentations (Turner et al., 1989). Because the first phase of experiments were with only glu-cose, perturbations were in the form of glucose pulses that were sufficient to saturate the cells.Contrary to expectation, the cell growth rate responded slowly to the pulses, implying that thekey enzymes, EG and EM, at the low growth rates prior to increased induction were below theirmaximum possible activities. This reveals a key cybernetic observation that the capacity of thecells to synthesize the required enzymes is need-based or resource-based, and not simply aninanimate biochemical phenomenon. The organism in these experiments and in the secondphase with multiple substrates was the K. oxytoca strain used earlier (Kompala et al., 1986).
In the experiments with a multi-substrate medium (Turner et al., 1988), combinations ofglucose with arbinose or fructose or xylose were used. In all cases, glucose was the preferredsubstrate, S1. An interesting observation was that if a saturating pulse of the less preferredcomponent, S2, was added to a batch culture growing in a glucose-fed medium, the key en-zymes for S2 were quickly synthesized; the biomass concentration rose until S2 had been de-pleted, whereupon the cells returned to growth on S1. It may also be noted that the glucose–fructose mixture promoted biphasic growth whereas the other two combinations produced adiauxic behavior (Kompala et al., 1986).
4. Taking stock
The cybernetic framework discussed so far is based on the matching law approach,whereby the allocation of resources to different cellular activities is according to the contri-butions of the activities to the ultimate goal. The goal throughout has been the maximizationof the rate of biomass production at every point in time. This raises the first limitation. Notalways is growth maximization the primary objective, and this objective may change duringthe course of fermentation. For instance, when there is inhibition by any of the substrates orproducts, there is an optimal level of growth beyond which productivity declines (Moser,1988). Second, sometimes cultivation of high cell density is followed by a stage of product
rG
mGeGsKG s+----------------.=
rGvG 0 vG 1< <;
rMvM 0 vM 1.< <;

276 P.R. Patnaik / Biotechnology Advances 18 (2000) 267–288
formation at low growth, as for some recombinant proteins (Ryu et al., 1991). Cyberneticmodeling has not yet accounted for such ‘non-maximal’ operations.
The possibility of mutations, leading to genotypic changes, was alluded to by Dhurjati et al.(1985). While introducing constitutive enzyme synthesis, Turner and Ramkrishna (1988) high-lighted this as a genotypic property as contrasted with inducible enzyme synthesis, a pheno-typic event. They argued that constitutive synthesis consumes a fixed fraction of the critical re-source(s). But if there are genotypic changes, their resource utilization will vary with time,which the modeling does not accommodate.
The studies reported thus far have been confined to substitutable substrates, meaning thatone substrate can perform the functions of another. For example, with a blend of glucose,lactose, and xylose (Kompala et al., 1986), K. oxytoca grows on all of them but at differentrates and in different orders of preference. However, this is not always so. Pavlou and Fre-drickson (1989) have classified 11 nutrient (substrate) use patterns, depending on the pres-ence or absence of inhibitions among the components and between nutrients and non-nutri-ents. Products are a common class of non-nutrients, and inhibition by them affects thestability of a culture in many cases.
Although Turner et al. (1988) demonstrated the validity of their modeling approach fortwo- and three-substrate systems with perturbations in batch culture, applicability to continu-ous and fed-batch fermentations with inflow disturbances and different pre-culturing condi-tions has not been proven. Earlier studies (Kompala et al., 1984, 1986) have shown that pre-culturing can have an effect on the subsequent course of fermentation and so it remains to beseen whether more developed models can express these effects better than the earlier simplerversions.
5. Resource limitation and resource synthesis
A major assumption of Turner et al. (1988) was that the cells possessed sufficient quantity ofinternal resource for enzyme synthesis. This assumption may be valid for batch and fed-batchfermentations under balanced growth or under slowly changing conditions but not when thereare sizable perturbations. One such situation was discussed in Section 3.1 where a saturatingpulse of glucose added to a culture growing slowly under glucose limitation did not produce theexpected rapid increase in cell density, indicating a limiting effect other than the lack of en-zymes and substrate. Just as enzyme synthesis rates are low in a substrate-starved culture andrequire time to increase in response to a surfeit of substrate (Turner et al., 1988), so do otherintracellular resources. For example, with ribosomes (Harder and Roels, 1982), only the ac-tive fraction of the total may be thought of as the relevant resource.
Turner et al.’s (1988) experiments also showed substrate consumption values for mainte-nance that were much larger than predicted. Large cellular maintenance requirements at lowgrowth rates have been attributed to high rates of protein degradation (Pine, 1972) and the ac-cumulation of regulatory nucleotides (Arbrige and Chesbro, 1982; Ingraham et al., 1983). Toreconcile these observations, Baloo and Ramkrishna (1991a) added two features to the modelof Turner and coworkers. One feature was a low maintenance system, in addition to normalmaintenance, and the other was a resource synthesis term; thus,

P.R. Patnaik / Biotechnology Advances 18 (2000) 267–288 277
(17)
The rate of resource (ribosomal) synthesis varies linearly with the concentration, eR, of itskey enzyme, ER, and Monodically with the substrate concentration, s. This form and a first-order degradation rate are similar to the growth and maintenance rates proposed earlier(Turner et al., 1988, 1989). Formulation of the cybernetic model then rested on five postu-lates. Two key postulates that distinguish the present formalism from earlier ones are: (1)growth and maintenance processes compete for the limited cellular resources in such a waythat substrate consumption rate is maximized; and (2) cellular demand for internal resourcesincreases with the growth rate. The second statement is physiologically expected, but notethat the first postulate shifts the emphasis from growth maximization to that of substrate uti-lization. Although Baloo and Ramkrishna (1991a) do not explicitly say so, it is possible toread the implication that maximal substrate utilization also maximizes cell growth. However,as stated before (Moser, 1988; Ryu et al., 1991), this is not always the case. The direct mean-ing of the first postulate is that when there are adequate quantities of substrates/nutrients andinternal resources, growth maximization is the cell’s primary objective (Dhurjati et al., 1985;Kompala et al., 1986) but when the substrates (and consequently the internally synthesizedribosomal resources) are in short supply, their efficient utilization supersedes the growth objec-tive (Tempest and Neijssel, 1976). With this background, the model formulated by Turner et al.(1988, 1989) was recast to include terms for low maintenance and resource synthesis, togetherwith corresponding cybernetic variables. So, now there are four cybernetic variables: (1) thefraction of resources allotted for (total) maintenance; (2) the fractional allocation for growth; (3)a maintenance modulaton variable to maximize the substrate consumption rate; and (4) a vari-able to regulate resource synthesis according to metabolic demand and substrate availability.
Baloo and Ramkrishna (1991a) applied the expanded cybernetic model to the growth of K.pneumoniae (ATCC13882). (The reasons for the choice of K. oxytoca in some studies and K.pneumoniae in others are not clear.) Results for batch and fed-batch cultures with perturba-tions in the form of glucose pulses showed that the model performed as well as, but appar-ently no better than, the simpler versions of Kompala et al. (1986) and Turner et al. (1988). Asimilar assessment also applies to steady-state continuous flow operation, for which an oldermodel (Pirt, 1982) also predicted the measured convex profiles of cell density versus the di-lution rate. The advantage of the additional features incorporated by Baloo and Ramkrishna(1991a) is realized in transient continuous cultures. Separate experiments with glucose andxylose media and increases or decreases in dilution rate by up to 400% showed that the newversion could predict the observed lag period during which the cells were adjusting to thechanged environment and the overshoot (or undershoot) in cell density before a new steadystate was reached. Neither phenomenological (Monodic) descriptions nor the earlier cyber-netic models could predict such behavior.
Baloo and Ramkrishna (1991b) generalized their modeling to multi-substrate systems byincreasing the number of cybernetic variables. To understand the magnitude of the increase,let us examine the resource synthesis rate expressed as follows:
(18)
rR α R
eR
eRm
-------- sKRs s+( )
----------------------.=
rR α R ,i
i 1=
n
∑eR ,i
eR ,im
----------si
KRs ,i si+( )--------------------------vR.=

278 P.R. Patnaik / Biotechnology Advances 18 (2000) 267–288
For n substrates, the number of parameters (cybernetic and biochemical) is (4n 1 1). Thus, achange from one to three substrates, which is not uncommon in many real media, increasesthe number of parameters to be estimated from 5 to 13. Using the heuristic guideline that 2n
data should be used to obtain unbiased estimates of n parameters (Draper and Smith, 1981),the number of data required jumps from 32 to 8192.
Despite the weakness of having many parameters, a redeeming feature of Baloo andRamkrishna’s (1991b) methodology is its ability to depict the observed transient profiles incontinuous cultures. Initially, a culture (K. pneumoniae) growing on glucose was switchedto xylose feed of the same concentration (0.5 g l21). In later experiments, a mixture of bothsubstrates was employed. Of particular interest in the steady-state results is the observationthat at low growth (and dilution) rates the two substrates are utilized simultaneously, whileat high dilution rates glucose is metabolized first. Slow growth results in higher cell density,suggesting that real reactor operation may have to strike a balance between the cell densitythat is possible and the rate of attainment. One set of transient studies focused on the re-sponse following a step-up in dilution rate from 0.352 to 1.04 h21. The model and the exper-iments showed the cells switching from simultaneous to sequential utilization. The reversetrend occurred upon a step-down in the dilution rate (from 1.0 to 0.281 h21).
6. Transport across the cell wall
The cybernetic framework developed so far is pictorially summarized in Fig. 2, which alsodistinguishes the cellular (biotic) phase from the extracellular (abiotic) environment. Con-centrations in the biotic phase are marked by a tilde (~). This distinction between phases wasintroduced by Straight and Ramkrishna (1991), and Fig. 2 pertains to lactose metabolism byK. oxytoca (not K. pneumoniae).
One significant departure of Fig. 2 from earlier formalisms is the inclusion of lactosetransport across the cell wall. There are three transport mechanisms. One is a high-affinityprocess which consumes a key resource p; the second is a low-affinity process (slip trans-port) that does not consume p but requires a higher level of extracellular lactose SL in orderto be fully active. Both processes are assumed to be catalyzed by the same induced enzyme,
T,L, and follow substrate-inhibition kinetics. The third mechanism is designed to compen-sate for the reduction in the concentration of p from its optimal value by activating anotherinflow route for lactose which does not consume p:
(19)
The enzyme TL,L is itself generated from TL without the involvement of p; Straight andRamkrishna (1991) suggest that this may be the conversion of the lac permease from dimericto monomeric form, as indicated by Villarejo (1980 ).
In the biotic phase, lactose is hydrolyzed to galactose and glucose. To prevent excessiveaccumulation of glucose in the biotic phase (Crabtree effect), feedback inhibition of lactosetransport by glucose and galactose is incorporated. As before (Kompala et al., 1986; Turneret al., 1988, 1989), there are reactions and enzymes for growth, resource synthesis and utili-
S~
S~
E~
S~
S~
SL B+ETL L,
→ SL B.+
E~ E~ S~

P.R. Patnaik / Biotechnology Advances 18 (2000) 267–288 279
zation, and maintenance. A novel feature is the efficiency of carbon consumption for p syn-thesis; this is represented as follows:
(20)
where rp,j is the synthesis rate of p, rx,j is the rate of biomass formation, and j stands for glu-cose (j 5 1) or galactose (j 5 2). vp and vx are cybernetic variables. Modeling the wholetransport–reaction system requires 13 cybernetic variables plus many others related to thebiochemical reactions.
Given its detailed biochemistry and metabolic controls, this model could express otherfeatures of complex growth dynamics such as intermittent changes in the rate of cell densityincrease and oscillatory behavior. The yield coefficient Yxs has been predicted to pass througha minimum as lactose concentration is varied from 0.5 to 5.0 g l21. However, the scatter ofthe experimental data and the predicted variations in the rates of change of cell density andthe yield coefficient are such that it is difficult to draw definite conclusions, as Straight andRamkrishna (1991) have done, that the variations are indeed significant. Moreover, oscilla-tory profiles can be produced also with much simpler models (Jarbezski, 1992).
7. Other applications
Since cybernetic modeling of microbial growth was introduced by Ramkrishna (1982), it isunderstandable that many of the initial developments were from this research group. How-ever, lately a number of other researchers have begun to apply cybernetic methods and somegroups have employed quite different approaches compared to earlier ones. Notable amongthese other groups is that of Venkatesh, who modified the modeling for easy determination
S~
rp , j rx , jvx+( )vp 0 vx 1< <;
S~
Fig. 2. Key enzyme mediated processes which determine the primary carbon flow into and through a cell. SL 5abiotic lactose; L 5 biotic lactose; Gl 5 biotic glucose; Ga 5 biotic galactose; P 5 abiotic key energyresource; X 5 a byproduct; B 5 biomass. Reproduced from Straight and Ramkrishna (1991) with the permis-sion of John Wiley & Sons, Inc., New York ©1991.
S~
S~
S~
S~
S~

280 P.R. Patnaik / Biotechnology Advances 18 (2000) 267–288
of the parameters, physical interpretation, and control. Gadgil and Venkatesh (1997) studiedthe batch cultivation of Lactobacillus bulgaricus with b-galactosidase as the product of inter-est. The model of Kompala et al. (1984) was modified to account for product inhibition andpH. It is known (Ishizaki et al., 1989) that lactate ions inhibit growth in an uncompetitivemanner; this was included in the model by reducing the maximum specific growth rate mm,by a factor (11p/KI) where p is the concentration of lactate and KI is an inhibition constant.Dependence of b-galactosidase activity on pH was introduced by making the enzyme degra-dation constant in Eq. (3) a function of pH. Unlike Ramkrishna and coworkers, Gadgil andVenkatesh (1997) used a complex medium whose composition is specified in their publica-tion; lactose was the main carbon source.
Even a simple model with nine parameters (of which three are cybernetic) accurately ex-pressed the cell concentration and substrate concentration profiles over a range of pH valuesfrom 4.2 to 5.6 and initial lactate concentration from 20 to 75 g l21. Because a complex sub-strate was used, it is not clear how its concentration was defined; presumably the lactose con-centration was measured at different times. The predicted enzyme concentration profiles ver-sus time passed through maxima, which is expected in view of the opposing effects ofsynthesis and degradation. The highest peak was attained for pH 5.6, a value supported bythe observation of Shah and Jelen (1990).
In another study (Gadgil et al., 1996) the same approach was applied to a somewhat morecomplex fermentation, the growth of recombinant Saccharomyces cerevisiae PJB 3 on meli-biose. While the cybernetic equations for cell growth, enzyme synthesis and degradation, andsubstrate uptake followed Kompala et al. (1986), an additional equation was introduced forthe enzymatic breakdown of melibiose to glucose and galactose; this was expressed by amodified Henri-Michaelis-Menten form:
(21)
where v2 and vm are cybernetic variables for galactose and melibiose respectively (v1 is forglucose), cmel and emel are the concentrations of melibiose and its key enzyme, while Kmel isthe usual equilibrium constant. It may be noted that although glucose does not occur in Eq.(21), it affects the degradation because, beyond a critical concentration, it represses the for-mation of a-galactosidase, the enzyme responsible for this reaction.
Subsequently, Venkatesh and associates questioned some aspects of the Ramkrishna for-malism and offered their own perspective of cybernetic growth. They objected to the largenumber of parameters, some of which related to intracellular phenomena that could not bemeasured easily, and the assumption that only the precursor concentration was controlling.In the first of two studies (Doshi et al., 1997), each bacterial cell was conceptually dividedinto as many compartments as the number of substrates, implying that the fraction ai of thetotal cell mass, X, utilized only the i-th substrate Si. So the overall growth rate could be ex-pressed as:
(22)
rd
v2vm emel emax
mel⁄( )cmel
Kmel cmel+-----------------------------------------------=
dXdt------- . α iri.
i∑

P.R. Patnaik / Biotechnology Advances 18 (2000) 267–288 281
where ri, the growth rate on Si, had the Monodic form suggested by Kompala et al. (1986). Be-cause the total enzyme pool is constant (in batch fermentation), the key enzyme synthesis ratefor Si was assumed to be proportional to the fraction of the total population utilizing Si. Thismeant that, unlike Kompala et al. (1986) and Turner et al. (1988), the consumption rate of Si
was multiplied by ai instead of ui 5 ri/Sri. Thus,
(23)
where ei is the dimensionless concentration of the key enzyme Ei, Si is the concentration ofthe i-th substrate, mmi and Ksi are the maximum specific growth rate and equilibrium constantfor Si, respectively, and b is the usual degradation constant. The cybernetic objective was todetermine the ai’s by maximizing the specific growth rate, (1/X)dX/dt. Doshi et al. (1997)showed that their model portrayed the results of Kompala et al. (1986) for K. oxytoca on glu-cose–xylose, glucose–lactose, and glucose–xylose–lactose mixtures, as well as their own ob-servations for S. cerevisiae PJB 3 growing on a combination of glucose and galactose. Themain benefit claimed is a two-fold reduction in the number of parameters compared to Ko-mpala et al. (1986) and Turner et al. (1988). However, this figure applies to extracellular con-centrations only because Doshi et al. (1997) did not include intracellular variables.
The validity of compartmentalizing the cell such that each portion is dedicated to one sub-strate and there is no interaction may also be questioned. Presumably to establish a physio-logically more reasonable basis for their model, a subsequent study (Venkatesh et al., 1997)adopted a very different perspective, that may be explained with the aid of Fig. 3. Every sub-strate is utilized by the microorganism through an independent pathway consisting of severalreactions producing a common intermediate. Each pathway has a driving force and a resis-tance, whose combined effect is reflected in the specific growth rate, mi, for that substrate.Beyond the formation of the intermediate is a common pathway characterized by a maximumspecific growth rate, mm. In the presence of many substrates, different fractions of the totalflux through the cell go through the individual pathways, and these fractions may changewith time. If ai is the fraction directed toward the i-th pathway and ri is the correspondinggrowth rate, the total biomass growth rate may be written as follows:
(24)
Note that Eq. (24) is the same as Eq. (22) but the cybernetic parameters ai have a differentphysical interpretation. As before, the microorganism was considered to organize its metabo-lism to maximize its total specific growth rate subject to conditions on the individual specificgrowth rates vis-à-vis the total rate (Venkatesh et al., 1997).
This model was tested satisfactorily with E. coli K-12 on combinations of glucose, acetate,lactate, and pyruvate, taken two at a time. It was shown that during the growth process therecan be a change from sequential to simultaneous utilization of the substrates and back to se-quential utilization, as observed by Narang (1994). This suggests the possibility of control-ling the utilization pattern (Pavlou and Fredrickson, 1989) by manipulating the operating condi-tions continually in fed-batch or continuous operation (O’Connor et al., 1992). However, theupdated model still does not address intracellular variables or more than two substrates. On a
dei
dt-------
α i mmi b+( )Si
Ksi Si+--------------------------------- ei
d X( )lndt
------------------ bei––=
rdXdt------- α iri.
i∑= =

282 P.R. Patnaik / Biotechnology Advances 18 (2000) 267–288
fundamental level, the authors have not explained why, from a common pool of reactants in acell, the biochemical pathways for different substrates must be mutually exclusive but con-verge to a common intermediate (Fig. 3). Because simplicity is claimed to be a major advan-tage, it would have been informative if Venkatesh et al. (1997) had compared the perfor-mance of their approach with that of Ramkrishna and coworkers in terms of CPU time,number of floating point operations (flops), etc. for different examples.
Like Venkatesh and coworkers, Yoo and Kim (1994) also differ with the Ramkrishnaschool in their concept of cybernetic modeling. They studied the formation of poly-b-hy-droxybutyric acid (PHB) intracellularly by Alcaligenes eutrophus NCIB11599. PHB is anenergy and carbon reserve material which can be synthesized by many prokaryotes under un-balanced growth conditions. Its potential applications include absorbable prosthetic devices,surgical sutures, food packaging, controlled release of drugs and piezoelectric sensors(Holmes, 1985). PHB synthesis by A. eutrophus is induced by stress in the environment, usu-ally by a high concentration of the carbon source, while the synthesis of protein and/or nu-cleic acid is inhibited by the lack of nitrogen (Oeding and Schlegel, 1973). So, Yoo and Kimbased their modeling on two postulates: (1) the cells can be divided into PHB and biomass(X) and the latter is the catalytically active component containing proteins and nucleic acid;and (2) the carbon source is optimally allocated to synthesis of the key enzymes so that thecells have a high degree of flexibility under nitrogen starvation.
Following earlier work (Dhurjati et al., 1985; Kompala et al., 1986), the growth rate ofbiomass and the synthesis rate of PHB were written in the form of modified Monod kinetics.Each step had a key enzyme, E1 or E2. Thereafter, Yoo and Kim departed from Herrnstein’s(1974) matching law in allocating resources. They considered that the fractional allocation ofa resource vi was given by:
(25)
where i 5 1 denotes the nitrogen source and i 5 2 the carbon source. The activities of the en-zymes E1 and E2 were regulated by the cybernetic variables as:
vi
exp ri( )exp r j
j∑( )
------------------------ i 1,2 j; 1,2==;=
Fig. 3. Schematic representation of converging biochemical pathways in cellular metabolism. Reproduced fromVenkatesh et al. (1997) with the permission of John Wiley & Sons, Inc., New York ©1997.

P.R. Patnaik / Biotechnology Advances 18 (2000) 267–288 283
(26)
In Eqs. (25) and (26), r1 is the rate of biomass formation and r2 that of PHB synthesis. Thesynthesis rates of the key enzymes were somewhat different from those employed by Turneret al. (1988, 1989) and Venkatesh et al. (1997) to allow simultaneous or sequential utilizationof the carbon and nitrogen sources in either the growth-associated or the non-growth-associ-ated phase.
For batch cultures in a medium containing glucose, ammonium sulfate, and nutrients, Yooand Kim (1994) compared their model with three classical unstructured models—a modifiedLuedking–Piret equation (Moser, 1988), a modified logistic equation (Mulchandani et al.,1989), and a Monod equation (Asenjo and Suk, 1985). The cybernetic model followed theexperimental data more accurately than the others over a 60-h period, and it expressed fea-tures such as the initial lag in PHB accumulation, which some of the other models could not.However, no perturbations were applied. As pointed out by Turner et al. (1988, 1989) andBaloo and Ramkrishna (1991a,b), it is in disturbed fermentations that the merits of a cyber-netic method are truly tested.
Alexander and Ramkrishna’s (1991) study of sideropore production by E. coli K-12 is asimilar departure from the earlier studies of other workers with Ramkrishna. Sideropores areiron-chelating agents produced in iron-limiting conditions to scavenge iron from the me-dium. The sideropores are excreted into the medium, where they form complexes with anyavailable ferric iron; receptors in the cell membrane help in the transport of the complexesinto the cells, where they are released in the ferrous state (Raymond and Carrano, 1979). E.coli K-12 produces the sideropore enterochelin, a trimeric molecule made up of 2,3-dihy-droxy benzoylserine (DHBS) monomer units.
The carbon substrate is utilized primarily for two respiration processes, and the resultingenergy is then used either for biomass production (growth) or for cell maintenance. Acetateis a key product of the respiratory pathway; it is produced moderately when there is suffi-cient iron in the medium and strongly when there is a shortage of iron. Sideropore synthesisis catalyzed by a specific key enzyme, which is induced by a lack of intracellular iron, butthere is also some constitutive synthesis. Alexander and Ramkrishna (1991) preferred aBlackman kinetic form for sideropore formation because their data indicated a faster satura-tion at lower iron levels than could be provided by a Monod form.
Transport of iron into the cell is the ultimate goal of sideropore synthesis. In the absenceof adequate understanding of the mechanisms of transport, Alexander and Ramkrishna(1991) explained their data by proposing a low-affinity transport and a high-affinity trans-port; each has a primary mechanism and a secondary mechanism. Within the cells, the sid-eropore-iron complex is degraded into its monomer units. Six cybernetic variables controlthe metabolic processes: (1) vM for the maintenance metabolism; (2) vLT and vHT for the low-and high-affinity transport processes, respectively; (3) vR1 and vR2 for the two respiration pro-cesses; and (4) vS for sideropore formation. Equations for them are similar to those proposedearlier (Kompala et al., 1986; Turner et al., 1988).
The complete model had 32 metabolic parameters in addition to the six for cybernetic control.To demonstrate the versatility of their model, Alexander and Ramkrishna (1991) showed that it
vi
ri( )expmax
jr j( )exp[ ]
------------------------------------ i 1,2 j 1,2.=;=;=

284 P.R. Patnaik / Biotechnology Advances 18 (2000) 267–288
could express adequately not only cell mass concentration but also the intracellular sid-eropore (expressed as DHBS) and iron concentrations as well as the iron in the medium overa 250-fold range in the initial iron concentration in batch cultures. Similar agreement with ex-perimental values was shown for continuous cultures for dilution rates between 0.1 and 0.8h21. An interesting result was a decrease in the washout dilution rate as the concentration of ironin the medium was reduced; however, no explanation was provided.
By going beyond the cell mass concentration, Alexander and Ramkrishna (1991) haveshown the possibility of applying a cybernetic model for intracellular variables. However,three difficulties remain. One is the large number of parameters, that other workers havepointed out as well (Yoo and Kim, 1994; Doshi et al., 1997). The second difficulty is that themodel has not been tested under disturbed conditions (Turner et al., 1988; Baloo andRamkrishna, 1991a,b). Third, given the first difficulty and the numerical convergence prob-lem elucidated by Yoo and Kim (1994), it is desirable to compare the cybernetic approachwith classical methods, as was done by Yoo and Kim (1994).
8. Concluding remarks
Cybernetic modeling was motivated by the need to explain in a simple and physiologicallysound manner the different types of behaviors shown by microbial cultures in real environ-ments. As discussed in this review, there has been notable success but significant limitationsremain. Application to a real environment has not been demonstrated satisfactorily. Manygrowth media used on a practically useful scale (rather than in small laboratory scale vessels)are either natural substrates of poorly defined compositions (e.g. bovine serum albumin) orare mixtures of well-defined substrates with complex additives (e.g. yeast extract). It is notclear how growth in such media can be modeled cybernetically. The behavior of microorgan-isms in complex media can be very sensitive to the operating conditions, as illustrated by thefermentative production of glyceraldehyde-3-phosphate dehydrogenase by E. coli C600 galK containing the plasmid pBR Eco gap (Nancib et al., 1993; Patnaik, 1996). Cybernetic mod-eling has not yet addressed this problem. Indeed, the presence of process noise becomes soubiquitous as one moves up the volumetric scale of cultivation that it is pertinent to askwhether microbes can always respond fast enough to the fluctuations created by the distur-bances and how a cybernetic approach can be developed to explain these responses. Recentwork has shown that inflow disturbances can radically change the concentration profiles[such as the generation of chaotic oscillations (Patnaik, 1994a)] and that the sensitivities varysignificantly with time and among the variables at a given time (Patnaik, 1994b).
Dhurjati et al. (1985) and Turner and Ramkrishna (1988) have referred to mutationalchanges and the distinction between genotypic and phenotypic events. These variations areparticularly significant if cellular composition changes during the fermentation, if this has aneffect on productivity and if the product is expensive. Recombinant fermentations obviouslysatisfy these criteria but so do long-duration fermentations such as for antibiotics (Vandamme,1984). In such cases, intracellular variables and the presence of cells differing in compositionand morphology are important. In recombinant fermentations, for instance, the concentrationsof plasmid-containing and plasmid-free cells and of the intracellular rDNA and proteolytic

P.R. Patnaik / Biotechnology Advances 18 (2000) 267–288 285
enzymes vary with time and determine both growth kinetics and protein synthesis and stability(Zhang et al., 1996). Here, maximizing the growth rate may not be beneficial because it reducesplasmid stability (Ryu et al., 1991; Summers, 1991). Maximizing recombinant protein synthesis,on the other hand, increases the sensitivity of the culture to disturbances (Patnaik, 1996). Thiskind of problem requires a complex objective function; only two studies (Alexander andRamkrishna, 1991; Yoo and Kim, 1994) have predicted intracellular concentrations while athird (Doshi et al., 1997) ignored it in a recombinant cultivation.
The cultivation of Penicillium sp. is a classic example of more than one kind of cellsgrowing at the same time in the broth. Kluge et al. (1992) postulated three kinds of cells, ac-tive, inactive and lysed, and constructed a model (not cybernetic) which accounted for a shiftfrom nitrogen-limitation to carbon-limitation upon moving from the initial growth phase tothe later production phase, in which there is little additional growth. Three kinds of cells alsooccur in Jarbezski’s (1992) description of ethanol synthesis by Zymomonas mobilis. Thismay not be a coincidence because three variables are the minimum necessary and sufficientto describe complex dynamics (Nicolis and Prigogine, 1989).
The problem of too many parameters has been pointed but by Venkatesh et al. (1997) andacknowledged by Straight and Ramkrishna (1991). The latter’s model had more than 40 pa-rameters, of which 13 were cybernetic. Some limitations of the matching law approach itselfhave been cited by various researchers in justifying cybernetic perspectives different fromthat of Ramkrishna. Doshi et al. (1997) explained their choice of different fractions of thecell mass as the cybernetic variables in preference to the fractional growth rates, which thematching law prescribes. Venkatesh et al. (1997) stated that it is important to include theATP concentration and fueling reactions. While their model was simpler than those formu-lated by Ramkrishna and associates, the suggestion about ATP not having been consideredbefore was not fair because Kompala et al. (1984, 1986) and Straight and Ramkrishna (1991)did acknowledge its role. Yoo and Kim (1994) chose to depart from the matching law be-cause, as they said, the resulting optimization function was very steep and required an ex-tremely small tolerance to ensure convergence. This was a numerical and not conceptual dif-ficulty; Yoo and Kim did not explain the basis of their choice of Eq. (25) as an alternative toEq. (8), except that it circumvented the numerical problem. Nevertheless, the fact that differ-ent ways of formulating cybernetic models are being explored is an indication of the growingpopularity and usefulness of this approach.
Nomenclature
B Biomass in Fig. 2cmel Concentration of melibioseEG Key enzyme for growth-associated processeseG Concentration of EG
Ei Key enzyme for i-th substrateei Concentration of Ei
emel Key enzyme for melibiose degradationemel
max Maximum value of emel

286 P.R. Patnaik / Biotechnology Advances 18 (2000) 267–288
EM Key enzyme for non-growth-associated processeseM Concentration of EM
ER Key enzyme for ribosome synthesis on a single substrateeR Concentration of ER
ER,i Key enzyme for ribosome synthesis on i-th substrateeR,i Concentration of ER,i
Kmel Equilibrium constant for melibiose degradationKRs Monod constant for ribosome synthesis on a single substrateKRs,i Monod constant for ribosome synthesis on i-th substrateKsi Monod constant for growth on i-th substratem Maintenance coefficientrd Rate of enzymatic degradation of melibiose in Eq. (21)rG Rate of growth based on EG
ri, rBi Rate of growth on i-th substraterM Rate of growth based on EM
rP,j Synthesis rate of P on j-th substraterR Rate of ribosome synthesis on a single substraterR,j Rate of ribosome synthesis on j-th substraterx,j Rate of biomass formation on j-th substrates Concentration of a single substrate Ssi, Si Concentration of i-th substrate
Ga Biotic galactose in Fig. 2Gl Biotic glucose in Fig. 2
SL Abiotic lactose in Fig. 2L Biotic lactose in Fig. 2P Biotic key energy source in Fig. 2X Byproduct in Fig. 2
t TimeuC,l Cybernetic variable regulating resource allocation for biomass synthesisuG,j Cybernetic variable regulating resource allocation for growth on glucose (j 5 1) or galac-
tose (j 5 2)uP,l Cybernetic variable regulating resource allocation for synthesis of P
uTA,l Cybernetic variable regulating resource allocation for active transport of SL
uTS,l Cybernetic variable regulating resource allocation for slip transport of SL
X Cell mass concentrationysi yield of cell mass per unit mass of i-th substrate
Greek letters
aR Rate constant for ribosome synthesis on a single substrateaR,i Rate constant for ribosome synthesis on i-th substrateb Degradation constant for the key enzyme Ei
m otal specific growth ratemmi Maximum specific growth rate on i-th substrate
S~
S~
S~
S~
S~
S~
S~

P.R. Patnaik / Biotechnology Advances 18 (2000) 267–288 287
vj Cybernetic variable for glucose (j 5 1) or galactose (j 5 2) in Eq. (21)vE Cybernetic variable activating synthesis of a key enzyme M,P
vG Cybernetic variable for EG
vG,j Cybernetic variable for activation of growth on glucose (j 5 1) or galactose (j 5 2)vi Cybernetic variable for i-th substratevM Cybernetic variable for EM
vm Cybernetic variable for melibiose in Eq. (21)vP Cybernetic variable activating synthesis of P
vP,j Cybernetic variable for rP,j
vR Cybernetic variable for rR
vX Cybernetic variable activating synthesis of X
vTA,l Cybernetic variable for activation of active transport of SL
vTS,l Cybernetic variable for activation of slip transport of SL
vTL,l Cybernetic variable for activation of leak transport of SL
vx,j Cybernetic variable for rx,j
References
Alexander ML, Ramkrishna D. Biotechnol Bioeng 1991;38:637–52.Apter MJ. Cybernetics and Development. Oxford: Pergamon Press, 1966.Arbrige M, Chesbro WR. Arch Microbiol 1982;132:338–44.Asenjo JA, Suk JS. Biotechnol Bioeng Symp Ser 1985;15:225–34.Baloo S, Ramkrishna D. Biotechnol Bioeng 1991a;38:1337–52.Baloo S, Ramkrishna D. Biotechnol Bioeng 1991b;38:1353–63.Bremmermann H. Prog Theor Biol 1967;1:59–68.Demain AL. Adv Biochem Eng 1971;1:113–42.Dhurjati P, Ramkrishna D, Flickinger C, Tsao GT. Biotechnol Bioeng 1985;27:1–9.Dijkhuizen L, van der Werf B, Harder W. Arch Microbiol 1980;124:261–8.Doshi P, Rengaswamy R, Venkatesh KV. Process Biochem 1997;32:643–50.Draper NR, Smith H. Applied Regression Analysis. New York: Wiley, 1981.Egli T, Bossard C, Hamer G. Biotechnol Bioeng 1986;28:1735–41.Esener AA, Roels JA, Kossen NWF. Biotechnol Bioeng 1983;25:2803–41.Gadgil CJ, Bhat PJ, Venkatesh KV. Biotechnol Prog 1996;12:744–50.Gadgil CJ, Venkatesh KV. J Chem Technol Biotechnol 1997;68:89–93.Harder A, Roels JA. Adv Biochem Eng 1982;21:55–107.Herrnstein RJ. J Exp Anal Behav 1974;21:159–64.Holmes PA. Phys Technol 1985;16:32–6.Ingraham JL, Maaloe O, Neidhardt FC. Growth of the Bacterial Cell. Sunderland, MA: Sinauer Associates, 1983.Ishizaki A, Ohta T, Kobayashi G. J Ferment Bioeng 1989;68:123–30.Jarbezski AB. Biotechnol Lett 1992;14:137–42.Kluge M, Siegmund D, Diekmann H, Thoma M. Appl Microbiol Biotechnol 1992;36:446–51.Kompala DS. Bacterial growth on multiple substrates: Experimental verification of cybernetic models. Ph.D. dis-
sertation. West Lafayette, IN: Purdue University, 1984.Kompala DS, Ramkrishna D, Tsao GT. Biotechnol Bioeng 1984;26:1272–81.Kompala DS, Ramkrishna D, Jansen NB, Tsao GT. Biotechnol Bioeng 1986;28:1044–55.Krulwich TA, Ensign JC. J Bacteriol 1969;97:526–31.Lee IH, Fredrickson AG, Tsuchiya HM. Appl Microbiol 1974;28:831–7.Lendenmann U, Snozzi M, Egli T. Appl Environ Microbiol 1996;62:1493–9.
E~
S~
S~

288 P.R. Patnaik / Biotechnology Advances 18 (2000) 267–288
Mandelstam J, McQuillen K. Biochemistry of Bacterial Growth. Oxford: Blackwell, 1968.Monod J. Actualites Scientifique et Industrielles 1942;911:1–21.Moser A. Bioprocess Technology. Kinetics and Reactors. New York: Springer-Verlag, 1988.Mulchandani A, Luong JHT, Groom C. Appl Microbiol Biotechnol 1989;30:11–7.Nancib N, Mosrati R, Boudrant J. Chem Eng J 1993;52:B35–B58.Narang A. The dynamics of microbial growth on mixtures of substrates. Ph.D. dissertation. West Lafayette, IN:
Purdue University, 1994.Nicolis G, Prigogine I. Exploring Complexity. San Francisco, CA: WH Freeman, 1989.O’Connor GM, Sanchez-Riera F, Cooney CL. Biotechnol Bioeng 1992;39:293–304.Oeding V, Schlegel HG. Biochem J 1973;134:239–48.Patnaik PR. Biotechnol Techniques 1994a;8:419–24.Patnaik PR. Appl Math Modelling 1994b;18:620–7.Patnaik PR. Bioprocess Eng 1996;15:215–9.Pavlou S, Fredrickson AG. Biotechnol Bioeng 1989;34:971–89.Pine MJ. Ann Rev Microbiol 1972;26:103–26.Pirt SJ. Principles of Microbe and Cell Cultivation. New York: Wiley, 1975.Pirt SJ. Arch Microbiol 1982;133:300–2.Ramkrishna D. A cybernetic perspective of microbial growth. In: Papoutsakis E, Stephanopoulos GN, Blanch
HW, editors. Foundations of Biochemical Engineering: Kinetics and Thermodynamics in Biological Systems.Washington DC: Am Chem Soc, 1982. pp. 161–78.
Ramkrishna D, Fredrickson AG, Tsuchiya HM. J Gen Microbiol 1966;12:311–9.Raymond KN, Carrano CJ. Accounts Chem Res 1979;12:183–94.Ryu DDY, Kim J-Y, Lee SB. Bioprocess kinetics and modelling of recombinant fermentation. In: Schugerl K, ed-
itor. Biotechnology. Weinheim: VCH, 1991, pp. 485–505.Shah N, Jelen P. J Food Sci 1990;55:506–9.Standing CN, Fredrickson AG, Tschuyia HM. Appl Microbiol 1972;23:354–60.Straight JV, Ramkrishna D. Biotechnol Bioeng 1991;37:895–909.Summers DK. Trends Biotechnol 1991;9:273–8.Tempest DW, Neijssel OM. In: Dean ACR, Ellwood DC, Evans CGT, Melling J, editors. Continuous Culture:
Applications and New Fields. Chichester: Ellis Horwood, 1976. p. 283.Turner BG, Ramkrishna D. Biotechnol Bioeng 1988;31:41–3.Turner BG, Ramkrishna D, Jansen NB. Biotechnol Bioeng 1988;32:46–54.Turner BG, Ramkrishna D, Jansen NB. Biotechnol Bioeng 1989;34:252–61.Vandamme EJ, editor. Biotechnology of Industrial Antibiotics, vol. 22. New York: Dekker, 1984.Venkatesh KV, Doshi P, Rengaswamy R. Biotechnol Bioeng 1997;56:635–44.Villarejo M. Biochem Biophys Res Commun 1980;93:16–23.Yoo S, Kim W-S. Biotechnol Bioeng 1994;43:1043–51.Zhang Z, Moo-Young M, Chisti Y. Biotechnol Adv 1996;14:401–35.














![[Lichtman Stuart] Cybernetic Transposition(BookZZ.org)](https://img.pdfslide.net/doc/110x75/55cf9443550346f57ba0bcf0/lichtman-stuart-cybernetic-transpositionbookzzorg.jpg)














