Embed Size (px)
Citation preview

B Cell Generation, Activation, And Differentiation
W. Robert Fleischmann, Ph.D.
Department of Urologic Surgery
University of Minnesota Medical School
(612) 626-5034

Objectives
• To understand B cell development• To understand the rearrangement of the
antibody variable region during B cell maturation
• To understand B cell activation and the signals involved in the activation process
• To understand the importance of somatic cell mutations
• To understand antibody class switching

B Cell Generation

B Cell Generation
• Progenitor B cells (pro-B cells) express c-Kit which binds to stem cell factor expressed on BM stromal cells, inducing them to proliferate and differentiate to Precursor B cells (pre-B cells).
• Pre-B cells express IL-7R and are stimulated to divide and differentiate.
• Pre-B cells rearrange antibody chain genes first, then chain genes.
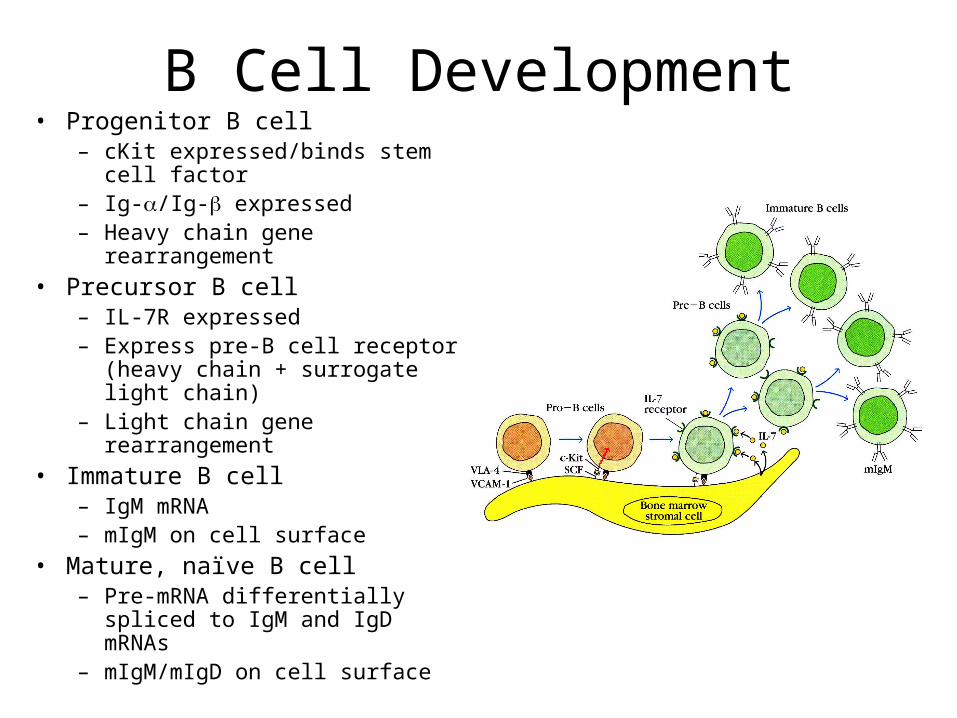
B Cell Development• Progenitor B cell
– cKit expressed/binds stem cell factor
– Ig-/Ig- expressed– Heavy chain gene
rearrangement
• Precursor B cell– IL-7R expressed– Express pre-B cell receptor
(heavy chain + surrogate light chain)
– Light chain gene rearrangement
• Immature B cell– IgM mRNA – mIgM on cell surface
• Mature, naïve B cell– Pre-mRNA differentially spliced
to IgM and IgD mRNAs– mIgM/mIgD on cell surface
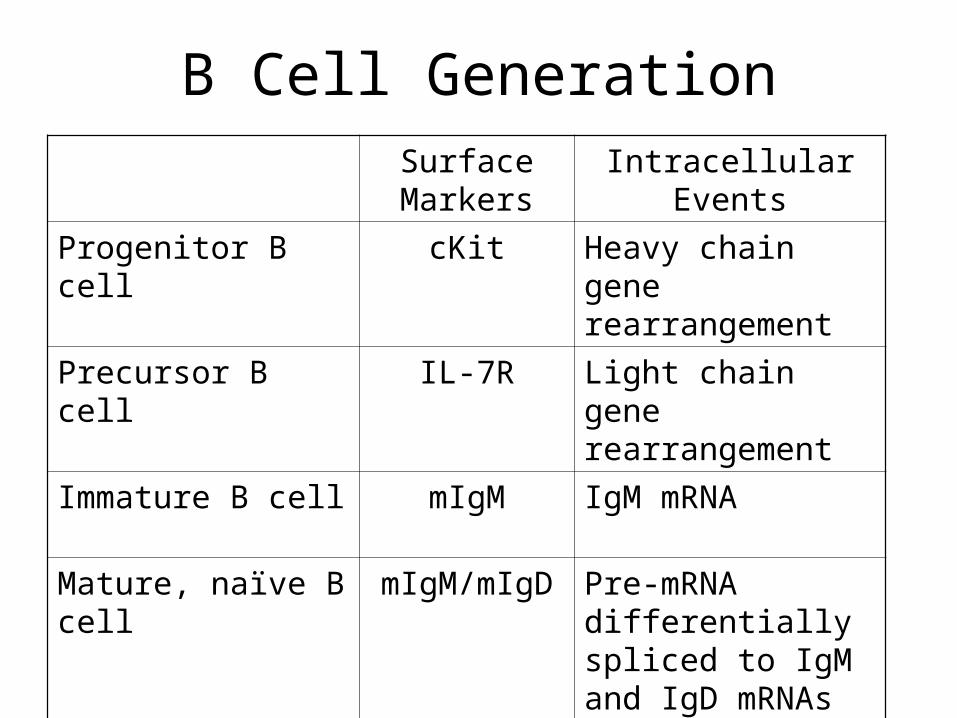
B Cell GenerationSurface Markers
Intracellular Events
Progenitor B cell cKit Heavy chain gene rearrangement
Precursor B cell IL-7R Light chain gene rearrangement
Immature B cell mIgM IgM mRNA
Mature, naïve B cell mIgM/mIgD Pre-mRNA differentially spliced to IgM and IgD mRNAs
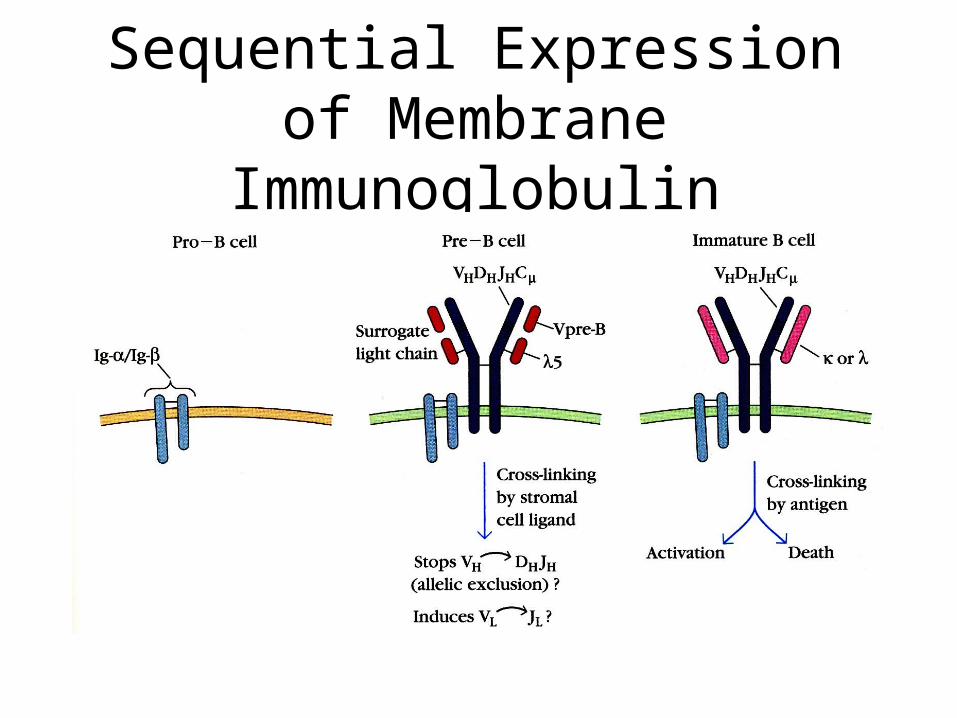
Sequential Expression of Membrane Immunoglobulin
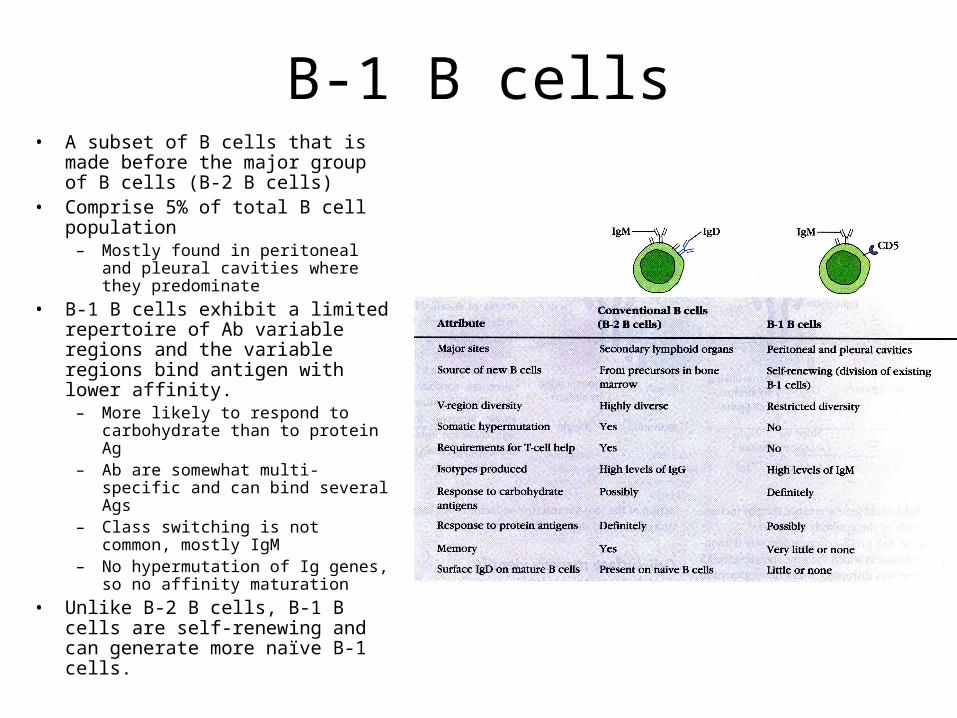
B-1 B cells• A subset of B cells that is made
before the major group of B cells (B-2 B cells)
• Comprise 5% of total B cell population
– Mostly found in peritoneal and pleural cavities where they predominate
• B-1 B cells exhibit a limited repertoire of Ab variable regions and the variable regions bind antigen with lower affinity.
– More likely to respond to carbohydrate than to protein Ag
– Ab are somewhat multi-specific and can bind several Ags
– Class switching is not common, mostly IgM
– No hypermutation of Ig genes, so no affinity maturation
• Unlike B-2 B cells, B-1 B cells are self-renewing and can generate more naïve B-1 cells.

Negative Selection of Self-Reactive B Cells
• Negative selection of many self-reactive B cells occurs in the bone marrow (clonal deletion).– This limits development of antibody-mediated autoimmunity.– If immature B cells, expressing mIgM recognize self-antigen,
• Some of the immature B cells undergo apoptosis.• Some of the immature B cells undergo editing of light chain
genes to produce a different light chain that, when combined with the heavy chain does not recognize self-antigen.
• Negative selection of some self-reactive B cells occurs in the periphery.– Not all self-reactive B cells are eliminated in the bone
marrow because not every self-antigen is expressed in the bone marrow.
– Mechanism not really understood.

B Cell Activation by Antigen

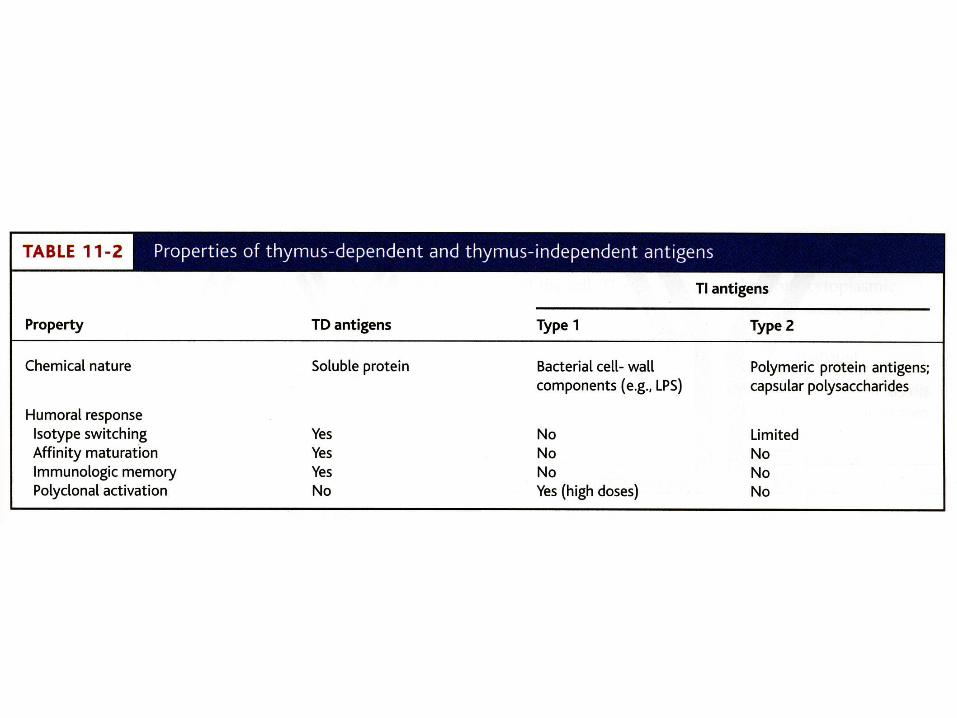
Thymus-Dependent vs. Thymus-Independent Antigens• Thymus-dependent antigens (TD Ags) require
direct contact of B2 cells with Th2 cells to induce an antibody response. – Generally, a stronger response than a thymus-
independent Ag response
• Thymus-independent antigens (TI Ags) induce B1 cells to produce antibodies without the need for Th2 cell activity.– Type 1 TI antigen = lipopolysaccharide and other
bacterial cell wall components– Type 2 TI antigen = highly repetitious molecules
such as polymeric proteins (flagellin) or bacterial cell wall polysaccharides with repeated subunits

B Cell Response to Type 1 TI Antigen
• B1 cells bind lipopolysaccharide via either toll-like receptor-4 (TLR4) or via the B cell receptor (specific antibody + Ig- and Ig-).– TLR4 binding is non-specific, so B cells with many different
antibodies will be activated (polyclonal activation). In this way, it acts as a B cell mitogen.
– Antibody binding is specific, so only B cells with specificity for the type of LPS will be activated.
• Type 1 TI antigen can activate both immature and mature B cells because it doesn’t require a surface antibody molecule (can activate through TLR4).
• Only IgM is produced.

B Cell Response to Type 2 TI Antigen
• B1 cells bind Type 2 TI antigen by crosslinking of the B cell receptor (specific antibody + Ig- and Ig-).– Antibody binding is specific, so only B1 cells with
specificity for the type of antigen will be activated.– Only mature (not immature B1 cells) will be
stimulated.
• Mostly IgM is produced.• While Th2 activity is not required, cytokine
production by Th2 cells are needed for full B cell response and for class switching to isotypes other than IgM.
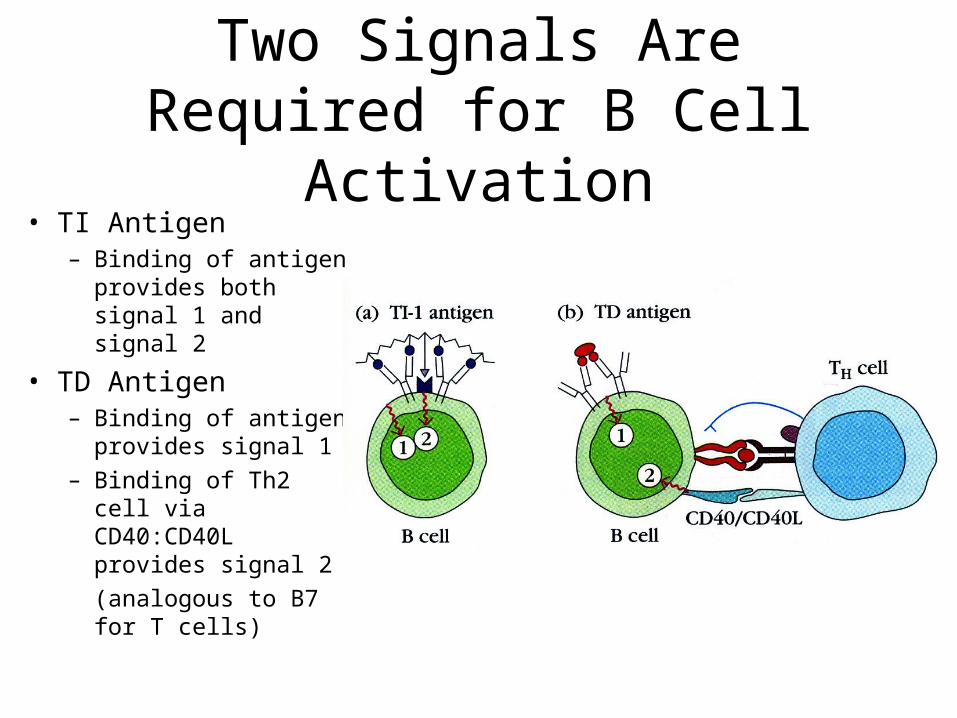
Two Signals Are Required for B Cell Activation
• TI Antigen– Binding of antigen
provides both signal 1 and signal 2
• TD Antigen – Binding of antigen
provides signal 1– Binding of Th2 cell
via CD40:CD40L provides signal 2
(analogous to B7 for T cells)

Role of Ig-/Ig- in Signaling
• mIgM and mIgD have short cytoplasmic tails that are too short to transduce a transmembrane signal.
• Ig-/Ig- has an ITAM motif (immunoreceptor tyrosine-based activation motif) on the cytoplasmic tails.
• The tyrosine based activation motif is activated by antigen cross-linking antibody on the surface of the B cell.
• This initiates a cascade of events.
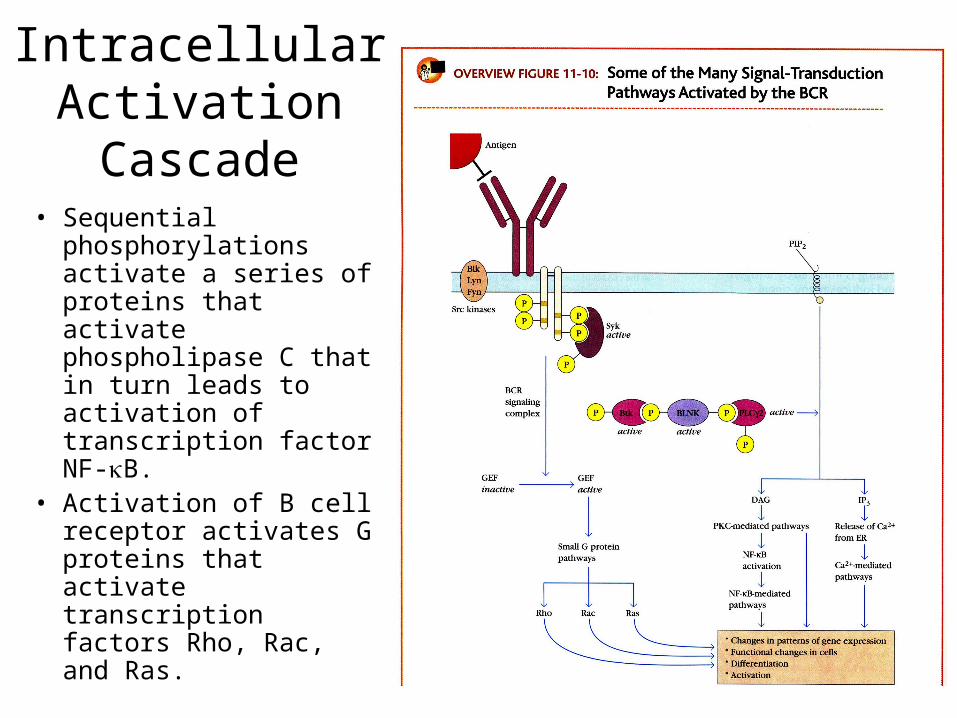
Intracellular Activation Cascade
• Sequential phosphorylations activate a series of proteins that activate phospholipase C that in turn leads to activation of transcription factor NF-B.
• Activation of B cell receptor activates G proteins that activate transcription factors Rho, Rac, and Ras.
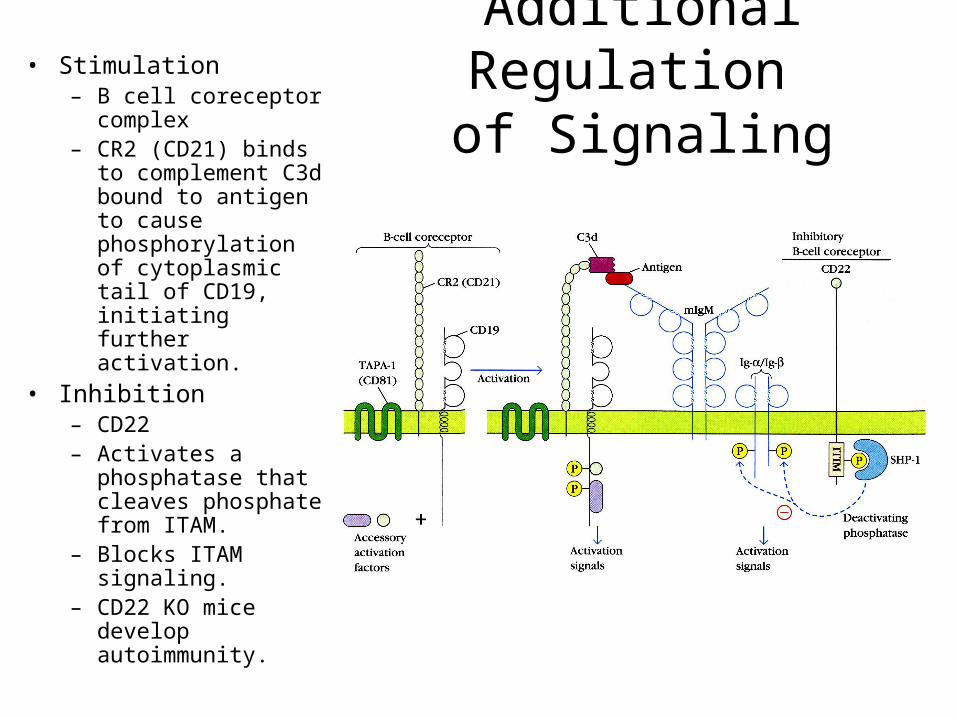
Additional Regulation of Signaling
• Stimulation– B cell coreceptor
complex– CR2 (CD21) binds to
complement C3d bound to antigen to cause phosphorylation of cytoplasmic tail of CD19, initiating further activation.
• Inhibition– CD22– Activates a
phosphatase that cleaves phosphate from ITAM.
– Blocks ITAM signaling.
– CD22 KO mice develop autoimmunity.

B Cell Activation by T Cell

B Cell Activation by T Cell
• The activation of B cells by antigen binding to receptor and by coreceptor binding creates conditions for proliferation and differentiation of B cells.
• However, important T cell interactions and T cell produced cytokines are necessary to complete the activation of B cells for thymus-dependent antigens.
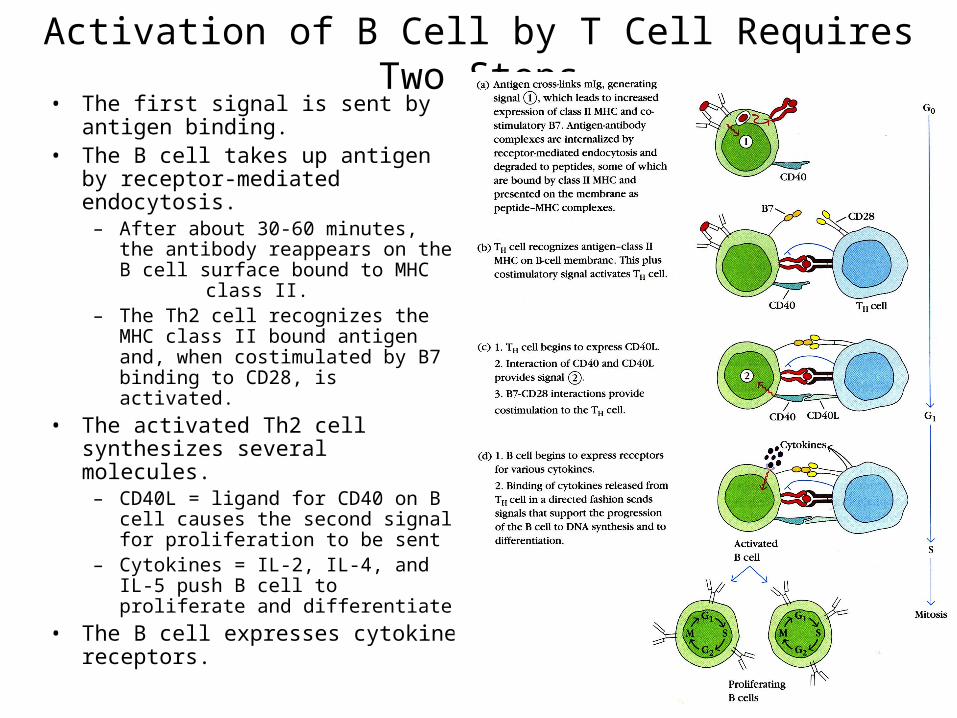
Activation of B Cell by T Cell Requires Two Steps• The first signal is sent by antigen
binding.• The B cell takes up antigen by
receptor-mediated endocytosis.– After about 30-60 minutes, the
antibody reappears on the B cell surface bound to MHC class II.
– The Th2 cell recognizes the MHC class II bound antigen and, when costimulated by B7 binding to CD28, is activated.
• The activated Th2 cell synthesizes several molecules.
– CD40L = ligand for CD40 on B cell causes the second signal for proliferation to be sent
– Cytokines = IL-2, IL-4, and IL-5 push B cell to proliferate and differentiate
• The B cell expresses cytokine receptors.
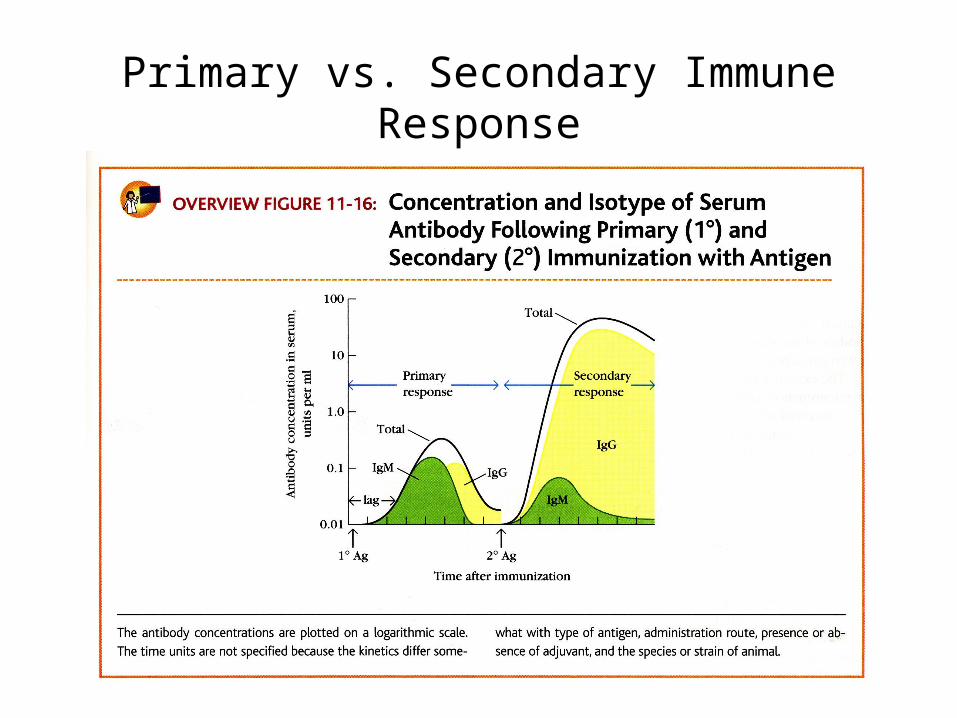
Primary vs. Secondary Immune Response
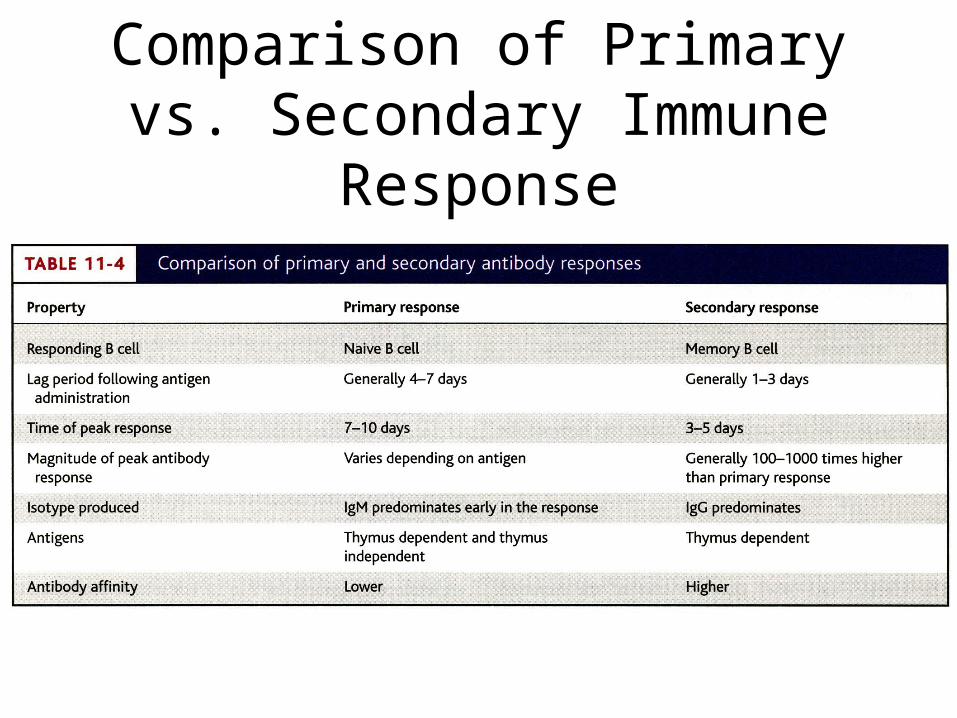
Comparison of Primary vs. Secondary Immune Response
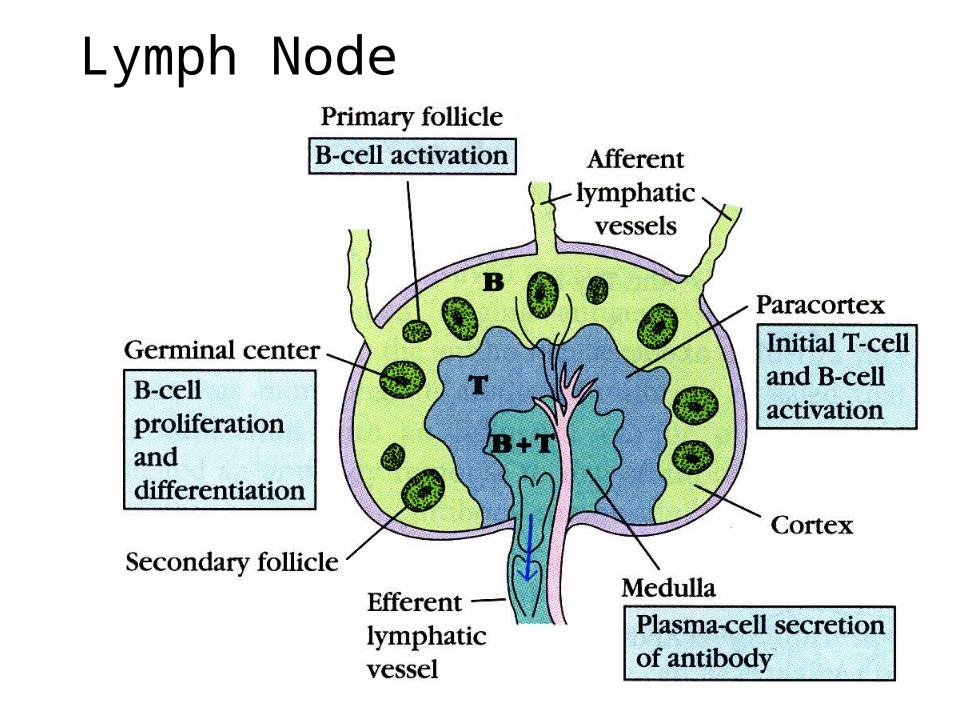
Lymph Node

Germinal Centers• Antigen flows through the lymph to
the lymph node either as free antigen or as antigen bound to an antigen-presenting cell.
• Free antigen binds to follicular dendritic cells in follicles and germinal centers, macrophages throughout the lymph node, dendritic cells in the paracortex, or B cells in the follicles and germinal centers.
• Processed antigen is presented to T cells and to B cells.
• T and B cells recognize each other at the periphery of the medullary region, bind, and move to the germinal centers.
• In the germinal centers, the B cells known as centrocytes undergo affinity maturation that is mediated by somatic cell mutation.
• In the germinal centers, there is class switching.
• The B cells become plasma cells and exit the germinal centers.
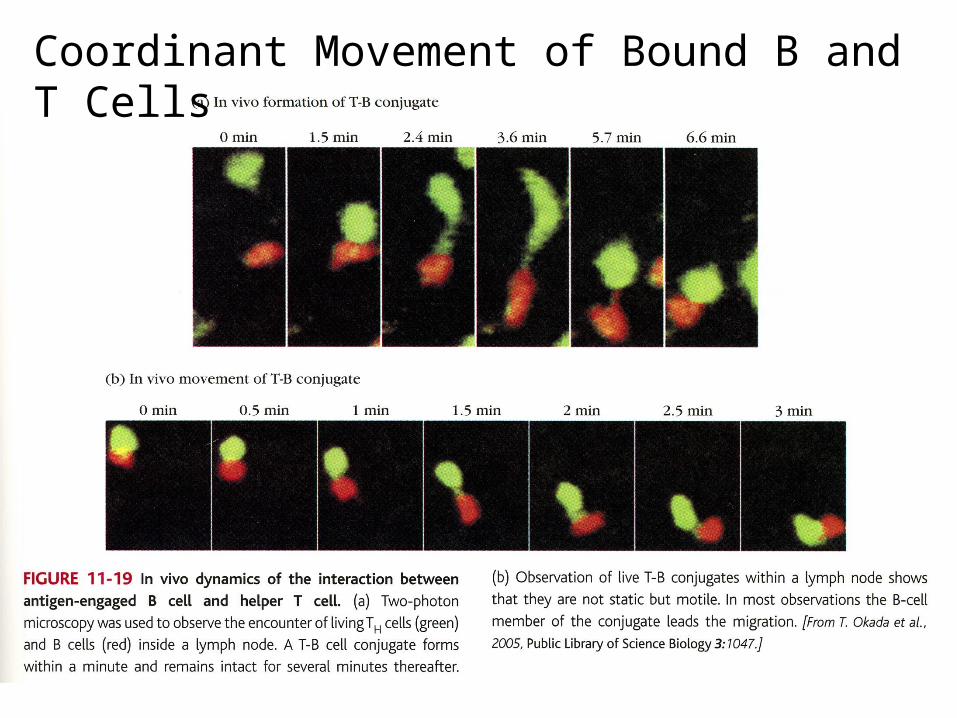
Coordinant Movement of Bound B and T Cells

Importance of Somatic Cell Mutations• Somatic hypermutation occurs within the VDJ
region with each cell division an activated B cell.
• The hypermutations give rise to a slightly different antibody molecule that may or may not have greater affinity for the antigen (affinity maturation).– If less affinity, then antibody will not be able to
remain bound to antigen and apoptosis will eliminate the B cell from the population.
– If greater affinity, the stronger interaction of the antigen for the antibody drives proliferation, until a clone of B cells with the greatest antigen affinity is founded.

Class Switching• An mIgM-bearing cell can produce secreted IgM or it can
undergo class switching by recombining the DNA at class switch recombination sites.
• Class switching exchanges the constant region of the antibody molecule . – IgM becomes IgG– IgG becomes IgE – IgG becomes IgA
• Class switching is driven by B/T cell interactions via the CD40/CD40L interaction and by specific cytokines produced by the T cell.
• Class switching occurs in the germinal centers of lymph nodes.• Individuals whose T cells lack CD40L cannot undergo class
switch and express a condition called X-linked hyper-IgM syndrome.– IgM only– No memory cell generation
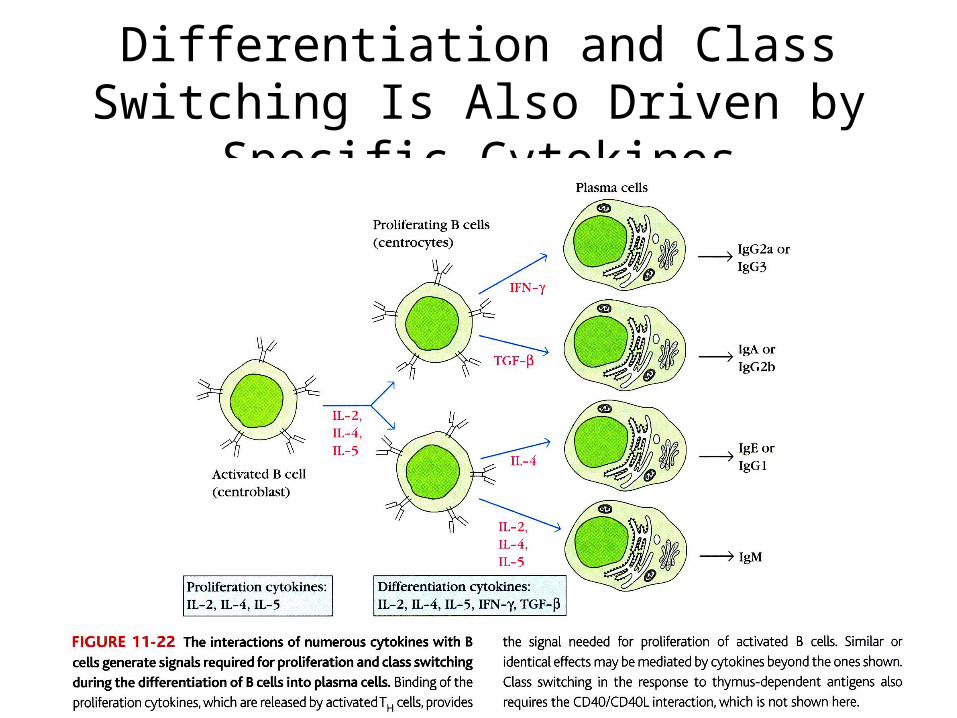
Differentiation and Class Switching Is Also Driven by Specific Cytokines

Immunoglobulin Production Down-Regulates the Immune Response
• As antibody is produced, it exerts a feedback inhibition of its own production.
• This occurs by two mechanisms– Soluble antibody binds antigen and prevents the antigen from
reaching the mIgM/mIgD antibody on the surface of other naïve B cells.
– Antibody/antigen complexes bind to Fc receptor molecules on the surface of the B cells making it ever more likely that the down-regulatory events mediated by CD22 will be triggered.
• There are few molecules of CD22 on a cell and many molecules of antibody.
• When few antibodies are bound to antigen, it is difficult for the antigen/antibody complex and CD22 to find each other to bind.
• When many antibodies are bound to antigen, the antigen/antibody complex and CD22 find each other easily.






























