Embed Size (px)
Citation preview

DEVELO
PMENT
1DEVELOPMENT AND DISEASE RESEARCH ARTICLE
INTRODUCTIONMaternal effect, which can refer to the dependence of earlyembryonic development on maternal products, is common in bothanimal and plant kingdoms. This dependence is on maternal proteinsand mRNAs that are recruited during oocyte maturation or followingfertilization. The dependence on maternal mRNA is because thefully grown oocyte and the one-cell embryo are essentiallytranscriptionally silent but translationally active, and the maternal-to-zygotic transition occurs during the late one-cell/early two-cellstages (Ko et al., 2000; Schultz, 2002; Wang et al., 2004).Mammalian maternal-effect genes have only been identifiedrecently, largely by gene-knockout approaches [e.g. Mater (Nalp5 –Mouse Genome Informatics), Hsf1, Zar1, Npm2 and Stella (Dppa3– Mouse Genome Informatics)]. Unlike their counterparts ininvertebrates and lower vertebrates, the molecular mechanisms ofmammalian maternal-effect genes have not been well understood.In the strictest sense, mutations in maternal-effect genes should notaffect oocyte development, maturation and ovulation, but shouldimpair embryo development. Indeed, null mutations of themammalian maternal-effect genes identified so far have little effecton ovulation and fertilization of the affected eggs, and interfere onlywith early embryo development (Tong et al., 2000; Christians et al.,2000; Wu et al., 2003; Burns et al., 2003; Payer et al., 2003). Hence,genes expressed in oocytes can be classified as required foroogenesis (i.e. oocyte development, maturation, ovulation andfertilization) and for embryonic development (i.e. maternal effect)(Dean, 2002).
A hallmark of oocyte development is the synthesis andaccumulation of macromolecules and organelles that constitute thematernal capital used to support early development prior toactivation of the embryonic genome (Schultz, 1993). In mammals,this maternal store includes proteins and RNAs, which are involved
in a wide range of processes, as evidenced by the variety ofimplicated functions of the currently identified maternal-effectgenes (Tong et al., 2000; Christians et al., 2000; Wu et al., 2003;Burns et al., 2003; Payer et al., 2003). Furthermore, it is still notclear how mammalian oocytes monitor and respond to a deficiencyin maternal-effect genes, even if malfunction of these genes doesnot directly affect oocyte maturation itself.
Basonuclin (Bnc1 – Mouse Genome Informatics) is a zinc-fingerprotein with a very restricted tissue distribution; it is mainly foundin keratinocytes of the stratified epithelium and germ cells (i.e. malegerm cells and oocytes) (Yang et al., 1997; Mahoney et al., 1998).There is highly suggestive evidence that basonuclin regulates rRNAtranscription (Iuchi and Green, 1999; Tseng et al., 1999). Forexample, the zinc fingers of basonuclin interact with threeevolutionarily conserved binding sites in the rDNA promoter (Tianet al., 2001). In mouse oocytes, basonuclin co-localizes with RNApolymerase I (Pol I) activity in the nucleolus, and a dominant-negative mutant of basonuclin interferes with Pol I transcription inoocytes (Tian et al., 2001). Basonuclin, however, differs from thededicated Pol I transcription factors (e.g. UBF), in that it is alsoabundant in nucleoplasm and can interact with Pol II promoters(Tseng et al., 1999). Such observations led to the proposal thatbasonuclin may have target genes other than rDNA (Tian et al.,2001). Together, these results led us to propose that basonuclin isnecessary for oocyte rRNA synthesis and possibly other processes,and perturbing basonuclin function in the oocyte will disruptoogenesis, leading to infertility. To test this hypothesis, we employeda recently developed transgenic-RNAi approach (Svoboda et al.,2000; Stein et al., 2003) to inhibit basonuclin function specificallyin mouse oocytes.
We report here that transgenic-RNAi effectively and specificallyreduces basonuclin level in mouse oocytes. The transgenic mice areapparently normal, except females are subfertile. Basonuclin-deficient oocytes display a reduced rate of PoI transcriptionand perturbation of a large number of Pol II-transcribed genes.Many basonuclin-deficient oocytes appear morphologically andbiochemically abnormal, but their maturation and ovulation proceedwithout apparent problems. Although normal numbers of eggs areovulated and inseminated, pre-implantation development is severelycompromised. Thus, basonuclin joins the short, but growing, list ofmammalian maternal-effect genes; among them, it is the first to alsoperturb oogenesis.
Basonuclin: a novel mammalian maternal-effect geneJun Ma1,2, Fanyi Zeng3, Richard M. Schultz2,4 and Hung Tseng1,4,5,*
Basonuclin is a zinc-finger protein found in abundance in oocytes. It qualifies as a maternal-effect gene because the source of pre-implantation embryonic basonuclin is maternal. Using a transgenic-RNAi approach, we knocked down basonuclin specifically inmouse oocytes, which led to female sub-fertility. Basonuclin deficiency in oocytes perturbed both RNA polymerase I- and II-mediated transcription, and oocyte morphology was affected (as evidenced by cytoplasmic and cell surface abnormalities). Some ofthe affected oocytes, however, could still mature to and arrest at metaphase II, and be ovulated. Nevertheless, fertilized basonuclin-deficient eggs failed to develop beyond the two-cell stage, and this pre-implantation failure accounted for the sub-fertilityphenotype. These results suggest that basonuclin is a new member of the mammalian maternal-effect genes and, interestingly,differs from the previously reported mammalian maternal-effect genes in that it also apparently perturbs oogenesis.
KEY WORDS: Female fertility, Oogenesis, Pre-implantation development, Transcription regulation, Transgenic RNAi
Development 133, 0000-0000 (2006) doi:10.1242/dev.02371
1Department of Dermatology, University of Pennsylvania, PA 19104, USA.2Department of Biology, University of Pennsylvania, PA 19104, USA. 3ShanghaiInstitute of Medical Genetics, Shanghai Jiao Tong University, Shanghai 200040,People’s Republic of China. 4Center for Research on Reproduction and Women’sHealth, University of Pennsylvania, PA 19104, USA. 5Department of Cell andDevelopmental Biology, University of Pennsylvania, PA 19104, USA.
*Author for correspondence (e-mail: [email protected])
Accepted 17 March 2006
Development ePress online publication date 19 April 2006http://dev.biologists.org/lookup/doi/10.1242/dev.02371Access the most recent version at First posted online on 19 April 2006 as 10.1242/dev.02371

DEVELO
PMENT
2
MATERIALS AND METHODSRNAi transgene constructTwo transgenic constructs differing in their vector backbones and sequenceof the basonuclin inverted repeats were employed in this study (see alsoResults). The Zp3 promoter directs oocyte-specific expression of thebasonuclin double-stranded (ds) RNA. To prepare the inverted repeat forgenerating the dsRNA hairpin, basonuclin-coding sequence was amplifiedby PCR and then ligated 3� to 3�. The inverted repeat was transferred topRNAi-ZP3 cassette (Stein et al., 2003), to yield pB1-rnai-zp3-1 or andpB1rani-zp3-2.
Generating transgenic miceThe transgene construct was used for microinjection at the Transgenic andChimeric Mouse Facility at University of Pennsylvania and TransgenicAnimal Model Core at University of Michigan with the approval of theInstitutional Animal Care and Use Committee at the respective universities.The zygotes for microinjection were produced by mating of B6SJLF1/Jmice. Transgenic out-crossed (TG-OC) mice were obtained by matingtransgenic founder males (B6SJLF1/J) with wild-type females (CF1), andgenotyped by PCR using the EGFP primer pair (5�-CGACGTA-AACGGCCACAAGT and 5�-CTCGTTGGGGTCTTTGCTCA) with 34cycles at 55°C for 20 seconds, 72°C for 40 seconds and 94°C for 20 seconds.
FertilityEach founder transgenic female (B6SJLF1/J) was mated with the same threestud males (B6SJLF1/J) in a random sequence. The same males were alsoused to assess the fertility of TG-OC females. Successful mating was verifiedby detecting a vaginal plug; the mated females were then housed separatelyfor observation and recording the number of pups delivered per litter, whichwas averaged to assess fertility. Other than the fertility tests, all furtherstudies were carried out with TG-OC females.
Collection and culture of oocytes and embryosGV, MII eggs and fertilized eggs were collected as previously described(Schultz et al., 1983). GV oocytes were cultured in CZB medium (Chatot etal., 1989) containing 0.2 mM IBMX (Sigma) to inhibit GV breakdown; MIIeggs were cultured in CZB medium and fertilized eggs cultured in KSOMmedium (Erbach et al., 1994) and their development was recorded at 36, 60and 84 hours post-coitum (hpc).
RNA isolation and real-time PCRTotal RNA was isolated from 20 oocytes and reverse transcribed withSuperscript II reverse transcriptase (Invitrogen) using oligo dT as primer.The resulting cDNA was quantified by real-time PCR using an ABI Prism7000 thermocycler (Applied Biosystems). The primers for basonuclin were:5�-ACTGGACGCTTCAGGATTACATC and 5�-GTCATGATGCTCCAG-TGATCCA. PCR conditions were 40 cycles at 95°C for 15 seconds and60°C for 60 seconds. UBF (internal standard), was amplified with primersprovided by ABI (assay ID: Mm00456972_m1). Each sample was analyzedin duplicate. The Ct values were converted to percentage of the wild-type(non-transgenic) level.
Run-on assay and quantification of the transcription lociRun-on assays were conducted on meiotically incompetent oocytes obtainedfrom 13- to 14-day-old mice as previously described (Tian et al., 2001).Briefly, oocytes were permeabilized and then incubated at 37°C for 1 hourwith 0.2 mM Br-UTP and 100 �g/ml �-amanitin. Oocytes were then fixedin 2% paraformaldehyde and the incorporated BrdU was detected withAlexa Fluor 488-conjugated anti-BrdU monoclonal antibody (MolecularProbes) at 1:20 dilution. The amount of incorporated BrdU was quantifiedby measuring the overall fluorescence intensity at the largest cross-sectionof the nucleolus (recorded on an epi-fluorescent digital micrograph) with the‘measurement’ function of the ImageJ, a software package created by W.Rasband (http://rsb.info.nih.gov/ij/). This value, in arbitrary units, wassubtracted by the background fluorescence of an area equal in size in thenucleoplasm, which was not labeled in the presence of �-amanitin. Thenumber of labeled transcription foci was scored using microphotographs,which represented a stack of selected optical planes of the nucleolus. Themicrophotograph stacks were produced with a Zeiss Axiovert 200 using a
100� objective under epifluorescence and by manual focusing. Theinformative optical planes containing unique transcription foci were selectedmanually and photographed. Scoring transcription foci in a nucleolusrequired usually 8 to 12 microphotographs. The foci that had been scored inthe previous plane were marked on photos to avoid repetition. This methodalso ensured that no informative optical plane was missed because the out-of-focus foci image of the previous plane could be seen.
RNase protection assayRNase protection assays were conducted to quantify rRNA in oocytes. [32P]-labeled antisense probe for rRNA was prepared using the Maxiscript T7 kitand pTRI-RNA-28S/pTRI-RNA-18S antisense control templates (Ambion).Fifty oocytes were transferred to the lysis buffer of the Direct Protect RPAkit (Ambion). The samples were then incubated with the probe, treated withnuclease and electrophoresed according to the manufacturer’s protocol.Protected fragments were detected by autoradiography and quantified bydensity measurement.
RNA isolation and microarray analysisTotal RNA was extracted from oocyte samples, each originated from one ofthe six TG-OC (T#50) and four wild-type eCG-stimulated mice, andsubjected to cRNA preparation (Pan et al., 2005; Zeng et al., 2004). Thecontrol 1, 3, 4 and transgenic 4, 5, 6 mice were from one litter, and the restfrom another. Each Affymetrix MOE 430 v2 GeneChip was probed with 15�g cRNA and processed according to the Affymetrix instructions. Rawmicroarray data (*.cel) (available at the Gene Expression OmnibusRepository, Accession Number GSE4029) were processed using the GC-RMA or MAS-5 algorithms to obtain the normalized expressionsummarization for each probe set. The expression level in our data set rangedbetween 1 and 66,582. Genes with an expression level of less than 10 wereexcluded before GeneSpring (Silicon Genetics) K-mean hierarchicalclustering and other filter analysis. Genes whose expression wassignificantly changed in the transgenic oocytes compared with the controlwere identified by Significance Analysis of Microarray (v1.21, two-class,unpaired analysis with the permutation parameter set at 500, FDR<1%) orby one-way ANOVA analysis (FDR <5%) and annotated in the GeneSpring.The statistically significant gene lists obtained by ANOVA were importedinto EASE v2.0 (Hosack et al., 2003) to test for overrepresentation ofbiological processes in transgenic versus wild-type samples. An EASE scorewas calculated for the likelihood of overrepresentation in the Gene OntologyConsortium annotation categories (i.e. biological process, cell component,molecular function), as well as KEGG pathways and SwissProt keywords.
Immunofluorescence staining and immunoblottingOocytes/embryos were fixed in 2.5% paraformaldehyde/PBS andimmunocytochemistry was conducted as previously described (Yu et al.,2004) using an affinity-purified anti-basonuclin antibody (MBP-1023) dilutedat 1:30. Affinity-purified pre-immune serum was used as a negative control.
For immunoblotting, 75 oocytes/embryos were lysed in 2� SDS-PAGEbuffer, run on a 7.5% SDS-PAGE and transferred to PVDF membrane(Hybond-P, Amersham Biosciences). Immunodetection was performedusing horseradish peroxidase-conjugated secondary antibodies and ECLAdvance reagents (Amersham) according to the manufacturer’s instructions.
DNA replication assayFertilized eggs were collected at 10 hpc and labeled with 10 �M BrdU for 6hours. The embryos were then fixed with paraformaldehyde, treated with 2N HCl at 37°C for 1 hour, and then neutralized in 0.1 M borate buffer (pH8.5), for 15 minutes. Incorporated BrdU was visualized with 1:30 dilutedAlexa fluor 488-conjugated anti-BrdU antibody (Invitrogen).
HistologyFor histological analysis, adult ovaries were fixed in Bouin’s fixative for 16hours before transfer to 70% ethanol. Samples were then embedded inparaffin, serially sectioned (5 �m) and stained with Hematoxylin and Eosin.
TUNEL labeling assayTUNEL (TdT-mediated dUTP nick end labeling) assays were carried outwith the In Situ Cell Death Detection Kit (Roche Diagnostics) according tothe manufacturer’s instructions.
RESEARCH ARTICLE Development 133 (10)

DEVELO
PMENT
Protein synthesis assay and transcription-requiring complex (TRC)quantificationOocytes were metabolically radiolabeled with [35S]-methionine and acid-insoluble radioactivity was then determined following TCA precipitation aspreviously described (Conover et al., 1991). Quantification of thetranscription-requiring complex (TRC) was also conducted as previouslydescribed (Conover et al., 1991).
RESULTSExpression profile of basonuclin during oogenesisand pre-implantation developmentImmunostaining showed that basonuclin was not detected in eight-cell embryos (Q. J. Tian and H.T., unpublished). Consistent withthis finding, a recent microarray study indicated that basonuclinmRNA was present in oocytes and one-cell embryos but its levelrapidly decreased by over 30-fold during pre-implantationdevelopment; by the eight-cell stage, it was at background levels(Zeng et al., 2004). We first confirmed this basonuclin mRNAexpression profile by RT-PCR (Fig. 1, broken line) and thenestablished the expression profile for basonuclin protein in oocytesand pre-implantation embryos by immunoblotting andimmunocytochemistry. Immunoblotting revealed an apparentdecline in basonuclin level from the immature oocyte (GV-stage) tothe mature metaphase II (MII) stages (Fig. 1). Similar amounts ofbasonuclin were present in MII eggs and one-cell embryos, but nobasonuclin was detected in two-cell and morulae/blastocysts, whichis consistent with the disappearance of its mRNA during pre-implantation development. Immunocytochemical analysis alsorevealed a similar and progressive decrease of immunoreactivematerial from oocytes to pre-implantation embryos (Fig. 1). Thus,
maternal basonuclin is not replaced by zygotic basonuclin duringpre-implantation development and therefore could have a maternaleffect.
Generating basonuclin-RNAi transgenic miceThe presence of basonuclin in oocytes and early embryos suggestsit performs: (1) the same function for both oocytes and embryos; (2)unrelated functions in oocyte maturation and/or in early embryos; or(3) an oocyte function, which is required by the embryos (maternaleffect). To resolve these possibilities, we employed a recentlydeveloped transgenic RNAi approach to inhibit basonuclin functionin oocytes (Svoboda et al., 2001). We constructed two types of RNAitransgene plasmids, pB1rnai-zp3-1 and pB1rani-zp3-2 (Fig. 2A); thedetails of generating transgenic mice are described in the Materialsand methods. Twenty-four founder mice (13 females and 11 males)were obtained, of which 14 were generated by pB1rnai-zp3-1 and10 by pB1rnai-zp3-2.
Transgenic basonuclin-RNAi females are sub-fertileWe analyzed the fertility of both founder transgenic females andfemale progeny of the transgenic founder males (Fig. 2B). To reducethe influence of male fertility on the outcome, three normal maleswere mated in a randomized sequence with each female being tested.Eight of the nine transgenic founder females showed a wide rangeof sub-fertility – from moderate to severe (infertile) – whencompared with the average of the seven non-transgenic littermatesin the control group (Fig. 2B). The sub-fertility was unlikely to bedue to the vector backbone and/or transgene integration sitesbecause the founder females were derived from multiple injectionsof the two transgene-constructs. Southern analysis also failed toestablish any correlation between the integration sites and transgenecopy number with the observed reduced fertility (data not shown).
The fertility of transgenic female progeny of the male foundersmirrored that of the transgenic female founders (Fig. 2C). Other thanfemale sub-fertility, the transgenic male and female mice appearednormal. Because transgenic females were sub-fertile, we usedtransgenic males to transmit the transgene and focused subsequentstudies on the female progeny of two transgenic lines, T50 and T8,which had ~10% and 50% normal fertility, respectively. To date, thefemale sub-fertility phenotype of these lines has been stable aftertwo generations of breeding.
Transgenic oocytes contain reduced amounts ofbasonuclin mRNA and proteinWe examined fully grown transgenic oocytes for basonuclin mRNAand protein to determine the cumulative effect of RNAi during thegrowth phase. Real-time PCR demonstrated that the amount ofbasonuclin mRNA in transgenic oocytes from both lines wasdramatically reduced. For T8 and T50 oocytes, respectively, anadditional 4 and 6 cycles were required to reach threshold, whencompared with non-transgenic counterparts (Fig. 2D). This increasein cycle number translates into a 94% and 98% reduction inbasonuclin mRNA. Indeed, immunocytochemical (Fig. 2F) andwestern blot (Fig. 2G) examinations confirmed the dramaticdecrease in basonuclin protein content in these transgenic oocytes.These assays, however, failed to distinguish the small difference inthe severely reduced basonuclin protein levels of the two transgeniclines. Conceivably, the exponential nature of the RT-PCR method ismore sensitive in detecting the small variation between the very lowbasonuclin levels, which were below the level of detection ofimmunocytochemistry and western blot. Notably, the small
3RESEARCH ARTICLEBasonuclin is a maternal-effect gene
Fig. 1. Basonuclin expression profile during oogenesis and pre-implantation development. The relative abundance of basonuclinmRNA was plotted using a log scale and data from a previousmicroarray analysis (Zeng et al., 2004) (unbroken line) and real-timePCR (broken line). Immunocytochemical staining of oocytes and pre-implantation embryos are shown in the insets in the graph. Thearrowheads indicate the positions of the germinal vesicle, the pronucleiand the nuclei of the respective developmental stages. An immunoblotof basonuclin at the corresponding developmental stages is shownbelow the graph (75 oocytes/embryos per lane). Developmental stages:GV, germinal vesicle-intact oocyte; Egg, metaphase II-arrested egg; 1C,one-cell embryo; 2C, two-cell embryo; M/B, morulae and blastocyst.

DEVELO
PMENT
4
difference in mRNA levels appeared to produce an effect on thefertility in T8 and T50 lines, which were 50% and 90%, respectively(Fig. 2E) and correlate with the 94% and 98% reduction ofbasonuclin mRNA content.
Our analysis also showed high targeting specificity of thetransgenic RNAi approach in oocytes, consistent with previousreports (Stein et al., 2003; Fedoriw et al., 2004; Yu et al., 2004). Intransgenic oocytes, no detectable change was observed in theamount of mRNA for UBF, a ubiquitous Pol I-specific transcriptionfactor (Fig. 2D). Furthermore, we observed no difference in theabundance of basonuclin 2 mRNA, which is a closely relatedhomolog and shares more than 57% of the nucleotide sequence withbasonuclin (Romano et al., 2004; Vanhoutteghem and Djian, 2004).This high targeting specificity is consistent with a previous reportthat transgenic RNAi targeting oocyte Msy2 mRNA has no effect on
the abundance of the closely related Msy4 transcript (Yu et al., 2004).These results demonstrate the efficacy and specificity of thetransgenic RNAi approach. Thus, as predicted, perturbingbasonuclin function in oocytes reduces fertility (Tian et al., 2001).
Increased oocyte developmental failure inbasonuclin-deficient oocytesTo characterize the cause of the sub-fertility phenotype, we firstexamined the histology of transgenic ovaries. Compared with thecontrol (Fig. 3A,B), histological sections revealed clear changes infollicular structures in adult transgenic ovaries (Fig. 3C,D), whichappeared normal in size and weight. Although the number offollicles appeared unaffected, some follicles exhibited abnormalmorphologies, which could be classified into two discrete categories.In the first category, the follicles were invariably large and the
RESEARCH ARTICLE Development 133 (10)
Fig. 2. Basonuclin-RNAi transgeneand its effect on female fertility,levels of basonuclin mRNA andprotein in oocytes. (A) Structures ofthe two transgene constructs withdifferent vector backbones, splicingsequences and basonuclin cDNAsequences. The nucleotide coordinatesof mouse basonuclin cDNA are shownabove one copy of the inverted repeats(black arrows). (B,C) Fertility of thetransgenic RNAi females was inferred bythe average litter-size. Transgenicfemales (founder, B, or out-crossedprogeny of the founder males, C) weremated with three normal males in arandomized sequence. (B) The data arebased on three matings for eachtransgenic founder, and the control(non-TG) is the average litter size of theseven non-transgenic females. (C) Themating/pregnancy number (n) is shownabove each bar. *P<0.001. BasonuclinmRNA was knocked-down in transgenicGV-stage oocytes as shown by real-timePCR (D), immunocytochemistry (F) andimmunoblot (G). (D) The mRNA for theupstream-binding factor (UBF), aubiquitous Pol I transcription factor, wasused as a control. (E) Real-time PCR datawere converted into the percentage ofwild-type basonuclin mRNA level andplotted against the fertility data from C.(F) Oocytes were stained with anti-basonuclin antibody (red) and DAPI(blue), and the two images are overlaidto indicate the position of germinalvesicle in the transgenic oocytes. Scalebar: 10 �m. (G) Two duplicate sampleswere analyzed for each transgenic line.Each lane contains protein extractedfrom 50 oocytes. After probing forbasonuclin, the membrane was strippedand re-probed with antibody against �-tubulin. TG, transgenic oocytes; non-TG,non-transgenic oocytes.

DEVELO
PMENT
oocytes contained cytoplasmic vesicles or cavities (Fig. 3C,D,arrows), which was not observed in non-transgenic ovaries (Fig.3A,B). In the second category, the follicles were small and irregularin shape, and the oocytes were apparently being degraded andabsorbed (Fig. 3C,D arrowheads). Moreover, cumulus cells incontact with the degenerating oocyte had enlarged nucleoli, whichwere intensely stained by hematoxylin (Fig. 3C,D). Transgenicovaries contained corpora lutea, suggesting ovulation had occurred.However, when compared with non-transgenic ovary (Fig. 3A,B),the transgenic corpora lutea contained more cells, which made themappear darker because of hematoxylin nuclear staining (Fig. 3C,D).The ovarian morphology suggested heterogeneity in the transgenicoocyte population, some oocytes failed, whereas others succeededin reaching maturity.
Maturation of basonuclin-deficient oocytesTo characterize the defect of basonuclin-deficient oocytes, GV-intactoocytes were isolated and observed in culture. The isolated oocytesmost probably belonged to the subpopulation that survivedbasonuclin-deficiency during the growth phase. A striking differencebetween the non-transgenic and transgenic GV-intact oocytes wasthat ~60% of the latter had dark, granular and even opaquecytoplasm (Fig. 3F, arrowhead shows an example). This cytoplasmicchange was accompanied by a roughing of the oocyte surface, asrevealed by differential interference contrast (DIC) microscopy (Fig.3G,H, arrowhead). To ascertain if oocytes with altered morphologydiffered in their ability to mature, GV-intact transgenic oocytes wereisolated and divided into two groups according to their cytoplasmtransparency (transparent and light; opaque and dark) and culturedin vitro. Surprisingly, both the transgenic GV-intact oocytes withlight or dark cytoplasm reached MII, as evidenced by emission ofthe first polar body, at incidence similar to the non-transgeniccontrols (Table 1). These observations strongly suggested that thecytoplasmic and cell surface changes did not interfered with the
maturation of this subpopulation. This conclusion was confirmedwhen the numbers of MII eggs isolated from PMSG/hCG-primedtransgenic mice were found to be the same when compared withcontrols (Table 1).
Pol I-mediated transcription is impaired inbasonuclin-deficient oocytesBecause of the suspected role of basonuclin in rRNA transcription,we used a run-on assay with BrUTP to assess Pol I transcription inbasonuclin-deficient growing oocytes, which were transcriptionallyactive (Fig. 4) as shown by incorporation of BrdU into bothnucleoplasm and nucleolus (Fig. 4A,B). In the presence of a highconcentration of �-amanitin, incorporation of BrU into nucleoplasmwas abolished, suggesting Pol II transcription was inhibited (Fig.4C,D). Under this condition, fluorescence intensity in the basonuclin-deficient oocytes was reduced by ~38% (Fig. 4E). Also reduced werethe numbers of transcription foci, where fluorescence is brighter thanother regions in the nucleolus, suggesting intensive incorporation ofBrUTP, i.e. a site of Pol I transcription. Because rDNA is a multi-copy gene, these foci probably represent the rDNA copies that areactively transcribed during the run-on labeling. Scoring the numberof transcription loci by optically sectioning the nucleolus (Fig. 4F,G)indicated that the number of foci was reduced by ~25% in T50transgenic oocytes when compared with controls (P<0.001) (Fig.4H). Nevertheless, we detected neither a significant change in mature28S and 18S rRNA level by RNase-protection assay (data notshown), nor an apparent change in the rate of protein synthesis in bothgrowing and fully grown transgenic oocytes (Table 1).
Perturbed Pol II transcription in basonuclindeficient-oocytesBecause of the potential of basonuclin as a Pol II transcriptionregulator, we investigated the effect of basonuclin deficiency on PolII transcription by analyzing the global gene expression pattern in
5RESEARCH ARTICLEBasonuclin is a maternal-effect gene
Fig. 3. Basonuclin-RNAi transgene affected the morphology of ovary and oocytes. Histology of non-transgenic (A,B) and T50 transgenic(C,D) ovaries are shown at two magnifications: the entire ovary (A,C) and an enlarged region (B,D). Arrows indicate follicles that contain abnormaloocytes; arrowheads indicate remnants of degenerating follicles. Follicles and oocytes are apparently normal in the transgenic ovary (*). cl, corpusluteum. GV-stage oocytes were isolated and observed under the phase-contrast (E,F) and differential interference contrast (DIC) (G,H) microscopy.Non-transgenic oocytes are shown in E,G, and transgenic ones in F,H. Arrowheads indicate examples of dark granules in the transgenic cytoplasm(F) and the bumps on transgenic cell surface (H). The transgenic population is heterogenous: both morphologically normal and abnormal oocytesare present. Scale bars: 50 �m in E; 10 �m in H.

DEVELO
PMENT
6
fully grown, GV-intact oocytes by microarray analysis. After anactive period of transcription during the first two-thirds of oocytegrowth, transcription decreases such that the fully grown oocyte isessentially transcriptionally quiescent, which stays so during oocytematuration until zygotic gene expression initiates in the late one-cellembryo (Hamatani et al., 2004; Zeng and Schultz, 2005). Therefore,the mRNA composition at the GV-intact stage reflects not only thecumulative effect of Pol II transcription during oocyte growth, butalso is the source of the mRNA at the beginning of oocytematuration and thus may have important bearings on earlyembryonic development.
RNAs of oocytes of four non-transgenic and six transgenic micewere analyzed individually on Affymetrix MOE 430 2.0 gene chips,which contains more than 34,000 transcripts. The resultingmicroarray data had a strong tendency to cluster within control andtransgenic groups (Fig. 5A), when compared with an unbiased genelist. There were 772 and 253 genes, the mRNA levels of which wereup- or downregulated more than twofold, respectively. As expected,basonuclin mRNA level was reduced by 210-fold, the largestmagnitude of change among the up- and downregulated genes; themRNA level of the related basonuclin 2 was not affected. An EASEanalysis (Hosack et al., 2003) (Fig. 5B) showed that the genes whose
RESEARCH ARTICLE Development 133 (10)
Table 1. Properties of basonuclin-RNAi transgenic oocytes and embryosNon-transgenic Transgenic (T50) P value
Ovulation (n=3)* 27.5±3.3 26.1±2.4 0.70
Nuclear maturation (%) (n=3)†
GVBD 88.5±6.7 93.0±1.2 0.58MII 57.9±10.3 61.5±6.3 0.79
Protein synthesis (n=3)‡ 5012±155 4846±224 0.53
Max male pronuclei diameter 3.63±0.06 (n=37) 2.95±0.12 (n=34) <0.001(arbitrary units at 12 hpc)§
Penetrated polyspermy at 12 hpc (%)¶ 2.8±2.8 (n=37) 41.1±12.5 (n=36) 0.04
*The average number of mature oocytes isolated from oviducts of three mice after super-ovulation.†Percentage of cultured oocytes in a particular maturation stage.‡Acid-precipitable [35S] methionine-incorporation (cpm/oocyte) of GV-intact oocytes isolated from 6-week-old mice.§The average of three sets of 12 hpc embryos: two observed with epi-fluorescence and one with confocal. DAPI-stained pronuclei were measured at their maximal diameter.n is the total number of embryos analyzed.¶Poly-pronuclei were used as an indicator to score polyspermy. The averages of three sets of embryos are shown. n is the total number of embryos analyzed.
Fig. 4. Pol I transcription is reduced in basonuclin-RNAi transgenic oocytes. Run-on assays wereperformed on growing oocytes (14 days) from control(A,C,F) and line T50 (B,D,G) in the absence (A,B) andpresence (C,D) of 100 �g/ml �-amanitin. Theincorporated BrdU was visualized by AlexFluor-488-labeled anti-BrdU antibody; nucleoplasm staining isindicated by arrowheads and that of nucleolus byarrows. The fluorescence intensity of nucleolus in run-onwith �-amanitin was measured from micrographs withImageJ (non-T, n=14; T50, n=10), and the average inarbitrary units is shown (E). Nucleoli of 20 oocytes fromeach group were optically sectioned (F, non-T control;G, T50), and the transcription foci (brightly labeledspots) were scored. The average number of foci peroocyte is shown (H). *P<0.05; **P<0.001.

DEVELO
PMENT
expression was upregulated more than twofold belonged to fourprocesses: transcription and DNA binding (39.1%), development(17.5%), intercellular junction/extracellular space (11.3%) and metalion binding (6.5%), whereas the genes downregulated more thantwofold were mostly related to cell motility/adhesion (37.8%),intracellular transport (30.0%) and protein binding (17.8%).Although it remains to be seen how these perturbations in geneexpression contributed to the phenotype, it is clear that: (1) ourRNAi approach was effective and highly specific; (2) Pol IItranscripts were evidently affected by basonuclin-deficiency; and (3)some of the transcript upregulation was probably a secondaryresponse to basonuclin deficiency (see also Discussion). Moreover,these results support the proposal that basonuclin regulates Pol IItranscription, in addition to its function as a Pol I transcriptionregulator.
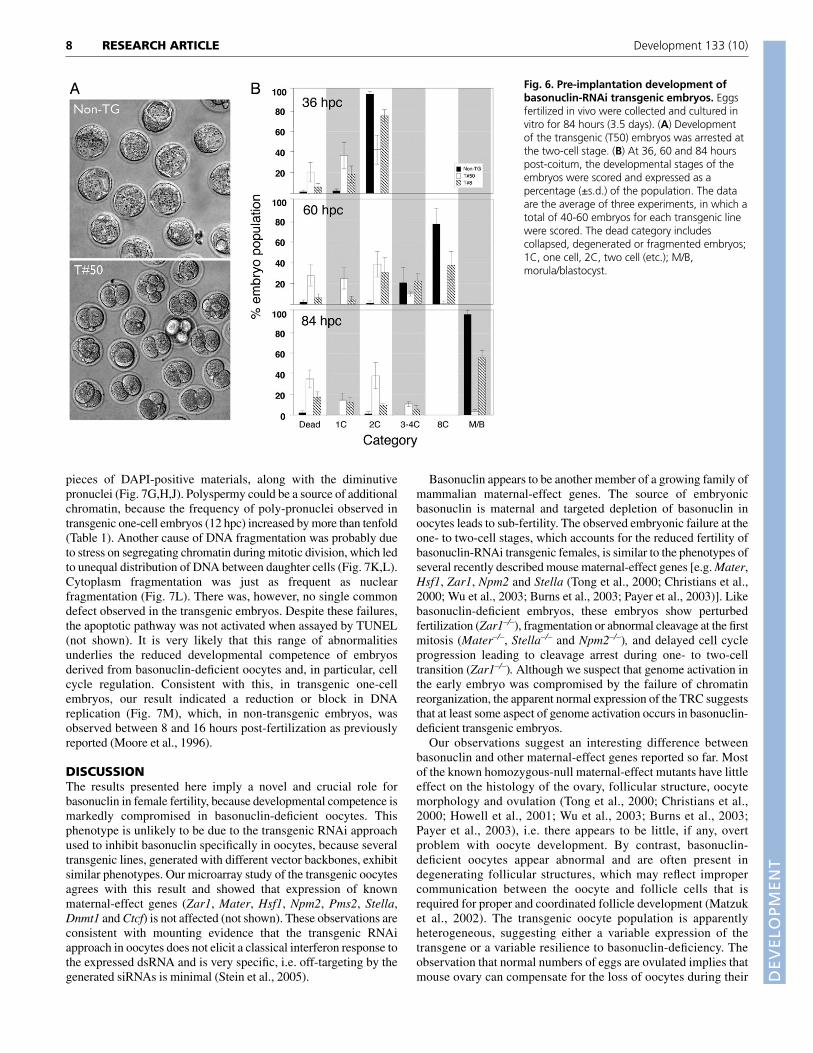
Basonuclin-deficient oocytes lead to earlyembryonic failureOur analysis of isolated oocytes suggested that sub-fertility occurredfollowing oocyte maturation and ovulation. Accordingly, weexamined the ability of transgenic oocytes to be fertilized andsupport pre-implantation development. Fertilized transgenic andcontrol eggs were collected at ~10 hpc, cultured and scored fordevelopmental stages at 36, 60 and 84 hpc. Developmentalprogression of transgenic embryos was clearly delayed or arrested(Fig. 6A). At 36 hpc, virtually all non-transgenic embryos reachedthe two-cell stage, whereas only 75% and 40% of T8 and T50embryos, respectively, did (Fig. 6B). At this point, 60% of the T50embryos were at the one-cell stage, and during the next 48 hours,few of them cleaved and progressed beyond the two-cell stage.Overall, very few T50 embryos could develop beyond the two-cellstage (Fig. 6A,B). Although development of T8 embryos was better,
~60% of them reached the blastocyst stage, the rate of developmentwas nonetheless slower than controls. Once the T8 transgenicembryos reached the eight-cell stage, when an increasing number ofzygotic genes are activated (Hamatani et al., 2004; Wang et al., 2004;Zeng et al., 2004), they readily developed to the blastocyst stagewithout further loss (Fig. 6B). We noted that, overall, at the end ofthis culture period (84 hpc or 3.5 days), 63±11% and 10±3% of theT8 and T50 embryos, respectively, developed beyond the four- toeight-cell stages, which could account for the fertility of these twotransgenic lines (53% and 10% respectively). Thus, early embryonicfailure caused the sub-fertility of the transgenic females.
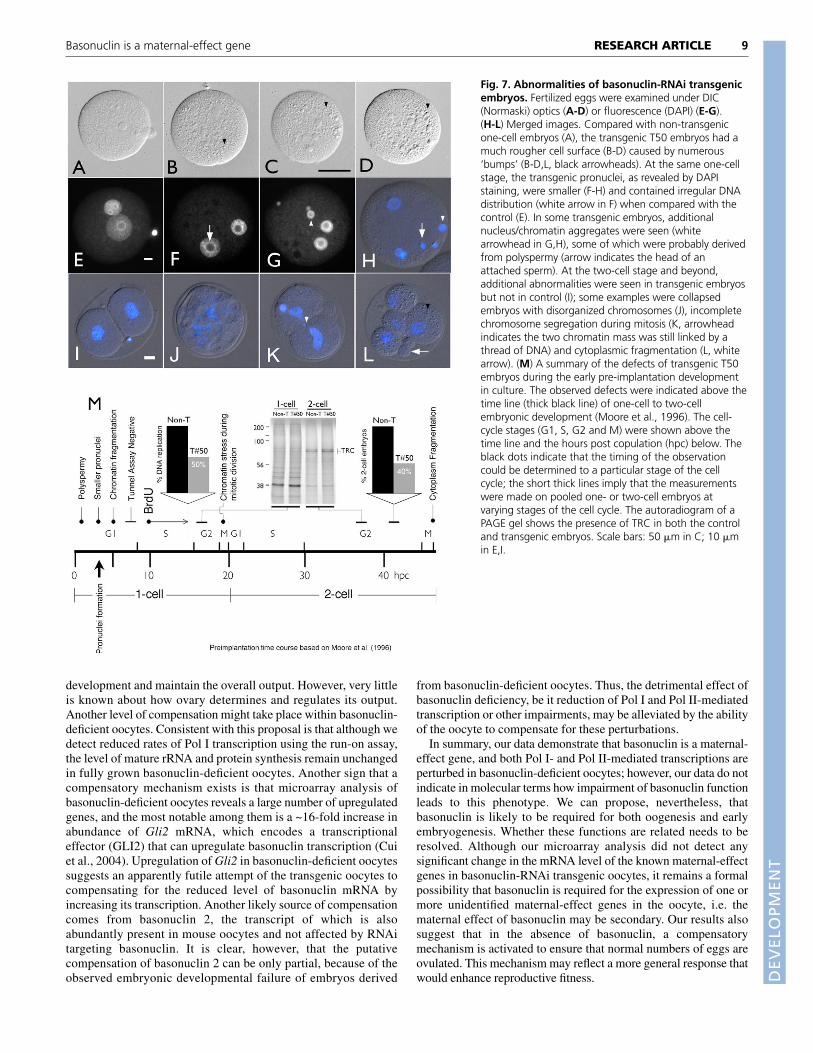
Because most of the embryo loss occurred during the one- andtwo-cell stages, we focused on development during this time. Wefirst assessed if genome activation, which is required fordevelopment beyond the two-cell stage, occurred in basonuclin-deficient two-cell embryos (Schultz, 2002). Genome activation, asassayed by expression of the transcription-requiring complex (TRC),a set of structurally related proteins that are accepted markers ofgenome activation (Conover et al., 1991), did occur in transgenicembryos to the same extent as in controls (Fig. 7M). Thus, it isunlikely that failure to activate the embryonic genome was the basisfor developmental failure and hence we looked at earlier times. Wenoted a variety of abnormalities in the one-cell transgenic embryos.First, and reminiscent of transgenic oocytes (Fig. 4G), transgenicone-cell embryos appeared ‘rough’ under Normaski optics (Fig. 7A-D). Nuclear morphology of the transgenic embryos was also altered;most notably, the male pronuclei were 18.7% smaller in diameterthan that of the control (Fig. 7E-H, Table 1), suggesting a chromatinde-condensation failure. The distribution of chromatin/DNAsurrounding nucleolus in the transgenic pronuclei also differedclearly from that of controls (Fig. 7F arrow). The most strikingdefect, however, was chromatin fragmentation, seen as additional
7RESEARCH ARTICLEBasonuclin is a maternal-effect gene
Fig. 5. Microarray analysis of basonuclin-deficient oocytes. Oocyte RNAs from fournon-transgenic and six transgenic mice wereanalyzed using Affymetrix Mouse GenomeGeneChip MOE 430 v2. Raw microarray datawere processed by the GC-RMA or MAS 5program and analyzed with SAM,GeneSpring and EASE software. (A) A sampleclustering analysis showed a high degree ofassociation within the control and transgenicgroups. (B) Pathway analysis grouped theupregulated genes into four processes, andthat of the downregulated into three. Thepercentage of each group in the up- ordownregulated category is indicated by thelength of the bar (black for upregulated andwhite for downregulated).
DEVELO
PMENT
8
pieces of DAPI-positive materials, along with the diminutivepronuclei (Fig. 7G,H,J). Polyspermy could be a source of additionalchromatin, because the frequency of poly-pronuclei observed intransgenic one-cell embryos (12 hpc) increased by more than tenfold(Table 1). Another cause of DNA fragmentation was probably dueto stress on segregating chromatin during mitotic division, which ledto unequal distribution of DNA between daughter cells (Fig. 7K,L).Cytoplasm fragmentation was just as frequent as nuclearfragmentation (Fig. 7L). There was, however, no single commondefect observed in the transgenic embryos. Despite these failures,the apoptotic pathway was not activated when assayed by TUNEL(not shown). It is very likely that this range of abnormalitiesunderlies the reduced developmental competence of embryosderived from basonuclin-deficient oocytes and, in particular, cellcycle regulation. Consistent with this, in transgenic one-cellembryos, our result indicated a reduction or block in DNAreplication (Fig. 7M), which, in non-transgenic embryos, wasobserved between 8 and 16 hours post-fertilization as previouslyreported (Moore et al., 1996).
DISCUSSIONThe results presented here imply a novel and crucial role forbasonuclin in female fertility, because developmental competence ismarkedly compromised in basonuclin-deficient oocytes. Thisphenotype is unlikely to be due to the transgenic RNAi approachused to inhibit basonuclin specifically in oocytes, because severaltransgenic lines, generated with different vector backbones, exhibitsimilar phenotypes. Our microarray study of the transgenic oocytesagrees with this result and showed that expression of knownmaternal-effect genes (Zar1, Mater, Hsf1, Npm2, Pms2, Stella,Dnmt1 and Ctcf) is not affected (not shown). These observations areconsistent with mounting evidence that the transgenic RNAiapproach in oocytes does not elicit a classical interferon response tothe expressed dsRNA and is very specific, i.e. off-targeting by thegenerated siRNAs is minimal (Stein et al., 2005).
Basonuclin appears to be another member of a growing family ofmammalian maternal-effect genes. The source of embryonicbasonuclin is maternal and targeted depletion of basonuclin inoocytes leads to sub-fertility. The observed embryonic failure at theone- to two-cell stages, which accounts for the reduced fertility ofbasonuclin-RNAi transgenic females, is similar to the phenotypes ofseveral recently described mouse maternal-effect genes [e.g. Mater,Hsf1, Zar1, Npm2 and Stella (Tong et al., 2000; Christians et al.,2000; Wu et al., 2003; Burns et al., 2003; Payer et al., 2003)]. Likebasonuclin-deficient embryos, these embryos show perturbedfertilization (Zar1–/–), fragmentation or abnormal cleavage at the firstmitosis (Mater–/–, Stella–/– and Npm2–/–), and delayed cell cycleprogression leading to cleavage arrest during one- to two-celltransition (Zar1–/–). Although we suspect that genome activation inthe early embryo was compromised by the failure of chromatinreorganization, the apparent normal expression of the TRC suggeststhat at least some aspect of genome activation occurs in basonuclin-deficient transgenic embryos.
Our observations suggest an interesting difference betweenbasonuclin and other maternal-effect genes reported so far. Mostof the known homozygous-null maternal-effect mutants have littleeffect on the histology of the ovary, follicular structure, oocytemorphology and ovulation (Tong et al., 2000; Christians et al.,2000; Howell et al., 2001; Wu et al., 2003; Burns et al., 2003;Payer et al., 2003), i.e. there appears to be little, if any, overtproblem with oocyte development. By contrast, basonuclin-deficient oocytes appear abnormal and are often present indegenerating follicular structures, which may reflect impropercommunication between the oocyte and follicle cells that isrequired for proper and coordinated follicle development (Matzuket al., 2002). The transgenic oocyte population is apparentlyheterogeneous, suggesting either a variable expression of thetransgene or a variable resilience to basonuclin-deficiency. Theobservation that normal numbers of eggs are ovulated implies thatmouse ovary can compensate for the loss of oocytes during their
RESEARCH ARTICLE Development 133 (10)
Fig. 6. Pre-implantation development ofbasonuclin-RNAi transgenic embryos. Eggsfertilized in vivo were collected and cultured invitro for 84 hours (3.5 days). (A) Developmentof the transgenic (T50) embryos was arrested atthe two-cell stage. (B) At 36, 60 and 84 hourspost-coitum, the developmental stages of theembryos were scored and expressed as apercentage (±s.d.) of the population. The dataare the average of three experiments, in which atotal of 40-60 embryos for each transgenic linewere scored. The dead category includescollapsed, degenerated or fragmented embryos;1C, one cell, 2C, two cell (etc.); M/B,morula/blastocyst.
DEVELO
PMENT
development and maintain the overall output. However, very littleis known about how ovary determines and regulates its output.Another level of compensation might take place within basonuclin-deficient oocytes. Consistent with this proposal is that although wedetect reduced rates of Pol I transcription using the run-on assay,the level of mature rRNA and protein synthesis remain unchangedin fully grown basonuclin-deficient oocytes. Another sign that acompensatory mechanism exists is that microarray analysis ofbasonuclin-deficient oocytes reveals a large number of upregulatedgenes, and the most notable among them is a ~16-fold increase inabundance of Gli2 mRNA, which encodes a transcriptionaleffector (GLI2) that can upregulate basonuclin transcription (Cuiet al., 2004). Upregulation of Gli2 in basonuclin-deficient oocytessuggests an apparently futile attempt of the transgenic oocytes tocompensating for the reduced level of basonuclin mRNA byincreasing its transcription. Another likely source of compensationcomes from basonuclin 2, the transcript of which is alsoabundantly present in mouse oocytes and not affected by RNAitargeting basonuclin. It is clear, however, that the putativecompensation of basonuclin 2 can be only partial, because of theobserved embryonic developmental failure of embryos derived
from basonuclin-deficient oocytes. Thus, the detrimental effect ofbasonuclin deficiency, be it reduction of Pol I and Pol II-mediatedtranscription or other impairments, may be alleviated by the abilityof the oocyte to compensate for these perturbations.
In summary, our data demonstrate that basonuclin is a maternal-effect gene, and both Pol I- and Pol II-mediated transcriptions areperturbed in basonuclin-deficient oocytes; however, our data do notindicate in molecular terms how impairment of basonuclin functionleads to this phenotype. We can propose, nevertheless, thatbasonuclin is likely to be required for both oogenesis and earlyembryogenesis. Whether these functions are related needs to beresolved. Although our microarray analysis did not detect anysignificant change in the mRNA level of the known maternal-effectgenes in basonuclin-RNAi transgenic oocytes, it remains a formalpossibility that basonuclin is required for the expression of one ormore unidentified maternal-effect genes in the oocyte, i.e. thematernal effect of basonuclin may be secondary. Our results alsosuggest that in the absence of basonuclin, a compensatorymechanism is activated to ensure that normal numbers of eggs areovulated. This mechanism may reflect a more general response thatwould enhance reproductive fitness.
9RESEARCH ARTICLEBasonuclin is a maternal-effect gene
Fig. 7. Abnormalities of basonuclin-RNAi transgenicembryos. Fertilized eggs were examined under DIC(Normaski) optics (A-D) or fluorescence (DAPI) (E-G).(H-L) Merged images. Compared with non-transgenicone-cell embryos (A), the transgenic T50 embryos had amuch rougher cell surface (B-D) caused by numerous‘bumps’ (B-D,L, black arrowheads). At the same one-cellstage, the transgenic pronuclei, as revealed by DAPIstaining, were smaller (F-H) and contained irregular DNAdistribution (white arrow in F) when compared with thecontrol (E). In some transgenic embryos, additionalnucleus/chromatin aggregates were seen (whitearrowhead in G,H), some of which were probably derivedfrom polyspermy (arrow indicates the head of anattached sperm). At the two-cell stage and beyond,additional abnormalities were seen in transgenic embryosbut not in control (I); some examples were collapsedembryos with disorganized chromosomes (J), incompletechromosome segregation during mitosis (K, arrowheadindicates the two chromatin mass was still linked by athread of DNA) and cytoplasmic fragmentation (L, whitearrow). (M) A summary of the defects of transgenic T50embryos during the early pre-implantation developmentin culture. The observed defects were indicated above thetime line (thick black line) of one-cell to two-cellembryonic development (Moore et al., 1996). The cell-cycle stages (G1, S, G2 and M) were shown above thetime line and the hours post copulation (hpc) below. Theblack dots indicate that the timing of the observationcould be determined to a particular stage of the cellcycle; the short thick lines imply that the measurementswere made on pooled one- or two-cell embryos atvarying stages of the cell cycle. The autoradiogram of aPAGE gel shows the presence of TRC in both the controland transgenic embryos. Scale bars: 50 �m in C; 10 �min E,I.

DEVELO
PMENT
10
The authors thank Jiangchao Qian, Petr Svoboda, Paula Stein, Martin Anger,Sergey Medvedev, Hua Pan and Thom Saunders for their assistance, and JohnTobias for analyzing the microarray data. The authors declare no financialconflict-of-interest in publication of this work. This work was supported bygrants from National Institute of Health to H.T. (HD44096) and R.M.S.(HD22681).
ReferencesBurns, K. H., Viveiros, M. M., Ren, Y., Wang, P., DeMayo, F. J., Frail, D. E.,
Eppig, J. J. and Matzuk, M. M. (2003). Roles of NPM2 in chromatin andnucleolar organization in oocytes and embryos. Science 300, 633-636.
Chatot, C. L., Ziomek, C. A., Bavister, B. D., Lewis, J. L. and Torres, I. (1989).An improved culture medium supports development of random-bred 1-cellmouse embryos in vitro. J. Reprod. Fertil. 86, 679-688.
Christians, E., Davis, A. A., Thomas, S. D. and Benjamin, I. J. (2000). Maternal-effect of Hsf1 on reproductive success. Nature 407, 693-694.
Conover, J. C., Temeles, G. L., Zimmermann, J. W., Burke, B. and Schultz, R.M. (1991). Stage-specific expression of a family of proteins that are majorproducts of zygotic gene activation in the mouse embryo. Dev. Biol. 144, 392-404.
Cui, C., Elsam, T., Tian, Q., Seykora, J. T., Grachtchouk, M., Dlugosz, A. andTseng, H. (2004). Gli proteins up-regulate the expression of basonuclin in basalcell carcinoma. Cancer Res. 64, 5651-5658.
Dean, J. (2002). Oocyte-specific genes regulate follicle formation, fertility and earlymouse development. J. Reprod. Immunol. 53, 171-180.
Erbach, G. T., Lawitts, J. A., Papaioannou, V. E. and Biggers, J. D. (1994).Differential growth of the mouse pre-implantation embryo in chemically definedmedia. Biol. Reprod. 50, 1027-1033.
Fedoriw, A. M., Stein, P., Svoboda, P., Schultz, R. M. and Bartolomei, M. S.(2004). Transgenic RNAi reveals essential function for CTCF in H19 geneimprinting. Science 303, 238-240.
Hamatani, T., Carter, M. G., Sharov, A. A. and Ko, M. S. (2004). Dynamics ofglobal gene expression changes during mouse preimplantation development.Dev. Cell 6, 117-131.
Hosack, D. A., Dennis, G., Jr, Sherman, B. T., Lane, H. C. and Lempicki, R. A.(2003). Identifying biological themes within lists of genes with EASE. GenomeBiol. 4, R70.
Howell, C. Y., Bestor, T. H., Ding, F., Latham, K. E., Mertineit, C., Trasler, J. M.and Chaillet, J. R. (2001). Genomic imprinting disrupted by a maternal-effectmutation in the Dnmt1 gene. Cell 104, 829-838.
Iuchi, S. and Green, H. (1999). Basonuclin, a zinc finger protein of keratinocytesand reproductive germ cells, binds to the rRNA gene promoter. Proc. Natl. Acad.Sci. USA 96, 9628-9632.
Ko, M. S., Kitchen, J. R., Wang, X., Threat, T. A., Hasegawa, A., Sun, T.,Grahovac, M. J., Kargul, G. J., Lim, M. K., Cui, Y. et al. (2000). Large-scalecDNA analysis reveals phased gene expression patterns during preimplantationmouse development. Development 127, 1737-1749.
Mahoney, M. G., Tang, W., Xiang, M. M., Moss, S. B., Gerton, G. L., Stanley,J. R. and Tseng, H. (1998). Translocation of the zinc finger protein basonuclinfrom the mouse germ cell nucleus to the midpiece of the spermatozoon duringspermiogenesis. Biol. Reprod. 59, 388-394.
Matzuk, M. M., Burns, K. H., Viveiros, M. M. and Eppig, J. J. (2002).Intercellular communication in the mammalian ovary: oocytes carry theconversation. Science 296, 2178-2180.
Moore, G. D., Ayabe, T., Kopf, G. S. and Schultz, R. M. (1996). Temporalpatterns of gene expression of G1-S cyclins and cdks during the first and secondmitotic cell cycles in mouse embryos. Mol. Reprod. Dev. 45, 264-275.
Pan, H., O’Brien, M. J., Wigglesworth, K., Eppig, J. J. and Schultz, R. M.
(2005). Transcript profiling during mouse oocyte development and the effect ofgonadotrophin priming and development in vitro. Dev. Biol. 286, 493-506.
Payer, B., Saitou, M., Barton, S. C., Thresher, R., Dixon, J. P., Zahn, D.,Colledge, W. H., Carlton, M. B., Nakano, T. and Surani, M. A. (2003). Stellais a maternal-effect gene required for normal early development in mice. Curr.Biol. 13, 2110-2117.
Romano, R. A., Li, H., Tummala, R., Maul, R. and Sinha, S. (2004).Identification of Basonuclin2, a DNA-binding zinc-finger protein expressed ingerm tissues and skin keratinocytes. Genomics 83, 821-833.
Schultz, R. M. (1993). Regulation of zygotic gene activation in the mouse.BioEssays 15, 531-538.
Schultz, R. M. (2002). The molecular foundations of the maternal to zygotictransition in the pre-implantation embryo. Hum. Reprod. Update 8, 323-331.
Schultz, R. M., Montgomery, R. R. and Belanoff, J. R. (1983). Regulation ofmouse oocyte meiotic maturation: implication of a decrease in oocyte cAMP andprotein dephosphorylation in commitment to resume meiosis. Dev. Biol. 97, 264-273.
Stein, P., Svoboda, P. and Schultz, R. M. (2003). Transgenic RNAi in mouseoocytes: a simple and fast approach to study gene function. Dev. Biol. 256, 187-193.
Stein, P., Zeng, F., Pan, H. and Schultz, R. M. (2005). Absence of non-specificeffects of RNA interference triggered by long double-stranded RNA in mouseoocytes. Dev. Biol. 286, 464-471.
Svoboda, P., Stein, P., Hayashi, H. and Schultz, R. M. (2000). Selectivereduction of dormant maternal mRNAs in mouse oocytes by RNA interference.Development 127, 4147-4156.
Svoboda, P., Stein, P. and Schultz, R. M. (2001). RNAi in mouse oocytes and pre-implantation embryos: effectiveness of hairpin dsRNA. Biochem. Biophys. Res.Commun. 287, 1099-1104.
Tian, Q., Kopf, G. S., Brown, R. S. and Tseng, H. (2001). Function of basonuclinin increasing transcription of the ribosomal RNA genes during mouse oogenesis.Development 128, 407-416.
Tong, Z. B., Gold, L., Pfeifer, K. E., Dorward, H., Lee, E., Bondy, C. A., Dean, J.and Nelson, L. M. (2000). Mater, a maternal-effect gene required for earlyembryonic development in mice. Nat. Genet. 26, 267-268.
Tseng, H., Biegel, J. A. and Brown, R. S. (1999). Basonuclin is associated withthe ribosomal RNA genes on human keratinocyte mitotic chromosomes. J. CellSci. 112, 3039-3047.
Vanhoutteghem, A. and Djian, P. (2004). Basonuclin 2, an extremely conservedhomolog of the zinc finger protein basonuclin. Proc. Natl. Acad. Sci. USA 101,3468-3473.
Wang, Q. T., Piotrowska, K., Ciemerych, M. A., Milenkovic, L., Scott, M. P.,Davis, R. W. and Zernicka-Goetz, M. (2004). A genome-wide study of geneactivity reveals developmental signaling pathways in the preimplantation mouseembryo. Dev. Cell 6, 133-144.
Wu, X., Viveiros, M. M., Eppig, J. J., Bai, Y., Fitzpatrick, S. L. and Matzuk, M.M. (2003). Zygote arrest 1 (Zar1) is a novel maternal-effect gene critical for theoocyte-to-embryo transition. Nat. Genet. 33, 187-191.
Yang, Z. H., Gallicano, G. I., Yu, Q.-C. and Fuchs, E. (1997). An unexpectedlocalization of basonuclin in the centrosome, mitochondria and acrosome ofdeveloping spermatids. J. Cell Biol. 137, 657-670.
Yu, J., Deng, M., Medvedev, S., Yang, J., Hecht, N. B. and Schultz, R. M.(2004). Transgenic RNAi-mediated reduction of MSY2 in mouse oocytes resultsin reduced fertility. Dev. Biol. 268, 195-206.
Zeng, F. and Schultz, R. M. (2005). RNA transcript profiling during zygotic geneactivation in the preimplantation mouse embryo. Dev. Biol. 283, 40-57.
Zeng, F., Baldwin, D. A. and Schultz, R. M. (2004). Transcript profiling duringpre-implantation mouse development. Dev. Biol. 272, 483-496.
RESEARCH ARTICLE Development 133 (10)





























