Embed Size (px)
Citation preview

®¹i häc quèc gia hµ néi
VIÖN VI SINH VËT Vµ C¤NG NGHÖ sinh häc
-----o0o-----
BÁO CÁO ĐỀ TÀI ĐẶC BIỆT CẤP ĐHQG
®Ò tµi nghiªn cøu khoa häc 2007-2008
NGHIÊN CỨU ĐẶC ĐIỂM SINH HỌC VÀ KHẢ NĂNG PHÂN GIẢI ĐỘC TỐ CẢU MỘT SỐ CHỦNG Microcystis PHÂN LẬP Ở HỒ
HOÀN KIẾM, HÀ NỘI
Mã số: QG.07.24
Chñ tr× ®Ò tµi : NguyÔn ThÞ Hoµi Hµ
Hµ néi - 2008

BÁO CÁO TÓM TẮT
1. Tên đề tài
Nghiên cứu đặc điểm sinh học và thăm dò khả năng phân giải độc tố của một
số chủng Microcystis phân lập ở hồ Hoàn Kiếm, Hà Nội
2. Các thành viên tham gia đề tài
Chủ trì đề tài
- Họ và tên: Nguyễn Thị Hoài Hà
Các thành viên:
Lƣu Thị Thùy Giang
Nguyễn Quang Huy
Lê Trung Thủy
Phạm Thị Bích Đào
Trần Thị Điệp
3. Tóm tắt tổng quan của đề tài
Trong những năm gần đây Hà Nội phát triển với tốc độ nhanh chóng, kéo
theo đó là sự ô nhiễm hàng loạt thủy vực nội thành và vùng lân cận. Hồ Hoàn Kiếm
một địa danh nổi tiếng của thủ đô cũng bị ô nhiễm. Chất lƣợng nƣớc thay đổi dẫn
tới sự phát triển thƣờng xuyên và không bình thƣờng của vi khuẩn lam, làm biến đổi
cấu trúc thành phần loài của hệ vi tảo trong hồ. Sự suy giảm một số loài vi tảo đặc
hữu đồng thời với sự gia tăng thành phần loài và mật độ của vi khuẩn lam gây độc ở
hồ Hoàn Kiếm đã đƣợc cảnh báo và gây nên những lo ngại về môi trƣờng cho khu
hệ động thực vật sống trong hồ.
Trong số các loài vi khuẩn lam gây nở hoa nƣớc hồ Microcystis aeruginosa
Kutzing là loài bị bắt gặp thƣờng xuyên và phổ biến nhất M.aeruginosa chứa độc tố
thuộc nhóm hepatotoxin (độc tố gan) cấu tạo từ các peptid mạch vòng có tên gọi là

microcystin.. Microcystin là chất ức chế đối với enzyme protein phosphatases
(nhóm PP1 và PP2A) [30]. Độc tố cũng gây ảnh hƣởng xấu lên hai loại protein
serine và threonin phosphatases. Đã có một số bài báo chứng minh hiện tƣợng nƣớc
nở hoa của vi khuẩn lam gây hại tới sinh vật thủy sinh, động vật và con ngƣời.
Microcystin thƣờng tích luỹ trong nội quan tế bào hoặc dƣới dạng tự do trong nƣớc.
Đã có không ít các công trình nghiên cứu hiện tƣợng nở hoa nƣớc và giảm thiểu tác
hại của độc tố vi khuẩn lam gây độc, tuy nhiên kết quả vẫn còn ở các mức độ khác
nhau. Để bảo vệ môi trƣờng nƣớc hồ, chúng tôi thực hiện đề tài: Nghiên cứu đặc
điểm sinh học và thăm dò khả năng phân giải độc tố của một số chủng Microcystis
phân lập ở hồ Hoàn Kiếm, Hà Nội.
4. Mục tiêu của đề tài
* Phân lập, định tên, nuôi cấy và lƣu giữ đƣợc các chủng vi khuẩn lam
Microcystis trong Bảo tang Vi tảo của Phòng Công nghệ Tảo và Sinh học Môi
trƣờng - Viện Vi sinh vật và Công nghệ Sinh học - ĐHQGHN.
* Thăm dò khả năng phân giải độc tố của chúng bằng vi khuẩn.
5. Tóm tắt nội dung nghiên cứu của đề tài
- Phân tích thành phần thuỷ lý, thuỷ hoá của nƣớc hồ Hoàn Kiếm tại
thời điểm thu mẫu.
- Phân lập, thuần khiết và nghiên cứu các điều kiện nuôi cấy thích hợp
của các chủng Microcystis.
- Phân loại đến loài các chủng Microcystis tuyển chọn đƣợc.
- Tách chiết và phân loại độc tố của các chủng Microcystis trên máy sắc
ký lỏng cao áp (HPLC).
- Thăm dò khả năng phân giải độc tố microcystin của vi khuẩn lam gây
độc bằng vi khuẩn bản địa (vi khuẩn nƣớc).
6. Kết quả chính của đề tài

- Hồ Hoàn Kiếm đang ở tình trạng ô nhiễm, hàm lƣợng COD, BOD vƣợt
tiêu chuẩn Việt Nam, TCVN 4942- 1995 (loại B) 20 lần. Với giá trị NH4+ từ 0,129-
0,699 (mg/l), NO2- từ 0.012- 0,05 (mg/l), NO3
- từ 0,047- 0,129 (mg/l), PO4
3- từ
0,08- 0.64 (mg/l) chứng tỏ hồ có hàm lƣợng dinh dƣỡng N, P khá cao.
- Đã phân lập thuần khiết và nuôi cấy 10 chủng vi khuẩn lam thuộc chi
Microcystis theo phƣơng pháp của Shirai có cải tiến. Môi trƣờng thạch agarose B12
với tỷ lệ 0,4% là môi trƣờng thích hợp nhất cho việc phân lập Microcystis.
- Các chủng Microcystis đều sinh trƣởng tốt trên môi trƣờng Bold 3N với số
lƣợng tế bào đạt từ 32,94 – 68,6 ×106 tế bào/ml
sau 15 đến 19 ngày nuôi cấy. Đặc
biệt chủng LT2 số lƣợng tế bào đạt cao nhất là 68,6 ×106 tế bào/ml sau ngày thứ 19.
Trên môi trƣờng J, các chủng Microcystis sinh trƣởng số lƣợng tế bào của các
chủng đạt từ 40,42 – 76,46 ×106 tế bào/ml
sau 15 đến 19 ngày nuôi cấy. Trong đó
chủng LT2 vẫn phát triển với số lƣợng tế bào đạt cao nhất 76,46 × 106 tế bào/ml
sau
ngày thứ 19. Trên môi trƣờng B12, số lƣợng tế bào đạt từ 35,36 – 54,78 ×106 tế
bào/ml sau 12 đến 15 ngày nuôi cấy. Ở môi trƣờng này chủng LT8 phát triển với
mật độ tế bào đạt cao nhất 54,78 ×106 tế bào/ml
sau ngày thứ 15.
- Có 7 loại axít béo không no dạng C15, C17 và C19 trong tế bào 2 chủng
Microcystis LT2 và LT8. trong đó thành phần axit palmitic (C16H32O2) chiếm lƣợng
lớn nhất với 1,302 μg/mg trọng lƣợng khô ở chủng LT2, chủng LT8 là 0,9683
μg/mg trọng lƣợng khô. Kết quả cho thấy có sự sai khác nhau về lƣợng của 7 loại
axit béo có ở trong tế bào 2 chủng LT2 và LT8.
- Dựa vào phƣơng pháp phân loại truyền thống theo phân loại hình thái học
và phƣơng pháp phân loại hiện đại dựa trên phân tích trình tự rADN 16S, 2 chủng
vi khuẩn LT 2, LT8 đƣợc xác định là Microcystis aeruginosa, 2 chủng vi khuẩn
nƣớc N4 và N7 đƣợc xác định thuộc các loài Sphingomonas mali và Sphingomonas
asaccharolytica
- Dựa vào kết quả phân tích trên HPLC, hàm lƣợng độc tố MC – RR đạt lớn
nhất là 512,8 ng/mg trọng lƣợng khô, độc tố MC – YR là 236,8 ng/mg và thấp nhất
là độc tố MC – LR: 114,25 ng/mg.

- Hai chủng vi khuẩn Sphingomonas tuyển chọn đƣợc có khả phân giải
microcystin khá nhanh. Ở nồng độ microcystin 3 µg/l thời gian bán phân giải là
1,34 giờ và với nồng độ 20µg/l, thời gian là 3,43 giờ.
- Nhiệt độ thích hợp cho sự phân giải microcystin của các chủng vi khuẩn
Sphingomonas mali N4 và Sphingomonas asaccharolytica N7 là 30°C. Sau 7 ngày
sự phân giải gần nhƣ hoàn toàn là 100%.
7. Các công trình công bố và sản phẩm đào tạo
Công trình công bố
- Phạm Thị Mai, Phạm Tiến Đức, Vũ Thị Lan Anh, Nguyễn Thị Hoài Hà
2008. Đánh giá sự biến động số lượng theo mùa và theo tầng nước của vi khuẩn
lam Microcystis aeruginosa ở hồ Hoàn Kiếm, Hà Nội, Tạp chí Khoa học Đại học
Quốc gia Hà Nội. Khoa học Tự nhiên và Công nghệ 24 số 15, tr 125 – 128.
- Phạm Thị Mai, Phạm Tiến Đức, Vũ Thị Lan Anh, Nguyễn Thị Hoài Hà
2008. Tìm hiểu khả năng phân giải độc tố của vi khuẩn lam Microcystis trong hồ
Hoàn kiếm bằng vi khuẩn. Tạp chí Khoa học Đại học Quốc gia Hà Nội. Khoa học
Tự nhiên và Công nghệ 24 số 15, tr 119 – 122.
Đào tạo 4 Cử nhân
- Bƣớc đầu phân lập, tuyển chọn một số chủng vi khuẩn có khả năng phân
giải độc tố của vi khuẩn lam Microcystis ở hồ Hoàn Kiếm, Hà Nội. 2007. Khoá
luận tốt nghiệp hệ đại học chính quy của Đặng Thị Kim Anh.
- Bƣớc đầu đánh giá khả năng phân giải độc tố microcystin của vi khuẩn lam
Microcystis aeruginosa bằng vi khuẩn Sphingomonas. 2008. Khoá luận tốt nghiệp
hệ đại học chính quy của Nguyễn Thị Hoài.
- Khảo sát sự biến động theo mùa của vi khuẩn lam Microcystis aeruginosa ở
hồ Hoàn Kiếm và khả năng ức chế sinh trƣởng của chúng bằng dịch chiết vỏ chuối,
vỏ quýt. 2009. Khoá luận tốt nghiệp hệ đại học chính quy của Lê Thị Trang.
- Phân lập, nghiên cứu một số đặc tính của tảo độc Microcystis tách chiết độc
tố và thăm dò biện pháp xử lý.2009. Đồ án tốt nghiệp của Nguyễn Hữu Hiếu

Đào tạo 1 Thạc sỹ
Phân giải tế bào Microcystis và sự phân huỷ độc tố microcystin bằng vi
khuẩn nƣớc 2007 - 2009.
Những đóng góp của đề tài:
- Là công trình đầu tiên tách và lƣu giữ đƣợc chủng vi khuẩn lam
Microcystis.
- Phân lập đƣợc 2 chủng vi khuẩn thuộc chi mới Sphingomonas có khả năng
phân giải độc tố microcystin của vi khuẩn lam Microcystis.
- Xây dựng đƣợc quy trình tách chiết độc tố microcystin.
8. Tình hình sử dụng kinh phí của đề tài
- Kinh phí đƣợc cấp: 100 000 000 đồng
- Kinh phí thực hiện: 100 000 000 đồng
Hà Nội , ngày 18 tháng 10 năm 2008
Chủ trì đề tài Thủ trƣởng đơn vị
(ký tên, đóng dấu)
TS. Nguyễn Thị Hoài Hà

ABSTRACT OF PROJECT
1. Title of the project: “Study on biological characteristis and capacity of
biodegradation toxin of some Microcystis isolated from Hoan Kiem Lake, Hanoi”
2. Coodinator: Nguyen Thi Hoai Ha
Member: Luu Thuy Giang
Nguyen Quang Huy
Le Trung Thuy
Pham Thi Bich Dao
Tran Thi Diep
3. Introduce
Hoan Kiem Lake is a famous place because of its historical and cultural
value. This is the habitat of valuable and rare species with endemicity and is also
diversifying about the component of microalgae species. In recent years, the
socioeconomic developmental process caused a lot of influences on the lake
ecosystem especially on water quality. Mesotrophic situation of this lake resulted in
the bloom water reducing specific microalgae species and concurrently increasing
the species component and density of toxic algae. Among toxic cyanobacterial
genera, Microcystis accounts for highest population (approximately 80% of total
cyanobacterial strains that cause the bloom water). Microcystis aeruginosa Krutzing
is the most popular species. This is a species which produces hepatotoxin, a type of
liver toxin, composed from cyclic peptides called microcystin.
They have effect on both animals and plants in waters. microcystin usually
accumulates in cells or releases to surrounding waters. Thus the research on toxic
cyanobacteria Microcystis in Hoan Kiem lake, Hanoi and finding effective methods
to diminish their negative effects are essential. In this report, we describe the

isolation, selection and classification location as well as biological characters of
some Microcystis strains from Hoan Kiem lake, as a basic for the next researches.
4.The airm of project
- Isolation, identification, inoculation and maitainance of Microcystis strains
(blue-green, Cyanobacteria) at Microalgal Technology and Enviromental Biology
Lab, Institute of Microbiology and Biotechnology, Vietnam National of University,
Hanoi.
- Capacity of toxin degradation by aquatic bacteria.
5. Brief summary:
- Biophysical and biochemical analysis of freshwater in Hoankiem lake at
sampling time.
- Isolation, screening, study on characteristics of Microcystis isolated.
- Identification of Microcystis isolated.
- Determination microcystin of same Microcystis strains by HPLC.
- Capacity of toxin degradation by fresh bacteria
6. Results obtained
- Hoankiem lake is in pollution condition in the aspect of chemical
evacuation such as COD and BOD concentration is over the corresponding ones in
Vietnamese standard system, TCVN 4942-1995 (type B) 20 times. Concentration of
ammonium cation 0,129- 0,699 (mg/l), nitrite 0.012- 0,05 (mg/l), nitrate 0,047-
0,129 (mg/l), and phosphate 0,08- 0.64 (mg/l) that demonstrate concentration of
nitrogen and phosphorus are relatively high.
- Based on modified Shirai method, 10 pure strains of Microcystis were
isolated and cultured. B12 medium (0.4% agarose) was selected as the most suitable
for Microcystis isolation.
- All of Microcystis strains were grown well on the Bold 3N medium that the
numbers of cell from 68,6 ×106
cells/ml after 15-19 days of culture. Especially, the

maximal cell number of LT2 strain increased to 68,6 ×106
cells/ml after 19th
culture.
For J medium, the cell density was from 40,42 – 76,46 ×106 cells/ml after 15-19
days of culture in which LT2 was the best growth strain with the numbers of cell
76,46 ×106 cells/ml after 19
th culture.
- Analytical results of unsaturated fatty acid in Microcystis LT2 and LT8
strains showed different 7 types of C15, C17 and C19 in which Palmitic acid
occupied as major part with 1,302 μg/mg and 0,9683 μg/mg (in dry weight) in LT2
and LT8, respectively. The results indicated the diference in the concentration
between 7 types of fatty acid.
- Based on morphological and molecular taxonomy (for example
comparision the sequence of 16S rDNA), LT2 and LT8 cyanobacterial strains were
identified as M. aeruginosa; aquatic N4 and N7 bacterial strains were type of
Sphingomonas mali và Sphingomonas asaccharolytica.
- HPLC analysis showed the maximal concentration of MC-RR of 512,8
ng/mg in dry weight, 236,8 ng/mg of MC-YR and the lowest 114,25 ng/mg MC-
LR.
- Sphingomonas strain was demonstrated to degrade microcystin in relatively
good level. Time for degradation of a half of 3 µg/l and 20µg/l microcystin was
1,34 h and 3,43 h; respectively.
- The suitable temperature for microcystin degradation by Sphingomonas
mali N4, Sphingomonas asaccharolytica N7 strains was at 30°C. The degradation
was completely after 7 day.
7. Publications and training activity

Publications
- Pham Thi Mai, Pham Tien Duc, Vu Thi Lan Anh, Nguyen Thi Hoai Ha.
Investigate water quality and the composition of algae in some lakes in Hanoi.
2008. Vietnam National University, Hanoi. Journal Natural Science. Vol 24, No 18,
pp 125 – 128.
- Pham Thi Mai, Pham Tien Duc, Vu Thi Lan Anh, Nguyen Thi Hoai Ha.
Study on toxin lysis in blue green algae by some strains bacteria isolated in Hoan
Kiem lake. 2008. Vietnam National University, Hanoi. Journal Natural Science. Vol
24, No 18, pp 119 – 122.
Training activity
Master thesis completed in 2009
- Lysis of Microcystis and degradation of microcystin by aquatic bacteria.
2007-2009. Tran Thi Diep‟s official undergraduate thesis.
Bacheler thesis completed
- Isolation and screening of Microcystis toxin-degrading bacterial strains in
Hoankiem lake, Hanoi, 2007. Dang Thi Kim Anh‟s official undergraduate thesis.
- Investigation of Microcystis aeruginosa microcystin degradation by
Sphingomonas. 2008. Nguyen Thi Hoai ‟s official undergraduate thesis.
- Study on the seasonal fluctuation of cyanobacterial Microcystis aeruginosa
in Hoankiem lake and growth restriction by banana skin and mandarin peel
extracted solution, 2009. Le Thi Trang‟s official undergraduate thesis.
- Isolation and study on Microcystis toxic cyanobacterial characteristics:
toxin extraction and treatment investigation, 2009. Nguyen Huu Hieu‟s official
undergraduate thesis.
8. Contributions of project:

- Here is the first report that described isolation, maintaince and preservation
of Microcystis strain.
- Two Sphingomonas bacterial strains were isolated and proved as potent
microcystin-degrading agent.
- Establishment of toxin microcystin extraction procedure.
9. Research grant
- Supplied expenditute: 100 000 000 Vnd
- Used expenditute: 100 000 000 Vnd
The project is supported by Vietnam, National University, Hanoi
Hanoi, 18th
October 2008
Project Coodinator Institute of Microbiology and Biotechnology
Nguyen Thi Hoai Ha
Vietnam, National University, Hanoi

MỤC LỤC
MỞ ĐẦU ..................................................................................................................... 1
Chƣơng 1. TỔNG QUAN TÀI LIỆU ......................................................................... 3
1.1. VÀI NÉT VỀ HỒ HOÀN KIẾM ......................................................................... 3
1.2. VI KHUẨN LAM (CYANOBACTERIA) GÂY ĐỘC ....................................... 3
1.2.1. Vi khuẩn lam Microcystis ................................................................................... 6
1.2.2. Độc tố microcystin của Microcystis .................................................................. 6
1.2.3. Cơ chế gây độc của microcystin ......................................................................... 8
1.3. SỰ PHÂN GIẢI ĐỘC TỐ MICROCYSTIN CỦA VI KHUẨN NƢỚC
Sphingomonas ............................................................................................................. 9
1.3.1. Vi khuẩn Sphingomonas ...................................................................................... 9
1.3.2. Cơ chế phân giải độc tố microcystin của Sphingomonas .............................. 10
1.4. ẢNH HƢỞNG CỦA ĐỘC TỐ TỚI NGƢỜI VÀ GIA SÚC ............................ 11
1.5. NHỮNG NGHIÊN CỨU VỀ VI KHUẨN LAM TRÊN THẾ GIỚI VÀ VIỆT
NAM.......................................................................................................................... 13
1.5.1. Trên thế giới ........................................................................................................ 13
1.5.2. Ở Việt Nam ......................................................................................................... 14
Chƣơng 2. NGUYÊN LIỆU VÀ PHƢƠNG PHÁP NGHIÊN CỨU ........................ 16
2.1. ĐỐI TƢỢNG NGHIÊN CỨU ........................................................................... 16
2.1.1. Đối tƣợng nghiên cứu ........................................................................................ 16
2.1.2. Địa điểm và thời gian nghiên cứu .................................................................... 16
2.1.3. Máy móc, dụng cụ .............................................................................................. 17
2.1.4. Hóa chất ............................................................................................................... 17
2.1.5. Môi trƣờng nuôi cấy vi khuẩn lam và vi khuẩn .............................................. 18
2.2. PHƢƠNG PHÁP NGHIÊN CỨU ...................................................................... 18
2.2.1. Phƣơng pháp điều tra thu mẫu vi khuẩn lam ở hồ Hoàn Kiếm và đánh giá
chất lƣợng nƣớc ............................................................................................................. 18
2.2.2. Phƣơng pháp phân lập và nuôi cấy vi khuẩn lam Microcystis ..................... 18
2.2.2.1. Làm giàu mẫu ....................................................................................... 18
2.2.2.2. Phƣơng pháp tách và thuần khiết trên thạch đĩa ................................... 18
2.2.3. Phân loại vi khuẩn lam Microcystis ................................................................. 19
2.2.3.1. Bằng phƣơng pháp hình thái ................................................................. 19
2.2.3.2 . Phƣơng pháp tách ADN ....................................................................... 19
2.2.3.3. Hóa phân loại phân tích thành phần acid béo ....................................... 20

2.2.4. Xác định mật độ tế bào ...................................................................................... 21
2.2.5. Phƣơng pháp tách chiết độc tố microcystin từ tế bào của Microc ystis ....... 22
2.2.6. Phƣơng pháp xác định hàm lƣợng trên máy quang phổ ................................ 22
2.2.7. Các phƣơng pháp sắc ký .................................................................................... 22
2.2.7.1. Phƣơng pháp sắc ký bản mỏng (TLC) .................................................. 23
2.2.7.2. Phƣơng pháp sắc ký lỏng cao áp (HPLC) ............................................ 23
2.2.8. Phƣơng pháp nuôi cấy chủng vi khuẩn Sphingomonas ................................. 24
2.2.9. Phƣơng pháp xác định khả năng phân giải microcystin ................................ 24
Chƣơng 3. KẾT QUẢ VÀ THẢO LUẬN ................................................................ 26
3.1. PHÂN TÍCH CÁC CHỈ TIÊU THỦY LÝ, THỦY HÓA NƢỚC HỒ HOÀN
KIẾM ......................................................................................................................... 26
3.1.1. Nhiệt độ ............................................................................................................... 26
3.1.2. pH ......................................................................................................................... 26
3.1.3. Các chất lơ lửng SS ............................................................................................ 26
3.1.4. Hàm lƣợng oxy hòa tan trong nƣớc (DO- Dissolve oxygen) ........................ 27
3.1.5. Nhu cầu oxy hóa học (COD- Chemical oxygen demand) ............................. 27
3.1.6. Nhu cầu oxy sinh hóa (BOD- Biochemical oxygen demand) ....................... 27
3.2. TUYỂN CHỌN CÁC CHỦNG VI KHUẨN LAM THUỘC CHI Microcystis
PHÂN LẬP ĐƢỢC ................................................................................................... 27
3.2.1. Đặc điểm nuôi cấy .............................................................................................. 27
3.2.2. Phân lập các khuẩn lạc của Microc ystis ......................................................... 29
3.2.3. Khả năng sinh trƣởng của 10 chủng Microcystis LT1, LT2, LT3, LT4,
LT5, LT6, LT7, LT8, LT9 và LT10 trên các môi trƣờng khác nhau ................. 32
3.2.4. Thành phần acid béo trong tế bào của 2 chủng LT2 và LT8 ........................ 35
3.2.5. Xác định và phân tích trình tự rADN 16S ...................................................... 36
3.3. TÁCH CHIẾT ĐỘC TỐ MICROCYSTIN .................................................... 38
3.3.1. Sắc ký bản mỏng (TLC) .................................................................................... 38
3.3.2. Xác định hàm lƣợng microcystin trên máy quang phổ .................................. 38
3.3.3. Hàm lƣợng microcystin trên sắc ký lỏng cao áp (HPLC) ............................. 39
3.4. PHÂN LẬP VÀ TUYỂN CHỌN CÁC CHỦNG VI KHUẨN NƢỚC CÓ KHẢ
NĂNG PHÂN GIẢI MICROCYSTIN ..................................................................... 44
3.4.1. Phân lập các chủng vi khuẩn nƣớc .................................................................. 44
3.4.2. Xác định và phân tích trình tự rADN 16S 2 chủng N4 và N7 ..................... 47
3.5. KHẢ NĂNG PHÂN GIẢI ĐỘC TỐ MICROCYSTIN CỦA 2 CHỦNG
Sphingomonas N4 VÀ N7 ......................................................................................... 49

3.5.1. Hiệu quả phân giải microcystin của 2 chủng N4 và N7 ................................ 49
3.5.2. Xác định điều kiện nhiệt độ thích hợp cho sự phân giải microcystin của
chủng Sphingomonas N4 và N7 .................................................................................. 51
KẾT LUẬN VÀ KIẾN NGHỊ................................................................................... 54
TÀI LIỆU THAM KHẢO ......................................................................................... 56
PHỤ LỤC 1 ............................................................................................................... 62
PHỤ LỤC 2 ............................................................................................................... 65
PHỤ LỤC 3 ............................................................................................................... 66
PHỤ LỤC 4 ............................................................................................................... 77
PHỤ LỤC 5 ............................................................................................................... 78
PHỤ LỤC 6 ................................................................................................................. 1

DANH MỤC HÌNH VÀ BẢNG
Danh mục hình
Hình 1.1. Hồ Hoàn Kiếm Hà Nội ................................................................................ 3
Hình 1.2. Microcystis flos-aquae [62] ........................................................................ 6
Hình 1.3. Microcystis aeruginosa [63] ....................................................................... 6
Hình 1.4. Mô hình cấu trúc hoá học của độc tố MC – LR [54] .................................. 7
Hình 1.5. Mô hình cấu trúc hoá học của độc tố MC – RR [54] .................................. 7
Hình 1.6. Mô hình cấu trúc hoá học của độc tố MC – YR [54] .................................. 8
Hình 1.7. Hình dạng tế bào của Sphingomonas [57] ................................................. 9
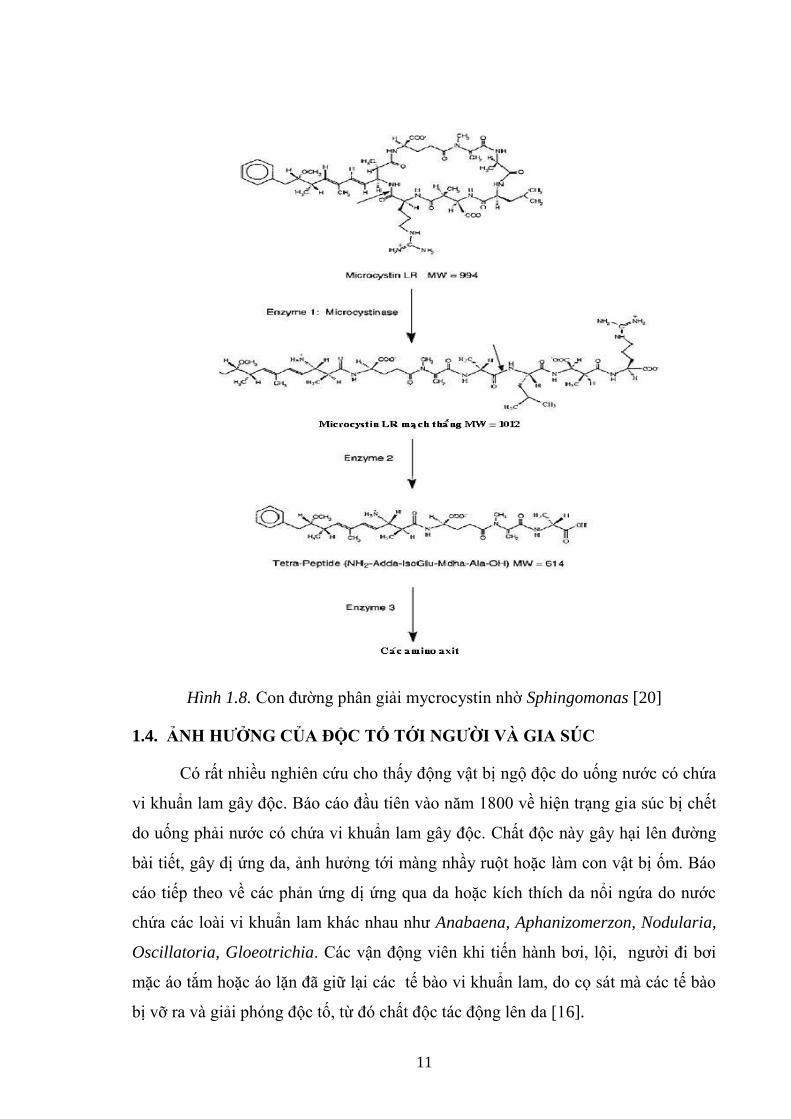
Hình 1.8. Con đƣờng phân giải mycrocystin nhờ Sphingomonas [20] ..................... 11
Hình 2.1. Sơ đồ vị trí thu mẫu ................................................................................... 16
Hình 3.1. Hình thái khuẩn lạc của Microcystis trên môi trƣờng C ......................... 28
Hình 3.2. Hình thái khuẩn lạc của Microcystis trên môi trƣờng Bold 3N ................ 28
Hình 3.3. Hình thái khuẩn lạc của Microcystis trên môi trƣờng B12 ....................... 28
Hình 3.4. Hình thái khuẩn lạc của Microcystis trên môi trƣờng J ........................... 28
Hình 3.5. Hình dạng tế bào chủng Microcystis sp., LT1 .......................................... 29
Hình 3.6. Hình dạng tế bào chủng Microcystis sp., LT2 .......................................... 29
Hình 3.7. Hình dạng tế bào chủng Microcystis sp., LT3 .......................................... 30
Hình 3.8. Hình dạng tế bào chủng Microcystis sp., LT4 .......................................... 30
Hình 3.9. Hình dạng tế bào chủng Microcystis sp., LT5 .......................................... 30
Hình 3.10. Hình dạng tế bào chủng Microcystis sp., LT6 ........................................ 30
Hình 3.11. Hình dạng tế bào chủng Microcystis sp., LT7 ........................................ 31
Hình 3.12. Hình dạng tế bào chủng Microcystis sp., LT8 ........................................ 31
Hình 3.13. Hình dạng tế bào chủng Microcystis sp., LT9 ........................................ 31
Hình 3.14. Hình dạng tế bào chủng Microcystis sp., LT10 ...................................... 31
Hình 3.15. Sinh trƣởng của các chủng Microcystis trong môi trƣờng dịch thể Bold
3N .............................................................................................................................. 33
Hình 3.16. Các chủng Microcystis phát triển tốt trong môi trƣờng dịch thể Bold 3N
................................................................................................................................... 33
Hình 3.17. Sinh trƣởng của các chủng Microcystis trong môi trƣờng dịch thể J ..... 34
Hình 3.18. Các chủng Microcystis phát triển tốt trong môi trƣờng dịch thể J .......... 34
Hình 3.19. Sinh trƣởng của các chủng Microcystis trong môi trƣờng dịch thể B12 35
Hình 3.20. Các chủng Microcystis phát triển tốt trong môi trƣờng dịch thể B12 ..... 35
Hình 3.21. Cây phả hệ dựa trên phân tích giải trình tự rADN 16S của 2 chủng LT2,
LT8 và các loài có quan hệ họ hàng gần ................................................................... 37
Hình 3.22. Kết quả chạy TLC trƣớc khi nhuộm nihyđrin ......................................... 38

Hình 3.23. Kết quả chạy TLC sau khi nhuộm nihyđrin ............................................ 38
Hình 3.24. Đồ thị đƣờng chuẩn microcystin ............................................................. 39
Hình 3.25a. Sắc ký đồ microcystin RR chuẩn ......................................................... 40
Hình 3.25b. Sắc ký đồ microcystin YR chuẩn ......................................................... 40
Hình 3.25c . Sắc ký đồ microcystin LR chuẩn .......................................................... 41
Hình 3.25d. Sắc ký đồ microcystin của chủng LT2 ................................................. 41
Hình 3.25e. Sắc ký đồ microcystin của chủng LT8 ................................................. 42
Hình 3.26. Nuôi cấy làm giàu các chủng vi khuẩn nƣớc trên môi trƣờng NA và M7
................................................................................................................................... 45
Hình 3.27. Cấy pha khuẩn lạc vi khuẩn trên môi trƣờng NA ................................... 45
Hình 3.28. Cây phả hệ dựa trên phân tích giải trình tự rADN 16S của chủng N4 và
các loài có quan hệ họ hàng gần................................................................................ 48
Hình 3.29. Cây phả hệ dựa trên phân tích giải trình tự rADN 16S của chủng N7 với
các loài có quan hệ họ hàng gần................................................................................ 49
Hình 3.30. Thời gian bán phân giải microcystin ở các nồng độ khác nhau của 2
chủng N4, N7. ........................................................................................................... 50
Hình 3.31. Khả năng phân giải microcystin của vi khuẩn Sphingomonas N4 ở các
nhiệt độ khác nhau .................................................................................................... 52
Hình 3.32. Khả năng phân giải của vi khuẩn Sphingomonas N7 ở nhiệt độ khác
nhau ........................................................................................................................... 53
Danh mục bảng
Bảng 1.1. Những loài vi khuẩn lam gây độc và sự phân bố của chúng [16] .............. 4
Bảng 3.1. Khả năng sinh trƣởng của 10 chủng Microcystis trên môi trƣờng Bold3N
................................................................................................................................... 32
Bảng 3.2. Khả năng sinh trƣởng của 10 chủng Microcystis trên môi trƣờng J ........ 33
Bảng 3.3. Khả năng sinh trƣởng của các chủng Microcystis trên môi trƣờng B12 .. 34
Bảng 3.4. Thành phần acid béo có trong tế bào của 2 chủng LT2 và LT8 .............. 36
Bảng 3.5. Hàm lƣợng microcystin của 2 chủng LT2 và LT8 ................................ 43
Bảng 3.6. Đặc điểm hình thái của 2 chủng vi khuẩn nƣớc N4 và N7 ....................... 46
Bảng 3.7. Đặc điểm sinh lý, sinh hoá của 2 chủng N4 và N7 ................................... 46
Bảng 3.8. Hằng số tốc độ phân giải (K) và thời gian bán phân giải microcystin ở các
nồng độ khác nhau .................................................................................................... 50
Bảng 3.9. Tỷ lệ % microcystin không bị phân giải do chủng Sphingomonas N4 ở
các nhiệt độ khác nhau .............................................................................................. 52
Bảng 3.10. Tỷ lệ % microcystin không bị phân giải do chủng Sphingomonas N7 ở
các nhiệt độ khác nhau .............................................................................................. 52
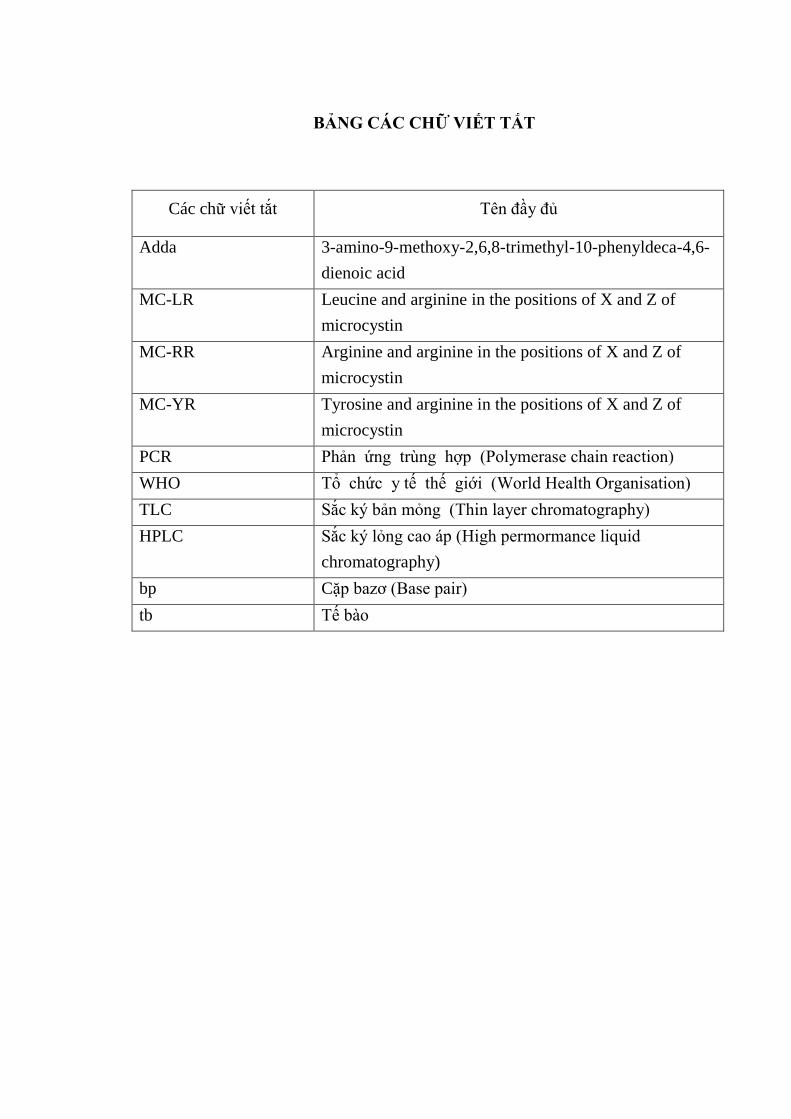
BẢNG CÁC CHỮ VIẾT TẮT
Các chữ viết tắt Tên đầy đủ
Adda 3-amino-9-methoxy-2,6,8-trimethyl-10-phenyldeca-4,6-
dienoic acid
MC-LR Leucine and arginine in the positions of X and Z of
microcystin
MC-RR Arginine and arginine in the positions of X and Z of
microcystin
MC-YR Tyrosine and arginine in the positions of X and Z of
microcystin
PCR Phản ứng trùng hợp (Polymerase chain reaction)
WHO Tổ chức y tế thế giới (World Health Organisation)
TLC Sắc ký bản mỏng (Thin layer chromatography)
HPLC Sắc ký lỏng cao áp (High permormance liquid
chromatography)
bp Cặp bazơ (Base pair)
tb Tế bào

1
MỞ ĐẦU
Trong những năm gần đây Hà Nội phát triển với tốc độ nhanh chóng, kéo
theo đó là sự ô nhiễm hàng loạt thủy vực nội thành và vùng lân cận. Hồ Hoàn Kiếm
một địa danh nổi tiếng của thủ đô cũng bị ô nhiễm. Chất lƣợng nƣớc thay đổi dẫn
tới sự phát triển thƣờng xuyên và không bình thƣờng của vi khuẩn lam, làm biến đổi
cấu trúc thành phần loài của hệ vi tảo trong hồ. Sự suy giảm một số loài vi tảo đặc
hữu đồng thời với sự gia tăng thành phần loài và mật độ của vi khuẩn lam gây độc ở
hồ Hoàn Kiếm đã đƣợc cảnh báo và gây nên những lo ngại về môi trƣờng cho khu
hệ động thực vật sống trong hồ [1, 2, 5, 9].
Trong số các loài vi khuẩn lam gây nở hoa nƣớc hồ Microcystis aeruginosa
Kutzing là loài bị bắt gặp thƣờng xuyên và phổ biến nhất M.aeruginosa chứa độc tố
thuộc nhóm hepatotoxin (độc tố gan) cấu tạo từ các peptid mạch vòng có tên gọi là
microcystin (MC) [14, 15, 19, 23, 29]. Microcystin là chất ức chế đối với enzyme
protein phosphatases (nhóm PP1 và PP2A) [30]. Độc tố cũng gây ảnh hƣởng xấu
lên hai loại protein serine và threonin phosphatases [15, 16, 17, 18, 22]. Đã có một
số bài báo chứng minh hiện tƣợng nƣớc nở hoa của vi khuẩn lam gây hại tới sinh
vật thủy sinh, động vật và con ngƣời [16,17, 19, 20, 21]. Microcystin thƣờng tích
luỹ trong nội quan tế bào hoặc dƣới dạng tự do trong nƣớc [16, 18, 35, 36, 42]. Đã
có không ít các công trình nghiên cứu hiện tƣợng nở hoa nƣớc và giảm thiểu tác hại
của độc tố vi khuẩn lam gây độc, tuy nhiên kết quả vẫn còn ở các mức độ khác
nhau. Để bảo vệ môi trƣờng nƣớc hồ, chúng tôi thực hiện đề tài: “Nghiên cứu đặc
điểm sinh học và thăm dò khả năng phân giải độc tố của một số chủng
Microcystis phân lập ở hồ Hoàn Kiếm, Hà Nội”.
Mục tiêu của đề tài
* Phân lập, định tên, nuôi cấy và lƣu giữ đƣợc các chủng vi khuẩn lam
Microcystis trong Bảo tàng Vi tảo của Phòng Công nghệ Tảo và Sinh học Môi
trƣờng - Viện Vi sinh vật và Công nghệ Sinh học - ĐHQGHN.
* Thăm dò khả năng phân giải độc tố của chúng bằng vi khuẩn.

2
Nội dung của đề tài
- Phân tích thành phần thuỷ lý, thuỷ hoá của nƣớc hồ Hoàn Kiếm tại
thời điểm thu mẫu.
- Phân lập, thuần khiết và nghiên cứu các điều kiện nuôi cấy thích hợp
của các chủng Microcystis.
- Phân loại đến loài các chủng Microcystis tuyển chọn đƣợc.
- Tách chiết và phân loại độc tố của các chủng Microcystis trên máy sắc
ký lỏng cao áp (HPLC).
- Thăm dò khả năng phân giải độc tố microcystin của vi khuẩn lam gây
độc bằng vi khuẩn bản địa (vi khuẩn nƣớc).

3
Chƣơng 1. TỔNG QUAN TÀI LIỆU
1.1. VÀI NÉT VỀ HỒ HOÀN KIẾM
Hồ Hoàn Kiếm nằm ở trung tâm thủ đô Hà Nội, xƣa là một đoạn của dòng
sông hồng và còn có tên gọi khác là sông Nhị. Diện tích hồ ƣớc tính khoảng 12
hecta, chiều dài Nam - Bắc là 700m. Chiều rộng Đông – Tây là 200m.
Hình 1.1. Hồ Hoàn Kiếm Hà Nội
Trong vài thập kỷ gần đây song
song với quá trình phát triển kinh tế
xã hội của Hà Nội,do không có biện
pháp ngăn chặn những tác động gây ô
nhiễm làm biến đổi hệ sinh thái hồ,
ảnh hƣởng tới chất lƣợng nƣớc hồ,
hàm lƣợng N và P tăng lên đã dẫn tới
sự phát triển bùng nổ của vi khuẩn
lam. Hồ Hoàn Kiếm trở thành một
trong những hồ phú dƣỡng ở Hà Nội.
Trong hồ có sự phát triển mạnh mẽ của một số vi khuẩn lam thuộc chi
Microcystis, tạo thành những váng lớn nổi bồng bềnh che kín mặt một góc hồ. Kết
thúc sự nở hoa, vi khuẩn lam chết hàng loạt gây mùi hôi lan xung quanh khu vực hồ
[6].
1.2. VI KHUẨN LAM (CYANOBACTERIA) GÂY ĐỘC
Vi khuẩn lam là nhóm cơ thể cổ xƣa nhất và mang nhiều tên gọi khác nhau
trong quá trình nghiên cứu: thực vật phân cắt (Schizophyceae), tảo nhầy
(Myxophyceae), tảo lam (Cyanophyta), vi khuẩn lam (Cyanobacteria). Từ năm
1854, Kopp đã coi vi khuẩn và tảo lam là một nhóm có chung nguồn gốc phát sinh.
Trong phân loại hiện đại, sinh vật chƣa có cấu trúc nhân hoàn hảo (Procaryota) gồm
vi khuẩn lam và vi khuẩn, căn cứ vào đặc điểm cả hai đều không có nhân điển hình,
chỉ có chất nhân và nhiễm sắc thể. Vi khuẩn lam và vi khuẩn còn có những nét
giống nhau nhƣ: màng tế bào đều cấu tạo từ murein, một loại glucozaminopeptit;

4
trong tế bào chất có nhiều riboxom; nhiều loài vi khuẩn và vi khuẩn lam đều có khả
năng đồng hoá đƣợc N2 tự do; cả hai đều có thể hình thành các pha cyanophase và
bacteriophase. Vi khuẩn lam khác với các ngành tảo và thực vật bậc cao ở cấu trúc
rất cổ xƣa, tế bào không có cấu trúc nhân điển hình và không có các giai đoạn phân
chia nhân. Vi khuẩn lam phân bố khắp nơi trên trái đất, đa số sống trong nƣớc ngọt,
một số phân bố trong thuỷ vực nƣớc mặn giàu chất hữu cơ hoặc trong nƣớc lợ [7].
Độc tố vi khuẩn lam phổ biến nhất là độc tố gan (hepatotoxin) và độc tố thần
kinh (neurotoxin). Một số loài vi khuẩn lam có chứa cả hai loại độc tố trên. Các
quần thể nở hoa trong thuỷ vực nƣớc ngọt chủ yếu thuộc về chi Microcystis thƣờng
gây độc [8, 9], nhƣng cũng có các chủng thuộc chi này không gây độc [29]. Có ý
kiến cho rằng độc tính không phải là tính trạng đặc hiệu ở những loài nhất định nào
đó, nhƣ với microcystin (hepatotoxin) thì độc tính của một chủng phụ thuộc vào
điều kiện là chủng đó có mang gen mã hóa cho microcystin hay không [38, 39]. Thí
nghiệm nuôi cấy vi khuẩn lam cũng chỉ ra rằng sản sinh microcystin là tính trạng
không đổi của một số chủng nhất định, hiếm khi bị tác động bởi các yếu tố môi
trƣờng [16, 49]. Trong khi các điều kiện dẫn tới sự nở hoa của vi khuẩn lam đã
đƣợc sáng tỏ, thì các yếu tố dẫn tới sự bùng nổ của các chủng gây độc so với các
chủng không độc vẫn chƣa đƣợc làm sáng tỏ [15, 20, 22, 34].
Bảng 1.1. Những loài vi khuẩn lam gây độc và sự phân bố của chúng [16]
Loài Loại độc tố Nơi phát sinh
Anabaena spp. (flos-
aquae, lemmermannii,
circinalis)
Microcystins Anh
Anabaena circinalis Microcystins Pháp
Anabaena flos-aquae Microcystins Nauy
Microcystis aeruginosa Microcystins Worldwide
M. viridis Microcystins Nhật Bản
M. botrys Microcystins Đan Mạch
Planktothrix agardhii Microcystins Trung Quốc
P. agardhii Microcystins Nauy
Oscillatoria limosa Microcystins Thuỵ Sỹ
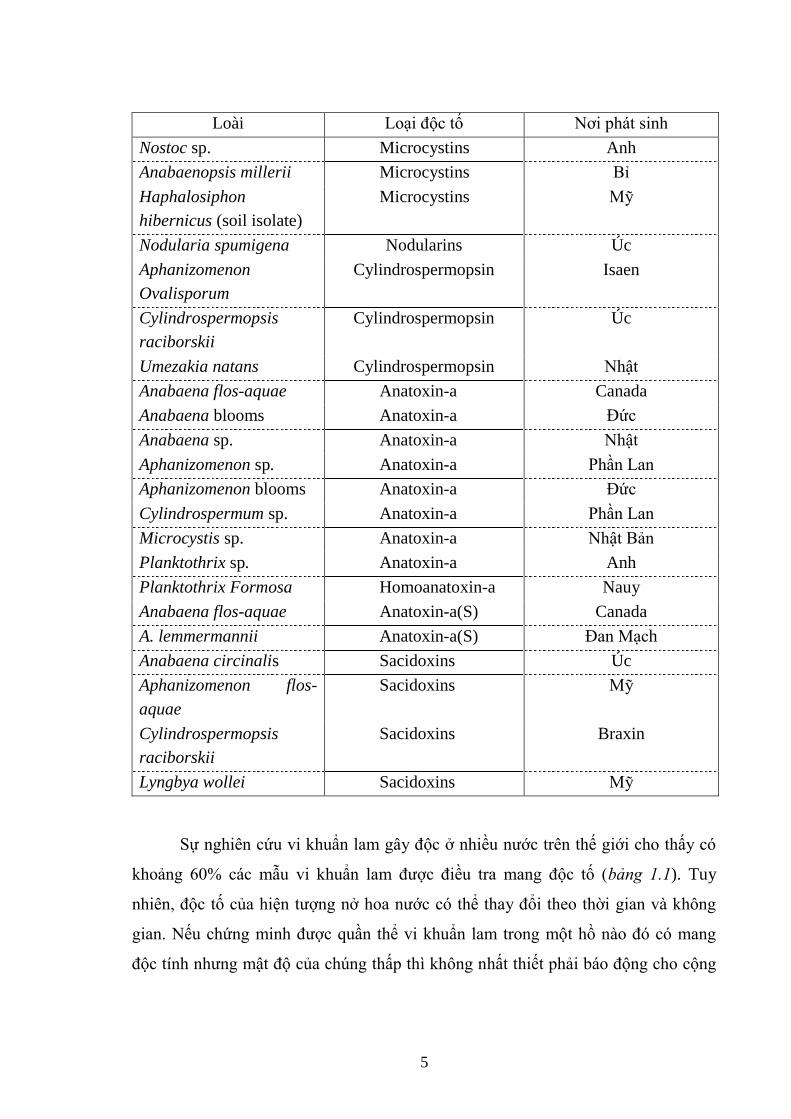
5
Loài Loại độc tố Nơi phát sinh
Nostoc sp. Microcystins Anh
Anabaenopsis millerii Microcystins Bỉ
Haphalosiphon
hibernicus (soil isolate)
Microcystins Mỹ
Nodularia spumigena Nodularins Úc
Aphanizomenon
Ovalisporum
Cylindrospermopsin Isaen
Cylindrospermopsis
raciborskii
Cylindrospermopsin Úc
Umezakia natans Cylindrospermopsin Nhật
Anabaena flos-aquae Anatoxin-a Canada
Anabaena blooms Anatoxin-a Đức
Anabaena sp. Anatoxin-a Nhật
Aphanizomenon sp. Anatoxin-a Phần Lan
Aphanizomenon blooms Anatoxin-a Đức
Cylindrospermum sp. Anatoxin-a Phần Lan
Microcystis sp. Anatoxin-a Nhật Bản
Planktothrix sp. Anatoxin-a Anh
Planktothrix Formosa Homoanatoxin-a Nauy
Anabaena flos-aquae Anatoxin-a(S) Canada
A. lemmermannii Anatoxin-a(S) Đan Mạch
Anabaena circinalis Sacidoxins Úc
Aphanizomenon flos-
aquae
Sacidoxins Mỹ
Cylindrospermopsis
raciborskii
Sacidoxins Braxin
Lyngbya wollei Sacidoxins Mỹ
Sự nghiên cứu vi khuẩn lam gây độc ở nhiều nƣớc trên thế giới cho thấy có
khoảng 60% các mẫu vi khuẩn lam đƣợc điều tra mang độc tố (bảng 1.1). Tuy
nhiên, độc tố của hiện tƣợng nở hoa nƣớc có thể thay đổi theo thời gian và không
gian. Nếu chứng minh đƣợc quần thể vi khuẩn lam trong một hồ nào đó có mang
độc tính nhƣng mật độ của chúng thấp thì không nhất thiết phải báo động cho cộng

6
đồng về những tác hại của chúng đối với sức khỏe con ngƣời và môi trƣờng [16, 22,
43, 48, 50].
1.2.1. Vi khuẩn lam Microcystis
Trong số các loài vi khuẩn lam đƣợc xác định gây độc cho con ngƣời thì
Microcystis aeruginosa đƣợc xem là thƣờng gặp nhất, là một trong những loài vi
khuẩn lam tham gia đáng kể hình thành hiện tƣợng nở hoa của nƣớc. Với đặc điểm
là tập đoàn bao gồm hàng nghìn tế bào riêng lẻ cỡ từ 2- 3 µm và mỗi tế bào đều
chứa một không bào khí. Chính sự tập hợp của hàng nghìn không bào khí định vị
trong vô số các tế bào này giúp chúng nổi trên mặt nƣớc và thu nhận đƣợc nguồn
ánh sáng mặt trời, thông qua quá trình quang hợp tổng hợp sắc tố diệp lục tạo nên
váng xanh trên mặt nƣớc [25].
Hình 1.2. Microcystis flos-aquae [62]
Hình 1.3. Microcystis aeruginosa [63]
1.2.2. Độc tố microcystin của Microcystis
Microcystin [16, 40] là những độc tố vi khuẩn lam gặp phổ biến và rộng rãi
nhất. Hợp chất này đƣợc tách ra đầu tiên từ loài vi khuẩn lam Microcystis
aeruginosa do vậy đƣợc gọi là microcystins. Chúng là các heptapeptide mạch vòng
do có chứa 1 amino acid đặc hiệu (ADDA) chuỗi bên [24]. Cho đến nay, đây là
những dạng cấu trúc chỉ gặp ở microcystins và nodularin (một loại pentapeptide
vòng của vi khuẩn lam ở trong nƣớc lợ). Ngƣời ta đã biết khoảng 70 loại độc tố
microcystins khác nhau. Chúng khác nhau ở các nhóm metyl và 2 amino acid. bên
trong chuỗi. Từ đó dẫn đến sự khác nhau về cấu trúc bậc 4, tính độc cũng nhƣ các
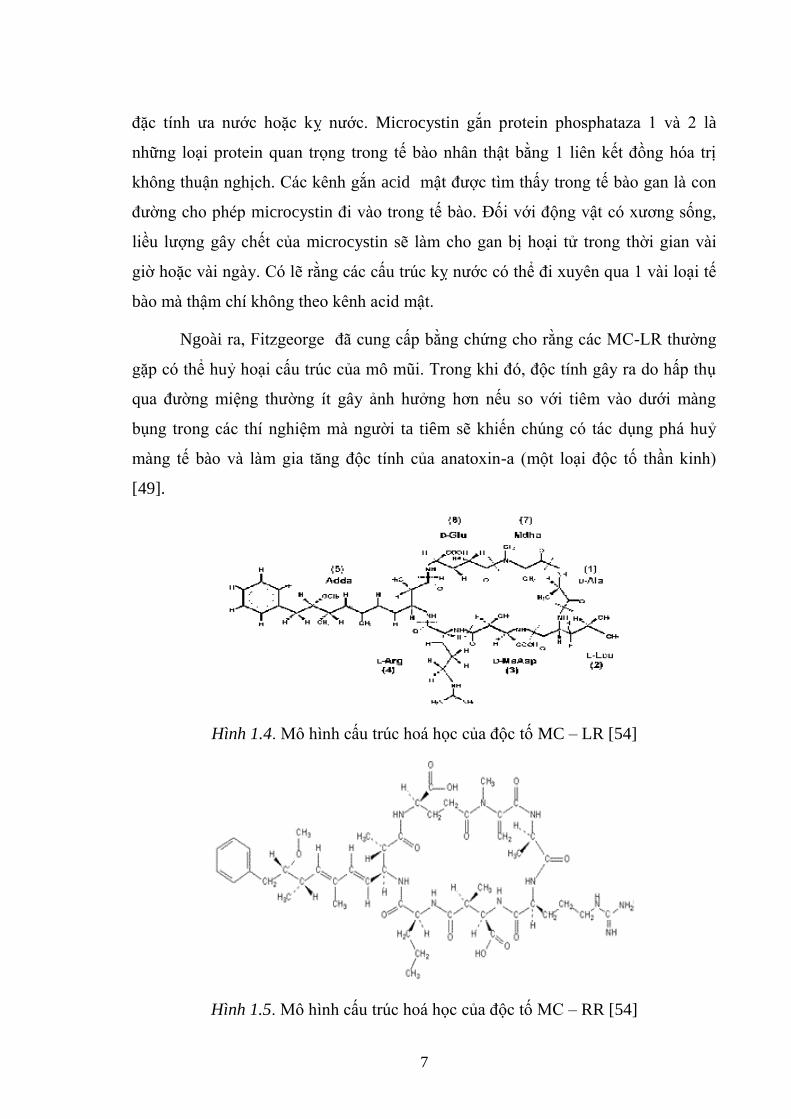
7
đặc tính ƣa nƣớc hoặc kỵ nƣớc. Microcystin gắn protein phosphataza 1 và 2 là
những loại protein quan trọng trong tế bào nhân thật bằng 1 liên kết đồng hóa trị
không thuận nghịch. Các kênh gắn acid mật đƣợc tìm thấy trong tế bào gan là con
đƣờng cho phép microcystin đi vào trong tế bào. Đối với động vật có xƣơng sống,
liều lƣợng gây chết của microcystin sẽ làm cho gan bị hoại tử trong thời gian vài
giờ hoặc vài ngày. Có lẽ rằng các cấu trúc kỵ nƣớc có thể đi xuyên qua 1 vài loại tế
bào mà thậm chí không theo kênh acid mật.
Ngoài ra, Fitzgeorge đã cung cấp bằng chứng cho rằng các MC-LR thƣờng
gặp có thể huỷ hoại cấu trúc của mô mũi. Trong khi đó, độc tính gây ra do hấp thụ
qua đƣờng miệng thƣờng ít gây ảnh hƣởng hơn nếu so với tiêm vào dƣới màng
bụng trong các thí nghiệm mà ngƣời ta tiêm sẽ khiến chúng có tác dụng phá huỷ
màng tế bào và làm gia tăng độc tính của anatoxin-a (một loại độc tố thần kinh)
[49].
Hình 1.4. Mô hình cấu trúc hoá học của độc tố MC – LR [54]
Hình 1.5. Mô hình cấu trúc hoá học của độc tố MC – RR [54]

8
Hình 1.6. Mô hình cấu trúc hoá học của độc tố MC – YR [54]
Biến thể xảy ra chủ yếu tại các vị trí 1 và 2. Ví dụ, Microcystin-LR chứa
leucine amino acid (L) và arginine (R) tại các vị trí 1 và 2 tƣơng ứng. Microcystin-
RR có arginine ở cả hai vị trí. Các nghiên cứu cơ chế độc học tế bào đã cho biết
microcystins ảnh hƣởng đến các cấu trúc tế bào và quá trình nguyên phân và điều
này giúp giải thích khả năng kích thích gây ung thƣ của chúng.
1.2.3. Cơ chế gây độc của microcystin
Cơ chế gây độc của microcystin ở mức độ phân tử là ức chế các hepatocyte
protein phosphatases P 1 (PP1) và 2A (PP2A) – hai enzyme điều chỉnh nhiều quá
trình diễn ra trong tế bào động vật và thực vật. Sự kìm hãm này dẫn tới sự siêu
phosphoryl hóa của tế bào cytoskeratin. Trong tế bào gan của ngƣời và động vật,
điều này còn làm tăng sự phá vỡ các vi sợi hình thành nên khung tế bào. Khi các vi
sợi trở lên tách rời màng tế bào sẽ làm xuất hiện các bóng nƣớc trong màng, đồng
thời làm màng tế bào bị co lại, dịch tế bào thoát ra. Đây đƣợc xem là nguyên nhân
gây ra chảy máu gan, phá vỡ cấu trúc của gan thƣờng thấy ở động vật. Sự tiếp xúc
lâu dài với nồng độ microcystin thấp trong nƣớc uống là nhân tố gây ra ung thƣ gan
ở ngƣời [14]. Các nghiên cứu cơ chế động học tế bào đã cho biết microcystin ảnh
hƣởng lớn đến cấu trúc tế bào và quá trình nguyên phân, điều này giúp giải thích
khả năng kích thích gây ung thƣ của chúng. Độc tố này cũng gây ảnh hƣởng xấu
đến hai loại enzyme là serin – photphatase và threonin – photphatase liên quan tới
sự điều hòa phát triển các tế bào nhân thật (Eukaryote), điều khiển sự hoạt động của
tế bào động thực vật trong quá trình phân chia, sinh trƣởng, trao đổi chất, sao chép

9
ADN và sự biểu hiện các tính trạng có liên quan. Điều đó cho thấy microcystin có
thể ảnh hƣởng đến các chức năng của tế bào nhân thật ở mức độ phân tử và từ đó có
thể ảnh hƣởng tới các động vật thủy sinh sống trong môi trƣờng có sự tiếp xúc với
độc tố này. Microcystin thƣờng tích lũy trong các tế bào vi khuẩn lam, khi nƣớc nở
hoa, độc tố dạng tan hoặc theo phƣơng thức tự nhiên hoặc do sự điều khiển của tế
bào mà giải phóng vào nƣớc. Trong trƣờng hợp nở hoa bởi Microcystis thì
microcystin ở dạng không hòa tan, khá bền vững, sẽ có thể tồn tại vài tuần trƣớc khi
bị phân giải sinh học [10].
1.3. SỰ PHÂN GIẢI ĐỘC TỐ MICROCYSTIN CỦA VI KHUẨN NƢỚC
Sphingomonas
1.3.1. Vi khuẩn Sphingomonas
Hình 1.7. Hình dạng tế bào của
Sphingomonas [57]
Sphingomonas là một vi khuẩn Gram
âm, không bào tử, dị dƣỡng, hoàn toàn
hiếu khí và sinh sắc tố xanh hoặc vàng
nhạt.
Chúng có khả năng di động nhanh nhờ
các lông roi và đa số có khả năng phát
quang dƣới ánh sáng cực tím.
Sphingomonas là vi khuẩn hình que có kích thƣớc 0.3-0.8 x 1.0-2.7µm, chúng có
màu vàng hoặc màu trắng. Sphingomonas hoàn toàn không có lipopolysaccarid
(LPS) – chứa nội độc tố. Thay vào đó, nó có một màng tế bào bao gồm có protein,
photphoslipid, quinone để hô hấp và màng ngoài chứa glycosphingolipit (GSLs).
GSLs giữ một vị trí giống nhƣ LPS trong các vi khuẩn Gram âm khác và xuất hiện
với nhiều chức năng tƣơng tự, nhƣ là một trở ngại đối với các thực thể kháng khuẩn.
Cũng bởi phần hydrocarbon của một GSL ngắn hơn của LPS nên bề mặt tế bào của
Sphingomonas có nhiều hydrophobic hơn các vi khuẩn Gram âm khác. Đây có thể
là một lí do giải thích cho việc loài vi khuẩn này có khả năng làm giảm các
hydrocarbon thơm, đa hydrophobic và tính nhạy cảm của nó đối với việc ngăn các

10
kháng khuẩn hydrophobic. Sphingomonas đƣợc tìm thấy phổ biến trong môi trƣờng
tự nhiên bao gồm trong nƣớc (nƣớc ngọt và nƣớc mặn), trên cạn, trong rễ cây, trong
các mẫu xét nghiệm bệnh và nhiều nơi khác. Sự phân bố rộng rãi của chúng trong
môi trƣờng là do chúng có khả năng sử dụng nhiều hợp chất hữu cơ, có thể sinh
trƣởng và tồn tại dƣới các điều kiện dinh dƣỡng thấp. Nhiều loài Sphingomonas
cũng đƣợc tìm thấy ở các nơi có chứa các hợp chất độc nhƣ PCBs, creozot,
pentachlorophenoL, thuốc diệt cỏ…Các nghiên cứu cho thấy trong môi trƣờng bị ô
nhiễm, Sphingomonas có khả năng sử dụng chất gây ô nhiễm làm nguồn năng
lƣợng để có thể cạnh tranh đƣợc với các sinh vật khác trong các môi trƣờng khác
nhau [20].
1.3.2. Cơ chế phân giải độc tố microcystin của Sphingomonas
Sphingomonas đã đƣợc David G. Bourne chứng minh là chủng vi khuẩn có
chứa các enzyme tham gia vào con đƣờng phân giải độc tố microcystin. Sự phân
giải độc tố này đƣợc xúc tác bởi enzyme nội bào [20].
Việc sử dụng các chất ức chế protease kinh điển cho phép: (1) phân loại các
enzyme này thành các họ protease cơ bản và (2) tích lũy tức thời trong điều kiện
invitro các sản phẩm trung gian từ sự phân hủy MC - LR. Vị trí phân cắt ban đầu
trên phân tử MC - LR bởi enzyme đƣợc xác định là liên kết peptides 3 - amino - 9 -
methoxy - 2,6,8 - trimethyl - deca - 4,6 dienoic acid (Adda) - Arg.
Hai sản phẩm phân giải trung gian đƣợc tìm thấy là MC - LR mạch thẳng:
NH2 - Adda - Glu(iso) -methyldehydroalanine - Ala - Leu - ß - methylaspartate -
Arg - OH (chất A) và sản phẩm còn lại là tetrapeptide: NH2 - Adda – Glu (iso) -
methyldehydroalanine - Ala -OH (chất B) là kết quả của sự làm giảm MC - LR nhờ
hai enzyme trung gian.
Chính hiện tƣợng mở vòng độc tố MC - LR nhờ enzyme microcystinase của
vi khuẩn khiến chất này trở nên không độc do đó giảm đáng kể tƣơng tác với các
protein phosphatase.
Con đƣờng phân hủy MC - LR bởi Sphingomonas đƣợc đề xuất nhƣ sau:
11
Hình 1.8. Con đƣờng phân giải mycrocystin nhờ Sphingomonas [20]
1.4. ẢNH HƢỞNG CỦA ĐỘC TỐ TỚI NGƢỜI VÀ GIA SÚC
Có rất nhiều nghiên cứu cho thấy động vật bị ngộ độc do uống nƣớc có chứa
vi khuẩn lam gây độc. Báo cáo đầu tiên vào năm 1800 về hiện trạng gia súc bị chết
do uống phải nƣớc có chứa vi khuẩn lam gây độc. Chất độc này gây hại lên đƣờng
bài tiết, gây dị ứng da, ảnh hƣởng tới màng nhầy ruột hoặc làm con vật bị ốm. Báo
cáo tiếp theo về các phản ứng dị ứng qua da hoặc kích thích da nổi ngứa do nƣớc
chứa các loài vi khuẩn lam khác nhau nhƣ Anabaena, Aphanizomerzon, Nodularia,
Oscillatoria, Gloeotrichia. Các vận động viên khi tiến hành bơi, lội, ngƣời đi bơi
mặc áo tắm hoặc áo lặn đã giữ lại các tế bào vi khuẩn lam, do cọ sát mà các tế bào
bị vỡ ra và giải phóng độc tố, từ đó chất độc tác động lên da [16].

12
Vào năm 1959, ở Canada mặc dù đã có gia súc bị chết và đƣợc cảnh báo về
nguồn nƣớc sử dụng song ngƣời ta vẫn bơi trong một hồ có chứa nhiều vi khuẩn
lam. Kết quả là có 13 ngƣời bị ốm, đau đầu, đau cơ, ỉa chảy nặng. Trong dịch tiết
của một bệnh nhân ngƣời ta đã phát hiện ra một số lƣợng lớn tế bào Microcystis
spp., và một số đoạn sợi tảo của Anabaena circinalis [19].
Năm 1979, ở Úc để ngăn chặn sự nở hoa của Cylindropermopsis raciborskii
trong nguồn nƣớc uống, ngƣời ta đã xử lý chúng bằng đồng sunfat. Điều này đã làm
cho độc tố trong tế bào đƣợc giải phóng gây ra ảnh hƣởng nghiêm trọng và làm 141
ngƣời phải nhập viện [34].
Năm 1988, ở Brazil tiếp sau trận lụt do vỡ con đập Itaparica thuộc bang
Bahia, khoảng 2000 ca bệnh về đƣờng tiêu hoá đã đƣợc báo cáo sau 42 ngày và có
88 ngƣời đã chết. Các điều tra nguyên nhân sau đó cho biết có hàm lƣợng độc tố vi
khuẩn lam rất cao [16].
Năm 1989, ở Anh có 10 trong số 20 lính đã bị ốm sau khi bơi tập trong nƣớc
bị ô nhiễm bởi vi khuẩn lam Microcystis spp., đã phải nhập viện [17].
Năm 1993, ở Trung Quốc một số ca bệnh ung thƣ gan đƣợc chẩn đoán là có
liên quan đến việc sử dụng nƣớc uống trên bề mặt bị nhiễm nhiều vi khuẩn lam [16,
18].
Năm 1994, ở Thuỵ Điển do quản lý không tốt nguồn nƣớc chƣa đƣợc xử lý
nhà máy đƣờng, nên khi chảy ra nó hoà lẫn vào nƣớc sông. Điều này khiến loài vi
khuẩn lam Planktothrix agardhii đã phát triển mạnh tạo nên hiện tƣợng nở hoa
nƣớc. Kết quả phân tích cho thấy rằng loài này chứa độc tố microcystin. Trong tổng
số 304 nông dân sử dụng nguồn nƣớc này thì có 121 ngƣời, kể cả động vật nuôi của
họ đã bị tiêu chảy, viêm cơ [16, 18].
Năm 1995, ở Úc, kết quả điều tra với 852 bệnh nhân tiếp xúc với nƣớc có
chứa vi khuẩn lam cho thấy rằng sau 2 –7 ngày nhiều bệnh nhân bị mắc tiêu chảy,
sốt dị ứng da…. Ở Monte Hermosa (Achentina) vào tháng 11/1995 đã có 45 triệu
con ngao Mesoderma marcoides đã bị chết do sự nở hoa của tảo gây hại. Cũng vào
năm này ở Việt Nam, loài Noctiluca scintilans nở hoa ở khu vực vịnh Vân Phong –

13
Khánh Hoà vào tháng 5 và 6 đã làm chết 20 tấn tôm hùm với thiệt hại lên đến 6 tỷ
đồng [5].
1.5. NHỮNG NGHIÊN CỨU VỀ VI KHUẨN LAM TRÊN THẾ GIỚI VÀ
VIỆT NAM
1.5.1. Trên thế giới
Những quan sát và phát hiện sớm nhất về vi tảo (trong đó có vi khuẩn lam)
đã đƣợc các nhà thực vật học mô tả. Các nhà nghiên cứu cho rằng nhận thức của
con ngƣời về sự nở hoa gây độc của vi khuẩn lam đƣợc biết đến từ thế kỷ 12 và thời
gian gần đây họ đã cảnh báo các cƣ dân ở nhiều quốc gia nhƣ Trung quốc, Bắc Phi,
Nam Phi về việc sử dụng nguồn nƣớc tại các thủy vực có váng tảo màu xanh [2, 47,
48].
Các biện pháp phân giải độc tố microcystin đã đƣợc nghiên cứu bởi Harada,
ông đã đƣa ra các biện pháp làm giảm hàm lƣợng microcystin trong nƣớc từ kết quả
của các quá trình pha loãng, hấp thụ các hạt vật chất, sự phân giải phụ thuộc nhiệt
độ, pH, sự quang phân và sự phân giải sinh học [13]. Năm 1997, Takenaka và
Watanabe đã kiểm tra các loài vi khuẩn phân lập từ một hồ ở Nhật Bản và phát hiện
ra rằng thông qua hoạt động của enzyme alkalil protease, Pseudomunas aeruginosa
(Sphingomonas) có khả năng làm giảm MC – LR. Đó là các loài có khả năng mở
vòng độc tố và tạo ra dạng mạch thẳng dẫn tới lƣợng độc tố giảm 160 lần. Nhiều
nghiên cứu về việc sử dụng ozon để giảm bớt hiện tƣợng nở hoa của vi khuẩn lam
đã chỉ ra: với tốc độ bơm ozon vào trong nƣớc là 1gm O3 /m3 nƣớc/giờ, hầu hết các
tế bào vi khuẩn có thể bị tiêu diệt trong 15 phút [8]. Tƣơng tự nhƣ vậy các nghiên
cứu về tính ổn định của microcystin dƣới tác động của tia UV cho thấy độc tố bị
phân giải nhanh bởi tia UV ở bƣớc sóng dài tƣơng ứng với sự hấp thụ tối đa lƣợng
độc tố [14].
Trong điều kiện tự nhiên, các vi sinh vật, đặc biệt là các vi khuẩn nƣớc đóng
vai trò quan trong trong việc phân giải độc tố không hòa tan mà đƣợc giải phóng
vào nƣớc. Ngƣời ta đã chứng minh đƣợc rằng microcystin có thể bị phân giải bởi
các vi sinh vật có trong hệ sinh thái thủy vực tự nhiên [12]. Đặc biệt, sự phân giải

14
sinh học microcystin còn xảy ra nhanh hơn ở các thủy vực có sự nở hoa bởi vi
khuẩn lam. Thực tế cho thấy các biện pháp cơ học nhằm lọc, vớt bỏ tảo tiêu tốn
nhiều thời gian, công sức lại không cho hiệu quả cao hoặc các biện pháp lý học, hóa
học (dùng CuSO4, vôi, Al2(SO4)3, ferric chlorit…) thƣờng không khả thi khi áp
dụng đối với các thủy vực sử dụng cho mục đích sinh hoạt bởi một lƣợng hóa chất
tồn dƣ [8]. Nhiều nghiên cứu đã khẳng định tro của lúa mạch, gỗ mục và một số loại
lá đặc biệt là lá sồi có tác dụng ức chế một cách đáng kể sự sinh trƣởng của vi
khuẩn lam gây độc [15]. Dịch chiết từ vỏ quýt và vỏ chuối lùn cũng có khả năng ức
chế hiện tƣợng nƣớc nở hoa bởi vi khuẩn lam [3].
1.5.2. Ở Việt Nam
Ở Việt Nam, nghiên cứu về tảo độc mới chỉ tiến hành trong 1-2 thập niên
vừa qua. Năm 1987, khi một lô ngao Việt Nam xuất sang Cộng đồng chung Châu
Âu (EU) bị phát hiện nhiễm độc tố tảo đã gây chú ý trong dƣ luận. Nghiên cứu thuỷ
triều đỏ do thực vật phù du biển gây ra mới đƣợc bắt đầu từ năm 1993 với những
kết quả ban đầu về thành phần loài và sự phân bố tảo độc hại ở một số vùng biển
của Việt Nam, trong đó đáng chú ý là công bố về “Thực vật phù du độc hại ở biển
Việt Nam” của 2 tác giả Nguyễn Ngọc Lâm và Đoàn Nhƣ Hải tại Hội nghị Quốc tế
lần thứ 7 về Tảo độc tổ chức tại Nhật Bản. Tiếp đó là công bố của Nguyễn Ngọc
Lâm và cộng sự năm 1996 tại Hội nghị Khoa học Biển Asean-Canada về sự bùng
nổ của loài Noctiluca scintilans liên quan đến hiện tƣợng phú dƣỡng ở vịnh Văn
Phong – Khánh Hoà. Nguyễn Thị Minh Huyền (1996) có những ghi nhận đầu tiên
về một số lòai tảo độc ở vùng biển Bạch Long Vĩ. Chu Văn Thuộc (1998) nghiên
cứu tảo biển độc hại ở vùng cửa sông ven biển miền bắc Việt Nam [2, 3, 4, 5, 6, 9].
Năm 1996-1999, đề tài quốc gia do Đặng Ngọc Thanh làm chủ trì đã đƣợc
thực hiện với nội dung nghiên cứu về thành phần loài ở biển Việt Nam và một số
vấn đề liên quan đến sinh thái tảo độc [6]. Theo Trịnh Thị Thanh và Đặng Thị Sy tại
một số đầm ao nuôi thả cá ở Thanh Trì, mặc dù ngành vi khuẩn lam (cyanobacteria)
có thành phần loài không lớn (khoảng 24 loài, chiếm khoảng 13%) nhƣng thƣờng
đạt sinh khối cao vào tháng 6 và 7, đặc biệt tại các ao có sử dụng phân bắc làm
nguồn thức ăn cho cá [6, 9]. Kết quả điều tra nghiên cứu các hồ chứa và một số thuỷ

15
vực thuộc các tỉnh phía Bắc đƣợc tiến hành bởi Đặng Đình Kim, Đặng Hoàng
Phƣớc Hiền (2006) cho thâý những loài tảo có tiềm năng độc gây nở hoa nƣớc
thuộc về các chi Microcystis, Anabaena, Aphanizomenon, Oscillatoria và
Gomphospheria [4]. Theo Dƣơng Đức Tiến (2001-2005) đã xác định trong hồ Hoàn
Kiếm và một số thuỷ vực miền Nam Việt Nam có các chi Microcystis,
Cylindrospermopsis, Anabaena, Oscillatoria, Lyngbya thuộc ngành vi khuẩn lam
(Cyanobacteria) chiếm đa số về thành phần loài gây nở hoa nƣớc. Trong đó có các
loài có khả năng gây độc nhƣ Microcystis aeruginosa, M. viridis, M. botrys, M.
wesenbergii và Cylindrospermopsis raciborskii [9].

16
Chƣơng 2. NGUYÊN LIỆU VÀ PHƢƠNG PHÁP NGHIÊN CỨU
2.1. ĐỐI TƢỢNG NGHIÊN CỨU
2.1.1. Đối tƣợng nghiên cứu
Đối tƣợng nghiên cứu bao gồm:
- Vi khuẩn lam Microcystis đƣợc phân lập từ hồ Hoàn Kiếm
- Vi khuẩn Sphingomonas đƣợc phân lập từ hồ Hoàn Kiếm
2.1.2. Địa điểm và thời gian nghiên cứu
Địa điểm thu mẫu: Hồ Hoàn Kiếm
Thời gian tiến hành nghiên cứu: Từ tháng 1/2007 đến tháng 12/2008 tại
phòng Công nghệ Tảo và Sinh học Môi trƣờng, Viện Vi sinh vật và Công nghệ Sinh
học, Đại học Quốc gia Hà Nội.
Thời điểm thu mẫu: Thu mẫu vào hai đợt. Đợt 1: 6/3/2007, Đợt 2: 8/5/2007
Vị trí các điểm thu mẫu: thu mẫu tại 6 điểm nhƣ trong hình sau.
Hình 2.1. Sơ đồ vị trí thu mẫu
D1: Cửa cống hàng Khay
D2: Đối diện ngân hàng
Incombank – Lê Thái Tổ
D3: Ngân hàng ANZ
D4: Nhà hàng thủy tạ
D5: Cửa cống đối diện tổng công
ty điện lực Việt Nam
D6: Đền Ngọc Sơn

17
2.1.3. Máy móc, dụng cụ
Sử dụng máy móc và dụng cụ có tại phòng Công nghệ Tảo và Sinh học môi
trƣờng, các phòng thí nghiệm thuộc Viện Vi sinh vật và Công nghệ Sinh học- Đại
học Quốc Gia.
Trung tâm Công nghệ Môi trƣờng và Phát triển bền vững- Đại học Khoa học
Tự nhiên.
Kính hiển vi quang học (Olympus, Nhật Bản)
Kính lúp quan sát khuẩn lạc (Olympus, Nhật Bản)
Máy đo pH (Osi, Pháp)
Máy sắc ký lỏng cao áp (High Performance Liquid Chromatography, HPLC
1100, Đức)
2.1.4. Hóa chất
Hóa chất dùng cho phân tích và thử nghiệm khi tiến hành đề tài gồm:
Hóa chất Xuất xứ Hóa chất Xuất xứ
CH3COONa Merk, Đức Nihyđrin Merk, Đức
Methanol Merk, Đức n- butanol Merk, Đức
Isopropanol Merk, Đức Ethyl axetat Merk, Đức
Hoá chất dùng trong phản ứng PCR:
Trình tự các đoạn mồi dùng trong nghiên cứu :
27f 5‟ - TTGATCCTGGCAG - 3‟
780f 5‟ - GATTAGATACCCTGGTAG - 3‟
920r 5‟ - GTCAATTCCTTTGAGTTT - 3‟
1525r 5‟ - AAAGGAGGTGATCCAGCC - 3‟
10 Taq buffer dNTP mix 2.5mM
Taq polymeraza (5U).

18
2.1.5. Môi trƣờng nuôi cấy vi khuẩn lam và vi khuẩn
Nuôi vi khuẩn lam Microcystis trên 6 loại môi trƣờng là Bold 3N, B12, J,
BG11, C, MA (Phụ lục 1)
Nuôi cấy vi khuẩn Sphingomonas trên 2 môi trƣờng NA và M7 (Phụ lục 2).
2.2. PHƢƠNG PHÁP NGHIÊN CỨU
2.2.1. Phƣơng pháp điều tra thu mẫu vi khuẩn lam ở hồ Hoàn Kiếm và đánh
giá chất lƣợng nƣớc
- Điều tra thu mẫu theo quy phạm của Bộ Tài nguyên và Môi trƣờng.
- Các chỉ tiêu đƣợc phân tích dựa theo „Standard methods for the
examination of water and waste water‟ của tổ chức Y tế Mỹ (Phụ lục 3).
2.2.2. Phƣơng pháp phân lập và nuôi cấy vi khuẩn lam Microcystis
2.2.2.1. Làm giàu mẫu
Phƣơng pháp phân lập Microcystis bƣớc 1 làm giàu mẫu [31].
Quan sát nƣớc nở hoa ở hồ Hoàn Kiếm ở thời điểm thu mẫu cho thấy loài
chủ yếu trong quần xã là Microcystis aeruginosa chiếm trên 80%. Hút 1000 µl mẫu
nƣớc hồ có các loài vi khuẩn lam khác nhau, cho vào ống Eppendorf, ly tâm
7000vòng/phút trong 10 phút và rửa 2 lần với dung dịch muối sinh lý 0,05% nhằm
mục đích loại bỏ bớt các loài tảo khác.
Lớp dịch nổi bên trên là vi khuẩn lam, sau đó hút 100 µl dịch nổi bên trên
cho vào nuôi cấy trong các bình tam giác dung tích 500 ml chứa 200 ml môi trƣờng
B12. Nuôi giữ trong điều kiện nhiệt độ 30oC dƣới ánh sáng đèn neon với cƣờng độ
ánh sáng là 1500lux theo quang chu kỳ là 12 giờ chiếu sáng và 12 giờ tối. Sau thời
gian 3 tuần nuôi cấy, quan sát khả năng sinh trƣởng của mẫu vi khuẩn lam đã đƣợc
làm giàu bằng kính hiển vi quang học ở độ phóng đại 400 – 1000 lần.
2.2.2.2. Phương pháp tách và thuần khiết trên thạch đĩa
Quy trình phân lập đƣợc tiến hành theo phƣơng pháp của Shirai có cải tiến
[31]. Các tập đoàn vi khuẩn lam sau khi làm giàu đƣợc xác định dƣới kính hiển vi là

19
các chủng thuộc chi Microcystis trong mẫu nƣớc đƣợc ly tâm 10.000 vòng/phút
trong 15 phút nhằm tiếp tục loại bỏ các loài tảo khác. Tập đoàn vi khuẩn lam nổi
phía trên đƣợc hút ra, đặt vào đĩa petri và tách riêng rẽ từng tập đoàn ra dƣới kính
lúp Olympus. Các tập đoàn đó đƣợc hút riêng vào từng ống Eppendorf, sau đó tiếp
tục sử dụng máy vortex mẫu nhằm mục đích phân tách tập đoàn thành các tế bào
riêng rẽ. Pha loãng mẫu theo phƣơng pháp của Bold và cấy trang trên các môi
trƣờng dinh dƣỡng agaroza có nhiệt độ tạo gel cực thấp (type IX, Sigma) với nồng
độ 0,4%, các hộp đĩa thạch đƣợc quấn kín bằng paraphin và đặt dƣới đèn neon với
cƣờng độ ánh sáng 1000 – 1500 lux ở nhiệt độ 30oC. Sau thời gian từ 07 – 30
ngày, quan sát khuẩn lạc bằng kính lúp Olympus. Những khuẩn lạc sạch sau khi
mọc trên môi trƣờng thạch agaroza đƣợc tách sang các lọ Penicillin chứa 5ml môi
trƣờng dịch thể. Theo dõi sự phát triển của tế bào vi khuẩn lam, quy trình đƣợc tiến
hành lặp đi lặp lại cho đến khi thu đƣợc các chủng thuần khiết.
2.2.3. Phân loại vi khuẩn lam Microcystis
2.2.3.1. Bằng phương pháp hình thái
Các chủng Microcystis sau khi đã phân lập thuần khiết đƣợc quan sát dƣới
kính hiển vi quang học Olympus có độ phóng đại 400-1000 lần để quan sát các đặc
điểm hình thái và kích thƣớc tế bào của chúng. Để định loại, chúng tôi sử dụng các
tài liệu chính sau:
- Phân loại thực vật học bậc thấp .
- Phân loại Vi khuẩn lam ở Việt Nam [07].
- The on-line database of Cyanobacterial genera [27].
2.2.3.2 . Phương pháp tách ADN
ADN đƣợc chiết xuất và tách từ sinh khối ƣớt theo phƣơng pháp [44]. Lấy 3
vòng que cấy sinh khối tế bào sau 14 giờ nuôi, ly tâm lạnh ở 4oC với tốc độ 8000
vòng trong15 phút và rửa 2 lần bằng 300µl dung dịch muối EDTA pH 8,0 (0,5M
Na2EDTA, 0,15M NaCl). Sau đó tạo dịch huyền phù trở lại trong 200µl đệm 10 x
TE (10mM Tris-HCl, 1mM EDTA, pH 8,0, tỷ lệ mẫu:đệm, 1: 2). Bổ sung 10µl của

20
lyzozym (Sigma, 1mg/ml Lysozym), giữ ở 37oC trong nồi cách thuỷ trong 15 phút.
Bổ sung dung dịch Tris-SDS (0,1M Tris-HCl pH 9,0 và 1% SDS) với thể tích bằng
thể tích mẫu. Dịch mẫu đƣợc giữ ở 60oC trong nồi cách thuỷ khoảng 10-30 phút.
Sự tan tế bào đƣợc xác định bằng sự trong suốt của dung dịch cùng với sự tăng độ
nhớt tƣơng ứng. Dịch tan tế bào đƣợc làm lạnh trong đá trong 10 phút, bổ sung
150µl cloroform-isoamyl alcohol, 24:1, v/v (giữ ở 4oC) với thể tích bằng 1/3thể tích
mẫu. Ly tâm lạnh ở 4oC với tốc độ 10.000 vòng trong 20 phút để phân lớp. Sau đó
chuyển phần nhớt ở phần đầu sang Eppendorf (Cỡ. 1,5ml) mới và ADN đƣợc tách
ra bằng bổ sung etanol lạnh (etanol 95,5% giữ ở-22oC) với thể tích gấp đôi thể tích
mẫu và 2µl của 3M Na acetate (pH=4,5).
Ngâm ADN trong 100µl ethanol 70% trong 30 phút và sau đó ngâm liên tiếp
trong ethnol 80, 90, 95% cho mỗi nồng độ trong 5 phút. Sợi ADN làm khô bằng
nhiệt độ phòng trong 10 phút và đƣợc làm tan trở lại trong 20µl đệm 0,1 x TE (10 x
TE) ở 4oC qua đêm.
Dịch đem phân tích bằng máy quang phổ kế ở các bƣớc sóng 230, 260 và
280nm. ADN hấp thụ rất nhanh ở bƣớc sóng 260nm. ở bƣớc sóng 280 chứng tỏ sự
có mặt của protêin. Bƣớc sóng 230nm biểu hiện sự có mặt của chất đệm, các muối
nhƣ EDTA và của ARN. Các tỷ lệ chuẩn về hấp thụ của ADN ở các bƣớc sóng 230
: 260 : 280 là 1,0 : 0,45 : 0,515. Hàm lƣợng ADN đƣợc tính nhƣ sau: 1 đơn vị OD
(mật độ quang, Optical Durity) ở 260nm tƣơng đƣơng với 50µg/ml ADN.
Cách tính: Đơn vị mẫu x độ loãng x chỉ số/1000 = ADN µg/µl.
Nếu ADN chƣa sạch cần khử protein và ARN bằng enzym proteaza K và
RNazaA
2.2.3.3. Hóa phân loại phân tích thành phần acid béo
Lấy 50mg mẫu cho vào ống thủy tinh nút vặn (cỡ 13mm x 100mm) [32].
Alkal hóa bằng 1.0ml chất phản ứng I (reagent I, gồm 15g sodium hydroxide
(NaOH), 50ml methanol và 50ml Milli- Q). Tế bào đƣợc thủy phân ở 1000C tại nồi
nhiệt khô trong 5 phút. Sau đó làm huyền phù bằng máy Vortex trong 5 giây rồi tiếp
tục cho thủy phân trong 25 phút. Dịch thủy phân đƣợc làm lạnh ở nhiệt độ phòng.

21
Methyl hóa dung dịch Alkal với 0.2ml chất phản ứng 2 (65ml 6N hydrochloric acid
và 55ml methanol). Sau khi metyl hóa, giữ ở 800C trong nồi cách thủy khoảng 10
phút. Dung dịch methyl hóa đƣợc bổ sung 1.25ml của chất thử 3 (gồm 50ml n-
hexane-methyl tert butyl ether, 1:1, v/v) và đƣợc trộn đều bằng máy lắc 2000rpm
trong 10 phút. Dịch nhớt phía trên đƣợc loại bỏ. Dung dịch đƣợc bổ sung 3ml thuốc
thử 4 (1.2g sodium hydroxide trong 100ml Milli-Q) và đƣợc trộn đều bằng tay trong
5 phút. Ly tâm ở 2500rpm trong 10 phút. Dịch trên cùng đƣợc chuyển sang ống
thủy tinh mới có nút vặn và cho bay hơi bằng khí nitrogen thông thƣờng tới khi khô.
Cuối cùng, toàn bộ dịch mẫu đƣợc làm tan với 50µl của diethyl ether 300 (hoặc
acetone – 300, n-hexane). Toàn bộ mẫu đƣợc chấm vào sắc ký bản mỏng (TCL) rồi
đƣợc đặt trong bồn thủy tinh chứa 100ml n-hexane-diethyl ether (4:1,v/v) trong 30 –
45 phút. Thành phần acid béo đƣợc hiên vệt bằng iot trong 30 – 45 phút. Acid béo
có cực và không cực xuất hiện nhƣ những vệt xanh. Những vệt đƣợc cạo và rửa
giải hai lần bằng diethyl ether 300. Trộn đều dung dịch silicagel trên máy lắc
2000rpm trong 10 phút và ly tâm 2500rpm trong 5 hoặc 10 phút. Dung dịch đƣợc
bay hơi bằng khí nitrogen thông thƣờng tới khi khô mẫu. Mẫu đƣợc hòa tan trở lại
với 50µl chất phản ứng 3. Dùng pipet nhỏ vào lọ nhỏ GC có nút đậy. Sau đó, đem
phân tích trên máy sắc ký khí (gas chromatography).
2.2.4. Xác định mật độ tế bào
Số lƣợng tế bào vi khuẩn lam trong một thể tích nƣớc đƣợc xác định bằng
cách đếm số tế bào trên 400 ô nhỏ thuộc 25 ô lớn của buồng đếm Goriaev.
Một ô lớn của buồng Goriaev có diện tích 1/25 mm2 và chiều cao 1/10 mm.
Nhƣ vậy thể tích của 1 ô là 1/25 × 1/10 (mm3), thể tích của 25 ô lớn là:
1/25 × 1/10 x 25 (mm3)
Nếu a là số lƣợng tế bào vi khuẩn lam đếm đƣợc trên buồng đếm thì số lƣợng
tế bào tảo trong 1 ml là:
X = a × 104 (tế bào).

22
2.2.5. Phƣơng pháp tách chiết độc tố microcystin từ tế bào của Microc ystis
Trong thí nghiệm của chúng tôi, microcystin của Microcystis đƣợc tách chiết
theo phƣơng pháp đông tan [44]. Thu sinh khối bằng cách ly tâm với tốc độ 7000
vòng/20 phút sau đó lọc dịch nổi bằng thiết bị lọc GF/C (Whatman). Dịch nổi đƣợc
lọc qua màng lọc kích thƣớc 0,2 μm (Milipore). Sinh khối thu đƣợc đông khô ở
nhiệt độ – 83oC, chiết rút độc tố bằng Methanol : nƣớc cất (80:20) và bằng
Methanol : n.Hexan (85:15). Lọc qua cột C18 để loại bỏ sắc tố chiết từ tế bào
Microcystis. Dịch chiết lỏng đƣợc giữ trong catrid Sep-Pak (Water) và rửa bằng
dung dịch methanol 20%. Dịch chiết đƣợc chạy qua sắc ký bản mỏng (TLC) để làm
sạch độc tố, cạo phần độc tố trên bản TLC. Microcystin sau đó đƣợc tách ra nhờ xử
lý với 3 ml methanol 90%, đem sấy khô qua dòng khí nitrogen và hoà tan trong một
lƣợng nhỏ hỗn hợp nƣớc- ethanol 20%. Thành phần và nồng độ của dịch chiết đƣợc
xác định bằng máy HPLC (Hewlett Packard model 1100 LC) qua cột ODS II (4,6
mm; 150 mm) với methanol chứa 0,05 M phosphatase, pH 3 (tỷ lệ 6:4 theo thể
tích), tách trong pha động.
2.2.6. Phƣơng pháp xác định hàm lƣợng trên máy quang phổ
Độc tố microcystin đƣợc phân tích trên máy quang phổ theo phƣơng pháp
[28 ]. Hàm lƣợng microcystin đƣợc tính toán theo phƣơng trình đƣờng chuẩn thu
đƣợc của microcystin chuẩn. Dựa vào đồ thị chuẩn của microcystin theo mật độ
quang, xác định hàm lƣợng microcystin có trong dịch chiết.
Phương pháp xây dựng đường chuẩn hàm lượng microcystin theo mật độ
quang: Dịch chiết MC - LR chuẩn có nồng độ 10ng/ml đƣợc pha loãng gấp 1000,
3000, 5000, 7000, 9000, 11000 lần với hỗn hợp dung môi methanon : nƣớc theo tỷ
lệ 80 : 20. Tiến hành đo ở bƣớc sóng 242 nm trên máy quang phổ ERMA 11
TOKYO. Căn cứ vào các giá trị mật độ quang đo ta lập đƣợc đồ thị biểu thị mối
quan hệ với nồng độ dịch chiết microcystin.
2.2.7. Các phƣơng pháp sắc ký
Là phƣơng pháp hiệu quả cho việc định tính, định lƣợng và tinh sạch các hợp
chất hữu cơ. Nguyên tắc chung của phƣơng pháp sắc ký là dựa trên sự phân bố các

23
chất giữa hai pha là pha động và pha tĩnh. Pha động có thể là chất khí hoặc lỏng để
đẩy mẫu qua vùng chứa pha tĩnh. Pha tĩnh chứa chất rắn hoặc lỏng gọi là các hợp
chất phụ có khả năng chứa các chất hòa tan. Mẫu phân tích chứa một hay nhiều cấu
tử, khi tiếp xúc với pha tĩnh và pha động các cấu tử khác nhau tách nhau ra do
chúng có áp lực khác nhau với cả hai pha.
2.2.7.1. Phương pháp sắc ký bản mỏng (TLC)
Việc tách các chất trong sắc ký bản mỏng [49] dựa trên sự phân tách các chất
giữa hai pha:
- Pha cố định là chất hấp phụ đƣợc trải rộng trên một phiến kính tạo thành
lớp mỏng.
- Pha động là dung môi thích hợp đựng trong bình có nắp đậy kín.
Trong quá trình sắc ký dung môi di chuyển làm dịch chuyển các thành phần
trong mẫu (đã đƣợc chấm thành từng vết trên phiến kính trải chất hấp phụ)
Tiến hành thí nghiệm:
- Pha mẫu sau cô quay thành 1ml bằng hỗn hợp dung môi methanol: nƣớc
(80:20), chấm toàn bộ mẫu lên trên bản silicagel. Đặt bản mỏng trong bồn thủy tinh
chứa 50ml hỗn hợp dung môi ethyl acetat: isopropanol: nƣớc = 8:5:3 (v/v), trong
45-60 phút. Sau khi làm khô tới nhiệt độ phòng, nhuộm TLC bằng nihydrin 2%
trong butanol bão hòa. Sấy khô, quan sát các vết xuất hiện trên nền TLC. Sau đó
đánh dấu, cạo các vết lên cùng với microcystin chuẩn. Tái chiết rút độc tố này bằng
hỗn hợp dung môi methanol : nƣớc (80:20). Đem dịch này ly tâm ở tốc độ 8000
vòng/ phút, trong 15 phút. Sau đó, lọc dịch qua cột C18.
2.2.7.2. Phương pháp sắc ký lỏng cao áp (HPLC)
Sắc ký lỏng cao áp [27 ] là phƣơng pháp chia hỗn hợp các chất trong cột sắc
ký trong đó pha động ở trang thái lỏng nhờ một áp lực lớn đƣợc đẩy qua pha tĩnh
Trong HPLC, mẫu phân tích đƣợc bơm vào cột sắc ký qua một van bơm sáu chiều,
sau đó nhờ bơm cao áp mà pha động đƣợc đẩy qua cột với tốc độ dòng không đổi
trong suốt thời gian chạy sắc ký. Ở đây pha động vừa đóng vai trò là chất mang,

24
mang mẫu phân tích vào cột tách, vừa là chất rửa giải trong suốt quá trình chạy sắc
ký. Pha động có thể là một dung môi hữu cơ hoặc hỗn hợp nhiều dung môi hữu cơ
khác nhau. Thiết kế hệ thống HPLC gồm:
Tiến hành thí nghiệm
Máy HPLC trên cột ODS II 4.6mm x 150mm, đƣờng kính hạt 5 μm. Bộ lọc
dung môi và bộ lọc mẫu đƣờng kính lỗ màng 100Ao. Detector UV-VIS, λ = 242
nm, máy vi tính ghi tín hiệu nối với máy in.
Pha tĩnh: Supelcosil LC –18
Pha động : Methanol chứa 0,5M phosphate = 6:4 (v/v)
Các mẫu đƣợc hòa tan trong hệ dung môi của pha động, tiến hành chạy MC
chuẩn và mẫu nghiên cứu. Tốc độ dòng là 1ml/ phút, pH = 3.
2.2.8. Phƣơng pháp nuôi cấy chủng vi khuẩn Sphingomonas
Chủng vi khuẩn Sphingomonas [22] đƣợc phân lập từ mẫu nƣớc hồ. Lấy 1
ml nƣớc hồ pha loãng ở các nồng độ pha loãng thích hợp. Nhỏ 1ml dịch pha loãng
trên môi trƣờng dịch thể NA và M7. Sau 24 giờ nhỏ 100µl dịch cấy trang đều trên
mặt thạch quan sát thấy các khuẩn lạc, tách và cấy ria nhóm khuẩn lạc cho tách
nhau riêng rẽ. Tách và cấy một khuẩn lạc trên môi trƣờng tƣơng ứng. Quá trình lặp
lại nhiều lần cho đến khi thu đƣợc các chủng thuần khiết.
Nuôi giữ chủng vi khuẩn này trên môi trƣờng thạch nghiêng, bảo quản ở
nhiệt độ 4°C dùng cho các nghiên cứu tiếp theo.
2.2.9. Phƣơng pháp xác định khả năng phân giải microcystin
Sự phân giải đƣợc bắt đầu bằng cách bổ sung dịch chiết microcystin thô vào
các mẫu nƣớc vô trùng để thu đƣợc các nồng độ microcystin khác nhau. Các chủng
vi khuẩn phân giải vi khuẩn lam đƣợc đƣa vào với mật độ 2.5 × 106 tế bào/ml. Sau
20 phút kiểm tra mẫu một lần bằng cách lấy 50ml dịch đem lọc qua màng lọc kích
thƣớc 0,22µm, sau đó đem đo ở bƣớc sóng 242nm để xác định nồng độ độc tố còn
lại. Mẫu đối chứng (không có vi khuẩn) đƣợc tiến hành song song. Các thí nghiệm

25
đƣợc lặp lại 3 lần. Hiệu quả phân giải microcystin đƣợc xác đinh nhờ phƣơng pháp
bán thời gian nhƣ mô tả của Kenefick [28]:
Ct = Co.e(-kt)
Trong đó: Ct – nồng độ microcystin tại thời điểm đo (µg/ml)
Co – nồng độ microcystin trƣớc phân hủy (µg/ml)
k – hằng số phân hủy bậc đầu tiên (ngày -1
)
t – thời gian bán phân hủy (ngày)

26
Chƣơng 3. KẾT QUẢ VÀ THẢO LUẬN
3.1. PHÂN TÍCH CÁC CHỈ TIÊU THỦY LÝ, THỦY HÓA NƢỚC HỒ HOÀN
KIẾM
Đặc điểm của các mùa có ảnh hƣởng tới các quần thể vi tảo và vi khuẩn lam.
nhƣ vi tảo silic kết hợp với việc phát triển nhanh roi trong mùa đông và mùa xuân,
tiếp theo là tảo lục vào cuối mùa xuân và đầu hè. Tại nơi phú dƣỡng và thuỷ vực
lớn, vi khuẩn lam thƣờng chiếm ƣu thế trong các loài thực vật nổi vào cuối xuân
đầu hè. Sự thay đổi về mùa là các nhân tố môi trƣờng thƣờng không đủ để gây ra sự
thay thế vi khuẩn lam bằng các loài thực vật nổi. Vi khuẩn lam là hiện hữu chiếm
ƣu thế hầu hết trong năm. Do vậy, để đánh giá chất lƣợng nƣớc hồ, chúng tôi đã tiến
hành phân tích tổng hợp các thành phần và các điều kiện ảnh hƣởng đến nƣớc.
Trong nghiên cứu này, chúng tôi tiến hành hai đợt thu mẫu vào tháng 03 và 05 năm
2007.
Kết quả phân tích chi tiết đƣợc trình bày ở phụ lục 5.
3.1.1. Nhiệt độ
Nhiệt độ ở một đợt thu mẫu trong cùng một hồ lại khác nhau tùy thuộc vào
thời gian và địa điểm thu mẫu. Tại các cống dẫn nƣớc thải vào hồ, nhiệt độ nƣớc
luôn cao hơn các điểm xa bờ và giữa hồ từ 1- 2°C.
3.1.2. pH
Trong thời gian chúng tôi thu mẫu, pH của hồ khá cao. Giá trị pH tại các
điểm lấy mẫu dao động từ 9,57- 9,84 vào đợt thu mẫu tháng 06/03/2007 và từ 9,77-
10,37 vào đợt thu mẫu tháng 08/05/2007.
3.1.3. Các chất lơ lửng SS
Hàm lƣợng các chất lơ lửng trong hồ tƣơng đối cao, dao động từ 230-
450mg/l trong hai đợt thu mẫu.

27
3.1.4. Hàm lƣợng oxy hòa tan trong nƣớc (DO- Dissolve oxygen)
Từ số liệu nghiên cứu cho thấy hàm lƣơng oxy hòa tan trong nƣớc cao, dao
động từ 9,42mg/l- 22,4mg/l.
Điều này phù hợp với thực tế là vào thời điểm chúng tôi tiến hành thu mẫu,
trong hồ có sự nở hoa rất nhiều của vi khuẩn lam Microcystis, chính sự quang hợp
của những váng nở hoa và các loài vi tảo khác dẫn tới hàm lƣợng oxy hòa tan trong
hồ tăng đáng kể.
3.1.5. Nhu cầu oxy hóa học (COD- Chemical oxygen demand)
Sự dao động giá trị COD trong hồ từ 80,85mg/l- 767,20mg/l. Giá trị COD
cao nhất gấp 20 lần so với tiêu chuẩn Việt Nam loại B về chất lƣợng nƣớc mặt.
3.1.6. Nhu cầu oxy sinh hóa (BOD- Biochemical oxygen demand)
Kết quả nghiên cứu của chúng tôi cho thấy hàm lƣợng BOD trong hồ dao
động thấp nhất là 55 mg/l, cao nhất là 260 mg/l vào tháng 3, ở tháng 5 thấp nhất là
100 mg/l, cao nhất là 548 mg/l. Hàm lƣợng BOD5 cao gấp 20 lần so với tiêu chuẩn
Việt Nam loại B về chất lƣợng nƣớc mặt.
Những nhận xét chung về nước ở hồ Hoàn Kiếm:
Từ các số liệu phân tích đƣợc trong bảng kết quả, có thể nhận xét hồ đang
trong tình trạng ô nhiễm. Với giá trị hàm lƣợng NH4+ dao động 0,129mg/l –
0,699mg/l, NO2- ở khoảng 0.012mg/l – 0,05mg/l, NO3
- từ 0,047mg/l – 0,129mg/l và
PO43-
là 0,08mg/l – 0.64mg/l chứng tỏ nƣớc hồ có hàm lƣợng dinh dƣỡng khá cao.
Sự phát triển của vi khuẩn lam là quanh năm nhƣng tháng 5 phát triển nhiều hơn
tháng 3 bởi thời tiết thƣờng ấm áp hơn. Vi khuẩn lam phát triển dày đặc trên mặt hồ
và đặc biệt tập trung ở khu vực cuối hƣớng gió.
3.2. TUYỂN CHỌN CÁC CHỦNG VI KHUẨN LAM THUỘC CHI
Microcystis PHÂN LẬP ĐƢỢC
3.2.1. Đặc điểm nuôi cấy
Lấy 1 lƣợng mẫu từ các lọ đựng mẫu trong môi trƣờng nuôi cấy làm giàu,
cho vào Eppendorf sạch, ly tâm ở tốc độ 10000 vòng/15 phút. Sau đó, tiến hành pha

28
loãng mẫu ở các nồng độ thích hợp (10-2; 10-3), nhỏ 100µl mẫu và trang đều trên
6 loại môi trƣờng thạch Bold 3N, B12, BG11, C, MA và J. Do M. aeruginosa có
không bào khí (gas vacuoles) nên chúng không tạo thành khuẩn lạc trên các loại
agar hoặc agarose thông thƣờng. Tuy nhiên, điều này có thể đƣợc khắc phục nhờ kỹ
thuật cải tiến của Watanabe [11, 49]. Dịch thể có chứa M. aeruginosa với môi
trƣờng sử dụng loại agarose có nhiệt độ tạo gel cực thấp nồng độ 0,4% (type IX,
Sigma) đƣợc sử dụng làm giá thể. Các đĩa thạch sau khi cấy đƣợc quấn paraphin,
đặt dƣới cƣờng độ ánh sáng 1500 Lux.
Sau 10 ngày quan sát khuẩn lạc.
Kết quả đƣợc trình bày ở hình 3.1 ; 3.2 ; 3.3 ; 3.4
Hình 3.1. Hình thái khuẩn lạc của
Microcystis trên môi trƣờng C
Hình 3.2. Hình thái khuẩn lạc của
Microcystis trên môi trƣờng Bold 3N
Hình 3.3. Hình thái khuẩn lạc của
Microcystis trên môi trƣờng B12
Hình 3.4. Hình thái khuẩn lạc của
Microcystis trên môi trƣờng J

29
Kết quả thử nghiệm cho thấy trên 3 môi trƣờng Bold 3N, B12 và J khuẩn lạc
của Microcystis có khả năng mọc tốt hơn so với các môi trƣờng BG11, C, MA. Ở 3
môi trƣờng sau, khuẩn lạc Microcystis sau 2 - 4 tuần nuôi mới thấy xuất hiện khuẩn
lạc. Điều này cho thấy trên các môi trƣờng cơ chất khác nhau, dẫn đến khả năng
sinh trƣởng của chúng cũng khác nhau.
3.2.2. Phân lập các khuẩn lạc của Microc ystis
Phân lập các khuẩn lạc Microcystis trên các môi trƣờng nuôi cấy, bƣớc đầu
chúng tôi thu đƣợc 10 chủng. Các chủng này có những đặc điểm hình thái tế bào
đƣợc nghiên cứu dƣới kính hiển vi quang học và đƣợc phát triển nuôi cấy trên các
loại môi trƣờng khác nhau. Kết quả đƣợc trình bày ở hình 3.5; 3.6; 3.7; 3.8; 3.9;
3.10; 3.11; 3.12; 3.13; 3.14
Hình 3.5. Hình dạng tế bào chủng
Microcystis sp., LT1
Tập đoàn với tế bào hình cầu, tế bào có
màu xanh lam với nhiều không bào khí,
đƣờng kính tế bào 3-7 μm. Bao nhầy
không màu, mờ bao quanh tế bào.
Hình 3.6. Hình dạng tế bào chủng
Microcystis sp., LT2
Tập đoàn với tế bào hình cầu,tế bào có
màu xanh nhạt, đƣờng kính tế bào 2-5 μm,
tập đoàn hình dáng không ổn định, các
khối nhỏ hình cầu hoặc thấu kính hơi dẹt.

30
Hình 3.7. Hình dạng tế bào chủng
Microcystis sp., LT3
Tập đoàn với tế bào hình cầu,tế bào có
màu xanh nhạt, đƣờng kính tế bào 2-5
μm, tập hợp thành từng tập đoàn nhỏ.
Hình 3.8. Hình dạng tế bào chủng
Microcystis sp., LT4
Tập đoàn với tế bào hình cầu,tế bào có
màu xanh lam với nhiều không bào khí,
đƣờng kính tế bào 5-7 μm, bao nhầy
không màu.
Hình 3.9. Hình dạng tế bào chủng
Microcystis sp., LT5
Tập đoàn với tế bào hình cầu,tế bào có
màu xanh lam với nhiều không bào khí,
đƣờng kính tế bào 3-7 μm, bao nhầy
không màu.
Hình 3.10. Hình dạng tế bào chủng
Microcystis sp., LT6
Tập đoàn với tế bào hình cầu,tế bào có
màu xanh nhạt, đƣờng kính tế bào 4-7 μm,
tập hợp thành từng tập đoàn nhỏ.

31
Hình 3.11. Hình dạng tế bào chủng
Microcystis sp., LT7
Tế bào hình cầu,tế bào có màu xanh
nhạt, đƣờng kính tế bào 3-6 μm, tập
hợp thành từng tập đoàn nhỏ.
Hình 3.12. Hình dạng tế bào chủng
Microcystis sp., LT8
Tập đoàn với tế bào hình cầu,tế bào có
màu xanh lam với nhiều không bào khí,
đƣờng kính tế bào 5-7 μm, thƣờng sắp xếp
dày đặc.
Hình 3.13. Hình dạng tế bào chủng
Microcystis sp., LT9
Tập đoàn với tế bào hình cầu,tế bào có
màu xanh nhạt, các khối nhỏ hình cầu
đƣờng kính tế bào 2-4 μm.
Hình 3.14. Hình dạng tế bào chủng
Microcystis sp., LT10
Tập đoàn với tế bào hình cầu,tế bào có
màu xanh lam, có đƣờng kính tế bào 2-4
μm, chứa không bào khí
Kết quả ở hình 3.5; 3.6; 3.7; 3.8; 3.9; 3.10; 3.11; 3.12; 3.13; 3.14 cho
thấy đặc điểm hình thái của 10 chủng trên có nhiều điểm tƣơng đồng, ở chủng LT2
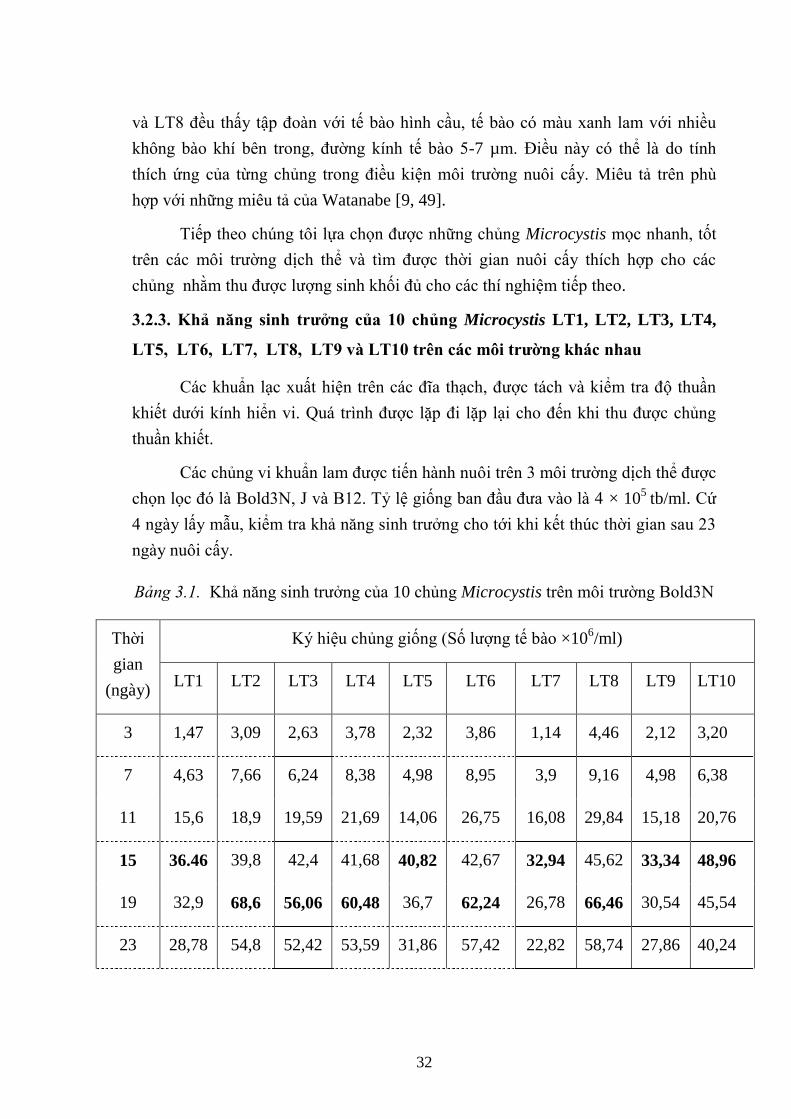
32
và LT8 đều thấy tập đoàn với tế bào hình cầu, tế bào có màu xanh lam với nhiều
không bào khí bên trong, đƣờng kính tế bào 5-7 µm. Điều này có thể là do tính
thích ứng của từng chủng trong điều kiện môi trƣờng nuôi cấy. Miêu tả trên phù
hợp với những miêu tả của Watanabe [9, 49].
Tiếp theo chúng tôi lựa chọn đƣợc những chủng Microcystis mọc nhanh, tốt
trên các môi trƣờng dịch thể và tìm đƣợc thời gian nuôi cấy thích hợp cho các
chủng nhằm thu đƣợc lƣợng sinh khối đủ cho các thí nghiệm tiếp theo.
3.2.3. Khả năng sinh trƣởng của 10 chủng Microcystis LT1, LT2, LT3, LT4,
LT5, LT6, LT7, LT8, LT9 và LT10 trên các môi trƣờng khác nhau
Các khuẩn lạc xuất hiện trên các đĩa thạch, đƣợc tách và kiểm tra độ thuần
khiết dƣới kính hiển vi. Quá trình đƣợc lặp đi lặp lại cho đến khi thu đƣợc chủng
thuần khiết.
Các chủng vi khuẩn lam đƣợc tiến hành nuôi trên 3 môi trƣờng dịch thể đƣợc
chọn lọc đó là Bold3N, J và B12. Tỷ lệ giống ban đầu đƣa vào là 4 × 105
tb/ml. Cứ
4 ngày lấy mẫu, kiểm tra khả năng sinh trƣởng cho tới khi kết thúc thời gian sau 23
ngày nuôi cấy.
Bảng 3.1. Khả năng sinh trƣởng của 10 chủng Microcystis trên môi trƣờng Bold3N
Thời
gian
(ngày)
Ký hiệu chủng giống (Số lƣợng tế bào ×106/ml)
LT1 LT2 LT3 LT4 LT5 LT6 LT7 LT8 LT9 LT10
3 1,47 3,09 2,63 3,78 2,32 3,86 1,14 4,46 2,12 3,20
7 4,63 7,66 6,24 8,38 4,98 8,95 3,9 9,16 4,98 6,38
11 15,6 18,9 19,59 21,69 14,06 26,75 16,08 29,84 15,18 20,76
15 36.46 39,8 42,4 41,68 40,82 42,67 32,94 45,62 33,34 48,96
19 32,9 68,6 56,06 60,48 36,7 62,24 26,78 66,46 30,54 45,54
23 28,78 54,8 52,42 53,59 31,86 57,42 22,82 58,74 27,86 40,24
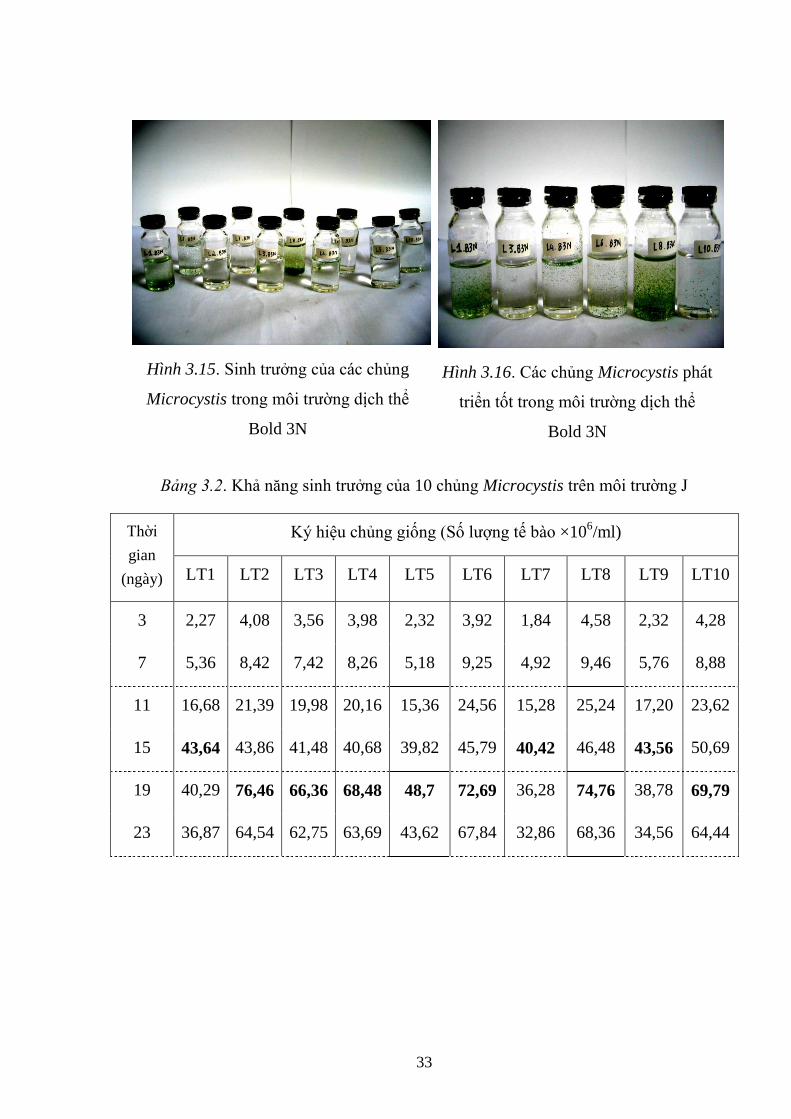
33
Hình 3.15. Sinh trƣởng của các chủng
Microcystis trong môi trƣờng dịch thể
Bold 3N
Hình 3.16. Các chủng Microcystis phát
triển tốt trong môi trƣờng dịch thể
Bold 3N
Bảng 3.2. Khả năng sinh trƣởng của 10 chủng Microcystis trên môi trƣờng J
Thời
gian
(ngày)
Ký hiệu chủng giống (Số lƣợng tế bào ×106/ml)
LT1 LT2 LT3 LT4 LT5 LT6 LT7 LT8 LT9 LT10
3 2,27 4,08 3,56 3,98 2,32 3,92 1,84 4,58 2,32 4,28
7 5,36 8,42 7,42 8,26 5,18 9,25 4,92 9,46 5,76 8,88
11 16,68 21,39 19,98 20,16 15,36 24,56 15,28 25,24 17,20 23,62
15 43,64 43,86 41,48 40,68 39,82 45,79 40,42 46,48 43,56 50,69
19 40,29 76,46 66,36 68,48 48,7 72,69 36,28 74,76 38,78 69,79
23 36,87 64,54 62,75 63,69 43,62 67,84 32,86 68,36 34,56 64,44
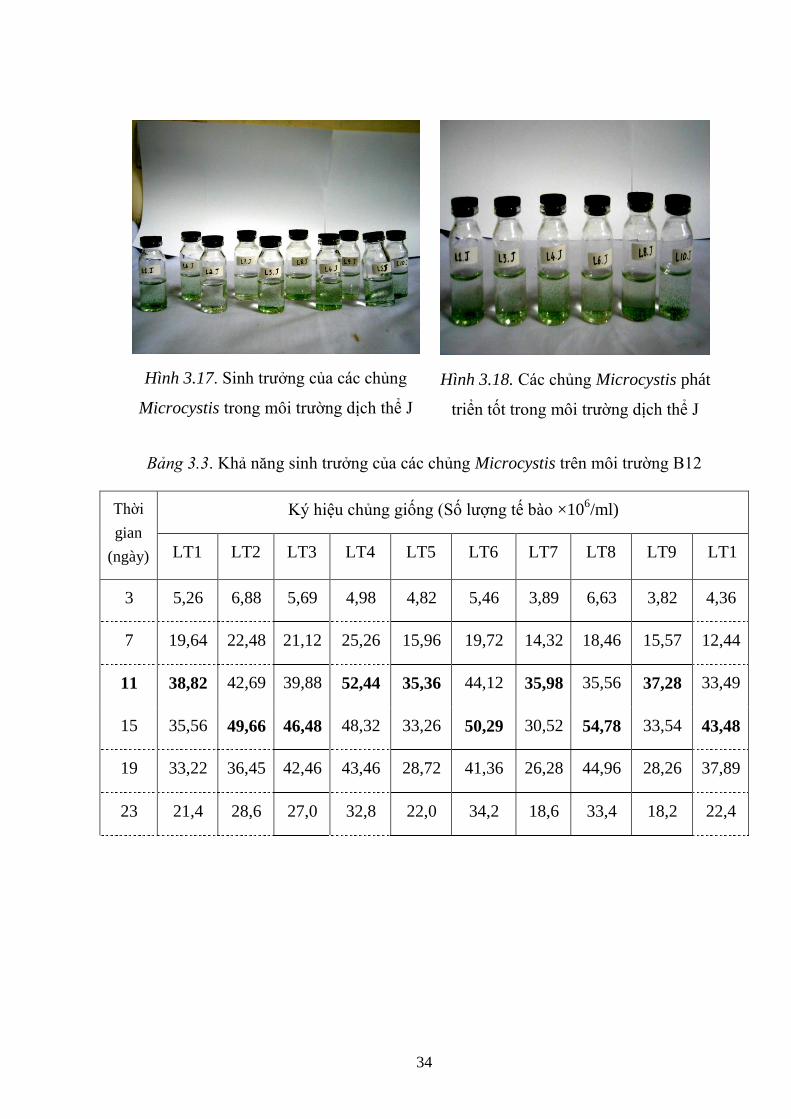
34
Hình 3.17. Sinh trƣởng của các chủng
Microcystis trong môi trƣờng dịch thể J
Hình 3.18. Các chủng Microcystis phát
triển tốt trong môi trƣờng dịch thể J
Bảng 3.3. Khả năng sinh trƣởng của các chủng Microcystis trên môi trƣờng B12
Thời
gian
(ngày)
Ký hiệu chủng giống (Số lƣợng tế bào ×106/ml)
LT1 LT2 LT3 LT4 LT5 LT6 LT7 LT8 LT9 LT1
3 5,26 6,88 5,69 4,98 4,82 5,46 3,89 6,63 3,82 4,36
7 19,64 22,48 21,12 25,26 15,96 19,72 14,32 18,46 15,57 12,44
11 38,82 42,69 39,88 52,44 35,36 44,12 35,98 35,56 37,28 33,49
15 35,56 49,66 46,48 48,32 33,26 50,29 30,52 54,78 33,54 43,48
19 33,22 36,45 42,46 43,46 28,72 41,36 26,28 44,96 28,26 37,89
23 21,4 28,6 27,0 32,8 22,0 34,2 18,6 33,4 18,2 22,4

35
Hình 3.19. Sinh trƣởng của các chủng
Microcystis trong môi trƣờng dịch thể
B12
Hình 3.20. Các chủng Microcystis phát
triển tốt trong môi trƣờng dịch thể B12
Kết quả ở bảng 3.1 cho thấy các chủng Microcystis sinh trƣởng trên môi
trƣờng Bold 3N, mật độ tế bào của các chủng đạt từ 32,94 – 68,6 × 106 sau 15 đến
19 ngày nuôi cấy. Đặc biệt chủng LT2 mật độ tế bào đạt cao nhất 68,6 × 106 /ml
sau ngày thứ 19.
Ở bảng 3.2 cho thấy các chủng Microcystis sinh trƣởng trên môi trƣờng J,
mật độ tế bào của các chủng đạt từ 40,42 – 76,46 × 106 /ml sau 15 đến 19 ngày nuôi
cấy. Trong đó chủng LT2 vẫn phát triển với mật độ tế bào đạt cao nhất 76,46 ×
106 /ml sau ngày thứ 19.
Ở bảng 3.3 cho thấy các chủng Microcystis sinh trƣởng trên môi trƣờng
B12, mật độ tế bào của các chủng đạt từ 35,36 – 54,78 × 106/ml sau 12 đến 15
ngày nuôi cấy. ở môi trƣờng này chủng LT8 phát triển với mật độ tế bào đạt cao
nhất 54,78 × 106/ml sau ngày thứ 15.
Từ những kết quả trên, chúng tôi nhận thấy khả năng sinh trƣởng của các
chủng khác nhau là khác nhau. Hai chủng LT2 và LT8 đều phát triển tốt nhất trên
các môi trƣờng dịch thể và thời gian sinh trƣởng của chúng chỉ trong 15 ngày, do
đó, chúng tôi lựa chọn 2 chủng này để tiếp tục nuôi thu sinh khối cho những nghiên
cứu tiếp theo.
3.2.4. Thành phần acid béo trong tế bào của 2 chủng LT2 và LT8
Bản chất hóa học của các chất dự trữ và thành tế bào của vi tảo đóng vai trò
quan trọng trong việc phân biệt các nhóm vi tảo khác nhau. Trong nghiên cứu này
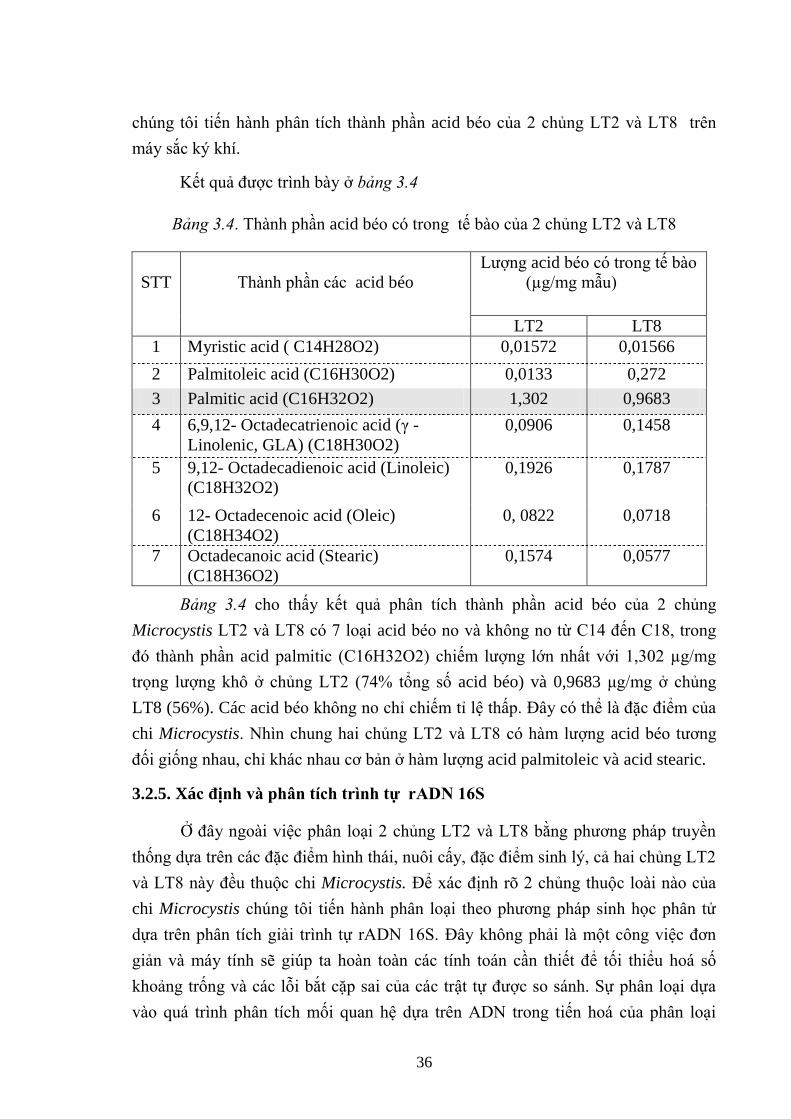
36
chúng tôi tiến hành phân tích thành phần acid béo của 2 chủng LT2 và LT8 trên
máy sắc ký khí.
Kết quả đƣợc trình bày ở bảng 3.4
Bảng 3.4. Thành phần acid béo có trong tế bào của 2 chủng LT2 và LT8
STT
Thành phần các acid béo
Lƣợng acid béo có trong tế bào
(µg/mg mẫu)
LT2 LT8
1 Myristic acid ( C14H28O2) 0,01572 0,01566
2 Palmitoleic acid (C16H30O2) 0,0133 0,272
3 Palmitic acid (C16H32O2) 1,302 0,9683
4 6,9,12- Octadecatrienoic acid (γ -
Linolenic, GLA) (C18H30O2)
0,0906 0,1458
5 9,12- Octadecadienoic acid (Linoleic)
(C18H32O2)
0,1926 0,1787
6 12- Octadecenoic acid (Oleic)
(C18H34O2)
0, 0822 0,0718
7 Octadecanoic acid (Stearic)
(C18H36O2)
0,1574 0,0577
Bảng 3.4 cho thấy kết quả phân tích thành phần acid béo của 2 chủng
Microcystis LT2 và LT8 có 7 loại acid béo no và không no từ C14 đến C18, trong
đó thành phần acid palmitic (C16H32O2) chiếm lƣợng lớn nhất với 1,302 µg/mg
trọng lƣợng khô ở chủng LT2 (74% tổng số acid béo) và 0,9683 μg/mg ở chủng
LT8 (56%). Các acid béo không no chỉ chiếm tỉ lệ thấp. Đây có thể là đặc điểm của
chi Microcystis. Nhìn chung hai chủng LT2 và LT8 có hàm lƣợng acid béo tƣơng
đối giống nhau, chỉ khác nhau cơ bản ở hàm lƣợng acid palmitoleic và acid stearic.
3.2.5. Xác định và phân tích trình tự rADN 16S
Ở đây ngoài việc phân loại 2 chủng LT2 và LT8 bằng phƣơng pháp truyền
thống dựa trên các đặc điểm hình thái, nuôi cấy, đặc điểm sinh lý, cả hai chủng LT2
và LT8 này đều thuộc chi Microcystis. Để xác định rõ 2 chủng thuộc loài nào của
chi Microcystis chúng tôi tiến hành phân loại theo phƣơng pháp sinh học phân tử
dựa trên phân tích giải trình tự rADN 16S. Đây không phải là một công việc đơn
giản và máy tính sẽ giúp ta hoàn toàn các tính toán cần thiết để tối thiểu hoá số
khoảng trống và các lỗi bắt cặp sai của các trật tự đƣợc so sánh. Sự phân loại dựa
vào quá trình phân tích mối quan hệ dựa trên ADN trong tiến hoá của phân loại

37
chủng loại phát sinh hoặc dựa vào những đặc điểm tƣơng tự nhau trong phân loại
hình thái đều đƣợc thể hiện ở những sơ đồ dạng cây gọi là cây phả hệ. Cây phả hệ
cho phép nhận biết mối quan hệ di truyền giữa các loài và giữa các chủng cùng
loài. Sử dụng Cluxtal 18.1 và chƣơng trình điện toán MEGA 2.1 cùng các thông số
tiến hoá cần thiết cho thấy mối quan hệ đã đƣợc thiết lập trên cơ sở phân tích phân
đoạn, so sánh giữa các vùng bảo toàn của những đoạn gen rADN 16S.
Hình 3.21. Cây phả hệ dựa trên phân tích giải trình tự rADN 16S của 2 chủng LT2,
LT8 và các loài có quan hệ họ hàng gần
Cây phả hệ (hình 3.21) cho thấy mối quan hệ gần giữa chủng LT2 (đoạn gen
456bp) và LT8 (đoạn gen 457bp) với 13 trật tự nucleotit của các chủng thuộc
Cyanobacteria trong ngân hàng dữ liệu gen. Chúng nằm trên cùng một nhánh và
cùng nhóm với loài Microcystis aeruginosa EU078503 với độ tƣơng đồng là 100%
(chủng LT2) và 99% (chủng LT8).
Cây phả hệ vẽ theo phƣơng pháp neighbour joining thƣớc đo = 0,05 Knor
trong trình tự nucleotid. Các giá trị hiển thị tại các nhánh cây là kết quả phân tích
boostrap với độ lập lại 1000 (với những giá trị >50% mới đƣợc thể hiển trên cây).
Sự khác biệt giữa 2 chủng là không đáng kể và nằm trong phạm vi khác biệt
cùng loài (conspecific variation). Dựa vào các kết quả phân tích đặc điểm sinh lý,
Lyngbya majuscula AF510964
Lyngbya majuscula AF510965
Anabaena circinalis EU780160
Anabaena circinalis EU780161
Anabaena circinalis EU078518 88
100
100
Microcystis aeruginosa AB023261
96
Microcystis ichthyoblabe AB012338
Microcystis viridis AY121357
Microcystis viridis AY065976
100
Microcystis wesenbergii AB038538
100
LT8
LT2
Microcystis aeruginosa EU078503
100
Microcystis aeruginosa EU078504
100
100
87
73
0.005
100 Microcystis aeruginosa EU078505

38
hoá và cây phả hệ cho thấy chúng có nhiều đặc điểm chung vì vậy 2 chủng LT2 và
LT8 đều thuộc loài Microcystis aeruginosa.
3.3. TÁCH CHIẾT ĐỘC TỐ MICROCYSTIN
Chúng tôi tiến hành tách chiết độc tố microcystin theo phƣơng pháp nhƣ đã
miêu tả ở phần phƣơng pháp (mục 2.2.5).
3.3.1. Sắc ký bản mỏng (TLC)
Hình 3.22. Kết quả chạy TLC
trƣớc khi nhuộm nihyđrin
Hình 3.23. Kết quả chạy TLC sau khi nhuộm
nihyđrin
Từ kết quả ở hình 3.22 và 3.23 ta thấy trên bản silicagel chạy mẫu tách chiết
microcystin xuất hiện các băng tƣơng đồng với băng MC chuẩn. Điều đó chứng tỏ
sự có mặt của độc tố trong mẫu chiết. Dựa vào kết quả này, chúng tôi tiến hành
phân tích và xác định hàm lƣợng microcystin trong dịch chiết trên máy quang phổ
và sắc ký lỏng cao áo HPLC.
3.3.2. Xác định hàm lƣợng microcystin trên máy quang phổ
Các băng chứa microcystin của dịch chiết chạy trên sắc ký bản mỏng đƣợc
cạo và tái chiết rút bằng methanol 80%. Sau đó lọc qua cột C18, lấy 1ml dịch lọc,
pha loãng 7000, 8000, 9000 lần. Đem các dung dịch đó đo trên máy quang phổ ở
bƣớc sóng 242 nm.
Kết quả đƣợc trình bày ở hình 3.24
Microcystin
chuẩn
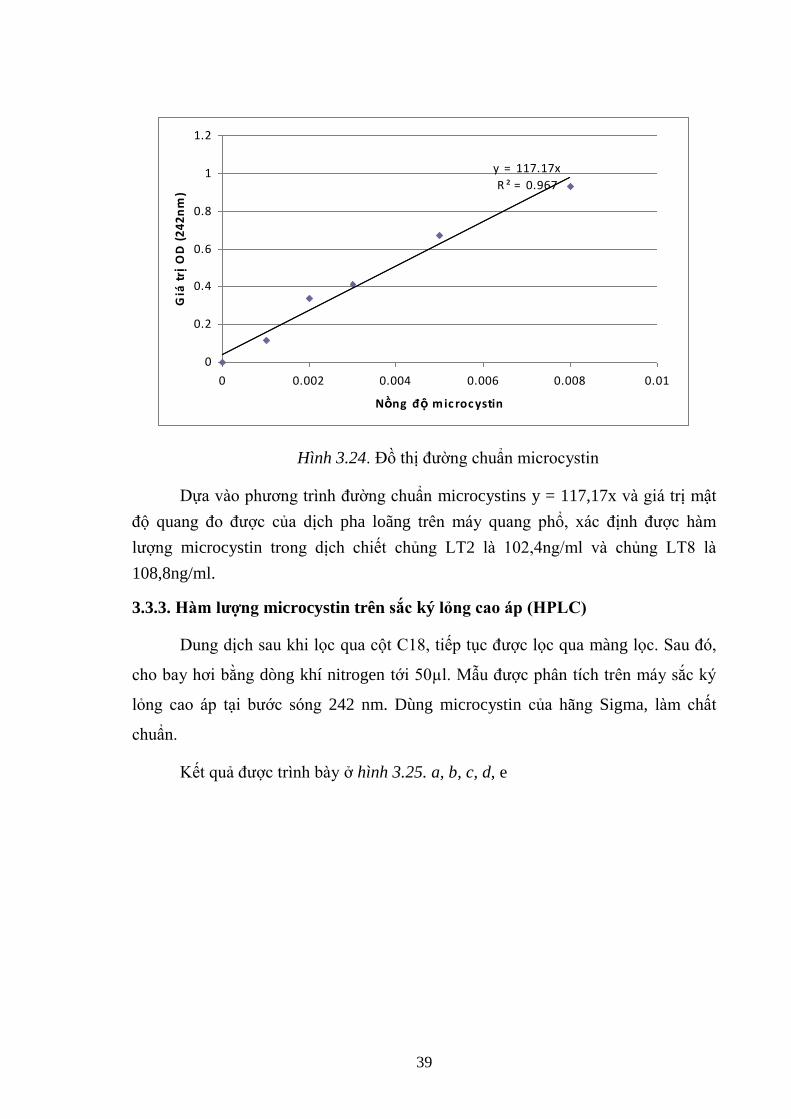
39
y = 117.17x
R ² = 0.967
0
0.2
0.4
0.6
0.8
1
1.2
0 0.002 0.004 0.006 0.008 0.01
Nồng độ mic roc ystin
Giá
trị
OD
(2
42
nm
)
Hình 3.24. Đồ thị đƣờng chuẩn microcystin
Dựa vào phƣơng trình đƣờng chuẩn microcystins y = 117,17x và giá trị mật
độ quang đo đƣợc của dịch pha loãng trên máy quang phổ, xác định đƣợc hàm
lƣợng microcystin trong dịch chiết chủng LT2 là 102,4ng/ml và chủng LT8 là
108,8ng/ml.
3.3.3. Hàm lƣợng microcystin trên sắc ký lỏng cao áp (HPLC)
Dung dịch sau khi lọc qua cột C18, tiếp tục đƣợc lọc qua màng lọc. Sau đó,
cho bay hơi bằng dòng khí nitrogen tới 50µl. Mẫu đƣợc phân tích trên máy sắc ký
lỏng cao áp tại bƣớc sóng 242 nm. Dùng microcystin của hãng Sigma, làm chất
chuẩn.
Kết quả đƣợc trình bày ở hình 3.25. a, b, c, d, e
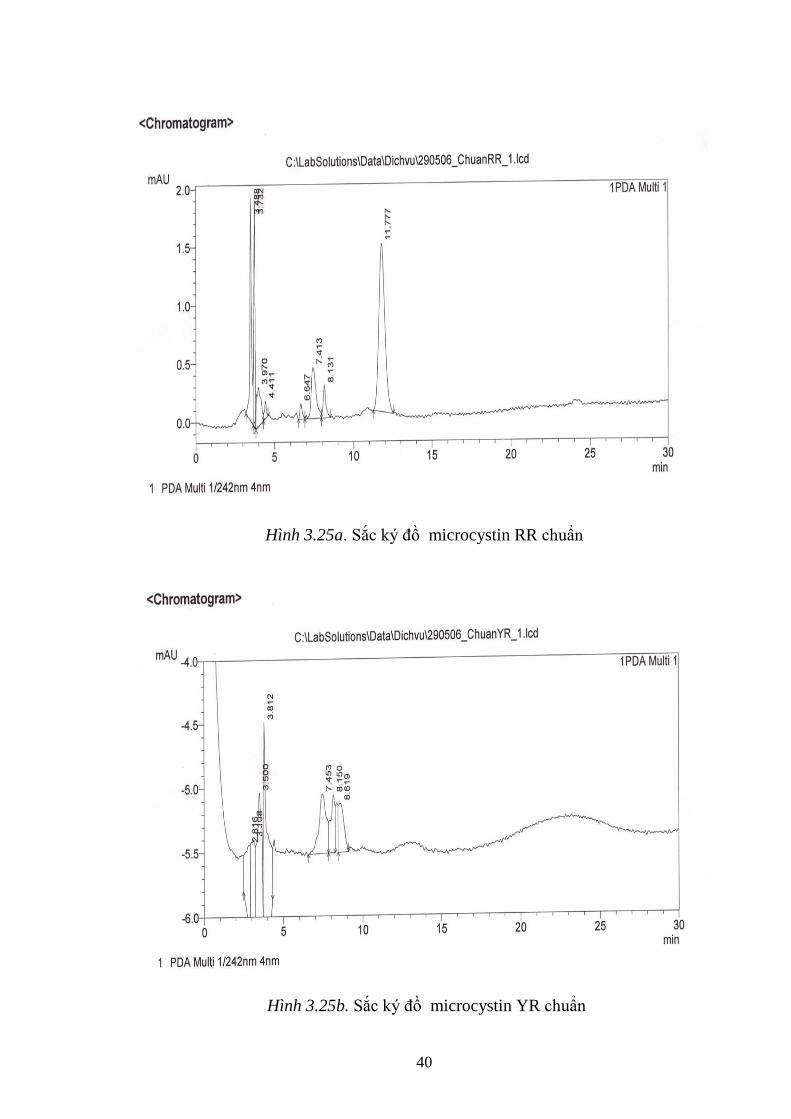
40
Hình 3.25a. Sắc ký đồ microcystin RR chuẩn
Hình 3.25b. Sắc ký đồ microcystin YR chuẩn
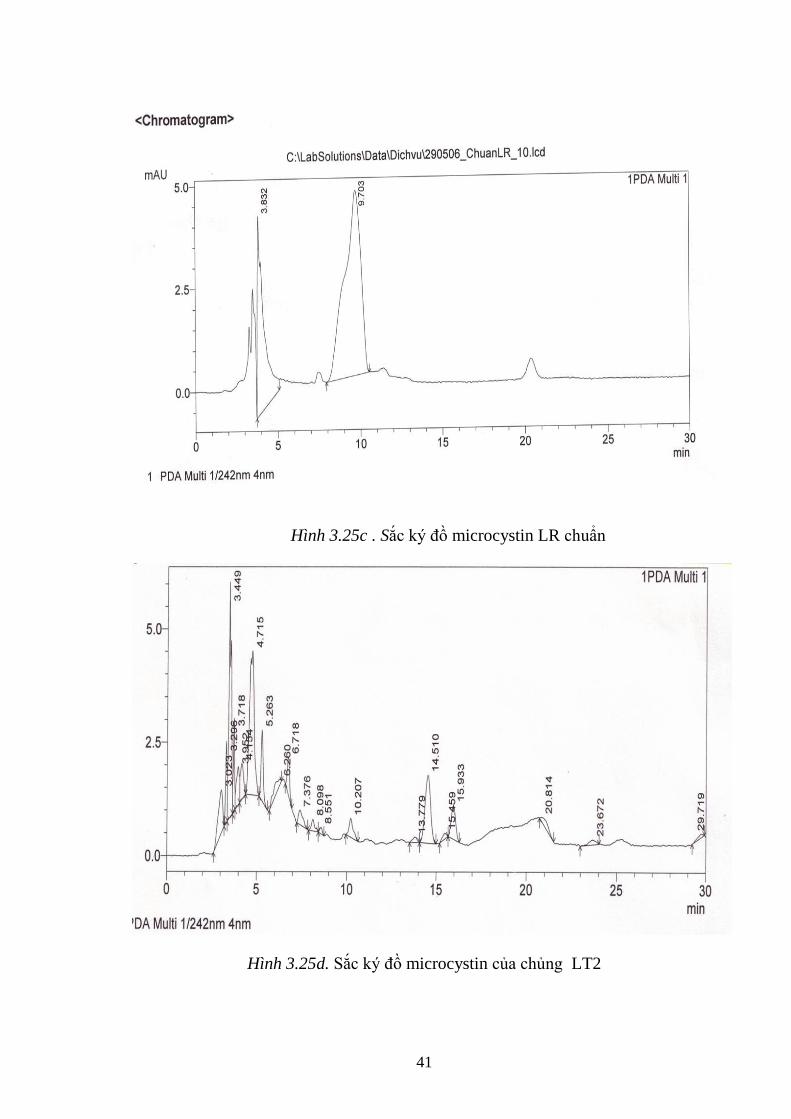
41
Hình 3.25c . Sắc ký đồ microcystin LR chuẩn
Hình 3.25d. Sắc ký đồ microcystin của chủng LT2
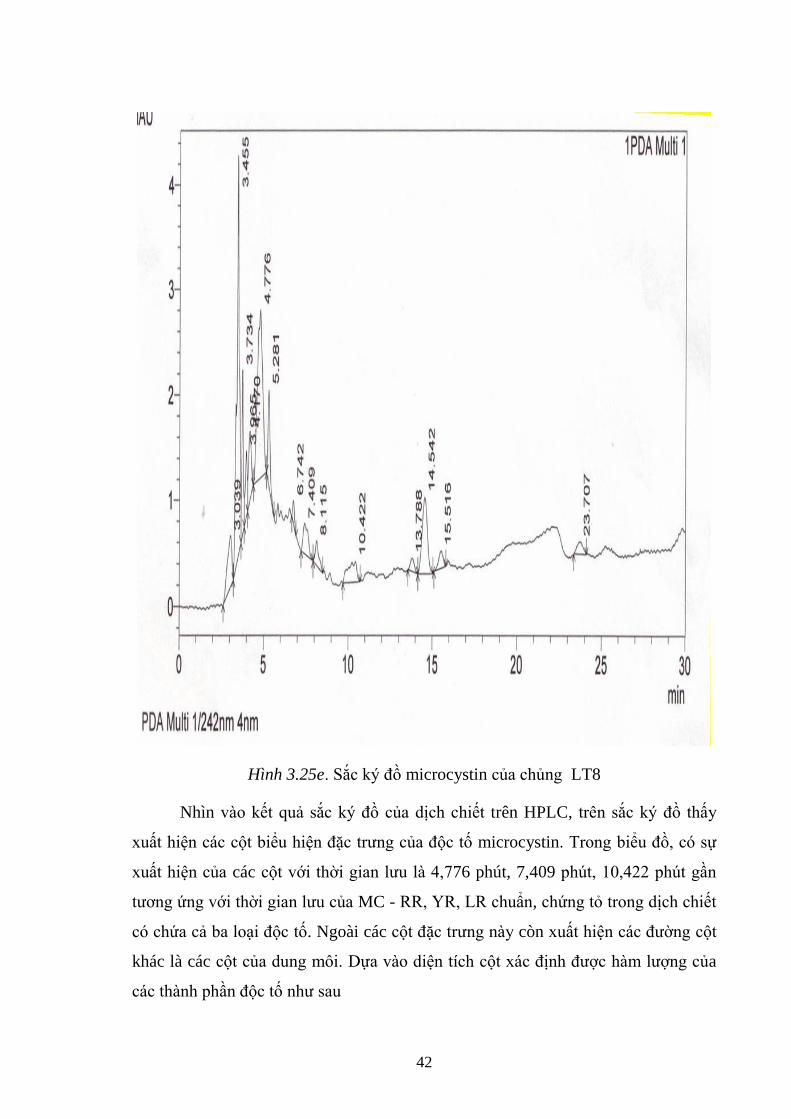
42
Hình 3.25e. Sắc ký đồ microcystin của chủng LT8
Nhìn vào kết quả sắc ký đồ của dịch chiết trên HPLC, trên sắc ký đồ thấy
xuất hiện các cột biểu hiện đặc trƣng của độc tố microcystin. Trong biểu đồ, có sự
xuất hiện của các cột với thời gian lƣu là 4,776 phút, 7,409 phút, 10,422 phút gần
tƣơng ứng với thời gian lƣu của MC - RR, YR, LR chuẩn, chứng tỏ trong dịch chiết
có chứa cả ba loại độc tố. Ngoài các cột đặc trƣng này còn xuất hiện các đƣờng cột
khác là các cột của dung môi. Dựa vào diện tích cột xác định đƣợc hàm lƣợng của
các thành phần độc tố nhƣ sau
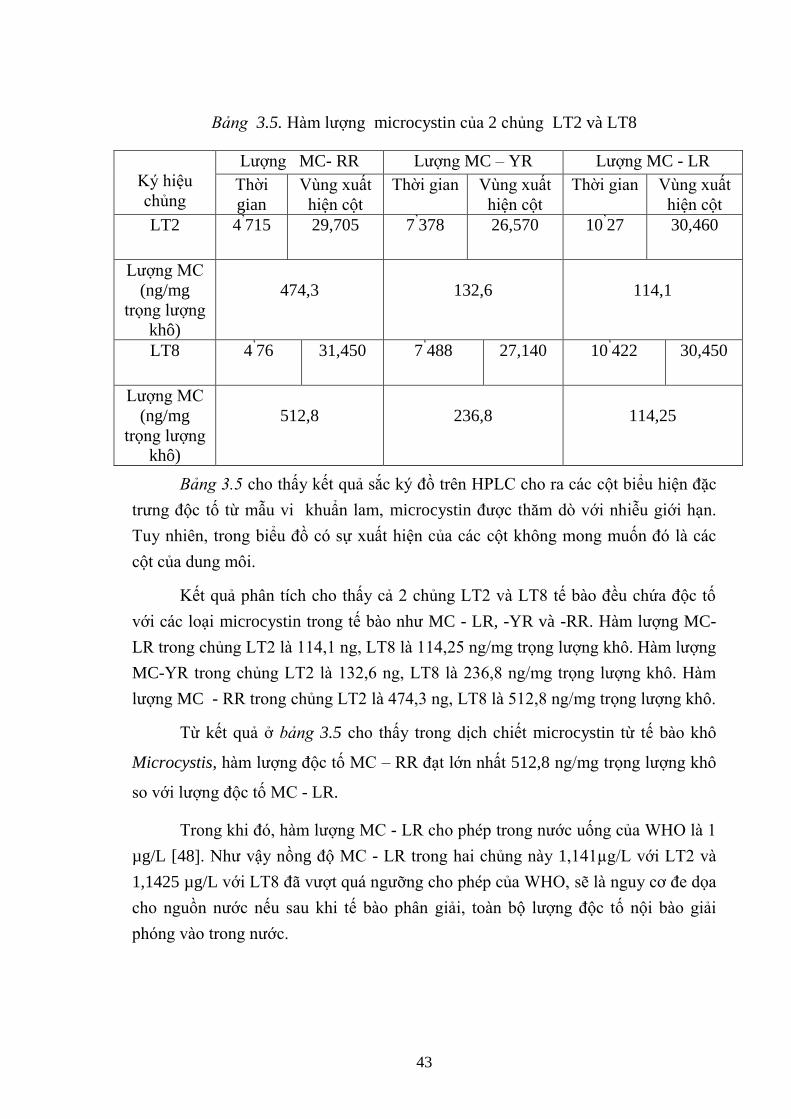
43
Bảng 3.5. Hàm lƣợng microcystin của 2 chủng LT2 và LT8
Ký hiệu
chủng
Lƣợng MC- RR Lƣợng MC – YR Lƣợng MC - LR
Thời
gian
Vùng xuất
hiện cột
Thời gian Vùng xuất
hiện cột
Thời gian Vùng xuất
hiện cột
LT2 4‟715 29,705 7
‟378 26,570 10
‟27 30,460
Lƣợng MC
(ng/mg
trọng lƣợng
khô)
474,3
132,6
114,1
LT8 4‟76 31,450 7
‟488 27,140 10
‟422 30,450
Lƣợng MC
(ng/mg
trọng lƣợng
khô)
512,8
236,8
114,25
Bảng 3.5 cho thấy kết quả sắc ký đồ trên HPLC cho ra các cột biểu hiện đặc
trƣng độc tố từ mẫu vi khuẩn lam, microcystin đƣợc thăm dò với nhiễu giới hạn.
Tuy nhiên, trong biểu đồ có sự xuất hiện của các cột không mong muốn đó là các
cột của dung môi.
Kết quả phân tích cho thấy cả 2 chủng LT2 và LT8 tế bào đều chứa độc tố
với các loại microcystin trong tế bào nhƣ MC - LR, -YR và -RR. Hàm lƣợng MC-
LR trong chủng LT2 là 114,1 ng, LT8 là 114,25 ng/mg trọng lƣợng khô. Hàm lƣợng
MC-YR trong chủng LT2 là 132,6 ng, LT8 là 236,8 ng/mg trọng lƣợng khô. Hàm
lƣợng MC - RR trong chủng LT2 là 474,3 ng, LT8 là 512,8 ng/mg trọng lƣợng khô.
Từ kết quả ở bảng 3.5 cho thấy trong dịch chiết microcystin từ tế bào khô
Microcystis, hàm lƣợng độc tố MC – RR đạt lớn nhất 512,8 ng/mg trọng lƣợng khô
so với lƣợng độc tố MC - LR.
Trong khi đó, hàm lƣợng MC - LR cho phép trong nƣớc uống của WHO là 1
µg/L [48]. Nhƣ vậy nồng độ MC - LR trong hai chủng này 1,141µg/L với LT2 và
1,1425 µg/L với LT8 đã vƣợt quá ngƣỡng cho phép của WHO, sẽ là nguy cơ đe dọa
cho nguồn nƣớc nếu sau khi tế bào phân giải, toàn bộ lƣợng độc tố nội bào giải
phóng vào trong nƣớc.

44
3.4. PHÂN LẬP VÀ TUYỂN CHỌN CÁC CHỦNG VI KHUẨN NƢỚC CÓ
KHẢ NĂNG PHÂN GIẢI MICROCYSTIN
3.4.1. Phân lập các chủng vi khuẩn nƣớc
Do tầm quan trọng và mức độ gây độc của các vi khuẩn lam nở hoa trong các
thuỷ vực ngày càng gia tăng nên các nghiên cứu về độ bền vững và phân giải độc tố
của chúng trở nên rất cần thiết. Trong điều kiện tự nhiên, các vi sinh vật, đặc biệt
các vi khuẩn nƣớc đóng vai trò quan trọng trong việc phân giải độc tố không hoà
tan đƣợc giải phóng vào trong nƣớc.
Ngƣời ta đã chứng minh rằng microcystin có thể bị phân giải bởi các vi sinh
vật có trong hệ sinh thái thuỷ vực tự nhiên [28]. Ngoài ra có các bằng chứng cho
rằng sự phân giải sinh học microcystin xảy ra nhanh hơn ở các thuỷ vực thƣờng bị
tảo lam độc nở hoa [49, 51]. Ngày nay khi hiện tƣợng nở hoa của tảo lam độc trong
các thuỷ vực trở nên nghiêm trọng thì các nghiên cứu tập trung vào vấn đề vi khuẩn
nƣớc là cách để giảm thiểu tác hại của vi khuẩn lam và độc tố của chúng. Do vậy,
các chủng vi khuẩn nƣớc đƣợc tuyển chọn và phân lập từ 6 địa điểm với sự xuất
hiện Microcystis theo tỷ lệ 1:1 (tính theo ml) đƣợc nuôi cấy làm giàu trong các ống
nghiệm dung tích 10 ml chứa 3 ml môi trƣờng NA, M7 [44].
Sau đó đem nuôi giữ trong điều kiện nhiệt độ 30oC dƣới ánh sáng đèn neon
yếu với cƣờng độ ánh sáng là 1000 lux theo quang chu kỳ là 12 giờ chiếu sáng : 12
giờ tối. Sau 24 nuôi cấy, đem ly tâm dịch thể với tốc độ 7000 vòng/10 phút thu sinh
khối. Rửa sinh khối 3 lần bằng dung dịch NA, M7 vô trùng. Sau đó đem nuôi cấy
lại trong thời gian 24 giờ dƣới các điều kiện đã mô tả ở trên (để kích thích các
chủng vi khuẩn thích ứng với sự phân giải tế bào M. aeruginosa và phân giải độc tố
microcystin).

45
Hình 3.26. Nuôi cấy làm giàu các chủng vi
khuẩn nƣớc trên môi trƣờng NA và M7
Hình 3.27. Cấy pha khuẩn lạc vi
khuẩn trên môi trƣờng NA
Hình 3.26 và 3.27 cho thấy sau 24 giờ nuôi cấy làm giàu trên 2 môi trƣờng.
Chúng tôi nhận thấy màu của dịch nuôi cấy xanh trên môi trƣờng MA và còn trên
môi trƣờng M7 không thấy xuẩt hiện màu xanh. Điều này cho thấy môi trƣờng.
MA thích hợp cho việc phân lập và nuôi cấy các chủng vi khuẩn nƣớc. Các mẫu
tiếp tục đƣợc cấy ria trên môi trƣờng thạch NA. Sau 12 giờ quan sát sự xuất hiện
các khuẩn lạc có tiết sắc tố xanh vào trong môi trƣờng. Tách các khuẩn lạc đơn cấy
ria trên môi trƣờng thạch nhiều lần cho đến khi tách đƣợc chủng thuần khiết. Trong
thí nghiệm này chúng tôi chọn đƣợc 2 chủng vi khuẩn có ký hiệu N4 và N7 có khả
năng sinh trƣởng tốt chỉ sau 12 giờ đã tiết sắc tố xanh vào môi trƣờng.
Nuôi giữ giống gốc vi khuẩn trên môi trƣờng thạch nghiêng (môi trƣờng
NA) và bảo quản ở nhiệt độ 4oC, dùng cho các nghiên cứu tiếp theo.
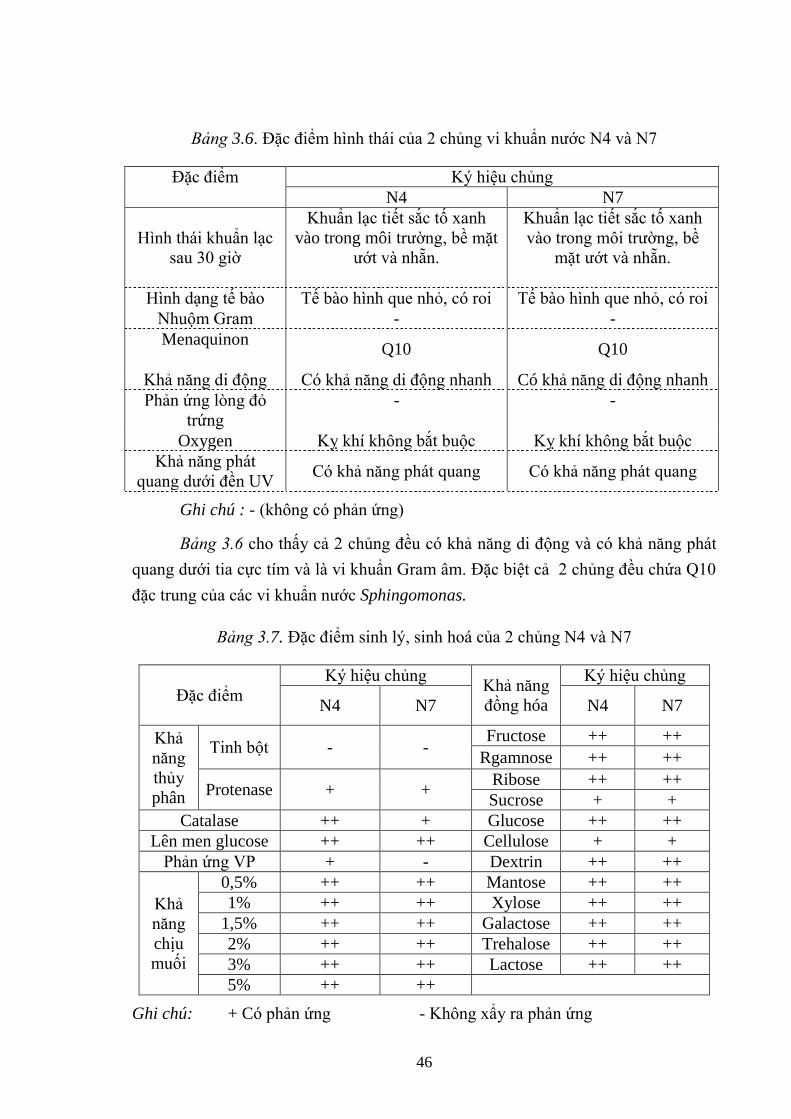
46
Bảng 3.6. Đặc điểm hình thái của 2 chủng vi khuẩn nƣớc N4 và N7
Đặc điểm Ký hiệu chủng
N4 N7
Hình thái khuẩn lạc
sau 30 giờ
Khuẩn lạc tiết sắc tố xanh
vào trong môi trƣờng, bề mặt
ƣớt và nhẵn.
Khuẩn lạc tiết sắc tố xanh
vào trong môi trƣờng, bề
mặt ƣớt và nhẵn.
Hình dạng tế bào Tế bào hình que nhỏ, có roi Tế bào hình que nhỏ, có roi
Nhuộm Gram - -
Menaquinon
Q10 Q10
Khả năng di động Có khả năng di động nhanh Có khả năng di động nhanh
Phản ứng lòng đỏ
trứng
- -
Oxygen Kỵ khí không bắt buộc Kỵ khí không bắt buộc
Khả năng phát
quang dƣới đền UV Có khả năng phát quang Có khả năng phát quang
Ghi chú : - (không có phản ứng)
Bảng 3.6 cho thấy cả 2 chủng đều có khả năng di động và có khả năng phát
quang dƣới tia cực tím và là vi khuẩn Gram âm. Đặc biệt cả 2 chủng đều chứa Q10
đặc trung của các vi khuẩn nƣớc Sphingomonas.
Bảng 3.7. Đặc điểm sinh lý, sinh hoá của 2 chủng N4 và N7
Đặc điểm
Ký hiệu chủng Khả năng
đồng hóa
Ký hiệu chủng
N4 N7 N4 N7
Khả
năng
thủy
phân
Tinh bột - - Fructose ++ ++
Rgamnose ++ ++
Protenase + + Ribose ++ ++
Sucrose + +
Catalase ++ + Glucose ++ ++
Lên men glucose ++ ++ Cellulose + +
Phản ứng VP + - Dextrin ++ ++
Khả
năng
chịu
muối
0,5% ++ ++ Mantose ++ ++
1% ++ ++ Xylose ++ ++
1,5% ++ ++ Galactose ++ ++
2% ++ ++ Trehalose ++ ++
3% ++ ++ Lactose ++ ++
5% ++ ++
Ghi chú: + Có phản ứng - Không xẩy ra phản ứng

47
Bảng 3.7 cho thấy 2 chủng vi khuẩn N4 và N7 đều không khả năng phân giải
tinh bột. Cả 2 chủng đều có khả năng phân hủy protein, nghĩa là có hoạt tính
protease. Ngoài ra cả 2 chủng này đều cho hoạt tính catalase rất mạnh. Chúng cũng
có khả năng chịu muối tốt từ 0,5- 5% và đồng hóa đƣợc hầu hết các loại đƣờng.
Điều này hoàn toàn phù hợp với nghiên cứu của David mô tả các chủng vi khuẩn
Sphingomonas phân giải độc tố MLR [20].
3.4.2. Xác định và phân tích trình tự rADN 16S 2 chủng N4 và N7
Để có đƣợc nhiều hơn nữa thông tin xác định về vị trí phân loại và phát sinh
chủng loại của 2 chủng N4, N7. ADN genom của 2 chủng N4 và N7 đƣợc dùng làm
khuôn để khuyếch đại đoạn ADN bằng phản ứng PCR, sử dụng cặp mồi 27f và
1252r. Chúng tôi đã xác định trình tự rADN 16S của nó và so sánh với trình tự có
sẵn trong Ribosomal Data Project. Tìm kiếm trình tự tƣơng đồng trực tuyến chỉ ra
rằng, trong số các loài đã công bố, trình tự rADN 16S (đoạn 700bp) của N4 giống
trình tự của Sphingomonas mali Y09638 (99%), Sphingomonas asaccharolytica
Y09639 (99%). Với chủng N7 (đoạn 564bp) giống Sphingomonas asaccharolytica
Y09639 (100%) và Sphingomonas mali Y09638 (99%).
Khoảng cách tiến hóa đƣợc tính toán để thiết lập cơ sở dữ liệu bao gồm trình
tự của chủng N4, N7 và 27 trình tự khác của nhóm Sphingomonas phân lớp alpha
của Proteobacteria với Rhodospirillum rubrum là outgroup (ngoài nhóm).
Cây phát sinh chủng loại đã đƣợc xây dựng lại trên cơ sở các số liệu ma trận
về khoảng cách thu đƣợc nhƣ (hình 3.28, 3.29). Chủng N4, N7 phân nhánh sâu từ 1
nhóm chính, nhóm này bao gồm một số thành viên của chi Sphingomonas. Các loài
đã công bố của chi Sphingomonas khác nhau về phát sinh chủng loại
Hai cây phả hệ vẽ theo phƣơng pháp neighbour joining thƣớc đo = 0,02 Knor
trong trình tự nucleotid. Các giá trị hiển thị tại các nhánh cây là kết quả phân tích
boostrap với độ lập lại 1000 (với những giá trị >50% mới đƣợc thể hiện trên cây
phả hệ).
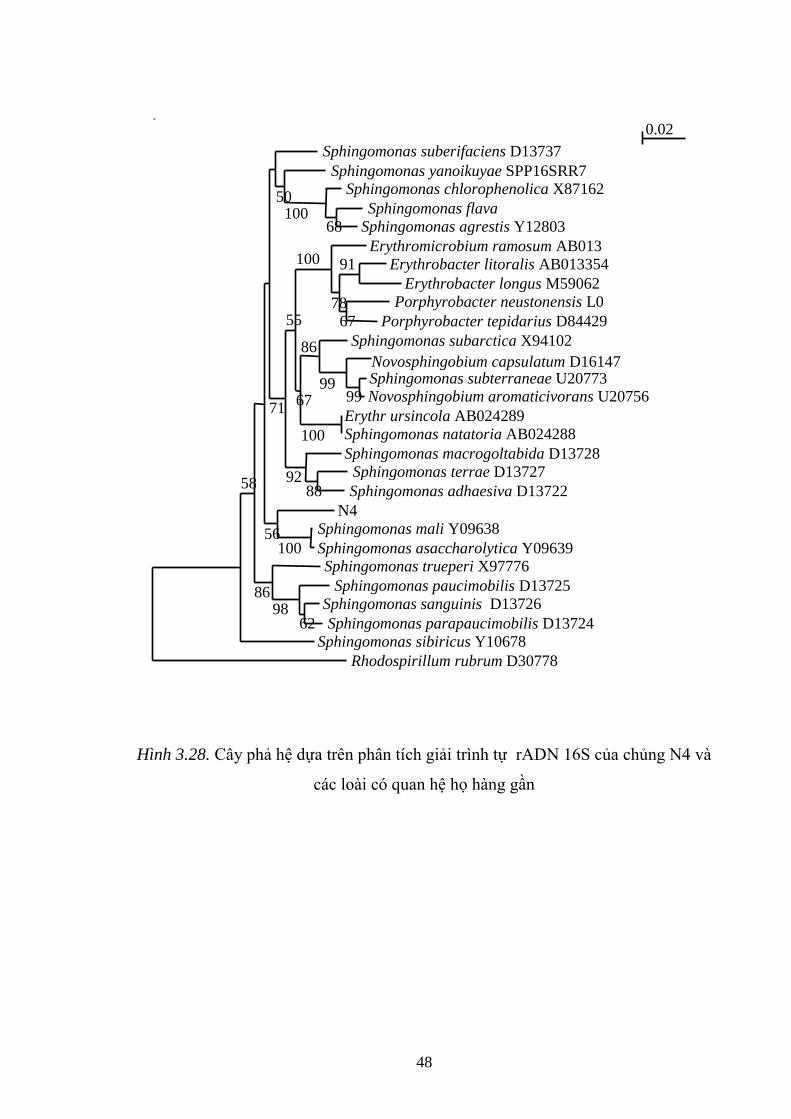
48
Hình 3.28. Cây phả hệ dựa trên phân tích giải trình tự rADN 16S của chủng N4 và
các loài có quan hệ họ hàng gần
Rhodospirillum rubrum D30778
Sphingomonas sibiricus Y10678
Sphingomonas parapaucimobilis D13724
Sphingomonas sanguinis D13726
Sphingomonas paucimobilis D13725
62
Sphingomonas trueperi X97776
98
Sphingomonas asaccharolytica Y09639
Sphingomonas mali Y09638
N4
100
Sphingomonas adhaesiva D13722
Sphingomonas terrae D13727
Sphingomonas macrogoltabida D13728
88
Sphingomonas natatoria AB024288
Erythr ursincola AB024289
Novosphingobium aromaticivorans U20756 Sphingomonas subterraneae U20773 Novosphingobium capsulatum D16147
99
Sphingomonas subarctica X94102
99
100
86
Porphyrobacter tepidarius D84429
Porphyrobacter neustonensis L0
Erythrobacter longus M59062
Erythrobacter litoralis AB013354
67
91
Erythromicrobium ramosum AB013
78
67
100
92
55
Sphingomonas agrestis Y12803
Sphingomonas flava
X87164
Sphingomonas chlorophenolica X87162
68
Sphingomonas yanoikuyae SPP16SRR7
100
Sphingomonas suberifaciens D13737
50
71
56
86
58
0.02
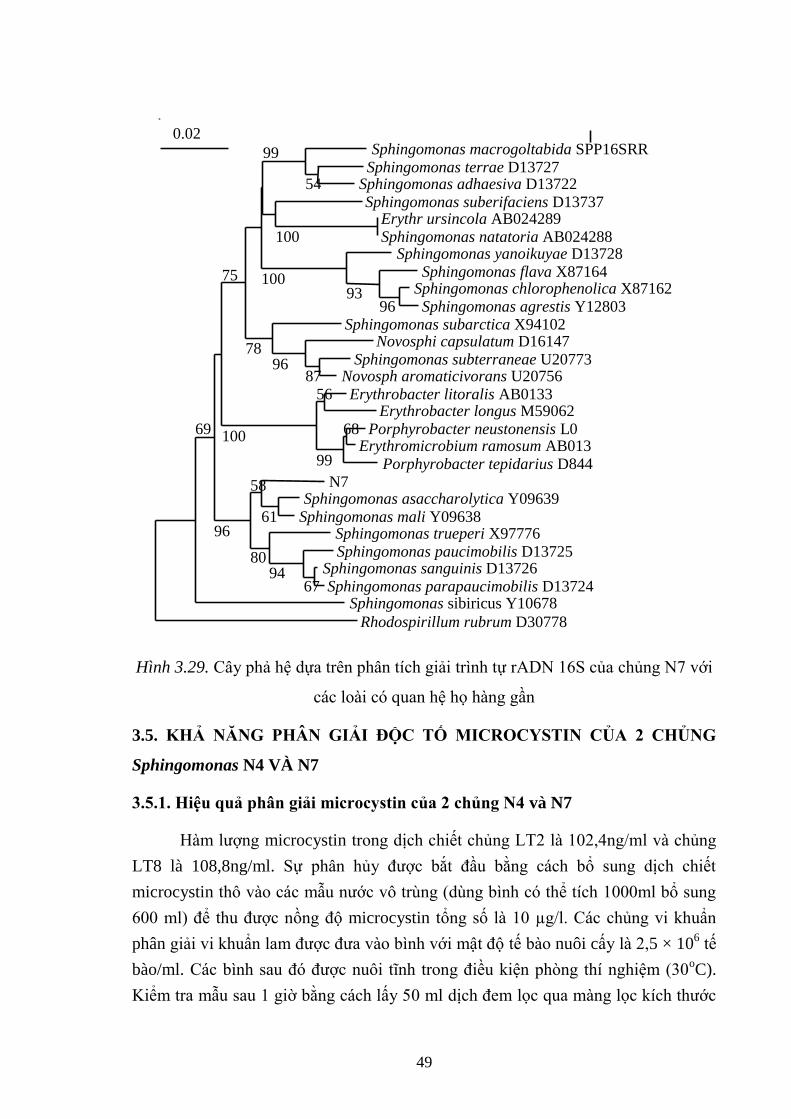
49
Hình 3.29. Cây phả hệ dựa trên phân tích giải trình tự rADN 16S của chủng N7 với
các loài có quan hệ họ hàng gần
3.5. KHẢ NĂNG PHÂN GIẢI ĐỘC TỐ MICROCYSTIN CỦA 2 CHỦNG
Sphingomonas N4 VÀ N7
3.5.1. Hiệu quả phân giải microcystin của 2 chủng N4 và N7
Hàm lƣợng microcystin trong dịch chiết chủng LT2 là 102,4ng/ml và chủng
LT8 là 108,8ng/ml. Sự phân hủy đƣợc bắt đầu bằng cách bổ sung dịch chiết
microcystin thô vào các mẫu nƣớc vô trùng (dùng bình có thể tích 1000ml bổ sung
600 ml) để thu đƣợc nồng độ microcystin tổng số là 10 µg/l. Các chủng vi khuẩn
phân giải vi khuẩn lam đƣợc đƣa vào bình với mật độ tế bào nuôi cấy là 2,5 × 106 tế
bào/ml. Các bình sau đó đƣợc nuôi tĩnh trong điều kiện phòng thí nghiệm (30oC).
Kiểm tra mẫu sau 1 giờ bằng cách lấy 50 ml dịch đem lọc qua màng lọc kích thƣớc
Rhodospirillum rubrum D30778
Sphingomonas sibiricus Y10678 Sphingomonas parapaucimobilis D13724
Sphingomonas sanguinis D13726 Sphingomonas paucimobilis D13725
67
Sphingomonas trueperi X97776
94
Sphingomonas mali Y09638 Sphingomonas asaccharolytica Y09639
N7
61
80
58
Porphyrobacter tepidarius D844 Erythromicrobium ramosum AB013
Porphyrobacter neustonensis L0 68 Erythrobacter longus M59062
Erythrobacter litoralis AB0133
99
56 Novosph aromaticivorans U20756
Sphingomonas subterraneae U20773 Novosphi capsulatum D16147
87
Sphingomonas subarctica X94102
96
Sphingomonas agrestis Y12803 Sphingomonas chlorophenolica X87162 Sphingomonas flava X87164
96
Sphingomonas yanoikuyae D13728
93
Sphingomonas natatoria AB024288 Erythr ursincola AB024289
Sphingomonas suberifaciens D13737
100
Sphingomonas adhaesiva D13722 Sphingomonas terrae D13727 Sphingomonas macrogoltabida SPP16SRR
54
99
100
78
100
75
96
69
0.02
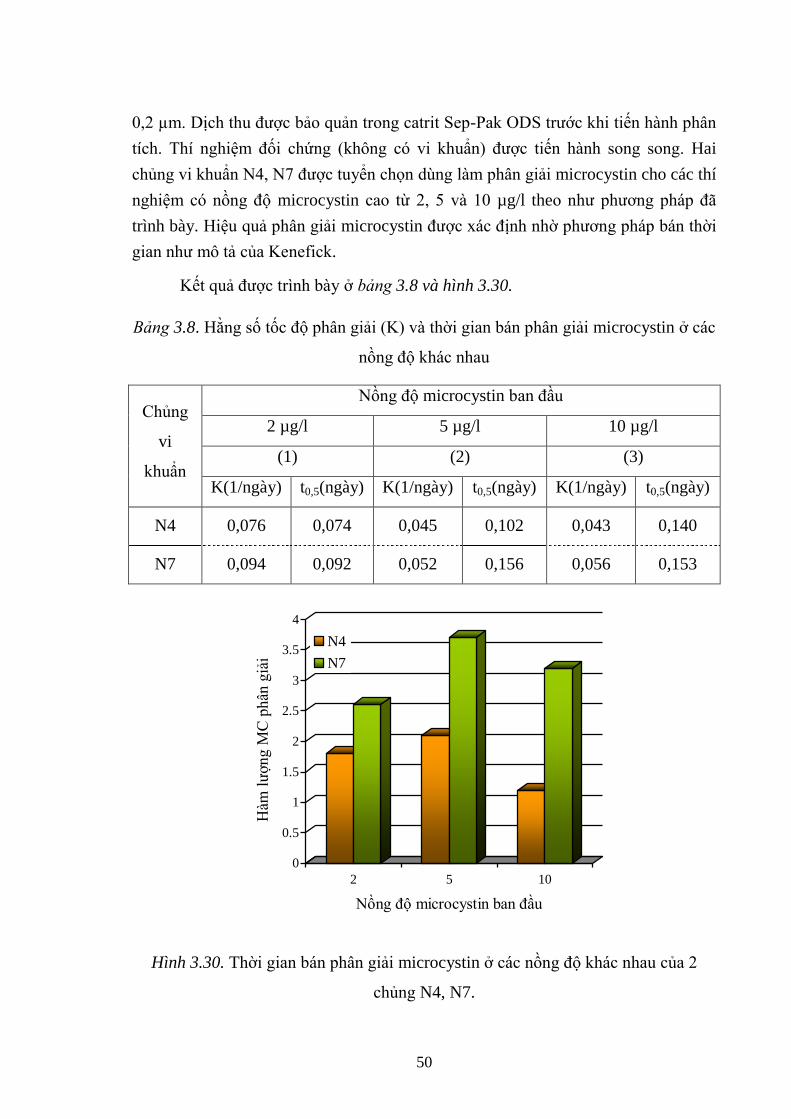
50
0,2 µm. Dịch thu đƣợc bảo quản trong catrit Sep-Pak ODS trƣớc khi tiến hành phân
tích. Thí nghiệm đối chứng (không có vi khuẩn) đƣợc tiến hành song song. Hai
chủng vi khuẩn N4, N7 đƣợc tuyển chọn dùng làm phân giải microcystin cho các thí
nghiệm có nồng độ microcystin cao từ 2, 5 và 10 µg/l theo nhƣ phƣơng pháp đã
trình bày. Hiệu quả phân giải microcystin đƣợc xác định nhờ phƣơng pháp bán thời
gian nhƣ mô tả của Kenefick.
Kết quả đƣợc trình bày ở bảng 3.8 và hình 3.30.
Bảng 3.8. Hằng số tốc độ phân giải (K) và thời gian bán phân giải microcystin ở các
nồng độ khác nhau
Chủng
vi
khuẩn
Nồng độ microcystin ban đầu
2 µg/l 5 µg/l 10 µg/l
(1) (2) (3)
K(1/ngày) t0,5(ngày) K(1/ngày) t0,5(ngày) K(1/ngày) t0,5(ngày)
N4 0,076 0,074 0,045 0,102 0,043 0,140
N7 0,094 0,092 0,052 0,156 0,056 0,153
0
0.5
1
1.5
2
2.5
3
3.5
4
Hàm
lƣ
ợng M
C p
hân
giả
i
2 5 10
Nồng độ microcystin ban đầu
N4
N7
Hình 3.30. Thời gian bán phân giải microcystin ở các nồng độ khác nhau của 2
chủng N4, N7.

51
Khả năng phân giải độc tố của chúng khá nhanh, đặc biệt ở nồng độ độc tố
thấp 2 µg/l, thời gian bán phân giải microcystin tƣơng đối ngắn, đối với chủng LT2
là 0,074 ngày (1 giờ 47 phút), đối với LT8 là 0,092 ngày (2 giờ 12 phút). Với nồng
độ microcystin cao gấp 5 lần (2 µg/l), thời gian bán phân giải của 2 chủng LT2, LT8
lần lƣợt là 0,14 ngày (3 giờ 21 phút) và 0,153 ngày (3 giờ 40 phút). Hằng số tốc độ
ban đầu trong các nghiên cứu của chúng tôi là từ 0,43-0,56 /ngày, so sánh với hằng
số tốc độ trong báo cáo của Jones [28] từ 0,05-0,3/ngày, Jones đã đo mức độ phân
giải microcystin bằng các thể tích khác nhau của tế bào còn lại không bị tách chiết
từ môi trƣờng nuôi cấy vi khuẩn phân giải microcystin. Hằng số tốc độ theo
Kenefick [28 ] là 0,25/ngày cũng phản ánh sự phân giải chậm hơn. Các kết quả trên
cho phép chúng tôi sơ bộ kết luận 2 chủng vi khuẩn Sphingomonas N4, N7 phân
lập từ nƣớc hồ có hoạt tính phân giải độc tố microcystin. Khả năng phân giải đối
với từng loại độc tố microcystin cụ thể sẽ đƣợc tiến hành trong các nghiên cứu tiếp
theo.
3.5.2. Xác định điều kiện nhiệt độ thích hợp cho sự phân giải microcystin của
chủng Sphingomonas N4 và N7
Một trong các yếu tố gây ảnh hƣởng nhiều nhất tới tốc độ phân giải độc tố
microcystin của vi khuẩn Sphingomonas đƣợc xác định là nhiệt độ. Chính vì vậy,
trong nghiên cứu này chúng tôi tiến hành các thử nghiệm khả năng phân giải độc tố
microcystin bởi vi khuẩn Sphingomonas ở các nhiệt độ khác nhau là 20, 25, 30°C,
dải nhiệt độ này cho phù hợp với điều kiện tự nhiên của hồ với nồng độ microcystin
ban đầu là 40 μg/l. Kết quả đƣợc trình bày trong bảng 3.9, 3.10 và hình 3.31, 3.32.
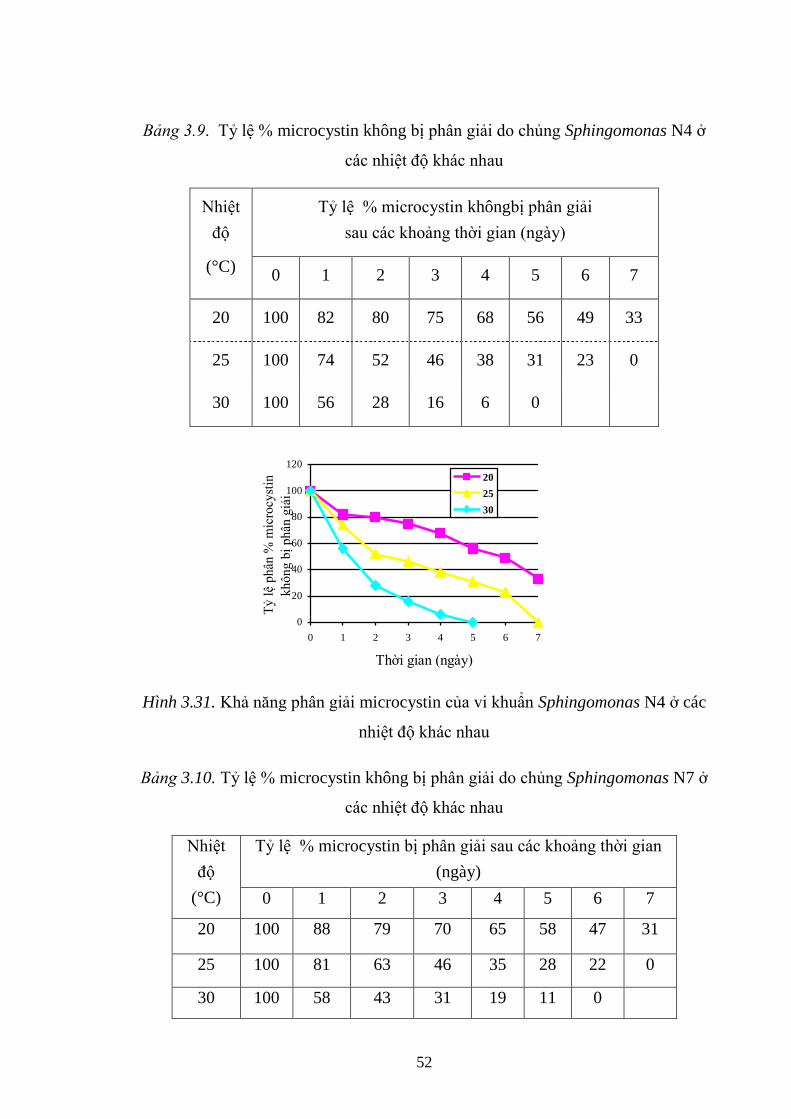
52
Bảng 3.9. Tỷ lệ % microcystin không bị phân giải do chủng Sphingomonas N4 ở
các nhiệt độ khác nhau
Nhiệt
độ
(°C)
Tỷ lệ % microcystin khôngbị phân giải
sau các khoảng thời gian (ngày)
0 1 2 3 4 5 6 7
20 100 82 80 75 68 56 49 33
25 100 74 52 46 38 31 23 0
30 100 56 28 16 6 0
0
20
40
60
80
100
120
0 1 2 3 4 5 6 7
Thời gian (ngày)
Tỷ
lệ
ph
ân %
mic
rocy
stin
kh
ôn
g b
ị p
hân
giả
i
20
25
30
Hình 3.31. Khả năng phân giải microcystin của vi khuẩn Sphingomonas N4 ở các
nhiệt độ khác nhau
Bảng 3.10. Tỷ lệ % microcystin không bị phân giải do chủng Sphingomonas N7 ở
các nhiệt độ khác nhau
Nhiệt
độ
(°C)
Tỷ lệ % microcystin bị phân giải sau các khoảng thời gian
(ngày)
0 1 2 3 4 5 6 7
20 100 88 79 70 65 58 47 31
25 100 81 63 46 35 28 22 0
30 100 58 43 31 19 11 0
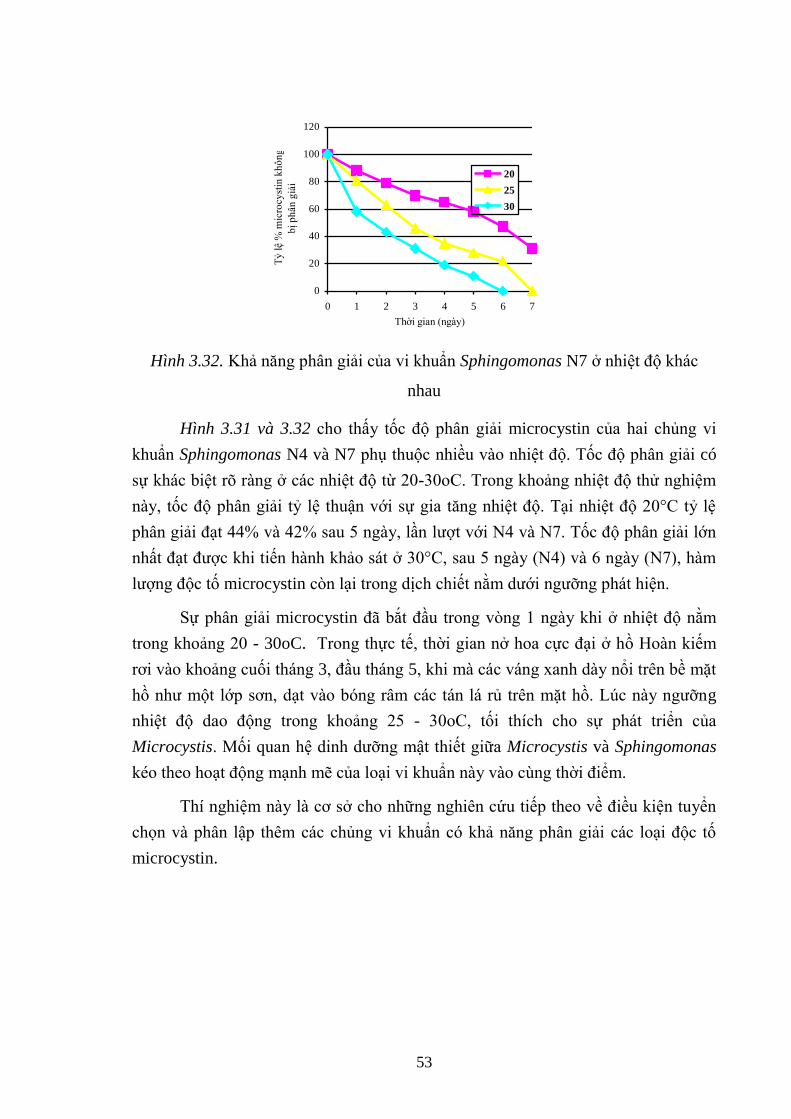
53
0
20
40
60
80
100
120
0 1 2 3 4 5 6 7
Thời gian (ngày)
Tỷ
lệ
% m
icro
cyst
in k
hô
ng
bị
ph
ân g
iải
20
25
30
Hình 3.32. Khả năng phân giải của vi khuẩn Sphingomonas N7 ở nhiệt độ khác
nhau
Hình 3.31 và 3.32 cho thấy tốc độ phân giải microcystin của hai chủng vi
khuẩn Sphingomonas N4 và N7 phụ thuộc nhiều vào nhiệt độ. Tốc độ phân giải có
sự khác biệt rõ ràng ở các nhiệt độ từ 20-30oC. Trong khoảng nhiệt độ thử nghiệm
này, tốc độ phân giải tỷ lệ thuận với sự gia tăng nhiệt độ. Tại nhiệt độ 20°C tỷ lệ
phân giải đạt 44% và 42% sau 5 ngày, lần lƣợt với N4 và N7. Tốc độ phân giải lớn
nhất đạt đƣợc khi tiến hành khảo sát ở 30°C, sau 5 ngày (N4) và 6 ngày (N7), hàm
lƣợng độc tố microcystin còn lại trong dịch chiết nằm dƣới ngƣỡng phát hiện.
Sự phân giải microcystin đã bắt đầu trong vòng 1 ngày khi ở nhiệt độ nằm
trong khoảng 20 - 30oC. Trong thực tế, thời gian nở hoa cực đại ở hồ Hoàn kiếm
rơi vào khoảng cuối tháng 3, đầu tháng 5, khi mà các váng xanh dày nổi trên bề mặt
hồ nhƣ một lớp sơn, dạt vào bóng râm các tán lá rủ trên mặt hồ. Lúc này ngƣỡng
nhiệt độ dao động trong khoảng 25 - 30oC, tối thích cho sự phát triển của
Microcystis. Mối quan hệ dinh dƣỡng mật thiết giữa Microcystis và Sphingomonas
kéo theo hoạt động mạnh mẽ của loại vi khuẩn này vào cùng thời điểm.
Thí nghiệm này là cơ sở cho những nghiên cứu tiếp theo về điều kiện tuyển
chọn và phân lập thêm các chủng vi khuẩn có khả năng phân giải các loại độc tố
microcystin.

54
KẾT LUẬN VÀ KIẾN NGHỊ
Kết luận
1. Hồ Hoàn Kiếm đang ở tình trạng ô nhiễm, hàm lƣợng COD, BOD vƣợt
tiêu chuẩn Việt Nam, TCVN 4942- 1995 (loại B) 20 lần. Với giá trị NH4+ từ 0,129-
0,699 (mg/l), NO2- từ 0.012- 0,05 (mg/l), NO3
- từ 0,047- 0,129 (mg/l), PO4
3- từ
0,08- 0.64 (mg/l) chứng tỏ hồ có hàm lƣợng dinh dƣỡng N, P khá cao.
2. Đã phân lập thuần khiết và nuôi cấy 10 chủng vi khuẩn lam thuộc chi
Microcystis theo phƣơng pháp của Shirai có cải tiến. Môi trƣờng thạch agarose B12
với tỷ lệ 0,4% là môi trƣờng thích hợp nhất cho việc phân lập Microcystis.
3. Các chủng Microcystis đều sinh trƣởng tốt trên môi trƣờng Bold 3N với số
lƣợng tế bào đạt từ 32,94 – 68,6 ×106 tb/ml sau 15 đến 19 ngày nuôi cấy. Đặc biệt
chủng LT2 số lƣợng tế bào đạt cao nhất là 68,6 ×106 tb/ml sau ngày thứ 19. Trên
môi trƣờng J, các chủng Microcystis sinh trƣởng số lƣợng tế bào của các chủng đạt
từ 40,42 – 76,46 ×106 tb/ml sau 15 đến 19 ngày nuôi cấy. Trong đó chủng LT2 vẫn
phát triển với số lƣợng tế bào đạt cao nhất 76,46 ×106 tb/ml sau ngày thứ 19. Trên
môi trƣờng B12, số lƣợng tế bào đạt từ 35,36 – 54,78 ×106 tb/ml sau 12 đến 15
ngày nuôi cấy. ở môi trƣờng này chủng LT8 phát triển với mật độ tế bào đạt cao
nhất 54,78 ×106 tb/ml sau ngày thứ 15.
4. Có 7 loại axít béo không no dạng C15, C17 và C19 trong tế bào 2 chủng
Microcystis LT2 và LT8. trong đó thành phần acid palmitic (C16H32O2) chiếm
lƣợng lớn nhất với 1,302 μg/mg trọng lƣợng khô ở chủng LT2, chủng LT8 là
0,9683 μg/mg trọng lƣợng khô. Kết quả cho thấy có sự sai khác nhau về lƣợng của
7 loại acid béo có ở trong tế bào 2 chủng LT2 và LT8.
5. Dựa vào phƣơng pháp phân loại truyền thống theo phân loại hình thái học
và phƣơng pháp phân loại hiện đại dựa trên phân tích trình tự rADN 16S, 2 chủng
vi khuẩn LT 2, LT8 đƣợc xác định là Microcystis aeruginosa, 2 chủng vi khuẩn
nƣớc N4 và N7 đƣợc xác định thuộc các loài Sphingomonas mali và Sphingomonas
asaccharolytica.

55
6. Phân tách trên TLC cho thấy dịch chiết từ sinh khối Microcystis khô có
chứa độc tố, hàm lƣợng microcystin thô phân tích trên quang phổ với chủng LT2 là
102,4ng/ml và chủng LT8 là 108,8ng/ml
7. Dựa vào kết quả phân tích trên HPLC, hàm lƣợng độc tố MC – RR đạt lớn
nhất là 512,8 ng/mg trọng lƣợng khô, độc tố MC – YR là 236,8 ng/mg và thấp nhất
là độc tố MC – LR: 114,25 ng/mg.
8. Nhiệt độ thích hợp cho sự phân giải microcystin của các 2 chủng vi khuẩn
Sphingomonas sp., N4, N7 là 30°C. Sau 7 ngày sự phân giải gần nhƣ hoàn toàn là
100%.
.Kiến nghị
- Cần tiếp tục giám sát sự nở hoa của các chủng vi khuẩn lam gây độc ở hồ
Hoàn Kiếm - Hà Nội và mối liên quan của chúng với các điều kiện môi trƣờng.
- Cần tiếp tục tiến hành những nghiên cứu về các biện pháp giảm thiểu và
ngăn ngừa tác hại do vi khuẩn lam gây độc gây ra, trong đó chú ý đến các biện pháp
sinh học vừa cho chi phí rẻ mà lại thân thiện với môi trƣờng.

56
TÀI LIỆU THAM KHẢO
Tài liệu tiếng Việt
1. Đặng Đình Kim, Đặng Hoàng Phƣớc Hiền (2006), Điều tra phát hiện tảo
độc tại các thuỷ vực trọng điểm của Hà Nội và một số tỉnh phía Bắc làm cơ sở cho
việc xây dựng chỉ tiêu quan trắc sinh học trong giám sát chất lượng nguồn nước,
Báo cáo Tổng kết nhiệm vụ khoa học công nghệ phục vụ quản lý nhà nƣớc và bảo
vệ môi trƣờng, Viện Công nghệ Môi trƣờng – Viện Khoa học và Công nghệ Việt
Nam.
2. Đặng Đình Kim, Đặng Hoàng Phƣớc Hiền (1999), Công nghệ Sinh học vi
tảo. Nhà xuất bản Nông nghiệp.Tr 120 – 134.
3. Đặng Đình Kim, Đặng Hoàng Phƣớc Hiền, Dƣơng Thị Thuỷ, Dƣơng Đức
Tiến (1997), Phát hiện được một số vi tảo độc trong các ao hồ Hà Nội. Kỷ yếu Viện
Công nghệ Sinh học, tr.408-412.
4. Chu Văn Thuộc & cs (1998), “Bước đầu nghiên cứu về tảo biển độc hại ở
vùng cửa sông ven biển miền bắc Việt Nam”, Tài nguyên và Môi trƣờng biển, tập 5,
tr. 155-163, Nhà xuất bản Khao học và Kỹ thuật.
5. Dƣơng Đức Tiến & cs (2002), Bảo vệ nguồn gen vi tảo (microalgae) đặc
hữu quý hiếm ở hồ Hoàn Kiếm – Hà Nội, Báo cáo khoa học trong chƣơng trình tài
trợ bảo vệ môi trƣờng và gìn giữ di sản văn hoá - Công ty Ford Motor.
6. Dƣơng Đức Tiến (1995), Phân loại Vi khuẩn lam ở Việt Nam. Nhà xuất
bản Nông nghiệp.
7. Dƣơng Đức Tiến, Võ Văn Chi (1978), “Phân loại Thực vật học bậc
thấp”. Nhà xuất bản Đại học và Trung học chuyên nghiệp.Tr 87-95.
8. Trịnh Thị Thanh (2000), Độc học Môi trường và Sức khoẻ con người. Nhà
xuất bản Đại Học Quốc gia Hà Nội. tr 39-45.
9. Doãn Đoan Trinh (2003), Hà Nội- địa chỉ du lịch văn hoá. Nhà xuất bản
Văn hoá thông tin.tr 36-40.
Tài liệu tiếng Anh

57
10. A.Munkhong, A.Mahakhant, P.Thavipoke, V.Arunpairojana (2005), Lysis
of Microcystis cells and degration of microcystins by soil bacteria, Algal Culture
Collection and the Environment, pp. 109-119.
11. American public health association. (1975), Standard methods for the
examination of water and wastewater, 4th
edition.
12. Ball As, Williams M, Vincent D and Robinson J (2001), Algal growth
control by a barley straw extract. Bioresour. Technol. 77, pp.177-181.
13. Barrett PRF, Curnow J and Little John JW (1996), The control of diatom
and cyanobacterial blooms in reservoirs using barley straw. Hydrobiol. 340, pp.
307-311.
14. Carmichael W (1995), Toxic Microcystis and the environment. In:
Watanabe M, Harada K.pp. 230-234.
15. Carmichael, W.W. (1992), A Status Report on Planktonic Cyanobacteria
(Blue-Green Algae) and their Toxins. EPA/600/R-92/079, United States
Environmental Protection Agency, Cincinnati, Ohio.
16. Chorus I, Bartram J, ed. (1999), Toxic cyanobacteria in water. A guide to
their public health consequences, monitoring and management. Published by E &
FN Spon on behalf of the World Health Organization.
17. Codd, G.A, Ward, C.J. and Bell, S.G. (1997), Cyanobacterial toxins:
occurrence, modes of action, health effects and exposure routes. In: J.P. Seiler and
E. Vilanova [Eds], Applied Toxicology: Approaches through Basic Science.
Archives of Toxicology Supplement 19, Springer-Verlag, Berlin, pp. 399-410.
18. Codd, G.A. (1983), Cyanobacterial poisoning hazard in British
freshwaters. Vet. Rec., 113. pp. 223-224.
19. Dillenberg HO, Dehnel MK (1960), Toxic waterbloom in Saskatchewan,
Canadian Medical Association Journal, 83: pp. 1151–1154.
20. Dvid G. Bourne (1996), Enzymatic pathway for the bacteria degradation
of the Cyanobacteria cyclic peptide toxin Microcystin LR. Pp.1086-4094
21. Falconer I (1996), Potential impact on human health of toxic
cyanobacteria. Phycologia, 35(Suppl. 6): pp. 6–11.

58
22. Falconer I, Buckley TH (1989), Tumor promotion by Microcystis sp., a
blue-green algae occurring in water supplies. Medical Journal of Australia, 150:
pp. 351–352.
23. Falconer I, ed. (1993), Algal toxins in seafood and drinking water. New
York, NY, Academic Press, pp. 224.
24. Fastner J, Neumann U, Wirsing B, Weckesser J, Wiedner C, Nixdorf B,
Chorus I (1999), Microcystins (heptapeptides) in German fresh water bodies.
Environmental Toxicology, 14(1):pp. 13–22.
25. Gibson MT, Welch IM, Barrett PRF and Ridge I (1990), Barley straw as
an inhibitor of algal growth II: Laboratory studies. J. Appl. Phycol. 2. pp. 241-248.
26. Jianzhong Chen, Zhili Liu, Guanju Ren, Pengfu Li and Yiwen Jiang
(2004), Control of Microcystis aeruginosa TH01109 with batangas mandarin skin
and dwarf banana peel, Technical note in School of Life Sciences, Nanjing
University, Nanjing 210093, China.pp. 1200 – 1211.
27. Jones, G. J., and P. T. Orr. (1994), Release and degradation of
microcystin following algicide treatment of a Microcystis aeruginosa bloom in a
recre- ational lake, as determined by HPLC and protein phosphatase inhibition
assay. Water Res. 28:pp. 871–876.
28. Kenefick, S.L., S.E. Hrudey, H.G. Peterson and E.E. Prepas. (1993) Toxin
release from Microcystis aeruginosa after chemical treatment. Water Sci. Tech. 27:pp.
433-440.
29. L.R. Song, N.Q.Gan, H.Pan (2005), Toxic Microcystis blooms and
microcystin composition in eutrophic lakes in China, Algal Culture Collection and
the Environment, pp. 89-108.
30. MacKintosh C, Beattie KA, Klumpp S, Cohen P, Codd GA (1990),
Cyanobacterial microcystin-LR is a potent and specific inhibitor of protein phosphatases 1
and 2a from both mammals and higher plants. Federation of the European Biochemical
Society Letters, 264:pp. 187–192.
31. Makoto Shirai, Katsumi Matumaru, Akio Ohotake, Yoshichika
Takamura, Tokujiro Aida and Masayasu Nakano (1989), Development of a Solid

59
Medium for Growth and Isolation of Axenic Microcystis Strains (Cyanobacteria), Appl
Environ Microbiol. 1989 October; 55(10): pp. 2569-2571.
32. Miller L.T (1982), Single derivatization method for routine analysis of
bacterial whole-cell fatty acid methyl esters, including hydroxy acids. J.Clin.
Microbiol.,16. pp. 584 – 586.
33. Nakagawa, M., Y. Takamura, and 0. Yagi. (1987), Isolation of the
slime from a cyanobacterium, Microcystis aeruginosa K-3A. Agric. Biol. Chem.
51:pp. 329-337.
34. Padisork J (1997), Cylindrospermopsis raciborskii (Woloszynska)
Seenayya et Subba Raju, an expanding, highly adaptive blue-green algal species:
worldwide distribution and review of its ecology. Archiv für Hydrobiologie, Suppl.,
107:pp. 563–593.
35. Pillinger JM, Cooper JA and Ridge I (1994), Role of phenolic
compounds in the antialgal activity of barley straw. J. Chem. Ecol. 20, pp. 1557-
1569.
36. Ridge I, Pillinger JM and Walters J (1995), Alleviating the problems of
excessive algal growth. In: DM Harper and AJD Ferguson (eds.) The Ecological
Basis for River Management. John Wiley. Chichester,pp. 211-218.
37. Ridge I, Walters J and Street M (1999), Algal growth control by
terrestrial leaf litter: a realistic tool, Hydrobiol. 395/396,pp. 173-180.
38. Rinehart KL, Namikoshi M, Choi BW (1994), Structure and biosynthesis
of toxins from bluegreen algae (cyanobacteria). Journal of Applied Phycology, 6:
pp. 159–176.
39. Rouhiainen L, Buikema W, Paulin L, Haselkorn R (1995), Cloning and
characterization of peptide synthetase genes from a hepatotoxic Anabaena strain.
40.Oral presentation at the 1st International Congress on Toxic Cyanobacteria,
Bornholm. pp. 127 -130..
40. Sivonen K, Jones J (1999), Cyanobacterial toxins. In: Chorus I, Bartram
J, ed. Toxic cyanobacteria in water. A guide to their public health on sequences,

60
monitoring and management. Published by E & FN Spon on behalf of the World
Health Organization, pp. 41–112.
41. Staley, J.T., Bryant, M.P., Pfennig, N. and Holt, J.G. [Eds] 1989 Bergey's
Manual of Systematic Bacteriology. Volume 3, Williams & Wilkins,
Baltimore.pp.58-62.
42. Tisdale ES (1931), Epidemic of intestinal disorders in Charleston, W.
Va., occurring simultaneously with unprecedented water supply conditions.
American Journal of Public Health, 21: pp. 198–200.
43. Turner PC, Gammie AJ, Hollinrake K, Codd GA (1990), Pneumonic
associated with cyanobacteria. British Medical Journal, 300: pp. 1440–1441.
44. Tzong-Huei Lee (2000), Isolation and identification of seven
microcystins from a culture M.TN-2 strain of Microcystis aeruginosa,
Bot.Bull.Acad.Sin., 41: pp. 197-202.
45. Vollenweider, R.A. (1968), Scientific Fundamentals of the
Eutrophication of Lakes and Flowing Waters with Particular Reference to Nitrogen
and Phosphorus as Factors in Eutrophication. Organisation for Economic
Cooperation and Development, Paris. pp. 120 -123.
46. WHO (1993), Guidelines for Drinking-water Quality, Second edition.
Volume 1, Recommendations. World Health Organization, Geneva. Pp. 46- 59.
47. WHO (1996), Guidelines for Drinking-water Quality, Second edition.
Volume 2, Health Criteria and Other Supporting Information. World Health
Organization, Geneva.pp. 267 – 268.
48. WHO (1998), Guidelines for Drinking-water Quality. Second edition,
Addendum to Volume 2, Health Criteria and Other Supporting Information. World
Health Organization, Geneva.pp. 245 -249.
49. Watanabe, M. M., and F. Kasai. (1985), NIES-collection list of strains,
1st ed., The National Institute for Environmental Studies, The Environmental
Agency, Tsukuba, Japan. pp. 97.

61
50. WRc (1996), The Fate of Intracellular Microcystin-LR During Water
Treatment. Report Ref. 96/DW/07/4, UK Water Industry Research Ltd., London.pp
135- 168.
51. Yoo S, Carmichael W, Hoehn R, Hrudey S (1995), Cyanobacterial (blue-
green algal) toxins: A resource guide. Denver, CO, American Water Works
Association Research Foundation, pp. 229.
52. Yu SZ (1995), Primary prevention of hepatocellular carcinoma. Journal
of Gastroenterology and Hepatology, 10: pp. 674–682.
Trang web tham khảo
53. http://en.wikipedia.org/wiki/Microcystin
54. http://www-cyanosite.bio.purdue.edu/cyanotox/toxins/microcystins.html
55. http://www.glerl.noaa.gov/pubs/brochures/mcystisflyer/mcystis.html
56. http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=1287722
57. http://microbewiki.kenyon.edu/index.php/Sphingomonas
58. http://elearning.hueuni.edu.vn/file.php/81/phanloaithuvat/htm/c2.htm
59. http://www.rimf.org.vn/bantin/tapchinewsdetail.asp?TapChiID=1&news
60. http://www.vnn.vn/khoahoc/2003/4/8715/
61. http://dnr.metrokc.gov/wlr/waterres/lakes/ToxicAlgae.htm
62. http://who.int/water_sanitation_health/resourcesquality/toxcyanchap1
63. http://etd.lib.ttu.edu/theses/available/etd-04272006-122817
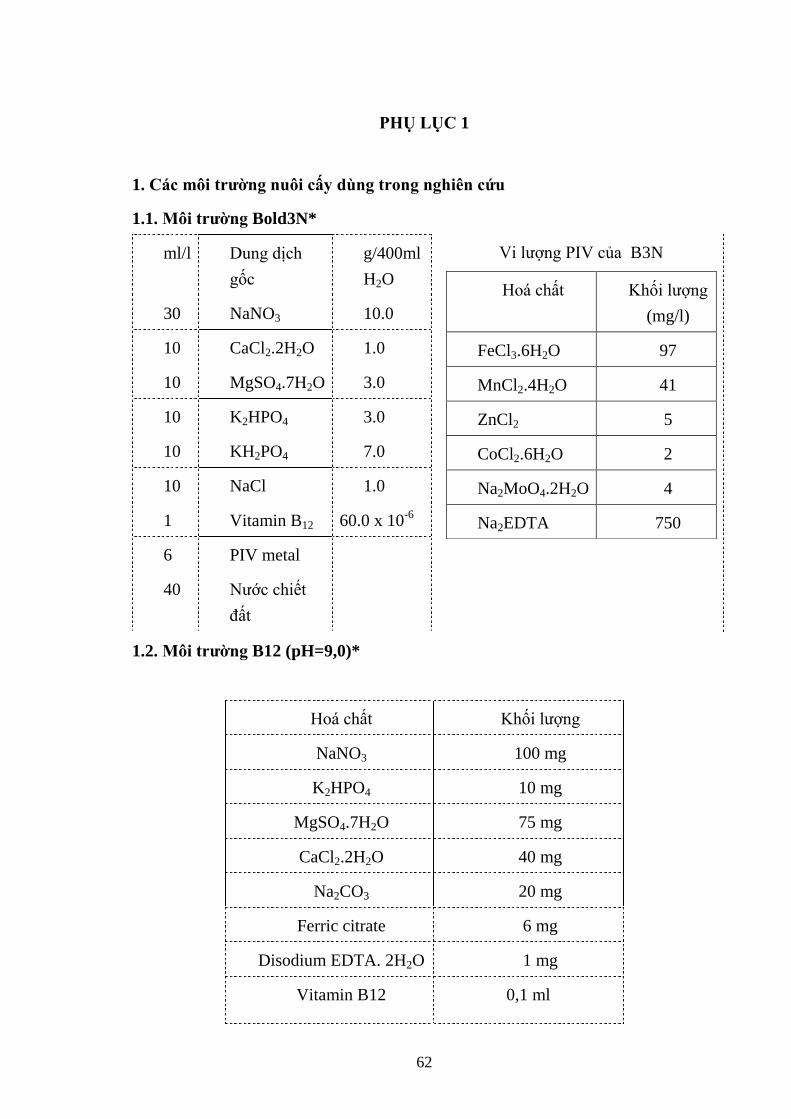
62
PHỤ LỤC 1
1. Các môi trƣờng nuôi cấy dùng trong nghiên cứu
1.1. Môi trƣờng Bold3N*
ml/l Dung dịch
gốc
g/400ml
H2O
30 NaNO3 10.0
10 CaCl2.2H2O 1.0
10 MgSO4.7H2O 3.0
10 K2HPO4 3.0
10 KH2PO4 7.0
10 NaCl 1.0
1 Vitamin B12 60.0 x 10-6
6 PIV metal
40 Nƣớc chiết
đất
Vi lƣợng PIV của B3N
Hoá chất Khối lƣợng
(mg/l)
FeCl3.6H2O 97
MnCl2.4H2O 41
ZnCl2 5
CoCl2.6H2O 2
Na2MoO4.2H2O 4
Na2EDTA 750
1.2. Môi trƣờng B12 (pH=9,0)*
Hoá chất Khối lƣợng
NaNO3 100 mg
K2HPO4 10 mg
MgSO4.7H2O 75 mg
CaCl2.2H2O 40 mg
Na2CO3 20 mg
Ferric citrate 6 mg
Disodium EDTA. 2H2O 1 mg
Vitamin B12 0,1 ml
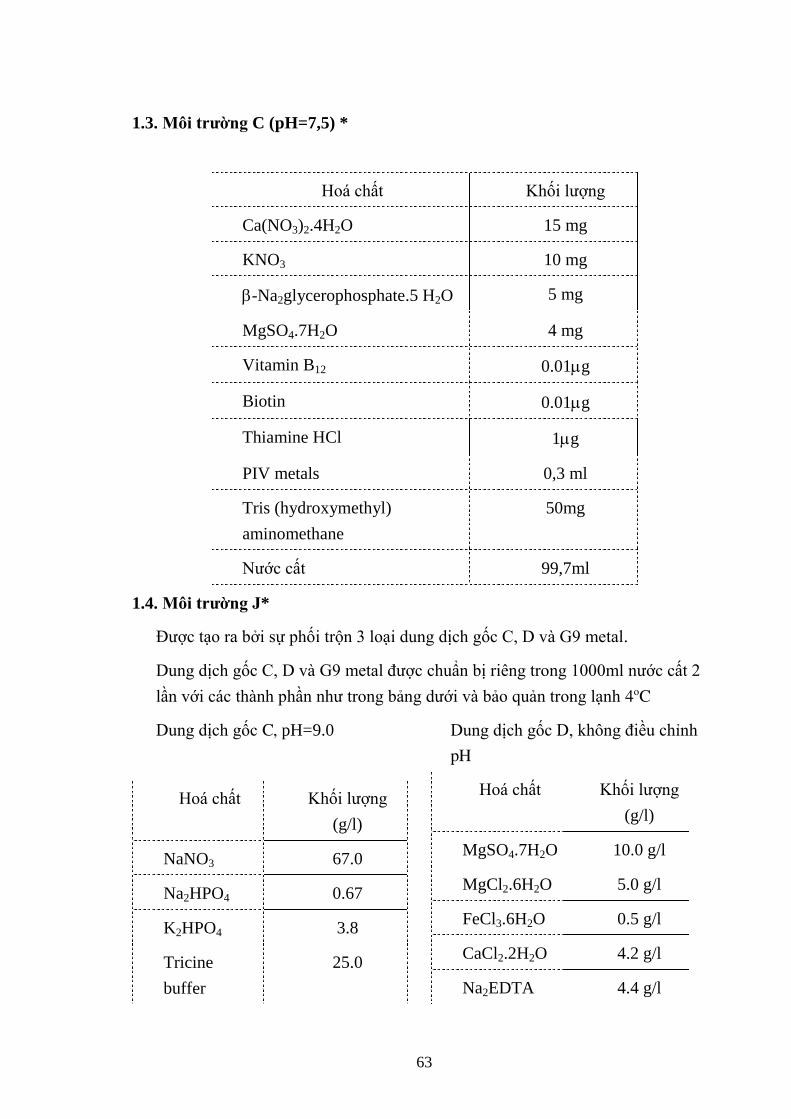
63
1.3. Môi trƣờng C (pH=7,5) *
Hoá chất Khối lƣợng
Ca(NO3)2.4H2O 15 mg
KNO3 10 mg
-Na2glycerophosphate.5 H2O 5 mg
MgSO4.7H2O 4 mg
Vitamin B12 0.01g
Biotin 0.01g
Thiamine HCl 1g
PIV metals 0,3 ml
Tris (hydroxymethyl)
aminomethane
50mg
Nƣớc cất 99,7ml
1.4. Môi trƣờng J*
Đƣợc tạo ra bởi sự phối trộn 3 loại dung dịch gốc C, D và G9 metal.
Dung dịch gốc C, D và G9 metal đƣợc chuẩn bị riêng trong 1000ml nƣớc cất 2
lần với các thành phần nhƣ trong bảng dƣới và bảo quản trong lạnh 4oC
Dung dịch gốc C, pH=9.0
Hoá chất Khối lƣợng
(g/l)
NaNO3 67.0
Na2HPO4 0.67
K2HPO4 3.8
Tricine
buffer
25.0
Dung dịch gốc D, không điều chỉnh
pH
Hoá chất Khối lƣợng
(g/l)
MgSO4.7H2O 10.0 g/l
MgCl2.6H2O 5.0 g/l
FeCl3.6H2O 0.5 g/l
CaCl2.2H2O 4.2 g/l
Na2EDTA 4.4 g/l
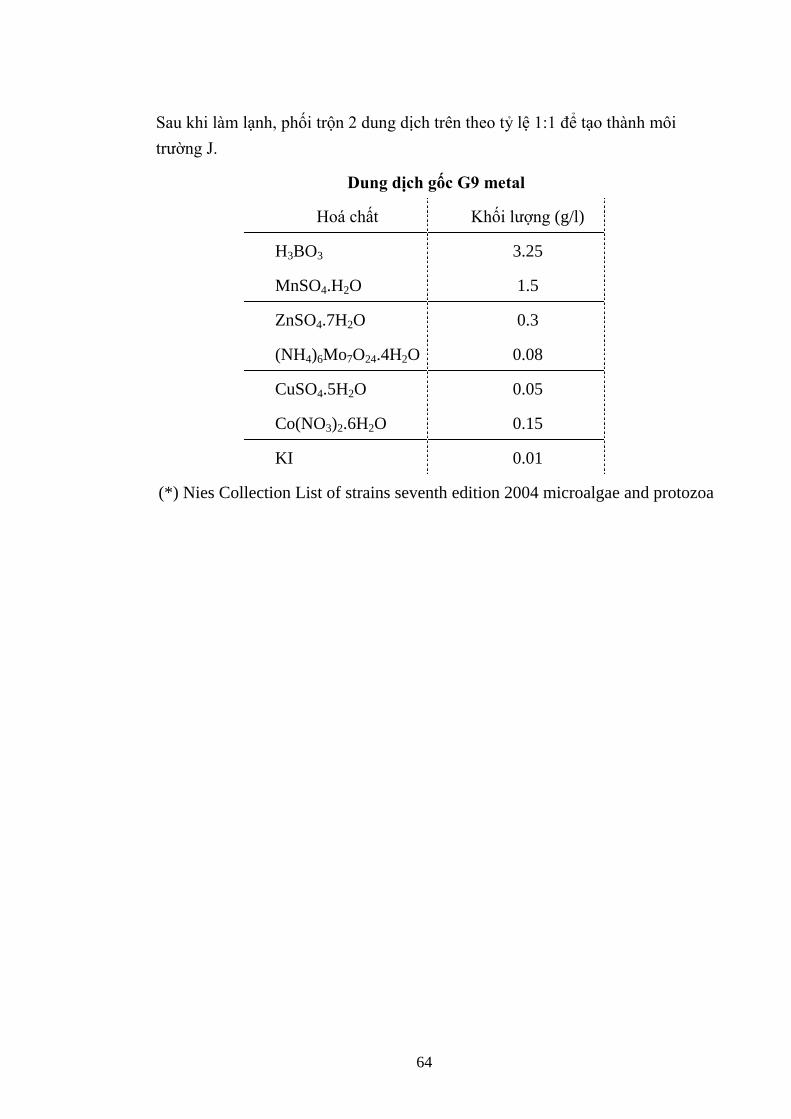
64
Sau khi làm lạnh, phối trộn 2 dung dịch trên theo tỷ lệ 1:1 để tạo thành môi
trƣờng J.
Dung dịch gốc G9 metal
Hoá chất Khối lƣợng (g/l)
H3BO3 3.25
MnSO4.H2O 1.5
ZnSO4.7H2O 0.3
(NH4)6Mo7O24.4H2O 0.08
CuSO4.5H2O 0.05
Co(NO3)2.6H2O 0.15
KI 0.01
(*) Nies Collection List of strains seventh edition 2004 microalgae and protozoa
65
PHỤ LỤC 2
Các môi trƣờng nuôi cấy vi khuẩn [39]
2.1. Môi trƣờng NA
Hóa chất Khối lƣợng (g)
Pepton 10,0
Cao thịt 1,0
NaCl 1,5
Thạch 15,0
Nƣớc cất 1 lít
Khử trùng ở 121oC trong 15 phút, pH=7
2.2. Môi trƣờng M7
Hóa chất Khối lƣợng (g)
Peptone 1,0
Yeast extract 1,0
Agar 15,0
Glucose 1,0
Nƣớc cất 1 lít
Khử trùng ở 121oC trong 15 phút, pH=7

66
PHỤ LỤC 3
Phƣơng pháp phân tích một số thông số thuỷ hoá
3.1. Xác định hàm lƣợng oxy hoà tan (DO) bằng phƣơng pháp Winkler
Nguyên tắc: phƣơng pháp xác định DO dựa vào khả năng của oxy hoà tan
oxy hoá Mn2+
thành Mn4+
trong môi trƣờng kiềm. Mn4+
lại có khả năng oxy hoá I-
thành I2 trong môi trƣờng axít. Do vậy lƣợng I2 đƣợc tạo ra có quan hệ định lƣợng
với DO và đƣợc xác định bằng Thiosulfat Natri.
Phương trình phản ứng:
Hoá chất cần dùng:
1) Dung dịch MnSO4: cân 480g MnSO4.4H2O không chứa sắt, dùng nƣớc
cất pha thành 1 lít
2) Dung dịch i-ô-dua kali kiềm: cân 350g KOH và 75g KI, dùng nƣớc cất
pha thành 500ml
3) Acid HCl đặc
4) Dung dịch Thiosulfat Natri 0,025N: Hoà tan 0,2g Na2S2O3.5H2O trong
1 lít nƣớc cất đã đun sôi để nguội. Dung dịch đƣợc đựng trong bình màu nâu, cho
thêm 0,2g Na2CO3
5) Dung dịch tinh bột
Cách tiến hành:
Lấy mẫu và cố định mẫu: dùng bình sạch lấy đầy mẫu nƣớc. Mẫu cần phải cố
định ngay, hút 1ml MnSO4, để đầu pipet dúng vào trong mặt nƣớc xong, rồi từ từ
cho thuốc thử vào bình. Sau đó hút 1ml dung dịch kiềm i-o-dua kali. Sau khi cố định
xong, đậy nút, lắc đều mẫu nƣớc đến khi thấy sinh ra kết tủa vàng nâu. Mẫu nƣớc đã
cố định nhƣ thế có thể giữ đƣợc vài giờ không thay đổi, đem về phòng thí nghiệm
rồi tiến hành công tác phân tích.
MnSO4 + 2KOH = Mn(OH)2 + K2SO4
2Mn(OH)2 + O2 = MnO2 + H2O
MnO2 + 2KI + 4H+
= Mn2+
+ 2K+ + I2 + 2H2O
2Na2S2O3 + I2 = Na2S4O6 + 2NaI (chuẩn độ)

67
Phân tích: trong mẫu nƣớc mà oxy hoà tan đã bị cố định và kết tủa hoàn toàn
lắng xuống, dùng pipet hút cho vào 1ml acid H2SO4 đậm đặc. Sau đó đậy nút lại, lắc
đều bình cho kết tủa vàng nâu hoà tan hết. Lấy 100ml dung dịch này vào bình tam
giác 250ml. Dùng dung dịch Na2S2O3 chuẩn độ cho đến màu vàng rất nhạt, cho
thêm 3 giọt dung dịch tinh bột, dung dịch có màu xanh lam. Tiếp tục dùng dung dịch
Na2S2O3 chuẩn độ cho tới khi màu lam vừa vặn biến mất. Ghi lƣợng Na2S2O3 đã
dùng.
Công thức tính toán kết quả:
8.W.N
DO / = 1000V
mg l
Trong đó: N - nồng độ dung dịch Na2S2O3 (N)
V - thể tích mẫu dùng chuẩn độ (ml)
W - thể tích dung dịch chuẩn Na2S2O3 (ml)
3.2. Xác định nhu cầu oxy sinh học BOD (Biology Oxygen Demand)
BOD là lƣợng oxy cần thiết để oxi hóa hoàn toàn các hợp chất hữu cơ
có trong nƣớc bằng vi sinh vật.
Nguyên tắc xác định: Định lƣợng nhu cầu oxi sinh hóa (BOD) bằng
thiết bị Oxitop (có gắn sensor BOD) dựa trên sự giảm áp do khí CO2 sinh ra trong
quá trình oxi hóa, đƣợc hấp thụ lại bởi các hạt kiềm mạnh . Sự giảm áp suất đƣợc bộ
vi xử lý trong sensor chuyển thành giá trị BOD tƣơng ứng.
Dụng cụ và hóa chất:
- Dung dịch đệm phốt phát pH = 7,2 gồm:
+ KH2PO4 : 8,5 g/l + K2HPO4 : 21,75 g/l
+ Na2HPO4.7H2O : 33,4 g/l + NH4Cl : 1,7 g/l
- Dung dịch MgSO4.7H2O : 22,5 g/l
- Dung dịch CaCl2 : 27,5 g/l
- Dung dịch FeCl3.6H2O : 0,25 g/l
- Dung dịch H2SO4 0,1N
- Dung dịch NaOH 0,1 N
- Dung dịch Na2SO3 0,025 N

68
- Nƣớc cất đã đƣợc cất với kalipemanganat
- KOH tinh thể
Xử lý mẫu:
- Loại bỏ các chất tẩy rửa bằng dung dịch Na2SO3 0,025N.
- Loại bỏ kim loại độc bằng cách điều chỉnh pH của mẫu đến 8,5 khi đó
hầu hết các kim loại sẽ kết tủa ở dạng oxit và hydroxit. Đem lọc bỏ kết tủa
- Trung hòa mẫu: Điều chỉnh pH của mẫu từ 6,5 – 7,5 bằng dung dịch
NaOH hoặc H2SO4.
- Bổ sung vi sinh vật nếu lƣợng vi sinh vật trong mẫu không đủ bằng
cách lấy nƣớc cống trên bề mặt, để lắng từ 1- 2 giờ. Nƣớc thu đƣợc có hàm lƣợng vi
sinh vật trong nƣớc khoảng 103 – 106 vi khuẩn/ ml . Thể tích nƣớc bổ sung có thể
dao động từ 3 – 30% thể tích mẫu, thông thƣờng bằng 10% thể tích mẫu.
- Nếu mẫu nƣớc không đủ dinh dƣỡng cần bổ sung thêm 1ml các dung
dịch đệm photphat, dung dịch MgSO4.7H2O, dung dịch CaCl2, dung dịch
FeCl3.6H2O vào một lít nƣớc pha loãng.
- Chuẩn bị nƣớc pha loãng cấy vi sinh vật: Nƣớc cất đã bổ sung chất
dinh dƣỡng, đƣợc sục đến bão hòa oxi pha với nƣớc có chứa vi sinh vật theo tỷ lệ
10/1.
Các bước tiến hành:
- Mẫu đƣợc ủ trong các điều kiện tiêu chuẩn: nhiệt độ là 200C, không có
ánh sáng, pH mẫu khoảng 6,5 – 7,5.
- Cho mẫu đã pha loãng, phù hợp với thang đo đã lựa chọn trong bảng
dƣới đây vào chai thí nghiệm.
Thể tích mẫu (ml) Khoảng BOD
(mg/l)
Hệ số pha loãng (f)
432 0 – 40 1
365 0-80 2
250 0-200 5

69
- Cho con từ vào mỗi chai ủ, đặt lẵng cao su vào miệng chai và cho hóa
chất hấp thụ CO2 (KOH hạt hoặc Soda hạt) vào lẵng cao su.
- Đặt và vặn chặt sensor BOD vào mỗi chai, cài đặt và bắt đầu quá trình
đo.
- Tiến hành làm với mẫu trắng bằng nƣớc pha loãng cấy vi sinh vật theo
trình tự tƣơng tự
- Xếp chai vào giá khuấy từ và đặt thiết bị khuấy từ vào tủ điều nhiệt và
nối với nguồn điện trong tủ
Tính toán kết quả:
5BOD = D- D´ f
Trong đó: BOD5 là giá trị BOD của mẫu cần xác định sau 5 ngày.
D là giá trị hiện trên sensor BOD sau 5 ngày lấy mẫu cần xác
định.
D‟ là giá trị hiện trên sensor BOD sau 5 ngày của mẫu trắng.
f là hệ số phụ thuộc vào thể tích mẫu.
3.3. Nhu cầu oxy hoá học ( COD) (Chemical Oxygen Demand)
COD la lƣơng oxy cân thiêt dung cho qua trinh oxy hoa hoan to àn các hợp
chât hƣu cơ co trong nguôn nƣơc băng cac chât oxy hoa manh nhƣ K 2Cr2O7 hay
KMnO4. Xác định bằng phƣơng pháp bicromat
Nguyên tắc: Khi cho dung dịch bicromat vào mẫu nƣớc, dƣới tác dụng của
nhiệt độ và H2SO4 đặc thì phần lớn các chất hữu cơ và vô cơ trong nƣớc bị oxy hoá.
Để phản ứng Oxy hoá xảy ra hoàn toàn ta cho thêm xúc tác Ag2SO4. Sau khi oxy
hoá, lƣợng Bicromat dƣ sẽ đƣợc xác định bằng phƣơng pháp chuẩn độ với dung
dịch muối Morh. Lƣợng chất hữu cơ bị oxy hoá sẽ tính ra bằng lƣợng oxy tƣơng
đƣơng qua Cr2O72-
bị khử, lƣợng oxy tƣơng đƣơng này chính là COD.
Phản ứng oxy của Bicromat đƣợc diễn ra theo phƣơng trình sau:
Cr2O72-
+ 14 H+
+ 6e = 2 Cr3+
+ 7 H2O
164 0-400 10
43.5 0-2000 50

70
Dụng cụ và hoá chất:
- Bình nón 50ml sạch và khô
- Pipet, nút bông sạch
- Dung dịch :
+ K2Cr2O7 đã sấy khô ở nhiệt độ 1050C trong 2 giờ, hoà tan trong
nƣớc cất, đƣa thể tích đến 1 lít;
+ Dung dịch H2SO4 đậm đặc;
+ Tinh thể Ag2SO4;
+ Vụn thuỷ tinh;
+ Chỉ thị màu Diphenylamin: 1g Diphenylamin trong 100ml H2SO4;
+ Dung dịch muối Morh 0,25N : Hoà tan 98g muối Fe (NH4)2(SO4)2 .6 H2O
trong nƣớc cất. Thêm 20ml H2SO4 đậm đặc, để nguội thêm nƣớc cất đến 1 lít.
Mỗi lần dùng muối Morh phải kiểm tra lại nồng độ bằng cách hút 25ml dung
dịch K2Cr2O7 0,25N cho vào bình tam giác, thêm nƣớc cất đến thể tích 200ml, thêm
15ml dung dịch H2SO4 1:1, thêm 5 giọt chỉ thị diphenylamin.
Dùng dung dịch muối Morh để chuẩn độ đến khi màu dung dịch chuyển sang
xanh lá cây. Ghi thể tích dung dịch muối Morh đã tiêu tốn (V ml). Nồng độ của
muối Morh đƣợc xác định theo công thức sau:
N =V
x 25,025
Cách tiến hành :
- Lấy 10ml nƣớc mẫu cho vào bình nón;
- Thêm 5ml K2Cr2O7 0,25N + 1ml H2SO4 đậm đặc + 200mg Ag2SO4
- Một ít vụn thuỷ tinh
- Lắc đều, đậy nút bông
- Làm tƣơng tự với mẫu nƣớc cất
- Đun hồi lƣu trong 2 giờ, sau 4 giờ đem ra làm lạnh
- Thêm 50ml nƣớc cất

71
- Thêm 2 giọt chỉ thị màu Diphenylamin, chuẩn độ bằng dung dịch muối
Morh 0,25N đến khi có màu xanh lá cây. Ghi lại lƣợng muối Morh đã dùng.
o Công thức tính lượng COD:
COD = V
xxNxVV 10008)21(
Trong đó :
V1 - là thể tích dung dịch muối Morh chuẩn mẫu trắng (m);
V2 - là thể tích dung dịch muối Morh chuẩn mẫu nƣớc cần xác định (ml);
N - là đƣơng lƣợng gam muối Morh (N).
3.4. Xác định hàm lƣợng NH4+ bằng phƣơng pháp so màu với thuốc thử Nessler
Nguyên tắc:
Ion NH4+ trong nƣớc tác dụng với thuốc thử Nessler cho hợp chất vàng nâu.
NH4+ + 2K2Hg2I4 + 3KOH NH2Hg2OI + 7KI + 2H2O.
Cƣờng độ màu của dung dịch chứa NH2Hg2OI tỷ lệ với hàm lƣợng NH4+
tham gia tạo thành hợp chất này.
Các yếu tố ảnh hưởng: các amin, cloramin, axeton, cồn và một số chất hữu
cơ hoà tan gây cản trở phép đo do chúng tác dụng với thuốc thử Nessler. Ngoài ra,
các chất gây nên độ cứng, sắt sunfua, độ đục ảnh hƣởng đến quá trình xác định. Loại
ảnh hƣởng cản trở bằng cách thêm muối Seignet.
Dụng cụ và hoá chất:
- Bình định mức 1 lít, 500 ml (1 cái), 50 ml (12 cái).
- Pipet các loại.
- Dung dịch muối Seignet 50%: hoà tan 50 g muối seignet trong 100 ml nƣớc
cất 2 lần.
- Dung dịch thuốc thử Nessler: cân 15 g HgI2 và 10 g KI hoà vào 500 ml
nƣớc cất, cho vào 40 g NaOH khuấy đều cho tan, để lắng vài ngày, gạn dung dịch
vào bình nâu.
Phương pháp lập đường chuẩn:

72
Dung dịch NH4Cl tiêu chuẩn: cân 0,382 g NH4Cl tinh khiết đã sấy khô ở
105oC trong 2 giờ, pha vào bình định mức 1 lít. Dung dịch có nồng độ là 0,1 mg N-
NH4+ trong 1 ml. Lấy 10 bình định mức có số thứ tự từ 1- 10. Dùng pipet hút dung
dịch chuẩn cho vào bình định mức với các thể tích là: 1 ml, 2 ml, 3 ml, 4 ml, 5 ml, 6
ml, 7 ml, 8 ml, 9 ml, 10 ml sau đó cho thêm 1 ml dung dịch seignet và 1 ml dung
dịch nessler, thêm nƣớc cất đến vạch định mức. Tiến hành so màu ở bƣớc sóng 420
nm trên máy so màu quang điện ERMA 11 TOKYO. Căn cứ vào các giá trị mật độ
quang đo đƣợc ta lập đƣờng chuẩn.
Cách tiến hành:
Lấy 5 ml nƣớc mẫu cần phân tích cho vào bình định mức 50 ml, pha loãng
bằng nƣớc cất tới 25 ml. Cho vào bình 1 ml muối seignet, tiếp tục cho vào 1 ml
thuốc thử nessler, thêm nƣớc cất đến vạch định mức. Làm tƣơng tự đối với mẫu
trắng và so màu ở bƣớc sóng 420 nm. Tra đƣờng chuẩn xác định hàm lƣợng N- NH-
4+ trong mẫu.
Công thức tính kết quả:
N-NH4+ = 1000
1,0
V
W
Trong đó: W- thể tích NH4Cl tiêu chuẩn tra trên đƣờng chuẩn.
V- thể tích mẫu nƣớc dùng phân tích (5 ml).
3.5. Xác định hàm lƣợng NO3- bằng acid phenoldisunfonic
Nguyên tắc:
Ion NO3- phản ứng với acid phenoldisunfonic tạo ra acid
nitrophenoldisunfonic. Acid này tác dụng với dung dịch kiềm cho muối màu vàng,
cƣờng độ màu tỷ lệ với hàm lƣợng NO3- trong dung dịch.
C6H3OH(SO3H)2 + HNO3 + 3NH4OH C6H2ONH4(SO3NH4)2NO2 + 4H2O.
(acid phenoldisunfonic) (màu vàng)
Dụng cụ, hoá chất:
- Bát sứ - Pipet các loại.
- Bình định mức 50 ml. - Dung dịch NH4OH 25%.
- Dung dịch NaOH 20%.

73
- Acid phenoldisunfonic: cân 3 g phenol nguyên chất kết tinh màu trắng cho
vào bình tam giác, thêm 20 ml H2SO4 đặc (d=1,84). Đậy bình bằng nút có ống thủy
tinh làm lạnh bằng không khí. Đun sôi cách thuỷ trong 6 giờ đƣợc dung dịch màu
vàng nhạt. Bảo quản trong bình sẫm màu.
Phương pháp lập đường chuẩn:
Cân 0,7216 g KNO3 đã sấy khô ở 105oC trong 2h, cho vào bình định mức 1
lit, dung nƣớc cất 2 lần hoà tan và thêm đến vạch định mức. 1 ml dung dịch này
chứa 0,1 mg N-NO3-. Lấy 50 ml dung dịch này cho vào bát sứ cô cạn, để nguội rồi
thêm 1 ml thuốc thử disunfonic, dùng đũa thuỷ tinh cạo nhẹ thành bát để trộn đều
thuốc thử, để yên trong 10 phút để thuốc thử ngấm đều. Sau đó thêm 15 ml nƣớc
cất, trung hoà bằng NaOH 20% đến pH kiềm, cho toàn bộ dung dịch màu vàng này
vào bình định mức 200 ml (dung dịch A).
Lấy 10 bình định mức 50 ml, cho dung dịch A vào bình định mức lần lƣợt
với các thể tích 1 ml, 3 ml, 5 ml, 7 ml, 9 ml rồi định mức đến vạch. So màu ở bƣớc
sóng 420 nm trên máy so màu quang điện ERMA 11 TOKYO, xây dựng đƣờng
chuẩn.
Cách tiến hành:
Lấy 50 ml nƣớc mẫu cần phân tích cho vào bát sứ, chƣng cạn, để nguội, cho
vào 1 ml dung dịch acid phenoldisunfonic, dùng đũa thuỷ tinh cạo thành bát để hoà
tan hết cặn, để yên trong 10 phút để acid ngấm đều. Cho thêm một ít nƣớc cất, cho
vào 7 ml dung dịch NH4OH 25%, đổ tất cả sang bình định mức 50 ml, tráng bát sứ
bằng nƣớc cất rồi đổ sang bình định mức. Dùng nƣớc cất định mức đến vạch. So
màu ở bƣớc sóng 420 nm trên máy so màu quang điện ERMA 11 TOKYO. Căn cứ
vào mật độ quang đo đƣợc và đƣờng chuẩn tính hàm lƣợng NO3- trong mẫu cần
phân tích.
Công thức tính kết quả:
N-NO3- (mg/l) = 1000
025,0
V
W
Trong đó: W- thể tích tra trên đƣờng chuẩn.
V- thể tích mẫu dùng phân tích (50 ml).
3.6. Phƣơng pháp xác định hàm lƣợng NO2-
Nguyên tắc:

74
Phƣơng pháp này dựa vào phản ứng Diajo hóa của nitrit trong môi trƣờng
acid với acid sunphanilamide tạo thành muối aro có màu đỏ hồng. Phƣơng pháp này
thích hợp với việc xác định NO2- có hàm lƣợng 1 mg/l N dạng NO2
- trở xuống.
Các yếu tố ảnh hưởng:
Các ion sau đây có thể gây cản trở: Fe3+
, Pb2+
, Hg2+
, Ag+, các ion có màu, khi
chúng có mặt trong dung dịch làm thay đổi màu của hệ. Các chất rắn cũng ảnh
hƣởng đến việc xác định NO2-.
Dụng cụ và hoá chất:
- Pipet các loại - Bình định mức 50 ml.
- Thuốc thử sunphanilamide: hoà tan 5g sunphanilamide trong hỗn hợp 50 ml
acid HCl đặc và 300 ml nƣớc cất. Pha loãng đến 500 ml bằng nƣớc cất.
- Dung dịch NED dihydroclorua: hoà tan 500 mg NED dihydroclorua trong
500 ml nƣớc, đựng trong chai tối màu, pha chế lại hàng tháng hoặc ngay tức khắc
khi thấy dung dịch chuyển sang màu nâu.
Các lập đường chuẩn:
Dung dịch NO2- tiêu chuẩn: cân 0,4927 g NaNO2 khan vào trong bình định
mức 1 lít, thêm nƣớc cất đến vạch, dung dịch này chứa 0,1 mg N-NO2- trong 1 ml.
Lấy 10 ml dung dịch này cho vào bình định mức 1 lít, thêm nƣớc cất đến vạch, dung
dịch này là dung dịch tiêu chuẩn có nồng độ 0,001 mg N-NO2- trong 1 ml. Cho vào
bình định mức 50 ml có thứ tự từ 1- 10 dung dịch chuẩn với các thể tích 1 ml, 2ml,
3ml, 4ml, 5 ml, 6 ml, 7 ml, 8 ml, 9 ml, 10 ml. Sau đó thêm nƣớc cất đến vạch, so
màu ở bƣớc sóng 530 nm trên máy so màu quang điện ERMA 11 TOKYO. Căn cứ
vào giá trị mật độ quang lập đƣờng chuẩn.
Cách tiến hành:
- Lấy 5 ml nƣớc mẫu cần phân tích cho vào bình định mức 50 ml, đối với
nƣớc thải thì pha loãng thành 25 ml.
- Dùng pipet cho vào bình 1ml dung dịch sunphanilamide, lắc trong 2 phút,
cho thêm 1ml dung dịch NED dihydroclorua, lắc trong 10 phút.
- Làm tƣơng tự với mẫu trắng. Tiến hành so màu trên máy so màu quang điện
ERMA 11 TOKYO ở bƣớc sóng 530 nm.
Công thức tính kết quả:

75
Hàm lƣợng N-NO2- đƣợc tính theo công thức:
N-NO2- (mg/l) = 1000
001,0
V
W
Trong đó: W- thể tích dung dịch N-NO2- tra trên đƣờng chuẩn.
V- thể tích mẫu nƣớc lấy phân tích (5 ml).
3.7. Phƣơng pháp xác định hàm lƣợng photphat PO43-
Nguyên tắc:
Khi đun nóng dung dịch PO43-
với molipdat amon có mặt H2SO4 đặc sẽ tạo
thành acid photpho molipdat, acid này tác dụng với acid ascorbic trong môi trƣờng
pH= 4 sẽ cho hợp chất photpho molipdat màu xanh lam. Căn cứ vào cƣờng độ màu
đo đƣợc và đƣờng chuẩn xác định hàm lƣợng PO43-
trong dung dịch cần phân tích.
Hoá chất và dụng cụ:
- Pipet các loại - Bình định mức 50 ml.
- Dung dịch (NH4)6Mo7O24.4H2O 2,5%: cân 25 g molipdat amon, hoà tan
trong bình định mức 1000 ml. Thận trọng cho thêm 175ml dung dịch H2SO4 đặc, để
nguội dung dịch rồi thêm nƣớc cất đến vạch định mức.
- Dung dịch acid ascorbic 1%: 1g acid ascorbic hoà tan trong 100ml dung
dịch CuSO4 0,05M (dung dịch này chỉ chuẩn bị trƣớc khi dùng).
Cách lập đường chuẩn:
- Dung dịch chuẩn photphat: cân 0,22131 g KH2PO4 đã sấy khô ở nhiệt độ
105oC trong 2h, hoà tan đến 1 lít bằng nƣớc cất 2 lần ta đƣợc dung dịch chuẩn có
hàm lƣợng 0,05 mg P-PO43-
trong 1 ml.
- Cho vào 6 bình định mức 50 ml lần lƣợt các thể tích dung dịch chuẩn
photphat nhƣ sau: 0,1 ml; 0,3 ml; 0,5 ml; 0,7 ml; 0,9 ml; 1,1 ml. Thêm 2 ml dung
dịch molipdat amon; 0,5 ml dung dịch acid ascorbic, thêm nƣớc cất đến vạch định
mức. Đun cách thuỷ ở nhiệt độ 30oC trong 10 phút. So màu trên máy so màu quang
điện ERMA 11 TOKYO ở bƣớc sóng 660 nm. Từ kết quả đo mật độ quang lập
đƣờng chuẩn.
Cách tiến hành phân tích:
Lấy 10 ml nƣớc mẫu cho vào bình định mức 50 ml, dùng pipet cho 2 ml dung
dịch thuốc thử molipdat amon và 0,5 ml dung dịch acid ascorbic, thêm nƣớc cất đến

76
vạch định mức. Đun cách thuỷ ở nhiệt độ 30oC trong 10 phút. So màu ở bƣớc sóng
660 nm trên máy so màu quang điện ERMA 11 TOKYO. Căn cứ vào giá trị mật độ
quang đo đƣợc và đƣờng chuẩn ta xác định đƣợc hàm lƣợng PO43-
trong mẫu.
Công thức tính kết quả:
P- PO43-
= 100005,0
V
W
Trong đó: W- thể tích tƣơng ứng với mật độ quang đo đƣợc.
V- thể tích mẫu đã dùng để phân tích (10 ml).
3.8. Xác định hàm lƣợng cặn lơ lửng SS
Nguyên tắc: Nƣớc mẫu đƣợc lắc đều đem lọc qua giấy lọc đã đƣợc biết trƣớc
khối lƣợng. Cặn còn lại trên giấy lọc đƣợc sấy đến trọng lƣợng không đổi ở nhiệt độ
1050C trong 2 giờ. Khối lƣợng chênh lệch trƣớc và sau khi sấy chính là cặn lơ lửng
SS hay còn gọi là chất rắn huyền phù
Các bước tiến hành:
Giấy lọc đƣợc sấy khô tới trọng lƣợng không đổi ở 1050C trong 2 giờ, để
nguội trong bình hút ẩm sau đó cân chính xác trọng lƣợng của giấy lọc. Lấy 100 ml
nƣớc mẫu lọc qua giấy lọc. Đem sấy giấy lọc và lớp cặn bên trên, để nguội trong
bình hút ẩm và cân chính xác trọng lƣợng giấy sau khi lọc.
Công thức tính kết quả:
A - B
SS / = 1000V
mg l
Trong đó: A là trọng lƣợng của giấy lọc và lớp cặn sau khi sấy (mg)
B là trọng lƣợng của giấy lọc ban đầu (mg)
V là thể tích mẫu nƣớc lọc qua giấy lọc (ml)

77
PHỤ LỤC 4
4.1. Phƣơng pháp nhuộn Gram
Nguyên tắc: Nhuộm Gram (phản ứng Gram) có vai trò đặc biệt trong việc
phân loại vi khuẩn. Trong quá trình nhuộm Gram, tế bào bƣớc đầu đƣợc xử lý với
tím kết tinh và iot nên có sự tạo thành phức chất tím kết tinh – iot bên trong tế bào.
Khi vi khuẩn Gram âm bị tẩy cồn, lipit của lớp màng ngoài bị hoà tan làm tăng tính
thấm của màng dẫn đến sự rửa trôi phức chất tím – iot và làm cho vi khuẩn mất
màu. Khi nhuộm bổ sung chúng sẽ bắt màu với thuốc này (Đỏ vàng với Safranin
hay đỏ tía với Fuchsin) . ở vi khuẩn Gram dƣơng, cồn làm cho các lỗ trong
peptidoglycan co lại do đó phức chất tim – iot bị giữ lại bên trong tế bào.
Cách tiến hành: Dùng que cấy vô trùng lấy dịch huyền phù tế bào vi khuẩn
đặt lên phiến kính. Cố định tiêu bản bằng ngọn lửa rồi nhuộn thuốc đầu bằng dung
dịch tím kết tinh (crystal violet) trong khoảng 1 phút. Rửa bằng nƣớc. Nhuộm tiếp
bằng dung dịch iốt (dung dịch Lugol) trong 1 phút. Rửa bằng nƣớc. Phủ lên vết bôi
dung dịch etanol 95%:axeton (1:1) trong khoảng 1 phút. Lại rửa bằng nƣớc. Sau đó
nhuộm tiếp bằng thuốc nhuộm màu đỏ (nhƣ Safranin hay Fuchsin Ziehl). Rửa qua
nƣớc, để khô, soi kính.
4.2. Oxidaza
Thử bằng kit (strip), chủng dƣơng tính cho màu tím, chủng âm tính không
màu.
4.3. Xác định khả năng di động (Motility test)
Phần lớn các vi khuẩn Bacillus và sphingomonas đều có khả năng di động,
chỉ 1 số ít loài là không có khả năng này.
Nguyên tắc: Tiên mao (lông roi) là những sợi lông dài, uốn khúc, mọc ở mặt
ngoài một số vi khuẩn và có tác dụng giúp chúng có thể chuyển động. Khả năng này
có thể đƣợc quan sát dựa vào sự sinh trƣởng và di dộng của vi khuẩn vào bên trong
môi trƣờng thạch mềm.
Môi trƣờng: Sử dụng môi trƣờng MRS bán lỏng (0,15 - 0,4% thạch). Mỗi
ống nghiệm chứa 5ml môi trƣờng. Khử trùng ở 121oC/15 phút. Làm nguội ở trạng
thái thẳng đứng.

78
Cách tiến hành: Đâm sâu đầu que cấy thẳng có chứa tế bào vi khuẩn lactic
xuyên vào giữa môi trƣờng trong ống nghiệm đến độ sâu khoảng 2ml. Chú ý đƣa
que cấy vào chính giữa ống nghiệm. Nuôi cấy ở nhiệt độ 30 - 370C trong 48 - 56
giờ. Quan sát vị trí sinh trƣởng của vi khuẩn trong ống nghiệm. Nếu tế bào mọc
thẳng theo vạch que cấy thì không có khả năng di động, ngƣợc lại nếu tế bào mọc
loang xung quanh vạch que cấy thì có khả năng di động.
4.4. Xác định khả năng phân giải tinh bột
Cấy chấm hoặc cấy vạch vi khuẩn trên môi trƣờng thạch thƣờng có bổ sung
1% tinh bột tan, để ở 370C trong 2 ngày. Đổ thuốc nhuộm Lugol. Chủng nào có hoạt
tính sẽ tạo vòng trong xung quanh hoặc ngay dƣới chân khuẩn lạc.
4.5. Phát hiện vi khuẩn Sphingomonas
Ngoài khả năng phát hiện vi khuẩn Sphingomonas dựa vào khả năng di động rất
nhanh của chúng khi soi dƣới kính hiển vi còn có thể phân biệt đƣợc chúng khi cấy
trên môi trƣờng King‟s B: Proteose peptone - 20g; Glycerol - 10ml; K2HPO4 -
1,5g; MgSO4 -1,5g; thạch - 18g; nƣớc - 1000ml. Trên môi trƣờng này, đa số các
chủng vi khuẩn Sphingomonas đều phát quang dƣới đèn cực tím (tuy nhiên vẫn có 1
số chủng không phát quang).

79
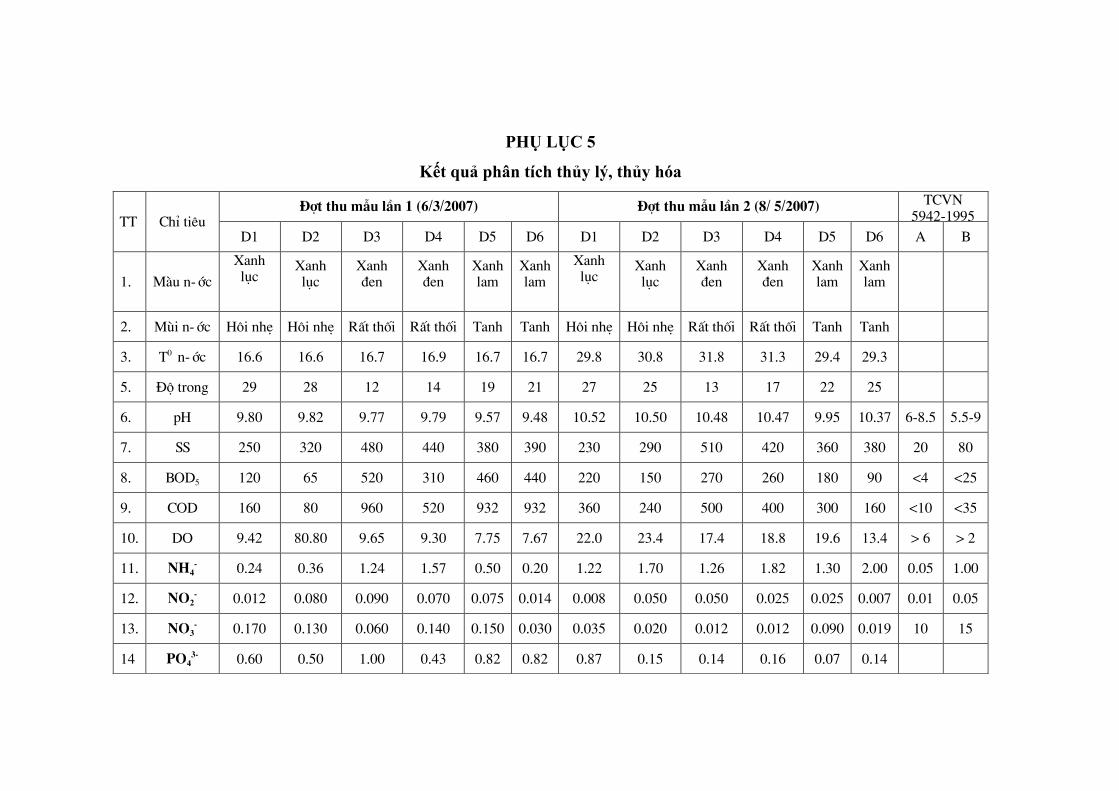
PHỤ LỤC 5
Kết quả phân tích thủy lý, thủy hóa
TT ChØ tiªu §ît thu mÉu lÇn 1 (6/3/2007) §ît thu mÉu lÇn 2 (8/ 5/2007) TCVN
5942-1995
D1 D2 D3 D4 D5 D6 D1 D2 D3 D4 D5 D6 A B
1. Mµu níc
Xanh lôc
Xanh lôc
Xanh ®en
Xanh ®en
Xanh lam
Xanh lam
Xanh lôc
Xanh lôc
Xanh ®en
Xanh ®en
Xanh lam
Xanh lam
2. Mïi níc H«i nhÑ H«i nhÑ RÊt thèi RÊt thèi Tanh Tanh H«i nhÑ H«i nhÑ RÊt thèi RÊt thèi Tanh Tanh
3. T0 níc 16.6 16.6 16.7 16.9 16.7 16.7 29.8 30.8 31.8 31.3 29.4 29.3
5. §é trong 29 28 12 14 19 21 27 25 13 17 22 25
6. pH 9.80 9.82 9.77 9.79 9.57 9.48 10.52 10.50 10.48 10.47 9.95 10.37 6-8.5 5.5-9
7. SS 250 320 480 440 380 390 230 290 510 420 360 380 20 80
8. BOD5 120 65 520 310 460 440 220 150 270 260 180 90 <4 <25
9. COD 160 80 960 520 932 932 360 240 500 400 300 160 <10 <35
10. DO 9.42 80.80 9.65 9.30 7.75 7.67 22.0 23.4 17.4 18.8 19.6 13.4 > 6 > 2
11. NH4- 0.24 0.36 1.24 1.57 0.50 0.20 1.22 1.70 1.26 1.82 1.30 2.00 0.05 1.00
12. NO2- 0.012 0.080 0.090 0.070 0.075 0.014 0.008 0.050 0.050 0.025 0.025 0.007 0.01 0.05
13. NO3- 0.170 0.130 0.060 0.140 0.150 0.030 0.035 0.020 0.012 0.012 0.090 0.019 10 15
14 PO43- 0.60 0.50 1.00 0.43 0.82 0.82 0.87 0.15 0.14 0.16 0.07 0.14
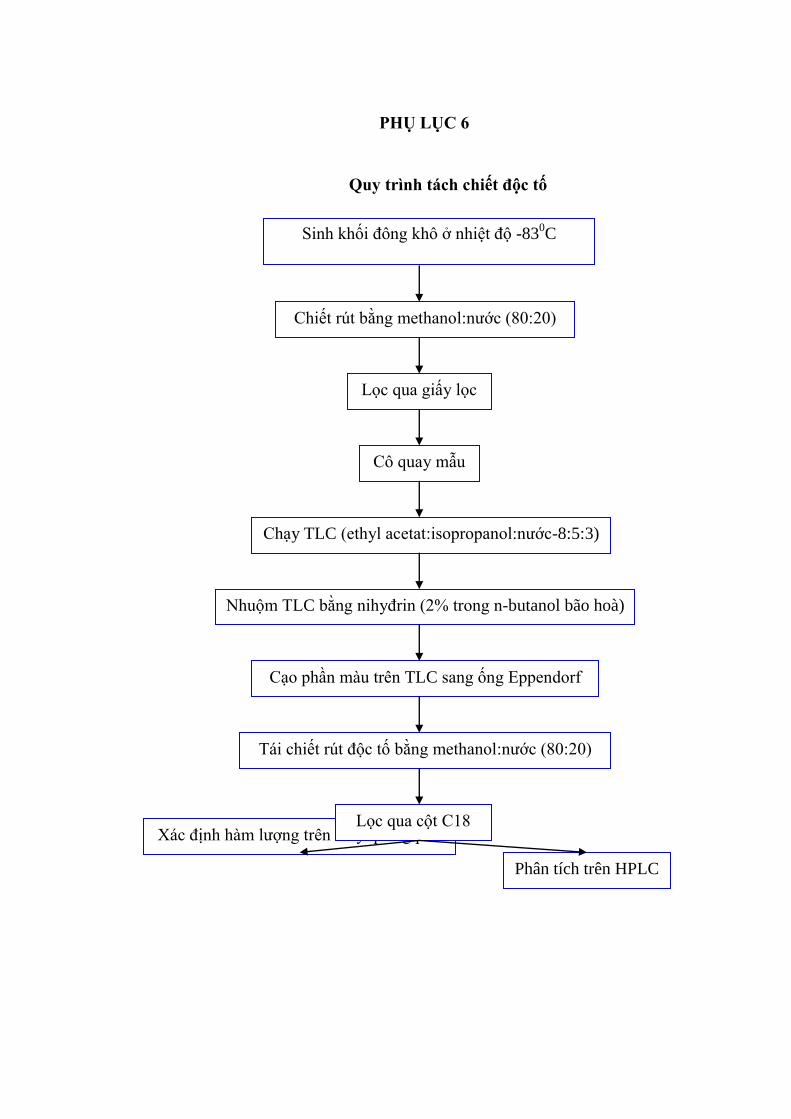
PHỤ LỤC 6
Quy trình tách chiết độc tố
Xác định hàm lƣợng trên máy quang phổ
Phân tích trên HPLC
Lọc qua cột C18
Tái chiết rút độc tố bằng methanol:nƣớc (80:20)
Cạo phần màu trên TLC sang ống Eppendorf
Nhuộm TLC bằng nihyđrin (2% trong n-butanol bão hoà)
Chạy TLC (ethyl acetat:isopropanol:nƣớc-8:5:3)
Cô quay mẫu
Lọc qua giấy lọc
Chiết rút bằng methanol:nƣớc (80:20)
Sinh khối đông khô ở nhiệt độ -830C
Sinh khối đông khô ở nhiệt độ -83ºC
PhiÕu ®¨ng ký
kÕt qu¶ nghiªn cøu c¸c ®Ò tµi khcn
Tªn ®Ò tµi: “Nghiªn cøu ®Æc ®iÓm sinh häc vµ th¨m dß kh¶ n¨ng ph©n gi¶i
sinh häc ®éc tè cña mét sè chñng Microcystis ph©n lËp ë hå Hoµn KiÕm, Hµ Néi”.
M· sè: QG 07 - 24
C¬ quan qu¶n lý ®Ò tµi: §¹i häc Quèc gia Hµ Néi
§Þa chØ: 144 Xu©n Thuû, CÇu GiÊy, Hµ Néi
§iÖn tho¹i: 7548664
C¬ quan chñ tr× ®Ò tµi: ViÖn vi sinh vËt vµ C«ng nghÖ Sinh häc
§Þa chØ: 144 Xu©n Thuû, CÇu GiÊy, Hµ Néi
§iÖn tho¹i: 7547907
Tæng kinh phÝ thùc chi: 100.000.000 ®
Trong ®ã: - Tõ ng©n s¸ch Nhµ níc: 100.000.000 ®
- Nguån kh¸c: 0
Thêi gian nghiªn cøu:
Thêi gian b¾t ®Çu: 05/2007
Thêi gian kÕt thóc: 05/ 2009
Tªn c¸c c¸n bé phèi hîp nghiªn cøu:
- Chñ tr× ®Ò tµi: TS. NguyÔn ThÞ Hoµi Hµ
- Nh÷ng ngêi tham gia: CN. Lu ThÞ Thuú Giang, CN. NguyÔn Quang Huy, CN.
Lª Trung Thuû, CN. Ph¹m ThÞ BÝch §µo, Th¹c sü. TrÇn ThÞ §iÖp
Sè ®¨ng ký ®Ò tµi :
Ngµy
Sè chøng nh©n ®¨ng ký
KQNC
Ngµy
B¶o mËt:
A. Phæ biÕn réng r·i
B. Phæ biÕn h¹n chÕ
C. B¶o mËt
Tãm t¾t ý nghÜa, kÕt qu¶ nghiªn cøu (kÓ c¶ kÕt qu¶ ®µo t¹o):
ý nghÜa:
Th«ng qua c¸c ho¹t ®éng cña ®Ò tµi sÏ gióp cho ngêi d©n xung quanh hå nãi
riªng vµ ngêi d©n Hµ Néi nãi chung nhËn thøc ®îc r»ng m«i trêng cña hå cÇn
®îc b¶o vÖ, tríc hÕt lµ v× lîi Ých cña chÝnh hä, cña Thñ ®« ngh×n n¨m v¨n hiÕn,
®ång thêi còng chÝnh lµ tµi s¶n chung mµ ai còng ph¶i cã tr¸ch nhiÖm cÇn ph¶i b¶o
vÖ vµ gi÷ g×n. C¸c loµi vi khuÈn lam ®îc ph©n lËp, víi lý lÞch cô thÓ sÏ lµm tiÒn ®Ò
quan träng cho nh÷ng nghiªn cøu tiÕp theo, nh»m gãp phÇn x©y dùng m«i trêng hå
s¹ch, an toµn vµ bÒn v÷ng.
MÆt kh¸c sÏ bæ sung vµo danh môc c¸c loµi vi khuÈn lam g©y ®éc níc ngät
cho ViÖt Nam. §Ò tµi ®îc thùc hiÖn sÏ gióp nhãm c¸n bé nghiªn cøu hiÓu s©u h¬n
vÒ ®Æc ®iÓm sinh häc cña vi khuÈn lam g©y ®éc Microcystis, tõ ®ã t×m c¸c kh¶ n¨ng
cã thÓ øng dông ®Ó h¹n chÕ t¸c ®éng xÊu g©y ra bëi Microcystis vµ ®éc tè
microcystin cña chóng trong c¸c hå, thuû vùc nuíc ngät trªn ®Þa bµn Hµ Néi vµ c¸c
tØnh miÒn B¾c nãi chung.
KÕt qu¶:
- Hồ Hoàn Kiếm đang ở tình trạng ô nhiễm, hàm lƣợng COD, BOD
vƣợt tiêu chuẩn Việt Nam, TCVN 4942- 1995 (loại B) 20 lần. Với giá trị NH4+ từ

0,129- 0,699 (mg/l), NO2- từ 0.012- 0,05 (mg/l), NO3
- từ 0,047- 0,129 (mg/l), PO4
3-
từ 0,08- 0.64 (mg/l) chứng tỏ hồ có hàm lƣợng dinh dƣỡng N, P khá cao.
- Đã phân lập thuần khiết và nuôi cấy 10 chủng vi khuẩn lam thuộc chi
Microcystis theo phƣơng pháp của Shirai có cải tiến. Môi trƣờng thạch agarose B12
với tỷ lệ 0,4% là môi trƣờng thích hợp nhất cho việc phân lập Microcystis.
- Các chủng Microcystis đều sinh trƣởng tốt trên môi trƣờng Bold 3N với số
lƣợng tế bào đạt từ 32,94 – 68,6 ×106 tb/ml
sau 15 đến 19 ngày nuôi cấy. Đặc biệt
chủng LT2 số lƣợng tế bào đạt cao nhất là 68,6 ×106
tb/ml sau ngày thứ 19. Trên
môi trƣờng J, các chủng Microcystis sinh trƣởng số lƣợng tế bào của các chủng đạt
từ 40,42 – 76,46 ×106 tb/ml
sau 15 đến 19 ngày nuôi cấy. Trong đó chủng LT2 vẫn
phát triển với số lƣợng tế bào đạt cao nhất 76,46 ×106 tb/ml
sau ngày thứ 19. Trên
môi trƣờng B12, số lƣợng tế bào đạt từ 35,36 – 54,78 ×106 tb/ml
sau 12 đến 15
ngày nuôi cấy. ở môi trƣờng này chủng LT8 phát triển với mật độ tế bào đạt cao
nhất 54,78 ×106 tb/ml
sau ngày thứ 15.
- Có 7 loại axít béo không no dạng C15, C17 và C19 trong tế bào 2 chủng
Microcystis LT2 và LT8. trong đó thành phần axit Palmitic acid (C17H34O2) chiếm
lƣợng lớn nhất với 1,302 μg/mg trọng lƣợng khô ở chủng LT2, chủng LT8 là
0,9683 μg/mg trọng lƣợng khô. Kết quả cho thấy có sự sai khác nhau về lƣợng của
7 loại axit béo có ở trong tế bào 2 chủng LT2 và LT8.
- Dựa vào phƣơng pháp phân loại truyền thống theo phân loại hình thái học
và phƣơng pháp phân loại hiện đại dựa trên phân tích trình tự rADN 16S, 2 chủng
vi khuẩn LT 2, LT8 đƣợc xác định là Microcystis aeruginosa, 2 chủng vi khuẩn
nƣớc N4 và N7 đƣợc xác định thuộc loài Sphin Sphingomonas mali và
Sphingomonas asaccharolytica
- Dựa vào kết quả phân tích trên HPLC, hàm lƣợng độc tố MC – RR đạt lớn
nhất là 512,8 ng/mg trọng lƣợng khô, độc tố MC – YR là 236,8 ng/mg và thấp nhất
là độc tố MC – LR: 114,25 ng/mg.
- Chủng vi khuẩn Sphingomonas tuyển chọn đƣợc có khả phân giải
microcystin khá nhanh. Ở nồng độ microcystin 3 µg/l thời gian bán phân giải là
1,34 giờ và với nồng độ 20µg/l, thời gian là 3,43 giờ.

- Nhiệt độ thích hợp cho sự phân giải microcystin của các chủng vi khuẩn
Sphingomonas mali N4 và Sphingomonas asaccharolytica N7. Sau 7 giờ sự phân
giải gần nhƣ hoàn toàn là 100%.
Các công trình công bố và sản phẩm đào tạo
Công trình công bố
- Phạm Thị Mai, Phạm Tiến Đức, Vũ Thị Lan Anh, Nguyễn Thị Hoài Hà
2008. Đánh giá sự biến động số lượng theo mùa và theo tầng nước của vi khuẩn
lam Microcystis aeruginosa ở hồ Hoàn Kiếm, Hà Nội, Tạp chí Khoa học Đại học
Quốc gia Hà Nội. Khoa học Tự nhiên và Công nghệ 24 số 15. Tr 125 – 128.
- Phạm Thị Mai, Phạm Tiến Đức, Vũ Thị Lan Anh, Nguyễn Thị Hoài Hà
2008. Tìm hiểu khả năng phân giải độc tố của vi khuẩn lam Microcystis trong hồ
Hoàn kiếm bằng vi khuẩn. Tạp chí Khoa học Đại học Quốc gia Hà Nội. Khoa học
Tự nhiên và Công nghệ 24 số 15. Tr 119 – 122.
Đào tạo 4 Cử nhân
- Bƣớc đầu phân lập, tuyển chọn một số chủng vi khuẩn có khả năng phân
giải độc tố của vi khuẩn lam Microcystis ở hồ Hoàn Kiếm, Hà Nội. 2007. Khoá
luận tốt nghiệp hệ đại học chính quy của Đặng Thị Kim Anh.
- Bƣớc đầu đánh giá khả năng phân giải độc tố microcystin của vi khuẩn lam
Microcystis aeruginosa bằng vi khuẩn Sphingomonas. 2008. Khoá luận tốt nghiệp
hệ đại học chính quy của Nguyễn Thị Hoài.
- Khảo sát sự biến động theo mùa của vi khuẩn lam Microcystis aeruginosa ở
hồ Hoàn Kiếm và khả năng ức chế sinh trƣởng của chúng bằng dịch chiết vỏ chuối,
vỏ quýt. 2009. Khoá luận tốt nghiệp hệ đại học chính quy của Lê Thị Trang.
- Phân lập, nghiên cứu một số đặc tính của tảo độc Microcystis tách chiết độc
tố và thăm dò biện pháp xử lý.2009. Đồ án tốt nghiệp của Nguyễn Hữu Hiếu
Đào tạo 1 Thạc sỹ
Phân giải tế bào Microcystis và sự phân huỷ độc tố microcystin bằng vi
khuẩn nƣớc. 2007 - 2009.
Những đóng góp của đề tài:
- Là công trình đầu tiên tách và lƣu giữ đƣợc chủng vi khuẩn lam
Microcystis.
- Phân lập đƣợc 2 chủng vi khuẩn thuộc chi mới Sphingomonas có khả năng
phân giải độc tố microcystin của vi khuẩn lam Microcystis.
- Xây dựng đƣợc quy trình tách chiết độc tố microcystin.
KiÕn nghÞ vÒ quy m« vµ ®èi tîng ¸p dông kÕt qu¶ nghiªn cøu:
Chøc vô Chñ nhiÖm ®Ò
tµi
Thñ trëng c¬
quan chñ tr× ®Ò tµi
Chñ tÞch héi
®ång ®¸nh gi¸
chÝnh thøc
Thñ trëng c¬ quan
qu¶n lý ®Ò tµi
Hä vµ tªn
NguyÔn ThÞ
Hoµi Hµ
Häc vÞ
TiÕn sü
Ký tªn
§ãng
dÊu




















![akciosujsag.hu - AlphaZoo, 2011.07.08-07.24[1]](https://img.pdfslide.net/doc/110x75/577d276a1a28ab4e1ea3de3a/akciosujsaghu-alphazoo-20110708-07241.jpg)







