Embed Size (px)
Citation preview

THE JOURNAL OF BIOLOGIC.~~ CHEMISTRY Vol. 248, No.22, Issue of November 25, pp.7754-7761, 1973
Printed in C.S.A.
Binding of Cytochalasin D to Platelet and Muscle Myosin*
(Received for publication, April 17, 1973)
ELENA I'USZKIS, SAUL PUSZKIN, LAN WEI Lo, AND S. W. TARTENBAUM~
From the Departments of Medicine and Pathology, Mount Sinai School of Medicine, New York 10029, and the Department of Microbiology, Columbia University, Sew YOTk 1003~?
SUMMARY
Human blood platelets exposed to 2 UM tritiated cyto- chalasin D showed maximum incorporation within 5 min. Tritiated cytochalasin D was found to be bound to thrombos- thenin M (platelet myosin), when subjected to gel exclusion chromatography in 0.6 M KCl. Using a Millipore filtration technique, binding at low ionic strength of tritiated cyto- chalasin D to purified preparations of contractile proteins, showed that platelet thrombosthenin M and muscle myosin, but not actin, was involved (0.9 mole of cytochalasin D per mole of protein). Prior treatment of these proteins with nonradioactive cytochalasin D, followed by exposure to the labeled alkaloid, showed little exchange under these condi- tions. Binding data gave a value of 0.99 f 0.11 sites per 4.6 X 10” g of muscle myosin, an average intrinsic associa- tion constant of 6.2 X lo7 M-I with an heterogeneity index of 1. Although the Ca2+- or Mg2+-stimulated ATPase activities of actomyosin from platelets (thrombosthenin) or muscle were not inhibited by 2 UM cytochalsin D, this concentration depressed thrombosthenin M and myosin ATPase. Recom- bination experiments indicated that actin and cytochalasin D compete for a binding region on myosin. The inhibitory effect of cytochalasin D on myosin ATPase was restored following dialysis against 0.6 M KCl; with concomitant re- moval of the drug. Furthermore, quantitative measurements of superprecipitation indicated that, although cytochalasin D did not interfere with dissociation of actomyosin by MgATP, increasing concentrations of cytochalasin D inhibited reasso- ciation. Labeled cytochalasin D associated with thrombos- thenin M could be removed by solvent extraction on thin layer chromatography and by sodium dodecyl sulfate gel electrophoresis, suggesting that the bond between cyto- chalasin D and thrombosthenin M or muscle myosin is not covalent.
The cytochalasins are a class of low molecular weight fungal metabolites that adversely affect a variety of cell functions (1, 2).
* This work was supported by Grants HL 12443, CA-11816, HD-03972, HL-05488, and Al-06801 from the United States Public Health Service and by Grant VC-76 from the American Cancer Society.
$ Present address, State University of New York, College of Environmental Science and Forestry, Syracuse, N. Y. 13210.
Four major cytochalasins, designated A, 13, C, and D, have been isolated and, although structurally similar, the C and D types exceed the A and B types in potency by at least 1 order of mag- nitude (3). Carter (3) first reported that exposure of fibro- blasts in culture to low concentrations of cytochalasin 13 resulted in inhibition of cytokinesis, membrane ruffling, and locomotion. Higher concentrations of these drugs can actually mediate nu- clear extrusion in a number of cultured mammalian cell lines (4). This led to several studies on the influence of cytochalasin B on those morphogenetic processes associated with cell motilit,y (5-7). A diverse number of morphological observations have accumulated concerning the action of cytochalasin B, including inhibition of secretory processes (8, 9), phagocytosis (lo-la), chemotasis (13), pigment migration (14, 15), and many others (.16-23). All of these functions, however, may be interrelated through the common denominator of the contractile mechanism of living cells.
Initial studies concerning the locus of action of cytochalasin B were morphological, and Schroeder (24), studying sea urchin and HeLa cells, suggested that cytochalasin deranged the 40- to 50-A contractile ring microfilaments. Bluemink (25) concluded, with drug-treated Xenopus eggs, that the site of cytochalasin 13 action was at the cytoplasmic membrane. Although a good deal of evidence was adduced in favor of inhibition of microfilament function by cytochalasin B, ot.her data raised doubts concerning this conclusion (17, 19). Sanger and Holtzer (26) demonstrated mucopolysaccharide biosynthesis to be inhibited by cytochalasin 13. Later experiments (27, 28) indicated that cytochalasin B inhibited transport of monosaccharides and amino sugars across cell membranes. It has also been noted that cytochalasin B inhibited formation of COa from labeled glucose (12, 27, 29).
The foregoing studies employed cytochalasin B concentrations in the range of 1 to 10 PM, and more recent reports dealt with the mode of action of 1 mM on the biochemistry of contraction. Spudich and Lin (30) showed both a drop in viscosity when cyto- chalasin B was added to muscle actomyosin and inhibition of the ATPase of an actin-heavy meromyosin (A-HMM) complex. Myosin was unaffected by cytochalasin 13. G-actin was able to polymerize in the presence of cytochalasin B, but reached only a low viscosity value comparable to F-actin plus drug. These data suggested that cytochalasin B reacted with actin rather than myosin, and actually competed with myosin for actin bind- ing. On the other hand, Forer el al. (31) did not demonstrate cytochalasin B inhibition of A-HMMI to form arrowheads and
1 The abbreviations used are: A-HMM, actin-heavy meromy- osin; HMM, heavy meromyosin.
7754
by guest on May 1, 2018
http://ww
w.jbc.org/
Dow
nloaded from

7755
did not notice breakdown of F-actin, inhibition of ATP-induced release of HMM from the A-HMM, or any effect of cytochalasin B on the G-actin to F-actin transformation.
Thus the mode of action of cytochalasins is as yet unclear and published data have led to conflicting assumptions. Experi- ments now reported concern the effect of the relatively unstudied congeneric cytochalasin D on a target site within muscle and platelets, the contractile protein myosin and thrombosthenin M. The availability of high specific activity tritiated cytochalasin D (32) has allowed the interaction of this compound to be studied stoichiometrically.
NATERIALS AND METHODS
Cytochalasin D was isolated from a strain of Zygosporium masonii, as described previously (32). The tritiated drug was prepared by the procedure of Hembree et al. (33), which involves microwave discharge-catalyzed exchange with tritium gas. It was purified by repeated preparative thin layer chromatography, utilizing three solvent systems, to constant specific activity (0.610 mCi per mg). The labeled drug was 98% homogeneous, as judged by analytical combined thin layer chromatography- radiochemical assay. Details of the preparation, purification, and pattern of labeling will be published elsewhere.2 This sample of [%]cytochalasin D was fully biologically active in a sensitive HeLa cell assay3 in which the dihydroderivative and the acetyl derivative, prepared from unlabeled cytochalasin D, were shown to possess at the same molar concentration, 12 and 4%, respectively, as the controls.
Normal human blood was collected into 34 of its volume of acid-citrate-dextrose (34), and platelets were prepared by differ- ential centrifugation (35). Thrombosthenin or thrombosthenin RI was extracted from platelets pooled from three or four donors. Sephadex G-200 and Sephadex G-25 were obtained from Phar- macia Co. Bovine serum albumin was obtained from Pentex. Adenosine 5’-triphosphate-disodium salt (ATP) was purchased from Sigma. Cleland’s reagent was obtained from Calbiochem.
Prolein Extraction-Thrombosthenin was extracted from plate- lets by the method of Bettex-Galland and Liischer (36). Throm- bosthenin M was prepared by the method of Puszkin et al. (37). These proteins were also extracted after prior exposure of plate- lets to 2 PHI [31!I]cytochalasin D. Muscle actomyosin (38), myo- sin (39), and actin (40) were prepared from fresh rabbit back muscle. Protein concentrations were measured by the method of Lowry et al. (41) and standardized with bovine serum albumin dissolved in 0.1 or 0.6 M KCl-0.05 M Tris, pH 7.2.
Determination oj ATPase Activity-ATPase activity of plate- let proteins was determined by the release of inorganic phos- phorus (Pi) from ATP according to Marsh (42) and adapted (43) to detect as little as 3 nmoles of Pi. Each milliliter of final assay mixture contained 0.04 M imidazole buffer, pH 6.8, 0.06 M KCl, 0.5 mM ATP, 1 mM MgClZ or CaClZ, and 0.1 mM ouabain. The amounts of proteins added to the final mixture were 0.1 mg of thrombosthenin or thrombosthenin M, 0.01 mg of actomyosin or myosin and 0.05 mg of actin. Assays were performed on each protein alone and after preincubation at room temperature for 2 min with 2 pM of cytochalasin D. The reaction was stopped by the addition of 0.4 ml of a 20% trichloroacetic acid per ml mixture. Blanks consisted of protein inactivated by trichloro- acetic acid prior to addition of ATP. ATPase activity was
2 L. W. Lo, R. E. Ehrenkaufer, S. W. Tanenbaum, and A. P. Wolf, manuscript in preparation.
3 J. Tannenbaum and S. W. Tanenbaum, unpublished obser- vations.
estimated as the difference between Pi at 30 min and Pi at zero time for platelets and zero to 15 min for muscle proteins.
Incorporation of [3H]Cytochalasin D into Platelets-Platelet concentrates from individual donors were incubated with [%- cytochalasin D at a final concentration of 2 P&Z. These platelet suspensions had a packed platelet volume of 7 to 10 “/;, and con- tained from 0.97 to 1.29 X 1O1” platelet per ml. ‘Two-milliliter aliquots removed at intervals between 0 and 60 min were added to 10 ml of Ringer’s solution. After centrifugation at 1465 x g, supernatant fiuid was removed and a platelet pellet was sus- pended in 5y0 trichloroacetic acid. The resulting precipitate was washed and dried, and glutamic acid, aspartic acid, gluta- mine, and asparagine were separated from the trichloroacet,ic acid supernatant fluid (35). Aliquots of plasma-Ringer’s super- natant fluid, platelets of trichloroacetic acid-soluble fraction, trichloroacetic acid-precipitated proteins, and separated amino acids were transferred to vials containing lO0/o irnphthalene, 0.4% 2,5-diphenyloxazole, and 0.03 y. bis-methylstyryl ben- zene, 10 ml per vial. Radioactivity was determined by a Pack- ard Tri-Carb liquid scintillation spectrometer n-here counting efficiency was 34% and quench corrections were made with internal standards.
Protein Binding oj Cytochalasin D-Intact platclcts preincu- bated with [3H]cytocllalasin D were suspended in Wcbcr-Eddsal (36) solution and disrupted in a Parr cell-disruption bomb for 30 min at 4” under NZ pressure of 1,200 p.s.i. [311]C~t,oc~llalasin D bound by extracted thrombosthenin or thrombosthcnin ;\I was determined after filtration on Sephadex G-200 by applying 2 to 3 mg of protein to a column (1.5 X 35 cm) previously equil- ibrated with 0.6 M KCI-Tris-HCI buffer, ~1-1 7.2. I~XIluent was collected in l-ml fractions and the degree of binding was calcu- lated as radioactivity per mole of protein. Capacity to bind [3H]cytochalasin D of previously isolated thrornbosthcnin or thrombosthenin Xl from platelets, and of actomyosin and myosin from muscle, was determined in a similar manner.
Binding of [3H]cytochalasin D to isolated thrombosthenin and thrombosthenin M from platelets and actomyosin, myosin, and actin from muscle was also studied under low ionic concentra- tions using 0.45-p Millipore filtration (Millipore Corporation). Each protein (0.1 mg) was incubated at room temperature with 0.04 M imidazole buffer (pH 6.8), 1 mM CaCl2, and 0.32 pn% [311]- cytochalasin D. After 5 min of incubation, the mixture was filtered through Millipore and both protein concentration and radioactivity were determined before and after filtration. Be- cause of the lower molecular weight of G-actin, polymerization was induced after incubation with 0.1 I\I KC1 plus 0.2 m&I ATP. In addition, muscle G-actin binding of [3H]cytocllalasin D was studied by gel filtration on Sephadex G-25 columns (1 x 20 cm) equilibrated and eluted with 0.2 mM ATP plus 0.2 m>r nscorbate, pH 7.5.
Experiments were also carried out to determine the concen- tration dependence of binding of [3H]cytochalasin I> to muscle myosin. Increasing concentrations of cytochalasin D were used to give a final concentration of 0.2 to 0.8 PM. From the [3H]- cytochalasin D in the absence of proteins and the corresponding concentration with protein present, the amount bound could be calculated and plotted according to the equation c/[cytochalasin D] = (n - ti)K,. ir is the average number of moles of cyto- chalasin D per mole of protein; [cytochalasin D] is the free drug concentration at equilibrium; K, is the apparent intrinsic asso- ciation constant; and n is the apparent number of equivalent binding sites on each protein molecule (44). As fl/[cytochalasin D] ---f 0, ii approaches n. The molecular weight of myosin was
by guest on May 1, 2018
http://ww
w.jbc.org/
Dow
nloaded from
taken as 460,000. The heterogeneity index a describes the dis- TABLE: I pension of association constants about the average constant; Effect of cytochalasin D on ATPase activity of platelet
K, was calculated from the generalized binding isotherm as contractile proteins
defined by Sips in 1948 (45). For each titration, the log a/ The incubation medium contained 0.04 M imidazolc buffer (pH n - ti was plotted versus log [cytochalasin D] by the method of 6.8), 0.06 M KCl, 0.05 pM ATP, 1 mM Ca2+ or &IgZ+, 0.1 mM ouabain, least squares and was obtained as the slope, and K, from the 0.1 mg of thrombosthenin or thrombosthenin M, and 0.05 mg of
intercent. When ac = 1 all sites have the same association actin. Incubation time was 30 min at 37”.
constant, and decreasing values of o( correspond either to in- creasing heterogeneity in respect to the association constant or to heterogeneity of the cytochalasin D used in the binding studies. Cation II
Reversibility of Cytochalusin D Action-One milligram of myo- sin was preincubated at room temperature with 0.1 M KC1 and 20 PM [311]cytochalasin D. After 5 min of incubation, KC1 concentration was increased to 0.6 M. The mixture was di- M~Z+ I alyzed for 24 hours against several changes of 0.6 M KCl-Tris buffer, pH 7.2, until no detectable radioactivity could be found in the external fluid. ATPase activity was determined on the myosin and reconstituted actomyosin (myosin + actin) prior to and after addition of cytochalasin D and on the 24-hour dialyzate. Ca2+
Acryiamide Disc Electrophoresis-Sodium dodecyl sulfate disc electrophorcsis was performed on single 0.1% sodium dodecyl sulfate, 5% acrylamide gels. Quadruplicates of 125.pg protein samples were run on a Buchler electrophoresis apparatus. ,4 current of 10 ma per gel was applied until the tracking dye reached the bottom of the gel. Duplicate gels were immersed
No. of ,repar- ation
1.45 1.60 0.38 0.32 0.40 1.18 1.23 1.61 1.58 0.45 0.39 0.35 1.15 1.05 0.95 1.05 0.33 0.36 0.51 0.90 0.95 1.07 1.19 0.55 0.49 0.38 1.35 1.28 1.23 1.10 0.41 0.46 0.61 1.50 1.37
1.50 1.70 1.63 0.63 o.ti5 0.93 0.90
1.48 1.53 1.93 0.75 0.71 1.10 1.03 1.05 1.13 1.82 O.G8 0.73 1.18 1.22 1.25 1.08 2.05 0.80 0.85 1.23 1.15
1.89 1.92 2.10 0.79 0.82 1.38 1.26
1 2 3 4
5
1 2
3 4 5
TABLE II
for 1 hour in 1 y0 Amido schwarz in 7 y0 acetic acid and destained Effect of cytochalasin D on Al’Pase activity of muscle aetomyosin, automatically with a diffusion destainer (Hoefer Scientific In- myosin, and reconstituted actomyosin
struments). The remaining pair of gels were cut into l-mm The incubation medium contained 0.04 M imidazole buffer (pH slices and each slice was assayed for its radioactivity. 6.8), 0.06 M KCI, 0.05 mM ATP, 1 rnM Ca2+ or Mg2+, 0.1 my oua-
Xuperprecipitation-Superprecipitation was recorded by meas- bain, 0.01 mg of myosin or actomyosin, and 0.05 mg of actin. urine absorbance of muscle actomvosin suspensions at 620 nm Incubation time was 15 min at 25”. in a Gilford recording spectrophotometer. In a final volume of 1 ml prior to addition of ATP, 0.5 mg of actomyosin was sus- pended in 0.15 M KC1 concentration buffered with 15 to 25 mM
Cation
Tris-acetate, pH 6.8, to obtain a zero setting. Increasing con- centration of cytochalasin D (2, 4, 6, 8 pM) were added to the protein prior to addition of MgATP. At zero time, 1 InM ATP Mg2+ plus 2 mM r\I; 1 .2+ were added to the suspension, and the reaction wsa allowed to proceed at 25”. Clearing was observed as a decrease in absorbance whereas superprecipitation which followed caused increased absorbance.
RESULTS
ATPase Activity and Xuperprecipitation-Changes in t,he ca2+
ATPase activity of the actomyosin complex can reflect altera- tions in the ATPase activity of myosin alone, without influencing the interaction between actin and myosin. Also ATPase ac- tivity can be inhibited by agents which cause actin and myosin
Kio. of prepar- ation
11.9 12.5 13.9 8.9
15.3
11.3 10.2 11.2
7.15 12.8
0.45 0.41 0.35 0.38 0.29 0.55 0.53 0.61 0.57 0.66 0.59 0.61
0.45 0.48 0.50 0.32 0.28 0.36
12.7 1.42 1.33 9.15 0.91 0.98
11.6 1.33 1.5
8.9 0.85 0.79 15.0 1.20 0.99
7.85 0.69 1.1 -
8.9 8.3 7.6
12.48
9.3 7.1
8.1
11.9
11.5 13.1 12.5
9.2:
14
10.2E 9.9
9.0 7.4
13.01
9.3 7.28 8.5 6.47
6.9 11.2
11.3 6.9 8.48
6.01 10.14
5.7
i
to be dissociated. The effect of cytochalasin D on both of these aspects was studied by measuring the influence of the drug on the ATPasc activity of platelet and muscle contractile protein and the effect of cytochalasin D on muscle actomyosin dissocia- tion and superprecipitation by ATP.
/ - -
ATPase activity of actin, myosin, and actomyosin from muscle and thrombosthenin, thrombosthenin M from platelets, and various combinations with and without prior treatment with cytochalasin D is shown in Tables I and II. Cytochalasin D had no effect on either thrombosthenin or muscle actomyosin in regard to Ca*f- and Mg2$-ATPase activity. Cytochalasin D inhibited approximately 90 and 62 y0 of the Ca2+ATPase activity of both myosin and platelet thrombosthenin M, respectively. When myosin and actin or thrombosthenin M and actin were
combined first and cytochalasin D added later to the mixture, no inhibitory effect on the ATPase activity of either actomyosin or actin-thrombosthenin M complexes was seen. In contrast, when untreated actin was added either to cytochalasin D-treated myosin or cytochalasin D-treated thrombosthenin M, actin failed to restore either actomyosin or actinthrombosthenin M ATPase activities. However, when untreated myosin and thrombosthenin M were combined with treated actin, ATPase activity of both actomyosin and actin-thrombosthenin M was restored.
Comparative behavior on superprecipitation of muscle acto- myosin in relation to treatment with or without cytochalasin D
by guest on May 1, 2018
http://ww
w.jbc.org/
Dow
nloaded from
prior addit,ion of Mg”fATP is described in the following esperi- mental data. Results of a typical experiment are presented in Fig. 1. At an ionic strength of 0.15, the clearing phase of acto- myosin was prolonged for 10 min, a condition found necessary to allow cytochalasin D to interact with the myosin moiety. In- creasing concentrations of cytochalasin D progressively pro- longed the clearing phase. Finally, at a concentration of 8 pM,
protein suspension superprecipitation was completely inhibited. Interference by cytochalasin D with formation of actomyosin complex is consistent with results of drug inhibition of ATPase activity.
Incorporalion and Protein-binding oj [3H]Cytochalasin D- Incubation of platelets with [3H]cytochalasin D resulted in rapid
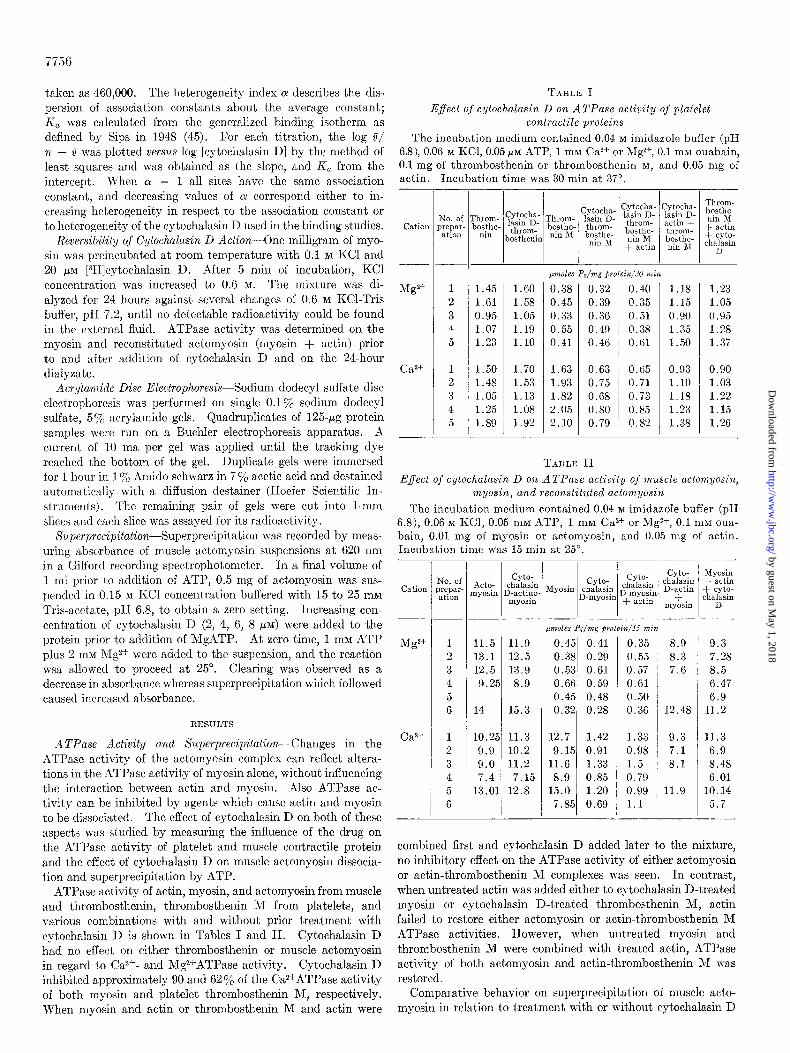
FIG. 1. The effect of cytochalasin D on the clearing phase and superprecipitation of actomyosin. *At zero time 1 rnM ATP and 2 mM Mg2+ were added in a final volume of 1 ml containing 0.5 mg of actomyosin in 0.15 nl KC1 and 15 rnnl Tris, pH 6.8.~ The time course of the turbidity change of each reaction mixture, compared to the initial turbidit,y, was followed. The time at which the clearing phase of each suspension ended is indicated by a small arrow. The large arrows indicate the different concentrations of cytochalasin D used. At 4 /*g of cytochalasin D per ml (8 p~c) superprecipit,ation \\-as completely inhibited. Control run was performed without cgtochalasin D (solid line).
FIG. 2. Incorporation of [3H]cytochalasin D in platelets. Platelet suspension (0.97 to 1.29 X lOlo cells per ml) with 2 PM [3H]cytochalasin D was incubated at 37”. Aliquots of 2 ml were taken at intervals as indicated in the figure and resuspended with 5y0 trichloroacetic acid. Plasma, trichloroacetic acid-soluble supernatant was precipitated and determined for its radioactivity.
7757
incorporation of the label into the cell (Fig. 2). At 5 min, masi- mum platelet labeling was achieved with 63% cellular incorpora- tion. The distribution of the 63% radioactivity within these platelets showed 78% in the trichloroacetic acid-soluble fraction and 22% in trichloroacetic acid-precipitated proteins. NO
radioactivity was found in the studied amino acids. Repeated washes of the precipitated proteins with 57, trichloroacetic acid showed release of radioactivity to the supernatant fluid.
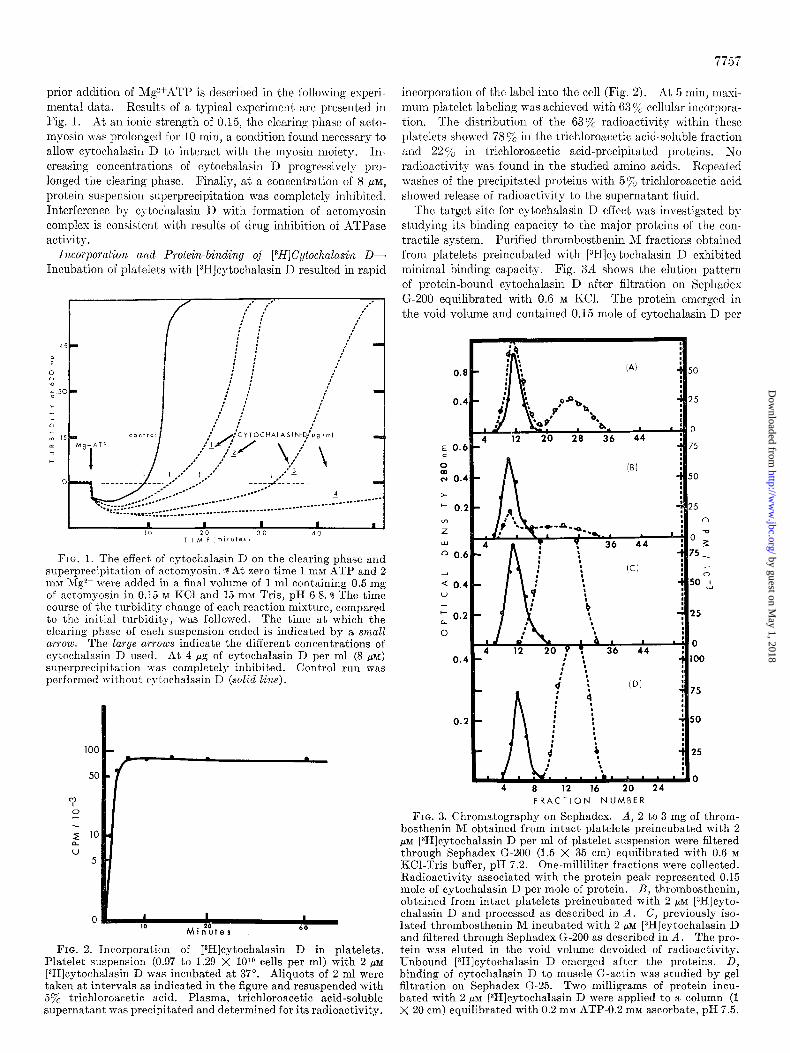
The target site for cytochalasin D effect was investigated by studying its binding capacity to the major proteins of the con- tractile system. Purified thrombosthenin M fractions obtained from platelets preincubat’ed with [3H]cytochalasin D exhibited minimal binding capacity. Fig. 3A shows the elution patt,ern of protein-bound cytochalasin D after filtration on Sephades G-200 equilibrated with 0.6 M KCI. The protein emerged in the void volume and contained 0.15 mole of cytochalasin D per
E 0.6 c
0
F4 0.4
>-
+ 0.1 - v1 Z IL
n 0.6
Q 0.4 ” -
L 0.2 0
4 12 20 28 36 44
4 8 12 lb 20 24
‘5
I
‘5
‘0
15
0
00
75
25
FRACTION NUMBER
FIG. 3. Chromatography on Sephadex. A, 2 to 3 mg of throm- bosthenin M obtained from intact platelets preincubated with 2 PM [3H]cytochalasin D per ml of platelet suspension were filtered through Sephadex G-200 (1.5 X 35 cm) equilibrated with 0.6 M KCl-Tris buffer, pH 7.2. One-milliliter fractions were collected. Radioactivity associated with the protein peak represented 0.15 mole of cytochalasin D per mole of protein. B, thrombosthenin, obtained from intact platelets preincubated with 2 PM [3H]cyto- chalasin D and processed as described in A. C, previously iso- lated thrombosthenin M incubated with 2 pM [3H]cytochalasin D and filtered through Sephadex G-200 as described in A. The pro- tein was eluted in the void volume devoided of radioactivity. Unbound [3H]cytochalasin D emerged after the proteins. D, binding of cytochalasin D to muscle G-actin was studied by gel filtrat,ion on Sephadex G-25. Two milligrams of protein incu- bated with 2 PM [3H]cytochalasin D were applied to a column (1 X 20 cm) equilibrated with 0.2 rnw ATP-0.2 mM ascorbate, pH 7.5.
by guest on May 1, 2018
http://ww
w.jbc.org/
Dow
nloaded from
7758
mole of protein. Unbound cytochalasin D emerged from the column after complete elution of the protein. Thrombosthenin (Fig. 3B) exhibited considerably less binding than thrombosthe- nin M (0.08 mole of cytochalasin D per mole of protein). When, on the other hand, thrombosthenin M was first extracted and purified and then incubated with cytochalasin D, a different pattern emerged. Fig. 3C shows the elution pattern on Sephadex G-200 of purified thrombosthenin M prepared in this way. No radioactivity was associated with the protein peak. Purified muscle actomyosin and myosin incubated with [3H]cytochalasin D also showed no protein binding when high ionic strength con- ditions were set. Sephadex G-25, equilibrated with ATP and ascorbate buffer, was used to study purified muscle G-actin incubated with [3H]cytochalasin D. However, no radioactivity was observed to be attached to the protein (Fig. 30).
Binding of [3H]cytochalasin D to myosin and thrombosthenin M under conditions of low ionic strength was shown by Milli- pore filtration (Table III). Muscle myosin and thrombosthenin
irreversible even after dissociation of cytochalasin D at high KC1 concentration. After incubation with [SH]cytochalasin D, myosin Ca*+ATPase activity was inhibited, and actin could not restore the Mg2+ATPase activity. After dialysis, no measurable residual radioactivity was found to be associated with the pro- tein. The Ca2+ATPase activity of the myosin and the ability of actin to activate MgzfATPase were restored (Table V).
Acrylamide Disc Electrophoresis-When thrombosthenin M obtained from platelets incubated in the presence of [3H]cyto- chalasin D was subjected to sodium dodecyl sulfate acrylamide disc electrophoresis, one major protein staining band of approxi- mately 220,000 daltons and two very weak staining bands of
TABLE IV
Representative data on binding of cytochalasin D to myosin
Incubation medium as in Table III, varying the cytochalasin
D (CD) concentration between 0.2 to 0.8 MM.
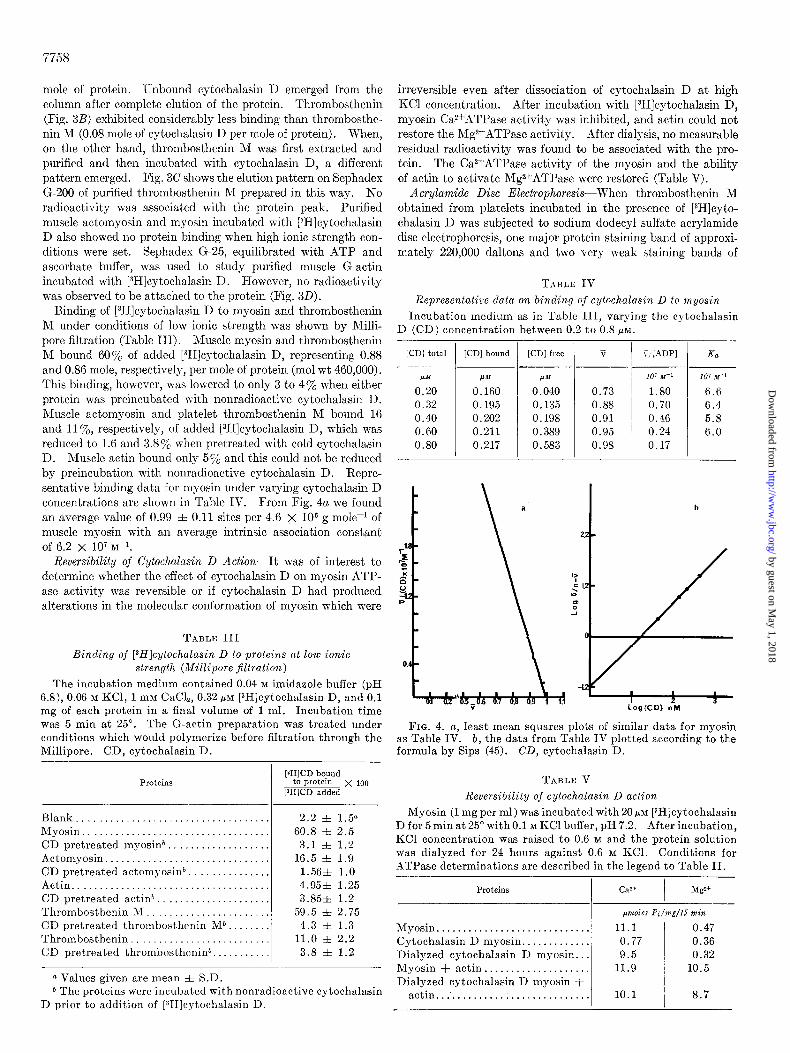
M bound 607% of added [3H]cytochalasin D, representing 0.88 and 0.86 mole, respectively, per mole of protein (mol wt 460,000). This binding, however, was lowered to only 3 to 47, when either protein w-as preincubated with nonradioactive cytochalasin D. Muscle actomyosin and platelet thrombosthenin M bound 16 and 1170, respectively, of added [3H]cytochalasin D, which was reduced to 1.6 and 3.8% when pretreated with cold cytochalasin D. Muscle actin bound only 570 and this could not be reduced by preincubation with nonradioactive cytochalasin D. Repre- sentative binding data for myosin under varying cytochalasin D concentrations are shown in Table IV. From Fig. 4a we found an average value of 0.99 f 0.11 sites per 4.6 X lo6 g mole-l of muscle myosin with an average intrinsic association constant of 6.2 X lo7 M-l.
[CD] total
P’M 0.20 0.32 0.40
0.60 0.80
Reversibility of Cytochalasin D Action-It was of interest to determine whether the effect of cytochalasin D on myosin ATP- ase activity was reversible or if cytochalasin D had produced alterations in the molecular conformation of myosin which were
TABLE III
Binding of [3H]cytochalasin D to proteins at low ionic
strength (Millipore $ltration)
The incubation medium contained 0.04 M imidazole buffer (pH 6.8), 0.06 M KCl, 1 mM CaC&, 0.32 PM [3H]cytochalasin D, and 0.1 mg of each protein in a final volume of 1 ml. Incubation time
was 5 min at 25”. The G-actin preparation was treated under conditions which would polymerize before filtration through the Millipore. CD, cytochalasin D.
FIG. 4. a, least mean squares plots of similar data for myosin as Table IV. b, the data from Table IV plotted according to the formula by Sips (45). CD, cytochalasin D.
Proteins
Blank.............................. 2.2 f 1.50 Myosin............................. 60.8 f 2.5 CD pretreated myosinb.. . . . . . 3.1 f 1.2
Actomyosin......................... 16.5 f 1.9 CD pretreated actomyosinb 1.56& 1.0 Actin............................... 4.95k 1.25
CD pretreated actin*. . . . . 3.85& 1.2
Thrombosthenin M.. _. _. . 59.5 zk 2.75 CD pretreated thrombosthenin Mb.. 4.3 f 1.3 Thrombosthenin . . 11.0 zk 2.2
CD pretreated thrombosthenin*. . 3.8 f 1.2
[ZHICD bound to protein x 100
LfHlCD added
Q Values given are mean f S.D. b The proteins were incubated with nonradioactive cytochalasin
D prior to addition of [3H]cytochalasin D.
[CD] bound
I.mJf
0.160 0.195 0.202
0.211 0.217
[CD] free v
P.W
0.040 0.73 0.135 0.88 0.198 0.91
0.389 0.95 0.583 0.98
V:[ADPl
10’ M-1
1.80 0.70 0.46
0.24 0.17
107 MI-1
6.6 6.4 5.8 6.0
b
/
TABLE V
Reversibility of cytochalasin D action
Myosin (1 mg per ml) was incubated with 20+$ [3H]cytochalasin D for 5 min at 25” with 0.1 M KC1 buffer, pH 7.2. After incubation, KC1 concentration was raised to 0.6 M and the protein solution
was dialyzed for 24 hours against 0.6 M KCl. Conditions for ATPase determinations are described in the legend to Table II.
Proteins I
ca2+ /
Mgz+
p?lZOleS Pi/mg/l5 ??iin
Myosin............................. 11.1 0.47
Cytochalasin D myosin.. . . . . . . . . 0.77 0.36 Dialyzed cytochalasin D myosin.. . 9.5 0.32 Myosin + actin.................... 11.9 10.5
Dialyzed cytochalasin D myosin + actin...‘.......................... 10.1 1 8.7
by guest on May 1, 2018
http://ww
w.jbc.org/
Dow
nloaded from

7759
smaller molecular weight were observed (Fig. 5). The [3H]- cytochalasin D was noted to migrate along with the tracking dye, to the bottom of the gels. Apparently, when sodium dodecyl sulfate depolymerized myosin into it.s subunits, L3H]- cytochalasin D was dissociated from the native protein sub- chain.
Thin Layer Chromatography-The entire radioactivity bound to thrombosthenin M could be removed from aqueous salt solu- tion by organic solvent extraction. Furthermore, thin layer chromatographic analysis of the [3H]cytochalasin D-treated thrombosthenin M complex directly applied to Eastman 6060 silica sheets (solvent, toluene-methanol, 75 : 10) revealed that 90% of the radioactivity placed initially at the origin was located at one spot, RF 0.50, coincident with authentic cytochalasin D migrating on parallel guide strips. These results demonstrated
FIG. 5. Disc gel electrophoresis. Sodium dodecyl sulfate polyacrylamide gel electrophoresis was performed in 5y0 gels. Thrombosthenin M (0.125 mg) obtained from platelets exposed to [3H]cytochalasin D was added to the top of the gel and moved downward toward the-anode.
that the alkaloid bond to the protein was not covalent in nature and also indicated that there had been no metabolism of the drug by the platelets during the uptake experiments.
DISCUSSION
Blood platelets have been reported to possess a considerable amount of an actomyosin-like protein, called thrombosthenin, which is believed to mediate clot retraction. Shepro et al. (46) and Wessells et al. (47), reported inhibition of clot retraction by cytochalasin B and recent work by White (48) extended these observations suggesting that cytochalasin B prevented changes in platelet shape as manifested by a prolongation of the primary aggregation phase that leads to inhibition of the platelet’s clot- retracting capacity.
In the present report, an attempt was made to correlate plate- let function with biochemical changes as manifested by the action of cytochalasins. Cytochalasin D was of interest because of its high potency and its availability as a tritiated compound, [3H]cytochalasin 1).
The effect of cytochalasin D on the contractile mechanisms was studied first by measuring ATPase activity of thrombosthe- nin, thrombosthenin M, myosin, and actomyosin both before and after actin activation and alone or combined with cyto- chalasin D. It was apparent that cytochalasin D exerted con- siderable effect because cytochalasin D-treated myosin and cytochalasin D-thrombosthenin M had their CazfATPase activ- ity inhibited. Furthermore, the addition of actin to either cytochalasin D-myosin or cytochalasin D-thrombosthenin M failed to restore Ca2+- or Mgz+ATPase activity. Interestingly, the reverse combination, untreated myosin or thrombosthenin M with cytochalasin D-treated actin presented normal ATPase activity of the complex molecule. Extracted platelet throm- bosthenin and muscle actomyosin, when incubated with cyto- chalasin D, showed no inhibition of ATPase activity. These data suggest that, when actin and myosin or thrombosthenin M and actin are combined, cytochalasin D does not affect ATPase activity of the complex.
The results obtained by mixing either untreated myosin or cytochalasin D-treated myosin with actin, and of mixing un- treated thrombosthenin M or cytochalasin D-treated throm- bosthenin M with actin suggest that actin and cytochalasin D bind to the same sites of both myosin and thrombosthenin M. Furthermore, it was apparent that when actin and cytochalasin D were added simultaneously to myosin or thrombosthenin M, actin was preferentially bound. However, when myosin or thrombosthenin M sites were previously blocked by cytochalasin D, actin was unable to react with either. These results differ from those of Spudich and Lin (30) who suggested that cyto- chalasin B appeared to interact with actin rather than with myosin. However, conditions of their assay were different; in particular, the high concentrations of cytochalasin B used could have been responsible for nonspecific effects. Spudich has further reported (49) that the activation of muscle HMM by platelet actin is not inhibited by cytochalasin B. Forer et al. (31), using the same drug concentration as Spudich and Lin (30) (1 mM cytochalasin B), concluded that F-actin was un- affected structurally or functionally, and that actin binding and release of HMM was normal. Their experiments were performed either by incubating actin with cytochalasin B to which HMM was added, or by mixing F-actin, cytochalasin B, and HMM. They found no blocking of HMM binding by actin nor inhibition of release of’ HMM from actin when ATP was added. This agrees with our results obtained when in the reaction mixture,
by guest on May 1, 2018
http://ww
w.jbc.org/
Dow
nloaded from

7760
myosin, actin, and cytochalasin D were added simultaneously. Cytochalasin D did not interfere with the formation of the acto- myosin complex. Apparently under these conditions myosin binds preferentially to actin rather than to cytochalasin D.
Restoration of the CaZ+ATPase activity and actin activation of the XIg2+ATI’ase on the dialyzed cytochalasin D-myosin indi- cated that the effect of cytochalasin D on myosin was reversible and that no irreversible alteration in the molecular conformation occurred.
From the results obtained by measuring the effect of cyto- chalasin D on superprecipitation, we found that cytochalasin D did not interfere wit.11 the dissociation of actomyosin into actin and myosin by A!Ig~ATP. Dissociation of actomyosin by hlg- ATP was evident by the decrease in optical density (clearing phase). When the clearing phase was prolonged at 0.15 M KCl, increasing amounts of cytochalasin D were found to inhibit pro- portionally reassociation of myosin to actin as manifested by complete inhibition of superprecipitation with 8 PM cytochalasin D. Wessels et al. (47), using cytochalasin B, failed to observe such an effect because superprecipitation was assessed only quali- tatively and because conditions for clearing could not be estab- lished. Alternatively, differences in potentiation between cyto- chalasin B and cytochalasin D could account for the discrepancy of effects.
Our results are also consistent with those obtained by measur- ing the effect of cytochalasin D on the ATPase activity of throm- bosthenin I\1 and myosin (Tables I and II). On the other hand, dissociation of actomyosin into actin and myosin by ATP at 0.06 M KC1 is unmeasurable. Under these conditions cytochala- sin D apparently could not bind to myosin; therefore, the ATPase activity of actomyosin w-as unaltered.
Intact platelets when incubated with [3H]cytocllalasin D showed rapid incorporation of label, without detectable (thin layer chromatography) metabolic transformation of the drug. Platelet amino acids, including aspartic and glutamic acids and their amides, were not radioactive, which precluded degradation of the alkaloid through the tricarboxylic acid cycle.
Of the total amount of tritiated drug sequestrated by the platelets, 22 y0 was initially located in trichloroacetic acid-precipi- tated fractions. Serial trichloroacetic acid washings from such precipitates were found to contain labeled cytochalasin D; thus, it appears that the denaturing act,ion of trichloroacetic acid af- fected the release of radioactivity from protein binding regions. The quantitative determination of cytochalasin D which could be bound in vitro by platelet- and muscle-derived proteins was difficult to assess by gel filtration because of the high ionic strength which was required (0.6) to maintain these proteins in solution; indeed, contractile proteins extracted from these sources and which were incubated with [3H]cytochalasin D failed to show coincident drug binding upon passage through Sephadex G-200. However, thrombosthenin and thrombosthenin M from platelets with prior exposure to the tritiated drug, when subjected to gel filtration, still contained measurable amounts of drug associated with these protein peaks. More radioactivity was associated with thrombosthenin ?\1 (0.15 mole of cytochalasin D per mole of protein) than with thrombosthenin (0.08 mole of cytochalasin D per mole of protein).
Consequently, other means were sought to determine binding at lower ionic strength. Initial attempts at equilibrium dialysis over a period of 24 hours proved unsuccessful, probably due to protein denaturation over this extended period of time. Binding of cytochalasin D by thrombost,henin, thrombosthenin M, acto-
Nillipore filtration. This evolved into the method of choice because it was rapid and, while it could be shown that free cyto- chalasin D minimally contaminated the filters, the proteins, as present in their aggregated states, were retained. To retain G-actin on the filter, it was necessary first to polymerize it to F-actin. 3Iuscle myosin and thrombosthenin RI were shown by this method to bind 0.88 and 0.86 mole of cytochalasin D per mole of protein, respectively. Proteins first exposed to non- radioactive cytochalasin D were completely blocked from further binding of [3H]cytochalasin D. These experiments indicated that cytochalasin D binding is nonexchangeable at low ionic strength and, more importantly, that binding of [3H]cytochnlasin D was not due to nonspecific adsorption. Yet, a small amount of cytochalasin D was found bound to actomyosin from muscle and thrombosthenin from platelets. This result was interpreted as probably being due to the presence of dissociated myosin or thrombosthenin 31 in these preparations. Negligible amounts of radioactive alkaloid binding was associated with actin.
The interdependence of the two actin-binding sites has been reported by Barany (50). Young’s (51) work has suggested that only one of the two subunits of the head portion of HMN may be able to bind to F-actin at any given time. The results ob- tained from kinetic studies of binding using different concentra- tions of cytochalasin D gave values of a maximal binding site of approximately 1. Therefore, it is possible that, if cytochalasin D binds to one of the actin-binding sites on myosin, the reactivity of the other actin binding site may be affected, resulting in the inability of actin to interact. Another possibility would be that cytochalasin D possesses two reactive groups capable of interact- ing with both sites on the myosin molecule. The fact that the heterogeneity index is 1 suggests that cytochalasin D binds to a homogeneous reactive site.
The effect of cytochalasin D on HMhI was not studied. Ef- forts to prepare this protein by adapting methods used for muscle to human platelets failed to yield the like protein fragment; although a limited amount of platelets from human donors can be obtained, efforts toward obtaining platelet H;\IN fragment are still in progress. Since subcellular localization of throm- bosthenin in platelets is still unclear, it therefore cannot be concluded as to whether cytochalasin D penetrates the cell mem- brane to bind to the cytoplasmic thrombost,henin III or whether it binds to thrombosthenin M possibly located at the cell surface. It should be noted parenthetically, in work done with HeLa and MDBK cells, that radioautographic microscopy has provided evidence that [3H]cytocllalasin D is present throughout the cyto- plasm of these cultured cell lines4
Thin layer chromatography of thrombosthenin ,II obtained after incubation of platelets with [3H]cytocl~alasin D showed that the label could be dissociated from the protein by organic sol- vents. In thin layer chromatography, the disscciated tritium label migrated identically to controls of free [3H]cytochalasin D on parallel lanes. The finding that cytochalasin D bound to thrombosthenin NI was dissociated from protein species in sodium dodecyl sulfate acrylamide electrophoresis also reinforces the notion that cytochalasin D binds noncovalently to throm- bosthenin i\I.
Some speculation on the nature of the binding of cxytochalasin D to myosins is in order. Since the drug is lipophilic, and indeed eontains neither functional nor potentially ionizable groups, hydrophobic bonding would appear to be one mechanism for an attachment which survives gel filtration and dialysis at low ionic
myosin, myosin, and actin was measured by the technique of 4 A. Miranda and G. C. Godman, personal communication.
by guest on May 1, 2018
http://ww
w.jbc.org/
Dow
nloaded from

strengths. It is hypothesized that a site on the multichain
myosin which might involve such bonding could be formed by a
cluster of amino acids at or near the ATPasc catalytic centers, and one which would be rich in residues containing nonpolar side chains. Uernatively, the alkaloid might be bound to myosin
by means of complex formation with a monovalent cation to- gether with a negatively charged group on the protein, since it has been shown t.hat cytochalasin I3 crystallizes as its silver fluoroborate comples (52).
The availability of high specific activity [3H]cytochalasin D has allowed the interaction of this compound to be studied stoichiometrically, thus explaining in part prior phenomena ob- served with congeneric cytochalasins, such as cytochalasin B.
Acknowledgments-We are very much indebted to Mrs. R. Maldonado Muiioz for her outstanding technical assistance and to Dr. S. Kochwn, Dr. Ralph Zaluskv, and Dr. R. Rosenfield for helpful suggestions concerning thismanuscript.
REFERENCES
1. ALDRIGE, D. C., AND TURNER, N. B. (1969) J. Chem. Sot. 22, 923
2. CARTER, S. B. (1972) Endeavor (Engl. Ed.) 31, 77 3. CARTER, S. B. (1967) iVature 213, 261 4. PRESCOTT. D. XI., MEYERSON. D., AND W.~LLICE. J. (1972)
5. Exp. Ce’ll Res. i1, 480 ‘
WESSELLS. N. K.. SPOONER. B. S.. Asn. J.. LUDUENA. M. A..
6. .\SD WR~NN, J.‘(l971) Sc&nce li3, 35i ’
SPOONER, 13. S., .\ND WI”SSELLS, N. K. (1970) Proc. Nat. Acad. Sci. U. 5’. A. 66, 360
7.
8. 9.
MANASEI~, F. J., BURNSIDE, B., AND STROMAN, J. (1972) Proc. Xat. Acad. Sri. U. S. A. 69, 308
SCHOFIELD, J. G. (1971) iVature New Biol. 234, 215 WILLIAMS, J. A., AND WOLFF, J. (1971) Biochem. Biophys. Res.
Commu,L. 44, 422 10.
11.
12.
MALAWIHTA, 8. E., GEE, J. B. L., AND BENSCH, K. (1971) Yale J. Biol. Med. 44, 286
DAVIS, A. T., I<STENSEN, R., AND QUIE, P. G. (1971) Proc. Sot. Ezp. Biol. Med. 137, 161
ZI~MOND, S. H., .IND HIRSCH, J. G. (1972) Exp. Cell Res. 73, 383
13. BI:CI<ER, I(:. I~., D.~vIs, A. T., ESTENSEN, Ii. D., AND QUI, P. G. (1972) J. In,nzu72ol. 108, 396
14. 15. 16. 17.
18.
19. 20.
MALA~ISTB, S. 14:. (1971) Nature 234, 354 MCGUIRE, j., AND MOELLMANN, G. (1972) Science 176, 642 RIDLER. M., AP~I) ShnTn. G. F. (1968) J. Cell Sci. 3, 595 &MM& ti., SHI.;IL1DAN, J. D.,‘ AND' ESTENSEN, RI D. (1971)
Proc. Sot. Erp. Biol. Med. 136, 1158 KRISHAN, A., .1x1) R.IY-CHAUDHURI, R. J. (1969) J. Cell Biol.
43, 618 GOLDMAN, 1:. D. (1972) J. Cell Biol. 62, 246 SPOONER, B. s., Y.\MADA, K. M., AND WESSELLS, N. K. (1971)
J. Cell Biol. 49, 595
21.
22.
23.
24. 25. 26.
27. 28. 29. 30.
31.
32.
“0 55.
34. ASTER, fi. H., AND JANDLE, J. H. (1964) J. Clin. Invest. 93, 843 35. PUSZICIN. E.. ALEDORT. L.. AND PUSZI~IN. S. (1970) J. Lab.
36.
37.
38.
39.
40.
41.
42. 43.
44. 45. 46.
47.
48.
49.
50.
51. 52.
YAMADA, K. M., SPOONER, B. S., AND WESSELLS, N. K. (1971) J. Cell Biol. 49, 614
YAM~DA, K. M., SPOONER, B. S., AND WESSELLS, N. K. (1970) Proc. Nat. Acad. Sci. U. S. A.66, 1206
WAGNI~R, R., ROSENBERG, M., AND ESTENSEN, R. (1971) J. Cell Biol. 60, 804
SCHBOEDER, T. 15. (1969) Biol. Bull. 137, 413 BLUJSMINIC. J. (19711 2. Zellforsch. MikroskoD. Anal. 121. 102 S.~NGER, J: W.,‘ AND'HOLTZE"R, H. (1972) Pro;. l\rat. Acad. Sci.
U. S. A. 69, 253 ZIGMOND, S. II., .\ND HIRSCH, J. G. (1972) Science 176, 1432 MIZEL, S. B., AND WII~SON, L. (1972) J. Biol. Chem. 247, 4102 HASLAM, IL. J. (1972) Proc. Biochem. Sot. 127, 434 SPUDICH, J. A., I\ND LIN, S. (1972) Proc. Nat. Acad. Sci.
U. S. A. 60, 442 FORER, A., EMMERSEN, J., AND BEnNIcE, 0. (1972) ScierLce 175,
774 TANENBAUM, S. W. (1971) Abstracts of the American Chemical
Society Meeting, Washington, D. C., September, Abstr. Biol. 128
HEMBREX, W. C., I<HRENI<BUFER, R. E., LIEBERMAN, S., .IXD WOLF, A. P. (1973) J. Biol. Chem., in press
Clin. iked‘76, 234 ‘ ' I .
BETTEX-GALL.~ND, M., AND L~~SCHER, R. E. (1965) AdvarL. Prot. Chem. 20, 1
PUSZKIN, 15., PUSZI~IN, S., AND ALEDORT, L. M. (1971) J. Biol. Chem. 246, 271
SZENT-GYORGYI, A. (1951) Chemistry of Muscle Contraction, p. 151, Academic Press, New York
RICHARDS, E. G., CHUNG, C. S., MENZEI., D. B., AND OLCOTT, H. S. (19671 Biochemislru 6. 528
CARSTEIN, M: Ii:., AND MO&M:~ERTS, W. F. H. M. (1963) Bio- chemistry 2, 28
LOWRY, 0. H., Itosm3nouon, N. J., F.~RR, A. L., AND RAND-ILL, R. J. (1951) J. Biol. Chem. 193, 265
MARSH, B. B. (1959) Biochim. Biophys. Acta 32, 357 PUSZICIN, S., BERL, S., PUSZI~IN, E., AND CL~RI~E, D. D.
(1968) Science 161, 170 SCATCHAXD, G. (1949) Ann. N. Y. Acad. Sci. 61, 660 SIPS, It. (1948) J. Chem. Phys. 16, 490 SHEPRO, D., BJ~LMARICH, F. A., ROBBLEE, L., AND CH.\O, F.
(1970) J. Cell Biol. 47, 544 WSSSSI,LS, N. K., SPOONER, B. S., As~I, J. F., BRADLV~Y, M. O.,
LUDUENA, M. A., TAYLOR, E. L., WRENN, J. T., AND YAM.IDI, K. M. (1971) Science 171, 135
WHITE, J. G. (1971) Platelet Aggregation (CAEN, J. ed) p. 15, Masson et Cie, Paris
SPUDICH, J. A. (1972) Cold Spring Harbor Symp. Quant. Biol. 37, 585
B.IR.~NY, M. (1959) Symposium on Sulfur and Proteins, p. 317, Academic Press, New York
YOUNG, M. (1967) J. Biol. Chem. 242, 2790 MCLAUGHLIN, G. A., SIM, G. A., RIECHI~L, J. R.., AND Tanrx,
CH. (1970) Chem. Commun. 1398
by guest on May 1, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Elena Puszkin, Saul Puszkin, Lan Wei Lo and S. W. TanenbaumBinding of Cytochalasin D to Platelet and Muscle Myosin
1973, 248:7754-7761.J. Biol. Chem.
http://www.jbc.org/content/248/22/7754Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/248/22/7754.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on May 1, 2018
http://ww
w.jbc.org/
Dow
nloaded from





























