Embed Size (px)
Citation preview

Biochemical Activities of Streptococcus pneumoniae Serotype 2Capsular Glycosyltransferases and Significance of SuppressorMutations Affecting the Initiating Glycosyltransferase Cps2E
David B. A. James,* Kanupriya Gupta, Jocelyn R. Hauser, Janet Yother
Department of Microbiology, University of Alabama at Birmingham, Birmingham, Alabama, USA
The capsular polysaccharide (CPS) is essential for Streptococcus pneumoniae virulence. Its synthesis requires multiple enzymes,and defects that block completion of the pathway can be lethal in the absence of secondary suppressor mutations. In this study,we examined the functions of three capsular glycosyltransferases (Cps2F, Cps2G, and Cps2I) involved in serotype 2 CPS synthe-sis, whose deletions select for secondary mutations. We demonstrate that Cps2F is a rhamnosyltransferase that catalyzes addi-tion of the third and fourth sugars in the capsule repeat unit, while Cps2G adds the fifth sugar (glucose). Addition of the terminalresidue (glucuronic acid) could not be detected; however, activities of the other glycosyltransferases together with bioinformaticanalyses suggest that this step is mediated by Cps2I. Most of the secondary suppressor mutations resulting from loss of theseenzymes occur in cps2E, the gene encoding the initiating glycosyltransferase. Examination of the 69 S. pneumoniae serotypescontaining Cps2E homologues yielded a consensus amino acid sequence for this protein and demonstrated that there is a highlysignificant association between the residues that are 100% conserved and those altered by suppressor mutations. Cps2E containsan extracytoplasmic loop whose function is unknown. Among our collection of mutants, six contained missense mutations af-fecting amino acids in the extracytoplasmic loop. These residues are highly conserved among S. pneumoniae Cps2E homologues,and mutations therein severely reduced CPS synthesis and Cps2E activity. The critical functions of these amino acids suggest arole for the Cps2E extracytoplasmic loop in initiation, and possibly regulation, of capsule synthesis.
Streptococcus pneumoniae (the pneumococcus) is an extracellu-lar bacterial pathogen that produces a protective surface struc-
ture known as the capsular polysaccharide (CPS). The CPS is anessential component for virulence, influencing both nasopharyn-geal colonization and systemic infection (1–8). CPS is also an im-munodominant surface antigen that serves as the basis for S. pneu-moniae serological typing and protective vaccines (3, 9–14). Over90 CPS serotypes have been identified; all but 2 utilize the Wzy-dependent pathway to synthesize CPS, while the remaining sero-types (serotypes 3 and 37) utilize the synthase-dependent pathway(15–18). This study focused on the Wzy-dependent pathway,which was first identified as a mechanism of synthesis for lipo-polysaccharide O antigen in Gram-negative bacteria (19–21) butis the principal mechanism for CPS synthesis in Gram-positivebacteria (18).
In the Wzy-dependent pathway, the capsule repeat unit is syn-thesized on the membrane acceptor undecaprenyl phosphate(Und-P) by multiple glycosyltransferases. Once completed, therepeat unit is transported across the cytoplasmic membrane by aWzx translocase and polymerized by a Wzy polymerase (Fig. 1).As a model for studying the Wzy-dependent pathway in S. pneu-moniae, our laboratory uses capsule serotype 2, which contains ahexasaccharide repeat unit comprised of glucose (Glc), rhamnose(Rha), and glucuronic acid (GlcUA) residues (22) that is poly-merized to at least 1,000 to 2,000 repeat units, based on the mo-lecular sizes of isolated polymers used in vaccine preparations (23)(Fig. 1). We previously identified and characterized the first twoglycosyltransferases involved in synthesizing the serotype 2 repeatunit: the glucose-1-phosphate (Glc-1-P) transferase Cps2E, whichinitiates synthesis (24), and the �1-4 rhamnosyltransferaseCps2T, which catalyzes the second and committed step (25).
There are a limited number of publications characterizing
pneumococcal glycosyltransferases involved in Wzy-dependentcapsule synthesis. In S. pneumoniae, homologues of the initiatingglycosyltransferase CpsE (also known as WchA) are present in atleast 69 serotypes (15). The glycosyltransferase activities of theseenzymes have been established, with enzyme functions demon-strated in serotypes 2, 8, 9v, and 14 (24, 26–28). CpsE is a memberof the polyprenyl-phosphate hexose-1-phosphate transferase(PHPT) family (20), in which the Salmonella enterica galactose-1-phosphate transferase WbaP serves as the prototypic enzyme (29–31). Three distinct regions are present in PHPT enzymes: the Nterminus, which in Cps2E contains 4 membrane-spanning do-mains; a central, extracytoplasmic loop; and a C-terminal cyto-plasmic tail following a membrane-spanning domain (31–34).Sugar transfer is catalyzed by residues in the cytoplasmic tail, asindependent studies have shown this region to be sufficient forglycosyltransferase activity (27, 31, 32, 35). The N-terminal regionis suggested to be required for protein insertion or stability in themembrane (31). The function of the extracytoplasmic loop is lessclear, but recent studies have suggested a role in modulation ofchain length distribution (31).
Three other genes in the cps2 locus—cps2F, cps2G, and cps2I—
Received 18 June 2013 Accepted 27 September 2013
Published ahead of print 4 October 2013
Address correspondence to Janet Yother, [email protected].
* Present address: David B. A. James, Department of Microbiology, New YorkUniversity School of Medicine, New York, New York, USA.
K.G. and J.R.H. contributed equally to this article.
Copyright © 2013, American Society for Microbiology. All Rights Reserved.
doi:10.1128/JB.00715-13
December 2013 Volume 195 Number 24 Journal of Bacteriology p. 5469 –5478 jb.asm.org 5469
on July 16, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

are predicted to encode the glycosyltransferases required for com-pletion of the repeat unit (Fig. 1) (25, 36, 37), but biochemicalevidence supporting their functions is lacking. In previous re-ports, we demonstrated that deletion of any one of these genes, orof the UDP-glucose dehydrogenase encoded by cps2K, the Wzxtranslocase (cps2J), or the Wzy polymerase (cps2H), was lethal inthe absence of secondary mutations, the majority of whichmapped to cps2E (25, 34). Each of the primary mutations affecteda point in the pathway past the committed step (Cps2T-catalyzedaddition of the first Rha). The lethality was attributed to eitherdestabilization of the membrane by the buildup of incompleterepeat units or sequestration of Und-P in the capsule pathway andaway from essential pathways such as peptidoglycan synthesis.Other reports have described similar suppressor mutations, someof which occurred in the initiating glycosyltransferase, when pri-mary mutations blocked the completion of polysaccharide path-ways in Pseudomonas aeruginosa lipopolysaccharide (LPS) Wzxflippase mutants (38), Xanthomonas campestris xanthan gum mu-tants (39), and Salmonella enterica serovar Typhimurium LPSmutants that failed to polymerize O-antigen subunits (30). Todate, we have obtained over 30 unique suppressor mutations incps2E that are distributed throughout the C-terminal cytoplasmictail and the central extracytoplasmic loop (25, 34). Suppressormutations mapping to the C-terminal region can be interpreted asa direct inhibition of Glc-1-P transferase activity. Suppressor mu-tations that map to the extracytoplasmic loop, however, are moredifficult to interpret but must have a function in limiting the ac-cumulation of incomplete repeat units.
In this report, we provide biochemical evidence for the previ-ously uncharacterized glycosyltransferases Cps2F and Cps2G that,together with bioinformatic analyses, completes the transferasespecificities for the type 2 CPS repeat unit of S. pneumoniae. Wealso further characterize our novel collection of secondary muta-tions in cps2E and demonstrate that residues in the extracytoplas-mic loop are critical for repeat unit initiation.
MATERIALS AND METHODSBacterial strains and growth conditions. S. pneumoniae strains (Table 1)were grown at 37°C in THY (Todd-Hewitt broth supplemented with 0.5%yeast extract) or on blood agar plates (BBL [Difco] or blood agar base[Becton and Dickinson] containing 3% defibrinated sheep blood [Colo-rado Serum Company]). Broth cultures were grown statically at 37°C.Plate cultures were incubated at 37°C in a candle jar. Escherichia colistrains (Table 1) were grown at 37°C in L broth (1% tryptone, 0.5% yeastextract, 0.5% NaCl, and 0.1% glucose) with shaking or on L agar plates (15g of agar/liter of L broth). Media were supplemented with the followingantibiotics as appropriate: ampicillin (Ap; 100 �g/ml), erythromycin(Em; 15 �g/ml for E. coli DB11 and 0.3 �g/ml for S. pneumoniae), andkanamycin (Km; 250 �g/ml).
Cloning of glycosyltransferase mutant enzymes. Sequences for cps2Ewith the mutations cps2E5718A¡G (H258R), cps2E5529G¡T (A162S),cps2E5544G¡T (D167Y), cps2E5539C¡T (L200F), and cps2E5632T¡G
(V196G) were PCR amplified from the chromosomal DNA of S. pneu-moniae strains DJ916, DJ921, BX547, BX552, and BX605, respectively.The primer pair used in PCRs was E8 (CATATGAATGGAAAAACAGTAAAGTC) and E11 (CTCGAGCTACTTCGCTCCATCTCTC), which am-plifies a 1.2-kb fragment containing the full-length cps2E (24). PCR prod-ucts were cloned into pET-20b and transformed into chemicallycompetent E. coli BL21-AI. The resulting strains DJ252 (Cps2EH258R),DJ253 (Cps2EA162S), DJ254 (Cps2ED167Y), DJ269 (Cps2EL200F), andDJ270 (Cps2EV196G) were used for expression of proteins for glycosyl-transferase assays, as described below.
Introduction of cps2E point mutations into S. pneumoniae D39. Tofacilitate identification of isolates transformed with cps2E point muta-tions, allelic exchange was used to insert a kanamycin resistance-encoding(Kmr) cassette (aphA-3) into the native cps2E. The aphA-3 gene was PCRamplified from the pneumococcal shuttle vector pSF151 (40) usingprimer pair DJ-03/DJ-04 and cloned into pGEM-T Easy cloning vector,resulting in pDJ271 (all plasmids were isolated and maintained in E. coliDH5� or DB11). Following digestion of pDJ271 with BstBI, the releasedaphA-3-containing fragment was ligated into BstBI-digested pBX146. Thelatter plasmid contains the full-length cps2E and partial upstream anddownstream sequences (cps2D and cps2T, respectively) cloned into thepneumococcal suicide vector pJY4164, which encodes Em resistance
FIG 1 S. pneumoniae type 2 capsular polysaccharide repeat unit structure and Wzy-dependent pathway. (Top) Repeat unit structure. Letters below eachglycosidic linkage represent the glycosyltransferase known or predicted to catalyze the linkage. (Bottom) Wzy-dependent model for synthesis of the type 2 repeatunit. The number of repeat units (n) is estimated to be 1,000 to 2,000, based on the molecular size of isolated polymer used in vaccine preparations (23). Figureadapted from references 25 and 55).
James et al.
5470 jb.asm.org Journal of Bacteriology
on July 16, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

(Emr) (41). BstBI cuts pBX146 at a single site within cps2E, allowing in-troduction of the aphA-3 fragment. The resulting construct, pJK101, wasused to transform competent D39 (42). Kmr isolates were demonstratedto be CPS negative and Em sensitive (Ems). The latter property indicatesthat Kmr was the result of allelic exchange and not single crossover result-ing in integration of the entire clone into the recipient chromosome. TheKmr isolate JK901 was used in subsequent studies.
Constructs containing the cps2E suppressor mutations encodingV196G and H258R were generated by PCR amplification of chromosomalDNA from strains DJ916 and BX605, respectively, using primer pair E8/E11 (primer sequences listed above) to amplify the full-length cps2E. Thefragments were cloned into the pGEM-T Easy cloning vector, and thesubsequent clone was then digested with MscI/BlpI. The released frag-ments were ligated to MscI/BlpI-digested pBX146. Digestion with MscI/BlpI releases a 751-bp internal fragment of cps2E that encompasses thesites of the mutations. The mutant plasmids resulting from these ligations(pDJ238, V196A, and pDJ241, H258R), along with the parent pBX146,were transformed into competent JK901 (cps2E::aph-3 Kmr), and reactionmixtures were plated on blood agar in the absence of antibiotic selection.Individual colonies were screened for Km sensitivity by patching on bloodagar with and without Km. Sensitive isolates were tested for Em sensitivityto confirm that the entire plasmid had not integrated into the chromo-some. The resulting strains were confirmed to have only the expectedmutations (JK906, V196, and JK907, H258) or the parental sequence(JK903) by DNA sequencing of cps2E. The frequencies of obtaining thesemutants were 0.3% (13/4,000) for JK906, 0.1% (6/6,000) for JK907, and0.7% (27/4,000) for JK903. Sequencing was performed by the UABGenomics Core Facility of the Heflin Center for Genomic Science.
Glycosyltransferase expression and membrane isolation. The previ-ously constructed E. coli strains expressing Cps2E (KJ4152), Cps2T(DJ009), Cps2F (DJ204), Cps2G (DJ011), and Cps2I (DJ005) were cul-tured and processed for membrane isolation as previously described (25).None of these constructs or those for other glycosyltransferases containedtags. For expression, 250-ml cultures were grown with the appropriateantibiotic selection to mid-exponential phase (cell density of approxi-mately 4.5 � 108 CFU/ml) and then induced with 0.2% arabinose for 30min (Cps2E and derivatives) or 4 h (all others). Cultures were centrifuged,and the cells were washed and lysed as previously described (25), followedby centrifugation at 100,000 � g to collect membranes. The final mem-brane pellet was suspended in 500 �l Tris-acetate (100 mM, pH 7.5)containing 10% glycerol and 1 mM dithiothreitol (DTT; buffer A) andstored at �80°C. Frozen membranes were thawed and vigorously vor-texed, and the homogenized solution was used to determine total mem-brane protein concentrations using the Bradford Bio-Rad protein dyeassay. Replicate samples (at least duplicate) were assayed. Membrane sam-ples from the parent and mutant strains demonstrated similar levels of anon-capsule-related protein, indicating homogeneity of the solutions (seeResults).
S. pneumoniae membranes were isolated based on previously de-scribed procedures (43, 44). Fifty-milliliter cultures were grown at 37°C inTHY to a density of 3 � 108 CFU/ml. Cultures were centrifuged at10,000 � g for 20 min. The cells were washed once in 50 ml of phosphate-buffered saline (137 mM NaCl, 2.7 mM KCl, 5.4 mM Na2HPO4·7H2O, 1.8mM KH2PO4 [pH 7.4]) and then suspended in 500 �l of protoplast buffer(20% sucrose, 50 mM Mg2SO4, 50 mM Tris-HCl [pH 7.4]). Samples wereincubated overnight at room temperature with gentle rotation (the S.pneumoniae autolysin LytA is active under these conditions [44]). Proto-plast formation was confirmed using phase-contrast light microscopy.The protoplasts were centrifuged at 16,000 � g for 10 min and washedonce in 500 �l of protoplast buffer. Protoplasts were osmotically lysed bysuspension in 1 ml of sterile water containing 10 mM EDTA, 1 mM DTT,and 1 mM phenylmethanesulfonyl fluoride, with incubation at room tem-perature for 30 min. Lysis was confirmed by light microscopy. Lysed cellswere centrifuged at 16,000 or 100,000 � g (with equivalent results) for 20min. The membrane-containing pellets were washed once in 1 ml of buf-
TABLE 1 Strains and plasmids used in this study
Strain orplasmid Propertiesa
Referenceor source
S. pneumoniaeBX547 D39 �cps2K Cpsr; cps2E5544 G¡T (D167Y) Kmr 34BX552 D39 �cps2H Cps� cps2E5539 C¡T (L200F); Kmr (in the
original publication, this mutant was incorrectlyidentified as L199F)
34
BX605 D39 �cps2K Cpsr cps2E5632T¡G (V196G) Kmr 34BX612 D39 �cps2K Cpsr cps2E5539T¡G (V165G) Kmr 34D39 Type 2 parent strain; Cps� 57DJ916 D39 �cps2F Cps� cps2E5718 A¡G (H258R) Kmr 25DJ918 D39 �cps2G Cpsr cps2E6059G¡C (M338I) Kmr 25DJ919 D39 �cps2G Cpsr cps2E6964C¡A (R307S) Kmr 25DJ921 D39 �cps2G Cpsr cps2E5529G¡T (A162S) Kmr 25JK901 pJK101 X D39; Cps�; aphA-3 inserted into cps2E; Kmr This studyJK903 pBX146 X JK901; cps2E repair; Cps� This studyJK906 pDJ238 X JK901; cps2E5632T¡G (V196G) Cpsr This studyJK907 pDJ241 X JK901; cps2E5718A¡G (H258R) Cpsr This studyKA1522 D39 �cps2E Cps� 24WU2 Type 3 parent strain; Cps� 58
E. coliBL21-AI F� ompT hsdSB(rB
� mB�) gal dcm araB::T7
RNAP-tetAInvitrogen
DB11 met thi gal hasdR nal rif; plasmid-cured derivative ofV854 from reference 59
59
DH5� F� 80lacZ�M15 �(lacZYA-argF) U169 recA1 endA1hsdR17 (rK
� mK�) phoA supE44 � thi-1 gyrA96
relA1
Invitrogen
DJ005 BL21-AI expressing Cps2I; Apr 25DJ009 BL21-AI expressing Cps2T; Apr 25DJ011 BL21-AI expressing Cps2G; Apr 25DJ204 BL21-AI expressing Cps2F, replaced GTG start with
ATG; Apr25
DJ238 DB11 (pDJ248); used to transform cps2E5718A¡G
(H258R)DJ241 DB11 (pDJ241); used to transform cps2E5632T¡G
(V196G)DJ252 BL21-AI (pDJ252) expressing Cps2EH258R; Apr This studyDJ253 BL21-AI (pDJ253) expressing Cps2EA162S; Apr This studyDJ254 BL21-AI (pDJ254) expressing Cps2ED167Y; Apr This studyDJ269 BL21-AI (pDJ269) expressing Cps2EL200F; Apr This studyDJ270 BL21-AI (pDJ269) expressing Cps2EV196G; Apr This studyDJ271 DH5� (pDJ271); aphA-containing clone; Kmr This studyJK101 DB11 (pJK101); contains aphA-3-disrupted cps2E;
KmrThis study
KJ4152 BL21-AI (pKJ4152) expressing full-length Cps2E; Apr 25RC124 BL21-AI (pET20b) vector control; Apr 24
PlasmidspBX146 pJY4164/cps2E; amplified from D39 using primer pair
cps2-D3/cps2-T1; contains full-length parentalcps2E. This plasmid is the same as the publishedpBX145.
34
pET20b Protein expression vector; Apr NovagenpDJ238 pBX146/cps2E5718A¡G (H258R) amplified from DJ916
using primers E8/E11bThis study
pDJ241 pBX146/cps2E5632T¡G (V196G) amplified fromBX605 using primers E8/E11b
This study
pDJ252 pET-20b/cps2E5718 A¡G (H258R) amplified fromDJ916 using primers E8/E11b
This study
pDJ253 pET-20b/cps2E5529G¡T (A162S) amplified from DJ921using primers E8/E11b
This study
pDJ254 pET-20b/cps2E5544 G¡T (D167Y) amplified fromBX547 using primers E8/E11b
This study
pDJ269 pET-20b/cps2E5539 C¡T (L200F) amplified fromBX552 using primers E8/E11b
This study
pDJ270 pET-20b/cps2E5632 T¡G (V196G) amplified fromBX605 using primers E8/E11b
This study
pDJ271 pGEM-T Easy/aphA-3-containing fragment amplifiedusing primer pair DJ-03/DJ-04
This study
pJK101 pBX146/aphA-3 from BstBI-digested product ofpDJ271
This study
pJY4164 S. pneumoniae suicide vector; Emr 41pKJ4152 pET-20b/cps2E amplified from S. pneumoniae D39
using primers E8/E11b25
a cps2E superscripts indicate locations of mutations based on GenBank accession no.AF026471. Amino acid changes are indicated in parentheses. Cpsr, reduced capsulelevels; ¡, nucleotide change; Apr, ampicillin resistant.b Primers are described in Materials and Methods.
Capsular Glycosyltransferases and Suppressor Mutations
December 2013 Volume 195 Number 24 jb.asm.org 5471
on July 16, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

fer A and suspended in 100 �l of buffer A. Total membrane protein con-centrations were quantified using the Bradford Bio-Rad protein dye assay,as described above.
Synthesis, purification, and quantification of dTDP-Rha. dTDP-Rha was synthesized in two steps essentially as previously described (25),with minor modifications to enzyme concentration and incubation time.Briefly, dTDP-Glc was synthesized in a 350-�l reaction mixture contain-ing 50 mM HEPES (pH 7.6), 30 mM MgCl2, 3.4 mM dTTP, 3.4 mMGlc-1-P, 2 U of inorganic pyrophosphatase, and 2 �M Cps2L. After incu-bation for 90 min at 37°C, the proteins were removed by filtration usingAmicon Ultra 10-kDa centrifugal filters (Millipore). The filtrate was sup-plemented with 0.1 mM NAD�, 6 mM NADH, 1 �M Cps2N, 1 �MCps2M, and 1 �M Cps2O in a final volume of 350 �l. The reaction mix-ture was incubated for 30 min at 37°C, the proteins were removed byfiltration, and dTDP-Rha was separated by high-performance liquidchromatography (HPLC) as previously described (25).
Glycosyltransferase assays. Glycosyltransferase assays to assess thefunctions of Cps2F, Cps2G, and Cps2I utilized recombinant enzymes ex-pressed separately in E. coli. For each enzyme, membranes containing 3 �gof total protein were incubated for 1 h at 10°C in reaction mixtures con-taining 10 mM MnCl2, 1 mM DTT, 0.008% NP-40 (in experiments uti-lizing multiple glycosyltransferases), 0.075 �Ci of UDP-[14C]Glc (300mCi/mmol), and, where indicated, 0.1 mM dTDP-Rha and/or 0.1 mMUDP-GlcUA. The final volume was brought to 75 �l with Tris-acetate(100 mM, pH 7.6). Reactions were stopped with 1 ml of 2:1 chloroform-methanol, and the organic phase was extracted three times with puresolvent upper phase (PSUP; 1.5 ml of chloroform, 25 ml of methanol, 23.5ml of H2O, and 0.183 g of KCl). Glycolipids were either preserved for lateruse as described for Und-P-P-14C-Glc-Rha-Rha-Rha (see below) or sub-jected to mild-acid hydrolysis by suspending the dried organic phase in 80�l of 20 mM HCl and incubating it at 70°C for 20 min. Hydrolyzedsamples were dried, suspended in 10 �l of chloroform-methanol (1:1),and applied as spots on silica-coated thin-layer chromatography (TLC;Whatman) plates. TLC plates were chromatographed for 8 h in butanol-ethanol-water (5:3:2). Dried plates were exposed to a phosphor screen(General Electric Healthcare) for 15 h, and bands were visualized in aStorm 820 phosphorimager (General Electric Healthcare).
Assays to quantify Cps2E activities of the parent and mutant proteinswere performed as previously described (24). Except as noted, S. pneu-moniae or E. coli membranes containing the indicated amounts of totalprotein (see figure legends) were incubated for 1 h at 10°C in 75-�l reac-tion volumes containing 10 mM MnCl2, 1 mM DTT, and 0.075 �Ci ofUDP-[3H]Glc (43.8 Ci/mmol) in Tris-acetate (100 mM, pH 7.6). Thereaction was stopped, and glycolipids were extracted and dried as de-scribed above for assays involving E. coli membranes. Incorporated[3H]Glc was analyzed by liquid scintillation counting.
Synthesis of Und-P-P-[14C]Glc-Rha-Rha-Rha (Cps2F product) andassay of Cps2G activity in the absence of other glycosyltransferases.Reaction mixtures of membranes containing Cps2E, Cps2T, and Cps2Fwere incubated as described above to synthesize the Cps2F product. Ex-tracted glycolipids were pooled, dried, and suspended in 100 �l of Tris-acetate (100 mM; pH 7.6) containing 0.1% NP-40. This solution was usedas the source of Und-P-P-[14C]Glc-Rha-Rha-Rha in subsequent glycosyl-transferase reactions. Membranes (3 �g of total protein) from DJ011 con-taining Cps2G or from RC124, a vector-only E. coli strain, were incubatedin reaction mixtures containing 0.008% NP-40, 10 mM MnCl2, 1 mMDTT, 25 �l of the Und-P-P-[14C]Glc-Rha-Rha-Rha solution, and 0.1 mMUDP-Glc. Reaction products were visualized as described above.
Bioinformatic analysis of the Cps2E homologues in S. pneumoniaeand other bacteria. Amino acid sequences from 94 serotypes (90 listed inreference 15 and the remaining 4 in references 45 and 46) were analyzed forCps2E homologues, which occur in 69 serotypes. These amino acid sequenceswere aligned by ClustalW (Geneious, version 5.6.4), and a consensus aminoacid sequence was generated. Using Fisher’s exact test, a significant correla-tion (P � 0.01) was observed between amino acids that are 100% conserved
among S. pneumoniae Cps2E homologues and those selected as suppressormutations. A protein basic local alignment search tool (BLAST [http://blast.ncbi.nlm.nih.gov/]) was used to identify Cps2E homologues in other bacte-ria. Homologous proteins from Clostridium phytofermentans ISDg (GenBankaccession number YP_001558318), Clostridium bolteae (WP_002576468),Streptococcus oralis (BAD22619), Streptococcus mitis (WP_001013652), andStreptococcus parasanguinis (WP_003009077) were among those identified.Amino acid sequences from these homologues, together with WbaP fromSalmonella enterica (YP_008266333) were aligned by ClustalW as describedabove.
RESULTS AND DISCUSSIONFunctional analyses of glycosyltransferases encoded in the cps2locus. We previously reported that Cps2E and Cps2T catalyze thefirst and second steps, respectively, in type 2 CPS synthesis (24,25). In addition to these enzymes, the cps2 locus encodes threeadditional predicted glycosyltransferases (Cps2F, Cps2G, andCps2I) that are expected to catalyze the remaining four glycosidiclinkages that complete the capsule repeat unit. The observationthat three enzymes might catalyze four linkages suggested that asingle glycosyltransferase encoded within the cps locus is capableof adding two sugar residues. We therefore sought to demonstratethe enzyme activities of Cps2F, Cps2G, and Cps2I and also clarifythe functions of the gene products whose deletions select for sec-ondary mutations in cps2E (25).
In in vitro glycosyltransferase assays, we utilized individualmembrane preparations obtained from E. coli expressing S. pneu-moniae glycosyltransferase Cps2F, Cps2G, or Cps2I. The absenceof functionally equivalent enzymes in E. coli was demonstrated bythe lack of activity when using E. coli membranes from a straincontaining only the plasmid vector, as described in the sectionsbelow. As previously reported (25), each of the S. pneumoniaeglycosyltransferases is membrane associated when expressed in E.coli despite lacking membrane-spanning domains. As for Cps2T(25), in vitro enzyme activity required the addition of nonionicdetergent (0.008% NP-40) at a concentration well below the crit-ical micelle concentration. This requirement may reflect a needfor temporal disassociation of enzymes from the membrane andaccess to membrane-bound glycolipid acceptors, possibly as partof the same micelle.
(i) Cps2F. To identify the enzyme that catalyzes the third stepin CPS synthesis, i.e., the second rhamnosyltransferase, we com-bined Cps2E-containing membranes in reactions with mem-branes containing Cps2T, nucleotide sugar substrates, and mem-branes containing the remaining glycosyltransferases. Glycolipidsformed in these reactions were extracted and the saccharide moi-ety was liberated by mild-acid hydrolysis and then separated byTLC. In the presence of UDP-[14C]Glc and dTDP-Rha, only thereaction mixture that contained Cps2F resulted in the formationof a new [14C]Glc-labeled product (Fig. 2), indicating that Cps2Fcatalyzes the third step in CPS synthesis. The Cps2F-dependentproduct was consistently generated in several independent reac-tions (data not shown).
The serotype 2 capsule has two identical inverting linkages(Rha-�-1,3-Rha-�-1,3) (22) (Fig. 1); however, Aanensen et al.identified only one gene (cps2F, also annotated as wchG) in the cpslocus that was predicted to encode an inverting transferase (36).This observation suggests that Cps2F may add both Rha residues.Using our standard TLC separation system, we were unable todistinguish two new products (Fig. 2). Alterations in the method,including adjustments to the polarity of the mobile phase, length
James et al.
5472 jb.asm.org Journal of Bacteriology
on July 16, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
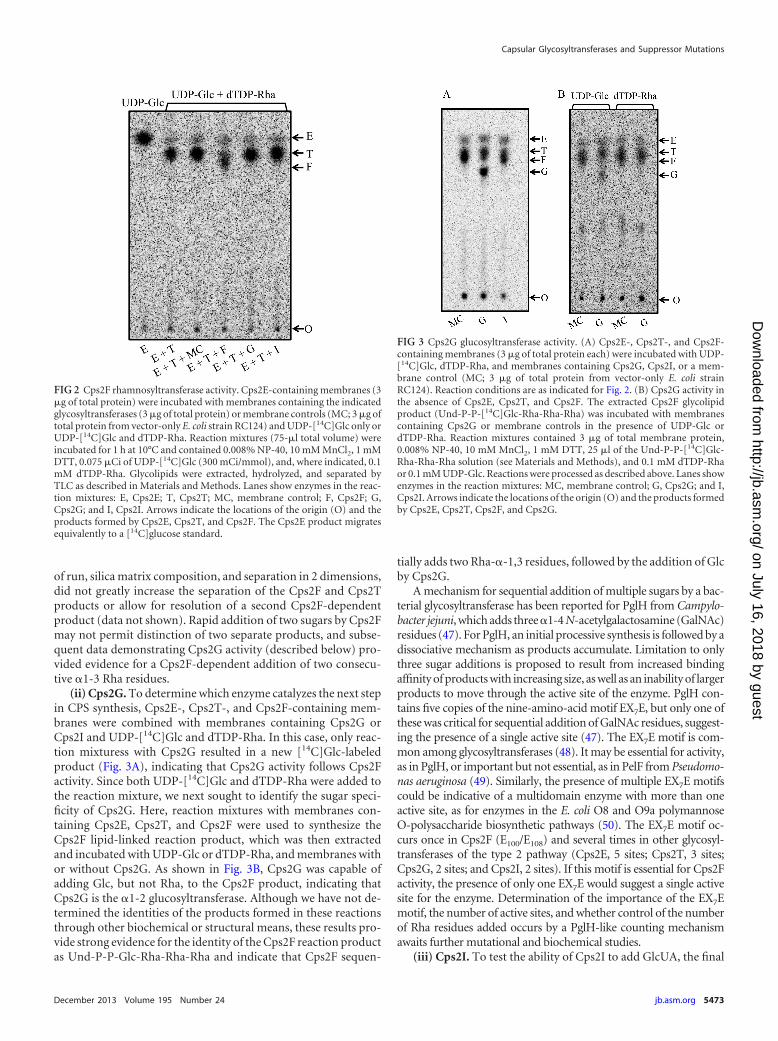
of run, silica matrix composition, and separation in 2 dimensions,did not greatly increase the separation of the Cps2F and Cps2Tproducts or allow for resolution of a second Cps2F-dependentproduct (data not shown). Rapid addition of two sugars by Cps2Fmay not permit distinction of two separate products, and subse-quent data demonstrating Cps2G activity (described below) pro-vided evidence for a Cps2F-dependent addition of two consecu-tive �1-3 Rha residues.
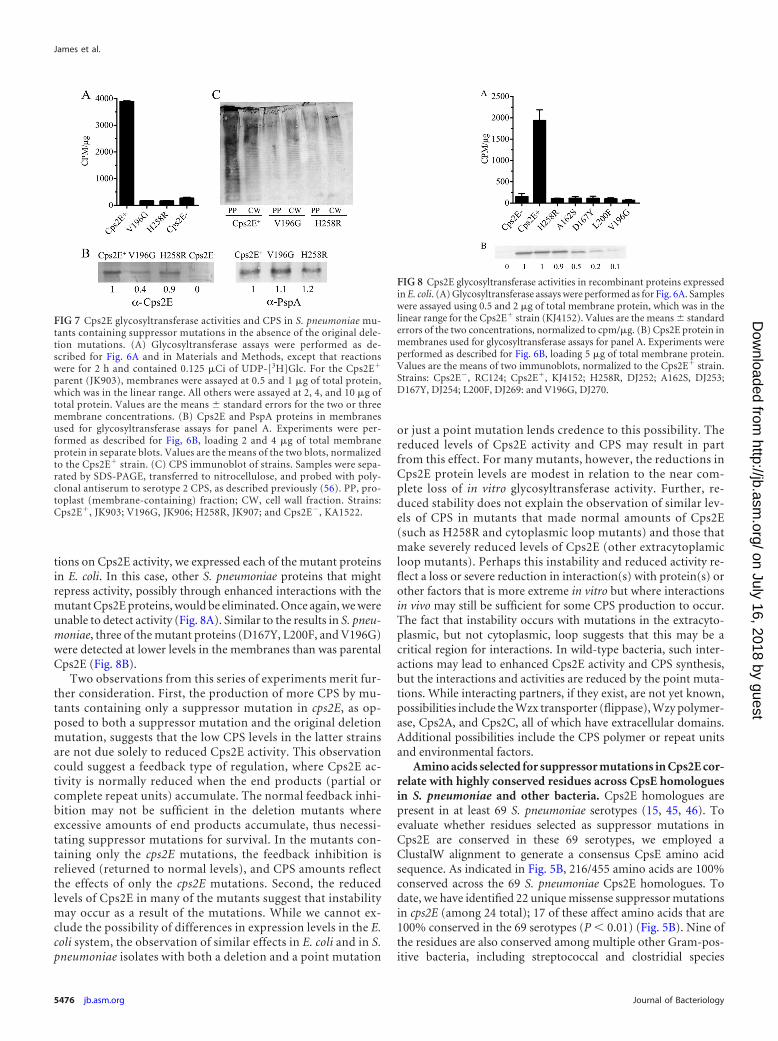
(ii) Cps2G. To determine which enzyme catalyzes the next stepin CPS synthesis, Cps2E-, Cps2T-, and Cps2F-containing mem-branes were combined with membranes containing Cps2G orCps2I and UDP-[14C]Glc and dTDP-Rha. In this case, only reac-tion mixturess with Cps2G resulted in a new [14C]Glc-labeledproduct (Fig. 3A), indicating that Cps2G activity follows Cps2Factivity. Since both UDP-[14C]Glc and dTDP-Rha were added tothe reaction mixture, we next sought to identify the sugar speci-ficity of Cps2G. Here, reaction mixtures with membranes con-taining Cps2E, Cps2T, and Cps2F were used to synthesize theCps2F lipid-linked reaction product, which was then extractedand incubated with UDP-Glc or dTDP-Rha, and membranes withor without Cps2G. As shown in Fig. 3B, Cps2G was capable ofadding Glc, but not Rha, to the Cps2F product, indicating thatCps2G is the �1-2 glucosyltransferase. Although we have not de-termined the identities of the products formed in these reactionsthrough other biochemical or structural means, these results pro-vide strong evidence for the identity of the Cps2F reaction productas Und-P-P-Glc-Rha-Rha-Rha and indicate that Cps2F sequen-
tially adds two Rha-�-1,3 residues, followed by the addition of Glcby Cps2G.
A mechanism for sequential addition of multiple sugars by a bac-terial glycosyltransferase has been reported for PglH from Campylo-bacter jejuni, which adds three �1-4 N-acetylgalactosamine (GalNAc)residues (47). For PglH, an initial processive synthesis is followed by adissociative mechanism as products accumulate. Limitation to onlythree sugar additions is proposed to result from increased bindingaffinity of products with increasing size, as well as an inability of largerproducts to move through the active site of the enzyme. PglH con-tains five copies of the nine-amino-acid motif EX7E, but only one ofthese was critical for sequential addition of GalNAc residues, suggest-ing the presence of a single active site (47). The EX7E motif is com-mon among glycosyltransferases (48). It may be essential for activity,as in PglH, or important but not essential, as in PelF from Pseudomo-nas aeruginosa (49). Similarly, the presence of multiple EX7E motifscould be indicative of a multidomain enzyme with more than oneactive site, as for enzymes in the E. coli O8 and O9a polymannoseO-polysaccharide biosynthetic pathways (50). The EX7E motif oc-curs once in Cps2F (E100/E108) and several times in other glycosyl-transferases of the type 2 pathway (Cps2E, 5 sites; Cps2T, 3 sites;Cps2G, 2 sites; and Cps2I, 2 sites). If this motif is essential for Cps2Factivity, the presence of only one EX7E would suggest a single activesite for the enzyme. Determination of the importance of the EX7Emotif, the number of active sites, and whether control of the numberof Rha residues added occurs by a PglH-like counting mechanismawaits further mutational and biochemical studies.
(iii) Cps2I. To test the ability of Cps2I to add GlcUA, the final
FIG 2 Cps2F rhamnosyltransferase activity. Cps2E-containing membranes (3�g of total protein) were incubated with membranes containing the indicatedglycosyltransferases (3 �g of total protein) or membrane controls (MC; 3 �g oftotal protein from vector-only E. coli strain RC124) and UDP-[14C]Glc only orUDP-[14C]Glc and dTDP-Rha. Reaction mixtures (75-�l total volume) wereincubated for 1 h at 10°C and contained 0.008% NP-40, 10 mM MnCl2, 1 mMDTT, 0.075 �Ci of UDP-[14C]Glc (300 mCi/mmol), and, where indicated, 0.1mM dTDP-Rha. Glycolipids were extracted, hydrolyzed, and separated byTLC as described in Materials and Methods. Lanes show enzymes in the reac-tion mixtures: E, Cps2E; T, Cps2T; MC, membrane control; F, Cps2F; G,Cps2G; and I, Cps2I. Arrows indicate the locations of the origin (O) and theproducts formed by Cps2E, Cps2T, and Cps2F. The Cps2E product migratesequivalently to a [14C]glucose standard.
FIG 3 Cps2G glucosyltransferase activity. (A) Cps2E-, Cps2T-, and Cps2F-containing membranes (3 �g of total protein each) were incubated with UDP-[14C]Glc, dTDP-Rha, and membranes containing Cps2G, Cps2I, or a mem-brane control (MC; 3 �g of total protein from vector-only E. coli strainRC124). Reaction conditions are as indicated for Fig. 2. (B) Cps2G activity inthe absence of Cps2E, Cps2T, and Cps2F. The extracted Cps2F glycolipidproduct (Und-P-P-[14C]Glc-Rha-Rha-Rha) was incubated with membranescontaining Cps2G or membrane controls in the presence of UDP-Glc ordTDP-Rha. Reaction mixtures contained 3 �g of total membrane protein,0.008% NP-40, 10 mM MnCl2, 1 mM DTT, 25 �l of the Und-P-P-[14C]Glc-Rha-Rha-Rha solution (see Materials and Methods), and 0.1 mM dTDP-Rhaor 0.1 mM UDP-Glc. Reactions were processed as described above. Lanes showenzymes in the reaction mixtures: MC, membrane control; G, Cps2G; and I,Cps2I. Arrows indicate the locations of the origin (O) and the products formedby Cps2E, Cps2T, Cps2F, and Cps2G.
Capsular Glycosyltransferases and Suppressor Mutations
December 2013 Volume 195 Number 24 jb.asm.org 5473
on July 16, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

sugar of the repeat unit, we used reactions with Cps2E-, Cps2T-,Cps2F-, Cps2G-containing membranes and the nucleotide sugarsUDP-[14C]Glc, dTDP-Rha, and UDP-GlcUA. Several attempts,with up to 20 �g of total membrane protein, resulted in no new[14C]Glc-labeled product (Fig. 4 and data not shown). We alsotested Cps2I activity directly by measuring the transfer of [14C]GlcUA from UDP-[14C]GlcUA to the Cps2G reaction product. Inthis case, we combined Cps2E-, Cps2T-, Cps2F-, Cps2G-, andCps2I-containing membranes with UDP-Glc, dTDP-Rha, andUDP-[14C]GlcUA. Glycolipids formed in these reactions were ex-tracted, dried, and analyzed for the incorporation of [14C]GlcUA.Membranes containing Cps2I did not incorporate [14C]GlcUAabove the negative control (identical reactions replacing Cps2Imembranes with control membranes). This result was in contrastto the positive control, which demonstrated [14C]GlcUA incorpo-ration by S. pneumoniae membranes containing the serotype 3synthase Cps3S (data not shown). This enzyme catalyzes the ad-dition of alternating Glc and GlcUA residues from the respectiveUDP sugars in the synthesis of the serotype 3 capsular polysaccha-ride (43, 51).
The inability to detect a Cps2I product was not due to reducedamounts of this protein in relation to the other glycosyltrans-ferases in the recombinant membranes, as we previously demon-strated that all of the glycosyltransferases are present at similarlevels (25). We also analyzed the effect of the addition of UDP-Glcand E. coli cell lysates containing functional Cps2K to the reactionsusing independent Cps2E-, Cps2T-, Cps2F-, Cps2G-, and Cps2I-containing membranes. Cps2K is the UDP-Glc dehydrogenasenecessary for synthesis of the Cps2I substrate UDP-GlcUA. Theseadditions also did not result in detection of a Cps2I-dependentproduct (data not shown).
Similar to our result, others have been unable to observe theaddition of terminal sugars on membrane-linked polysaccharidesfrom E. coli (GlcUA) and Klebsiella aerogenes (GlcUA) using invitro assays with membrane-associated glycosyltransferases (52,53). These results were attributed to limitations of the in vitrosystems. Factors that may be important in these systems includepossible protein modifications or the requirement for a chaper-one. An analysis of the S. pneumoniae phosphoproteome has iden-tified a serine-threonine phosphorylated peptide that correspondsto Cps2I (54), suggesting that modification may be important inCps2I function.
With only one enzyme and one linkage unaccounted for, weconclude that Cps2I adds the terminal �1-6 GlcUA sugar to thetype 2 repeat unit. Support for such a prediction comes frombioinformatic analyses, where cps2I is predicted to encode a re-taining glycosyltransferase (36), and �1-6 GlcUA is a retaininglinkage.
Suppressor mutations in the extracytoplasmic loop reduceCps2E activity and CPS levels. Deletions of the genes encodingglycosyltransferases (Cps2F, Cps2G, or Cps2I), the UDP-glucosedehydrogenase (Cps2K), the Wzx translocase (Cps2J), or the Wzypolymerase (Cps2H) select for isolates carrying secondary muta-tions, suggesting that the primary mutations are lethal in the ab-sence of suppression (25, 34). Most of these secondary mutations(32 of 35) map to cps2E and are missense mutations (24 of 32) thatoccur in both the extracytoplasmic and cytoplasmic regions (25,34) (Fig. 5A). Suppressor mutations in the cytoplasmic loop likelydirectly inhibit Cps2E Glc-1-P transferase activity, but the specificeffects of suppressor mutations in the extracytoplasmic loop, aswell as the function of this region, are less evident. Six strains fromour collection contain different missense mutations that map tothe Cps2E extracytoplasmic loop (Table 1 and Fig. 5A). Each ofthese mutations results in a nonconservative amino acid changeaffecting either residue polarity or the presence of an aromatic sidechain. No capsule is detectable with either of the S. pneumoniaestrains DJ916 (suppressor mutation H258R) and BX552 (L200F),whose primary mutations (�cps2F and �cps2H, respectively)eliminate the ability to synthesize the repeating capsule backboneand polymerize the repeat units, respectively (25, 34). However,the �cps2K mutants (BX547 [D167Y], BX605 [V196G], andBX612 [V165G]) and the �cps2G mutant (DJ921 [A162S]), eachof which lacks the resulting side chain in the polymerized polymer,can synthesize high-molecular-weight polymer, albeit at very lowlevels. This CPS is detectable only in the membrane, perhaps be-cause of the higher fraction of polysaccharide normally locatedthere (25, 34). Because high levels of membrane-associated poly-saccharide can accumulate in the parent without harm, we haveinterpreted this result to be indicative of the ability to transfer CPSfrom Und-P not only to peptidoglycan but also to another mem-brane acceptor in the parent, allowing the Und-P to be recycled(34). The low level of CPS produced by the �cps2K and �cps2Gmutants may reflect a reduced ability to make this transfer, withthe cps2E mutations resulting in less functional enzymes that slowsynthesis of repeat units, thereby allowing the transfer to keeppace. The reduced transfer from Und-P could occur because ofaltered recognition of the incomplete repeat unit by one or moreenzymes in the pathway (i.e., the Wzx transporter, the Wzy poly-merase, or the as-yet-unknown enzyme that moves the polymer toit final destination).
To test the effects of the extracytoplasmic loop mutations on
FIG 4 Assay for Cps2I glycosyltransferase activity. Cps2E-containing mem-branes (3 �g of total protein) were incubated with membranes containing theindicated glycosyltransferases (3 �g of total protein) and nucleotide sugars.Reactions were processed as described for Fig. 2 and contained 0.008% NP-40,10 mM MnCl2, 1 mM DTT, 0.075 �Ci of UDP-[14C]Glc (300 mCi/mmol),and, where indicated, 0.1 mM dTDP-Rha or 0.1 mM dTDP-Rha and 0.1 mMUDP-GlcUA. Lanes show enzymes in the reaction mixtures: E, Cps2E; T,Cps2T; F, Cps2F; G, Cps2G; and I, Cps2I. Arrows indicate the locations of theorigin (O) and the products formed by Cps2E, Cps2T, Cps2F, and Cps2G.
James et al.
5474 jb.asm.org Journal of Bacteriology
on July 16, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

Cps2E activity, we used the strains described above in in vitroglycosyltransferase assays. Each of the mutants exhibited activitybarely above that observed for the Cps2E-negative control strain(Fig. 6A, mutations H258R, A162S, D167Y, L200F, and V196G).This result was similar to what we observed previously for �cps2Kmutants that contained mutations in the Cps2E cytoplasmic loop(34). We again tested the latter effect using recently isolated�cps2G mutants containing mutations in the Cps2E cytoplasmicloop (25). Again, minimal activity was observed (M338I andR307S in Fig. 6A). In immunoblots of Cps2E, we noted reduced
levels of protein for most of the extracytoplasmic loop mutants,but no reduction was evident for the cytoplasmic mutants in ei-ther this or the previous study (Fig. 6B and reference 34). Thereduction in protein levels alone should not be sufficient to ac-count for the severe reductions in Cps2E activity, indicating thatother effects may be involved.
To address the possibility that Cps2E activities for the extracy-toplasmic loop mutants might be reduced as a result of the pri-mary deletion mutations, we introduced two of the mutations(H258R, derived from the �cps2F strain DJ916, and V196G, de-rived from the �cps2K strain BX605) into the parent, D39. Usingmembranes from these mutants, we were unable to detect Cps2Eactivity, even with increased amounts of membranes (Fig. 7A). Asin the original deletion strains, the H258R mutant exhibitedCps2E protein levels similar to those of the parent, whereas theV196G mutant was reduced (Fig. 7B). In contrast, the non-cap-sule-related protein PspA, which fractionates with the membranes(44), was present in similar amounts in the parent and mutantstrains (Fig. 7B). By the Quellung reaction, which uses reactivitywith antiserum for microscopic observation of CPS, the mutantsappeared to be CPS negative (data not shown). In the more sen-sitive immunoblot assay used to detect CPS in membrane and cellwall fractions, however, CPS was evident and displayed the fullrange of low- to high-molecular-weight polymers, albeit at re-duced amounts (Fig. 7C). The amount of CPS produced wasgreater than that seen for the original strains containing both acps2E mutation and a deletion (25), an effect we observed previ-ously with mutations in the cytoplasmic loop (34).
To further test the effect of the extracytoplasmic loop muta-
FIG 5 S. pneumoniae Cps2E topology and CpsE consensus sequence. (A) S.pneumoniae Cps2E topology and suppressor mutations (25, 34). The to-pology was determined using TMpred program from the ExPASy Proteom-ics website (http://www.expasy.org/tools), as previously reported (34).Only missense mutations are shown. The P363T and T378R mutationswere each identified in two independent mutants. Five nonsense muta-tions (E191, L244, L369, W407, and Q408) and three frameshift mutations(M59, E240, and G293) have also been identified. In the original publica-tion (34), L200F and K313N were incorrectly listed as L199F and K312N,respectively. (B) Conservation of Cps2E homologues in S. pneumoniae andother bacteria. Amino acid sequences from the 69 S. pneumoniae serotypescontaining Cps2E homologues were used in a ClustalW alignment to gen-erate a consensus sequence. Bars indicate the degree of conservation for therespective residues, where the height of the bar correlates to a greaterconservation (0 to 100%; dark gray bars indicate 100% conservation). Starsabove amino acids indicate relative positions of residues in the consensusCpsE where suppressor mutations were identified in the S. pneumoniaeCps2E. Triangles indicate residues conserved among Cps2E homologues inmultiple other bacteria, including Clostridium phytofermentans ISDg(GenBank accession number YP_001558318), Clostridium bolteae(WP_002576468), Streptococcus mitis (WP_001013652), Streptococcus ora-lis (BAD22619), and Streptococcus parasanguinis (WP_003009077). Twelveresidues (A177, D182, G310, G321, K331, F361, P385, G393, E407, G433,D446, and W467) are conserved in WbaP from Salmonella enterica subsp.enterica serovar 4 (YP_008266333). Note that residue numbers are shiftedby one from panel A due to insertion of a gap (denoted by a hyphen) foralignment of consensus sequence. X, no conservation of residue.
FIG 6 Cps2E glycosyltransferase activities in S. pneumoniae mutants contain-ing original deletion mutations and cps2E suppressor mutations. (A) Mem-branes were used to measure incorporation of [3H]Glc from UDP-[3H]Glc toan organically soluble product. Results are the means � standard errors fortwo independent membrane preparations assayed in separate experiments(with two replicates per sample). Samples were assayed using 0.2 and 0.5 �g oftotal membrane protein, which was in the linear range for the parent Cps2Eactivity. Activities are expressed as CPM/�g of total membrane protein. (B)Cps2E protein in membranes used for glycosyltransferase assays of panel A.Samples containing 5 �g of total membrane protein were separated by SDS-12% PAGE, transferred to a nitrocellulose membrane, and probed with poly-clonal antiserum to Cps2E (25). Relative protein levels were determined bydensitometry and normalized to that of the Cps2E� parent (D39). The valuesbelow the blot represent the means from three independent membrane prep-arations, two of which were used for the glycosyltransferase assays in panel A.A representative blot is shown. All samples were run on the same gel/blot; areplicate sample of M338I was spliced out of the image. Strains: Cps2E�,KA1522; Cps2E�, D39; H258R, DJ916; A162S, DJ921; D167Y, BX547; L200F,BX552; V196G, BX605; M338I, DJ918; and R307S, DJ919.
Capsular Glycosyltransferases and Suppressor Mutations
December 2013 Volume 195 Number 24 jb.asm.org 5475
on July 16, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
tions on Cps2E activity, we expressed each of the mutant proteinsin E. coli. In this case, other S. pneumoniae proteins that mightrepress activity, possibly through enhanced interactions with themutant Cps2E proteins, would be eliminated. Once again, we wereunable to detect activity (Fig. 8A). Similar to the results in S. pneu-moniae, three of the mutant proteins (D167Y, L200F, and V196G)were detected at lower levels in the membranes than was parentalCps2E (Fig. 8B).
Two observations from this series of experiments merit fur-ther consideration. First, the production of more CPS by mu-tants containing only a suppressor mutation in cps2E, as op-posed to both a suppressor mutation and the original deletionmutation, suggests that the low CPS levels in the latter strainsare not due solely to reduced Cps2E activity. This observationcould suggest a feedback type of regulation, where Cps2E ac-tivity is normally reduced when the end products (partial orcomplete repeat units) accumulate. The normal feedback inhi-bition may not be sufficient in the deletion mutants whereexcessive amounts of end products accumulate, thus necessi-tating suppressor mutations for survival. In the mutants con-taining only the cps2E mutations, the feedback inhibition isrelieved (returned to normal levels), and CPS amounts reflectthe effects of only the cps2E mutations. Second, the reducedlevels of Cps2E in many of the mutants suggest that instabilitymay occur as a result of the mutations. While we cannot ex-clude the possibility of differences in expression levels in the E.coli system, the observation of similar effects in E. coli and in S.pneumoniae isolates with both a deletion and a point mutation
or just a point mutation lends credence to this possibility. Thereduced levels of Cps2E activity and CPS may result in partfrom this effect. For many mutants, however, the reductions inCps2E protein levels are modest in relation to the near com-plete loss of in vitro glycosyltransferase activity. Further, re-duced stability does not explain the observation of similar lev-els of CPS in mutants that made normal amounts of Cps2E(such as H258R and cytoplasmic loop mutants) and those thatmake severely reduced levels of Cps2E (other extracytoplamicloop mutants). Perhaps this instability and reduced activity re-flect a loss or severe reduction in interaction(s) with protein(s) orother factors that is more extreme in vitro but where interactionsin vivo may still be sufficient for some CPS production to occur.The fact that instability occurs with mutations in the extracyto-plasmic, but not cytoplasmic, loop suggests that this may be acritical region for interactions. In wild-type bacteria, such inter-actions may lead to enhanced Cps2E activity and CPS synthesis,but the interactions and activities are reduced by the point muta-tions. While interacting partners, if they exist, are not yet known,possibilities include the Wzx transporter (flippase), Wzy polymer-ase, Cps2A, and Cps2C, all of which have extracellular domains.Additional possibilities include the CPS polymer or repeat unitsand environmental factors.
Amino acids selected for suppressor mutations in Cps2E cor-relate with highly conserved residues across CpsE homologuesin S. pneumoniae and other bacteria. Cps2E homologues arepresent in at least 69 S. pneumoniae serotypes (15, 45, 46). Toevaluate whether residues selected as suppressor mutations inCps2E are conserved in these 69 serotypes, we employed aClustalW alignment to generate a consensus CpsE amino acidsequence. As indicated in Fig. 5B, 216/455 amino acids are 100%conserved across the 69 S. pneumoniae Cps2E homologues. Todate, we have identified 22 unique missense suppressor mutationsin cps2E (among 24 total); 17 of these affect amino acids that are100% conserved in the 69 serotypes (P � 0.01) (Fig. 5B). Nine ofthe residues are also conserved among multiple other Gram-pos-itive bacteria, including streptococcal and clostridial species
FIG 8 Cps2E glycosyltransferase activities in recombinant proteins expressedin E. coli. (A) Glycosyltransferase assays were performed as for Fig. 6A. Sampleswere assayed using 0.5 and 2 �g of total membrane protein, which was in thelinear range for the Cps2E� strain (KJ4152). Values are the means � standarderrors of the two concentrations, normalized to cpm/�g. (B) Cps2E protein inmembranes used for glycosyltransferase assays for panel A. Experiments wereperformed as described for Fig. 6B, loading 5 �g of total membrane protein.Values are the means of two immunoblots, normalized to the Cps2E� strain.Strains: Cps2E�, RC124; Cps2E�, KJ4152; H258R, DJ252; A162S, DJ253;D167Y, DJ254; L200F, DJ269: and V196G, DJ270.
FIG 7 Cps2E glycosyltransferase activities and CPS in S. pneumoniae mu-tants containing suppressor mutations in the absence of the original dele-tion mutations. (A) Glycosyltransferase assays were performed as de-scribed for Fig. 6A and in Materials and Methods, except that reactionswere for 2 h and contained 0.125 �Ci of UDP-[3H]Glc. For the Cps2E�
parent (JK903), membranes were assayed at 0.5 and 1 �g of total protein,which was in the linear range. All others were assayed at 2, 4, and 10 �g oftotal protein. Values are the means � standard errors for the two or threemembrane concentrations. (B) Cps2E and PspA proteins in membranesused for glycosyltransferase assays for panel A. Experiments were per-formed as described for Fig, 6B, loading 2 and 4 �g of total membraneprotein in separate blots. Values are the means of the two blots, normalizedto the Cps2E� strain. (C) CPS immunoblot of strains. Samples were sepa-rated by SDS-PAGE, transferred to nitrocellulose, and probed with poly-clonal antiserum to serotype 2 CPS, as described previously (56). PP, pro-toplast (membrane-containing) fraction; CW, cell wall fraction. Strains:Cps2E�, JK903; V196G, JK906; H258R, JK907; and Cps2E�, KA1522.
James et al.
5476 jb.asm.org Journal of Bacteriology
on July 16, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

(Fig. 5B). Twelve of these residues (A162, D167, G292, G303,K313, F339, P363, G371, E385, G411, D425, and W435) are alsoconserved in their relative positions in WbaP (A177, D182, G310,G321, K331, F361, P385, G393, E407, G433, D446, and W467), theinitiating glycosyltransferase for the S. enterica O antigen. Theextracytoplasmic loops for both Cps2E and WbaP are 148 aminoacids in length but share limited sequence identity (23/152 resi-dues [15%]; positives 49/152 [32%]; gaps 8/152 [5%]). Theseresults indicate that the selection for suppressor mutations inCps2E identifies important and highly conserved functional resi-dues.
Conclusions. The assembly of surface polysaccharides re-quires multiple enzymes in a complex process that is undoubtedlyhighly coordinated. The results obtained in this and other studiesfrom our laboratory (24, 25, 34) and others (31) continue to sug-gest that Cps2E and related enzymes play central roles in the reg-ulation of this process. The existence of the extracytoplasmic loopand the effect of mutations therein suggest a function that is nec-essary for interaction with other surface enzymes, repeat units/polymer, or with an environmental factor.
In S. enterica WbaP mutants in which only the extracytoplas-mic loop is lacking, O antigen is produced at levels similar to thatof the parent but with an increased frequency of polymers in the10- to 20-repeat-unit range, and in vitro glycosyltransferase activ-ity is comparable to that of the parent (31). The present resultssuggest that in addition to any potential role in modulation ofchain length, the extracytoplasmic loop may have a role in regu-lating initiation of capsule synthesis. The cytoplasmic loop aloneappears capable of initiating synthesis, however, as one of the ex-tracytoplasmic loop mutations obtained in our original studycontained a nonsense mutation (E191*) just prior to a secondarytranslation start that effectively separated the N- and C-terminalregions. This mutant was able to produce CPS, albeit at low levels(reference 34 and unpublished data). This result is consistent withthe observation that the cytoplasmic region and the final trans-membrane domain are sufficient for activity in other glycosyl-transferases of the PHPT family (27, 31, 32, 35). In the native S.pneumoniae system, however, that activity may not be optimal inthe absence of a fully functional outer loop.
The requirement for suppressor mutations when capsule syn-thesis is blocked at a point past the committed step provides apowerful tool to identify critical residues of Cps2E. Despite thefact that mutations in many other genes (including cps2LMNO[dTDP-Rha synthesis], cps2C and cps2D [tyrosine phosphoregu-latory pathway], and cps2T [first rhamnosyltransferase]) couldreduce or eliminate CPS synthesis, 32 of 35 suppressor mutationshave thus far occurred in cp2E. These results suggest that there area greater number of effective target sites in cps2E than in all othersites combined, even though the collective target size for the genesabove is more than 4 times that of cps2E. We have not yet reachedsaturation mutagenesis; thus, many other residues likely remain tobe identified, providing the potential to better understand themultiple roles of Cps2E and related enzymes in capsule synthesis.
ACKNOWLEDGMENTS
This work was supported by Public Health Service grant AI28457 from theNational Institutes of Health and by the Mizutani Foundation for Glyco-science.
We thank Ella Robinson for graphics assistance.
REFERENCES1. Avery OT, Dubos R. 1931. The protective action of a specific enzyme
against type III pneumococcus infection in mice. J. Exp. Med. 54:73– 89.2. Brown EJ, Joiner KA, Cole RM, Berger M. 1983. Localization of com-
plement component 3 on Streptococcus pneumoniae: anti-capsular anti-body causes complement deposition on the pneumococcal capsule. Infect.Immun. 39:403– 409.
3. Griffith F. 1928. The significance of Pneumococcal types. J. Hyg. (Lond.)27:113–159.
4. Hardy GG, Magee AD, Ventura CL, Caimano MJ, Yother J. 2001.Essential role for cellular phosphoglucomutase in virulence of type 3Streptococcus pneumoniae. Infect. Immun. 69:2309 –2317.
5. Magee AD, Yother J. 2001. Requirement for capsule in colonization byStreptococcus pneumoniae. Infect. Immun. 69:3755–3761.
6. Nelson AL, Roche AM, Gould JM, Chim K, Ratner AJ, Weiser JN. 2007.Capsule enhances pneumococcal colonization by limiting mucus-mediated clearance. Infect. Immun. 75:83–90.
7. Weiser JN, Austrian R, Sreenivasan PK, Masure HR. 1994. Phase vari-ation in pneumococcal opacity: relationship between colonial morphol-ogy and nasopharyngeal colonization. Infect. Immun. 62:2582–2589.
8. Wood WB, Jr, Smith MR. 1949. The inhibition of surface phagocytosis bythe capsular slime layer of pneumococcus type III. J. Exp. Med. 90:85–96.
9. Francis T, Tillett WS. 1930. Cutaneous reactions in pneumonia. Thedevelopment of antibodies following the intradermal injection of type-specific polysaccharide. J. Exp. Med. 52:573–585.
10. Macleod CM, Hodges RG, Heidelberger M, Bernhard WG. 1945. Pre-vention of pneumococcal pneumonia by immunization with specific cap-sular polysaccharides. J. Exp. Med. 82:445– 465.
11. van Dam JE, Fleer A, Snippe H. 1990. Immunogenicity and immuno-chemistry of Streptococcus pneumoniae capsular polysaccharides. AntonieVan Leeuwenhoek 58:1– 47.
12. Centers for Disease Control and Prevention. 2011. Epidemiology andprevention of vaccine-preventable diseases, 12th ed, p 233–248. PublicHealth Foundation, Washington, DC.
13. Weiser J, Nahm MH. 2008. Immunity to extracellular bacteria, p 1182–1203. In Paul WE (ed), Fundamental immunology, 6th ed. Wolters Klu-wer Health/Lippincott Williams & Wilkins, Philadelphia, PA.
14. Abramson JS, Baker CJ, Fisher MC, Gerber MA, Meissner HC, MurrayDL, Overturf GD, Prober CG, Rennels MB, Saari TN, Weiner LB,Whitley RJ. 2000. American Academy of Pediatrics. Committee on Infec-tious Diseases. Policy statement: recommendations for the prevention ofpneumococcal infections, including the use of pneumococcal conjugatevaccine (Prevnar), pneumococcal polysaccharide vaccine, and antibioticprophylaxis. Pediatrics 106:362–366.
15. Bentley SD, Aanensen DM, Mavroidi A, Saunders D, Rabbinowitsch E,Collins M, Donohoe K, Harris D, Murphy L, Quail MA, Samuel G,Skovsted IC, Kaltoft MS, Barrell B, Reeves PR, Parkhill J, Spratt BG.2006. Genetic analysis of the capsular biosynthetic locus from all 90 pneu-mococcal serotypes. PLoS Genet. 2:e31.
16. Dillard JP, Vandersea MW, Yother J. 1995. Characterization of thecassette containing genes for type 3 capsular polysaccharide biosynthesisin Streptococcus pneumoniae. J. Exp. Med. 181:973–983.
17. Llull D, Garcia E, Lopez R. 2001. Tts, a processive beta-glucosyltransferase of Streptococcus pneumoniae, directs the synthesis ofthe branched type 37 capsular polysaccharide in pneumococcus and othergram-positive species. J. Biol. Chem. 276:21053–21061.
18. Yother J. 2011. Capsules of Streptococcus pneumoniae and other bacteria:paradigms for polysaccharide biosynthesis and regulation. Annu. Rev. Mi-crobiol. 65:563–581.
19. Raetz CR, Whitfield C. 2002. Lipopolysaccharide endotoxins. Annu. Rev.Biochem. 71:635–700.
20. Valvano MA. 2003. Export of O-specific lipopolysaccharide. Front. Bio-sci. 8:s452–s471.
21. Whitfield C. 2006. Biosynthesis and assembly of capsular polysaccharidesin Escherichia coli. Annu. Rev. Biochem. 75:39 – 68.
22. Jansson PE, Lindberg B, Anderson M, Lindquist U, Henrichsen J. 1988.Structural studies of the capsular polysaccharide from Streptococcus pneu-moniae type 2, a reinvestigation. Carbohydr. Res. 182:111–117.
23. Yother J, Bentley SD, Hennessey JP. 2008. Genetics, biosynthesis, andchemistry of pneumococcal capsular polysaccharides, p 33– 46. In SiberGR, Klugman KP, Makela PH (ed), Pneumococcal vaccines: the impact ofconjugate vaccine. ASM Press, Washington, DC.
Capsular Glycosyltransferases and Suppressor Mutations
December 2013 Volume 195 Number 24 jb.asm.org 5477
on July 16, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

24. Cartee RT, Forsee WT, Bender MH, Ambrose KD, Yother J. 2005. CpsEfrom type 2 Streptococcus pneumoniae catalyzes the reversible addition ofglucose-1-phosphate to a polyprenyl phosphate acceptor, initiating type 2capsule repeat unit formation. J. Bacteriol. 187:7425–7433.
25. James DBA, Yother J. 2012. Genetic and biochemical characterizations ofenzymes involved in Streptococcus pneumoniae serotype 2 capsule synthe-sis demonstrate that Cps2T (WchF) catalyzes the committed step by ad-dition of beta1-4 rhamnose, the second sugar residue in the repeat unit. J.Bacteriol. 194:6479 – 6489.
26. Kolkman MA, Morrison DA, Van Der Zeijst BA, Nuijten PJ. 1996. Thecapsule polysaccharide synthesis locus of Streptococcus pneumoniae sero-type 14: identification of the glycosyl transferase gene cps14E. J. Bacteriol.178:3736 –3741.
27. Pelosi L, Boumedienne M, Saksouk N, Geiselmann J, Geremia RA.2005. The glucosyl-1-phosphate transferase WchA (Cap8E) primes thecapsular polysaccharide repeat unit biosynthesis of Streptococcus pneu-moniae serotype 8. Biochem. Biophys. Res. Commun. 327:857– 865.
28. van Selm S, Kolkman MA, van der Zeijst BA, Zwaagstra KA, Gaastra W,van Putten JP. 2002. Organization and characterization of the capsulebiosynthesis locus of Streptococcus pneumoniae serotype 9V. Microbiology148:1747–1755.
29. Jiang XM, Neal B, Santiago F, Lee SJ, Romana LK, Reeves PR. 1991.Structure and sequence of the rfb (O antigen) gene cluster of Salmonellaserovar typhimurium (strain LT2). Mol. Microbiol. 5:695–713.
30. Yuasa R, Levinthal M, Nikaido H. 1969. Biosynthesis of cell wall lipo-polysaccharide in mutants of Salmonella. V. A mutant of Salmonella ty-phimurium defective in the synthesis of cytidine diphosphoabequose. J.Bacteriol. 100:433– 444.
31. Saldias MS, Patel K, Marolda CL, Bittner M, Contreras I, Valvano MA.2008. Distinct functional domains of the Salmonella enterica WbaP trans-ferase that is involved in the initiation reaction for synthesis of the Oantigen subunit. Microbiology 154:440 – 453.
32. Wang L, Liu D, Reeves PR. 1996. C-terminal half of Salmonella entericaWbaP (RfbP) is the galactosyl-1-phosphate transferase domain catalyzingthe first step of O-antigen synthesis. J. Bacteriol. 178:2598 –2604.
33. Wang L, Reeves PR. 1994. Involvement of the galactosyl-1-phosphatetransferase encoded by the Salmonella enterica rfbP gene in O-antigensubunit processing. J. Bacteriol. 176:4348 – 4356.
34. Xayarath B, Yother J. 2007. Mutations blocking side chain assembly,polymerization, or transport of a Wzy-dependent Streptococcus pneu-moniae capsule are lethal in the absence of suppressor mutations and canaffect polymer transfer to the cell wall. J. Bacteriol. 189:3369 –3381.
35. Patel KB, Ciepichal E, Swiezewska E, Valvano MA. 2011. The C-terminaldomain of the Salmonella enterica WbaP (UDP-galactose:Und-P galac-tose-1-phosphate transferase) is sufficient for catalytic activity and speci-ficity for undecaprenyl monophosphate. Glycobiology 22:116 –122.
36. Aanensen DM, Mavroidi A, Bentley SD, Reeves PR, Spratt BG. 2007.Predicted functions and linkage specificities of the products of the Strep-tococcus pneumoniae capsular biosynthetic loci. J. Bacteriol. 189:7856 –7876.
37. Iannelli F, Pearce BJ, Pozzi G. 1999. The type 2 capsule locus of Strepto-coccus pneumoniae. J. Bacteriol. 181:2652–2654.
38. Burrows LL, Lam JS. 1999. Effect of wzx (rfbX) mutations on A-band andB-band lipopolysaccharide biosynthesis in Pseudomonas aeruginosa O5. J.Bacteriol. 181:973–980.
39. Katzen F, Ferreiro DU, Oddo CG, Ielmini MV, Becker A, Puhler A,Ielpi L. 1998. Xanthomonas campestris pv. campestris gum mutants: ef-fects on xanthan biosynthesis and plant virulence. J. Bacteriol. 180:1607–1617.
40. Tao L, LeBlanc DJ, Ferretti JJ. 1992. Novel streptococcal-integrationshuttle vectors for gene cloning and inactivation. Gene 120:105–110.
41. Yother J, Handsome GL, Briles DE. 1992. Truncated forms of PspA that
are secreted from Streptococcus pneumoniae and their use in functionalstudies and cloning of the pspA gene. J. Bacteriol. 174:610 – 618.
42. Hardy GG, Caimano MJ, Yother J. 2000. Capsule biosynthesis and basicmetabolism in Streptococcus pneumoniae are linked through the cellularphosphoglucomutase. J. Bacteriol. 182:1854 –1863.
43. Cartee RT, Forsee WT, Schutzbach JS, Yother J. 2000. Mechanism oftype 3 capsular polysaccharide synthesis in Streptococcus pneumoniae. J.Biol. Chem. 275:3907–3914.
44. Yother J, White JM. 1994. Novel surface attachment mechanism of theStreptococcus pneumoniae protein PspA. J. Bacteriol. 176:2976 –2985.
45. Calix JJ, Nahm MH. 2010. A new pneumococcal serotype, 11E, has avariably inactivated wcjE gene. J. Infect. Dis. 202:29 –38.
46. Calix JJ, Porambo RJ, Brady AM, Larson TR, Yother J, Abeygunward-ana C, Nahm MH. 2012. Biochemical, genetic, and serological character-ization of two capsule subtypes among Streptococcus pneumoniae serotype20 strains: discovery of a new pneumococcal serotype. J. Biol. Chem. 287:27885–27894.
47. Troutman JM, Imperiali B. 2009. Campylobacter jejuni PglH is a singleactive site processive polymerase that utilizes product inhibition to limitsequential glycosyl transfer reactions. Biochemistry 48:2807–2816.
48. Coutinho PM, Deleury E, Davies GJ, Henrissat B. 2003. An evolvinghierarchical family classification for glycosyltransferases. J. Mol. Biol. 328:307–317.
49. Ghafoor A, Jordens Z, Rehm BH. 2013. Role of PelF in pel polysaccharidebiosynthesis in Pseudomonas aeruginosa. Appl. Environ. Microbiol. 79:2968 –2978.
50. Greenfield LK, Richards MR, Li J, Wakarchuk WW, Lowary TL, Whit-field C. 2012. Biosynthesis of the polymannose lipopolysaccharide O-an-tigens from Escherichia coli serotypes O8 and O9a requires a unique com-bination of single- and multiple-active site mannosyltransferases. J. Biol.Chem. 287:35078 –35091.
51. Dillard JP, Yother J. 1994. Genetic and molecular characterization ofcapsular polysaccharide biosynthesis in Streptococcus pneumoniae type 3.Mol. Microbiol. 12:959 –972.
52. Drummelsmith J, Whitfield C. 1999. Gene products required for surfaceexpression of the capsular form of the group 1 K antigen in Escherichia coli(O9a:K30). Mol. Microbiol. 31:1321–1332.
53. Sutherland IW, Norval M. 1970. The synthesis of exopolysaccharide byKlebsiella aerogenes membrane preparations and the involvement of lipidintermediates. Biochem. J. 120:567–576.
54. Sun X, Ge F, Xiao CL, Yin XF, Ge R, Zhang LH, He QY. 2010.Phosphoproteomic analysis reveals the multiple roles of phosphorylationin pathogenic bacterium Streptococcus pneumoniae. J. Proteome Res.9:275–282.
55. Yother J. 2006. Integration of capsular polysaccharide biosynthesis withmetabolic and virulence pathways in Streptococcus pneumoniae, p 51– 65.In Brogden KA, Minion FC, Cornick N, Stanton TB, Zhang Q, Nolan LK,Wannemuehler MJ (ed), Virulence mechanisms of bacterial pathogens,4th ed. ASM Press, Washington, DC.
56. Bender MH, Cartee RT, Yother J. 2003. Positive correlation betweentyrosine phosphorylation of CpsD and capsular polysaccharide produc-tion in Streptococcus pneumoniae. J. Bacteriol. 185:6057– 6066.
57. Avery OT, MacLeod CM, McCarty M. 1944. Studies on the chemicalnature of the substance inducing transformation of pneumococcal types:induction of transformation by a deoxyribonucleic acid fraction isolatedfrom pneumococcus type III. J. Exp. Med. 79:137–158.
58. Briles DE, Nahm M, Schroer K, Davie J, Baker P, Kearney J, Barletta R.1981. Antiphosphocholine antibodies found in normal mouse serum areprotective against intravenous infection with type 3 Streptococcus pneu-moniae. J. Exp. Med. 153:694 –705.
59. Macrina FL, Tobian JA, Jones KR, Evans RP, Clewell DB. 1982. Acloning vector able to replicate in Escherichia coli and Streptococcus san-guis. Gene 19:345–353.
James et al.
5478 jb.asm.org Journal of Bacteriology
on July 16, 2018 by guesthttp://jb.asm
.org/D
ownloaded from





























