Embed Size (px)
Citation preview

BIOSYSTEMATICS OF THE GENUS IPS (COLEOPTERA: SCOLYTIDAE) IN NORTH AMERICA. HOPPING'S GROUPS IV AND X
GERALD N. LANIER' Forest Research Laboratory, Canadian Forestry Service, Edmonton, Alberta
Abstract Can. Ent. 104: 361-388 (1972) The taxonomic concepts of the Ips species of groups IV and X were tested
by controlled breeding, bioassay of sex pheromone specificity, and examination of morphological and karyological details. No changes were made in the status of Ips pini, I. bonanseai, and I . avulsus of group IV. I. interstitialis of group X is removed from synonymy with I. calligraphus, and I. ponderosae, also pre- viously considered a synonym of I. calligraphus, is assigned subspecific rank. Assertions of close phylogenetic relationship of the Eurasian I. duplicatus and I. sexdentatus with groups IV and X, respectively, are rejected.
I . p i t~ i from widely separated localities were interfertile although egg hatchability was subnormal in some crosses involving F, from matings of beetles from western and eastern North America. I. pini and I. bonanseai readily mated but less than 3% of the eggs hatched and the larvae died. There was no evidence of breeding incompatibility in crosses of I . c. calligraphus from Florida and California or I. c. ponderosae from New Mexico and South Dakota. How- ever, hatchability differed drastically between reciprocal pairings of the I. calli- graphus subspecies, possibly as a result of cytoplasmic incompatibility. All eggs produced in pairings of I. interstitialis with either of the I. calligraphus subspecies failed to hatch.
First meiotic metaphase of I. pini, I. bonanseai, and both I. calligraphus subspecies invariably showed the formula 15AA + Xy,. This karyotypic formula also applied to I. avulsus except for a few individuals that had 2 small supernumerary chromosomes. The karyotype of I. interstitialis was not deter- mined. Heteromorphic bivalents and univalents were detected in hybrids of the I. calligraphus subspecies.
Cross attractiveness to sex pheromones was demonstrated for I. pirti from different regions, for I . pini and I. bonanseai, and for the I. calligraphus sub- species. When samples of attractive frass from eastern and western I. pini males were presented simultaneously, eastern females responded equally to both samples but western females favored western frass by a margin of 2 to 1. Similarly, I . c. calligraphus clearly discriminated in favor of consubspecific pheromone while I . c. ponderosae did not.
The locality of several of Eichhoff's holotypes is noted.
INTRODUCTION This paper is the third of a series on North American Ips reporting the results
of intensive controlled breeding experiments, study of karyological and morpho- logical detail, and tests of sex pheromone specificity. Previous reports dealt with groups IX (Lanier 1970b) and I11 (Lanier 1 9 7 0 ~ ) . Groups 111, IV, IX and X constitute a discrete subdivision of the genus which includes most of its best known and economically important members.
Review of Group ZV In his revision of the genus Ips, Hopping (1963, 1964) placed the North
American species Ips pini (Say), I. oregonis (Eichhoff) , I. bonanseai (Hopkins) , I. avulsus (Eichhoff) and, tentatively, I. duplicatus Sahlberg of Eurasia in his group IV. Characteristics used to define this group were four spines on each lateral margin of the elytral declivity, bisinuate antenna1 club sutures, and male genitalia narrow and elongate with median struts more than twice the length of the median lobe. =Present address: Department of Forest Entomology, State University College of Forestry at Syracuse University, Syracuse, N.Y. 13210.
361
Can
. Ent
omol
. Dow
nloa
ded
from
pub
s.es
c-se
c.ca
by
Nat
ural
Res
ourc
es C
anad
a on
07/
11/1
1Fo
r pe
rson
al u
se o
nly.

3 62 THE CANADIAN ENTOMOLOGIST March 1972
I . oregonis and pini, the most similar of the group IV species, were dis- tinguished by the former's western distribution, slightly larger size, and coarser sculpturing (Swaine 19 18; Chamberlin 1939). Hopping ( 1964) recognized these differences but synonymized the two species under pini because they intergraded in western Canada.
Similarity in parasites supports this synonymy. Eastern and western forms of a mite (Iponemus truncatus truncatus Lindquist) parasitic on the eggs of pini and oregonis, respectively, also intergrade in the arid regions of southern British Columbia and eastern Washington (Lindquist 1 9 6 9 ~ ) . In addition, certain internally parasitic nematodes in beetles from eastern and western regions are indistinguishable (Nickle, pers. comm.). Nevertheless, Smith (1952) reported pini from eastern Canada to have 15 pairs of chromosomes (meiotic formula = 14AA + Xy,) while Lanier (1966) observed 16 pairs (15AA + Xy,) in pini (oregonis) from California.
Lindquist ( 1 9 6 9 ~ ) agreed that pini, bonanseai, and avulsus are of a common phyletic line because their parasitic Iponemus mites, all subspecies of I. truncatus (Ewing) , are very similar. However, the only Iponemus recorded on duplicatus was a species not clearly related to truncatus.
Total inviability of eggs resulting from matings between pini (oregonis) and I . plastographus (Leconte) of group I11 has been demonstrated (Lanier 1966), but the interfertility between the various group IV species and between disjunct populations of pini, sensu lato, has not been tested prior to this investigation.
Review of Group X Hopping's (1963, 19653) group X was composed of Ips species with 6 spines
on each side of the elytral declivity, including I. calligraphus (Germar) of the eastern United States and southeastern Canada, I. ponderosae Swaine of the interior western United States, I. interstitialis (Eichhoff) of Mexico, Central America and the Caribbean Islands, and I. sexdentatus (Boerner) of Eurasia.
Leconte (1868, 1876) recognized a large form of calligraphus from New Mexico. Swaine (1925) described this entity as I. ponderosae, citing, in addition to size, its coarse sculpturing and deeply impressed punctation. I. interstitialis from Jamaica (Eichhoff 1869) has been differentiated from calligraphus by the former's smaller size and its propensity to attack small, rather than large, diameter host material (Schwerdtfeger 1956). Schedl (1955) doubted that these three species were distinct and Hopping (1965b) synonymized them under calligraphus.
One of the recently discussed problems in the concept of calligraphus (Hopping 1965b; Wood and Stark 1968a; Lindquist 1969a) is that specimens from California are similar to calligraphus from the southeastern states rather than to the geo- graphically nearer ponderoaae, as had been assumed by Chamberlin (1939) and Keen ( 1953). Hopping (pers. comm.) and Lindquist ( 1 9 6 9 ~ ) independently suggested that the former was introduced in California, possibly with pines taken to the forest nursery in Placerville-the site of the first recorded collection of a 6-spined Ips in California (Keen 1929). This opinion is supported by the very limited distribution of calligraphus in California (Wood and Stark 1948a, b) and Lindquist's ( 1 9 6 9 ~ ) discovery that one subspecies of mite, Iponemus calligraphi calligraphi Lindquist, parasitizes calligraphus s. str. in California and the south- eastern states while another, I. c. cordillerae Lindquist, is associated with ponderosae and interstitialis.
Can
. Ent
omol
. Dow
nloa
ded
from
pub
s.es
c-se
c.ca
by
Nat
ural
Res
ourc
es C
anad
a on
07/
11/1
1Fo
r pe
rson
al u
se o
nly.

Volume 104 THE CANADIAN ENTOMOLOGIST 363
In grouping sexdentatus with North American 6-spined Ips, Schedl (1955) and Hopping ( 1963, 1965 b) apparently overlooked Leconte's ( 1 876) important observation that the 6 spines of the Old and New World Ips are not homologous; the former has 2 spines below the most prominent spine which arises from the fifth stria1 interspace while the latter have 3 spines below the principal spine. Moreover, Lindquist (1969a) considered the Iponemus on these taxa to be remotely related while mites on certain more clearly related Ips showed Holarctic afiities.
This paper further demonstrates the remoteness of the relationship of Old and New World 6-spined Ips, raises interstitialis from synonymy, and designates ponderosae a subspecies of calligraphus.
METHODS Controlled breeding experiments, study of morphological and cytological
details, and tests of sex pheromone specificity were employed in this investigation. Insects used for breeding and cytological study were taken as callow adults
from laboratory colonies established from field populations (Table I, Figs. 1, 2). Procedures for breeding experiments and for preparation and examination of slides showing chromosomes or morphological features were described previously (Lanier 1966,1970b).
Newly-emerged females were used in tests of pheromone specificity. Pheromone-laden frass for bioassay was collected in gelatin capsules A x e d over the entrance holes of males individually introduced in Jeffrey or ponderosa pine. A test saniple weighed about .1 g and was taken from the aggregate collected from
Table I. Host and collection locality of Ips species of groups IV and X used in mating and cytological studies
No. O 9 paired examined
Species Collection locality Host Pinus with d 8 of: cytologically
1. pini
"oregonis"
2. bonanseai
3. avulsus
1. calligraphus calligraphus
2. calligraphus ponderosae
Group IV
a. Ont., Sault Ste. Marie b. Ont., Angus c. B.C., Armstrong d. Alta., Banff e. Calif., Hat Creek P.O. f . Calif., Cottage Springs g. Calif., Big Bear P.O. h. Nev., Genoa i. Ariz., Flagstaff
res{nosa res%nosa ponderosa
jeffreyi jefreyi jeffreyi jefreyi ponderosa
a. Ariz., Santa Catalina Mts. strobiformis
a. Va., Keysville virginiana
Group X a. Calif., Avery ponderosa b. Calif., Murphies ponderosa c. Fla., Gainesville elliottii
a. N.M., Ruidoso ponderosa b. S.D., Sheridan Lake ponderosa
l a lh, i; 2a lg, i; 2a If, g, h
- lc; 2a; 3a 1 b; 2a; 3a
lb, c; 2b; 3a l b ; 2a
3, interslitialis a. Ariz., Chiricahua Mts. arizonica 2a -
Can
. Ent
omol
. Dow
nloa
ded
from
pub
s.es
c-se
c.ca
by
Nat
ural
Res
ourc
es C
anad
a on
07/
11/1
1Fo
r pe
rson
al u
se o
nly.
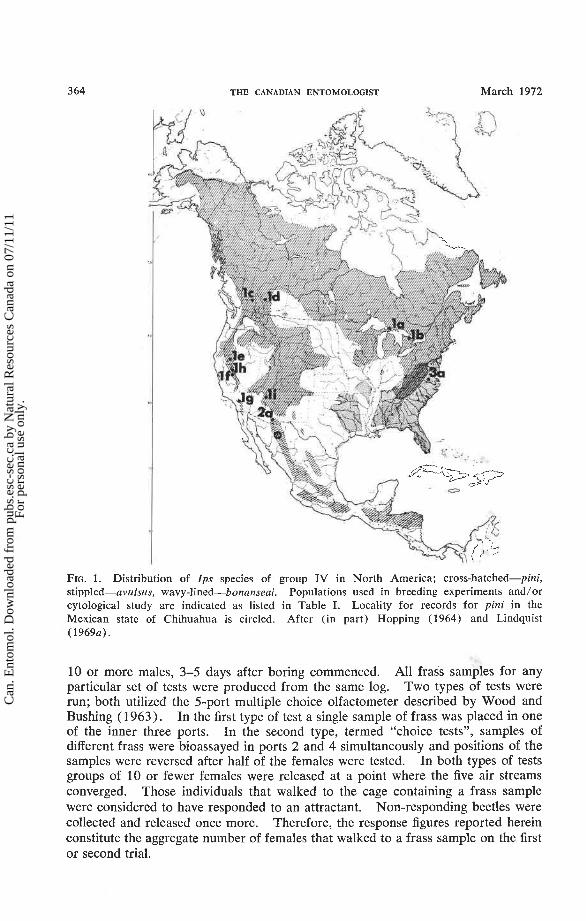
3 64 THE CANADIAN ENTOMOLOGIST March 1972
FIG. 1. Distribution of Ips species of group N in North America; cross-hatched-pini, stippled-avulsus, wavy-lined-bonanseai. Populations used in breeding experiments and/or cytological study are indicated as listed in Table I. Locality for records for pini in the Mexican state of Chihuahua is circled. After (in part) Hopping (1964) and Lindquist (1969~).
10 or more males, 3-5 days after boring commenced. All frass samples for any particular set of tests were produced from the same log. Two types of tests were run; both utilized the 5-port multiple choice olfactometer described by Wood and Bushing (1963). In the first type of test a single sample of frass was placed in one of the inner three ports. In the second type, termed "choice tests", samples of different frass were bioassayed in ports 2 and 4 simultaneously and positions of the samples were reversed after half of the females were tested. In both types of tests groups of 10 or fewer females were released at a point where the five air streams converged. Those individuals that walked to the cage containing a frass sample were considered to have responded to an attractant. Non-responding beetles were collected and released once more. Therefore, the response figures reported herein constitute the aggregate number of females that walked to a frass sample on the first or second trial.
Can
. Ent
omol
. Dow
nloa
ded
from
pub
s.es
c-se
c.ca
by
Nat
ural
Res
ourc
es C
anad
a on
07/
11/1
1Fo
r pe
rson
al u
se o
nly.

Volume 104 THE CANADIAN ENTOMOLOGIST 365
FIG. 2. Distribution of Ips species and subspecies of group X in North America; stippled- calligraphus calligraphus, cross-hatched-c. ponderosae, wavy-lined-interstitialis. Populations used in breeding experiments and/or cytological study are indicated as listed in Table I. After (in part) Hopping (1965) and Lindquist ( 1 9 6 9 ~ ) .
TAXONOMY Group IV
The synonymy, description, key, and illustration of group IV species by Hopping ( 1964) need not be repeated here. Supplemental information is provided below.
Ips pini (Say) TAXONOMIC LITERATURE
Bostrichus pini Say 1826, p. 257. Tomicus praefictus Eichhoff 1867, p. 401, syn. by Leconte 1876, p. 365 was over-
looked by Hopping 1964, p. 974. Ips interpunctus, (nec? Tomicus interpunctus Eichhoff 1878, p. 241) Swaine 191 8,
pp, 110, 11 6-1 17; Chamberlin 1939, pp. 429-430; 1958, pp. 164, 172-173; Hopping 1965a, p. 533. See Remarks.
Ips pini Hopping 1964; Lanier 1966. BIOLOGICAL LITERATURE (Selected)
Tomicus pini, Fitch 1858; Felt 1906. Ips pini, Clemens 19 16; Craighead and Middleton 1930, p. 6; Doane et al. 1936,
pp. 92-93; Anderson 1948; Craighead 1950, p. 335; Reid 1955, 1957; Thomas 1961, 1967; Johnson 1967; Wygant and Lara 1967; Barr 1969; Schenk and Benjamin 1969; Lanier and Cameron 1969.
Ips oregonis, Keen 1929; Doane et al. 1936, pp. 93-94; Keen 1953, pp. 150-151; Miller and Keen 1960, pp. 104-105; Wood 1961.
Can
. Ent
omol
. Dow
nloa
ded
from
pub
s.es
c-se
c.ca
by
Nat
ural
Res
ourc
es C
anad
a on
07/
11/1
1Fo
r pe
rson
al u
se o
nly.
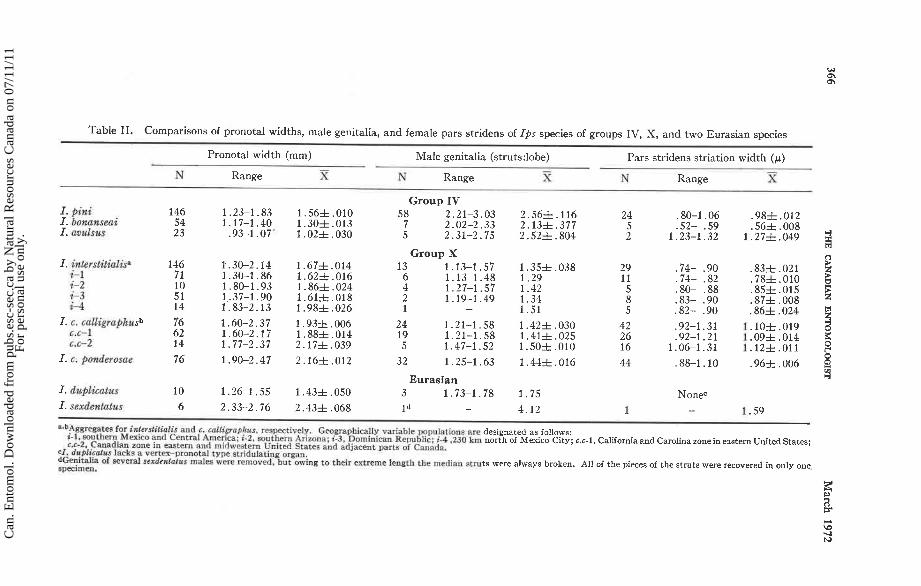
Table 11. Comparisons of pronotal widths, male genitalia, and female pars stridens of Ips species of groups IV, X, and two Eurasian species
Pronotal width (mm) Male genitalia (struts:lobe) Pars stridens striation width (p) - - -
N Range X N Range X N Range S
Group IV
Group X 13 1.13-1.57
Eurasian 3 1.73-1.78 1 * -
a"Aggrpgatm for iniersfiiialis and c. ca l l i~opkus . respectively. Geographically variable populations urn designated as follows: 1-1, southern Mexico and Centtal America; i - 2 . southern Arizona; i-3. Dominican Ker\ublic; t-4.230 km north of Mexico City; 6.6-1, California and Carolina zone in eastern United States; C.C-2, Cznadiiln zone in eastern and midwwtern United Statra and adjacent parts ni Canada.
cf. da8licafars lacks a vertex-pronot11 type striduhting orpan. aGe+talia of ~ v e r a l ~ e x d ~ a l o i t a s males were removed. bu t owing to their extreme length the median struts were always broken. All of the pieces of the struts were recovered in only one, Bpeclnlen.
z e-. w 4 N C
an. E
ntom
ol. D
ownl
oade
d fr
om p
ubs.
esc-
sec.
ca b
y N
atur
al R
esou
rces
Can
ada
on 0
7/11
/11
For
pers
onal
use
onl
y.
Volume 104 THE CANADIAN ENTOMOLOGIST 3 67
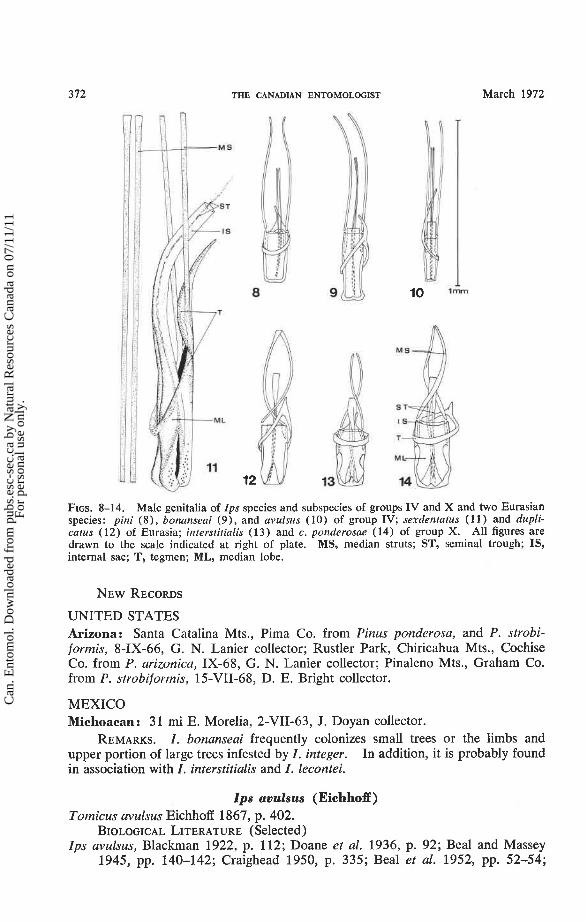
DIAGNOSIS. The larger size of pini easily differentiates it from avulsus and bonanseai (Table 11). The largest chromosomal bivalent of pini usually forms a ring at PMI while those of avulsus and bonanseai are frequently cross-shaped. In addition, pini differs from avulsus in having a strongly sexually dimorphic declivital armature, narrower striations on the pars stridens, and a median lobe of the male genitalia which is no more than 3 times longer than wide (Fig. 8) . The punctures on the pronotum of I . pini are more than a puncture's diameter apart, the median lobe of the male genitalia is rounded caudally, and the antennae are amber-colored. The pronotal punctures on bonanseai are less than their own diameter apart, the median lobe is flared caudally, and the antennae have a greenish cast.
KARYOLOGY. The mitotic number is 32 and the male meiotic formula is 15AA + Xy, (this designates 15 pairs of autosomes and a parachute-shaped sex bivalent).
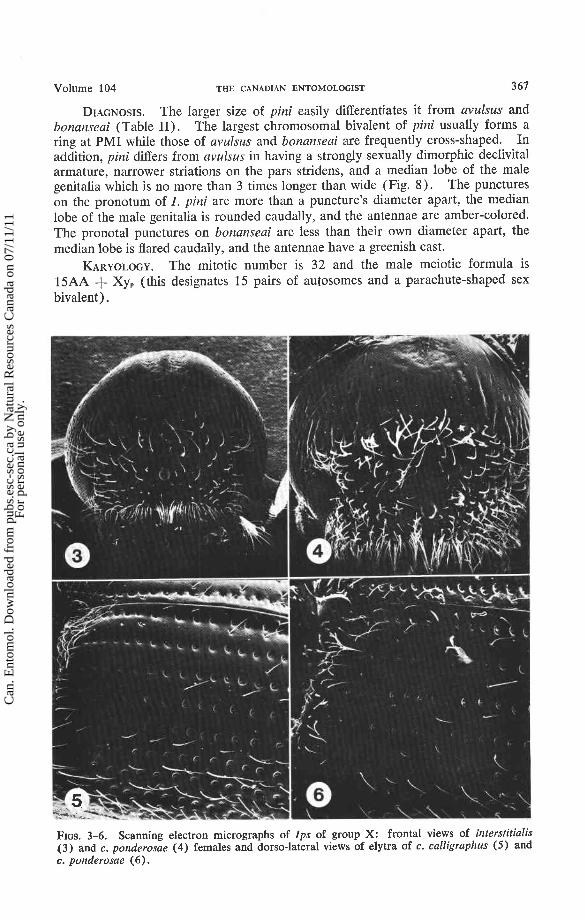
FIGS. 3-6. Scanning electron micrographs of Ips of group X: frontal views of interstitialis (3) and c. ponderosae (4) females and dorso-lateral views of elytra of c. calligraphus (5) and c. ponderosae (6 ) .
Can
. Ent
omol
. Dow
nloa
ded
from
pub
s.es
c-se
c.ca
by
Nat
ural
Res
ourc
es C
anad
a on
07/
11/1
1Fo
r pe
rson
al u
se o
nly.

368 THE CANADIAN ENTOMOLOGIST March 1972
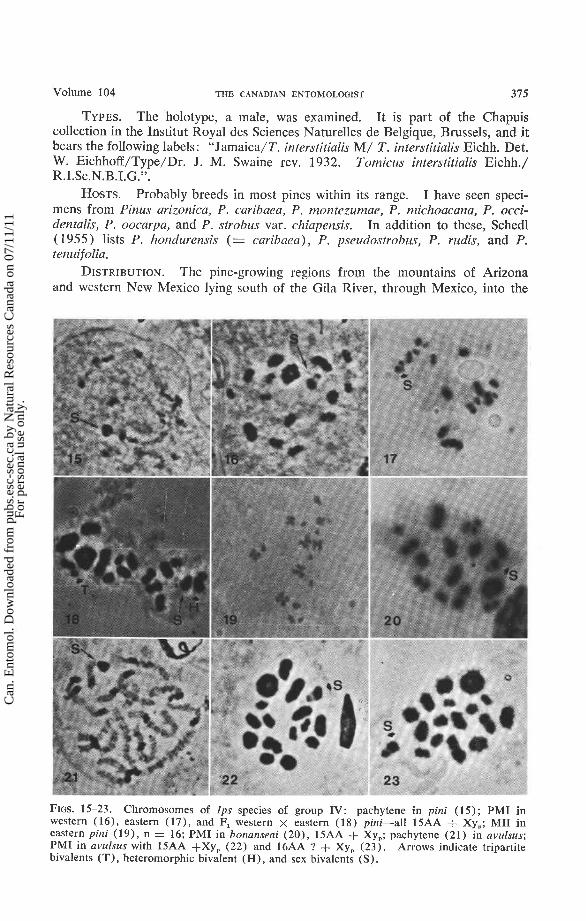
Pachytene bivalents are marked by heteropycnotic segments of varying sizes (Fig. 15). On the larger chromosomes these segments appear to be procentric; in other chromosomes they seem to compose an entire arm. The X and y coalesce around the nucleolus to form a moderately staining sphere.
By first prometaphase (PMI) heteropycnosis is no longer apparent and the Xy, is clearly defined (Figs. 16-18). Bivalents Nos. 1 and 2 (in order of de- creasing size) are distinctly larger than the remaining autosomes. Other bivalents are not individually recognizable by size although slight breaks occur between bivalents 4-5, 5-6, and 11-12. Bivalent No. 1 forms a ring or, rarely a thick rod (as in Fig. 17) or a cross. No. 2 is ring-shaped in about 60% of the meiocites and cross- or rod-shaped in the remainder. Bivalent No. 5 forms a tripartite configura- tion (Lanier 1966) in individuals from California or Nevada, but this formation was infrequent in specimens from other localities. The remaining bivalents are shaped like crosses, rods, or tiny dumbbells.
At MI1 the chromatid forms are free; their confluence marks the position of the centromeres (Fig. 19) . No. 1 is a heterobrachial cross with an arm ratio (AR) of about 4:s . Nos. 2-6 are isobrachial crosses while the succeeding autosomes are heterobrachial or cephalobrachial. The X and y chromosomes were not in- dividually identified, but by comparison of X- and y-bearing MIIs, it was ascertained that the X was an isobrachial in the size range of autosomes 3-5 while the y was similar in size and appearance to Nos. 12-1 5.
GEOGRAPHIC VARIATION. Hopping (1964) wrote "there is considerable variation between series of I. pini from different localities and different hosts but the distribution is continuous from east to west and the variations intergrade one into the other. Some eastern seres, especially from Pinus resinosa, are smoother, more shining and have flatter elytral interspaces than most from the west. The punc- tation of the pronotum also varies from fine to rather coarse. The third declivital spine in the male varies from strongly capitate and straight to sub-capitate and bent ventrad". I agree with this description except that I noted no variation between series from different hosts at the same locality. In addition, I found specimens from Mexico and the southern Rocky Mountains to be significantly larger (P < .01) than those from eastern North America, British Columbia, and California. HOW- ever, there were no significant differences between regions for width of striations on the pars stridens or struts-lobe ratio (S/L) of the male genitalia (Table 11).
TYPES. The location of the pini holotype and the type locality are unknown to me. I have seen the holotypes of the following species and I agree that all are synonyms of pini: Ips laticollis Swaine (1918) "Ottawa Ont., 1912" is in the Canadian National Collection, Ottawa; Tomicus praefrictus Eichhoff bearing the labels "Eichh. Am. Bor./Type/det Tomicus praefrictus Eich./R.I.Sc.N.B.I.G." is a male, apparently of the eastern variety, in the Chapuis collection held at the Institut Royal des Sciences Naturelles de Belgique : Tomicus oregonis Eichhoff, bearing the labels "Oregon Leconte/T. oregonis M./Type/det Tomicus oregonis Eich./ R.1.Sc.N.B.I.G." is a male, larger than most western pini and also in the Chapuis collection.
HOSTS. Chamberlin (1939, 1958) and Hopping (1964) believed that pini utilized all species of pines within this range. In addition to the hosts recorded by these authors, I have seen specimens from Pinus edulis, Pinus washoensis, Pinus engelmannii, Picea pungens, Picea mariana, and Picea chihuahuaensis.
Can
. Ent
omol
. Dow
nloa
ded
from
pub
s.es
c-se
c.ca
by
Nat
ural
Res
ourc
es C
anad
a on
07/
11/1
1Fo
r pe
rson
al u
se o
nly.

Volume 104 THE CANADIAN ENTOMOLOGIST 369
DISTRIBUTION. I. pini occurs throughout the pine and spruce forests of North America except: 1-on the Pacific Coast west of the Sierra Nevada, Cascade, and Canadian Coast Range mountains; 2-the southern pine zone (which is occupied by avulsus); 3-most of southern Arizona, Mexico, and Central America (which is occupied by bonanseai) (Fig. 1 ) . The geographic limit of pini in the southern Appalachians has recently been extended well beyond that recorded by Hopping (1964). Berisford et al. (1970) collected pini in Virginia, and Ciesla and Bell (1968) recorded this species in North and South Carolina. However, Ciesla (pers. comm.) notes that pini and avulsus are separated in the southern Appalachians by elevation; the former occurs at high elevations where it infests principally white pine (Pinus strobus) while the latter occurs at lower elevations where it infests southern "hard" pines. Figure 1 includes surprising new records for pini in the Sierra Madre Occidental of northern Mexico. Previous references to pini in northern Mexico (Hopping 1964; Wygant and Lara 1967) are apparently based entirely upon specimens taken from the San Pedro Martir Range of Baja California. These Mexican localities are not related floristically; pines of Baja California are species that extend from California while the species composition of Chihuahua is distinctly Mexican (Mirov 1967).
NEW RECORDS. Over 2000 specimens from more than 200 localities were seen. The only records listed here are those that add significantly to Hopping's (1965) distribution map for pini.
CANADA Northwest Territories: Ft. Liard, Ft. Providence, 15 mi W. James River, Pine Point, and Yellowknife. Nova Scotia: Mersey River in Queens CO. Ontario: Gogama, Iroquois Falls, Kashabowie, Lockalsh, McIntosh, Nakina, Ryland, Sioux Falls, Upsala and White River. Saskatchewan : Nipawin Peak.
MEXICO Chihuahua: Mesa del Huracan, 108" 15': 30°4', Pinus engelmannii, 21-VII-64, E. E. Lindquist collector (50 spm.). Same data, J. B. Thomas collector (26 sprn.). La Laja, Pice0 chihuahuaensis, 16-VII-60, S. L. Wood collector (2 sprn.). See Remarks.
UNITED STATES Alaska : Juneau in Douglas Co. Arizona : Sitgreaves Natl. For. Colorado : Antonito in Conejos Co.; Castle Rock in Douglas Co. (this series was erroneously reported as Ips plastographus by Lindquist (1969a), p. 61, and Boss and Thatcher (1970), pp. 2, 4 ) ; Granby in Grand Co.; 12 mi N. Gardner in Huerfano CO.; Buckhorn Canyon and Ft. Collins in Larimer Co. Nebraska: Halsey in Thomas CO. Nevada: Mt. Rose in Washo Co., Pinus washoensis. New Mexico: Santa Fe Ski Area in Santa Fe Co., Picea pungens; Capitan (2 series, 1 from Pinus edulis) and Ruidoso in Lincoln Co.; Cloudcroft ( 3 series) in Otero Co. Utah: Logan Cyn. in Cash Co.; Boneta in Duchesne Co.; Panguitch in Garfield Co.; La Sal Mts. in San Juan Co., Pinus edulis; Kamas and 4 mi S. Mirror Lake in Summit Co.; Tabbie Mtn. in Uinta Nat. For. Wyoming: Upton in Weston CO.; Buffalo District in Bighorn Nat. For.; Frontier Cr. and Wiggins Fork in Shoshone Nat. For.
REMARKS. The true identity of I. interpunctus (Eichhoff) remains an enigma. Most authors have followed Swaine (1918) in referring to a northern form of pini
Can
. Ent
omol
. Dow
nloa
ded
from
pub
s.es
c-se
c.ca
by
Nat
ural
Res
ourc
es C
anad
a on
07/
11/1
1Fo
r pe
rson
al u
se o
nly.

370 THE CANADIAN ENTOMOLOGIST March 1972
under this name. Hopping ( 1 9 6 5 ~ ) believed that the original description best fit a group VI species.
The Mexican records for pini listed here were completely unexpected in view of the previously published records for this species and the general rule that northern Ips and their hosts do not penetrate into interior Mexico. Moreover, pini appears to be invariably replaced by bonanseai south of the Gila River in Arizona. Never- theless, two specimens of pini from Picea chihuahuaensis were typical in all aspects of external morphology. Seventy-six specimens from Pinus engelmannii, also in the Mexican state of Chihuahua, had been identified by other workers as bonanseai, but in their size (mean pronotal width = 1.66 mm) pars stridens, male genitalia, and other morphological characteristics, they were identical to pini from the southern Rocky Mountains. Fifty of these specimens in the Canadian National Collection, Ottawa, were collected by E. E. Lindquist; 26 in the Forestry Labora- tory collection, Edmonton, bear the alcohol vial label of J. B. Thomas. These two men often collected from the same material during an expedition of Canadian entomologists to Mexico in 1964. The dual collection, together with the fact that the expedition took no specimens north of Mexico (Lindquist and Thomas, pers. comm.), confirms the authenticity of this record. Nevertheless, other series-1 of 14 specimens taken by J. B. Thomas bearing the same collection data as the above except for lot number and date, and another series of 31 specimens from Pinus arizonica-were all clearly bonanseai. These collections were made over a period of several days and at different elevations (Lindquist, pers. comm.). Perhaps pini replaces bonanseai at high elevations in the same way it replaces avulsus in the southern Appalachian region.
The difference in chromosome number reported for eastern and western pini by Smith (1952) and Lanier (1966), respectively, was not apparent in this investigation. Small differences in relative size of homologous chromosomes were revealed by the appearance of heteromorphic bivalents in inter-regional hybrids (see Results, karyology of hybrids), but spermatogenesis was normal and the hybrids were fertile. I. pini should therefore be considered to be a generally chromosomally stable species which may have one of the small chromosomes (bivalents) missing in certain individuals. An intrapopulational difference in chromosome number is reported here for avulsus.
Ips bonanseai (Hopkins) Tomicus bonanseai Hopkins 1905, p. 76.
BIOLOGICAL LITERATURE Ips bonanseai, Schwerdtfeger 1956; Thomas 1966, 1967; Barr 1969, pp. 641, 651;
Lanier and Cameron 1969; Lindquist 1969a. DIAGNOSIS. Characters differentiating bonanseai and pini are summarized
under the latter species. I. bonanseai differs from avulsus in having denser pro- notal punctation, greenish brown rather than amber antennae, narrower striations on the pars stridens, and male genitalia with a flared rather than straight median lobe (Figs. 9, 10).
KARYOLOGY. Mitotic (spermatagonial) divisions were not observed. The male meiotic formula is 15AA + Xy, and MI1 cells contained 16 chromosomes.
Pachytene cells appeared similar to those of pini. At PMI (Fig. 20) only the largest bivalent (No. 1) was identifiable by its size.
Minor breaks in size sequence occurred between autosomes 6-7 and 11-12. No. 1
Can
. Ent
omol
. Dow
nloa
ded
from
pub
s.es
c-se
c.ca
by
Nat
ural
Res
ourc
es C
anad
a on
07/
11/1
1Fo
r pe
rson
al u
se o
nly.
Volume 104 THE CANADIAN ENTOMOLOGIST
- I. calligraphus calligraphus 1 I. calligraphus calligraphus 2
0 I, calligraphus ponderosae '0 I. calligraphus ponderosae (coincident polnts) V I. interstitialis 1
I. interstitialis 2 • ' A I, interstitialis 3 ¤ D 0 I, interstitialis 4 • • . . . 0 . m
Pronotal Width (mm)
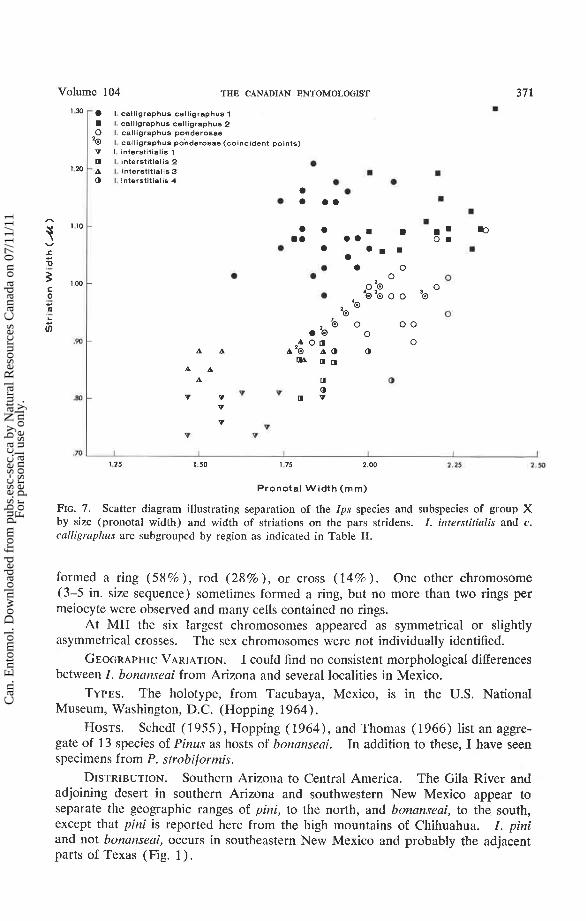
FIG. 7. Scatter diagram illustrating separation of the Ips species and subspecies of group X by size (pronotal width) and width of striations on the pars stridens. I . interstitialis and c. calligraphus are subgrouped by region as indicated in Table 11.
formed a ring (58% ), rod (28%) , or cross (14%). One other chromosome (3-5 in. size sequence) sometimes formed a ring, but no more than two rings per meiocyte were observed and many cells contained no rings.
At MI1 the six largest chromosomes appeared as symmetrical or slightly asymmetrical crosses. The sex chromosomes were not individually identified.
GEOGRAPHIC VARIATION. I could find no consistent morphological differences between I. bonanseai from Arizona and several localities in Mexico.
TYPES. The holotype, from Tacubaya, Mexico, is in the U.S. National Museum, Washington, D.C. (Hopping 1964).
HOSTS. Schedl (1955), Hopping (1964), and Thomas (1966) list an aggre- gate of 13 species of Pinus as hosts of bonanseai. In addition to these, I have seen specimens from P. strobiformis.
DISTRIBUTION. Southern Arizona to Central America. The Gila River and adjoining desert in southern Arizona and southwestern New Mexico appear to separate the geographic ranges of pini, to the north, and bonanseai, to the south, except that pini is reported here from the high mountains of Chihuahua. I. pini and not bonanseai, occurs in southeastern New Mexico and probably the adjacent parts of Texas (Fig. 1 ) .
Can
. Ent
omol
. Dow
nloa
ded
from
pub
s.es
c-se
c.ca
by
Nat
ural
Res
ourc
es C
anad
a on
07/
11/1
1Fo
r pe
rson
al u
se o
nly.
372 THE CANADIAN ENTOMOLOGIST March 1972
FIGS. 8-14. Male genitalia of Ips species and subspecies of groups IV and X and two Eurasian species: pini ( a ) , bonanseai (9), and avulsus (10) of group IV; sexdentatus (1 1) and dupli- catus (12) of Eurasia; interstitialis ( 1 3 ) and c. ponderosae (14) of group X. All figures are drawn to the scale indicated at right of plate. MS, median struts; ST, seminal trough; IS, internal sac; T, tegmen; ML, median lobe.
UNITED STATES Arizona: Santa Catalina Mts., Pima Co. from Pinus ponderosa, and P. strobi- formis, 8-IX-66, G. N. Lanier collector; Rustler Park, Chiricahua Mts., Cochise Co. from P. arizonica, IX-68, G. N. Lanier collector; Pinaleno Mts., Graham Co. from P. strobiformis, 15-VII-68, D. E. Bright collector.
MEXICO Michoacan: 31 mi E. Morelia, 2-VII-63, J. Doyan collector.
REMARKS. I . bonanseai frequently colonizes small trees or the limbs and upper portion of large trees infested by I. integer. In addition, it is probably found in association with I. interstitialis and I. lecontei.
Ips avulsus (Eichhoff) Tomicus avulsus Eichhoff 1867, p. 402.
BIOLOGICAL LITERATURE (Selected) Ips avulsus, Blackman 1922, p. 112; Doane et al. 1936, p. 92; Beal and Massey
1945, pp. 140-142; Craigliead 1950, p. 335; Beal el al. 1952, pp. 52-54;
Can
. Ent
omol
. Dow
nloa
ded
from
pub
s.es
c-se
c.ca
by
Nat
ural
Res
ourc
es C
anad
a on
07/
11/1
1Fo
r pe
rson
al u
se o
nly.

Volume 104 THE CANADIAN ENTOMOLOGIST 373
Bennett et al. 1958, pp. 7-9; Thatcher 1960; Wilkinson 1962, 1963; Vit6 et al. 1964; Yearian and Wilkinson 1967; Thomas 1967; Lanier and Cameron 1969; Lindquist 1969a; Mason 1969, 1970; Berisford et al. 1970. DIAGNOSIS. Characters differentiating avulsus from pini and bonanseai art
summarized under those species. KARYOLOGY. Four individuals from two broods had gonial cells with 32
chromosomes or PMI with 15AA + Xy, (Fig. 22). Three other specimens from a third brood had 34 gonial chromosomes and 16AA + Xy, (Fig. 23) or 15AA + Xy, + 2 univalents. The extra chromosomes are considered supernumerary.
Pachytene chromosomes are quite clearly defined (Fig. 21). Bivalent No. 1 has two well-separated heteropycnotic segments; other bivalents are marked by a single conspicuous dark segment and a number of small dots. The Xy, + nucleolus forms a moderately staining sphere.
At PMI bivalent No. 1 is conspicuous as a large ring (86% ) or cross (14% ). Other bivalents form rods or crosses (rarely rings) with minor breaks in size sequence between 2-3, 3-4, and 8-9. Supernumerary chromosomes usually formed a bivalent that was not distinguishable from the smaller autosomal bivalents. In several cases the supernumerary chromosomes remained unpaired but they did not form multivalent configurations with other chromosomes.
At MI1 chromosomes Nos. 1 and 3 were isobrachial crosses while No. 2 was distinctly heterobrachial with an AR of approximately 1:3. The larger of the remaining chromosomes were heterobrachial while the smaller appeared cephalo- brachial. The sex chromosomes were not individually identified.
GEOGRAPHIC VARIATION. None apparent in the material examined. TYPES. According to Hopping (1964) the holotype was probably destroyed
in World War 11. HOSTS. All southern pines and occasionally Pinus strobus (Beal and Massey
1945). DISTRIBUTION. Southeastern United States. The geographic range of avulsus
overlaps that of pini in the southern Appalachian Mountains, but the former infests pines in the Carolinian and transition zones while the latter is found in P. strobus in the higher Canadian zone (Ciesla, pers. comm.). Nevertheless, Beal and Massey (1945) report avulsus from P. strobus so it seems possible that this Ips could occur in cohabitation with pini.
NEW RECORDS. None. REMARKS. Recently I. avulsus has been characterized as an aggressive tree-
killer (Mason 1969, 1970). Under endemic conditions this species frequently utilizes the limbs and upper bole of trees infested by Dendroctonus spp., I. calli- graphus, and/or I. grandicollis.
Hopping (1964) and Lanier and Cameron (1969) noted no sex-associated differences in the declivital armature of avulsus. However, Wilkinson (pers. comm.) found that the second spine is well developed and connected to the third by a nearly straight ridge in males while in the females the second spine is less bulbous and well separated from the third. Using this character (designated b2 by Lanier and Cameron, 1969), I sexed 25 specimens with 100% accuracy. Separation of old adults should be less accurate because their declivital armature is apt to be abraded.
Can
. Ent
omol
. Dow
nloa
ded
from
pub
s.es
c-se
c.ca
by
Nat
ural
Res
ourc
es C
anad
a on
07/
11/1
1Fo
r pe
rson
al u
se o
nly.

3 74 THE CANADIAN ENTOMOLOGIST March 1972
Group X Key to the Species and Subspecies
1 . Distance of frontal tubercle from epistomal margin less than twice the width of the tubercle. Pronotal width usually greater than 2 mm. Interior western United States . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . calligraphus ponderosa,e Swaine
'1. Distance of frontal tubercle from epistomal margin 2 or more times the width of the tubercle. Pronotal width usually less than 2 mm; larger if specimens are from northeastern United States or adjacent parts of Canada . . . . . . . . . . . . . . 2
2 . Punctures on interstriae of elytra less than ?h the diameter of stria1 punctures. Width of the striation on pars stridens .91-1.21 p. Eastern North America and central California . . . . . . . . . . . . . . . . . . . . . calligraphus calligraphus (Germar)
'2. Punctures on interstriae of elytra equal to, or larger than, ?h the diameter of strial punctures. Width of striation on pars stridens .74-.90 p. Southern Arizona, southwestern New Mexico, Mexico, Central America, Caribbean Islands . . . . . . . . ................................................. interstitialis (Eichhoff)
Ips interstitialis (Eichhoff ) TAXONOMIC LITERATURE.
Tomicus interstitialis Eichhoff 1869, p. 273; Blandford 1895, p. 187. Ips interstitialis, Hagedorn 1910, p. 55; Schedl 1939, pp. 320-321; Hopping 1963,
p. 516; Schedl 1955, pp. 36-39. Ips calligraphus, Hopping 1965b (in part).
BIOLOGICAL LITERATURE (Selected) Ips interstitialis, Schwerdtfeger 1956; Lindquist 1969a "calligraphus (interstitialis)." Ips calligraphus, Etheridge 1968, pp. 75, 76.
DIAGNOSIS. I. interstitialis conforms closely to Hopping's ( 1965b) descrip- tion of I. calligraphus s. lato, but it differs from calligraphus s. str. in its geographic range and in the following characteristics: striations on the pars stridens of the female are .74-.90 p wide (51; = .832.021, Table 11) in interstitialis while they are usually more than .93 p wide in calligraphus; the pronotal width and the struts-lobe ratio of the male genitalia (Fig. 13) are generally less than those for the geo- graphically nearest calligraphus. In addition, interstitialis differs from calligraphus ponderosae in having a higher frontal tubercle (Figs. 3, 4) and from calligraphus calligraphus in having larger and more abundant punctures on the elytral interstriae.
KARYOLOGY. Unknown. A laboratory colony (Table I) was established but lost before cytological examination was attempted. Attempts with specimens preserved in carnoy were unproductive.
GEOGRAPHIC VARIATION. Specimens within or between series from the same general locality often differed considerably in size, but other morphological charac- teristics were quite consistent. Between-locality variation is apparent in Table I1 and Fig. 7. Mean pronotal width for series i-4 from middle Mexico (interstitialis-4 in Fig. 7 ) were very significantly (P < .01) greater than those for specimens from southern Arizona (i-2), the Dominican Republic (i-3), or southern Mexico and Central America (i-1). The second group, i-2, averaged significantly larger in size than i-1 or i-3. The mean width of striations on the pars stridens of i-1 females was less than those for other groups which showed no further significant differences in this characteristic. Other variation included the frontal tubercle which was uncharacteristically near the epistomal margin in some i-4 specimens and the interstrial punctures which were unusually small in a few of the i-3 specimens.
Can
. Ent
omol
. Dow
nloa
ded
from
pub
s.es
c-se
c.ca
by
Nat
ural
Res
ourc
es C
anad
a on
07/
11/1
1Fo
r pe
rson
al u
se o
nly.
Volume 104 THE CANADIAN ENTOMOLOGIST 375
TYPES. The holotype, a male, was examined. It is part of the Chapuis collection in the Institut Royal des Sciences Naturelles de Belgique, Brussels, and it bears the following labels: "Jamaica/T. interstitialis M/ T. interstitialis Eichh. Det. W. Eichhoff/Type/Dr. J. M. Swaine rev. 1932. Tomicus interstitialis Eichh./ R.1.Sc.N.B.I.G.".
HOSTS. Probably breeds in most pines within its range. I have seen speci- mens from Pinus arizonica, P. caribaea, P. nzontezumae, P. michoacana, P. occi- dentalis, P. oocarpa, and P. strobus var. chiapensis. In addition to these, Schedl (1955) lists P. hondurensis (= caribaea), P. pseudostrobus, P. rudis, and P. tenuifolia.
DI~TRIBUTION. The pine-growing regions from the mountains of Arizona and western New Mexico lying south of the Gila River, through Mexico, into the
FIGS. 15-23. Chromosomes of Ips species of group IV: pachytene in pini (15); PMI in western (16), eastern (17), and F, western x eastern (18) pini-all 15AA + Xy,; MI1 in eastern pir~i (19), n = 16; PMI in bonanseai (20), 15AA + Xy,; pachytene (21) in avulsus; PMI in avulsus with 15AA +Xy, (22) and 16AA ? + Xy, (23). Arrows indicate tripartite bivalcnts (T), heterornorphic bivalent (H), and sex bivalents (S).
Can
. Ent
omol
. Dow
nloa
ded
from
pub
s.es
c-se
c.ca
by
Nat
ural
Res
ourc
es C
anad
a on
07/
11/1
1Fo
r pe
rson
al u
se o
nly.

376 THE CANADIAN ENTOMOLOGIST March 1972
Caribbean Antilles (but not the Bahamas) (Fig. 2) . Hopping (19656) reported that this species has become established in the Philippine Islands where it attacks P. insularis.
MATERIAL EXAMINED. 22 1 specimens.
BRITISH HONDURAS Augustine (D. E. Etheridge collector).
BRITISH WEST INDIES Jamaica: Claredon, Christine (3 spm. collected by K. Hall, Feb. '66).
DOMINICAN REPUBLIC Jarabacoa (R. F. Billings collector).
GUATEMALA San Geronimo (This series of 4 specimens from the CNC includes the holotype, allotype, and 1 paratype of Ips blandfordi-a name which has no status in nomen- clature as it has not been published with a description of the species.)
MEXICO Chiapas: Altamirano; 5 mi N. Bochil; El Bosque; Lagos de Colores; jct. hwys. 190 and 195; 25 mi E. San Cristobal. Hidalgo: 230 km N. Mexico City near Zimapan. Mexico : Temascaltepec.
UNITED STATES Arizona: 20 mi S. Chiricahua Nat. Monument in Cochise Co.; Santa Catalina Mts. in Pima Co.
REMARKS. The concept of this species should be viewed with some skepticism owing to the considerable variation between specimens from widely separated localities and the relatively small number of localities represented. Reproductive isolation between interstitialis from the northern extreme of range and both calli- graphus subspecies is documented here. However, the potential for interbreeding between specimens from localities such as southern Mexico and the Caribbean Islands, and between these populations and calligraphus s. lato, has yet to be tested.
Ips calligraphus (Germar) TAXONOMIC LITERATURE
Bostricus calligraphus Germar 1824, p. 461. Tomicus calligraphus, Fitch 1858, p. 721 ; Leconte 1868, p. 162; 1876, pp. 362-
363. Ips calligraphus, Smith 1900, p. 363; Swaine 1909, pp. 120-121 ; Hagedorn 1910,
pp. 50-51; Swaine 1918, pp. 107, 112; Chamberlin 1939, pp. 410, 416-417; Schedl 1955, pp. 36-37; Hopping 1963, pp. 509, 516; 1965b (includes I. interstitialis Eichhoff) .
Bostrichus exesus Say 1826, p. 266. Syn. by Leconte 1876, p. 363. Bostrichus chloroticus Dejean 1837, p. 232. Syn. by Eichhoff 1878, p. 224. Bostrichus conformis Dejean 1837, p. 232. Syn. by Eichhoff 1878, p. 224. Tomicus praemorsus Eichhoff 1867, p. 401. Syn. by Eichhoff 1876, p. 378. Ips ponderosae Swaine 1925, p. 197; Chamberlin 1939, pp. 410, 417; Schedl 1955,
pp. 36-37;Hopping 1963,p. 516. Syn. by Hopping 19656.
Can
. Ent
omol
. Dow
nloa
ded
from
pub
s.es
c-se
c.ca
by
Nat
ural
Res
ourc
es C
anad
a on
07/
11/1
1Fo
r pe
rson
al u
se o
nly.
Volume 104 THE CANADIAN ENTOMOLOGIST 377
DIAGNOSIS. I . calligraphus conforms to Hopping's (1965b) description except as noted under the subspecies. The striations on the pars stridens of the female calligraphus are usually more than .93 /i wide while they are less than .90 p wide in interstitialis (Table 11). The pronotal width and the struts/lobe ratio of the male genitalia (Fig. 14) are generally greater than those for the geographically nearest interstitialis.
KARYOLOGY. The mitotic number is 32 and the meiotic formula of the male is 15AA + Xy,.
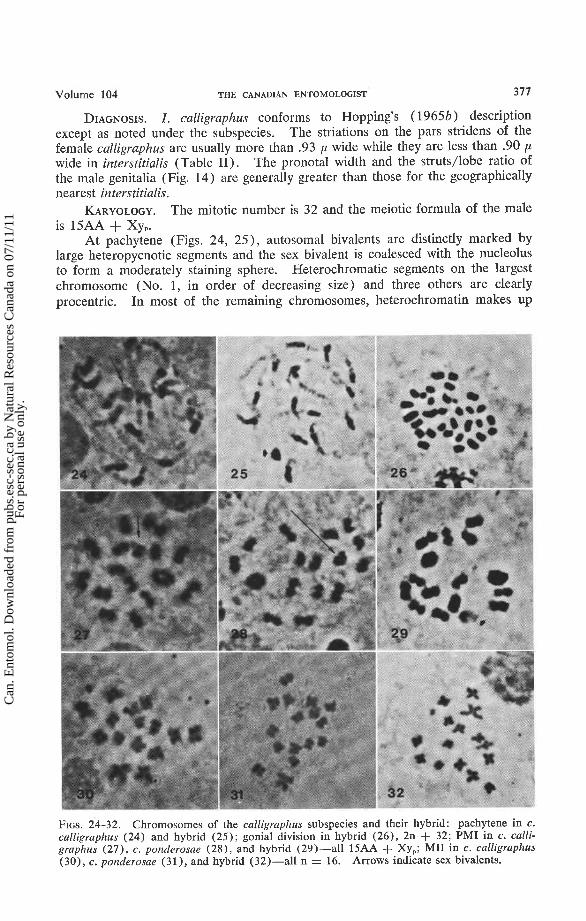
At pachytene (Figs. 24, 25)' autosomal bivalents are distinctly marked by large heteropycnotic segments and the sex bivalent is coalesced with the nucleolus to form a moderately staining sphere. Heterochromatic segments on the largest chromosome (No. 1, in order of decreasing size) and three others are clearly procentric. In most of the remaining chromosomes, heterochromatin makes up
FIGS. 24-32. Chromosomes of the calligraphus subspecies and their hybrid: pachytene in c . calligraphus (24) and hybrid (25); gonial division in hybrid (26), 2n + 32; PMI in c . calli- graphus (27), c . ponderosae (28), and hybrid (29)-all 15AA + Xy,; MI1 in c . calligraphus (30), c . ponderosae (31), and hybrid (32)-all n = 16. Arrows indicate sex bivalents.
Can
. Ent
omol
. Dow
nloa
ded
from
pub
s.es
c-se
c.ca
by
Nat
ural
Res
ourc
es C
anad
a on
07/
11/1
1Fo
r pe
rson
al u
se o
nly.

378 THE CANADIAN EPJTOMOLOGYST March 1972.
one arm, euchromatin the other. There are tiny darkly staining segments ("beads") within the euchromatic arms.
At diplotene-diakinesis every euchromatic-appearing arm is usually chiasmate. Thus, four bivalents may form rings while the remaining bivalent forms crosses or rods.
The bivalents contract unequally during diakinesis-PMI (Figs. 27-29), and by full MI only the largest (No. 1 ) and the two smallest (Nos. 14, 15) clearly differ from the others in size.
At PMII (Figs. 30-32), chromosome No. 2 forms an asymmetrical cross. No. 1 and other autosomes form symmetrical or slightly asymmetrical crosses. The X chromosome is isobrachial but the arms are usually contiguous and the y chromo- some appears as a dot.
REMARKS. Biological literature, geographic variation, types, hosts, distribu- tion, and material examined are treated under the subspecies.
Ips calligraphus calligraphus (Germar) BIOLOGICAL LITERATURE (Selected)
Ips calligraphus, Blackman 1922, p. 114; Craighead 1950, pp. 334-335; Doane et al. 1936, pp. 85, 86; Beal and Massey 1945, pp. 144-145; Beal et al. 1952, pp. 52-54; Bennett et al. 1958, pp. 7-9; Thatcher 1960; Wilkinson 1962, 1963; Yearian and Wilkinson 1963; VitC et al. 1964; Wilkinson 1964, pp. 63- 64; Yearian and Wilkinson 1967; Wilkinson et al. 1967; Thomas 1967; Wood and Stark 1968a, b; Barr 1969; Lanier and Cameron 1969 (refers in part to ponderosae s. str. ) ; Lindquist 1969a, b.
Ips ponderosae (California only), Keen 1929; Keen 1953, p. 146 (refers in part to c. ponderosae); Wood 1961, p. 187; VitC et al. 1964. DIAGNOSIS. I . calligraphus calligraphus conforms to Hopping's (1965b)
description. This subspecies differs from c. ponderosae by its generally smaller size, wider striations on the pars stridens (Table 11), and higther frontal tubercle. In addition, the punctures in the elytral interstriae in c. calligraphus are usually less than ?h the diameter of the stria1 punctures but greater than ?h this diameter in c. ponderosae (Figs. 5, 6 ) .
KARYOLOGY. AS described under calligraphus s. lato, the only apparent dif- ferences in the chromosomes of this subspecies and those of c. ponderosae were sequence and shape of heterochromatic "beads" on pachy-diplotene chromosomes.
GEOGRAPHIC VARIATION. Specimens from the Great Lakes region (c.c.-2, Table I1 and Fig. 7 ) were remarkably larger and more coarsely sculptured than those from the southeastern states and California (c.c.-1). Size variation is con- siderable within series from California or the southern pine regions but there seemed to be no consistent morphological or chromosomal differences between series from these localities.
TYPES. The holotype of I. calligraphus (Germar) is unknown to me. Type locality is Kentucky.
HOSTS. Attacks most pines within its range but apparently prefers thick- barked species. I have seen specimens from Pinus echinata, P. elliottii, P. ponderosa, P. resinosa, P. rigida, P. strobus, and P. taeda. In addition, Hopping (1965b) gives P. palustris and P. pungens, and Schedl (1955) lists P. hondurensis (= caribaea) , P. sabiniana, and P. silvestris (planted).
Can
. Ent
omol
. Dow
nloa
ded
from
pub
s.es
c-se
c.ca
by
Nat
ural
Res
ourc
es C
anad
a on
07/
11/1
1Fo
r pe
rson
al u
se o
nly.

Volume 104 THE CANADIAN ENTOMOLOGIST 379
DISTRIBUTION. This subspecies occurs in the extreme southern parts of eastern Canada, south through the lake states into the Appalachian Mountains to the Atlantic Coast and the Gulf of Mexico (Fig. 2) . It also occurs at low eleva- tions on the west slope of the central Sierra Nevada in California where it was apparently introduced (see text); other California localities given by Hopping (1965b) have been discounted (Wood and Stark 1968a, b).
MATERIAL EXAMINED. Over 500 specimens.
CANADA Ontario: Angus, Hamilton, Nairn, St. Williams, and 1000 Islands. Quebec: Province label only.
UNITED STATES California: Avery and Murphies in Calaveras Co.; Grass Valley in Nevada Co.; Placerville in El Dorado Co. Florida: Gainesville in Alachua Co.; Lake City in Columbia Co. Georgia: Athens (2 series) in Clarke Co.; Cairo in Grady CO.; Soperton in Treutlen Co. Massachusetts: Hyannis. Minnesota: St. Anthony Park and state label only. Mississippi: A & M (probably Mississippi State University at Starkville); Meridian in Lauderdale Co.; Saucier and County record only in Harrison Co. New York: Albany (2 series) in Albany Co.; Lake Placid in Essex Co.; Syracuse (2 series) in Onondaga Co.; Saratoga Springs in Saratoga Co. North Carolina: Asheville (4 series) and Bent Creek in Buncombe Co.; Durham in Durham Co. South Carolina: Boykin in Kershaw Co.
REMARKS. This subspecies is not known as an important tree-killer. It prefers thick-barked host material and normally attacks lower on the bole than sympatric Ips species which include grandicollis (Eichhoff) and avulsus (Eichhoff) in the southern pine region; grandicollis (= chagnoni Swaine) and pini (Say) in the eastern mountains, midwestern states, and southeastern Canada; paraconfusus Lanier, latidens (Leconte), and possibly sabinianae (Hopping) in California. I. c. calligraphus can easily be distinguished from these other Ips by its six declivital spines, by its larger size, and by its long, relatively large diameter galleries with coarse egg niches.
Ips calligraphus ponderosae Swaine BIOLOGICAL LITERATURE
Ips calligraphus, Hopkins 1902, p. 11; Lanier and Cameron 1969 (refers in part to c. calligraphus) .
Ips ponderosae, Doane et al. 1936, p. 86; Keen 1953, p. 146 (refers in part to c. calligraphus); Lindquist 19690, b. ("calligraphus (ponderosae)"). DIAGNOSIS. I. calligraphus ponderosae is based upon the concept of I.
ponderosae Swaine (1925). It conforms to Hopping's (1965b) description of I. calligraphus s. lato, except his analysis of the frontal tubercle, which was apparently based entirely upon I. calligraphus calligraphus. I. c. ponderosae differs from c. calligraphus by its generally larger size, narrower striations on the pars stridens (Table 11), and lower frontal tubercle (Fig. 4). The punctures in the interstrial spaces of the elytra are about !h the diameter of the stria1 punctures in c. ponderosae (Fig. 6) but less than this relative diameter in c. calligraphus (Fig. 5 ) .
Can
. Ent
omol
. Dow
nloa
ded
from
pub
s.es
c-se
c.ca
by
Nat
ural
Res
ourc
es C
anad
a on
07/
11/1
1Fo
r pe
rson
al u
se o
nly.

380 THE CANADIAN ENTOMOLOGIST March 1972
KARYOLOGY. Described under calligraphus s. lato. The karyology of this subspecies appeared to differ only in sequence and shape of heterochromatic "beads" on pachydiplotene chromosomes.
GEOGRAPHIC VARIATION. Size and other morphological characteristics were remarkably consistent within and between series from widely separated localities such as the Black Hills of South Dakota and Ruidoso, New Mexico.
TYPES. The holotype of I . ponderosae, a male, was examined. The speci- men is in the CNC and bears the following data: "Coconino N. F., Ariz. VI-1915/ Pinus ponderosa, lot 364". The allotype, also in the CNC, is labeled as follows: "Tusayan N. F. Ariz. VI-19 16/Pinus ponderosa, lot 348".
HOSTS. The vast majority of the collections of this subspecies were from Pinus ponderosa. In addition, I have seen specimens from P. banksiana planted near Halsey, Neb.
DISTRIBUTION. The arid ponderosa pine forests of the interior United States, east of the Continental Divide; southern Utah and Arizona, north of the Gila River (Fig. 2).
MATERIAL EXAMINED. Over 1000 specimens.
UNITED STATES Arizona: Prescott in Yavapai Co. (4 series); Mormon Lake in Coconino CO. Colorado: Allens Park in Boulder Co.; Castle Rock in Douglas Co.; Colorado Springs in El Paso Co.; Buckhorn Canyon in Larimer Co. Montana: Lodge Grass in Big Horn Co.; Clancy in Jefferson Co.; Nelson in Lewis and Clark CO.; 7 mi E. Ashland and Otter in Powder River Co.; Ashland and Lamedeer in Rosebud CO. Nebraska: Halsey in Thomas Co. (2 series). New Mexico: 5 mi SW. Capitan, Ruidoso and Cloudcroft in Lincoln Co.; Banderlier Nat. Monument in Santa Fe Co. South Dakota: Custer in Custer Co.; Camp Cook in Harding Co.; Deadwood in Lawrence Co.; Mt. Rushmore P. 0. and Sheridan Lake in Pennington Co.; Rosebud in Todd Co. Wyoming: Dayton in Sheridan Co.; Upton in Weston Co.
REMARKS. Like the other entities of group X, this subspecies attacks weakened or windthrown thick-barked trees. Owing to their large diameter and length, the gallery systems of c. ponderosae may be confused with those of I. integer (Eichhoff) which also breeds in large diameter ponderosa pine. However, the galleries are more coarsely sculptured and the egg niches are larger and more widely spaced than those of the latter species. In addition to integer, c. ponderosae may be associated with I. pini, latidens, lecontei (in Arizona), and cribricollis (in New Mexico).
BIOSYSTEMATIC EXPERIMENTS Breeding Tests
Group IV I . pini (oregonis) from Arizona, southern California, and three Sierra Nevada
populations were interfertile but egg hatchability differed significantly ( P < .05) for some reciprocal pairings. For example, 12 Arizona females mated by males of populations If, lg, and l h (Table I ) had an aggregate 1arvae:egg niche ratio (L/E) of .52 while the L/E for 9 reciprocal pairings was .81. Hatchability for 4 pairings of the F, inter se was .56 and L/Es for backcrosses to California and Arizona males were .70 and .27, respectively. However, the latter figure was based
Can
. Ent
omol
. Dow
nloa
ded
from
pub
s.es
c-se
c.ca
by
Nat
ural
Res
ourc
es C
anad
a on
07/
11/1
1Fo
r pe
rson
al u
se o
nly.
Volume 104 THE CANADIAN ENTOMOLOGIST 381
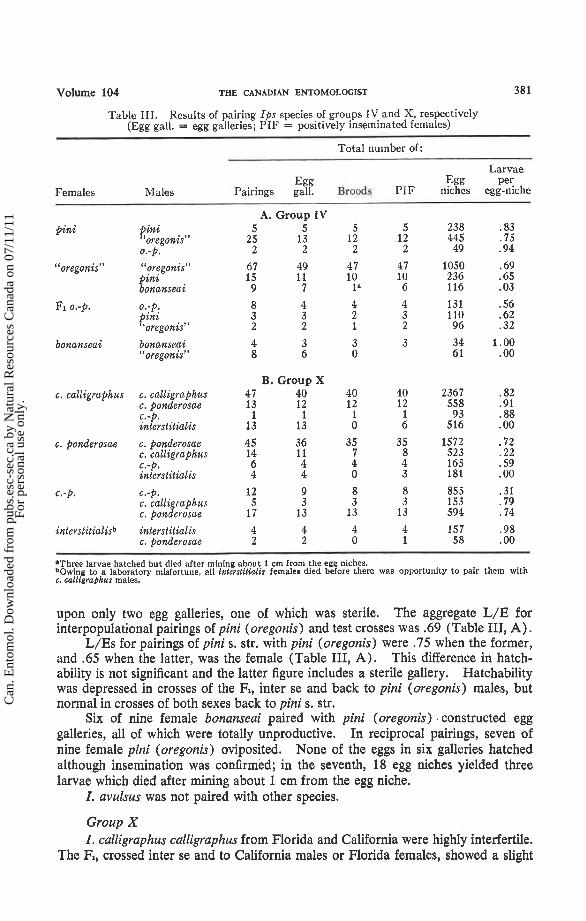
Table 111. Results of pairing Ips species of groups IV and X, respectively (Egg gall. = egg galleries; PIF = positively inseminated females)
Total number of:
Egg Egg Females Males Pairings gall. Broods PIF niches
A. Group IV pini t i n i 5 5 5 5 238
oregonis" 2 5 13 12 12 445 0.-p. 2 2 2 2 49
"oregonis" " prggonis" 67 49 4 7 47 1050 pznz 15 11 10 10 236 bonanseai 9 7 la 6 116
F1 0.-p. 0.-p. pini "oregonis"
bonanseai bonanseai 4 3 3 3 34 "oregonis" 8 6 0 6 1
B. Group X c. calligraphus c. calligraphus 47 40 40 40 2367
c. ponderosae 13 12 12 12 558 c.-p. 1 1 1 1 93 interstitielis 13 13 0 6 516
c. ponderosae c. ponderosae 45 36 35 35 1572 c. calligraphus 14 11 7 8 523 c.-p. 6 4 4 4 165 interstitialis 4 4 0 3 181
c.-P. 6.-p. 12 9 8 8 855 c. calli raphus 5 3 3 3 153 c, ponferosae 17 13 13 13 594
inlerslitialisb interstitialis 4 4 4 4 157 c. ponderosae 2 2 0 1 58
Larvae pe':
egg-nlche
'Three larvae hatched but died after mining about 1 cm from the egg niches. to a laboratory misfortune, all inlerslilialis females died before there was opportunity to pair them with
c. wlligraphus males.
upon only two egg galleries, one of which was sterile. The aggregate L/E for interpopulational pairings of pini (oregonis) and test crosses was .69 (Table 111, A).
L/Es for pairings of pini s. str. with pini (oregonis) were .75 when the former, and .65 when the latter, was the female (Table 111, A). This difference in hatch- ability is not significant and the latter figure includes a sterile gallery. Hatchability was depressed in crosses of the PI, inter se and back to pini (oregonis) males, but normal in crosses of both sexes back to pini s. str.
Six of nine female bonanseai paired with pini (oregonis) .constructed egg galleries, all of which were totally unproductive. In reciprocal pairings, seven of nine female pini (oregonis) oviposited. None of the eggs in six galleries hatched although insemination was confirmed; in the seventh, 18 egg niches yielded three larvae which died after mining about 1 cm from the egg niche.
I , avulsus was not paired with other species.
Group X I . calligraphus calligraphus from Florida and California were highly interfertile.
The F,, crossed inter se and to California males or Florida females, showed a slight
Can
. Ent
omol
. Dow
nloa
ded
from
pub
s.es
c-se
c.ca
by
Nat
ural
Res
ourc
es C
anad
a on
07/
11/1
1Fo
r pe
rson
al u
se o
nly.

382 THE CANADIAN ENTOMOLOGIST March 1972
reduction in fertility (.84-.95 for P, vs. .62-.69 for F,). The aggregate L/E for intrasubspecific pairings was .82 (Table 111, B).
I. c. ponderosae from New Mexico and South Dakota were also interfertile although the L/Es for both crosses were .69. No significance can be attributed to this low hatchability because the figure for intrapopulational (control) pairings of South Dakota beetles was also .69. Hatchability in the F2 and backcrosses were higher (L/Es = .73-.78), but still slightly below that of New Mexico controls (.85). The aggregate L/E for these pairings was .72 (Table 111, B) .
Fertility of the reciprocal crosses of the calligraphus subspecies differed drastically. The L/E was .91 when c. calligraphus was the female while that for the reciprocal cross was .22 (Table 111, B) . In the latter case, all of the 154 eggs in four galleries excavated by c. ponderosae females failed to hatch while individual L/Es for seven brood-producing galleries were .77, .76, .55, .49, .45, .15, and .09. This unequal fertility appears to have been maintained in backcrosses of F, males, although the degree of difference (.88 vs. .59) was less marked. Eggs laid by Fl females backcrossed to both subspecies showed intermediate fertility (.74-.79) and the L/E for the F? was .3 1.
I. interstitialis and both calligraphus subspecies appeared to be totally incapable of interbreeding. All of the 19 interspecific pairings resulted in egg galleries, but none of the eggs hatched even though 10 of 11 females recovered alive had sperm in their spermathecae. Four control pairings of interstitialis had a L/E of .98 (Table 111, B). The interstitialis females were accidentally frozen before there was opportunity to pair them with c. calligraphus males.
Karyology of the Hybrids Group IV Squash preparations of testes from two F, pini of the Arizona X southern
California (populations li, and lg, Table I) cross contained numerous clear meiotic cells. Bivalent formation was invariably complete and chiasmata number was normal, but two or more of the middle-sized bivalents appeared heteromorphic (Fig. 18) . This condition masked the break in size sequence between bivalents 4 and 5 which was observed in both parent populations. Evidently, some of the chromosomes from the same size rank in the two populations are not homologous. Similarly, four Fl of the Ontario and British Columbia colonies (i.e., pini s. str. and "oregonis"; l b and lc, Table I ) showed normal meiosis but a few of the middle- sized bivalents appeared heteromorphic. This inequality in size does not neces- sarily reflect genetic divergence because the difference may reside in the quasi-inert heterochromatic arms (Lanier 1 9 7 0 ~ ) .
Group X There were no disruptions in pairing and no heteromorphic bivalents evident
in 12 preparations from Fl of interpopulational pairings (California-lb X Florida- l c ) of c. calligraphus. Heterochromatic "beads" on pachydiplotene chromosomes appeared to match perfectly.
Meiosis in Fl of interpopulational pairings of c. ponderosae (New Mexico-2a and South Dakota-2b) was also normal except for putative tetravalent crosses observed in one of the six individuals examined. No differences in alignment of "beads" were detected although the early stages of meiosis were not very clear in this material.
Can
. Ent
omol
. Dow
nloa
ded
from
pub
s.es
c-se
c.ca
by
Nat
ural
Res
ourc
es C
anad
a on
07/
11/1
1Fo
r pe
rson
al u
se o
nly.
Volume 104 THE CANADIAN ENTOMOLOGIST 383
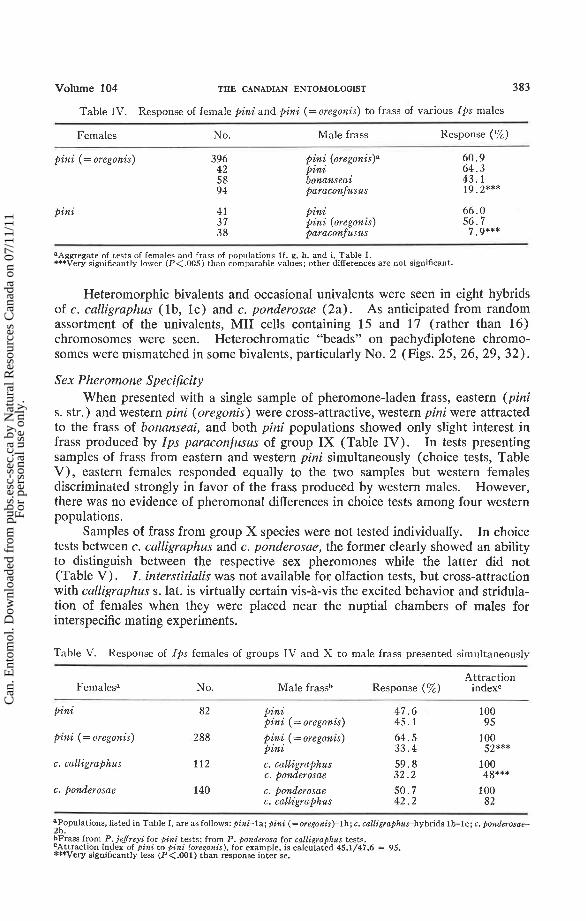
Table IV. Response of female pin i and pini (=oregonis) t o frass of various I p s males
Females No. Male frass Response (%)
pini (= oregonis)
pini
pin! (oregonis)" pinz bonanseai paraconfusus
4 1 pini 37 pini (oregonis) 3 8 paraconfusus
aAggregate of tests of females and frass of populations If, g, h, and i. Table I. ***Very significantly lower (P<.005) than comparable values; other differences are not significant.
Heteromorphic bivalents and occasional univalents were seen in eight hybrids of c. calligraphus ( l b , l c ) and c. ponderosae (2a). As anticipated from random assortment of the univalents, MI1 cells containing 15 and 17 (rather than 16) chromosomes were seen. Heterochromatic "beads" on pachydiplotene chromo- somes were mismatched in some bivalents, particularly No. 2 (Figs. 25, 26, 29, 32).
Sex Pheromone Specificity When presented with a single sample of pheromone-laden frass, eastern (pini
s. str.) and western pini (oregonis) were cross-attractive, western pini were attracted to the frass of bonanseai, and both pini populations showed only slight interest in frass produced by Ips paraconfusus of group IX (Table IV). In tests presenting samples of frass from eastern and western pini simultaneously (choice tests, Table V), eastern females responded equally to the two samples but western females discriminated strongly in favor of the frass produced by western males. However, there was no evidence of pheromonal differences in choice tests among four western populations.
Samples of frass from group X species were not tested individually. In choice tests between c. calligraphus and c. ponderosae, the former clearly showed an ability to distinguish between the respective sex pheromones while the latter did not (Table V). I. interstitialis was not available for olfaction tests, but cross-attraction with calligraphus s. lat. is virtually certain vis-8-vis the excited behavior and stridula- tion of females when they were placed near the nuptial chambers of males for interspecific mating experiments.
Table V. Response of I p s females of groups IV and X to male frass presented simultaneously
Attraction Femalesa No. Male frassb Response (%) indexc
pini 82 pin i 47.6 100 pini (= oregonis) 45.1 95
pini (= oregonis) 288 pini (= oregonis) 64.5 100 pin i 33.4 52***
c. calligraphus 112 c. calligraphus 59.8 100 c. ponderosae 32.2 48***
c. ponderosae 140 c. pondevosae 50.7 100 c. calligraphus 42.2 82
~populations, listed in Table I, are as follows: pini-la; pini (=oregonis)-lh; c. calligraphus-hybrids lb-lc; c. ponderosae 2b. bFrass from P. jeffreyi for pini tests. from P. pondevosa for calligraphus tests. cAttraction index of pini to pini (orkgonis). for example. is calculated 45.1/47.6 = 95. ***Very significantly less (P<.001) than response inter se.
Can
. Ent
omol
. Dow
nloa
ded
from
pub
s.es
c-se
c.ca
by
Nat
ural
Res
ourc
es C
anad
a on
07/
11/1
1Fo
r pe
rson
al u
se o
nly.

384 THE CANADIAN ENTOMOLOGIST March 1972
Elgect of the Host One of the characteristic differences between the taxa within groups IV and X
is their sizes. All laboratory colonies were reared in ponderosa pine under near optimal conditions. Size variation within colonies appeared relatively slight as compared to that within some of my series of wild beetles such as pini from Sault Ste. Marie, Ont., or c. calligraphus from Avery, Calif. However, differences between taxa, as reflected in Table 11, were clearly maintained. A previous study (Lanier 1970c) showed that host species, per se, had no significant effect on the size of I. plastographus or integer.
DISCUSSION AND CONCLUSIONS Species Validity
The interfertility of individuals from widely separated localities, together with cross-attractiveness to the respective pheromones and the lack of morphological and cytological distinctiveness, confirm Hopping's (1964) synonomization of oregonis with pini. Converse results clearly demonstrate the validity of bonanseai, as distinct from pini. I. avulsus is so different from the other group IV species that breeding experiments seemed unnecessary. Nevertheless, I consider the grouping of avulsus with pini and bonanseai (Hopping 1963) to be valid.
I. interstitialis is reinstated to species status because of its total lack of inter- fertility with calligraphus s. lat. and because of the consistent differences in width of striations on the pars stridens of the respective species. However, the concept of this species cannot be considered to be totally objective because a single colony, and that from an extreme of the species range, was tested in breeding experiments. 1. c. ponderosae is elevated from synonymy (Hopping 1965b) to subspecies because it represents a morphologically and cytologically defined, geographically isolated entity.
The hypothesis that c. calligraphus from California and the southeastern states is of the same origin is clearly confirmed by the complete interfertility, by the extreme similarities in morphological and cytological detail, and by the identity of parasitic mites on these beetles.
Non-Reciprocal Fertility The striking inequality in hatchability between reciprocal matings of the calli-
graphus subspecies and the increase in fertility of one backcross (c. ponderosae female X F1 male), relative to that of the original mating (c. ponderosae female X c. calligraphus male) (Table 111, B), do not have an adequate cytogenetic explana- tion. Embryo mortality can not be due to imbalance of autosomal genes because the same combinations were possible in both crosses. Nor can this unequal hatchability be attributed to sex-linked genes-this would distort the ratio of males to females in the Fl and/or backcross generations, but the sex ratio in these broods rarely deviated from 1 : 1.
I believe these results reflect cytoplasmic incompatibility similar to that between 1. integer and I . plastographus of group I11 (Lanier 1970c, 1971). Specifically, it appears that c. ponderosae ooplasm is antagonistic toward the c. calligraphus sperm and/or the hybrid embryo, but there is no such interaction in the reciprocal cross. Furthermore, the absolute inability of interstitialis and calligraphus s. lat. to cross could be a result of strong cytoplasmic incompatibility in both directions. Unfortunatery, breeding data do not permit decision between cytoplasmic and genetic explanations in cases of complete intersterility.
Can
. Ent
omol
. Dow
nloa
ded
from
pub
s.es
c-se
c.ca
by
Nat
ural
Res
ourc
es C
anad
a on
07/
11/1
1Fo
r pe
rson
al u
se o
nly.

Volume 104 THE CANADIAN ENTOMOLOGIST 385
Species Relationships Within group IV, pini and bonanseai are most closely related. The male
genitalia of avulsus (Fig. 10) show greatest similarity to those of bonanseai (Fig. 9 ) , but the elongate structure and well-developed internal sac also suggest phylogenetic ties to group I1 (emarginatus Leconte illustrated by Hopping, 1963). In other characters such as length/width of the elytra, punctation, and width of striations on the pars stridens, avulsus is most similar to pini.
The relative closeness of the relationships between interstitialis and the calli- graphus subspecies is difficult to establish. Lindquist (1969a) considered the Iponemus mites parasitizing interstitialis in southern Mexico to be calligraphus cordillerae Lindquist, the same subspecies that is found on Ips c. ponderosae. Pars stridens measurements (Table 11) support the inference that intersritialis is closest to c. ponderosae but comparison of frontal tubercles and the elytral puncturing suggests that interstitialis is most closely allied to c. calligraphus.
I . duplicatus and sexdentatus should not be included in groups IV and X, res- pectively. I. duplicatus lacks a vertex-pronotal type stridulating mechanism and its male genitalia (Fig. 12) show little similarity to those of group IV species (Figs. 8-10). Striations on the pars stridens of sexdentatus are much wider than those on group X species (Table 11) and the male genitalia are so different that com- parisons are preposterous (Figs. 11, 13, 14) . In addition to these differences, it was previously pointed out that parasitic lponemus mites on the Eurasian species show no clear affinities for those on the Ips of groups IV and X (Lindquist 1969a) and that the declivital spines on sexdentatus and North American 6-spined Ips are not homologous (Leconte 1876).
I concur with Lindquist's ( 1 9 6 9 ~ ) opinion that groups 111, IV, IX, and X evolved from common stock in North America; therefore, barring emigration, one should not expect members of these groups to be found in Eurasia.
Sex Pheromones and Interbreeding in Nature The clear preference of western pini females for frass produced by western
(vs. eastern) males and that of female c. calligraphus for the frass of its own males (vs. c. poderosae) (Table V ) demonstrates differences in both pheromone and pheromone receptors of the respective entities. This same type of unilateral dis- crimination occurs between several group IX Ips (Wood 1970; Lanier and Wood, unpub. data). However, there is no reason to expect that these slight differences would prevent interspecific mating in nature because the preferences are not absolute and the discrimination is unilateral. Progeny from these unions should show even less propensity to segregate; Lanier (1970a, c ) demonstrated the breakdown of specificity in hybrids of non-crossattractive species of Ips. The role of sex phero- mones in speciation will be discussed in more detail in a subsequent paper.
ACKNOWLEDGMENTS Facilities and support for this study were provided by the Canada Department
of Fisheries and Forestry and, to a lesser extent, the University of California at Berkeley and the State University of New York College of Forestry at Syracuse University. D. L. Wood of the University of California guided the early part of this work, and technical assistance was ably provided by G. A. Schofer of the Canada Department of Fisheries and Forestry.
Living specimens were provided by J. B. Thomas of the Canada Department of Fisheries and Forestry, Sault Ste. Marie, Ont.; R. C. Wilkinson of the University
Can
. Ent
omol
. Dow
nloa
ded
from
pub
s.es
c-se
c.ca
by
Nat
ural
Res
ourc
es C
anad
a on
07/
11/1
1Fo
r pe
rson
al u
se o
nly.

386 THE CANADIAN ENTOMOLOGIST March 1972
of Florida, Gainesville; and D. T. Jennings of the Rocky Mountain Forest and Range Experiment Station, Albuquerque, N.M. Preserved specimens were loaned by R. Damoiseau of the Institut Royal des Sciences Naturelles de Belgique; J. P. Vit6 of the Boyce Thompson Institute, Beaumont, Texas; D. E. Bright of the Entomology Research Institute, Ottawa; and S. L. Wood of Brigham Young Univer- sity, Provo, Utah; and several institutions.
To all, a sincere "thank you".
REFERENCES Anderson, R. F. 1948. Host selection by the pine engraver. J. econ. Ent. 41: 596-602. Barr, B. A. 1969. Sound production in Scolytidae (Coleoptera) with emphasis on the genus
Ips. Can. Ent. 101: 636-672. Beal, J. A., W. Haliburton, and F. B. Knight. 1952. Forest insects of the southeast: with
special reference to species occurring in the Piedmont Plateau of North Carolina. Duke Univ. School Forestry Bull. 14, 168 pp.
Beal, J. A. and C. L. Massey. 1945. Bark beetles and ambrosia beetles (Coleoptera: Scolytidae) with special reference to species occurring in North Carolina. Duke Univ. School Forestry Bull. 10, 178 pp.
Bennett, W. H., C. W. Chellman, and W. R. Holt. 1958. Insect enemies of southern pines. U.S. For. Serv. Sth. For. Exp. Stn Occ. Pap. 164, 130 pp.
Berisford, C. W., H. M. Kulman, and R. L. Pienkowski. 1970. Notes on the biologies of hymenopterous parasites of Ips spp. bark beetles in Virginia. Can. Ent. 102: 484-490.
Blackman, M. W. 1922. Mississippi Bark Beetles. Miss. Exp. Strz Bull. 11, 130 pp. Blandford, W. F. H. 1895. Fam. Scolytidae. In Biologia Centrali-Americana, Vol. 6, pp.
81-298. Boss, G. D. and T. 0. Thatcher. 1970. Mites associated with Ips and Dendroctonus in
southern Rocky Mountains with special reference to lponemus truncatus (Acarina: Tarsonemidae). U.S. Dep. Agric. For. Serv. Res. Note RM-171.
Chamberlin, W . J. 1939. The bark and timber beetles of North America. Ore. St. Coll. Coop. Assoc., Corvallis. 513 pp.
1958. The Scolytoidea of the Northwest: Oregon, Washington, Idaho and British Columbia. Ore. St. Monog. Stud. Ent., No. 2, 205 pp.
Ciesla, W. M. and J. C. Bell. 1968. The pine engraver, Ips pirli (Coleoptera: Scolytidae), in the southern Appalachian Mountains. Ann. ent. Soc. Am. 61: 235-236.
Clemens, W. A. 1916. The pine bark beetle (Ips pini (Say)). Corrtell Uriiv. agric. Exp. Stn Bull. 383, pp. 287-298.
Craighead, F. C. 1950. Insect enemies of eastern forests. U.S. Dep. Agric. Misc. Publ. 657. Craighead, F. C. and W. Middleton. 1930. An annotated list of the important North
American forest insects. U.S. Dep. Agric. Misc. Publ. 74. Dejean. 1837. Catalogue des Col6opt&res de la collection de le baron Dejean. Ed. IV.
503 pp. Doane, R. W., et al. 1936. Forest insects. McGraw-Hill, New York and London. Eichhoff, W. 1867. Neue amerikanische Borkenkafer-Gattungen und Arten. Berl. ent. Z.
11: 399-402. - 1869. Neue Amerikanische Borkenkafer-Gattung und Arten. Berl. ertt. Z. 12: 273-
280. 1876. Synoptical observations. Stettin. ent. Ztg. 37: 378. 1878. Ratio descriptio, emendatio, eorum Tomicinorum. Mern. Soc. r. Sci. LiLge
7: 1-531. Etheridge, D. E. 1968. Preliminary observations on the pathology of Pirzus caribaea Morelet
in British Honduras. Commorzw. For. Rev. 47: 131. Felt, E. P. 1906. Pine bark beetle. In Insects affecting park and woodland trees. Mern.
N.Y. St. Mus. 8: 351-354. '
Fitch, A. 1858. Fourth report on the noxious, beneficial and other insects of the state of New York.
Germar, E. F. 1824. Insectorum Species-Novae Hut Minus Cignitae, Descriptionibus Illustratae.
Hagedorn, M. 1910. Coleopterum Catalogus, Pars IV, Ipidae. Junk, Berlin.
Can
. Ent
omol
. Dow
nloa
ded
from
pub
s.es
c-se
c.ca
by
Nat
ural
Res
ourc
es C
anad
a on
07/
11/1
1Fo
r pe
rson
al u
se o
nly.

Volume 104 THE CANADIAN ENTOMOLOGIST 387
Hopkins, A. D. 1902. Insect enemies of the pine in the Black Hills Forest Reserve. U.S. Dep. Agric. Div. Ent. Bull. 32, 24 pp.
1905. Notes on some Mexican Scolytidae with descriptions of some new species. Proc. ent. Soc. Wash. 7: 71-79.
Hopping, G. R. 1963. The natural groups of species in the genus Ips De Geer (Coleoptera: Scolytidae) in North America. Can. Ent. 95: 508-516.
1964. The North American species in groups IV and V of Ips De Geer (Coleoptera: Scolytidae). Can. Ent. 96: 970-978.
1965a. The North American species of group VI of Ips De Geer (Coleoptera: Scolytidae). Can. Ent. 97: 533-541.
1965b. The North American species in group X of Ips De Geer (Coleoptera: Scolytidae). Can. Ent. 97: 803-809.
Johnson, P. C. 1967. Distribution of bark beetle attacks on ponderosa pine trees in Montana. U.S. For. Serv. Res. Note Int-62, 7 pp.
Keen, F. P. 1929. Insect enemies of California pines and their control. St. Calif. Dep. Nut. Res., Div. For. Bull. 7 .
1953. Insect enemies of western forests. Misc. Publs U.S. Dep. Agric., NO. 273, 2nd ed.
Lanier, G. N. 1966. Interspecific mating and cytological studies of closely related species of Ips De Geer and Orthotomicus Ferrari (Coleoptera: Scolytidae). Can. Ent. 98: 175-188.
1970a. Sex pheromones: abolition of specificity in hybrid bark beetles. Science 169: 71-72.
19706. Biosystematics of North American Ips (Coleoptera: Scolytidae). Hopping's group IX. Can. Ent. 102: 1 139-1 163.
1970c. Biosystematics of the genus Ips (Coleoptera: Scolytidae) in North America. Hopping's group 111. Can. Ent. 102: 1404-1423.
1971. Cytoplasmic incompatibility and breeding isolation in bark beetles (Coleop- tera: Scolytidae). Can. I . Genet. Cytol. 13: 160-163.
Lanier, G. N. and E. A. Cameron. 1969. Secondary sex characters in North American species of the genus Ips (Coleoptera: Scolytidae). Can. Ent. 101: 862-870.
Leconte, J. L. 1868. Synopsis of Scolytidae of North America (taxonomic). Discussion, keys to species, 19 genera, 96 species. Trans. Am. ent. Soc. 2: 150-178.
1876. Family IX. Scolytidae. In J. L. Leconte and G. H. Horn, The Rhyncho- phora of America north of Mexico. Proc. Am. philos. Soc. 15: 341-391.
Lindquist, E. E. 1969a. Review of holarctic tarsonemid mites (Acarina: Prostigmata) para- sitizing eggs of ipine bark beetles. Mem. ent. Soc. Can., No. 60.
19696. New species of Tarsonemus (Acarina: Tarsonemidae) associated! with bark beetles. Can. Ent. 101: 1291-1314.
Mason, R. R. 1969. Behavior of Ips populations after summer thinning in a loblolly pine plantation. For. Sci. 15: 390-398.
1970. Comparison of flight aggregation in two species of southern Ips (Coleoptera: Scolytidae). Can. Ent. 102: 1036-1041.
Miller, I. M. and F. P. Keen. 1960. Biology and control of the western pine beetle. U.S. Gov. Printing Off., Washington, D.C. 381 pp.
Mirov, N. T. 1967. The genus Pinus. Ronald Press, New York. Reid, R. W. 1955. The bark beetle complex associated with lodgepole pine slash in Alberta.
I. Notes on the biologies of some Scolytidae attacking lodgepole pine slash. Can. Ent. 57: 311-323.
1957. The bark beetle complex associated with lodgepole pine slash in Alberta. IV. Distribution, population densities, and effects of several environmental factors. Can. Ent. 89: 437-447.
Say, T. 1826. Descriptions of new species of coleopterous insects inhabiting the United States. I. Acad. Nut. Sci. Philad. 5: 257, 266.
Schedl, K. E. 1939. Scolytidae, Coptonotidae Platypodidae mexicanos. An. Es. Nac. Cienc. Biol. 1: 317-379.
1955. Die Kiefern-Borkenkafer Guatemalas. 2. angew. Ent. 38: 1-48. Schenk, J. A. and D. M. Benjamin. 1969. Notes on the biology of Ips pini in central
Wisconsin jack pine forests. Ann. ent. Soc. Am. 62: 480-485. Schwerdtfeger, F. 1956. Scolytidae (Col.) on Pinus-Arten in Mittelamerika. I. Das Genus
Ips De Geer. 2. angew. En!. 39: 34-57.
Can
. Ent
omol
. Dow
nloa
ded
from
pub
s.es
c-se
c.ca
by
Nat
ural
Res
ourc
es C
anad
a on
07/
11/1
1Fo
r pe
rson
al u
se o
nly.

388 THE CANADIAN ENTOMOLOGIST March 1972
Smith, J. B. 1900. Insects of New Jersey. N.J. St. Bd Agric. 755 pp. Smith, S. G. 1952. The cytology of Sitophilus (= Calandra) oryzae (L.), S. grarlarius (L.)
and some other Rhynchophora (Coleoptera). Cytologia 17: 50-70. Swaine, J. M. 1909. Catalogue of the described Scolytidae of America, north of Mexico.
Appendix B in Report of the State Entomologist on injurious and other insects of the state of New York. N.Y. St. Mus. Bull. 134: 76-159.
1918. Canadian bark beetles. 11. A preliminary classification, with an account of the habits and means of control. Tech. Bull. Can. Dep. Agric., No. 14.
1925. New species of Ipidae (Coleoptera). Can. Ent. 58: 192-197. Thatcher, R. C. 1960. Bark beetles affecting southern pines: A review of current knowledge.
U.S. Dep. Agric. For. Serv. Sth. For. Exp. Stn, Occ. Pap. 180. Thomas, J. B. 1961. The life history of Ips pini (Say) (Coleoptera: Scolytidae). Can. Ent.
93: 384-390. - 1966. Some Scolytidae from the Sierra Madre Occidental in Mexico. Can. Ent.
98: 871-875. 1967. A comparative study of gastric caeca in adult and larval stages of bark
beetles (Coleoptera: Scolytidae). Proc. ent. Soc. Ont., Vol. 97, pp. 71-90. Vit6, J. P., R. I. Gara, and H. D. von Scheller. 1964. Field observations on the response to
attractants of bark beetles infesting southern pines. Contr. Boyce Thompson Inst. P1. Res. 22: 461-470.
Wilkinson, R. C. 1962. Stridulating organs in three southeastern Ips bark beetles. Flu Ent. 45: 43-44.
1963. Larval instars and head capsule morphology in three southeastern Ips bark beetles. Flu Ent. 46: 19-22. - 1964. Attraction and development of Ips bark beetle populations in artificially
infested pine bolts exposed on fire towers and turntables in Florida. Flu Ent. 47: 57-64. Wilkinson, R. C., W. T. McClelland, R. M. Murillo, and E. 0. Ostmark. 1967. Stridulation
and behavior in two southeastern Ips bark beetles (Coleoptera: Scolytidae). Flu Ent. 50: 185-195.
Wood, D. L. 1961. Stridulation in the genus Ips De Geer. Pan-Pacif. Ent. 37: 187-188. - 1970. Pheromones of bark beetles. In D. L. Wood, R. M. Silverstein and M.
Nakajima, Control of insect behavior by natural products. Academic Press, New York. Wood, D. L, and R. W. Bushing. 1963. The olfactory response of Ips confusus (LeConte)
(Coleoptera: Scolytidae) to the secondary attraction in the laboratory. Can. Ent. 95: 1066-1078.
Wood, D. L. and R. W. Stark. 1968a. The life history of Ips calligrapl~us (Coleoptera: Scolytidae) with notes on its biology in California. Carl. Ent. 100: 145-151.
1968b. Addendum. Can. Ent. 100: 548. Wygant, N. D. and R. R. Lara. 1967. Pine engraver Ips pini (Say), pp. 117-119. In
Important forest insects of mutual concern to Canada, the United States and Mexico. Queen's Printer, Ottawa.
Yearian, W. C. and R. C. Wilkinson. 1963. An artificial rearing medium for Ips calligraphus Germ. Flu Ent. 46: 319-320.
1967. Development of three Ips bark beetles on a phloem-based rearing medium. Flu Ent. 50: 43-45.
(Received 22 June 1971)
Can
. Ent
omol
. Dow
nloa
ded
from
pub
s.es
c-se
c.ca
by
Nat
ural
Res
ourc
es C
anad
a on
07/
11/1
1Fo
r pe
rson
al u
se o
nly.





























