Embed Size (px)
Citation preview

Ct
Ma
b
a
ARRAA
KTCAOTR
1
craTllit2Ea(
Pcsflpo
0d
Toxicology Letters 204 (2011) 2–11
Contents lists available at ScienceDirect
Toxicology Letters
journa l homepage: www.e lsev ier .com/ locate / tox le t
-Phycocyanin: An effective protective agent against thymic atrophy byributyltin
onika Guptaa, Upendra Nath Dwivedib, Shashi Khandelwala,∗
Indian Institute of Toxicology Research, P. Box 80, Mahatma Gandhi Marg, Lucknow 226001, IndiaDepartment of Biochemistry, Lucknow University, Lucknow 226001, India
r t i c l e i n f o
rticle history:eceived 10 February 2011eceived in revised form 28 March 2011ccepted 28 March 2011vailable online 6 April 2011
eywords:ributyltin-Phycocyanin
a b s t r a c t
Spirulina platensis, used worldwide as a food supplement, is a natural source of protein, vitamins, car-bohydrates and polyunsaturated fatty acids. C-Phycocyanin (C-Pc), its major biliprotein, is known topossess anti-oxidant, anti-inflammatory and radical scavenging properties. Our present study showedthat treatment with C-Pc protects the rats from Tributyltin (TBT) induced thymic atrophy. The resultsreveal TBT-induced oxidative stress mediated apoptosis in rat thymocytes in vivo and its attenuationby C-Pc. This ameliorative effect could be attributed to antioxidant activity of the biliprotein. C-Pc alsoincreased TBTC reduced thymic weight and cellularity as well. TBTC-induced ROS generation and loweredGSH levels were restored by C-Pc, suggesting its radical scavenging properties. The various apoptotic
poptosisxidative stresshymic atrophyat
determinants such as mitochondrial membrane potential, Bax/Bcl-2 ratio, caspase-3 activity and apo-ptotic cell population were effectively modulated by C-Pc treatment. We make this first observation toillustrate the effectiveness of C-Pc in reducing TBTC-induced thymic atrophy. The morphology of thymictissue was restored to near normal by this biliprotein. The present study, therefore, suggests that C-Pc could serve as an effective natural antioxidant for efficient management of TBTC induced oxidativedamage.
. Introduction
Organotins are man-made compounds with numerous commer-ial applications like heat stabilisers in PVC, catalysts in chemicaleactions, glass coatings, agricultural pesticides, biocides in marinentifoulant paints, wood treatments and preservatives (Batt, 2006).heir extensive use to control biofouling tends to contaminateakes, rivers and coastal environments and pose a threat to marineife (Choi et al., 2009; Eguchi et al., 2010). TBT is reported to bemmuno-, neuro-, hepato-, nephro-, gastro- toxic and also causesesticular damage (Snoeij et al., 1987; Kotake, 2007; Zhang et al.,009). The most sensitive organ to tributyltin toxicity is thymus.
xperimental animal studies indicate that tributyltin compoundsre immunotoxic at doses below those affecting other target organsKrajnc et al., 1984; Boyer, 1989). In rats, organotin compoundsAbbreviations: CMF-DA, 5′-chloromethylfluorescein diacetate; C-Pc, C-hycocyanin; DCFH-DA, 2′7′-dichlorofluorescein diacetate; DEVD-AFC, benzyloxy-arbonyl-Asp-Glu-Val-Asp-7-amino-4-trifluoromethyl-coumarin; DMSO, dimethylulfoxide; EDTA, ethylenediamine tetracetic acid; FBS, fetal bovine serum; FITC,uorescein-5-isothiocyanate; GSH, glutathione; HE, hematoxylin eosin; PBS, phos-hate buffered saline; PI, propidium iodide; Rh 123, rhodamine 123; ROS, reactivexygen species; TBTC, tributyltin chloride; TE, Tris–EDTA.∗ Corresponding author. Tel.: +91 0522 2627586; fax: +91 0522 2628227.
E-mail address: skhandelwal [email protected] (S. Khandelwal).
378-4274/$ – see front matter © 2011 Elsevier Ireland Ltd. All rights reserved.oi:10.1016/j.toxlet.2011.03.029
© 2011 Elsevier Ireland Ltd. All rights reserved.
reduce cortical thymocytes leading to reduced thymus weight andsubsequently suppress T cell mediated immune responses (Seinenand Willems, 1976; Seinen et al., 1977). Non-specific immunefunctions like natural killer (NK) cells (Aluoch et al., 2007) andmacrophage activity (Vos et al., 1984; Kergosien and Rice, 1998)are also affected. The possible mechanism involved in tributyltintoxicity in thymocytes is apoptosis via oxidative damage, involv-ing a substantial increase of intracellular calcium (Chow et al.,1992; Liu et al., 2006; Nakatsu et al., 2007). Several in vitro experi-ments show that organotins induced toxicity involve activation ofMAP kinase signaling pathways (Mundy and Freudenrich, 2006; Leeet al., 2010). A recent study demonstrated cell death by autophagyin neurons by tributyltin (Nakatsu et al., 2010).
Considering the alarming usage of tributyltin and subsequenthuman dietary exposure to this compound, effective strategies areneeded to reduce the threat by TBT. A few studies are availabledepicting the role of natural compounds in mitigating the toxiceffect caused by tributyltin both in vitro and in vivo. Liu and Xu(2007) used garlic oil to prevent tributyltin induced oxidative dam-age in vitro and in vivo and in another similar study by Liu et al.(2008), green tea polyphenol was taken to protect rat model against
tributyltin induced oxidative damage.Spirulina platensis, a blue-green alga, has emerged as a promis-ing and highly nutritious food source during recent years as itcontains high quantity and quality proteins as well as carotenoids,

M. Gupta et al. / Toxicology L
Fig. 1. Structure of C-Phycocyanin.
vt(
b(ah
acgIee((Fbeme
ib2siipomof2tci
with a 12 h light/dark cycle at 25 ± 2 ◦C and fed with standardrodent pellet and water ad libitum. IITR animal house and breeding
TE
E
itamins, and minerals. It has been commercialized in several coun-ries as health food supplements and for therapeutic purposesMani et al., 2007; Ravi et al., 2010).
Spirulina’s antioxidant potential is attributed to phycocyanin,-carotene, tocopherol, g-linolenic acid and phenolic compoundsRavi et al., 2010). Its manifold applications include: anticancer,ntibacterial, and antiparasitic and also in allergies, ulcers, anaemia,eavy metal poisoning and radiation poisoning.
C-Phycocyanin is composed of two moieties: phycobilisomend its apoprotein counterpart as depicted in Fig. 1. Phycocyaninontaining Se has been shown to have strong superoxide and hydro-en peroxide radical scavenging activities (Huang et al., 2007).n oxidative stress related studies, Spirulina has been shown toxhibit protective efficacy (Lu et al., 2006; Ponce-Canchihuamant al., 2010). Spirulina platensis reduces Cd induced hepatoxicityKaradeniz et al., 2009) and gentamicin induced nephrotoxicityKaradeniz et al., 2008) possibly by its antioxidant properties.urthermore, Spirulina platensis lowers DMBA-induced hamsteruccal pouch carcinogenesis (Grawish et al., 2010) and selenium –nriched Spirulina induces G1 cell cycle arrest and mitochondria-ediated apoptosis in MCF-7 human breast carcinoma cells (Chen
t al., 2009).One of the most widely studied effects of C-Phycocyanin (C-Pc)
s its antioxidant capacity and its free radical scavenging ability,oth in vitro and in vivo (Romay et al., 1998; Bhat and Madyastha,000). Its inhibitory effect was also observed on hepatic micro-omal lipid peroxidation (Halliwell, 1990). The anti-inflammatory,mmunomodulatory, antiproliferative effects and apoptosis induc-ng ability in tumor cells of C-Pc are mainly mediated by itsotent antioxidant property. This can explain the anticancer effectf C-Pc, since most neoplastic processes have a chronic inflam-atory component and other elements that express a marked
xidative stress. Also, C-Pc is a selective inhibitor of COX-2, thusavoring programmed cell death of tumor cells (Reddy et al.,000). A study carried out by Subhashini et al. (2004), showedhat C-Pc induces apoptosis in K-562 cell line by Cytochrome-
release from mitochondria to cytosol and causing a decreasen Bcl-2.
able 1ffect of C-Pc on TBTC altered body weight, thymus weight, cellularity and cell viability.
Parameters Control
Body wt. changed +2.0 ± 0.4Relative thymus wt. (mg/g b. wt.) 3.2 ± 0.5Thymic cellularity (×106) 90.01 ± 18.2Cell viability (% of control) 100 ± 15.2
ach value represents mean ± S.D. (n = 6).*** p < 0.001 as compared to control, using one-way ANOVA.** p < 0.01 as compared to control, using one-way ANOVA.* p < 0.05 as compared to control, using one-way ANOVA.a p < 0.001 as compared to TBTC group, using one-way ANOVA.b p < 0.01 as compared to TBTC group, using one-way ANOVA.c p < 0.05 as compared to TBTC group, using one-way ANOVA.d Difference between terminal and predisposing body wt.
etters 204 (2011) 2–11 3
Considering the traditional therapeutic usage of Spirulina andthe potential effectiveness of its biliprotein C-Phycocyanin, thepresent study was aimed to determine if C-Phycocyanin could pos-sibly prevent the rats from thymic atrophy induced by tributyltinchloride (TBTC).
This study was so designed to include various cellular deter-minants to explore the cytoprotective efficacy of C-Pc. Theanti-oxidative and anti-apoptotic efficacy of C-Pc was determinedby mapping of oxidative stress (ROS and GSH) and apoptotichallmarks (mitochondrial depolarization, caspase-3 activity, phos-phatidylserine externalization, hypodiploid population, Bax/Bcl-2ratio). The thymic histology was also undertaken to decifer linkbetween cell death and lymphoid morphology, if any. Addition-ally, the uptake of C-Pc by thymocytes was monitored to know itsbioavailability.
This study clearly demonstrates effective mitigation by C-Pc andantioxidant and free radical scavenging nature of the biliproteincould mainly contribute to its ameliorative efficacy in TBTC inducedthymic atrophy.
2. Materials and methods
2.1. Chemicals
All the chemicals were of highest grade purity available.Acrylamide, antibiotic–antimycotic solution, C-Phycocyanin,2′,7′-dichlorofluorescein diacetate (DCFH-DA), Dulbecco’sphosphate buffered saline (PBS), Fetal Bovine Serum (FBS),2-Mercaptoethanol, N,N′-methylene bis-acrylamide, N,N,N′,N′-tetramethyl ethylenediamine (TEMED), Propidium Iodide (PI),Rhodamine 123 (Rh123), RNase A, RPMI 1640, Sodium DodecylSulphate, Trypan Blue and all other chemicals were purchased fromSigma–Aldrich. 5′-chloromethylfluorescein diacetate (CMF-DA)were purchased from Molecular Probes and Annexin V-FITC fromBD Pharmingen (Becton Dickinson Company). Alamar Blue waspurchased from Invitrogen and tributyltin chloride was purchasedfrom Merck. HRP-conjugated anti-mouse IgG, HRP-conjugatedanti-rabbit IgG were from Sigma and anti-Bax, anti-Bcl-2 antibodieswere from Biovision.
2.2. Experimental layout
Male Wistar rats were maintained in IITR animal house understandard conditions. Four to six weeks old male rats were used forin vivo studies. Rats were housed in plastic polypropylene cages
facility are registered with Committee for the Purpose of Controland Supervision of Experiments on Animals (CPCSEA), Govern-
TBTC TBTC + C-Pc C-Pc
−1.0 ± 0.2 +1.0 ± 0.4 +2.4 ± 0.71.7 ± 0.4*** 2.6 ± 0.3*,b 3.1 ± 0.9a
43.80 ± 10.2*** 78.99 ± 12.8**,a 91.29 ± 21.9a
71.1 ± 11.9** 89.8 ± 9.9*,c 109.2 ± 7.1a

4 logy Le
mo
2
Oooib
vbm1as
2
pacts1b
2
ha
2
aor
2
inemaE
M. Gupta et al. / Toxico
ent of India and CPCSEA guidelines were followed (IAEC approvalbtained).
.3. Treatment schedule of TBTC
The rats were divided in four groups of six animals each.ut of which, one group was administered corn oil (0.1 ml/50 g,rally) for 48 h and rest of the groups received TBTC (50 mg/kg,rally, in corn oil) for 48 h and/or C-Phycocyanin (70 mg/kg, i.p.,n dH2O) was given 3 h prior to TBTC. The grouping chart is aselow:
Group I (Control)
Group II (TBTC alone)
Group III (TBTC+C-Pc)
Group IV (C-Pc alone)
Corn oil - 48h
TBTC - 48h
Corn oil-48h
TBTC - 48h
C-Pc - 3h
C-Pc - 3h
TBTC dose selected for our investigation was based on the pre-ious study of Raffray and Cohen (1993). Rats were weighed ineginning of the experiment and at time of sacrifice (48 h). Thy-uses were excised and a small portion was immediately put in
0% neutral buffered formalin for histopathological examinationnd remaining portion was used for biochemical estimations theame day.
.4. Preparation of single cell suspension
Thymocyte single cell suspension was prepared aseptically byassing through 100 �m nylon mesh. It was then centrifugedt 200 × g at 4 ◦C for 10 min and resuspended in complete cellulture medium (RPMI 1640 containing HEPES and 2 mM glu-amine, supplemented with 10% FBS and 1% antibiotic–antimycoticolution). For biochemical assays, cell density was adjusted to.5 × 106 cells/ml and viability of the freshly isolated cells testedy trypan blue exclusion method was > 95%.
.5. Assessment of cellularity
Total number of thymocytes per animal were counted usingaemocytometer on an inverted light microscope (Leica DM IL),fter preparing single cell suspension.
.6. Assessment of cell viability
Briefly, cells (10,000 cells/well) were seeded in 96 well platend Alamar Blue solution (10 �l) was added and incubated at 37 ◦Cvernight. Absorbance was measured at 570 nm in a microplateeader (FluoStar Omega).
.7. Histopathological examination
Collected thymic tissues were processed for histological exam-nation. At the time of sacrifice, the tissues were fixed in 10%eutral buffered formalin for 24 h. Then, they were dehydrated with
thanol and embedded in paraffin wax using standard histologicalethods. Five mm sections were cut and stained with hematoxylinnd eosin and examined under light microscope (Nikon Eclipse600).
tters 204 (2011) 2–11
2.8. ROS levels
ROS generation was detected by DCF fluorescence by using 2,7-dichlorofluorescein diacetate (DCFH-DA). Thymocytes were thenloaded with DCFH-DA (100 �M final conc.) for 30 min at 37 ◦C in CO2incubator. After washing twice with PBS, the cells were suspendedin PBS and DCF fluorescence was monitored in a microplate readerat Ex: 488 nm and Em: 510 nm.
2.9. C-Phycocyanin uptake in primary thymocytes
Thymus was dissected from male Wistar rat (3–4 weeks old)and single cell suspension of thymocytes was prepared as men-tioned earlier. Thymocytes in RPMI medium were exposed toC-Phycocyanin (1.0 �g/ml) with (out) TBTC (0.3 �M) and incubatedat 37 ◦C in CO2 incubator for indicated time points. Cells were thenharvested and resuspended in PBS. C-Pc fluorescence was moni-tored in a microplate reader at Ex: 584 nm and Em: 640 nm.
2.10. Bcl-2 and Bax protein levels
For western blotting, thymocytes were harvested to obtain cellpellet. Cells were lysed by lysis buffer (Tris 0.125 M, pH 6.8, SDS3%, Glycerol 20%, Bromophenol blue 0.1% and 2-mercaptoethanol5%). Cell lysate was separated on 12% SDS-PAGE and electrophoret-ically transferred on to a nitrocellulose membrane. Membraneswere blocked in solution containing 5% non-fat dry milk inPBST buffer (PBS buffer containing 0.1% Tween-20) for 4 h atroom temperature, probed with anti-Bax and anti-Bcl-2 antibodies(1:2000) in PBST buffer overnight at 4 ◦C, followed by horseradishperoxidase-labeled anti-mouse secondary antibody (1:7000) for2 h and visualised on gel doc system (Versadoc) by enhancedchemiluminescent detection method according to manufacturer’sprotocol.
2.11. Apoptotic assays by flow cytometry
All the apoptotic assays were performed on a BD-LSR cytometer.For each sample, data from 10,000 cells were recorded in list modeon logarithmic scales. Analysis was performed with Cell Quest soft-ware (BDIS) on the cells characterized by their forward/side scatter(FSC/SSC) parameters. Cell debris with a low FSC/SSC was excludedfrom the analysis.
2.11.1. Apoptotic and necrotic population by Annexin V and PIuptake
Positioning of quadrants on Annexin V/PI dot plots was per-formed and living cells (Annexin V−/PI−), early apoptotic/primaryapoptotic cells (Annexin V+/PI−), late apoptotic/secondary apo-ptotic cells (Annexin V+/PI+) and necrotic cells (Annexin V−/PI+)were distinguished. The total apoptotic population, therefore,included cells with AnnexinV+/PI− and Annexin V+/PI+ fluores-cence.
Briefly, cells were suspended in binding buffer (1×) and analiquot of 100 �l was incubated with 5 �l Annexin V-FITC and 10 �lPI for 15 min in dark at RT and 400 �l PBS was then added to eachsample. The FITC and PI fluorescence were measured through FL-1 filter (530 nm) and FL-2 filter (585 nm) respectively, and 10,000events were acquired.
2.11.2. Hypodiploid population by cell cycle studyThe cells with hypodiploid DNA (Sub-G1) were determined by
cell cycle studies. Cells were washed with PBS, fixed in ice cold 70%ethanol and kept overnight at 4 ◦C. Cells were harvested, washedand suspended in 1 ml PBS. Phosphate citrate buffer (200 �l, pH7.8) was added and the cells were incubated for 60 min at RT. After
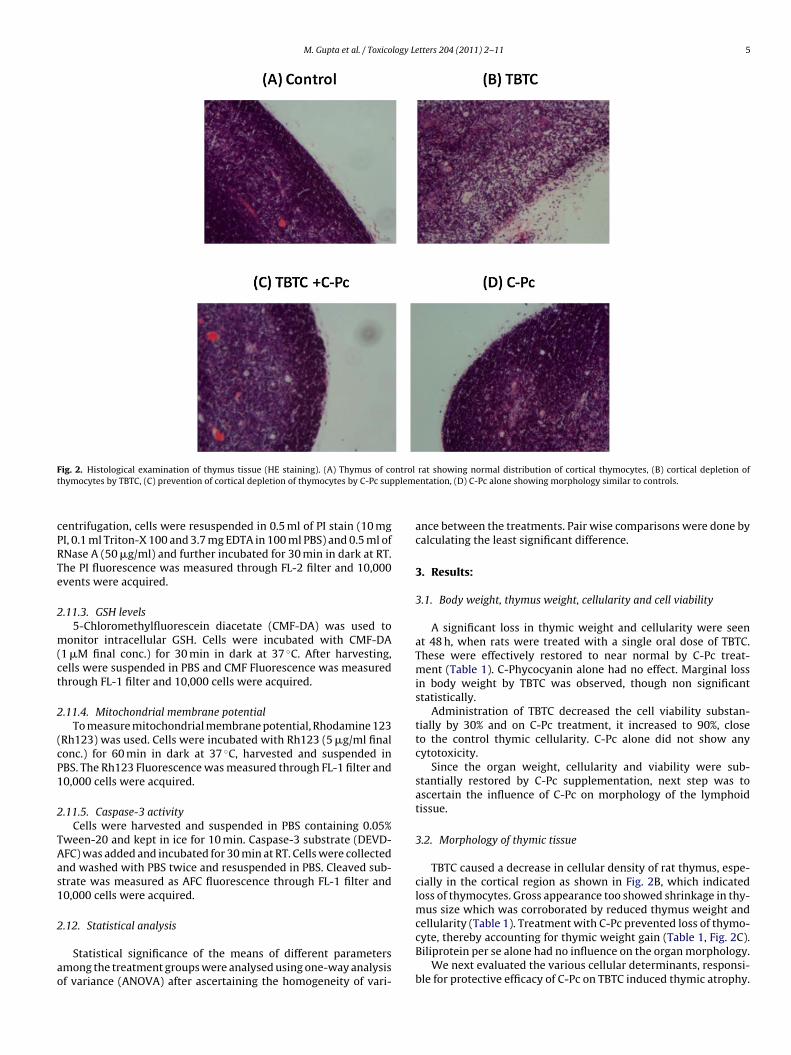
M. Gupta et al. / Toxicology Letters 204 (2011) 2–11 5
F ntrolt plem
cPRTe
2
m(ct
2
(cP1
2
TAas1
2
ao
ig. 2. Histological examination of thymus tissue (HE staining). (A) Thymus of cohymocytes by TBTC, (C) prevention of cortical depletion of thymocytes by C-Pc sup
entrifugation, cells were resuspended in 0.5 ml of PI stain (10 mgI, 0.1 ml Triton-X 100 and 3.7 mg EDTA in 100 ml PBS) and 0.5 ml ofNase A (50 �g/ml) and further incubated for 30 min in dark at RT.he PI fluorescence was measured through FL-2 filter and 10,000vents were acquired.
.11.3. GSH levels5-Chloromethylfluorescein diacetate (CMF-DA) was used to
onitor intracellular GSH. Cells were incubated with CMF-DA1 �M final conc.) for 30 min in dark at 37 ◦C. After harvesting,ells were suspended in PBS and CMF Fluorescence was measuredhrough FL-1 filter and 10,000 cells were acquired.
.11.4. Mitochondrial membrane potentialTo measure mitochondrial membrane potential, Rhodamine 123
Rh123) was used. Cells were incubated with Rh123 (5 �g/ml finalonc.) for 60 min in dark at 37 ◦C, harvested and suspended inBS. The Rh123 Fluorescence was measured through FL-1 filter and0,000 cells were acquired.
.11.5. Caspase-3 activityCells were harvested and suspended in PBS containing 0.05%
ween-20 and kept in ice for 10 min. Caspase-3 substrate (DEVD-FC) was added and incubated for 30 min at RT. Cells were collectednd washed with PBS twice and resuspended in PBS. Cleaved sub-trate was measured as AFC fluorescence through FL-1 filter and0,000 cells were acquired.
.12. Statistical analysis
Statistical significance of the means of different parametersmong the treatment groups were analysed using one-way analysisf variance (ANOVA) after ascertaining the homogeneity of vari-
rat showing normal distribution of cortical thymocytes, (B) cortical depletion ofentation, (D) C-Pc alone showing morphology similar to controls.
ance between the treatments. Pair wise comparisons were done bycalculating the least significant difference.
3. Results:
3.1. Body weight, thymus weight, cellularity and cell viability
A significant loss in thymic weight and cellularity were seenat 48 h, when rats were treated with a single oral dose of TBTC.These were effectively restored to near normal by C-Pc treat-ment (Table 1). C-Phycocyanin alone had no effect. Marginal lossin body weight by TBTC was observed, though non significantstatistically.
Administration of TBTC decreased the cell viability substan-tially by 30% and on C-Pc treatment, it increased to 90%, closeto the control thymic cellularity. C-Pc alone did not show anycytotoxicity.
Since the organ weight, cellularity and viability were sub-stantially restored by C-Pc supplementation, next step was toascertain the influence of C-Pc on morphology of the lymphoidtissue.
3.2. Morphology of thymic tissue
TBTC caused a decrease in cellular density of rat thymus, espe-cially in the cortical region as shown in Fig. 2B, which indicatedloss of thymocytes. Gross appearance too showed shrinkage in thy-mus size which was corroborated by reduced thymus weight andcellularity (Table 1). Treatment with C-Pc prevented loss of thymo-
cyte, thereby accounting for thymic weight gain (Table 1, Fig. 2C).Biliprotein per se alone had no influence on the organ morphology.We next evaluated the various cellular determinants, responsi-ble for protective efficacy of C-Pc on TBTC induced thymic atrophy.
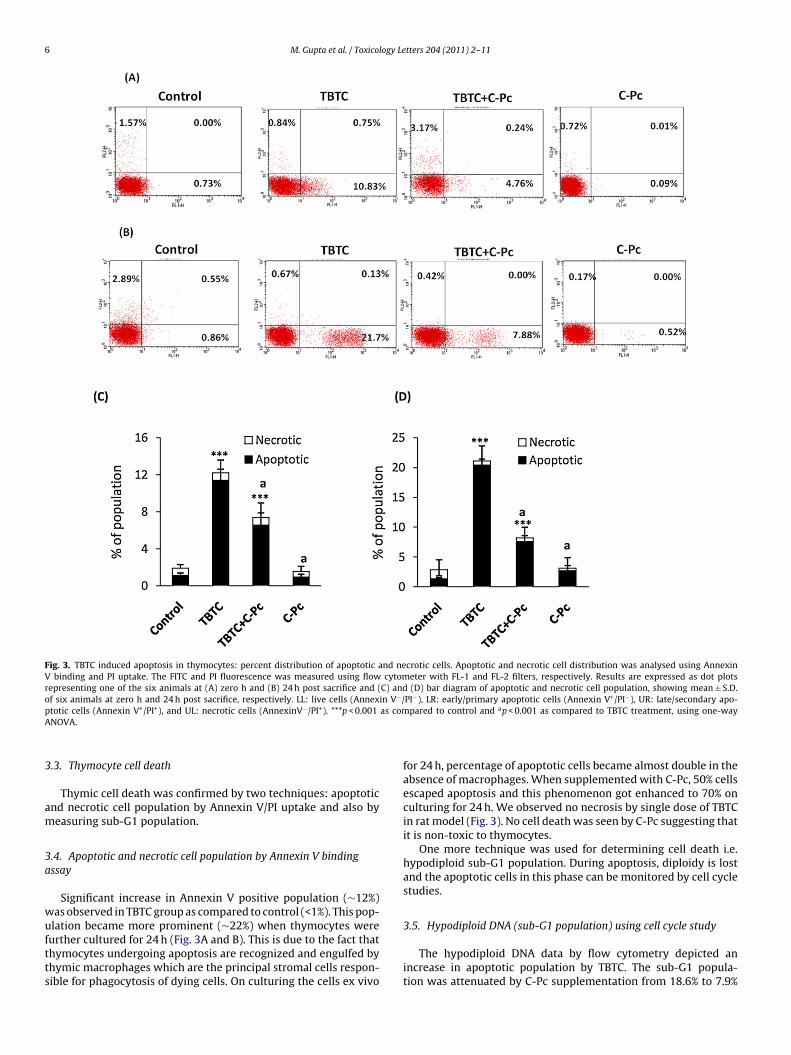
6 M. Gupta et al. / Toxicology Letters 204 (2011) 2–11
Fig. 3. TBTC induced apoptosis in thymocytes: percent distribution of apoptotic and necrotic cells. Apoptotic and necrotic cell distribution was analysed using AnnexinV binding and PI uptake. The FITC and PI fluorescence was measured using flow cytometer with FL-1 and FL-2 filters, respectively. Results are expressed as dot plotsrepresenting one of the six animals at (A) zero h and (B) 24 h post sacrifice and (C) and (D) bar diagram of apoptotic and necrotic cell population, showing mean ± S.D.of six animals at zero h and 24 h post sacrifice, respectively. LL: live cells (Annexin V−/PI−), LR: early/primary apoptotic cells (Annexin V+/PI−), UR: late/secondary apo-p as comA
3
am
3a
wuftts
totic cells (Annexin V+/PI+), and UL: necrotic cells (AnnexinV−/PI+). ***p < 0.001NOVA.
.3. Thymocyte cell death
Thymic cell death was confirmed by two techniques: apoptoticnd necrotic cell population by Annexin V/PI uptake and also byeasuring sub-G1 population.
.4. Apoptotic and necrotic cell population by Annexin V bindingssay
Significant increase in Annexin V positive population (∼12%)as observed in TBTC group as compared to control (<1%). This pop-lation became more prominent (∼22%) when thymocytes were
urther cultured for 24 h (Fig. 3A and B). This is due to the fact thathymocytes undergoing apoptosis are recognized and engulfed byhymic macrophages which are the principal stromal cells respon-ible for phagocytosis of dying cells. On culturing the cells ex vivopared to control and ap < 0.001 as compared to TBTC treatment, using one-way
for 24 h, percentage of apoptotic cells became almost double in theabsence of macrophages. When supplemented with C-Pc, 50% cellsescaped apoptosis and this phenomenon got enhanced to 70% onculturing for 24 h. We observed no necrosis by single dose of TBTCin rat model (Fig. 3). No cell death was seen by C-Pc suggesting thatit is non-toxic to thymocytes.
One more technique was used for determining cell death i.e.hypodiploid sub-G1 population. During apoptosis, diploidy is lostand the apoptotic cells in this phase can be monitored by cell cyclestudies.
3.5. Hypodiploid DNA (sub-G1 population) using cell cycle study
The hypodiploid DNA data by flow cytometry depicted anincrease in apoptotic population by TBTC. The sub-G1 popula-tion was attenuated by C-Pc supplementation from 18.6% to 7.9%
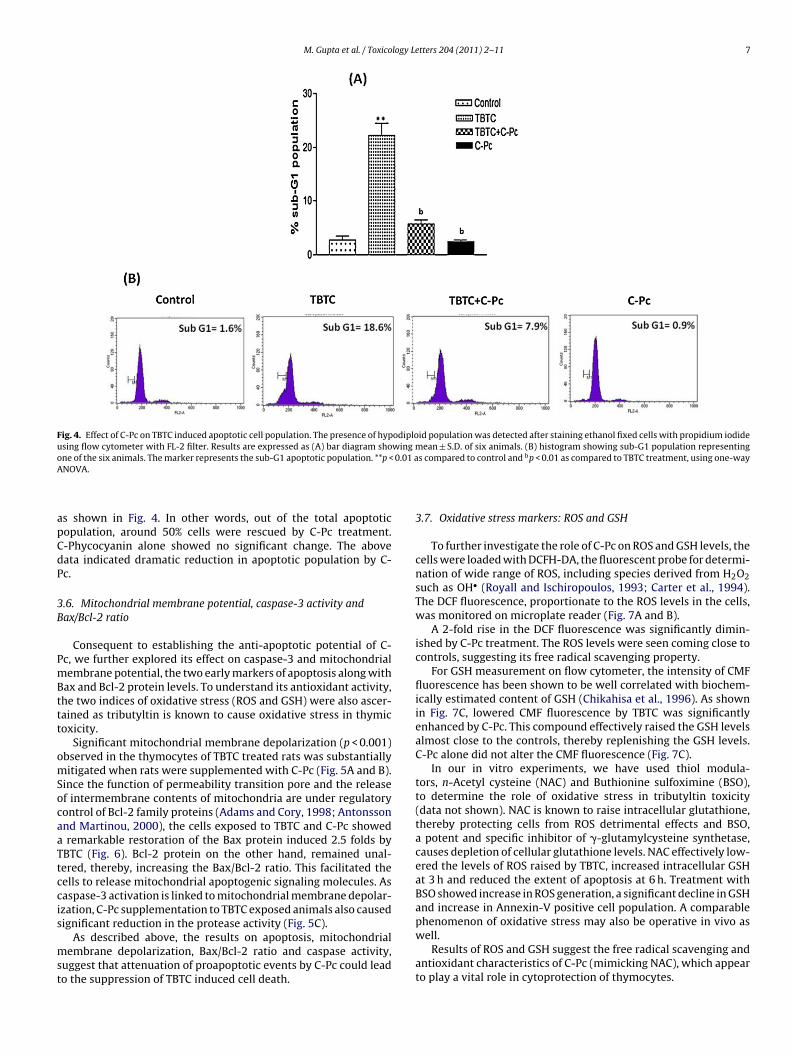
M. Gupta et al. / Toxicology Letters 204 (2011) 2–11 7
Fig. 4. Effect of C-Pc on TBTC induced apoptotic cell population. The presence of hypodiploid population was detected after staining ethanol fixed cells with propidium iodideusing flow cytometer with FL-2 filter. Results are expressed as (A) bar diagram showing mean ± S.D. of six animals. (B) histogram showing sub-G1 population representingone of the six animals. The marker represents the sub-G1 apoptotic population. **p < 0.01 as compared to control and bp < 0.01 as compared to TBTC treatment, using one-wayA
apCdP
3B
PmBttt
omSocaaTtccis
mst
NOVA.
s shown in Fig. 4. In other words, out of the total apoptoticopulation, around 50% cells were rescued by C-Pc treatment.-Phycocyanin alone showed no significant change. The aboveata indicated dramatic reduction in apoptotic population by C-c.
.6. Mitochondrial membrane potential, caspase-3 activity andax/Bcl-2 ratio
Consequent to establishing the anti-apoptotic potential of C-c, we further explored its effect on caspase-3 and mitochondrialembrane potential, the two early markers of apoptosis along with
ax and Bcl-2 protein levels. To understand its antioxidant activity,he two indices of oxidative stress (ROS and GSH) were also ascer-ained as tributyltin is known to cause oxidative stress in thymicoxicity.
Significant mitochondrial membrane depolarization (p < 0.001)bserved in the thymocytes of TBTC treated rats was substantiallyitigated when rats were supplemented with C-Pc (Fig. 5A and B).
ince the function of permeability transition pore and the releasef intermembrane contents of mitochondria are under regulatoryontrol of Bcl-2 family proteins (Adams and Cory, 1998; Antonssonnd Martinou, 2000), the cells exposed to TBTC and C-Pc showedremarkable restoration of the Bax protein induced 2.5 folds by
BTC (Fig. 6). Bcl-2 protein on the other hand, remained unal-ered, thereby, increasing the Bax/Bcl-2 ratio. This facilitated theells to release mitochondrial apoptogenic signaling molecules. Asaspase-3 activation is linked to mitochondrial membrane depolar-zation, C-Pc supplementation to TBTC exposed animals also causedignificant reduction in the protease activity (Fig. 5C).
As described above, the results on apoptosis, mitochondrialembrane depolarization, Bax/Bcl-2 ratio and caspase activity,
uggest that attenuation of proapoptotic events by C-Pc could leado the suppression of TBTC induced cell death.
3.7. Oxidative stress markers: ROS and GSH
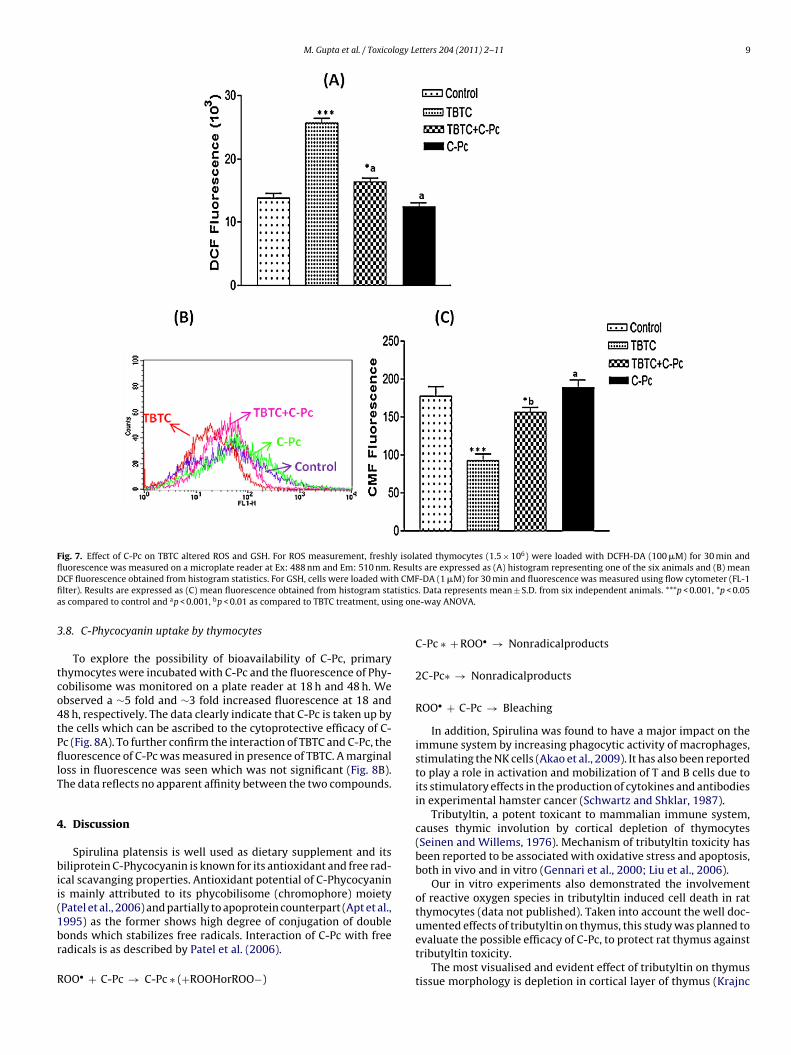
To further investigate the role of C-Pc on ROS and GSH levels, thecells were loaded with DCFH-DA, the fluorescent probe for determi-nation of wide range of ROS, including species derived from H2O2such as OH• (Royall and Ischiropoulos, 1993; Carter et al., 1994).The DCF fluorescence, proportionate to the ROS levels in the cells,was monitored on microplate reader (Fig. 7A and B).
A 2-fold rise in the DCF fluorescence was significantly dimin-ished by C-Pc treatment. The ROS levels were seen coming close tocontrols, suggesting its free radical scavenging property.
For GSH measurement on flow cytometer, the intensity of CMFfluorescence has been shown to be well correlated with biochem-ically estimated content of GSH (Chikahisa et al., 1996). As shownin Fig. 7C, lowered CMF fluorescence by TBTC was significantlyenhanced by C-Pc. This compound effectively raised the GSH levelsalmost close to the controls, thereby replenishing the GSH levels.C-Pc alone did not alter the CMF fluorescence (Fig. 7C).
In our in vitro experiments, we have used thiol modula-tors, n-Acetyl cysteine (NAC) and Buthionine sulfoximine (BSO),to determine the role of oxidative stress in tributyltin toxicity(data not shown). NAC is known to raise intracellular glutathione,thereby protecting cells from ROS detrimental effects and BSO,a potent and specific inhibitor of �-glutamylcysteine synthetase,causes depletion of cellular glutathione levels. NAC effectively low-ered the levels of ROS raised by TBTC, increased intracellular GSHat 3 h and reduced the extent of apoptosis at 6 h. Treatment withBSO showed increase in ROS generation, a significant decline in GSHand increase in Annexin-V positive cell population. A comparablephenomenon of oxidative stress may also be operative in vivo as
well.Results of ROS and GSH suggest the free radical scavenging andantioxidant characteristics of C-Pc (mimicking NAC), which appearto play a vital role in cytoprotection of thymocytes.
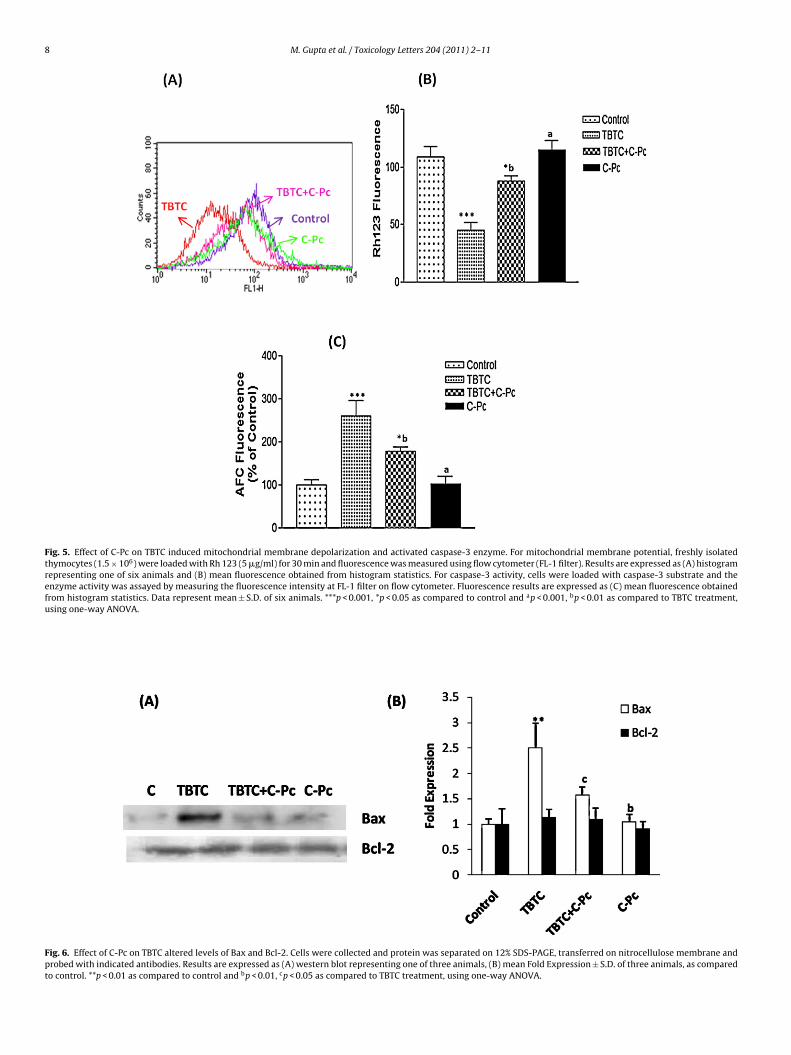
8 M. Gupta et al. / Toxicology Letters 204 (2011) 2–11
Fig. 5. Effect of C-Pc on TBTC induced mitochondrial membrane depolarization and activated caspase-3 enzyme. For mitochondrial membrane potential, freshly isolatedthymocytes (1.5 × 106) were loaded with Rh 123 (5 �g/ml) for 30 min and fluorescence was measured using flow cytometer (FL-1 filter). Results are expressed as (A) histogramrepresenting one of six animals and (B) mean fluorescence obtained from histogram statistics. For caspase-3 activity, cells were loaded with caspase-3 substrate and theenzyme activity was assayed by measuring the fluorescence intensity at FL-1 filter on flow cytometer. Fluorescence results are expressed as (C) mean fluorescence obtainedfrom histogram statistics. Data represent mean ± S.D. of six animals. ***p < 0.001, *p < 0.05 as compared to control and ap < 0.001, bp < 0.01 as compared to TBTC treatment,using one-way ANOVA.
Fig. 6. Effect of C-Pc on TBTC altered levels of Bax and Bcl-2. Cells were collected and protein was separated on 12% SDS-PAGE, transferred on nitrocellulose membrane andprobed with indicated antibodies. Results are expressed as (A) western blot representing one of three animals, (B) mean Fold Expression ± S.D. of three animals, as comparedto control. **p < 0.01 as compared to control and bp < 0.01, cp < 0.05 as compared to TBTC treatment, using one-way ANOVA.
M. Gupta et al. / Toxicology Letters 204 (2011) 2–11 9
Fig. 7. Effect of C-Pc on TBTC altered ROS and GSH. For ROS measurement, freshly isolated thymocytes (1.5 × 106) were loaded with DCFH-DA (100 �M) for 30 min andfluorescence was measured on a microplate reader at Ex: 488 nm and Em: 510 nm. Results are expressed as (A) histogram representing one of the six animals and (B) meanD h CMFfi tisticsa ng on
3
tco4tPfllT
4
bii(1br
R
CF fluorescence obtained from histogram statistics. For GSH, cells were loaded witlter). Results are expressed as (C) mean fluorescence obtained from histogram stas compared to control and ap < 0.001, bp < 0.01 as compared to TBTC treatment, usi
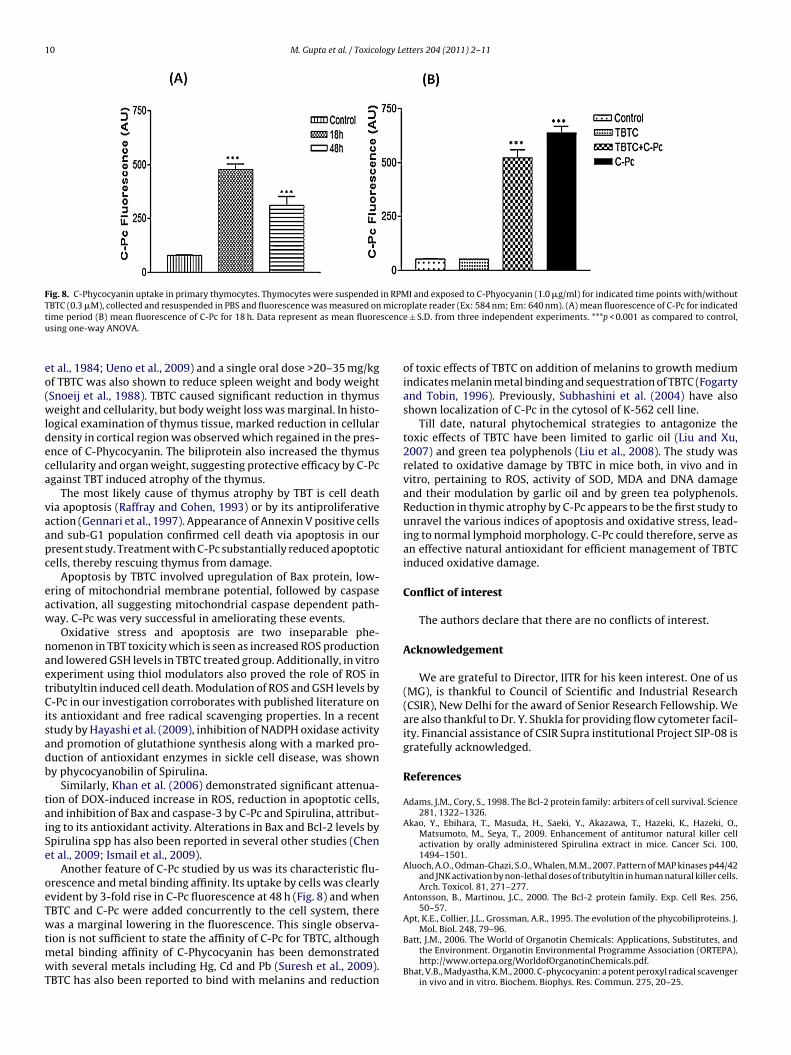
.8. C-Phycocyanin uptake by thymocytes
To explore the possibility of bioavailability of C-Pc, primaryhymocytes were incubated with C-Pc and the fluorescence of Phy-obilisome was monitored on a plate reader at 18 h and 48 h. Webserved a ∼5 fold and ∼3 fold increased fluorescence at 18 and8 h, respectively. The data clearly indicate that C-Pc is taken up byhe cells which can be ascribed to the cytoprotective efficacy of C-c (Fig. 8A). To further confirm the interaction of TBTC and C-Pc, theuorescence of C-Pc was measured in presence of TBTC. A marginal
oss in fluorescence was seen which was not significant (Fig. 8B).he data reflects no apparent affinity between the two compounds.
. Discussion
Spirulina platensis is well used as dietary supplement and itsiliprotein C-Phycocyanin is known for its antioxidant and free rad-
cal scavanging properties. Antioxidant potential of C-Phycocyanins mainly attributed to its phycobilisome (chromophore) moietyPatel et al., 2006) and partially to apoprotein counterpart (Apt et al.,995) as the former shows high degree of conjugation of double
onds which stabilizes free radicals. Interaction of C-Pc with freeadicals is as described by Patel et al. (2006).OO• + C-Pc → C-Pc ∗ (+ROOHorROO−)
-DA (1 �M) for 30 min and fluorescence was measured using flow cytometer (FL-1. Data represents mean ± S.D. from six independent animals. ***p < 0.001, *p < 0.05
e-way ANOVA.
C-Pc ∗ + ROO• → Nonradicalproducts
2C-Pc∗ → Nonradicalproducts
ROO• + C-Pc → Bleaching
In addition, Spirulina was found to have a major impact on theimmune system by increasing phagocytic activity of macrophages,stimulating the NK cells (Akao et al., 2009). It has also been reportedto play a role in activation and mobilization of T and B cells due toits stimulatory effects in the production of cytokines and antibodiesin experimental hamster cancer (Schwartz and Shklar, 1987).
Tributyltin, a potent toxicant to mammalian immune system,causes thymic involution by cortical depletion of thymocytes(Seinen and Willems, 1976). Mechanism of tributyltin toxicity hasbeen reported to be associated with oxidative stress and apoptosis,both in vivo and in vitro (Gennari et al., 2000; Liu et al., 2006).
Our in vitro experiments also demonstrated the involvementof reactive oxygen species in tributyltin induced cell death in ratthymocytes (data not published). Taken into account the well doc-umented effects of tributyltin on thymus, this study was planned to
evaluate the possible efficacy of C-Pc, to protect rat thymus againsttributyltin toxicity.The most visualised and evident effect of tributyltin on thymustissue morphology is depletion in cortical layer of thymus (Krajnc
10 M. Gupta et al. / Toxicology Letters 204 (2011) 2–11
Fig. 8. C-Phycocyanin uptake in primary thymocytes. Thymocytes were suspended in RPMI and exposed to C-Phyocyanin (1.0 �g/ml) for indicated time points with/withoutTBTC (0.3 �M), collected and resuspended in PBS and fluorescence was measured on microplate reader (Ex: 584 nm; Em: 640 nm). (A) mean fluorescence of C-Pc for indicatedt scencu
eo(wldeca
vaapc
eaw
naetCisadb
taiSe
oeTwtmwT
ime period (B) mean fluorescence of C-Pc for 18 h. Data represent as mean fluoresing one-way ANOVA.
t al., 1984; Ueno et al., 2009) and a single oral dose >20–35 mg/kgf TBTC was also shown to reduce spleen weight and body weightSnoeij et al., 1988). TBTC caused significant reduction in thymuseight and cellularity, but body weight loss was marginal. In histo-
ogical examination of thymus tissue, marked reduction in cellularensity in cortical region was observed which regained in the pres-nce of C-Phycocyanin. The biliprotein also increased the thymusellularity and organ weight, suggesting protective efficacy by C-Pcgainst TBT induced atrophy of the thymus.
The most likely cause of thymus atrophy by TBT is cell deathia apoptosis (Raffray and Cohen, 1993) or by its antiproliferativection (Gennari et al., 1997). Appearance of Annexin V positive cellsnd sub-G1 population confirmed cell death via apoptosis in ourresent study. Treatment with C-Pc substantially reduced apoptoticells, thereby rescuing thymus from damage.
Apoptosis by TBTC involved upregulation of Bax protein, low-ring of mitochondrial membrane potential, followed by caspasectivation, all suggesting mitochondrial caspase dependent path-ay. C-Pc was very successful in ameliorating these events.
Oxidative stress and apoptosis are two inseparable phe-omenon in TBT toxicity which is seen as increased ROS productionnd lowered GSH levels in TBTC treated group. Additionally, in vitroxperiment using thiol modulators also proved the role of ROS inributyltin induced cell death. Modulation of ROS and GSH levels by-Pc in our investigation corroborates with published literature on
ts antioxidant and free radical scavenging properties. In a recenttudy by Hayashi et al. (2009), inhibition of NADPH oxidase activitynd promotion of glutathione synthesis along with a marked pro-uction of antioxidant enzymes in sickle cell disease, was showny phycocyanobilin of Spirulina.
Similarly, Khan et al. (2006) demonstrated significant attenua-ion of DOX-induced increase in ROS, reduction in apoptotic cells,nd inhibition of Bax and caspase-3 by C-Pc and Spirulina, attribut-ng to its antioxidant activity. Alterations in Bax and Bcl-2 levels bypirulina spp has also been reported in several other studies (Chent al., 2009; Ismail et al., 2009).
Another feature of C-Pc studied by us was its characteristic flu-rescence and metal binding affinity. Its uptake by cells was clearlyvident by 3-fold rise in C-Pc fluorescence at 48 h (Fig. 8) and whenBTC and C-Pc were added concurrently to the cell system, thereas a marginal lowering in the fluorescence. This single observa-
ion is not sufficient to state the affinity of C-Pc for TBTC, althoughetal binding affinity of C-Phycocyanin has been demonstratedith several metals including Hg, Cd and Pb (Suresh et al., 2009).
BTC has also been reported to bind with melanins and reduction
e ± S.D. from three independent experiments. ***p < 0.001 as compared to control,
of toxic effects of TBTC on addition of melanins to growth mediumindicates melanin metal binding and sequestration of TBTC (Fogartyand Tobin, 1996). Previously, Subhashini et al. (2004) have alsoshown localization of C-Pc in the cytosol of K-562 cell line.
Till date, natural phytochemical strategies to antagonize thetoxic effects of TBTC have been limited to garlic oil (Liu and Xu,2007) and green tea polyphenols (Liu et al., 2008). The study wasrelated to oxidative damage by TBTC in mice both, in vivo and invitro, pertaining to ROS, activity of SOD, MDA and DNA damageand their modulation by garlic oil and by green tea polyphenols.Reduction in thymic atrophy by C-Pc appears to be the first study tounravel the various indices of apoptosis and oxidative stress, lead-ing to normal lymphoid morphology. C-Pc could therefore, serve asan effective natural antioxidant for efficient management of TBTCinduced oxidative damage.
Conflict of interest
The authors declare that there are no conflicts of interest.
Acknowledgement
We are grateful to Director, IITR for his keen interest. One of us(MG), is thankful to Council of Scientific and Industrial Research(CSIR), New Delhi for the award of Senior Research Fellowship. Weare also thankful to Dr. Y. Shukla for providing flow cytometer facil-ity. Financial assistance of CSIR Supra institutional Project SIP-08 isgratefully acknowledged.
References
Adams, J.M., Cory, S., 1998. The Bcl-2 protein family: arbiters of cell survival. Science281, 1322–1326.
Akao, Y., Ebihara, T., Masuda, H., Saeki, Y., Akazawa, T., Hazeki, K., Hazeki, O.,Matsumoto, M., Seya, T., 2009. Enhancement of antitumor natural killer cellactivation by orally administered Spirulina extract in mice. Cancer Sci. 100,1494–1501.
Aluoch, A.O., Odman-Ghazi, S.O., Whalen, M.M., 2007. Pattern of MAP kinases p44/42and JNK activation by non-lethal doses of tributyltin in human natural killer cells.Arch. Toxicol. 81, 271–277.
Antonsson, B., Martinou, J.C., 2000. The Bcl-2 protein family. Exp. Cell Res. 256,50–57.
Apt, K.E., Collier, J.L., Grossman, A.R., 1995. The evolution of the phycobiliproteins. J.Mol. Biol. 248, 79–96.
Batt, J.M., 2006. The World of Organotin Chemicals: Applications, Substitutes, andthe Environment. Organotin Environmental Programme Association (ORTEPA),http://www.ortepa.org/WorldofOrganotinChemicals.pdf.
Bhat, V.B., Madyastha, K.M., 2000. C-phycocyanin: a potent peroxyl radical scavengerin vivo and in vitro. Biochem. Biophys. Res. Commun. 275, 20–25.

logy L
B
C
C
C
C
C
E
F
G
G
G
H
H
H
I
K
K
K
K
K
K
L
L
M. Gupta et al. / Toxico
oyer, I.J., 1989. Toxicity of dibutyltin, tributyltin and other organotin compoundsto humans and to experimental animals. Toxicology 55, 253–298.
arter, W.O., Narayanan, P.K., Robinson, J.P., 1994. Intracellular hydrogen perox-ide and superoxide anion detection in endothelial cells. J. Leukoc. Biol. 55,253–258.
hen, T., Wong, Y.S., Zheng, W., 2009. Induction of G1 cell cycle arrest andmitochondria-mediated apoptosis in MCF-7 human breast carcinoma cells byselenium-enriched Spirulina extract. Biomed. Pharmacother. 27, Epub ahead ofprint 2009.
hikahisa, L., Oyama, Y., Okazaki, E., Noda, K., 1996. Fluorescent estimation of H2O2-induced changes in cell viability and cellular nonprotein thiol level of dissociatedrat thymocytes. Jpn. J. Pharmacol. 71, 299–305.
hoi, M., Moon, H., Yu, J., Eom, J., Choi, H., 2009. Butyltin contamination in industri-alized Bays associated with intensive marine activities in Korea. Arch. Environ.Contam. Toxicol. 57, 77–85.
how, S.C., Kaas, G.E.N., McCabe, M.J., Orrenius, S., 1992. Tributyltin increasescytosolic free Ca2+ concentration in thymocytes by mobilizing intracellular Ca2+,activating a Ca2+ entry pathway, and inhibiting Ca2+ efflux. Arch. Biochem. Bio-phys. 298, 143–149.
guchi, S., Harino, H., Yamamoto, Y., 2010. Assessment of antifouling biocides con-taminations in Maizuru Bay. Japan. Arch. Environ. Contam. Toxicol. 58, 684–693.
ogarty, R.V., Tobin, J.M., 1996. Fungal melanins and their interactions with metals.Enzyme Microb. Technol. 19, 311–317.
ennari, A., Potters, M., Seinen, W., Pieters, R., 1997. Organotin-induced apoptosisas observed in vitro is not relevant for induction of thymus atrophy at antipro-liferative doses. Toxicol. Appl. Pharmacol. 147, 259–266.
ennari, A., Viviani, B., Galli, C.L., Marinovich, M., Pieters, R., Corsini, E., 2000. Organ-otins induce apoptosis by disturbance of [Ca2+]i and mitochondrial activity,causing oxidative stress and activation of caspases in rat thymocytes. Toxicol.Appl. Pharmacol. 169, 185–190.
rawish, M.E., Zaher, A.R., Gaafar, A.I., Nasif, W.A., 2010. Long-term effect of Spir-ulina platensis extract on DMBA-induced hamster buccal pouch carcinogenesis(immunohistochemical study). Med. Oncol. 27, 20–28.
alliwell, B., 1990. How to characterize a biological antioxidant. Free Radic. Res.Commun. 9, 1–32.
ayashi, O., Katayanagi, Y., Ishii, K., Kato, T., 2009. Flow cytometric analysisof age-related changes in intestine intraepithelial lymphocyte subsets andtheir functional preservation after feeding mice on spirulina. J. Med. Food 12,982–989.
uang, Z., Guo, B.J., Wong, R.N.S., Jiang, Y., 2007. Characterization and antioxidantactivity of selenium-containing phycocyanin isolated from Spirulina platensis.Food Chem. 100, 1137–1143.
smail, M.F., Ali, D.A., Fernando, A., Abdraboh, M.E., Gaur, R.L., Ibrahim, W.M., Raj,M.H.G., Ouhtit, A., 2009. Chemoprevention of rat liver toxicity and carcinogen-esis by Spirulina. Int. J. Biol. Sci. 5, 377–387.
aradeniz, A., Yildirim, A., Simsek, N., Kalkan, Y., Celebi, F., 2008. Spirulina platensisprotects against gentamicin-induced nephrotoxicity in rats. Phytother. Res. 22,1506–1510.
aradeniz, A., Cemek, M., Simsek, N., 2009. The effects of Panax ginseng and Spir-ulina platensis on hepatoxicity induced by cadmium in rats. Ecotoxicol. Environ.Safety 72, 231–235.
ergosien, D.H., Rice, C.D., 1998. Macrophage secretory function is enhanced bylow doses of tributyltin-oxide (TBTO), but not tributyltin-chloride (TBTCl). Arch.Environ. Contam. Toxicol. 34, 223–228.
han, M., Varadharaj, S., Shobha, J.C., Naidu, M.U., Parinandi, N.L., Kutala, V.K., Kup-pusamy, P., 2006. C-Phycocyanin ameliorates doxorubicin-induced oxidativestress and apoptosis in adult rat cardiomyocytes. J. Cardiovasc. Pharmacol. 47,9–20.
otake, Y., 2007. Neurotoxicty induced by environmental low molecular weightsubstances. J. Health Sci. 53, 639–643.
rajnc, E.I., Wester, P.W., Loeber, J.G., Van Leeuwen, F.X.R., Vos, J.G., Vaessen,H.A.M.G., Van der Heijden, C.A., 1984. Toxicity of bis(tri-n-butyltin) oxide inthe rat. I. Short-term effects on general parameters and on the endocrine andlymphoid systems. Toxicol. Appl. Pharmacol. 75, 363–386.
ee, C., Chen, I., Lee, C., Chi, C., Tsai, M., Tsai, J., Lin, H., 2010. Inhibition of gap
junctional intercellular communication in WB-F344 rat liver epithelial cells bytriphenyltin chloride through MAPK and PI3-kinase pathways. J. Occup. Med.Toxicol. 5, 17–27.iu, H., Xu, L., 2007. Garlic oil prevents tributyltin-induced oxidative damage in vivoand in vitro. J. Food Prot. 70, 716–721.
etters 204 (2011) 2–11 11
Liu, H., Wang, Y., Lian, L., Xu, H., 2006. Tributyltin induces DNA damage as well asoxidative damage in rats. Environ. Toxicol. 21, 166–171.
Liu, H., Guo, Z., Xu, L., Hsu, S., 2008. Protective effect of green tea polyphenols ontributyltin-induced oxidative damage detected by in vivo and in vitro models.Environ. Toxicol. 23, 77–83.
Lu, H., Hsieh, C., Hsu, J., Yang, Y., Chou, H.N., 2006. Preventive effects of Spirulinaplatensis on skeletal muscle damage under exercise-induced oxidative stress.Eur. J. Appl. Physiol. 98, 220–226.
Mani, U.V., Iyer, U.M., Dhruv, S.A., Mani, I.U., Sharma, K.S., 2007. Therapeutic utilityof spirulina. In: Gershwin, M.E., Belay, A. (Eds.), Spirulina in Human Nutritionand Health. CRC Press. Taylor & Francis group, Tokyo, pp. 71–95.
Mundy, W.R., Freudenrich, T.M., 2006. Apoptosis of cerebellar granule cells inducedby organotin compounds found in drinking water: involvement of MAP kinases.Neurotoxicology 27, 71–81.
Nakatsu, Y., Kotake, Y., Ohta, S., 2007. Concentration dependence of the mechanismsof tributyltin-induced apoptosis. Toxicol. Sci. 97, 438–447.
Nakatsu, Y., Kotake, Y., Takai, N., Ohta, S., 2010. Involvement of autophagy via mam-malian target of rapamycin (mTOR) inhibition in tributyltin-induced neuronalcell death. J. Toxicol. Sci. 35, 245–251.
Patel, A., Mishra, S., Ghosh, P.K., 2006. Antioxidant potential of C-Phycocyaninisolated from cyanobacterial species Lyngbya, Phormidium and Spirulina spp.Indian J. Biochem. Biophys. 43, 25–31.
Ponce-Canchihuaman, J.C., Perez-Mendez, O., Hernandez-Munoz, R., Torres-Duran,P.V., Juarez-Oropeza, M.A., 2010. Protective effects of Spirulina maxima onhyperlipidaemia and oxidative-stress induced by lead acetate in the liver andkidney. Lipids Health Dis. 9, 35.
Raffray, M., Cohen, G.M., 1993. Thymocyte apoptosis as a mechanism for tributyltin-induced thymic atrophy in vivo. Arch. Toxicol. 67, 231–236.
Ravi, M., De, S.L., Azharuddin, S., Paul, S.F.D., 2010. The beneficial effects of spirulinafocusing on its immunomodulatory and antioxidant properties. Nutr. DietarySuppl. 2, 73–83.
Reddy, C.M., Bhat, V.B., Kiranmai, G., Reddy, M.N., Reddanna, P., Madyastha, K.M.,2000. Selective inhibition of cyclooxygenase-2 by C-phycocyanin, a biliproteinfrom Spirulina platensis. Biochem. Biophys. Res. Commun. 277, 599–603.
Romay, C., Armesto, J., Remirez, D., González, R., Ledon, N., Garc�́a, I., 1998. Antioxi-dant and anti-inflammatory properties of C-phycocyanin from blue-green algae.Inflamm. Res. 47, 36–41.
Royall, J.A., Ischiropoulos, H., 1993. Evaluation of 2′ ,7′-dichlorofluorescein and dihy-drorhodamine 123 as fluorescent probes for intracellular H2O2 in culturedendothelial cells. Arch. Biochem. Biophys. 302, 348–355.
Schwartz, J., Shklar, G., 1987. Regression of experimental hamster cancer by betacarotene and algae extracts. J. Oral Maxillofac. Surg. 45, 510–515.
Seinen, W., Willems, M.I., 1976. Toxicity of organotin compounds. I. Atrophyof thymus and thymus-dependent lymphoid tissue in the rat fed di-n-octyltindichloride. Toxicol. Appl. Pharmacol. 35, 63–75.
Seinen, W., Vos, J.G., Van Krieken, R., Penninks, A.H., Brands, R., Hooykaas, H., 1977.Toxicity of organotin compounds. III. Suppression of thymus-dependent immu-nity in rats by di-n-butyltindichloride and dison, n-octyltindichloride. Toxicol.Appl. Pharmacol. 44, 213–224.
Snoeij, N.J., Penninks, A.H., Seinen, W., 1987. Biological activity of organotin com-pounds – an overview. Environ. Res. 44, 335–353.
Snoeij, N.J., Penninks, A.H., Seinen, W., 1988. Dibutyltin and tributyltin compoundsinduce thymus atrophy in rats due to a selective action on thymic lymphoblasts.Int. J. Immunopharmacol. 10, 891–899.
Subhashini, J., Mahipal, S.V., Reddy, M.C., Mallikarjuna Reddy, M., Rachamallu, A.,Reddanna, P., 2004. Molecular mechanisms in C-Phycocyanin induced apoptosisin human chronic myeloid leukemia cell line-K562. Biochem. Pharmacol. 68,453–462.
Suresh, M., Mishra, S.K., Mishra, S., Das, A., 2009. The detection of Hg2+ by Cyanobac-teria in aqueous media. Chem. Commun. 14, 2496–2498.
Ueno, S., Kashimoto, T., Susa, N., Asai, T., Kawaguchi, S., Takeda-Homma, S., Terada, Y.,Sugiyama, M., 2009. Reduction in peripheral lymphocytes and thymus atrophyinduced by organotin compounds in vivo. J. Vet. Med. Sci. 71, 1041–1048.
Vos, J.G., De Klerk, A., Krajnc, E.I., Kruizinga, W., van Ommen, B., Rozing, J., 1984. Tox-icity of bis(tri-n-butyltin)oxide in the rat. II. Suppression of thymus-dependent
immune responses and of parameters of nonspecific resistance after short-termexposure. Toxicol. Appl. Pharmacol. 75, 387–408.Zhang, J., Zuo, Z., He, C., Cai, J., Wang, Y., Chen, Y., Wang, C., 2009. Effect of tributyltinon testicular development in Sebastiscus marmoratus and the mechanisminvolved. Environ. Toxicol. Chem. 28, 1528–1535.





























