Embed Size (px)
Citation preview

Investigative Ophthalmology & Visual Science, Vol. 31, No. 11, November 1990Copyright © Association for Research in Vision and Ophthalmology
Calcium-Induced Opacification and Profeolysisin the Intact Rat Lens
R. J. W. Truscorr,* J. M. Marcanfonio,t J- Tomlinson,-(- and G. Duncanf
When intact rat lenses were incubated in artificial aqueous humor in the presence of 1 mM calcium anda sulfhydryl reagent p-chloromercuriphenyl sulfonate (pCMPS) a visible annular opacity developedwithin 4 hours. Combined photographic and ion-sensitive microelectrode investigations of the lensesdemonstrated that the subsequent linear increase in opacification was accompanied by an increase ininternal free calcium. Opacities were not observed in lenses incubated in the absence of either pCMPSor calcium. Gel electrophoresis of the soluble and urea-soluble fractions from lenses exposed to 1 mMcalcium for periods of up to 14 hours showed no evidence for crystallin degradation and only minorproteolysis of cytoskeletal proteins. When lenses were incubated under identical conditions, but with 5mM calcium, the degree of opacification increased up to approximately 8 hours and then remainedconstant. A progressive loss in cytoskeletal proteins was observed which correlated with a furtherincrease in free calcium such that by 14 hours of incubation, when the internal calcium approached 1mM, most of the spectrin and vimentin present in the cortex of the lens had disappeared. An unidenti-fied 110-kilodalton protein also disappeared from lenses incubated in 5 mM calcium. These resultsindicate that proteolysis by calcium-dependent enzymes such as calpain may play a significant role incytoskeletal regulation and metabolism in the lens. A role for cytoskeleton/membrane/crystallininteraction in calcium-induced opacification is discussed. Invest Ophthalmol Vis Sci 31:2405-2411, 1990
Recent investigations highlight the role played bycalcium in the development of senile cortical cataractin humans.1'2 The importance of internal calcium-ion concentration has also been established in severalanimal model systems. In organ-cultured bovinelenses, for example, a decrease in transparency wasobserved only in lenses with increased levels of cal-cium.3 In selenite cataract, calcium levels in the nu-clear region of the rat lens, where the opacity is local-ized, are increased more than tenfold over controlvalues.4 The development of selenite cataract is alsoassociated with an increase in insoluble protein5 andextensive proteolysis.6
It is known that increased concentrations of cal-cium lead to the activation of proteases such as cal-pain7 and that calpain I, calpain II, and calpastatin, a
From the *Australian Cataract Research Foundation, Universityof Wollongong, Wollongong, New South Wales, Australia, and thefSchool of Biological Sciences, University of East Anglia, Norwich,United Kingdom.
Supported in part by grants from the Royal Society/AustralianAcademy of Science, the Wellcome/Ramaciotti Foundation(R.J.W.T.) and the Nuffield Foundation. J.M.M. is an RNIB Se-nior Research Fellow and J.T. an SERC-Pfizer CASE student.
Reprint requests: Dr. R. J. W. Truscott, Australian CataractResearch Foundation, University of Wollongong, Wollongong,NSW 2500, Australia.
specific endogenous inhibitor of calpain, are presentin the lens.8 Calcium has also been found to produceaggregation of proteins when added to solutions ofsoluble lens proteins.9
There is currently much interest in the potentialuse of calpain inhibitors as agents for the preventionof cataract. Central to an understanding of calcium-dependent cataract formation, however, is the eluci-dation of the mechanism whereby calcium inducesopacification and whether proteolysis is a necessaryprecondition for opacification in the intact lens. Thisstudy was undertaken to investigate these issues: (1)the relationship between intracellular free calciumand lens opacities and (2) the concomitant inductionof lens proteolyses.
Materials and Methods
Lenses were dissected from Wistar rats killed bycervical dislocation. Each lens was washed in 4 ml ofmodified artificial aqueous humor (AAH) solution10
containing 1 or 5 mM CaCl2 together with 40 ^M ofthe sulfhydryl reagent p-chloromercuriphenyl sulfo-nate (pCMPS). The pCMPS was chosen since it hasbeen shown to stimulate calcium influx into the ratlens without penetrating the cytoplasm.10 Lenseswere then incubated in a capped vial at 36°C in 8 ml
2405
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933154/ on 02/12/2018

2406 INVESTIGATIVE OPHTHALMOLOGY & VISUAL SCIENCE / November 1990 Vol. 31
of A AH containing 1 or 5 mM CaCl2 and pCMPS fortimes up to 18 hr. Free-calcium levels in the lenseswere measured using calcium-sensitive microelec-trodes, inserted 150-200 fim into the posterior cor-tex, as described previously.'' Lenses were transferredfrom the incubation vials to small plastic petri dishesand photographed in AAH by transmitted light ontoIlford 100 ASA black-and-white film (Cheshire,U.K.). Negatives were scanned with 626-nm light in adensitometer (Joyce Loebl, Chromoscan 3, Joyce-Loebl, Gareshead, U.K.). This procedure was foundto give more reproducible results than those obtainedfrom lenses photographed against a black back-ground (data not shown).
At the end of incubation the nucleus was removedfrom each lens using a trephine (2-mm internal diam-eter), and the cortex was frozen in liquid nitrogen andstored at —20°C. The cortex was homogenized in 400n\ of 40 mM Tris, pH 7.5, containing 5 mM Ethylene,glycol-bis (Baminoethyl ether)N,N,N',N'-tetraceticacid (EGTA), 5 mM ethylenediaminetetraacetic acid(EDTA), 1 mM N-ethyl maleimide, and 0.5 mMPhenylmethyl sulfonyl fluoride (PMSF) and centri-fuged (10,000 X g for 15 min). The pellet was reex-tracted as described and then dissolved in 60 ix\ of 8M urea containing 1 mM EDTA by shaking for 2 hr.A sample (20 /ul) was mixed with an equal volume ofsample buffer (6.25 mM Tris pH 6.8, 1.0% sodiumdodecyl sulfate [SDS], 15% glycerol, and 5% v/v 2-mercaptoethanol), and 20 ;ul was used for electropho-resis. Discontinuous one-dimentional gel electropho-resis was done as described by Laemmli.12 Proteinswere stained with colloidal Coomassie blue13 and thegels scanned at 626 nm, like the photographic nega-tives.
Proteins were transferred to nitrocellulose mem-branes for immunoblot analysis, essentially as de-scribed by Towbin et al.14 After blocking the mem-branes with 3% bovine serum albumin, the blots wereprobed as follows: (1) for spectrin, we used a mono-specific polyclonal antibody to sheep erythrocytespectrin,15 diluted 1:400, overnight at 4°C, (2) forvimentin, we used anti-IFA, a panspecific monoclo-nal antibody which recognizes a determinant com-mon to all intermediate filament proteins,16 undi-luted, overnight at 4°, (3) for actin, we used monoclo-nal antiactin (Amersham, Aylesbury, Bucks, U.K.),diluted 1:1000, overnight at 4°C, and (4) for protein115, we used a monoclonal antibody to the bovinelens fiber cell Mrl 15-kilodalton (kD) protein,17 di-luted 1:1000, for 4 hr at room temperature. Peroxi-dase-conjugated second antibodies (Sigma, St. Louis,MO) were diluted 1:2000 and applied for 1 hr atroom temperature, with 3-amino-9-ethylcarbazole asthe chromophoric agent.
All animals were treated in accordance with the
ARVO Resolution on the Use of Animals in Re-search.
ResultsSince pCMPS was being used under two conditions
(1 mM and 5 mM Ca2+) to induce opacification, theeffect of the sulfhydryl complexing agent on lensmembrane potentials was monitored. The data(Table 1) confirm that a very large depolarization wasinduced, which appeared to be independent of thelevel of external calcium. This indicates that pCMPSinduces a large change in membrane permeability, ashas previously been suggested,10 with a consequentialchange in internal sodium and calcium. Since Ca2+
rather than Na+ is responsible for the major change intransparency during cataract formation, we investi-gated, in detail, the relationship between internalCa2+ changes and opacification.
In the intact rat lens a visible whitesannular opacitycould be observed after approximately a 4 hr incuba-tion in AAH containing 40 /xM pCMPS and 1 mMcalcium. The degree of opacification appeared to in-crease with time. Lenses incubated in the absence ofeither calcium or pCMPS remained transparent forover 18 hr. This is illustrated in Figure 1 where a lensincubated in AAH containing 1 mM calcium andpCMPS for 14 hr (Fig. la) is shown together with alens incubated for the same time in the absence ofpCMPS(Fig. lb).
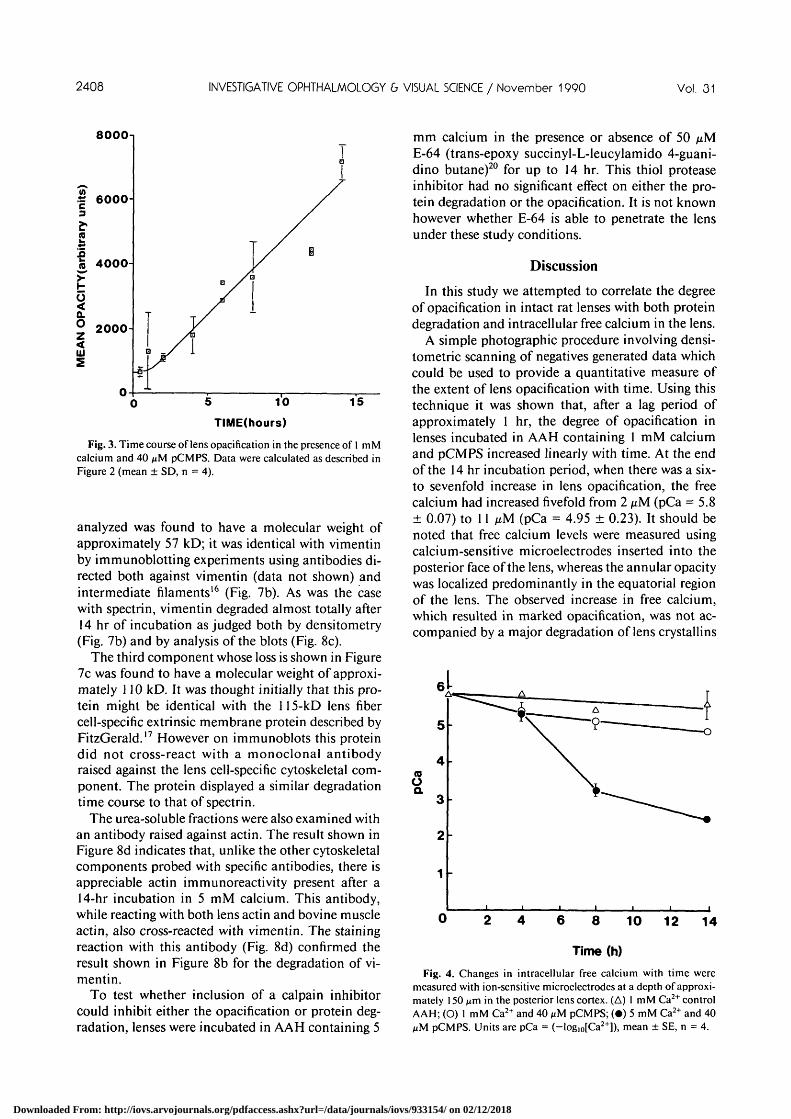
To quantify the extent of the cataract as a functionof time, lenses were photographed and analyzed bydensitometric scanning as described. The data ob-tained for the lenses shown in Figure 1 are depicted inFigure 2. The degree of opacification of each lens wascalculated by summing the areas of the peaks in thedensitometric profile. Using this procedure the degreeof lens opacification as a function of the time of in-cubation could be determined. The results obtainedfor lenses incubated in solutions containing pCMPSand 1 mM calcium are shown in Figure 3. The degreeof opacification increased steadily with time after alag period of approximately 1 hr. This increase inopacification was accompanied by a small increase inintracellular free calcium as determined by calciumsensitive microelectrodes (Fig. 4).
Table 1. Membrane voltage measurementscorresponding to pCa values given in Figure 4.
Time(hr)
048
14
Control(mV)
-73.9 ± 3.3-70.3 ± 1.3-64.1 ±4.4-60.9 ± 4.8
(4)(6)(3)(3)
40 fiM pCMPS+ 1 mM Ca2+
(mV)
-18.1 ±4.4(4)-14.1 ± 1.8(3)-6.3 ± 1.1 (2)
40 fiM pCMPS+ 5 mM Ca2+
(mV)
_-20.2 ± 4.95 (2)-21.7 ±3 .5 (4)
-3 .5 ±0 .9 (3)
The voltage values are expressed as mean ± SEM (n).
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933154/ on 02/12/2018
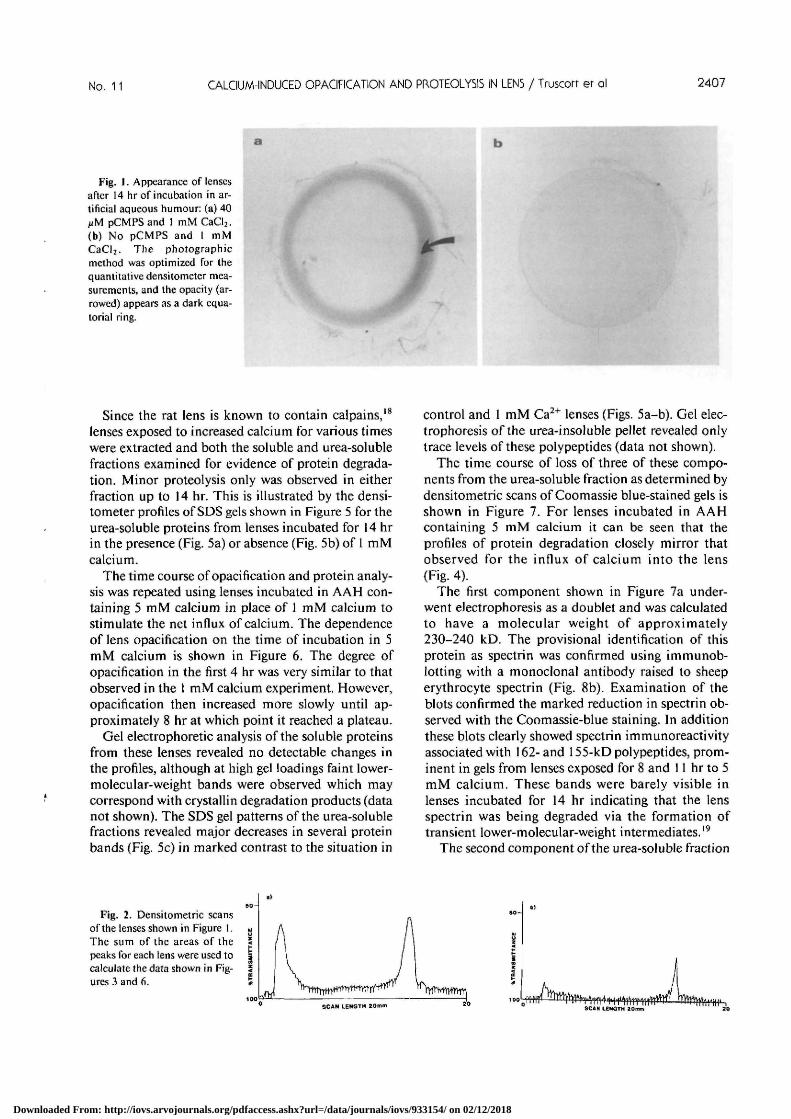
No. 11 CALCIUM-INDUCED OPACIFICATION AND PROTEOLY5IS IN LENS / Truscorr er al 2407
Fig. 1. Appearance of lensesafter 14 hr of incubation in ar-tificial aqueous humour: (a) 40iiM pCMPS and 1 mM CaCl2.(b) No pCMPS and I mMCaCl2. The photographicmethod was optimized for thequantitative densitometer mea-surements, and the opacity (ar-rowed) appears as a dark equa-torial ring.
Since the rat lens is known to contain calpains,18
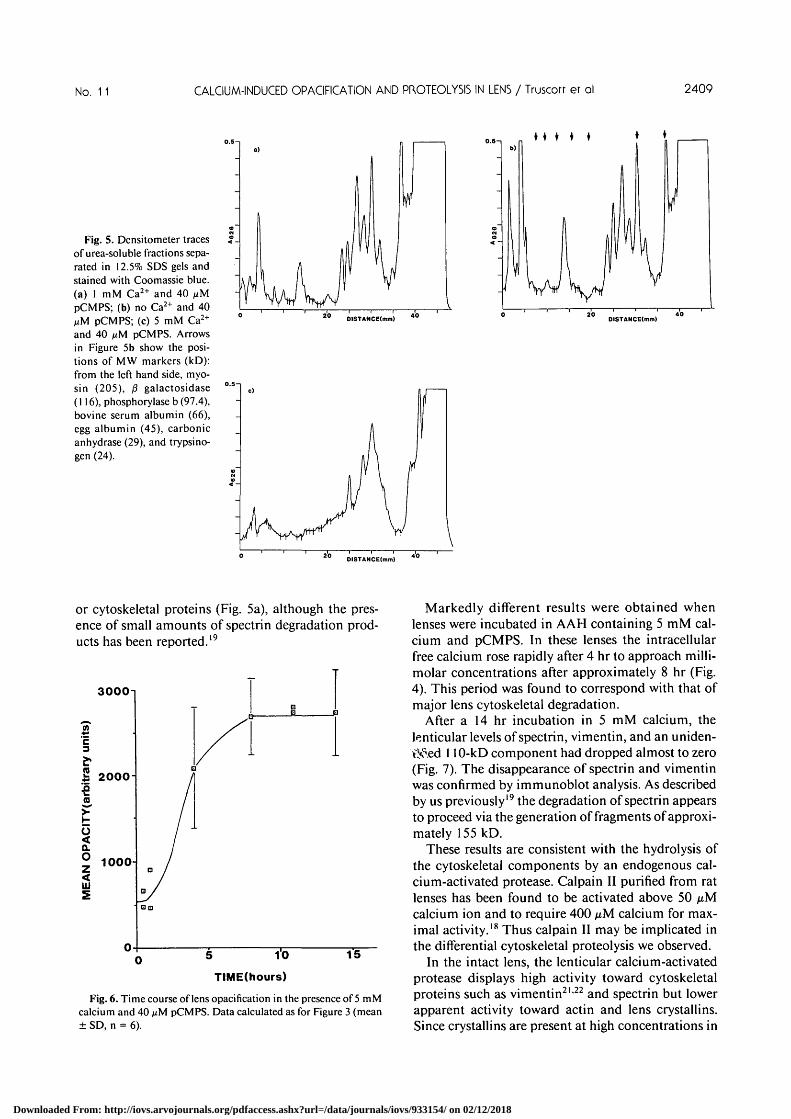
lenses exposed to increased calcium for various timeswere extracted and both the soluble and urea-solublefractions examined for evidence of protein degrada-tion. Minor proteolysis only was observed in eitherfraction up to 14 hr. This is illustrated by the densi-tometer profiles of SDS gels shown in Figure 5 for theurea-soluble proteins from lenses incubated for 14 hrin the presence (Fig. 5a) or absence (Fig. 5b) of 1 mMcalcium.
The time course of opacification and protein analy-sis was repeated using lenses incubated in AAH con-taining 5 mM calcium in place of 1 mM calcium tostimulate the net influx of calcium. The dependenceof lens opacification on the time of incubation in 5mM calcium is shown in Figure 6. The degree ofopacification in the first 4 hr was very similar to thatobserved in the 1 mM calcium experiment. However,opacification then increased more slowly until ap-proximately 8 hr at which point it reached a plateau.
Gel electrophoretic analysis of the soluble proteinsfrom these lenses revealed no detectable changes inthe profiles, although at high gel loadings faint lower-molecular-weight bands were observed which maycorrespond with crystallin degradation products (datanot shown). The SDS gel patterns of the urea-solublefractions revealed major decreases in several proteinbands (Fig. 5c) in marked contrast to the situation in
control and 1 mM Ca2+ lenses (Figs. 5a-b). Gel elec-trophoresis of the urea-insoluble pellet revealed onlytrace levels of these polypeptides (data not shown).
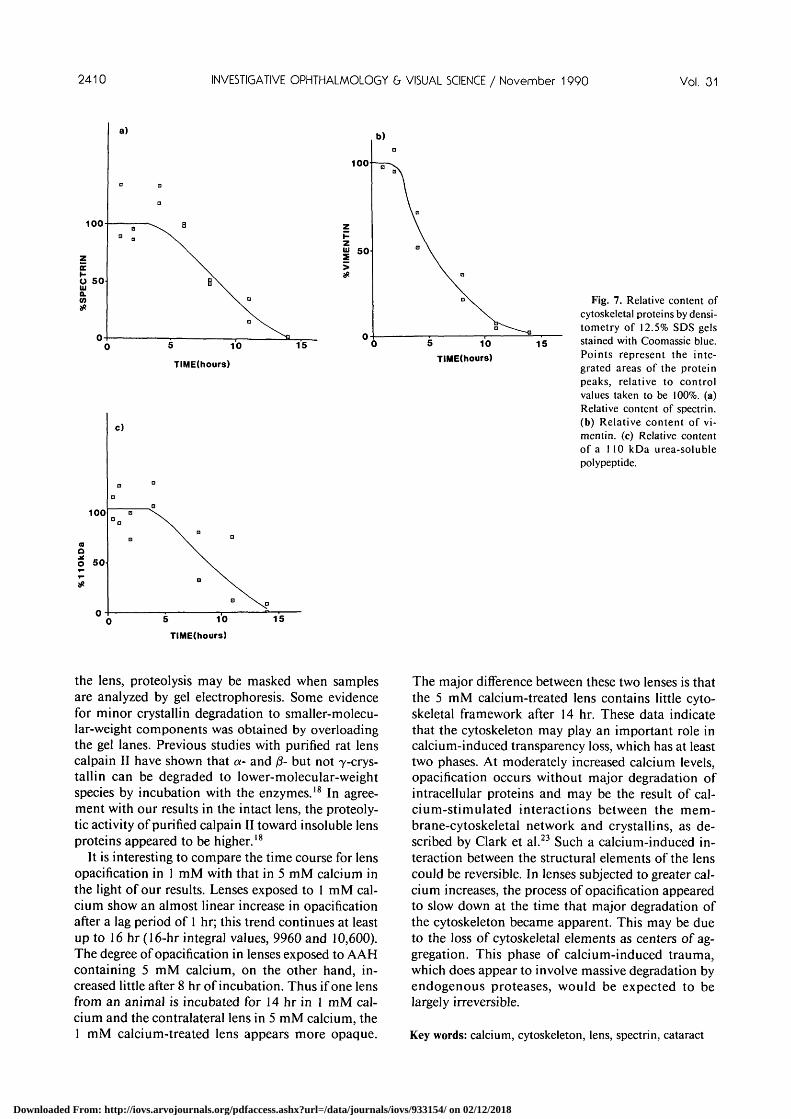
The time course of loss of three of these compo-nents from the urea-soluble fraction as determined bydensitometric scans of Coomassie blue-stained gels isshown in Figure 7. For lenses incubated in AAHcontaining 5 mM calcium it can be seen that theprofiles of protein degradation closely mirror thatobserved for the influx of calcium into the lens(Fig. 4).
The first component shown in Figure 7a under-went electrophoresis as a doublet and was calculatedto have a molecular weight of approximately230-240 kD. The provisional identification of thisprotein as spectrin was confirmed using immunob-lotting with a monoclonal antibody raised to sheeperythrocyte spectrin (Fig. 8b). Examination of theblots confirmed the marked reduction in spectrin ob-served with the Coomassie-blue staining. In additionthese blots clearly showed spectrin immunoreactivityassociated with 162- and 155-kD polypeptides, prom-inent in gels from lenses exposed for 8 and 11 hr to 5mM calcium. These bands were barely visible inlenses incubated for 14 hr indicating that the lensspectrin was being degraded via the formation oftransient lower-molecular-weight intermediates.19
The second component of the urea-soluble fraction
Fig. 2. Densitometric scansof the lenses shown in Figure 1.The sum of the areas of thepeaks for each lens were used tocalculate the data shown in Fig-ures 3 and 6.
SCAN LENGTH 20 SCAN LENGTH 10
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933154/ on 02/12/2018
2408 INVESTIGATIVE OPHTHALMOLOGY & VISUAL SCIENCE / November 1990 Vol. 31
8000-
| 6000-3
Ii 4000-
2000H
5 10TIME(hours)
15
Fig. 3. Time course of lens opacification in the presence of I mMcalcium and 40 fiM pCMPS. Data were calculated as described inFigure 2 (mean ± SD, n = 4).
analyzed was found to have a molecular weight ofapproximately 57 kD; it was identical with vimentinby immunoblotting experiments using antibodies di-rected both against vimentin (data not shown) andintermediate filaments16 (Fig. 7b). As was the casewith spectrin, vimentin degraded almost totally after14 hr of incubation as judged both by densitometry(Fig. 7b) and by analysis of the blots (Fig. 8c).
The third component whose loss is shown in Figure7c was found to have a molecular weight of approxi-mately 110 kD. It was thought initially that this pro-tein might be identical with the 115-kD lens fibercell-specific extrinsic membrane protein described byFitzGerald.17 However on immunoblots this proteindid not cross-react with a monoclonal antibodyraised against the lens cell-specific cytoskeletal com-ponent. The protein displayed a similar degradationtime course to that of spectrin.
The urea-soluble fractions were also examined withan antibody raised against actin. The result shown inFigure 8d indicates that, unlike the other cytoskeletalcomponents probed with specific antibodies, there isappreciable actin immunoreactivity present after a14-hr incubation in 5 mM calcium. This antibody,while reacting with both lens actin and bovine muscleactin, also cross-reacted with vimentin. The stainingreaction with this antibody (Fig. 8d) confirmed theresult shown in Figure 8b for the degradation of vi-mentin.
To test whether inclusion of a calpain inhibitorcould inhibit either the opacification or protein deg-radation, lenses were incubated in AAH containing 5
mm calcium in the presence or absence of 50E-64 (trans-epoxy succinyl-L-leucylamido 4-guani-dino butane)20 for up to 14 hr. This thiol proteaseinhibitor had no significant effect on either the pro-tein degradation or the opacification. It is not knownhowever whether E-64 is able to penetrate the lensunder these study conditions.
Discussion
In this study we attempted to correlate the degreeof opacification in intact rat lenses with both proteindegradation and intracellular free calcium in the lens.
A simple photographic procedure involving densi-tometric scanning of negatives generated data whichcould be used to provide a quantitative measure ofthe extent of lens opacification with time. Using thistechnique it was shown that, after a lag period ofapproximately 1 hr, the degree of opacification inlenses incubated in AAH containing 1 mM calciumand pCMPS increased linearly with time. At the endof the 14 hr incubation period, when there was a six-to sevenfold increase in lens opacification, the freecalcium had increased fivefold from 2 ixM (pCa = 5.8± 0.07) to 11 fiM (pCa = 4.95 ± 0.23). It should benoted that free calcium levels were measured usingcalcium-sensitive microelectrodes inserted into theposterior face of the lens, whereas the annular opacitywas localized predominantly in the equatorial regionof the lens. The observed increase in free calcium,which resulted in marked opacification, was not ac-companied by a major degradation of lens crystallins
sa.
Time (h)
Fig. 4. Changes in intracellular free calcium with time weremeasured with ion-sensitive microelectrodes at a depth of approxi-mately 150 /urn in the posterior lens cortex. (A) 1 mM Ca2+ controlAAH; (O) 1 mM Ca2+ and 40 nM pCMPS; (•) 5 mM Ca2+ and 40MM pCMPS. Units are pCa = (-log,o[Ca2+]), mean ± SE, n = 4.
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933154/ on 02/12/2018
No. 11 CALCIUM-INDUCED OPACIFICATION AND PROTEOLYSIS IN LENS / Truscorr er ol 2409
Fig. 5. Densitometer tracesof urea-soluble fractions sepa-rated in 12.5% SDS gels andstained with Coomassie blue,(a) 1 mM Ca2+ and 40 /xMpCMPS; (b) no Ca2+ and 40MM pCMPS; (c) 5 mM Ca2+
and 40 nM pCMPS. Arrowsin Figure 5b show the posi-tions of MW markers (kD):from the left hand side, myo-sin (205), /3 galactosidase(116), phosphorylase b (97.4),bovine serum albumin (66),egg albumin (45), carbonicanhydrase (29), and trypsino-gen (24).
ns • • • • •).o—i
-
<o
-
b)
. I .
I'1/
VDISTANCE(mm) DISTANCE(mm)
DISTANCE(mm)
or cytoskeletal proteins (Fig. 5a), although the pres-ence of small amounts of spectrin degradation prod-ucts has been reported.19
3000-
E3
S 2000-
(arb
iV
CIT
Y
a.° 1000
in
n
t
/
/ -
Ia /
D /
• a
0
DB C
/
/
5 i'O
TIME(hours)
]
15
Fig. 6. Time course of lens opacification in the presence of 5 mMcalcium and 40 MM pCMPS. Data calculated as for Figure 3 (mean± SD, n = 6).
Markedly different results were obtained whenlenses were incubated in AAH containing 5 mM cal-cium and pCMPS. In these lenses the intracellularfree calcium rose rapidly after 4 hr to approach milli-molar concentrations after approximately 8 hr (Fig.4). This period was found to correspond with that ofmajor lens cytoskeletal degradation.
After a 14 hr incubation in 5 mM calcium, thelenticular levels of spectrin, vimentin, and an uniden-t>?ed 110-kD component had dropped almost to zero(Fig. 7). The disappearance of spectrin and vimentinwas confirmed by immunoblot analysis. As describedby us previously19 the degradation of spectrin appearsto proceed via the generation of fragments of approxi-mately 155 kD.
These results are consistent with the hydrolysis ofthe cytoskeletal components by an endogenous cal-cium-activated protease. Calpain II purified from ratlenses has been found to be activated above 50 /xMcalcium ion and to require 400 ^M calcium for max-imal activity.18 Thus calpain II may be implicated inthe differential cytoskeletal proteolysis we observed.
In the intact lens, the lenticular calcium-activatedprotease displays high activity toward cytoskeletalproteins such as vimentin21'22 and spectrin but lowerapparent activity toward actin and lens crystallins.Since crystallins are present at high concentrations in
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933154/ on 02/12/2018
2410 INVESTIGATIVE OPHTHALMOLOGY b VISUAL SCIENCE / November 1990 Vol. 01
100
z£O 50UJa.<n
0-
a)
D Q
D
a \ B
\
B\\ . El
100
5 iO
TIME(hours)
15
100
aO 50
c)
5 10TIME(hours)
15
5 10TIME(hours)
Fig. 7. Relative content ofcytoskeletal proteins by densi-tometry of 12.5% SDS gelsstained with Coomassie blue.Points represent the inte-grated areas of the proteinpeaks, relative to controlvalues taken to be 100%. (a)Relative content of spectrin.(b) Relative content of vi-mentin. (c) Relative contentof a 110 kDa urea-solublepolypeptide.
the lens, proteolysis may be masked when samplesare analyzed by gel electrophoresis. Some evidencefor minor crystallin degradation to smaller-molecu-lar-weight components was obtained by overloadingthe gel lanes. Previous studies with purified rat lenscalpain II have shown that a- and (3- but not y-crys-tallin can be degraded to lower-molecular-weightspecies by incubation with the enzymes.18 In agree-ment with our results in the intact lens, the proteoly-tic activity of purified calpain II toward insoluble lensproteins appeared to be higher.18
It is interesting to compare the time course for lensopacification in 1 mM with that in 5 mM calcium inthe light of our results. Lenses exposed to 1 mM cal-cium show an almost linear increase in opacificationafter a lag period of 1 hr; this trend continues at leastup to 16 hr (16-hr integral values, 9960 and 10,600).The degree of opacification in lenses exposed to A AHcontaining 5 mM calcium, on the other hand, in-creased little after 8 hr of incubation. Thus if one lensfrom an animal is incubated for 14 hr in 1 mM cal-cium and the contralateral lens in 5 mM calcium, the1 mM calcium-treated lens appears more opaque.
The major difference between these two lenses is thatthe 5 mM calcium-treated lens contains little cyto-skeletal framework after 14 hr. These data indicatethat the cytoskeleton may play an important role incalcium-induced transparency loss, which has at leasttwo phases. At moderately increased calcium levels,opacification occurs without major degradation ofintracellular proteins and may be the result of cal-cium-stimulated interactions between the mem-brane-cytoskeletal network and crystallins, as de-scribed by Clark et al.23 Such a calcium-induced in-teraction between the structural elements of the lenscould be reversible. In lenses subjected to greater cal-cium increases, the process of opacification appearedto slow down at the time that major degradation ofthe cytoskeleton became apparent. This may be dueto the loss of cytoskeletal elements as centers of ag-gregation. This phase of calcium-induced trauma,which does appear to involve massive degradation byendogenous proteases, would be expected to belargely irreversible.
Key words: calcium, cytoskeleton, lens, spectrin, cataract
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933154/ on 02/12/2018

No. 11 CALCIUM-INDUCED OPACIFICATION AND PROTEOLY515 IN LENS / Truscorr er ol 2411
c) d)
Fig. 8. Identification of poly-peptides. In all figures, lane 1,no pCMPS and 1 mM CaCl2:lane 2, 40 fiM pCMPS and 5mM CaCl2, after 14 hr of incu-bation, (a) Urea-soluble pro-teins separated in 12.5% SDSgels and stained with Coomas-sie blue, (b) Western blot from a6% SDS gel, probed with an an-tibody to sheep erythrocytespectrin. (c) Western blot from12.5 SDS gel, probed with anantibody to intermediate fila-ments, (d) As for c, probed withanti-actin.
205-
205—
-205
-116- 97- 66
- 45- 36- 29- 24
1 2 1 2 1 2 1 2
AcknowledgmentsThe authors thank Dr. Alan Hargreaves of the John
Innes Institute for providing the antibodies to spectrin andvimentin, Dr. Paul FitzGerald of the University of Califor-nia, Davis, for providing the monoclonal antibody to thefiber cell-specific extrinsic membrane protein, Dr. NoelEllis for the use of the densitometer, Sheila Davies for thephotography, and Jill Gorton for producing the manu-script.
References1. Duncan G and Bushell AR: Ion analysis of human cataractous
lenses. Exp Eye Res 20:223, 1975.2. Hightower KR and Reddy VN: Calcium content and distribu-
tion in human cataract. Exp Eye Res 34:413, 1982.3. Marcantonio JM, Duncan G, and Rink H: Calcium-induced
opacification and loss of protein in the organ-cultured bovinelens. Exp Eye Res 42:617, 1986.
4. Hightower KR, David LL, and Shearer TR: Regional distribu-tion of free calcium in selenite cataract: Relation to calpain II.Invest Ophthalmol Vis Sci 28:1702, 1987.
5. Bunce GE and Hess JL: Biochemical changes associated withselenite-induced cataract in the rat. Exp Eye Res 33:505, 1981.
6. David LL and Shearer TR: Calcium-activated proteolysis inthe lens nucleus during selenite cataractogenesis. InvestOphthalmol Vis Sci 25:1275, 1984.
7. Suzuki K: Calcium activated neutral protease: Domain struc-ture and activity regulation. Trends in Biochemical Science12:103, 1987.
8. Yoshida H, Murachi T, and Tsukahara I: Distribution of cal-pain I, calpain II and calpastatin in the bovine lens. InvestOphthalmol Vis Sci 26:953, 1985.
9. Jedziniak JA, Kinoshita JH, Yates EM, Hocker LO, and Ben-edek GB: Calcium-induced aggregation of bovine lens alphacrystallins. Invest Ophthalmol Vis Sci 11:905, 1972.
10. Hightower K: Superficial membrane SH groups inaccessible byintracellular GSH. Curr Eye Res 5:421, 1986.
11. Duncan G and Jacob TJC: Influence of external calcium andglucose on internal total and ionised calcium in the rat lens. JPhysiol (Lond) 357:485, 1984.
12. Laemmli UK: Cleavage of structural proteins during the as-sembly of the head bacteriophage T4. Nature 227:680, 1970.
13. NeuhofT V, Arnold N, Taube D, and Ehrhardt W: Improvedstaining of proteins in PAGE including IEF with clear back-ground at nanogram sensitivity using Coomassie brilliant blueG25Oand R25O. Electrophoresis 9:255, 1988.
14. Towbin H, Staehelin T, and Gordon J: Electrophoretic transferof proteins from acrylamide gels to nitrocellulose sheets: Pro-cedure and some applications. Proc Natl Acad Sci USA76:4350, 1979.
15. Hesketh JE, HarmarAJ, Armstrong AA, and ArbuthnottGW:Immunortistochemical localisation of a spectrin-like protein(fodrin) in nerve cells in culture. Neurosci Lett 63:33, 1986.
16. Pruss RM: Efficient detection of intermediate filament pro-teins using a panspecific monoclonal antibody: Anti IFA. JNeuroimmunol 8:293, 1985.
17. FitzGerald PG: Immunochemical characterisation of a Mr 115lens fiber cell-specific extrinsic membrane protein. Curr EyeRes 7:1243, 1988.
18. David LL and Shearer TR: Purification of calpain II from ratlens and determination of endogenous substrates, Exp Eye Res42:227, 1986.
19. Truscolt RJW, Marcantonio JM, Tomlinson J, and DuncanG: Calcium-induced cleavage and breakdown of spectrin in therat lens. Biochem Biophys ResCommun 162:1472, 1989.
20. Barrett AJ, Kembhavi AA, Brown MA, Kirschke H, KnightCG, Tamai M, and Hanada K: L-trans-epoxysuccinyl-leucyla-mido(4-guanidino)butane (E64) and its analogues as inhibitorsof cysteine proteinases including cathepsins B, H and L. Bio-chem J 201:189, 1982.
21. Ireland M and Maisel H: Evidence for a Ca2+ activated pro-tease specific for lens cytoskeletal proteins. ARVO Abstracts.Invest Ophthalmol Vis Sci 24(Suppl):l29, 1983.
22. Lorand L, Conrad SM, and Velasco PT: Degradation of vi-mentin in a cataract model: The Ca2+-treated lens. Ann N YAcad Sci 455:703, 185.
23. Clark JI, Danford-Kaplan ME, and Delaye M: Calcium de-creases transparency of homogenate from cortex and has noeffect on nucleus. Exp Eye Res 47:447, 1988.
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933154/ on 02/12/2018





























