Embed Size (px)
Citation preview

Citar como: Gamboa-Gaitán MA. 2016. Capítulo 11: Fotosíntesis, pp: 233-252 en: Botánica general:
Introducción al estudio de las plantas. Universidad Nacional de Colombia, Departamento de Biología,
Laboratorio de Biología Tropical. En revisión.
Capítulo 11: Fotosíntesis
Introducción
El proceso de captación de la luz solar y su conversión en energía química es fundamental para la existencia de la vida tal
como se conoce. Este fenómeno al parecer apareció independientemente en varias líneas evolutivas de los dominios
Eukarya y Bacteria, pues está presente en las plantas, algunas algas y las bacterias verdes y rojas. La fotosíntesis es quizá la
propiedad más distintiva de todas las plantas, pues es el proceso que les confiere su capacidad autotrófica. Este capítulo está
dedicado a conocer las bases del proceso de la fotosíntesis, estudiando la estructura del aparato fotosintético, las rutas
bioquímicas que lo caracterizan y el destino de los productos finales de dicho proceso: los fotoasimilados o fotosintatos.
Contenido
11.1. Para la fotosíntesis se requiere una serie de sustancias capaces de absorber luz: los pigmentos.
11.2. Con el fin de aumentar la eficiencia fotosintética, los pigmentos se agregan en complejos supramoleculares dentro de
organelos especializados.
11.3. Una vez absorbida la luz se inician reacciones químicas redox que en conjunto constituyen la fase lumínica de la
fotosíntesis.
11.4. La síntesis no cíclica de ATP se da en un complejo enzimático transmembranal que usa el gradiente de protones entre
el lumen y el estroma.
11.5. Con la energía producida en la fase lumínica se lleva a cabo la fase de fijación del CO2 atmosférico.
11.6. La fotorrespiración disminuye la eficiencia fotosintética.
11.7. Las plantas tienen alternativas fotosintéticas: fotosíntesis C4 y metabolismo ácido de Crasuláceas (MAC).
11.8. La fotosíntesis es un proceso regulado, que puede ser afectado por muchos factores.
11.9. Los productos de la fotosíntesis tienen diversos destinos.
11.10. Para el transporte a larga distancia de los fotosintatos y otras sustancias se requiere un tejido especializado.

Capítulo 11: Fotosíntesis
234
11.1. Para la fotosíntesis se requiere una serie de
sustancias capaces de absorber luz: los pigmentos.
La luz que llega al cuerpo vegetal puede ser reflejada,
transmitida o absorbida, siendo sólo en este último caso
utilizable en algún proceso fisiológico de la planta. Para
que sea absorbida con eficiencia las plantas usan
moléculas especializadas denominadas pigmentos, las
cuales dan color a las diferentes partes del cuerpo, como
hojas o pétalos, según el tipo de luz visible que absorban
o reflejen. A la curva que muestra la cantidad de luz
absorbida por un pigmento en función de la longitud de
onda de la luz incidente, se le denomina espectro de
absorción de dicho pigmento (Figura 11.1). Tal espectro
es de gran utilidad para el estudio de las propiedades y
funciones de los pigmentos. Un motivo químico
recurrente en ello es la presencia de dobles enlaces
alternos, los cuales presentan propiedades de resonancia
cuando son impactados por luz visible, lo que les permite
absorber la energía para tornarla biológicamente
utilizable.
En las plantas los principales pigmentos son las
clorofilas (a y b), así como los carotenoides (Figura
11.2), los cuales se encuentran dispuestos de forma muy
organizada en el interior de los cloroplastos. En efecto,
las moléculas de clorofila y de carotenoides están ligadas
a proteínas en los complejos fotosintéticos. La clorofila a
es la principal molécula fotosintética según queda
evidenciado en la coincidencia de su espectro de
absorción con la mayor actividad fotosintética por
longitud de onda, a lo cual se le denomina espectro de
acción. La clorofila b y los carotenoides son
denominados pigmentos accesorios, ya que sirven como
complemento fotosintético a la clorofila a, como puede
deducirse del hecho que la curva de absorción de esta
molécula no calza perfectamente con la curva del
espectro de acción de la fotosíntesis, por lo que parte de
ella se debe al trabajo de otros pigmentos.
La clorofila a está constituida por una anillo de
tetrapirrol que posee un átomo de magnesio en el centro y
una serie de sustituyentes en la periferia de los anillos,
entre los cuales el más grande es una cadena de fitol. Las
dos longitudes de onda de máxima absorción de la
clorofila a corresponden a 430 y 662 nm, mientras que la
clorofila b posee sus máximos de absorción a los 453 y
642 nm. El rango de absorción óptimo de los
carotenoides suele estar entre 400 y 550 nm, ampliando
así la gama energética de fotones que puede ser absorbida
por el aparato fotosintético.
Los pigmentos tienen un estado basal de mínima
energía (S0), en el cual pueden absorber fotones y pasar a
un estado excitado de mayor energía (S1, S2, etc.), el cual
es inestable y tiende a ceder su energía para regresar a su
estado basal. Este retorno energético puede suceder de
varias formas, como por la pérdida de energía por
emisión de calor, por conversión interna, por emisión de
fotones visibles de menor energía (fluorescencia), o por
fosforescencia. En estos casos, sin embargo, la energía no
es utilizable en procesos fotosintéticos. Para que la
fotosíntesis se dé, existe otra forma de cesión de energía,
llamada transferencia excitónica, en la cual la molécula
excitada transfiere su energía a otra molécula cercana, lo
cual es posible debido a la ordenada disposición de los
pigmentos fotosintéticos.
11.2. Con el fin de aumentar la eficiencia fotosintética,
los pigmentos se organizan en complejos
supramoleculares dentro de organelos especializados.
La fotosíntesis vegetal se realiza en un organelo
especializado denominado cloroplasto. Estos organelos
poseen sus propios ribosomas y ADN, por lo que se cree
que alguna vez fueron microorganismos de vida libre,
aunque actualmente parte de su material genético migró
al núcleo de la célula vegetal. En su interior se

Capítulo 11: Fotosíntesis
235
encuentran las membranas tilacoidales, las cuales son un
sistema endomembranoso formado por una bicapa
lipídica rica en galactolípidos, sulfolípidos y fosfolípidos,
con muchos ácidos grasos poliinsaturados que le dan gran
fluidez. Estos tilacoides se organizan apiladamente
constituyendo los granas (Figura 11.3), que se
interconectan por medio de los tilacoides estromáticos y
albergan de manera muy ordenada los elementos
necesarios para la fotosíntesis, incluyendo los pigmentos.
El sistema endomembranoso tilacoidal es
cerrado y deja en su interior un lumen completamente
separado del estroma, que es la matriz acuosa del
cloroplasto y es equiparable al citoplasma de la célula.
Los tilacoides presentan polaridad transversal (los lípidos
que se orientan hacia el estroma son diferentes a los que
lo hacen hacia el lumen), así como una distribución
lateral heterogénea (los elementos transmembranales del
tilacoide estromático son diferentes de los del grana). Por
ello los tilacoides son estructural y funcionalmente
heterogéneos, ya que su función fotosintética se basa en
cuatro complejos supramoleculares que pueden incluir
proteínas y pigmentos, los cuales se insertan
ordenadamente en las membranas tilacoidales. De
acuerdo con su posición funcional, el primer complejo
consta del sistema recolector de luz II (SRLII) y un
fotosistema II denominado agua-plastoquinona óxido-
reductasa; el segundo complejo se forma por el sistema
recolector de luz I (SRLI) y el fotosistema I, que se
denomina plastocianina-ferredoxina óxido-reductasa; el
tercer complejo se llama citocromo b6f, cuya función es
conectar los dos complejos anteriores mediante
reacciones de óxido reducción; el cuarto complejo se
encarga de producir ATP a partir del gradiente de
protones generado por el flujo de electrones y se
denomina ATP sintasa (Figuras 11.4 y 11.5).
Los SRL están constituidos por polipétidos
pequeños (20-25 kDa), pigmentos clorofílicos (a y b) y
carotenoides (luteínas, violaxantinas, neoxantinas y β-
carotenos), que se disponen formando dos
subcomponentes funcionales: el complejo antenal y el
centro de reacción. El primero absorbe los fotones por los
numerosos pigmentos que posee y dirige la energía de
excitación hacia el centro de reacción, donde es
transformada en energía química debido a unas clorofilas
especiales que permiten la separación de carga y la
transferencia de electrones. Estas clorofilas se denominan
P700 y P680, por las longitudes de onda a las cuales
tienen la máxima absorbancia, ubicándose la primera en
el SRLI y la P680 en el SRLII.
11.3. Una vez absorbida la luz se inician reacciones
químicas redox que en conjunto constituyen la fase
lumínica de la fotosíntesis.
La energía del centro de reacción de la P680 se usa para
impulsar el flujo de electrones desde el donante, la
molécula de agua, hacia el receptor final del fotosistema
II, la plastoquinona. Al comienzo de este proceso
interviene una molécula proteica con átomos de
manganeso (Mn), que descompone la molécula de agua
por oxidación (fotólisis del agua), liberando electrones y
produciendo O2 que es expulsado a la atmósfera. Los
electrones pasan por una serie de intermediarios que
sufren procesos cíclicos de reducción oxidación: la
feofitina y las quinonas. A continuación los electrones
atraviesan el complejo citocromo b6f, que es un agregado
proteico transmembranal con grupos hemos, para llegar a
la plastocianina, la cual los transmitirá finalmente a la
ferredoxina habiendo pasado por el P700, la vitamina K1
y varios grupos sulfoférricos. Esta ruta recorrida por los
electrones se representa con el denominado esquema Z
(Figura 11.4), pudiéndose describir la reacción global de
la transferencia de electrones de la siguiente manera
2 H2O + 2 NADP+ → O2 + 2 NADPH + 2 H
+.

Capítulo 11: Fotosíntesis
236
Este proceso implica el flujo de cuatro
electrones (4 e-) desde el agua hasta el NADP
+, acoplado
al paso de ocho protones (8 H+) del estroma al lumen del
tilacoide, lo cual puede generar diferencias de hasta 4
unidades de pH entre estos compartimentos, siendo el
lumen más ácido. Los estimados indican que por cada
dos electrones que se movilizan en la cadena
transportadora, se fosforilan 1.33 moléculas de ADP y se
traslocan cuatro protones hacia el lumen. Alter-
nativamente a este flujo lineal de electrones, existe la
posibilidad de flujos cíclicos en los que el NADP+ no es
el receptor final, sino que en algún punto de la cadena los
electrones fluyen hacia atrás, a una molécula con
potencial redox más positivo. El fin de estos flujos es
producir ATP sin que se produzca NADPH y parece que
tales vías cíclicas existen en ambos fotosistemas. Este
tipo de producción de ATP se denomina fosforilación
cíclica y es menos eficiente que la resultante del
movimiento lineal de los electrones. Tanto el ATP como
el NADPH son moléculas energéticas, empleadas en las
siguientes etapas de la fotosíntesis para producir azúcares
a partir de la fijación del carbono atmosférico. Es
importante entender que la proporción ATP/NADPH es
regulada constantemente por la célula para mantener un
balance adecuado.
11.4. La síntesis no cíclica de ATP se da en un
complejo enzimático transmembranal que usa el
gradiente de protones entre el lumen y el estroma.
El movimiento lineal de electrones hasta el NADP+ hace
que los protones pasen al lumen generando un gran
aumento en su concentración. La ATP sintasa, complejo
enzimático tilacoidal, conformado por una asociación de
polipétidos que se agrupan en dos dominios, uno
estromático y otro transmembranal, aprovecha el paso de
tales protones a favor de su gradiente electroquímico para
sintetizar ATP a partir de ADP y fosfato inorgánico:
ADP + Pi → ATP + H2O.
Esta es una reacción de fosforilación impulsada por un
gradiente de protones (fuerza protón motriz), que se
origina como consecuencia del paso de protones inducido
por el flujo de electrones de la cadena fotosintética, por
lo que se denomina fotofosforilación. El dominio de la
ATP sintasa que está anclado en la membrana
(intrínseco), se denomina CF0 y permite el paso de los
protones desde el lumen tilacoidal hacia el estroma,
mientras que el dominio estromático (extrínseco),
llamado CF1, está encargado de la síntesis del ATP. El
paso de los protones hacia el estroma (tres H+ por cada
ciclo de síntesis), induce una rotación en el dominio CF0
que se transmite al dominio CF1, provocando en éste
cambios de conformación que permiten la fosforilación
del ADP (Figura 11.5).
11.5. Con la energía producida en la fase lumínica se
lleva a cabo la fase de fijación del CO2 atmosférico.
El objetivo de la fase lumínica de la fotosíntesis es usar la
energía del sol para producir energía química, la cual se
almacena en forma de ATP y NADPH. Con estas
sustancias se logra fijar el carbono atmosférico en
compuestos orgánicos útiles para la planta, que son la
base de la producción primaria -y por tanto de toda la
vida- en este planeta. La ecuación global de este proceso
es:
6 CO2 + 6 H2O + E → C6H12O6 + 6 O2,
donde E representa la energía requerida para el proceso
(2879 kJ·mol-1
). La estequiometría de la ecuación indica
que se requieren 6 moles de dióxido de carbono y seis de
agua para producir una mol de glúcido (hexosa) y seis de
oxígeno molecular. Con estudios usando 14
C se pudo
demostrar que el CO2 se une a una molécula de cinco
carbonos, la ribulosa-1,5-bisfosfato (RuBP), para formar

Capítulo 11: Fotosíntesis
237
una molécula inestable de seis carbonos que rápidamente
se divide para formar dos moléculas estables de tres
carbonos cada una: el ácido 3-fosfoglicérico (AFG) y su
isómero interconvertible, la dihidroxiacetona fosfato
(DHAF). Debe tenerse presente que el CO2 es un gas en
el que el carbono está muy oxidado y en forma no
orgánica, pues no forma parte de compuestos propios de
los seres vivos. A esta forma se le llama carbono
atmosférico y para que pueda incorporarse a los seres
vivos debe estar en una forma reducida en una molécula
de uso biológico. Esta reacción recientemente descrita, en
la que el carbono pasa de ser un gas atmosférico a ser un
componente de una molécula orgánica biológica, se
denomina fijación del carbono. A partir del AFG y la
DHAF se inicia una serie de reacciones enzimáticas que
regeneran la RuBP, mientras que el carbono del CO2
atmosférico es llevado a la molécula en la que quedará
permanentemente fijado. Por ello este proceso fue
denominado fase o ciclo de fijación del carbono (CFC),
ilustrado en la figura 11.6.
Esta fase posee tres pasos claves: 1) fijación del
CO2 mediante la carboxilación de la RuBP, 2) reducción
del carbono a glúcidos y 3) regeneración de la RuBP.
Para que todo esto se verifique, es necesario usar la
energía del ATP y del poder reductor del NADPH
obtenida en la fase lumínica de la fotosíntesis. Dado que
esta fase no requiere luz directa sino la energía química
producida en la fase anterior, se consideró que se
realizaba en la oscuridad y por lo tanto fue llamada
inicialmente fase oscura de la fotosíntesis. En realidad
puede llevarse a cabo con o sin luz, pues sólo necesita la
energía mencionada, por lo que es más correcto
denominarla fase de fijación del carbono, como se
mencionó.
La primera reacción del CFC, que es la
carboxilación de la RuBP, se lleva a cabo por la enzima
ribulosa-1,5-bisfosfato carboxilasa oxigenasa, mejor
conocida como rubisco. Se ha estimado que esta es la
enzima más abundante en la biósfera, dado que las
plantas poseen muchas copias en sus cloroplastos,
probablemente en respuesta al hecho de ser una proteína
de muy baja eficiencia catalítica. Además de esta
proteína hay otras enzimas involucradas en el CFC, el
cual es una ruta bioquímica compleja que consume 6
moléculas de ATP y 6 de NADPH para fijar 3 moléculas
de CO2. Adicionalmente, se consumen 3 moléculas de
ATP en el proceso de regeneración de la RuBP.
Detallando un poco la ecuación de la fotosíntesis
se obtiene:
6 CO2 + 6 H2O + 6RuBP +12 NADPH + 18 ATP →
1 AFG + 1 DHAF + 6 RuBP + 12 NADP+ + 12 H
+ +18
ADP + 16 Pi + 6 O2,
donde se aprecia que la ganancia en términos de carbono
está en las dos moléculas de triosas fosfato (AFG y
DHAF), que se originaron a partir de seis moléculas de
CO2. El oxígeno molecular, que es esencial para la
inmensa mayoría de seres vivos del planeta, es en
realidad un subproducto, un desecho, que se acumuló en
la atmósfera durante miles de millones de años, desde la
aparición de los primeros organismos fotosintéticos
oxigénicos. Esta acumulación llevó los niveles
atmosféricos de dicho gas desde casi cero hasta el actual
21%, convirtiendo nuestra atmósfera en un ambiente
altamente oxidante.
Pero lo que más interesa de esta fase es el
destino de las triosas, las cuales son enviadas al
citoplasma por el mecanismo de antiporte con fosfato
inorgánico, lo que se verifica mediante un traslocador de
fosfato. Allí son usadas para la síntesis de sacarosa,
almidón, aminoácidos esenciales, ácidos grasos e
isoprenoides. Cuando hay exceso de fotosíntesis en el
cloroplasto, es posible procesar tales triosas allí mismo y

Capítulo 11: Fotosíntesis
238
almacenar la energía resultante en forma de almidón en
su interior (Figura portada del capítulo). Desde allí puede
ser luego exportado según las necesidades de la planta.
11.6. La fotorrespiración disminuye la eficiencia
fotosintética.
La rubisco, ribulosa-1,5-bisfosfato carboxilasa oxigenasa,
recibe ese nombre porque además de carboxilar es capaz
de oxigenar, es decir, puede usar O2 como sustrato en el
mismo centro activo que fija el CO2. Como estos gases
compiten por unirse al mismo sitio de la rubisco, se dice
que el oxígeno es un inhibidor competitivo de la enzima.
Cuando es el O2 el que se incorpora, su unión con la
ribulosa-1,5-bisfosfato produce ácido fosfoglicérico
(AFG) y ácido fosfoglicólico, en lugar de las dos triosas
fosfatadas normalmente obtenidas.
El ácido fosfoglicólico posee dos carbonos, por
lo que la utilización de O2 en lugar de CO2 por parte de la
rubisco no implica ganancia de carbono. Luego de esta
oxigenación se dan más reacciones químicas que
conforman un proceso de naturaleza cíclica, ya que se
regenera la molécula de ribulosa-1,5-bisfosfato. Este
complicado proceso consume ATP y poder reductor y se
lleva a cabo en tres organelos: el cloroplasto, el
peroxisoma y la mitocondria. A este proceso se le conoce
en conjunto como el ciclo de la fotorrespiración y la
explicación de su existencia es motivo de amplio debate
en la comunidad científica.
Algunos autores plantean que su función es
servir como vía disipadora de energía en condiciones
adversas para el aparato fotosintético, como cuando hay
alta luminosidad y baja concentración de CO2. La
fotorrespiración deriva su nombre del hecho que depende
de la luz (de ahí el prefijo foto), además de su similitud
con la respiración mitocondrial, ya que consume oxígeno
y genera CO2. Hay que tener en cuenta, sin embargo, que
a diferencia de la respiración que sucede en la
mitocondria, la fotorrespiración consume energía en vez
de producirla (Figura 11.7).
11.7. Las plantas tienen alternativas fotosintéticas:
fotosíntesis C4 y metabolismo ácido de Crasuláceas
(MAC).
La fijación del carbono que se ha descrito se denomina
fotosíntesis C3 porque el primer producto estable es un
compuesto orgánico de tres carbonos, el ácido
fosfoglicérico (AFG). Si bien esa es la ruta más
abundante en el reino vegetal, hay rutas fotosintéticas
alternativas que poseen diferentes mecanismos para
aumentar la concentración del CO2 en la cercanía de la
rubisco, que fueron desarrolladas por plantas que viven
en ambientes cálidos y secos. Se trata de la fotosíntesis
C4 y de la fotosíntesis del metabolismo ácido de
crasuláceas (MAC). La C4 se denomina así porque el
primer producto estable tras la fijación del CO2
atmosférico es un ácido de cuatro carbonos, mientras que
la fotosíntesis MAC fue nombrada por la familia de
plantas en la cual se descubrió: Crassulaceae. Tanto el
metabolismo C4 como el MAC están presentes en
especies que poseen diferencias morfofisiológicas
notables con respecto a las plantas con fotosíntesis C3.
La anatomía del mesófilo en las plantas C4 se
denomina anatomía Kranz (Figura 11.8), la cual consta
de una capa de células muy grandes, denominadas células
de la vaina, que se ubican alrededor de los haces
vasculares foliares. Estas células poseen grandes
cloroplastos, los cuales encuentran adosados a las paredes
externas y son capaces de almacenar almidón. Las
paredes de estas células poseen muchos plasmodesmos
que las comunican con las células del mesófilo. La
variante fisiológica más notable con respecto a la
fotosíntesis C3 se debe a una enzima especial, la
fosfoenolpiruvato carboxilasa (FEPC), que cataliza la

Capítulo 11: Fotosíntesis
239
unión del CO2 atmosférico con una molécula de tres
carbonos, el fosfoenolpiruvato, en las células del
mesófilo. El producto de esta reacción es el oxaloacetato,
el cual es rápidamente convertido a ácido málico o
aspártico (Figura 11.9).
Estos ácidos son transportados a las células de la
vaina del haz, donde son almacenados en grandes
concentraciones, para luego ser descarboxilados
generando un ambiente muy rico en CO2, en el cual la
rubisco inicia el proceso de fijación y reducción del
carbono ya descrito (el ciclo C3). La fotorrespiración es
de esta manera prácticamente abolida, ya que la elevada
concentración de CO2 evita que la rubisco incorpore O2.
Luego de esta descarboxilación, los compuestos de tres
carbonos son regresados a las células del mesófilo donde
regeneran el aceptor inicial, la molécula de
fosfoenolpiruvato, para continuar la captación de dióxido
de carbono.
Las plantas C4 se clasifican de acuerdo a la
enzima que lleva a cabo la descarboxilación, las cuales
son: la enzima málica dependiente de NADP+
(cloroplasto), la fosfoenol piruvato carboxiquinasa
(citoplasma) y la enzima málica dependiente de NAD+
(mitocondria). Al primer tipo pertenecen el maíz, la caña
de azúcar y el sorgo. El metabolismo C4 se ha hallado en
unas 17 familias de angiospermas, pero se sabe de la
existencia de especies con características intermedias C3-
C4, como sucede en algunas especies neotropicales de la
familia Clusiaceae. El mayor costo energético del
metabolismo C4 comparado con las plantas C3 se ve
ampliamente compensado con la casi total anulación de
la fotorrespiración, por lo que en definitiva es más barato
fijar una molécula de CO2 en plantas C4 que en las C3.
Además, las plantas C4 presentan mayor eficiencia en el
uso del nitrógeno y del agua, por lo que se desempeñan
mejor que las C3 en ambientes secos y calurosos, razón
por la cual la mayoría de ellas habita zonas geográficas
con climas semiáridos y tropicales de altas temperaturas.
Las plantas con metabolismo ácido de
crasuláceas (MAC), son generalmente suculentas, con
células ricas en vacuolas llenas de agua y habitan lugares
muy áridos. Fijan el CO2 en ácidos orgánicos al igual que
las plantas C4, pero a diferencia de ellas la separación de
la primera y segunda carboxilaciones es temporal, no
espacial, ya que la primera carboxilación se hace durante
la noche en las células del mesófilo. Esto produce una
acumulación de los ácidos de cuatro carbonos
provenientes de la fijación del CO2, ya que éste no es
usado para sintetizar glúcidos sino hasta el otro día
cuando haya ATP producido por la fase lumínica. Los
ácidos acumulados, principalmente malato, dan un
aumento notable a la acidez a la planta -de ahí el nombre
de esta vía fotosintetizadora- y son transformados en
piruvato por descarboxilación durante el día. Esto genera
un ambiente rico en CO2 en la cercanía de la rubisco
(Figura 11.10). En el día los estomas permanecen
cerrados por lo que no hay pérdida de agua, mientras que
en la noche, cuando se abren para permitir la entrada de
CO2, la transpiración es baja porque la temperatura
ambiental también lo es. Por ello es que el MAC es la
forma más eficiente de hacer fotosíntesis, al menos en
cuanto al uso del agua se refiere.
En ambientes acuáticos, donde el CO2 se
encuentra en solución, se verifica la situación
CO2 + H2O ↔ HCO3- + H
+,
con el equilibrio desplazado hacia la molécula del
bicarbonato (HCO3-). Allí hay plantas con mecanismos
de bombeo de este compuesto que, asociados a la
anhidrasa carbónica, generan una concentración de CO2
óptima alrededor de la rubisco. En el trópico, plantas

Capítulo 11: Fotosíntesis
240
acuáticas como Isoetes y Egeria presentan estos y otros
mecanismos similares de concentración de CO2.
Al igual que hay plantas con características
intermedias C3-C4, hay intermedios C3-MAC. Se cree
que el metabolismo C4 es de reciente aparición entre las
angiospermas, pero como son grupos lejanamente
emparentados, es posible que se trate de un fenómeno
polifilético. El metabolismo MAC está asociado a plantas
más primitivas, por lo que su origen parece más antiguo y
también polifilético. Se puede pensar que las plantas
MAC aparecieron como una respuesta adaptativa a
ambientes áridos o a situaciones de aridez transitorias.
11.8. La fotosíntesis es un proceso regulado, que
puede ser afectado por muchos factores.
El buen funcionamiento de la fotosíntesis depende de la
coordinación de sus dos fases, la lumínica y la de fijación
del carbono. La tasa global de crecimiento de una planta
está regulada por el desempeño de las fases mencionadas
y está directamente relacionada con la tasa de
crecimiento del organismo. El principal factor limitante
de la primera etapa es la luz, mientras que el agua, el
CO2, los nutrientes inorgánicos y la regulación de la
rubisco son los factores claves de la segunda fase. Tanto
los defectos como los excesos de todos estos factores
pueden afectar la tasa fotosintética global (Figura 11.11).
Dado que las plantas no pueden desplazarse para
buscar más luz en condiciones de oscuridad, o para
ocultarse en condiciones de exceso de radiación, han
desarrollado mecanismos para usar óptimamente el
recurso lumínico cuando está escaseando o para disiparlo
cuando está en exceso. El uso adecuado y la disipación
están en un continuo equilibrio dinámico que hace
aumentar el primera y disminuir la segunda y viceversa.
La tasa fotosintética aumenta linealmente con la densidad
de flujo fotónico (DFF), hasta alcanzar una saturación en
la cual el exceso de luz no es utilizable. El punto de
saturación depende de la especie y de las circunstancias
fisiológicas del individuo, pero en todo caso la radiación
sobrante debe ser disipada o dañará la planta, ya que
puede inducir el proceso denominado fotoinhibición, que
lleva a una disminución significativa de la tasa
fotosintética de la planta.
El fotosistema II es el punto más sensible en la
actividad reguladora fotosintética, ya que es capaz de
sufrir mecanismos de desacoplamiento y desactivación
para maximizar la disipación de energía lumínica. Un
mecanismo muy reconocido se debe a un polipéptido
particular del fotosistema II, denominado D1, que es una
proteína con una muy alta tasa de recambio aun en
condiciones normales de luminosidad. Por ser una
molécula que se desactiva para evitarle daños al
fotosistema, se ha comparado su función con la de un
fusible en un sistema eléctrico convencional. También
hay otras formas de desacoplamiento para disipar energía
a nivel de la transferencia de electrones en las antenas,
como en los mecanismos de fluorescencia y en los
centros de reacción, por flujos cíclicos de electrones
alrededor de éstos.
En la fase de fijación y reducción del carbono
(CFC), un aspecto de gran importancia es la actividad de
la rubisco, enzima grande de relativamente poca
eficiencia que puede usar como sustratos tanto al CO2
como al O2. Si hay unas condiciones óptimas y un aporte
normal de CO2, su tasa de asimilación dependerá
principalmente del estado de activación de la rubisco, en
el cual participan el propio CO2 (uniéndose a un lugar
diferente del sitio activo), el ión Mg++
, un pH alto, la
presencia de la enzima rubisco activasa y otros factores
menos claramente entendidos. Tanto el CO2 como la
ribulosa-1,5-bisfosfato son sustratos indispensables para
el funcionamiento de la rubisco, por lo que el exceso de
uno de nada sirve si el otro está en cantidades limitantes.

Capítulo 11: Fotosíntesis
241
Otras enzimas del CFC están también comprometidas en
su regulación, conociéndose que factores como la
reducción del grupo tiol, el pH, la concentración de
magnesio y las concentraciones de los sustratos, ejercen
un papel regulatorio muy importante de la tasa global del
CFC.
La producción de los glúcidos a partir del
carbono fijado en la fotosíntesis experimenta también una
regulación de acuerdo a la tasa fotosintética, lo cual es
particularmente conocido para la sacarosa y el almidón.
En la síntesis de estos productos existen enzimas claves
que son activadas por procesos de fosforilación de
aminoácidos específicos, según la intensidad del CFC.
11.9. Los productos de la fotosíntesis tienen diversos
destinos.
Las sustancias producidas por la fotosíntesis, llamados
fotosintatos o fotoasimilados, pueden ser usados para
diversos procesos fisiológicos según las circunstancias
celulares. En primera instancia, se usan para la
respiración celular y para la construcción de sustancias
necesarias para la propia célula, como 3-fosfoglicerato,
aminoácidos, azúcares y otros productos. Otro posible
destino de los fotoasimilados es su almacenamiento
dentro del mismo cloroplasto (Figura portada del
capítulo), lo cual sucede cuando hay exceso de
producción. Un tercer destino de los fotosintatos es su
transporte a larga distancia, desde los órganos
productores (fuentes) hasta los órganos que los usan o
almacenan (sumideros). La distancia entre fuentes y
sumideros varía según la especie, pudiendo ser de pocos
centímetros en hierbas como las orquídeas miniaturas
cuyo cuerpo completo no supera los 20 cm de longitud
(ej: Pleurothallis spp.), o de muchos metros en grandes
árboles maderables (ej: Nectandra spp.).
Los estudios realizados para tratar de entender el
transporte por el floema a larga distancia, llevaron a
comprender que la difusión simple no podía ser la
responsable de este fenómeno, requiriéndose de una
explicación fisiológica alterna, conocida actualmente
como la hipótesis de Münch (sección 12.11).
11.10. Para el transporte a larga distancia de los
fotoasimilados y otras sustancias se requiere un tejido
especializado.
El floema es un tejido encargado de la conducción de
sustancias, que se encuentra generalmente más externo
que el xilema en los tallos. Su presencia fue notada desde
hace mucho tiempo aunque su papel en la conducción de
sustancias orgánicas fue reconocido sólo hasta mediados
del siglo XX. Los estudios pioneros se basaron en el
anillamiento, técnica que consiste en retirar la corteza del
tallo a cierta altura y observar los cambios. La
consecuencia más notoria es la acumulación de
fotosintatos en la parte superior del anillo
(hinchamiento), y la disminución de ellos en la parte
inferior (agotamiento). Esto demuestra que el transporte
basípeto en el tallo se produce en la corteza. Los estudios
posteriores que usaron marcadores radiactivos
demostraron que unas células especializadas
denominadas elementos cribosos, son las encargadas de
transportar los fotosintatos en la planta.
El floema es un sistema conductor continuo
(Figura 11.12), constituido por células vivas en estado
funcional, el cual se extiende por todo el cuerpo vegetal y
llega hasta unas pocas células de distancia de los ápices
de la raíz y el tallo. La composición del floema es
diferente entre las plantas con flores (angiospermas) y las
plantas sin flores (gimnospermas). En estas últimas la
unidad estructural y funcional es la célula cribosa,
mientras que en las angiospermas es el elemento criboso,
el cual se une por los extremos con otros elementos

Capítulo 11: Fotosíntesis
242
cribosos alineados longitudinalmente formando el tubo
criboso. Las paredes terminales de los elementos cribosos
poseen perforaciones mayores que los plasmodesmos,
que en conjunto reciben el nombre de placa cribosa. Las
células cribosas de las angiospermas no están alineadas
longitudinalmente y las interconexiones se dan mediante
áreas cribosas ubicadas en las paredes laterales. Una
descripción más completa de la composición del floema
la encuentra en la sección 5.6.2.
Existen muchas variaciones en la composición
del fluido floemático entre especies, tanto cuantitativas
como cualitativas. Esto sucede porque son numerosas las
variables que pueden afectarlo, tales como la tasa
fotosintética, el órgano de la planta, el grado de
desarrollo y las condiciones ambientales. Algunas
características generales relativamente comunes a
cualquier fluido floemático son el pH alcalino (ca. 8), la
alta viscosidad, el potencial osmótico negativo (-1 MPa a
-2 MPa) y la abundancia de sustancias disueltas de bajo
peso molecular. Los azúcares provenientes de la
fotosíntesis son el mayor componente de los solutos del
floema, siendo la sacarosa el azúcar más abundante.
También se transportan aminoácidos, nucleótidos y son
abundantes el potasio, el ión cloruro y el fosfato e incluso
se sabe que algunos virus son capaces de movilizarse por
el floema. La forma en que las sustancias son cargadas y
movilizadas por el floema es explicada en detalle en la
sección 12.11.
BIBLIOGRAFÍA
Azcón-Bieto J & Talón M. 1993. Fisiología y
bioquímica vegetal. Interamericana & McGraw-Hill,
Nueva York, E.U.A.
Azcón-Bieto J & Talón M. 2000. Fundamentos de
fisiología vegetal. McGraw-Hill Interamericana, Barce-
lona, España.
Bowes G, Rao SK, Estavillo GM & Reiskind JB. 2002.
C4 mechanisms in aquatic angiosperms: Comparisons
with terrestrial C4 systems. Functional Plant Biology 29:
379-392.
Ellis RJ. 1979. The most abundant protein in the world.
Trends in Biochemical Science 4: 241-244.
Ghanotakis DF & Yocum CF. 1990. Photosystem II
and the oxygen-evolving complex. Annual Review of
Plant Physiology and Plant Molecular Biology 41: 255-
276.
Govindjee & Coleman WJ. 1990. How plants make
oxygen. Scientific American 50-58.
Hall DO & Rao KK. 1994. Photosynthesis, 5 ed.
Cambridge University Press, Cambridge, Inglaterra.
Hatch MD & Slack CR. 1966. Photosynthesis by
sugarcane leaves. A new carboxylation reaction and the
pathway of sugar formation. Biochemical Journal 101:
103-111.
Rost TL, Barbour MG, Thornton RM, Weier TE &
Stocking CR. 1985. Botánica. Introducción a la biología
vegetal. Editorial Limusa, México DF, México.
Taiz L & Zeiger E. 1998. Plant physiology, 2 ed.
Sinauer Associates, Inc., Publishers, Sunderland, E.U.A.
Van-Bel AJE. 1993. Strategies of phloem loading.
Annual Review of Plant Physiology and Plant Molecular
Biology 44: 253-281.

Capítulo 11: Fotosíntesis
243
Figura de la portada del capítulo
Figura 11.1. Espectro de absorción de las clorofilas a y b. En general, la luz verde y amarilla es poco usada por las plantas,
si bien los pigmentos accesorios ayudan a aumentar la eficiencia en la captación de fotones por parte del cloroplasto.
Gráfica en dominio público por cortesía de un autor anónimo <<Kurzon>>, a través de Wikipedia Commons.

Capítulo 11: Fotosíntesis
244
Figura 11.2. Pigmentos encontrados en las plantas. A) Los carotenoides son moléculas que absorben luz y actúan
complementariamente con las clorofilas, B) clorofila a. Una de las características más notables de los pigmentos es la
presencia de dobles enlaces alternos.
Figura 11.3. Ampliación de una fotografía electrónica de transmisión de una célula vegetal mostrando el cloroplasto con
tilacoides apilados formando los granas. En estas estructuras membranales es que se ubican los fotosistemas y por lo tanto
es el sitio exacto de la fotosíntesis. Foto en dominio público por cortesía de Dartmouth Electron Microscope Facility,
Dartmouth College.

Capítulo 11: Fotosíntesis
245
Figura 11.4. Esquema de la disposición de los fotosistemas (FS) y las moléculas involucradas en el transporte de electrones
en la membrana tilacoidal del cloroplasto. También llamado esquema Z. A) Plastoquinona, B) complejo de citocromos, C)
plastocianina, D) proteína ferrosulfurada, E) ferredoxina, F) NADP+ reductasa, la enzima que transfiere los electrones al
NADP+ para producir NADPH. El flujo de electrones por estas moléculas produce una acumulación de protones en el
interior del tilacoide, lo que hace de éste un espacio ácido (con pH bajo). Este gradiente de protones es usado por la ATP
sintasa para generar ATP.

Capítulo 11: Fotosíntesis
246
Figura 11.5. Representación esquemática de la ATP sintasa, la cual está compuesta por dos subunidades. A) La subunidad
CF0 está embebida en la membrana tilacoidal y a través de ella pasan los protones hacia el estroma, B) la subunidad CF1 se
encuentra en el estroma y es la encargada de acoplar la fosforilación del ADP para producir ATP.

Capítulo 11: Fotosíntesis
247
Figura 11.6. Ciclo de fijación del carbono. Tres moléculas de CO2 se unen a tres moléculas de ribulosa 1,5-bisfosfato y
producen seis moléculas de 3-fosfoglicerato. Luego seis ATP son usados para producir seis moléculas de 1,3-
bisfosfoglicerato, las cuales generan seis moléculas de gliceraldehído 3-fosfato mediante el uso de seis moléculas de
NADPH. Una molécula de gliceraldehído 3-fosfato se usa para la síntesis orgánica, mientras que las restantes cinco se usan
para regenerar las tres moléculas de ribulosa 1,5-bisfosfato con las que se reinicia el ciclo. Así, tres moléculas de CO2
fijadas producen una molécula de gliceraldehído 3-fosfato, con un costo neto de 9 ATP y 6 NADPH, los cuales fueron
producidos durante la fase lumínica de la fotosíntesis.

Capítulo 11: Fotosíntesis
248
Figura 11.7. En la fotorrespiración la ribulosa 1,5-bisfosfato se une al oxígeno y genera una molécula de 3-fosfoglicerato y
una de 2-fosfoglicolato, un compuesto de dos carbonos. Esta molécula pasa del cloroplasto al peroxisoma donde genera
glicina que a su vez es exportada a la mitocondria donde se genera serina. Esta molécula pasa al peroxisoma y genera
glicerato, que entra al cloroplasto y produce 3-fosfoglicerato que entra al ciclo de regeneración de la ribulosa 1,5-bisfosfato.
Todo este proceso consume O2, ATP y NADH a la vez que genera CO2, por lo que fue llamado fotorrespiración. A)
Reacciones en el cloroplasto, B) reacciones en el peroxisoma, C) reacciones en la mitocondria.

Capítulo 11: Fotosíntesis
249
Figura 11.8. Corte transversal de hoja de maíz en la que se nota la anatomía Kranz. A) Haz vascular, B) células grandes de
la vaina del haz con cloroplastos gigantes, C) cutícula, capa de sustancias hidrófobas depositada sobre la pared externa de
las células de la epidermis. Foto cortesía de X. Marquínez.

Capítulo 11: Fotosíntesis
250
Figura 11.9. En la fotosíntesis C4 la enzima fosfoenolpiruvato carboxilasa (FEPC), cataliza la unión del CO2 al
fosfoenolpiruvato para producir oxaloacetato, el cual es rápidamente convertido a ácido málico o aspártico. Estos ácidos de
cuatro carbonos son llevados a la célula de la vaina del haz donde se descarboxilan generando un ambiente rico en CO2 para
la rubisco, así el ciclo de fijación del carbono (CFC) es más eficiente. Los compuestos C3 son devueltos a la célula del
mesófilo donde regeneran fosfoenolpiruvato. A) Célula del mesó-filo, B) célula de la vaina del haz vascular.

Capítulo 11: Fotosíntesis
251
Figura 11.10. El metabolismo ácido de crasuláceas (MAC), es un mecanismo de concentración de CO2 presente en plantas
suculentas que viven en ambientes secos y soleados. La estrategia es fijar el carbono de noche, cuando al abrir los estomas
se pierde poco agua debido a la baja temperatura, almacenándolo en ácidos de cuatro carbonos dentro de la vacuola.
Durante el día esos ácidos se descarboxilan para aumentar la concentración de CO2 en las cercanías de la rubisco,
permitiendo la fijación definitiva del carbono a través del ciclo de fijación del carbono (CFC).
Capítulo 11: Fotosíntesis
252
Figura 11.11. Varios factores ambientales influyen directamente en la tasa fosotintética de las plantas. La cantidad y
calidad de luz incidente, la temperatura del aparato fotosintético, la tasa CO2/O2 y las condiciones intracelulares como la
necesidad de los fotosintatos y la actividad de la rubisco o de otros componentes individuales de la fase lumínica o del CFC,
están entre los factores que más influyen al proceso fotosintético.
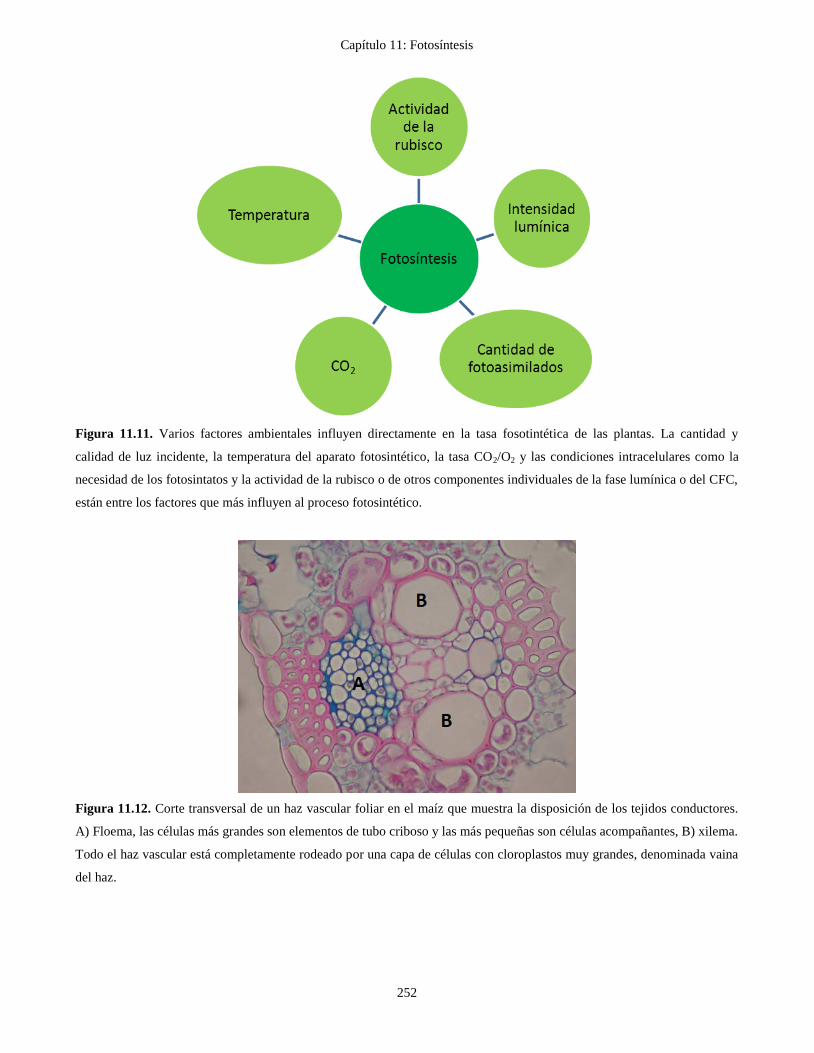
Figura 11.12. Corte transversal de un haz vascular foliar en el maíz que muestra la disposición de los tejidos conductores.
A) Floema, las células más grandes son elementos de tubo criboso y las más pequeñas son células acompañantes, B) xilema.
Todo el haz vascular está completamente rodeado por una capa de células con cloroplastos muy grandes, denominada vaina
del haz.





























