Embed Size (px)
Citation preview

Centre for Agrobiological Research (CABO), Wageningen, The Netherlands
Catalase and Peroxidase in Primary Bean Leaves during Development and Senescence
JOHANNA M. BRABER
With 2 figures
Received August 22, 1979 . Accepted September 11, 1979
Summary
Catalase and peroxidase activity and isoenzyme composition are assayed in extracts of the developing and senescing primary leaves of Phaseolus vulgaris which are also analysed for their chlorophyll content, organic nitrogen content and soluble protein content. Photosynthetic rate and dark respiration rate are measured in leaves of intact plants. Catalase activity level is highest in the young mature leaf, when also peaks are observed in photosynthetic rate level, dark respiration rate level, chlorophyll content, organic nitrogen content and soluble protein content. Peroxidase activity level gradually increases during development and senescence until the leaves turn brown.
The number of anodic peroxidase isoenzymes increases from two in the young growing leaf to four in the senescent leaf. Three cathodic peroxidases were detected in mature and senescent leaves which were most intense at senescence. Only one band with catalase activity was detected which was most intense in the young mature leaf.
Catalase and peroxidase activity level in primary bean leaves show an inverse relationship during development and senescence. It is hypothesized from this, from the molecular structure of both enzymes and from the in vitro relationship between catalase and peroxidase activity that the tetrameric molecules of catalase in vivo might des integrate into monomeric units with peroxidase activity.
Key words: catalase, peroxidase, senescence, Phaseolus vulgaris L.
Introduction
The activity levd of the heme protein enzymes catalase and peroxidase and the peroxidase isoenzyme composition has been related to leaf senescence (PATRA et a!., 1978; PARISH, 1968; LAZAR and FARKAS, 1970). The biochemical function of both enzymes and the role in senescence is not clear.
Catalase which is predominantly found at the peroxisom~s is thought to convert the hydrogen peroxide evolving in photorespiration (HALLIWELL, 1977). The catalase activity level is species specific with respect to the occurrence of photorespiration and is higher in leaves of plants with a Cl-type of photosynthesis compued to C-plants,
z. Pjlanzenphysiol. Bd. 97. S. 135-144. 1986.

136 JOHANNA M. BRABER
the difference being more pronounced in monocotyledons than in dicotyledons (PATRA et aI., 1978). The activity level shows a maximum during development and senescence in mature leaves of Oryza sativa (PATRA et aI., 1978) and the course follows that of the photo respiration rate level in Glycine max and Phaseolus vulgaris (MULCHI et aI., 1971; PEET et aI., 1977) and more general in C 3-plants (SALIN and
HOMANN, 1971; FAIR et aI., 1973). Plant catalase is an unstable enzyme in aqueous solution with a molecular weight of ca. 225,000 for catalase from Cucumis sativus (LAMB et aI., 1978) and Lens culinaris (SCHIEFER et aI., 1976) and four similar
subunits of ca. 54,000 (SCHONBAUM and CHANCE, 1976). The peroxidase activity level and the number of isoenzymes increases during leaf
senescence (BATES and CHANT, 1970; LAZAR and FARKAS, 1970). Same changes in leaf peroxidase are observed following virus-infection (FARKAS and STAHMANN, 1966; CHANT, 1967; SOLYMOSY et aI., 1967; NOVACKY and HAMPTON, 1968), which has
been interpreted as an involvement of peroxidases in a defense-mechanism towards palthogens (FRIC, 1976). In vitro many phenols and aromatic rings which occur
naturally in plant tissue can serve as a hydrogen donor. The compounds which are converted in vivo by peroxidases are not known, neither is the source of the substrate hydrogen peroxide. Plant peroxidase is a stable enzyme occurring in up to 20 isoenzymes (DELINCEE and RADOLA, 1970), well soluble in aqueous medium and present in the whole celL The molecular weight varies from 40,000 (SAUNDERS et aI., 1964), 55,000 (PAUL, 1963) to 65,000 (KOKKINAKIS and BROOKS, 1979).
Peroxidase contains protohematin IX as a prosthetic group, as does one catalase subunit (PAUL, 1963; SCHONBAUM and CHANCE, 1976). Catalase from beef liver reversibly can be split into monomeric subunits with peroxidatic activity at alkali treatment with loss of catalatic activity (SAMEJIMA et aI., 1967; MARKLUND, 1973).
The poorly understood biochemical function of peroxidases (FRIC, 1976), the in vitro relationship between catalatic and peroxidatic activity and the molecular
structure of both enzymes suggest that plant catalase and plant peroxidase are related to each other in vivo in such a way that catalase degradation at senescence can lead to subunits with peroxidase activity.
In this article the results are presented of analysis of developing and senescing primary leaves of the C 3-plant Phaseolus vulgaris. The change in activity levels of peroxidase and catalase and the isoenzyme composition is discussed in relation to the photosynthetic rate, the dark respiration rate, the chlorophyll content, the orgamc nitrogen content and the soluble protein content and in relation to each other.
Materials and Methods
Experimental plants Seeds of brown bean plants (PhaseDlus vulgaris L. var. BERNA) were sown in Perlite.
Eight-day seedlings were transplanted to aerated Hoagland nutrient solution. The plants were grown in the phytotron at 17 h light period (HPL 56 J . m-2 • sec-I), 20°C and a relative humidity of 70 to 80 Ofo. Primary leaves were sampled at successive stages of devel-
Z. PJlanzenphysiDI. Bd. 97. S. 135-144. 1980.

Catalase and peroxidase in primary bean leaves 137
opment at intervals of 1 to 4 days, at two hours after the start of the light period. The leaf age is expressed in days after sowing. Each time two replicates were sampled, each sample consisting of 3 to 8 plants (6 to 16 primary leaves) depending on leaf size. Only one replicate was assayed in the catalase activity test. At an age of ca. 25 days the leaves turned yellow, after ca. 30 days they were yellow for 50 % and after 35 days the leaves were yellow to brown or shed.
Extraction of fresh leaves
100 grams of fresh primary leaves were cut into pieces of 1 cm2• 10 grams of this material were homogenized in a Braun mixer in 50 ml 0.1 mol· I-I phosphate buffer pH 7.8 for 1 to 2 minutes. The suspension was filtered through a double layer of cheese cloth and centrifuged in a MSE-25 type centrifuge at 110,000 g for 45 minutes and the supernatant was used for analysis, either immediately or after storage in the freezer.
Acetone powder
Leaf samples for electrophoresis experiments were conserved as acetone powder according to the method described by CLEMENTS (1965) and stored in a freezer after drying.
Extraction of acetone powder
0.5 grams of the powder were extracted in a prechilled mortar and pestle at 4 °C with 10 ml cold 0.1 mol· I-I Tris-buffer pH 7.8 containing 17 Ofo sucrose, 0.1 Ofo ascorbic acid and 0.1 % cysteine-HC!. The extract was transferred to a centrifuge tube with an additional amount of 10 ml buffer and centrifuged at 10,000 g in a MSE-17 type centrifuge for 30 minutes. A 10 ml-portion of the supernatant was further centrifuged at 110,000 g for 45 minutes in a MSE-25 type centrifuge. The supernatant was used for electrophoresis experiments.
Leaf analysis
Total soluble protein content was assayed in fresh leaf extracts using the Folin method (LOWRY et a!., 1951) with bovine serum albumine as a reference. Total nitrogen content was determined in oven dried material (105°C) with the Kjeldahl destruction method with added salicylic acid. Nitrate nitrogen content was analysed using an Orion ionselective nitrate electrode. Organic nitrogen content was calculated as the difference between total nitrogen and nitrate nitrogen. Chlorophyll content was assayed in fresh leaves after ARNON (1949). Leaf area was determined with an automatic area meter model AAM 5 (Hayashi Denko Co.). Dry weight was determined by loss of weight after drying at 105°C. The rates of photosynthesis and dark respiration were determined at 25°C with intact plants as described by LOUWERSE and VAN OORSCHOT (1969).
Enzyme activities
Peroxidase activity was assayed at 25°C in a 1 cm-cuvette in a reaction mixture containing 1.7 ml 0.07 mol· I-I phosphate buffer pH 5.9, 1.0 ml 0.1 mol· 1-1 guaiacol, 0.1 ml fresh leaf extract (well diluted) and 0.2 ml 1 Ofo hydrogen peroxide solution. The absorbancy increases at 420 nm were followed with a Pye Unicam SP 1800 recording spectrophotometer. The blank consisted of the same mixture without H 20 2 •
Peroxidase activity is expressed as conversion of guaiacol or hydrogen peroxide per minute per leaf calculated from the amount of tetraguaiacol, the coloured reaction product developed under the ~.ssay conditions. One mol tetraguaiacol developed corresponds with the conversion of four mol guaiacol and four mol hydrogen peroxide. Peroxidase activity is calculated from the absorbancy changes with a molecular extinction coefficient of tetraguaiacol of 26 cm2 • ,Ilmol-I (MAEHLY and CHANCE, 1959; PUTTER, 1970).
Z. Pflanzenphysiol. Bd. 97. S. 135-144. 1980.

138 JOHANNA M. BRABER
Catalase activity was assayed by a modified method of PATRA et al. (1978). 0.2 ml freshly prepared extract was added to 10 ml 0.035 % H 20 2 and 3 ml extraction buffer at 25°C. The reaction was stopped by adding 10 ml 2 Ofo H 2S04 after 1 to 10 minutes. The residual H 20 2
was titrated with 0.01 mol· 1-1 KMn04• The blank consisted of the reaction mixture in which the extract was added to the acidified solution. Catalase activity was expressed as mmol H 20 2 used per minute per leaf.
Polyacrylamide gelelectrophoresis
Acetone powder extracts were analysed with disc electrophoresis in a Shandon apparatus (DAVIS and ORNSTEIN, 1961). 0.1 ml extract was placed on top of a 71/ 2 Ofo separation gel with 21/2 Ofo concentration gel. Bromophenolblue was used a front marker. The elution buffer was Tris-buffer pH 8.6. The applied current was 41/ 2 rnA per tube. When the top reservoir was connected with the negative pole, the anodic isoenzymes were separated in the gel. The cathodic isoenzymes were separated by reversing the current. The protein bands were stained with Amidoblack. Isoenzymcs of peroxidase were detected by immersing the gels in phosphate buffer pH 5.3 and then in an acidic H 20 2-benzidine mixture (SAFONOVA et aI., 1970). Catalase activity was made visible in the peroxidase test by development of dioxygen bubbles in the gel.
Results
1. Photosynthetic rate, dark respiration rate, leaf area and contents of chlorophyll, organic nitrogen, soluble protein and dry matter
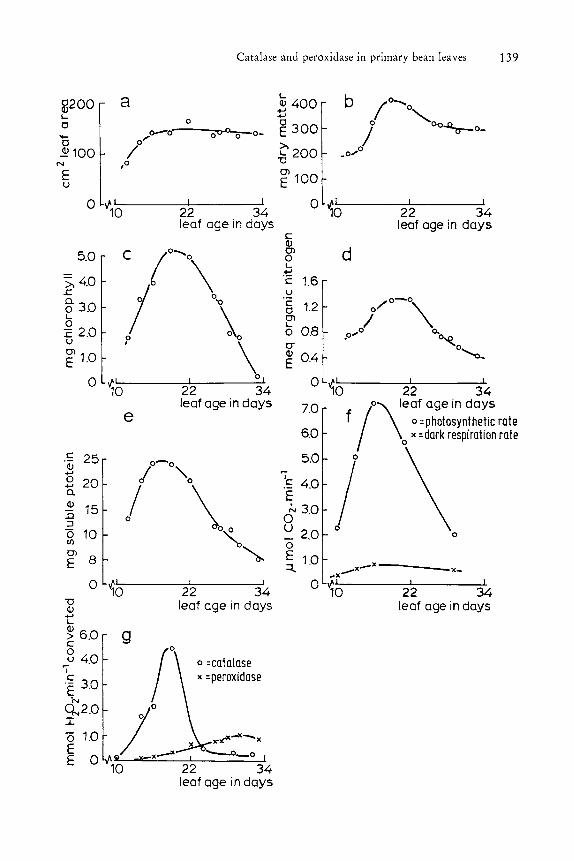
The results of the analysis are presented in figure 1. Photosynthetic rate, dark respiration rate, chlorophyll content, soluble protein content and organic nitrogen content show maxima at a leaf age of 16 to 20 days. At that time the leaf is fully grown, leaf area and dry matter content do not increase further. At the same time the first trifoliar leaf starts to develop and the primary leaf changes its function to a material exporting organ. At a leaf age of 30 days all parameters, except for the dark respiration rate, have declined more than 50 0/0. The soluble protein content decreases from about 30 Ofo to some 10 Ofo of the total amount of organic nitrogen in the period of a leaf age of 14 to 35 days.
2. Catalase and peroxidase activities
The enzyme activities were assayed in fresh extracts of leaves of an age of 10 to 33 days. The results are presented in figure 1. Maximum catalase activity is observed between 16 and 20 days with a sharp decrease between 20 and 24 days. Peroxidase activity level increases gradually during the whole life cycle of the leaf with a decrease at the final stage and is at 30 days five times higher than Ithe activity at 16 days and increases more than the soluble protein content decreases.
Fig. 1: Analysis of primary bean leaves of 10 to 33 days after sowing. a) leaf area. b) dry matter content. c) chlorophyll content. d) organic nitrogen content. e) soluble protein content. f) photosynthetic rate and dark respiration rate. g) catalase activity and peroxidase activity.
Z. PJlanzenphysiol. Bd. 97. S. 135-144. 1980.
~200 L-a -a ~100
N
E u
5.0
>. 4.0 .s:: c. o 3.0 L-0 :c 2.0 u Ol E 1.0
0
.S: 25 Q) +'
.8 20 c. Q)
15 ::c ::J (5 III
10 Ol
8 E
0 "C 111 +' L-
~ 6.0 e:
84.0 I e: .- 30 E . N
Q..,2.o I
'0 1.0
Catalase and peroxidase in primary bean leaves 139
a o
~ 0"
/ ,0
I I
22 34 leaf age in days
C (-\
j ~ o ~ ,
\ ~O
1 0 1
22 34 leaf age in days
e
0-0
I '\.0
I \~ 'b.
~O 1 1
22 34 leaf age in days
g
/ o =cataiase x =peroxidase
... X"' .... -x
L-111400 ~
~ 300
~200 "C
E100
o ~O'-I ----22'-1 ----'314 leaf age in days
e: Q) Ol 0 L-:!:
1.6 e: u e: 1.2 a Ol
5 0.8 c-~ 0.4
0
7.0
6.0
5.0
Ie: 4.0 'E .:.. 30 o .
U _ 2.0 0
~1.o 0
d
o\eaf age in days flo = photosynthetic rate x =dark respiration rate o
/ o
x-x---__ -x_ ~x- , I
22 34 leaf age in days
J' E o "'iii lIo::X--" 1 E
10 22 34 leaf age in days
140 JOHANNA M. BRABER
3. Polyacrylamide gelelectrophoresis
Acetone powder extracts of leaves of different ages were analysed for their protein composition and isoenzyme pa~tern of peroxidase and catalase.
Protein composition
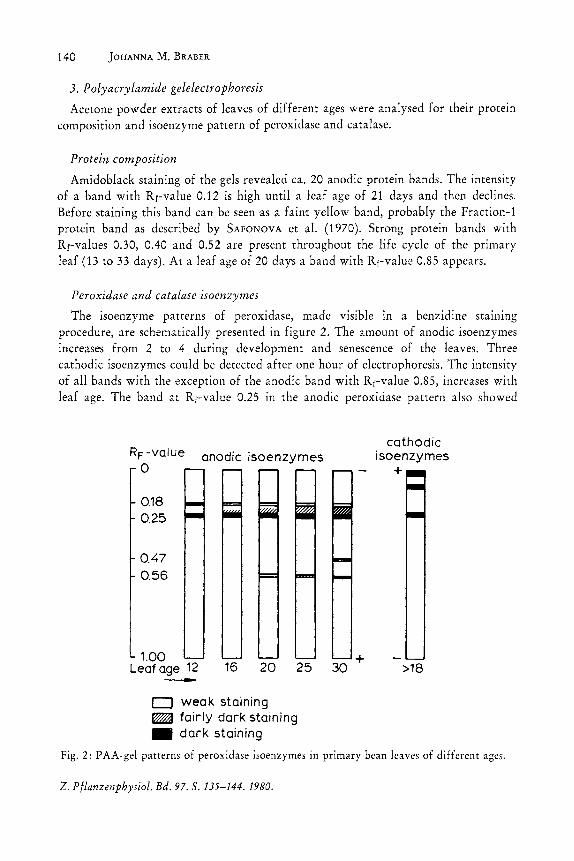
Amidoblack staining of the gels revealed ca. 20 anodic protein bands. The intensity of a band with Rf-value 0.12 is high until a leaf age of 21 days and then declines. Before staining this band can be seen as a faim yellow band, probably the Fraction-l protein band as described by SAFONOVA et al. (1970). Strong protein bands with Rr-values 0.30, 0.40 and 0.52 are present throughout the life cycle of the primary leaf (13 to 33 days). At a leafage of 20 days a band with Rf-value 0.85 appears.
Peroxidase and catalase isoenzymes
The isoenzyme patterns of peroxidase, made visible in a benzidine stammg procedure, are schematically presented in figure 2. The amount of anodic isoenzymes increases from 2 to 4 during development and senescence of the leaves. Three cathodic isoenzymes could be detected after one hour of electrophoresis. The intensity of all bands with the exception of the anodic band with Rr-value 0.85, increases with leaf age. The band at Rf-value 0.25 in the anodic peroxidase pattern also showed
RF -value anodic isoenzymes o
0.18 0.25
0.47 0.56
1.00 Leaf age 12 --- 16 20
o weak staining
25
~ fairly dark staining _ dark staining
+ 30
cathodic isoenzymes
+
>18
Fig. 2: PAA-gel patterns of peroxidase isoenzymes in primary bean leaves of different ages.
Z. Pjlanzenphysiol. Bd. 97. S. 135-144. 1980.

Catalase and peroxidase in primary bean leaves 141
catalase activity. This was most intense at a leaf age of 15 to 21 days and could not be detected after a leaf age of about 30 days. No other bands with catalase activity were revealed.
Discussion
The electrophoresis experiments show an increasing activity level and number of isoenzymes of peroxidase at detection with benzidine in primary bean leaves during development and senescence, while the total amount of soluble protein decreases. The Amidoblack staining shows a complicated pattern in which the disappearance of the band with Rf-value 0.12, probably Fraction-l protein, predominates. The peroxidase isoenzyme pattern shows the greatest change after the leaf is fully grown. The p;:ttern is similar to that described by BATES and CHANT (1970) at detection with guaiacol and quinol, and by RACUSEN and FOOTE (1966) in bean leaves, and by SHAIO-LIM, CHEN et al. (1970) at senescence in Xanthium leaves with several substrates. Although more isoenzymes are detected with benzidine than with guaiacol, all guaiacol bands are detected with benzidine. The increase in number of isoenzymes at senescence detected with benzidine is thus in agreement with the increase in peroxidase activity level assayed with guaiacol.
The change in intensity of the catalase band is in agreement with the catalase activity level at development and senescence. Plant catalase often shows only one enzyme band which differs in IEP or Rf-value, depending on the source (SCANDALIOS, 1965; RACUSEN and FOOTE, 1966; SCHIEFER et aI., 1976; LAMB et aI., 1978).
The par.allel course of the catalase activity level and essential leaf functions like photosynthetic rate and dark respiration rate (Fig. 1) and photorespiration rate in developing leaves of Ca-plants (MULCHI et aI., 1971; SALIN and HOMANN, 1971; PEET et aI., 1977) suggest an important role for this enzyme in the well functioning of a leaf, most probably in photorespiration.
A comparison of the possible amount of hydrogen peroxide developed in photorespiration and peroxidase and catalase activities in vitro shows that the total conversion capacity of the enzymes for hydrogen peroxide during development and senescence is at any time much higher than the expected maximum amount of hydrogen peroxide developed in the leaf which can be estimated from the photosynthetic rate. Photorespiration may produce CO2 at a rate up to 50 % of the net CO2 uptake during photosynthesis (ANDREWS and LORIMER, 1978).
At a leaf age of 16 days peroxidase and catalase activities are 0.20 and 2.16 mmol H 20 2 per minute per leaf, respectively. Photosynthetic rate is 0.0072 mmol CO2 per minute per leaf and the maximum photo respiration rate thus 0.0036 mmol O2 per minute per leaf. The overcapacity for peroxidase activity is 60-fold and for catalase activity 700-fold, assuming that the enzyme activities are of the same level in vivo as in vitro. At a leaf age of 30 days the overcapacity is even higher. Other hydrogen
z. P/lanzenphysiol. Bd. 97. S. 135-144. 1980.

142 JOHANNA M. BRABER
peroxide generating processes in leaves cannot be considered quantitatively because of
lack of information. Peroxidase activity is generally not related to photo respiration and because the
source of Ithe hydrogen peroxide for the peroxidase reaction is not known, it would be interesting to relate the capacity to the hydrogen donor. Because the nature, source and relative extent of different donors are not well-known, it is not possible to make a calculation which could contribute to the comprehension of the biochemical function of peroxidase in the leaf. There is an inverse relationship between the activities of catalase and peroxidase in primary bean leaves during development and senescence. Catalase is present as a single enzyme, peroxidase shows a multiple isoenzyme pattern. From this, from the molecular structure of both enzymes and from the in vitro relationship between catalatic and peroxidatic activity it can be hypothesized that the tetrameric catalase molecules in vivo degrade into monomeric molecules with peroxidase activity.
The peroxidase activity level in the bean leaf still increases when the catalase activity has dropped to a low level. This can be explained by a degradation via dimeric molecules lacking catalase activity (SUND et al., 1967) and peroxidase activity, because plant peroxidases are not known to be dimeric.
The hypothesis is in agreement with the observation of an increased rate of appearance of isoenzymes of peroxidase in virus-infected leaves which are also developed at senescence in healthy plants. The synthesis of peroxidases can be explained as a result of the virus, successfully attacking the respiration system of the plant cell and inducing an increased degradation of catah:se, instead of an active plant defense mechanism.
The hypothesis of catalase degradation into peroxidase is not easily experimentally confirmed. One of the main arguments lies in ,the amino acid sequence of the protein parts of the isoenzymes of peroxidase and the subunits of catalase in the same plant. Plant catalases have hardly been studied for their amino acid sequence because of isolation problems.
Study of the relation between catalase and peroxidase from this point of view could provide a better comprehension of the biochemical function of both enzymes.
Acknowledgements
The author wishes to thank Mr. W. LOUWERSE for the data from photosynthesis experiments, and Mr. G. W. VAN DEN BORN for technical assistance.
References
ANDREWS, T. ]. and G. H. LORIMER: Photorespiration-still unavoidable? FEBS Lett. 90, 1-9 (1978).
ARNON, D. I.: Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physio!. 24,1-15 (1949).
Z. PJlanzenphysiol. Bd. 97. S. 135-144. 1980.

Catalase and peroxidase in primary bean leaves 143
BATES, P. C. and S. R. CHANT: Alterations in peroxidase activity and peroxidase isoenezymes in virus-infected plants. Ann. Appl. BioI. 65, 105-110 (1970).
CHANT, S. R.: Respiration rates and peroxidase activity in virus-infected Phaseolus vulgaris. Experientia (Basel) 23, 676-677 (1967).
CHEN SHAIO-LIM, L. R. TOWILL, and J. R. LOEWENBERG: Isoenzyme patterns in developing Xanthium leaves. Physiol. Plant. 23, 434-443 (1970).
CLEMENTS, R. L.: Fruit proteins: extraction and electrophoresis. Anal. Biochem. 13, 390-401 (1965).
DAVIS, B. J. and L. ORNSTEIN: Disc electrophoresis. Distillation products industries, Rochester, USA, 1961.
DELlNcEE, H. and B. J. RADOLA: Thin-layer isoelectric focusing on Sephadex layers of horseradish peroxidase. Biochim. Biophys. Acta 200, 404-407 (1970).
DEKoCK, P. C., A. HALL, and R. H. E. INKSON: A study of peroxidase and catalase distribution in the potatoe tuber, Ann. Bot. 43, 295-298 (1979).
FAIR, P., J. TEW, and C. F. CRESSWELL: Enzyme activities associated with carbondioxide exchange in illuminated leaves of Hordeum vulgare L. II. Effects of external concentration of carbondioxide and oxygen. Ann. Bot. 37, 1035-1039 (1973).
FARKAS, G. L. and M. A. STAHMANN: On the nature of changes in peroxidase isoenzymes in bean leaves infected by southern bean mosaic virus. Phytopathology 56, 669-677 (1966).
FRU'::, F.: Oxidative enzymes. In: PIRSON, A. and M. H. ZIMMERMANN (Ed.): Encyclopedia of plant physiology, New Series, Vol. 4: Physiological plant pathology, 617-631. Springer Verlag, New York, 1976.
HALLIWELL, B.: A personal assessment of the current state of knowledge of photorespiration and C4-metabolism. In: HALL, D.O., J. COOMBS, and T. W. GOODWIN (Eds.): Proceedings of the 4th international congress on photosynthesis, 347-355. The biochemical society, London, 1978.
KOKKINAKIS, D. M. and J. L. BROOKS: Tomato peroxidase. Purification, characterization and catalytic properties. Plant Physiol. 63, 93-99 (1979).
LAMB, J. E., H. RIEZMAN, W. M. BECKER, and C. J. LEAVER: Regulation of glyoxysomal enzymes during germination of cucumber. Biochim. Biophys. Acta 159, 429-439 (1978).
LAZAR, G. and G. L. FARKAS: Patterns of enzyme changes during leaf senescence. Acta BioI. Acad. Sci. Hung. 21, 389-396 (1970).
LOUWERSE, W. and J. L. P. VAN OORSCHOT: An assembly for routine measurements of photosynthesis, respiration and transportation of intact plants under controlled conditions. Photosynthetica (Prague) 3, 305-315 (1969).
LOWRY, O. H., N. J. ROSENBROUGH, A. LEWIS FARR, and R.J.RANDALL: Protein measurement with the Folin phenol reagent. J. BioI. Chern. 193, 265-275 (1951).
MAEHLY, A. C. and B. CHANCE: The assay of catalases and peroxidases. In: GLICK, D. (Ed.): Methods of biochemical analysis, Vol. 1, 357-425. Interscience publishers, New York, 1959.
MARK LUND, S.: Tryptic digestion and alkaline denaturation of catalase. Biochim. Biophys. Acta 321, 90-97 (1973).
MULCHI, C. L., R. J. VOLK, and W. A. JACKSON: Oxygen exchange of illuminated leaves of carbon dioxide compensation. In: HATCH, M. D., C. B.OSMOND, and R.O.SLATYER (Eds.): Photosynthesis and photorespiration, 35-50. Wiley-Interscience, New York, 1971.
NOVACKY, A. and R. E. HAMPTON: Peroxidase isozymes in virus-infected plants. Phytopathology 58, 301-305 (1968).
PATRA, H. K., M. KAR, and D. MISHRA: Catalase activity in leaves and cotyledons during plant development and senescence. Biochem. Physiol. Pflanz. (BPP) 172, 385-390 (1978).
PARISH, R. W.: Studies on senescing tobacco leafdiscs with special reference to peroxidase.
Z. P/lanzenphysiol. Bd. 97. S. 135-144. 1980.

144 JOHANNA M. BRABER
I. The effects of cutting, and of inhibition of nucleic acids and protein synthesis. P]anta 82, 1-13 (1968).
PAUL, K. G.: Peroxidases. In: The Enzymes, Vol. 8,2nd ed., 232. Academic press, New York, 1963.
PEET, M. M., A. BRAVO, D. H. WALLACE, and J. L. OZBUN: Photosynthesis, stomatal resistance and enzyme activities in relation to yield of field-grown dry bean varieties. Crop Sci. 17, 287-293 (1977).
PUTTER, J.: Peroxydasen. In: BERGMEYER, H. U. (Ed.): Methoden der Enzymatischen AnaIysen, Vo!' 1,648-653, Academic press, New York, 1970.
RACUSEN, D. and M. FOOTE: Peroxidase isoenzymes in bean leaves by preparative disc electrophoresis. Can. J. Bot. 44, 1633-1638 (1966).
SAFONOVA, M., R. DEJAEGERE, and V. SAFONOV: Specificity of protein components and isoenzyme spectra in functionally different organs of plants. Ann. Physio!. Univ. Brux. 15, 31-49 (1970).
SALIN, M. L. and P. H. HOMANN: Changes of photorespiratory activity with leaf age. Plant Physiol. 48, 193-196 (1971).
SAMEJIMA, T., W. J. MCCABE, and J. TSI YANG: Reconstitution of alkaline-denatured catalase. Arch. Biochem. Biophys. 127, 354-360 (1968).
SAUNDERS, B. c., A. G. HOLMES-SlEDLE, and B. P. STARK: Peroxidase, Butterworths, London, 1964.
SCANDALIOS, J. G.: Subunit dissociation and recombination of catalase isozymes. Proc. Natl. Acad. Sci. USA 53, 1035-1040 (1965).
SCHIEFER, S., W. TEIFEL, and H. KINDL: Plant microbody proteins. I. Purification and characterization of catalase from leaves of Lens culinaris. Hoppe-Seyler's Z. physio!. Chern. 357,163-175 (1976).
SCHONBAUM, G. R. and B. CHANCE: Catalase. In: BOYER, P. D. (Ed.): The Enzymes, Oxidation-Reduction Part C, 3rd ed., Vol. 13, 363-408. Academic press, New York, 1976.
SUND, H., K. WEBER, and E. MOLBERT: Dissoziation der Rinderleber-Katalase in ihre Untereinheiten. Eur. J. Biochem. 1,400-410 (1967).
SOLYMOSY, F., J. SZIRMAI, L. BECZNER, and G. L. FARKAS: Changes in peroxidaseisozyme patterns induced by virus infection. Virology 32, 117-121 (1967).
JOHANNA M. BRABER, Centre for Agrobiological Research (CABO), Postbus 14, 6700 AA, Wageningen, The Netherlands.
Z. Pflanzenphysiol. Bd. 97. S. 135-144. 1980.





























