Embed Size (px)
Citation preview

1570 Dispatch
Cell biology: Alternatives to baker’s yeastBenjamin S. Glick
Saccharomyces cerevisiae is an excellent modelorganism for addressing questions in cell biology, butother yeast systems are also providing new insightsinto several fundamental cellular processes.
Address: Department of Molecular Genetics and Cell Biology, TheUniversity of Chicago, 920 East 58th Street, Chicago, Illinois 60637,USA. E-mail: [email protected]
Current Biology 1996, Vol 6 No 12:1570–1572
© Current Biology Ltd ISSN 0960-9822
Cell biologists often use the term ‘yeast’ as a synonym forSaccharomyces cerevisiae. The reason is that S. cerevisiae, thebaker’s yeast, is by far the most-studied and best-characterized unicellular eukaryote. Early skepticism aboutusing S. cerevisiae as a model system turned into over-whelming enthusiasm as it became clear that, in manyrespects, yeast cells are very similar to higher eukaryoticcells. Yeast genetics combined with increasingly versatilebiochemical methods has illuminated processes rangingfrom signal transduction to organelle biogenesis. Recently,the sequencing of the entire genome of S. cerevisiae wascompleted, opening the way for a comprehensive molec-ular understanding of this organism [1].
Given these tremendous resources, why study cell biologyin any other model system? One reason is that certain path-ways and organelles are not present in yeast cells. Exam-ples include regulated secretion, which can be studied inprotozoa such as Tetrahymena thermophila [2], and flagellarassembly, which is being characterized using the green algaChlamydomonas reinhardtii [3]. Even when a process occursin S. cerevisiae, the mechanism in another yeast species maybe sufficiently different that significant insights can beobtained by comparing the two systems. Perhaps the mostfamous example is cell division in the fission yeastSchizosaccharomyces pombe [4]. I shall focus here on anaspect of cell biology that has only recently benefited fromthe use of yeasts other than S. cerevisiae — the biogenesisand dynamics of membrane-bounded organelles.
Non-Saccharomyces yeastsFor the purposes of this discussion, non-Saccharomycesyeasts can be divided into two categories: other buddingyeasts and S. pombe. S. pombe is evolutionarily quitedivergent from budding yeasts. This divergence has twoadvantages for research [5]. First, in some respects, S.pombe resembles mammalian cells more closely than doesS. cerevisiae. Second, if a process is similar in S. pombe andS. cerevisiae, it is likely to be conserved throughouteukaryotes. Many molecular genetic techniques are
available for S. pombe [5], and sequencing of the S. pombe genome is well under way (see the web site athttp://www.nih.gov/sigs/yeast/fission.html).
Cell biologists have made use of several budding yeastsaside from S. cerevisiae, including Pichia pastoris, Hansenulapolymorpha, Yarrowia lipolytica and Kluyveromyces lactis.Like S. cerevisiae, these yeasts can be manipulated usingmethods such as mating and sporulation, transformationwith integrating and replicating vectors, and gene replace-ment by homologous recombination [6]. Moreover, genesand antibodies obtained from research with S. cerevisiaecan often be used to study other budding yeasts.
Protein translocation and folding in the ERIn most cells, nascent secretory proteins associate with thesignal recognition particle (SRP) and then undergocotranslational translocation across the endoplasmic reticu-lum (ER) membrane [7]. S. cerevisiae is unusually adept atpost-translational translocation into the ER, and cansurvive in the absence of the SRP [7]. The SRP is essen-tial, however, for the viability of Y. lipolytica and S. pombe.Wise and colleagues [8] took advantage of this observationto perform a genetic study in S. pombe of the SRP54subunit, which contains a GTPase domain. Mutants ofSRP54 that were trapped in the GTP-bound conformationhad a dominant lethal phenotype, whereas mutantstrapped in the GDP-bound conformation did not interferewith the function of wild-type SRP54. These data led to aworking model for the role of GTP hydrolysis in the SRPreaction cycle.
After translocation into the ER, secretory proteins interactwith folding and quality-control machinery. One compo-nent of this machinery is calnexin, a molecular chaperonethat recognizes misfolded proteins that have glucoseresidues on their N-linked oligosaccharide side chains [9].The enzyme responsible for maintaining the glucosyla-tion state of misfolded proteins is UDP–glucose:glycopro-tein glucosyltransferase. Genetic analysis of thisquality-control system has been limited because S. cere-visiae lacks the glucosyltransferase and its calnexin-likeprotein is not essential. The ER of S. pombe, however,contains both UDP–glucose:glycoprotein glucosyltrans-ferase and a bona fide calnexin homolog that is essential forcell growth [10,11].
Organization and positioning of the Golgi apparatusThe processes that generate the unique organization ofthe Golgi apparatus are still mysterious. Although theGolgi apparatus in S. cerevisiae has been productively
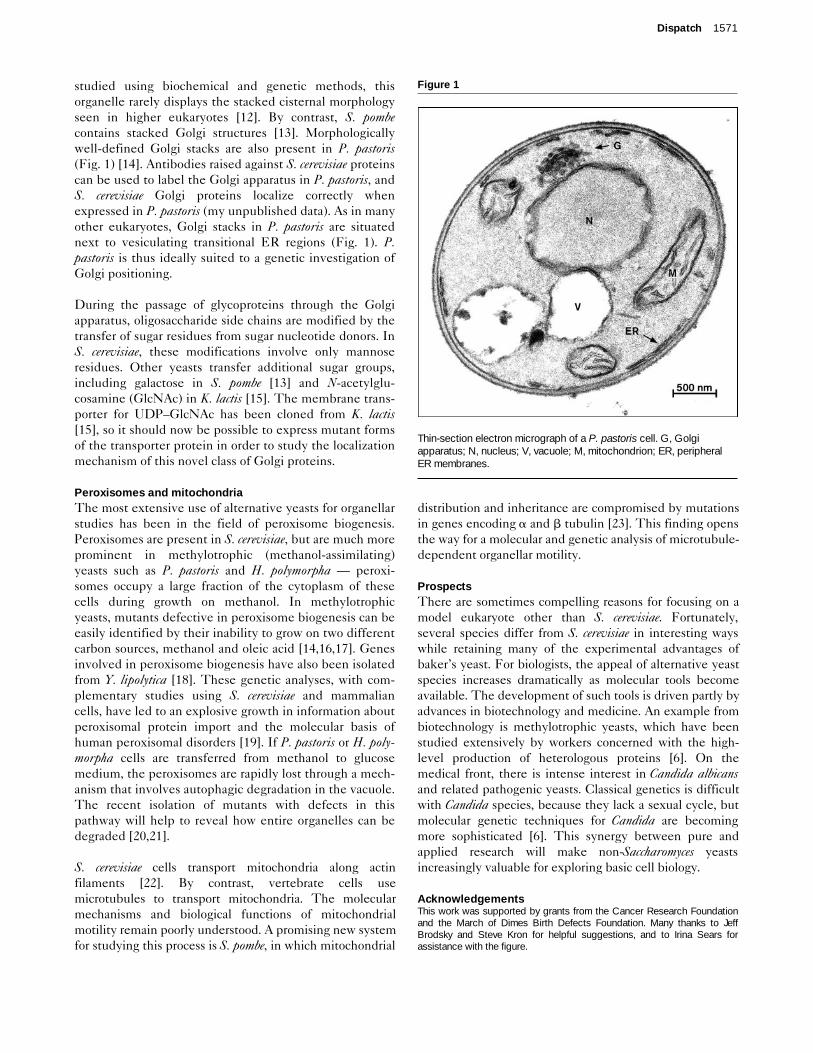
studied using biochemical and genetic methods, thisorganelle rarely displays the stacked cisternal morphologyseen in higher eukaryotes [12]. By contrast, S. pombecontains stacked Golgi structures [13]. Morphologicallywell-defined Golgi stacks are also present in P. pastoris(Fig. 1) [14]. Antibodies raised against S. cerevisiae proteinscan be used to label the Golgi apparatus in P. pastoris, andS. cerevisiae Golgi proteins localize correctly whenexpressed in P. pastoris (my unpublished data). As in manyother eukaryotes, Golgi stacks in P. pastoris are situatednext to vesiculating transitional ER regions (Fig. 1). P.pastoris is thus ideally suited to a genetic investigation ofGolgi positioning.
During the passage of glycoproteins through the Golgiapparatus, oligosaccharide side chains are modified by thetransfer of sugar residues from sugar nucleotide donors. InS. cerevisiae, these modifications involve only mannoseresidues. Other yeasts transfer additional sugar groups,including galactose in S. pombe [13] and N-acetylglu-cosamine (GlcNAc) in K. lactis [15]. The membrane trans-porter for UDP–GlcNAc has been cloned from K. lactis[15], so it should now be possible to express mutant formsof the transporter protein in order to study the localizationmechanism of this novel class of Golgi proteins.
Peroxisomes and mitochondriaThe most extensive use of alternative yeasts for organellarstudies has been in the field of peroxisome biogenesis.Peroxisomes are present in S. cerevisiae, but are much moreprominent in methylotrophic (methanol-assimilating)yeasts such as P. pastoris and H. polymorpha — peroxi-somes occupy a large fraction of the cytoplasm of thesecells during growth on methanol. In methylotrophicyeasts, mutants defective in peroxisome biogenesis can beeasily identified by their inability to grow on two differentcarbon sources, methanol and oleic acid [14,16,17]. Genesinvolved in peroxisome biogenesis have also been isolatedfrom Y. lipolytica [18]. These genetic analyses, with com-plementary studies using S. cerevisiae and mammaliancells, have led to an explosive growth in information aboutperoxisomal protein import and the molecular basis ofhuman peroxisomal disorders [19]. If P. pastoris or H. poly-morpha cells are transferred from methanol to glucosemedium, the peroxisomes are rapidly lost through a mech-anism that involves autophagic degradation in the vacuole.The recent isolation of mutants with defects in thispathway will help to reveal how entire organelles can bedegraded [20,21].
S. cerevisiae cells transport mitochondria along actinfilaments [22]. By contrast, vertebrate cells usemicrotubules to transport mitochondria. The molecularmechanisms and biological functions of mitochondrialmotility remain poorly understood. A promising new systemfor studying this process is S. pombe, in which mitochondrial
distribution and inheritance are compromised by mutationsin genes encoding a and b tubulin [23]. This finding opensthe way for a molecular and genetic analysis of microtubule-dependent organellar motility.
ProspectsThere are sometimes compelling reasons for focusing on amodel eukaryote other than S. cerevisiae. Fortunately,several species differ from S. cerevisiae in interesting wayswhile retaining many of the experimental advantages ofbaker’s yeast. For biologists, the appeal of alternative yeastspecies increases dramatically as molecular tools becomeavailable. The development of such tools is driven partly byadvances in biotechnology and medicine. An example frombiotechnology is methylotrophic yeasts, which have beenstudied extensively by workers concerned with the high-level production of heterologous proteins [6]. On themedical front, there is intense interest in Candida albicansand related pathogenic yeasts. Classical genetics is difficultwith Candida species, because they lack a sexual cycle, butmolecular genetic techniques for Candida are becomingmore sophisticated [6]. This synergy between pure andapplied research will make non-Saccharomyces yeastsincreasingly valuable for exploring basic cell biology.
AcknowledgementsThis work was supported by grants from the Cancer Research Foundationand the March of Dimes Birth Defects Foundation. Many thanks to JeffBrodsky and Steve Kron for helpful suggestions, and to Irina Sears forassistance with the figure.
Dispatch 1571
Figure 1
Thin-section electron micrograph of a P. pastoris cell. G, Golgiapparatus; N, nucleus; V, vacuole; M, mitochondrion; ER, peripheralER membranes.

1572 Current Biology 1996, Vol 6 No 12
References1. Dujon B: The yeast genome project: what did we learn? Trends
Genet 1996, 12:263–270.2. Chilcoat ND, Melia SM, Haddad A, Turkewitz AP: Grl1p, an acidic,
calcium-binding protein in Tetrahymena thermophila dense-coresecretory granules, influences granule size, shape, content,organization and release but not protein sorting or condensation.J Cell Biol 1996, 35:
3. Dutcher SK: Flagellar assembly in two hundred and fifty easy-to-follow steps. Trends Genet 1995, 11:398–404.
4. Forsburg SL, Nurse P: Cell cycle regulation in the yeastsSaccharomyces cerevisiae and Schizosaccharomyces pombe.Annu Rev Cell Biol 1991, 7:227–256.
5. Moreno S, Klar A, Nurse P: Molecular genetic analysis of fissionyeast Schizosaccharomyces pombe. Methods Enzymol 1991,194:795–823.
6. Sudbery PE: The non-Saccharomyces yeasts. Yeast 1994,10:1707–1726.
7. Walter P, Johnson AE: Signal sequence recognition and proteintargeting to the endoplasmic reticulum membrane. Annu Rev CellBiol 1994, 10:87–119.
8. Althoff SM, Stevens SW, Wise JA: The Srp54 GTPase is essentialfor protein export in the fission yeast Schizosaccharomycespombe. Mol Cell Biol 1994, 14:7839–7854.
9. Helenius A: How N-linked oligosaccharides affect glycoproteinfolding in the endoplasmic reticulum. Mol Biol Cell 1994,5:253–265.
10. Parlati F, Dignard D, Bergeron JJM, Thomas DY: The calnexinhomologue cnx1+ in Schizosaccharomyces pombe, is an essentialgene which can be complemented by its soluble ER domain.EMBO J 1995, 14:3064–3072.
11. Fernández F, Jannatipour M, Hellman U, Rokeach LA, Parodi AJ: Anew stress protein: synthesis of Schizosaccharomyces pombeUDP-Glc:glycoprotein glucosyltransferase mRNA is induced bystress conditions but the enzyme is not essential for cell viability.EMBO J 1996, 15:705–713.
12. Preuss D, Mulholland J, Franzusoff A, Segev N, Botstein D:Characterization of the Saccharomyces Golgi complex throughthe cell cycle by immunoelectron microscopy. Mol Biol Cell 1992,3:789–803.
13. Chappell TG, Hajibagheri NMA, Ayscough K, Pierce M, Warren G:Localization of an a 1,2 galactosyltransferase activity to the Golgiapparatus of Schizosaccharomyces pombe. Mol Biol Cell 1994,5:519–528.
14. Gould SJ, McCollum D, Spong AP, Heyman JA, Subramani S:Development of the yeast Pichia pastoris as a model organism fora genetic and molecular analysis of peroxisome assembly. Yeast1992, 8:613–628.
15. Abeijon C, Robbins PW, Hirschberg CB: Molecular cloning of theGolgi apparatus uridine diphosphate-N-acetylglucosaminetransporter from Kluyveromyces lactis. Proc Natl Acad Sci USA1996, 93:5963–5968.
16. Liu H, Tan X, Veenhuis M, McCollum D, Cregg JM: An efficientscreen for peroxisome-deficient mutants of Pichia pastoris. JBacteriol 1992, 174:4943–4951.
17. Nuttley WM, Szilard RK, Smith JJ, Veenhuis M, Rabuchinski RA: ThePAH2 gene is required for peroxisome assembly in themethylotrophic yeast Hansenula polymorpha. Gene 1995,160:33–39.
18. Szilard RK, Titorenko VI, Veenhuis M, Rachubinski RA: Pay32p of theyeast Yarrowia lipolytica is an intraperoxisomal component of thematrix protein translocation machinery. J Cell Biol 1995,131:1453–1469.
19. Subramani S: Convergence of model systems for peroxisomebiogenesis. Curr Opin Cell Biol 1996, 8:513–518.
20. Tuttle DL, Dunn WA, Jr: Divergent modes of autophagy in themethylotrophic yeast Pichia pastoris. J Cell Sci 1995, 108:25–35.
21. Titorenko VI, Keizer I, Harder W, Veenhuis M: Isolation andcharacterization of mutants impaired in the selective degradationof peroxisomes in the yeast Hansenula polymorpha. J Bacteriol1995, 177:357–363.
22. Simon VR, Swayne TC, Pon LA: Actin-dependent mitochondrialmotility in mitotic yeast and cell-free systems: identification of amotor activity on the mitochondrial surface. J Cell Biol 1995,130:345–354.
23. Yaffe MP, Harata D, Verde F, Eddison M, Toda T, Nurse P:Microtubules mediate mitochondrial distribution in fission yeast.Proc Natl Acad Sci USA 1996, 93:11664–11668.