Embed Size (px)
Citation preview

Sepsi et al. 2016. Meiotic chromosome dynamics in wheat. Plant Journal. Page 1.
CENH3 morphogenesis reveals dynamic centromere associations during synaptonemal complex formation and the progression through male meiosis in hexaploid wheat Adel Sepsi1), James D. Higgins, J.S. (Pat) Heslop-Harrison, Trude Schwarzacher*) Department of Genetics, University of Leicester, University Road, Leicester, LE1 7RH, UK 1) address from 1 July 2016: Agricultural Institute, Centre for Agricultural Research, Hungarian Academy of Sciences, Brunszvik u. 2., 2462, Martonvásár, Hungary *) corresponding author Authors’ email addresses: [email protected]; [email protected]; [email protected]; [email protected] This article has been accepted for publication by the Plant Journal and undergone full peer review but has not been through the copyediting, typesetting, pagination and proofreading process, which may lead to differences between this version and the Version of Record. Please cite this article as Sepsi A, Higgins JD, Heslop-Harrison JS, Schwarzacher T. 2016. CENH3 morphogenesis reveals dynamic centromere associations during synaptonemal complex formation and the progression through male meiosis in hexaploid wheat. Plant Journal. http://dx.doi.org/10.1111/tpj.13379 Video material is available on YouTube (user pathh1) which is also linked through https://molcyt.org/2016/09/29/cenh3-morphogenesis-delimits-dynamic-centromere-associations-during-synaptonemal-complex-formation-in-male-meiosis-of-hexaploid-wheat/ Plant Journal: Received Date : 10-Jun-2016; Revised Date : 04-Sep-2016; Accepted Date : 08-Sep-2016 ; Article type : Original Article SUMMARY During meiosis, centromeres in some species undergo a series of associations, but the processes and progression to homologous pairing is still a matter of debate. Here, we aimed to correlate meiotic centromere dynamics and early telomere behaviour to the progression of synapotonemal complex (SC) construction in hexaploid wheat (2n=42) by triple immunolabelling of CENH3 protein marking functional centromeres, and SC proteins ASY1 (unpaired lateral elements) and ZYP1 (central elements in synapsed chromosomes). We show that single or multiple centromere associations formed in meiotic interphase undergo a progressive polarisation (clustering) at the nuclear periphery in early leptotene, leading to formation of the telomere bouquet. Critically, immunolabelling shows the dynamics of these presynaptic centromere associations and a structural reorganisation of the centromeric chromatin coinciding with key events of synapsis initiation from the subtelomeric regions. As short stretches of subtelomeric synapsis emerged at early zygotene, centromere clusters lost their strong polarization, gradually resolving as individual centromeres indicated by more than 21 CENH3 foci associated with unpaired lateral elements. Only following this centromere depolarisation were homologous chromosome arms connected, as observed by the alignment and fusion of interstitial ZYP1 loci elongating at zygotene so synapsis at centromeres is a continuation of the interstitial synapsis. Our results thus reveal that centromere associations are a component of the timing and progression of chromosome synapsis, and the gradual release of the individual centromeres from the clusters correlates with the elongation of interstitial synapsis between the corresponding homologues.

Sepsi et al. 2016. Meiotic chromosome dynamics in wheat. Plant Journal. Page 2.
SIGNIFICANCE STATEMENT During meiosis, centromeres in some species undergo a series of associations, but the role of these associations to subsequent chromosome pairing is still debated. Here, we used immunolocalization of centromere and synaptonemal complex proteins to correlate meiotic centromere dynamics and early telomere behaviour to the progression of chromosome pairing in wheat. We found that centromere clusters gradually resolve after subtelomeric synapsis, and that this centromere depolarisation and individualization precedes elongation of interstitial arm synapsis. KEY WORDS Triticum aestivum L., CENH3, Meiosis, Synaptonemal complex, ASY1, ZYP1, centromere associations, centromere pairing, chromatin expansion-contraction

Sepsi et al. 2016. Meiotic chromosome dynamics in wheat. Plant Journal. Page 3.
INTRODUCTION Centromeres are specific chromosomal regions where kinetochore proteins assemble and connect to the bipolar spindle; they are thus essential in ensuring genome stability and accurate chromosome segregation both at mitosis and meiosis (Verdaasdonk and Bloom, 2011; Brar and Amon, 2008). Plant centromeres contain variable repetitive DNA sequences that are generally genome-specific (Heslop-Harrison et al., 1999; Heslop-Harrison, 2000; Lermontova et al., 2015) while the functional centromere can be defined epigentically by the chromosomal region encompassing centromeric nucleosomes where the canonical histone H3 has been substituted by CENH3, the centromere-specific histone H3 variant (CENP-A, CID or Cse4 in other organisms) (Houben and Schubert, 2003; Dalal et al., 2007). Kinetochore formation depends on these nucleosomes and loss of CENH3 causes centromere inactivity and chromosome elimination (Jiang et al., 2003; Dawe and Henikoff, 2006; Henikoff and Furuyama, 2012).
Centromere-centromere interactions occur during the meiotic process (Da Ines and White, 2015), a phenomenon suggested to control homologue pairing and proved to promote the fidelity of achiasmate chromosome segregation (Dawson et al., 1986; Karpen et al., 1996; Gladstone et al., 2009). Centromere associations from early meiotic interphase I to late prophase I have been reported in many organisms, and classified into three main classes 1.) centromere coupling: involving non-homologous, pairwise associations, (Church and Moens, 1976; Dernburg et al., 1996; Prieto et al., 2004) 2.) centromere clustering: the associations of the centromeres in one or very few groups at a restricted area of the nuclear periphery (Wen et al., 2012; Christophorou et al., 2013), and 3.) centromere pairing: consisting the homologous association of centromeres facilitated by meiotic proteins (Kemp et al., 2004).
Allohexaploid wheat (Triticum aestivum L.) carries three homoeologous genomes (AABBDD, 2n=6X=42) and each pair of homologous chromosomes has two pairs of homoeologous counterparts. In order to ensure genome integrity, chromosomes need to recognize the correct (homologous) pairing partner, and avoid recombination with a homoeologue. Indeed, irrespective of the partial homology between the A, B and D genomes of wheat (Sears, 1976), pairing, crossing over and chiasmata exhibit strict disomic inheritance, occurring only between homologous chromosomes. This diploid-like inheritance is controlled by the Ph1 (Pairing homoeologous 1) locus, mapped to the long arm of chromosome 5B (Riley and Chapman, 1958; Griffiths et al., 2006). Centromere associations in wheat were first reported from reconstruction of thin electron micrograph sections, showing that groups of two to seven centromeres were associated in early meiotic interphase (Bennett 1979). It was not clear however, whether these associations involve homologous chromosomes. The interaction of homologous chromosomes in early meiotic interphase has been reported when analysing the behaviour of a pair of rye chromosome arms in the wheat background and has been proposed as part of the homologue recognition process (Schwarzacher, 1997).
A series of subsequent studies directly visualised centromere-specific sequences in wild type and ph1 mutant wheat and showed early centromere associations followed by the clustering of the centromeres at a restricted area of the nuclear periphery (Martinez-Perez et al., 2000; Martinez-Perez et al., 2003). Centromere clustering coincided with the formation of the telomere bouquet: the gathering of the telomeres at one pole of the nuclear periphery (Gelei 1921). Although the precise timing of bouquet arrangement has been challenging due to the absence of independent developmental markers (Dernburg et al., 1995), telomere cluster formation has been found to coincide with initiation of synapsis and was placed to the leptotene-to-zygotene transition (Zickler and Kleckner, 1998; Ronceret and Pawlowski, 2010). Similarly, it has been difficult to visualise simultaneously the progression of centromere- and chromosome pairing from meiotic prophase I in wheat without appropriate immunoprobes, and there is no direct evidence about the nature and progression of presynaptic centromere interactions from association to homologous pairing.
Homologous chromosome pairing in meiotic prophase I is facilitated by the formation of the synaptonemal complex (SC), a tripartite proteinaceous structure organized along the length of meiotic chromosomes (Zickler and Kleckner, 1999). Axial elements (AE) of the SC first appear as discrete dispersed foci during G2 that then extend and coalesce. By leptotene, AEs form continuous threads along the whole length of the chromosomes that by zygotene become progressively connected by transverse filament (TF) proteins, implying the intimate

Sepsi et al. 2016. Meiotic chromosome dynamics in wheat. Plant Journal. Page 4.
synapsis of the homologues. The structure formed by the interconnection of the TF constitutes the central element of the SC, while axial elements once connected become the lateral elements of the SC (Schwarzacher, 2003; Page and Hawley, 2004). Proteins associated with the SC have been reported from fungi to humans (Kironmai et al., 1998; Meuwissen et al., 1997). ASY1 has been described as an axial element associated protein in Arabidopsis thaliana (Caryl et al., 2000; Armstrong et al., 2002) and is loaded onto the unsynapsed chromosome axes before synapsis. ZYP1 is a TF protein that polymerizes between the unsynapsed axial cores of homologous chromosomes during zygotene until synapsis is complete at pachytene (Higgins et al., 2005). Antibodies raised against the Arabidopsis ASY1 and ZYP1 have been used to detect the corresponding proteins in barley and rye and were shown to localise to the asynapsed- and synapsed chromosome axes, respectively (Mikhailova et al., 2006; Higgins et al., 2012).
In this study, our aim was to investigate centromere dynamics (coupling, clustering and pairing) during prophase I of meiosis in hexaploid bread wheat. We have raised a specific antibody to Triticum aestivum CENH3 and performed dual immunolocalisation with SC proteins ASY1 and ZYP1. Using confocal and widefield fluorescence microscopy we have measured centromere associations and the development of the SC with the formation of the telomere bouquet, thus providing insight into centromere and chromosome arm pairing, as well as an accurate timing for meiotic centromere dynamics.
RESULTS Staging of early meiosis Events occurring during meiotic homologue pairing of the 2n=6x=42 chromosomes in wheat follow a specific time-course and must be integrated with exactly defined stages of meiosis to assess the sequence of centromere movements. To investigate the progression of meiosis and the dynamics of chromatin and centromere behaviour, we used an antibody to CENH3 marking active centromeres simultaneously with antibodies to the SC-axial and central element related proteins, ASY1 and ZYP1. In additional experiments, telomeres were marked by fluorescent in situ hybridization. With well-defined staging of pollen mother cells (PMCs), telomere and centromere movements could be followed during chromosome alignment and pairing events, encompassing meiotic interphase and prophase I (Figures 1-4, Table 1). Progression through meiosis I and II was also studied to show that bivalent formation and chromosome disjunction is normal under our experimental conditions and the line used.
Using classical morphological criteria (Bennett et al., 1973; Holm 1986) combined with ASY1 and ZYP1 immunostaining (Armstrong et al., 2002; Mikhailova et al., 2006; Higgins et al., 2012; this study), the time course of meiotic progression was defined and is decribed in Table 1. Nuclei in PMCs at the interphase before meiotic prophase (meiotic interphase) are larger than somatic interphase nuclei (Figure S1) and harboured as expected two or three nucleoli of various sizes within the nucleus. Appearance of ASY1 and ZYP1 signal (Figures 1-2) defined the sub-stages of meiotic prophase that were then used to stage the nuclei when analysing CENH3 signal and relating them to the bouquet stage and clustering of centromeres and telomeres (Table 1). In diplotene, desynapsis was indicated by the progressive decomposition of the linear ZYP1 signal, but also reappearance of fuzzy ASY1 signal (Figure 2f). By diakinesis, the breakdown of the nuclear envelope resulted in the loss of the sub-spherical shape of the nuclei and defined bivalents emerged, as homologous centromeres separated and both ASY1 and ZYP1 disappeared from the contracted chromosomes. CENH3 dynamics in early meiosis Meiotic nuclei had much brighter CENH3 fluorescence compared to the smaller somatic/mitotic tapetal nuclei from the young anthers. The difference in fluorescence intensity was seen from mid-meiotic interphase throughout meiosis (Figure S1).

Sepsi et al. 2016. Meiotic chromosome dynamics in wheat. Plant Journal. Page 5.
Meiotic interphase: early centromere associations When ASY1 first appeared in meiotic interphase, centromeres were observed with CENH3 signals of varying sizes in one hemisphere of the nucleus (Figure 1a; Movie S1) while telomeres tended to be located in the opposite hemisphere (Table 1). We observed a considerable variation in the total number of CENH3 foci per nucleus during meiotic interphase, ranging between 15 and 33 (Mean=23.3, SD =4.1, N= 47), indicative of variable centromere groupings or arrangements, and possibly centromere movement (Figures 1a, 3a), with some single centromeres along with associations of three or more. Leptotene: clustering of centromeres followed by the telomere bouquet Concomitant with the initial loading of ASY1 onto the chromatin axes as short stretches and their subsequent elongation, centromeres, either single or in associations, underwent a progressive polarization in a restricted area of the nuclear periphery (Figure 1b; Figure S2; Movie S2). The movement was accompanied by further centromere associations (clustering), reflected by the substantial decrease in the number of CENH3 foci to a mean of 9.6 (SD=3.3, N=21; Figure 3a) and increase in size of individual foci (compare Figure 1a with 1b), while the total volume of CENH3 signal remained similar to interphase (Figure 3b). Centromere polarization during leptotene was followed by the formation of the telomere bouquet at the opposite pole of the nucleus (Figure 1b; Table 1). Importantly, during this dynamic movement, centromeres remained in close proximity with the nuclear envelope. Early zygotene: resolution of centromere clusters coincided with emergence of homologous synapsis At the onset of zygotene, the centromere clustering resolved at the same time as the continuous chromosome axes became visible (Figures 1c, 2a, b). During the separation of the centromeres, the telomere bouquet was fully formed with tight telomere clustering (Table 1). Evidence for initiation of synapsis came from short, linear ZYP1 stretches that were organised in a conical structure at the telomere pole of the nucleus opposite to the centromeres (Figure S3; Movie S3). This is consistent with the ZYP1 polymerization to start from the subtelomeric regions in barley (Higgins et al., 2012). Coincident with the emergence of linear ZYP1 at the telomeric regions, we observed numerous interstitial punctate ZYP1 foci associated with the ASY1 strands, in some cases intermitting the continuity of the ASY1 signal (Figures 2a, b; Table 1). We interpret these ZYP1 foci as the initiation point of interstitial synapsis, but elongation was not observed in early zygotene.
Previous studies involving species of the Triticeae noted that once lateral elements are connected by the synaptonemal complex, the immunodetection of ASY1 is reduced and progressively replaced by the ZYP1 signal on the now-synapsed chromosomes (Mikhailova et al., 2006; Khoo et al., 2012). To study the colocalisation of CENH3, ASY1 and ZYP1 proteins in detail on wheat meiocytes, we used high-resolution confocal laser scanning microscopy (Figure 4). ZYP1 elongation progressively connected two parallel but separated ASY1 threads with the ASY1 signal reduced at the ‘connection’ point (Figure 4c, also visible at late zygotene in Figure 4e). If centromere clusters resolved as homologous centromere pairs, then homologously associated centromeres would have a central role in homologue recognition and would be connected with the assembled SC (ZYP1). In contrast, our three-dimensional rendering of confocal stacks showed that all CENH3 foci co-localised with ASY1 staining only, but not ZYP1 and were thus unpaired. Furthermore, isolated single centromeres were found to co-localise with a single ASY1 thread (Figure 4a, balso visible at late zygotene, Figure 2c, Movie S4, and Figure S6a;), evidence that centromeres become released individually and not necessarily as paired homologues from the clusters.
Although the mean number of CENH3 foci per zygotene nucleus was 21 (SD=5, N=45; Figure 3a), the number varied from 14 to 33, reflecting progressive release of centromeres from the clusters (Figure 1c, 2a, b; Figure S4). Almost half the early zygotene nuclei (21 of 45) had more than 21 (the number of centromere pairs) CENH3 foci, supporting our observation that centromeres do not resolve as pairs at early zygotene (Figure 3a).

Sepsi et al. 2016. Meiotic chromosome dynamics in wheat. Plant Journal. Page 6.
Mid-late zygotene: centromere clusters and homologous synapsis progressed to centromere synapsis Mid-zygotene was characterised by elongation of synapsis from the interstitial punctate ZYP1 foci (Figure 2c). Unpaired centromeres, including singlets and larger associations were labelled with the ASY1 antibody (Figures 2c, d; 4d, Movie S4), but also homologously paired centromeres could be unequivocally identified by ZYP1 strands (Figures 4e, also visible at pachytene, Figures 2e; 4f) and became more frequent during zygotene progression. The mean number of centromeric foci at mid-late zygotene was 21.3 (SD=3.2, N=55) with a significant fraction of nuclei (58%) showing 21 or fewer loci (Figure 3a) and 21 bivalents formed at the end of zygotene. A high number of these unsynapsed chromatin threads observed at late zygotene involved interlocks (Figure 4e). Pachytene-diplotene: synaptonemal complex disassembly, interstitial regions separated first At pachytene, both chromosome arms and centromeric foci showed full synapsis (Figure 2e; Table 1). Twenty-one compact CENH3 foci could be detected, representing all the paired centromeres (Figure 3a). ASY1 signal, already much diminished at late zygotene (Figure 2d), was not detected at chromatin threads in pachytene, but at the peripherally located nucleolus (Figure 2e). Interstitial regions desynapsed at early diplotene (Table 1), seen by the fractional breakdown of the ZYP1 signal at multiple points on the chromosome arms (Figure 2f) and exhibited a characteristic, fuzzy ASY1 signal (Figure 4h, i). As diplotene progressed and ASY1 reduced, paired centromere foci started to form bipartite structures (Figure S5a, b), and subsequently separated and dispersed (Figure S5c, d) resulting in 42 individual centromeres by diakinesis (Figure 5a). Diakinesis-metaphase I and meiosis II: ASY1 signal returned No ASY1 or ZYP1 signal localized on bivalents during diakinesis and metaphase I (Figure 5a, b). Centromeres became bi-oriented as chromosomes lined up at the metaphase plate. Sister centromeres were held together as bipartite foci during anaphase I and telophase I (Figure 5c-e) and separated as two individual foci representing individual sister centromeres at metaphase II (Figure 5f). By mid-anaphase I, while homologous centromeres and chromosomes were pulled apart, ASY1 signal reappeared around the dividing chromatin. It was first detected as strong punctate foci (Figure 5d), which then transformed to heavy parallel bundles (Figure 5e), parting from the zone of the centromeres towards the equatorial plate, remaining in close association with the chromosomes. It encompassed the space of the meiotic division, as a frame for the dividing chromatin.
ASY1 was also detected during meiosis II: as dispersed punctate signal in metaphase II, with two heavy foci at the opposite poles of the nucleus (Figure 5f); and in anaphase II, where it first enveloped the dividing chromatin as a dispersed signal, with the same two heavy foci at the poles as seen in metaphase II (Figure 5g). In late anaphase II parallel bundles of ASY1 reappeared, but with much lower frequency and intensity than in anaphase I. (Figure 5h). Prominent ASY1 staining persisted in the tetrads and co-localised with the nucleoli (Figure 5i). The presence of ASY1 during meiosis long after the disassembly of the synaptonemal complex is interesting. ASY1 is not an SC component itself (Armstrong et al. 2002) but associates with the chromatin at the SC and thus a role as chromatin chaperon during meiotic cell division could be possible.
At metaphase I, 42 CENH3 foci were visualised (Figure 5b, c). At telophase I, 21 double foci were detected (Figure 5e) that split into smaller foci during metaphase II to telophase II (Figure 5f-g). The number of centromeres in the tetrad stage ranged from 7 to 10, indicating centromere coupling (Figure 5i). Centromere expansion-contraction cycles and association with dispersed ZYP1 signal Our observations of centromeres in meiosis I revealed expansion coincident with changes in their associations during prophase while there were also changes in nuclear volume (Figure 3). We measured area ( m2) occupied by the totality of CENH3 foci within the nucleus throughout meiotic interphase- and prophase I. There is evidence that total centromere size and the size of the global chromatin are strongly correlated (Zhang and Dawe, 2012) and the nucleus (Dawe et

Sepsi et al. 2016. Meiotic chromosome dynamics in wheat. Plant Journal. Page 7.
al. 1994) (or chromatin) undergoes programmed expansion during meiosis. In the relatively flat, sub-spherical preparations, nuclear area was found to increase by 87% (P=1.2×10-16) during zygotene, while centromere area increased by a similar amount (108%, P=3.2×10-10) slightly but notably earlier between late interphase and early zygotene (Figure 3b). Confocal image stacks collected from meiotic nuclei immunostained for CENH3-ASY1-ZYP1 proteins, detected a dispersed ZYP1 signal around the centromeric regions during centromere clustering. The signal co-localised with CENH3 foci, but clearly differed from the linear and punctate ZYP1 marking SC assembly (Figure 2a), suggesting a mechanistically distinct role for ZYP1 other than in connection with the SC assembly. Early, presynaptic centromere associations in yeast and Drosophila have been shown to depend on the components of the SC central elements Zip1 and C(3)G), respectively (Tsubouchi and Roeder, 2005; Christophorou et al., 2013). To exclude cross excitation or bleed through of red signal into the far red excitation channel, experiments were conducted with and without CENH3 and also using a ZYP1 antibody raised in different animals (Figure 2; Figure S6). Interestingly the dispersed ZYP1 could not be detected during chromosome arm pairing (mid zygotene, Figure 2c,d) while by late zygotene-early pachytene it was present at the synapsed centromeres (Figure 2e, Figure S6) and persisted to late diplotene (Figure 2f). Unsynapsed centromeres showing ASY1 did not exhibit the dispersed ZYP1 signal (Figure S6a). It is not clear however whether the dispersed ZYP1 is not present at the centromeres by mid zygotene or the sensitivity of our detection could not visualise the minor ZYP1 population. DISCUSSION The organisation of the centromeres (centromeric DNA and histone proteins), as chromosomal sites responsible for kinetochore assembly and essential for chromosome segregation, has been described in detail (Heslop-Harrison and Schwarzacher, 2011). Here, we are able to examine the progress of wheat pollen mother cell nuclei to show key aspects of how centromeres behave during male meiosis. It is critical that the centromeres, chromatin, and progression of pairing are linked closely in an analysis of the dynamic progression and temporal stages of meiotic events and to the completion of meiosis, achieved here by combining immuno-staining of CENH3 to mark the centromeres with simultaneous visualisation of ASY1 and ZYP1 of the synaptonemal complex.
SC antibodies ASY1 and ZYP1 have been used in a number of plants to follow the progression of chromosome pairing and their presence was detected from meiotic interphase to late prophase I (Armstrong et al., 2002; Lohmiller et al., 2008). Triticeae show a reduced ASY1 level along the synapsed chromosome axes from zygotene to early diplotene (this study and Phillips et al., 2012) which is sometimes, but not always seen in Arabidopsis and tomato and may reflect a different mechanism for homologous synapsis or may be caused by the masking of the protein epitope.
Chromosome pairing in wheat and its pseudo-diploid behaviour has been of great interest since wheat’s polyploid nature was first revealed (Kihara 1930; Sears 1939). Many studies showed aspects of chromosome pairing behaviour from the 1950s to 1980s including the important role of the Ph1 gene locus in restricting pairing of homoeologous chromosomes which were later supported by electron microscopy (Jenkins, 1983; Holm, 1988). All current models of chromosome pairing in wheat emphasize the importance of centromere movement and clustering starting from early meiotic prophase, eventually resolved into the pairing of strictly homologous chromosomes by pachytene. Here, we can add the exact timing of dynamic events of centromere clustering with respect to the well-accepted progression of lateral element appearance from leptotene, and paired SC formation during zygotene. The role of presynaptic centromere associations and their transition to homologous pairing Individual chromosome pairs visualised by in situ hybridisation (Schwarzacher et al., 1992) can be studied in alien introgression lines of hexaploid wheat, carrying whole chromosomes or chromosome arms from a related species (Molnár et al., 2015; Ali et al., 2016). A study on a

Sepsi et al. 2016. Meiotic chromosome dynamics in wheat. Plant Journal. Page 8.
wheat-rye translocation line revealed that cognition of homologous chromosome partners initiates at the centromeres in meiotic interphase and subsequently, homologous chromosome segments align in leptotene with synapsis starting at the telomeres (Schwarzacher, 1997). Subsequent studies on wild type and ph1 mutant wheat reported centromere clustering at the telomere bouquet stage and suggested that this clustering plays a critical role in the homologous pairing process. It has been proposed that presynaptic centromere associations are under the control of the Ph1 locus and serve as an initial chromosome sorting mechanism that facilitates the cognition of the homologues, and consequently influences the specificity of synapsis (Martinez-Perez et al., 2001; Martinez-Perez et al., 2003). According to this model, homologous centromeres recognize each other during clustering, and subsequently become released from the clusters as homologous centromere pairs.
Other investigations on hybrid lines of Triticeae (wheat-rye, oat-maize) (Bass et al., 2000; Jenkins, 1983; Maestra et al., 2002) concluded that transition from non-homologous to homologous centromere interactions in meiotic prophase is driven by synapsis, and centromere associations in early meiosis are not based on homology and thus cannot promote homologous chromosome recognition. Consistent with these, a study involving chromosomes with centromere introgressions from rye both in the presence and in the absence of Ph1 suggested that homologous centromere pairing was rather a result of the synaptonemal complex expansion than the presynaptic association of the centromeres. The frequency of homologous associations of rye centromeres both in the ph1 mutant and in the wild type wheat background increased with the progression of meiosis, reaching 100% at pachytene (Corredor et al., 2007). Bhullar et al., (2014) identified a Ph1 candidate gene in wheat, that if silenced by RNAi silencing in Arabidopsis enhanced centromere coupling at leptotene, followed by multivalent formation, and postulate its early functioning in enabling association of homologues before meiosis.
Our observations on centromere and chromosome pairing revealed that, centromere clustering coincides with the initial formation of SC axial elements during meiotic leptotene, and is temporally tightly followed by the formation of the telomere bouquet. In accordance with other studies (Gelei 1921, Holm 1986, Higgins et al., 2012), we show that synapsis initiation and elongation are spatiotemporally separated in two classes (Figures 6, Table 1): 1.) Synapsis emergence at the subtelomeric regions occurring at the leptotene-zygotene transition, with the telomere bouquet fully developed at the opposite pole of the centromere foci, and 2.) Elongation of interstitial synapsis at mid-zygotene initiated at multiple points of the chromosome arms. The initiation and progressive release of individual centromeres from the clusters coincides with the emergence of subtelomeric synapsis (ZYP1) during which no linear or punctate ZYP1 colocalise with the centromeres.
Several independent studies on wheat-alien hybrid lines suggested that homologous centromeres recognize each other and associate at some point during the late interphase-early prophase transition, most likely during the clustering stage (Aragón-Alcaide et al., 1997; Martínez-Pérez et al., 1999; Schwarzacher, 1997). Our observations clearly show that instead of remaining connected, homologous centromeres became released individually from the clusters, presumably at the same time point, as they might be included into the same or adjacent clusters. Resolution of the centromere clusters infers the release of the centromeres from the restricted area at the nuclear periphery and the end of the centromere polarization. The obvious gradual progression of centromere separation from the clusters and of the arm and centromere synapsis suggest that centromere clusters might play a role in the timing of the emergence and elongation of interstitial synapsis. Those chromosomes whose centromeres became released earlier, are in advance to initiate interstitial elongation compared to those being released at a later time point (Figure 6). The presence of larger centromere associations at zygotene might act to keep centromeres turned off and released individually only those undergoing pairing. This pairing strategy would enable the cell to deal with the large genome size and with the presence of the homoeologous genomes and thus minimize the number of interlocks. If homologous centromeres and consequently homologous chromosomes become released at the same time point they have a better opportunity to pair with each other and to avoid the pairing with a homologue released at a different time-slot. Kleckner et al. (2004) proposed that chromosomes play a ‘governing role’ not only via their information content but also via internal mechanical properties generated by chromatin expansion and contraction. The release of individual

Sepsi et al. 2016. Meiotic chromosome dynamics in wheat. Plant Journal. Page 9.
centromeres from the polarised clusters that we see may be a result of mechanical forces applied by the telomere bouquet and the subsequent subtelomeric synapsis (Figure 6).
Our observations on the expansion-contraction state of the centromeric regions during meiosis further support this model. Centromere clustering at leptotene implied a clear change in the packaging of the centromeric chromatin. Centromeres entered the clusters as highly compact, point-like structures, and left as significantly expanded elongated threads. Such a major chromatin reorganisation prior to the initiation of chromosome synapsis occurs in maize and includes elongation of knob heterochromatin, increase in chromosome volume, and alignment and peripheral localisation of telomeric regions (Dawe et al., 1994). While centromeres clustered during leptotene, chromatin including centromeres undergoes reorganisation.
Centromere clustering might thus be involved in the timing of the meiotic process, it keeps the chromatin in a polarized configuration -together with the telomere bouquet- while the centromeres structurally reorganise and subtelomeric synapsis initiates (Figure 6). Subtelomeric synapsis provides the chromatin an orderly arrangement, whereby homologous chromosomes are attached together by their end to a restricted area of the nuclear periphery, and this organisation is maintained until large parts of the interstitial regions are synapsed. In this highly strict chromatin context, the release of the centromeres from the clusters and nuclear periphery provide an enhanced motility of the corresponding homologues, enabling homology search and thus precedes the elongation of interstitial synapsis.
Meiotic control of the dynamism of centromere associations To decipher the mechanism controlling presynaptic centromere associations and dissociations is beyond the scope of the present work, however, the finely tuned sequence of events in centromere behaviour and synapsis suggest that the two pathways are under the same genetic control. In budding yeast (Saccharomyces cerevisiae), centromere associations have been linked to key events of meiotic recombination and homologous synapsis (Jin et al., 1998; Hayashi et al., 1998); while many features of yeast meiosis are conserved in other eukaryotes, it is not clear how chromosome cognition and pairing behaviour can be extrapolated from the small genome and point centromeres.
The discovery of non-homologous centromere coupling (NCC), which follows centromere clustering in yeast spo11 mutants - lacking the gene encoding the conserved topoisomerase Spo11, responsible to introduce DNA double strand-breaks (DSBs) and thus promoting homologous recombination - pointed a link between presynaptic centromere associations and meiotic recombination (Tsubouchi and Roeder, 2005; Obeso and Dawson, 2010). Although the presence of yeast Zip1 gene (synonym to Zyp1 in Arabidopsis) was found critical for the initiation of NCC, simultaneous immunolabelling of the centromeres and ZIP1 protein, showed no or very little co-localisation. This indicated that ZIP1 regulates centromere coupling in a different way than the formation of chromosome synapsis (Obeso and Dawson, 2010; Chuong and Dawson, 2010). The evidence, that ZIP1 controls centromere associations in a synapsis-independent manner, has been reported by studying phosphatase PP4 that dephosphorylates many Mec-1 (a DSB-activated checkpoint serine/threonine kinase) dependent phospho-proteins after DSB repair, a process critical for the cell to pass the recombination checkpoint. In pp4 mutants phosphorylated Mec-1 targets are accumulated, and in this checkpoint active state both DSB repair and homology independent centromere coupling were blocked. Zip1 Serine 75 has been identified as a consensus site for Mec1, and it has been shown that its hyper-phosphorylation destabilizes centromere coupling. Zip1-S75 phosphorylation by Mec-1 occurs in response to recombination initiation, which connects meiotic centromere dynamics to DSB repair (Falk et al., 2010). These observations imply that the separation of NCCs and the initiation of SC formation are regulated by the same molecular signal, i.e. DSB formation and subsequent homologous recombination. Early non-homologous centromere associations have been proposed to sequester centromeres from their homologs at the time when recombination is initiated, in order to inhibit recombination to occur around the centromeres (Bardhan, 2012). A high resolution 3D-reconstruction of early centromere associations and SC formation in barley provided the first direct evidence in plants that presynaptic centromere associations are non-homologous. These results raised the possibility that early centromere-centromere interactions

Sepsi et al. 2016. Meiotic chromosome dynamics in wheat. Plant Journal. Page 10.
play a similar role in higher plants as proposed for yeast: i.e. the inhibition of centromeric recombination (Phillips et al., 2012). Our observations on wheat meiocytes revealed that apart from the major linear and punctate ZYP1 signal, a minor population of dispersed ZYP1 colocalises with the CENH3 protein at specific intervals of meiotic prophase I. This suggests a role for ZYP1 in the regulation of meiotic centromere dynamics, which in turn influences the timing of chromosome synapsis. Although further studies are required to reveal the mechanisms through which ZYP1 controls centromere associations, the present work established a reliable timing for meiotic centromere movement and revealed a link between presynaptic centromere associations and the initiation and progression of synapsis. SC formation has been shown to be stringently coupled with Spo11-induced meiotic DNA DSB formation and consequent interhomolog recombination in Arabidopsis and barley (Higgins et al., 2005; Barakate et al., 2014). It is thus critical to include centromeres in further studies on synapsis and meiotic recombination to elucidate how centromere associations are involved and controlled during the recombination process. Understanding the role of the centromere dynamism in key meiotic processes might shed light to mechanisms involved in the repression of centromeric recombination, so far representing a genetic bottleneck for wheat breeders. EXPERIMENTAL PROCEDURES Nuclei preparation Spikes of Triticum aestivum ‘Chinese Spring’ (CS) estimated to be entering meiosis were collected approx. three hours after the light came on. The first and second florets of each spikelet from the bottom half of the spike were dissected. One of the three anthers per floret was measured and squashed in 50% acetic acid to identify the approximate meiotic stage (Bennett et al., 1973). To minimize cell distortion, isotonic paraformaldehyde (PFA, Agar Scientific, Stansted, UK, No. R1026, 16%; diluted to 4% with PBS, pH=7.4) was used for fixation for 30-60 min, washed and stored at 4°C overnight. Sampled anthers were then digested with an enzyme cocktail (Houben et al., 2003); 2.5 % pectinase, 1U/mg, No. 17389, 2.5 % pectolyase, 0.3 U/mg, No. P3026, 2.5 % cellulase, 0.3 U/mg, No. C1184, 2 % cytohelicase, No. C8274; all Sigma-Aldrich, St Louis, USA). Pollen mother cell loculi were squeezed out in a drop of 0.1% Triton-X-100 and nuclei were counterstained by adding a drop of DAPI (4´, 6-diamidino-2-phenylindole; 4mg/l). Preparations were covered with a coverslip and the quality of the cells was determined by epifluorescent microscopy. Slides were frozen on dry ice, coverslips removed and transferred to PBS buffer at 4°C. Preparations were then used for immunolabelling or in situ hybridisation immediately or after storage for up to 24h at 4°C. Antibody production and immunolabelling The wheat anti-CENH3 antibody was raised using the synthetic peptide CARTKHPAVRKTK (12 amino acids from the N-terminus of wheat CENH3 starting with a cysteine for conjugation (Li et al., 2013). The peptide was coupled to keyhole limpet hemocyanine protein carrier and injected into rabbits for immunisation (Diagenode, Seraing, Belgium). SC protein antibodies, At-ASY1 (rat) and At-ZYP1 (guinea pig and rat) have been described and produced earlier (Armstrong et al., 2002; Higgins et al., 2005).
For immunolabelling nuclei preparations were blocked in TNB blocking buffer (0.1M Tris-HCl, pH 7.5, 0.15M NaCl, 0.5% (w/v) Blocking Reagent, Sigma-Aldrich) for one hour at room temperature, then incubated with CENH3 (rabbit), ASY1 (rat) and ZYP1 (guinea pig) primary antibodies for 3h at 37°C. Slides were washed in PBS and incubated for 45min at 37°C with anti-rabbit-Alexa 594 or 488 (Invitrogen, California, USA, No. A-1172 or A-11008, 1:300), anti-rat-Alexa 488 or 594 (Invitrogen, No. A-21208, or Abcam No. ab150160, 1:300) and anti-guinea pig-Alexa 647 (Abcam, No. ab150187, 1:150) depending on the primary antibody used. Slides were mounted in 1µg/ml DAPI-50% Antifade (Citifluor, London, UK).

Sepsi et al. 2016. Meiotic chromosome dynamics in wheat. Plant Journal. Page 11.
In situ hybridization (FISH) The universal plant telomeric repeat sequences (TRS) were amplified by PCR using the oligomer primers T1 (5’-TTTAGGG-3’) 5 and T2 (5’-CCCTAAA-3’) 5 (Schwarzacher and Heslop-Harrison, 1991)(Cox, 1993), and labelled with biotin-14-dATP by nick-translation (BioNick Labelling System, Invitrogen,). PFA-fixed nuclei were permeabilized in a 0.1% (v/v) Triton-X - 0.1% (w/v) CHAPS hydrate solution (Sigma, No. C9426), and digested with 100µg/ml pepsin (Sigma, No. P6887) before fixing in ready-made saline buffered 4% PFA (Fisher Scientific, Loughborough, UK, No. F/1520/21). Hybridisation mixture contained 60 ng labelled TRS probe, 50% deionised formamide and 10% dextran sulphate in 2x SSC. Slides with the probe mixture were denatured at 75°C for 6 minutes and incubated at 37°C overnight. Biotin signals were detected using streptavidin Alexa 488 (Roche Diagnostics). Slides were mounted as above. Imaging Immunofluorescence images were taken on a Nikon Eclipse 80i epifluorescent microscope DS-QiMc monochromatic camera (Nikon, Tokyo, Japan). Series of grayscale images for each colour were captured along the Z-axis, taking 15-20 stacks per colour. Single channel images were pseudocoloured and merged in NIS elements (Nikon). All-in-focus images were created using the Extended Depth of Focus (EDF) module.
Confocal images were collected using an Olympus IX81 FV1000 confocal laser scanning microscope with a 60×1.35 NA UPlanSAPO objective (Olympus, Tokyo, Japan). Z-series of Alexa 594, 488, 647 and DAPI images were taken in sequential scan mode (50-100 stacks per image, 200nm distance). Image stacks were deconvolved using Huygens Essential software (Scientific Volume Imaging, Hilversum, The Netherlands). For 3D reconstruction of centromeres, and SC axial and central elements were processed using the Imaris surface reconstruction tool (Bitplane Scientific Software, Zurich, Switzerland). Measurements were carried out on EDF images by using the area measurement tool of NIS elements. Swelling has been observed on some of the nuclei subject to in situ hybridisation, therefore these were excluded from any nuclear measurement data. ACKNOWLEDGEMENTS The research leading to these results has received funding from the People Programme (Marie Curie Actions) of the European Union’s Seventh Framework Programme (FP7/2007-2013) under REA grant agreement no 625835. We thank the Advanced Imaging Facility at the University of Leicester (Dr. Kees Straatman) for training with confocal microscopy and subsequent image analysis. We thank Dr. John Bailey for help with the nuclei preparations. Federica Raccis and Manica Balant, ERASMUS students, are acknowledged for technical assistance. CONFLICT OF INTEREST The authors declare no conflict of interest. SUPPORTING INFORMATION Additional Supporting Information may be found in the online version of this article. Supporting figures: Figures S1: CENH3 fluorescence intensity of meiotic and mitotic nuclei of hexaploid wheat. EDF images of double immunostaining to visualise active centromeres with anti-CENH3 and SC axial elements with anti-ASY1 at meiotic interphase, leptotene, zygotene and diplotene. Figure S2: Three-dimensionally reconstructed confocal images of a leptotene nucleus immunostained for CENH3 and ASY1 taken from Movie S2. Figure S3: Three-dimensionally reconstructed confocal image of an early zygotene nucleus immunostained for CENH3, ASY1 and ZYP1 proteins taken from Movie S3.

Sepsi et al. 2016. Meiotic chromosome dynamics in wheat. Plant Journal. Page 12.
Figure S4: Progressive resolution of centromere clusters during zygotene. EDF images of immunostaining to visualise active centromeres with anti-CENH3 and SC axial elements with anti-ASY1 and DAPI staining. Figure S5: Separation of the synapsed centromeres at diplotene. EDF images of immunostaining to visualise active centromeres with anti-CENH3 and SC axial elements with anti-ASY1 and DAPI staining. Figure S6: The presence of the dispersed ZYP1 at the centromeres. EDF images of immunostaining with ZYP1 (rat) and ZYP1 (guinea pig) detected with red or far red respectively. Supporting animations Movie S1: Three-dimensionally reconstructed meiotic interphase nucleus immunostained for CENH3 and ASY1 proteins. Movie S2: Three-dimensionally reconstructed confocal image of a leptotene nucleus immunostained for CENH3 and ASY1 proteins. Movie S3: Three-dimensionally reconstructed confocal image of an early zygotene nucleus immunostained for CENH3, ASY1 and ZYP1 proteins. Movie S4: Sequence of individual confocal image stacks collected from the nucleus presented in Figure 2c. REFERENCES Ali, N., Heslop-Harrison, J.S., Ahmad, H., Graybosch, R.A., Hein, G.L. and
Schwarzacher, T. (2016) Introgression of chromosome segments from multiple alien species in wheat breeding lines with wheat streak mosaic virus resistance. Heredity, 117, 114-123.
Aragón-Alcaide, L., Reader, S., Beven, A., Shaw, P., Miller, T. and Moore, G. (1997) Association of homologous chromosomes during floral development. Curr Biol, 7, 905–908.
Armstrong, S.J., Caryl, A.P., Jones, G.H. and Franklin, F.C. (2002) Asy1, a protein required for meiotic chromosome synapsis, localizes to axis-associated chromatin in Arabidopsis and Brassica. J Cell Sci, 115, 3645–3655.
Barakate, A., Higgins, J.D., Vivera, S., Stephens, J., Perry, R.M., Ramsay, L., Colas, I., Oakey, H., Waugh, R., Franklin, F.C.H., Armstrong, S.J., and Halpin, C. (2014) The synaptonemal complex protein ZYP1 is required for imposition of meiotic crossovers in barley. Plant Cell, 26, 729–740.
Bardhan, A. (2012) Meiotic chromosome interactions: nonhomologous centromere (un)coupling and homologous synapsis. ISRN Cell Biology, 2012, 1–9.
Bass, H.W., Riera-Lizarazu, O., Ananiev, E.V., Bordoli, S.J., Rines, H.W., Phillips, R.L., Sedat, J.W., Agard, D.A. and Cande, W.Z. (2000) Evidence for the coincident initiation of homolog pairing and synapsis during the telomere-clustering (bouquet) stage of meiotic prophase. J Cell Sci, 113 ( P, 1033–1042.
Bennett, M.D. (1979) Centromere arrangements in Triticum aestivum and their relationship to synapsis. Heredity, 43, 157.
Bennett, M.D., Rao, M.K., Smith, J.B. and Bayliss, M.W. (1973) Cell Development in the Anther, the Ovule, and the Young Seed of Triticum aestivum L. Var. Chinese Spring. Philosophical Transactions of the Royal Society B: Biological Sciences, 266, 39–81.
Bhullar, R., Nagarajan, R., Bennypaul, H., Sidhu, G.K., Sidhu, G., Rustgi, S., Wettstein, D. von and Gill, K.S. (2014) Silencing of a metaphase I-specific gene results in a phenotype similar to that of the Pairing homeologous 1 (Ph1) gene mutations. Proc Natl Acad Sci U S A, 111, 14187–14192.
Brar, G.A. and Amon, A. (2008) Emerging roles for centromeres in meiosis I chromosome segregation. Nat Rev Genet, 9, 899–910.
Caryl, A.P., Armstrong, S.J., Jones, G.H. and Franklin, F.C. (2000) A homologue of the yeast HOP1 gene is inactivated in the Arabidopsis meiotic mutant asy1. Chromosoma, 109, 62–71.

Sepsi et al. 2016. Meiotic chromosome dynamics in wheat. Plant Journal. Page 13.
Christophorou, N., Rubin, T. and Huynh, J.R. (2013) Synaptonemal complex components promote centromere pairing in pre-meiotic germ cells. PLoS Genet, 9, e1004012.
Chuong, H. and Dawson, D.S. (2010) Meiotic cohesin promotes pairing of nonhomologous centromeres in early meiotic prophase. Mol Biol Cell, 21, 1799–1809.
Church, K. and Moens, P.B. (1976) Centromere behavior during interphase and meiotic prophase in Allium fistulosum from 3-D, E.M. reconstruction. Chromosoma, 56, 249–263.
Corredor, E., Lukaszewski, A.J., Pachón, P., Allen, D.C. and Naranjo, T. (2007) Terminal regions of wheat chromosomes select their pairing partners in meiosis. Genetics, 177, 699–706.
Cox, A. (1993) Comparison of plant telomere locations using a PCR-generated synthetic probe. Ann Bot, 72, 239–247.
Dalal, Y., Furuyama, T., Vermaak, D. and Henikoff, S. (2007) Structure, dynamics, and evolution of centromeric nucleosomes. Proc Natl Acad Sci U S A, 104, 15974–15981.
Dawe, R.K. and Henikoff, S. (2006) Centromeres put epigenetics in the driver’s seat. Trends Biochem Sci, 31, 662–669.
Dawe, R.K., Sedat, J.W., Agard, D.A. and Cande, W.Z. (1994) Meiotic chromosome pairing in maize is associated with a novel chromatin organization. Cell, 76, 901–912.
Dawson, D.S., Murray, A.W. and Szostak, J.W. (1986) An alternative pathway for meiotic chromosome segregation in yeast. Science, 234, 713–717.
Dernburg, A.F., Sedat, J.W. and Hawley, R.S. (1996) Direct evidence of a role for heterochromatin in meiotic chromosome segregation. Cell, 86, 135–146.
Dernburg, Sedat, A.F., Cande, J.W., Bass, Z.W. and W, H.A.N.K. (1995) Cytology of Telomeres. In: Telomeres (ed. E.H. Blackburn and C.W. Greider), Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, 295–337.
Falk, J.E., Chan, A.C., Hoffmann, E. and Hochwagen, A. (2010) A Mec1- and PP4-dependent checkpoint couples centromere pairing to meiotic recombination. Dev Cell, 19, 599–611.
Gelei, J. (1921) Weitere Studien über die Oogenese des Dendrocoelum lacteum. II. Die Längskonjugation der Chromosomen. Archiv für Zellforschung, 16, 88–169.
Gladstone, M.N., Obeso, D., Chuong, H. and Dawson, D.S. (2009) The synaptonemal complex protein Zip1 promotes bi-orientation of centromeres at meiosis I. PLoS Genet, 5, e1000771.
Griffiths, S., Sharp, R., Foote, T.N., Bertin, I., Wanous, M., Reader, S., Colas, I. and Moore, G. (2006) Molecular characterization of Ph1 as a major chromosome pairing locus in polyploid wheat. Nature, 439, 749–752.
Hayashi, A., Ogawa, H., Kohno, K., Gasser, S.M. and Hiraoka, Y. (1998) Meiotic behaviours of chromosomes and microtubules in budding yeast: relocalization of centromeres and telomeres during meiotic prophase. Genes Cells, 3, 587–601.
Henikoff, S. and Furuyama, T. (2012) The unconventional structure of centromeric nucleosomes. Chromosoma, 121, 341–352.
Heslop-Harrison, J.S. (2000) Comparative genome organization in plants: from sequence and markers to chromatin and chromosomes. Plant Cell, 12, 617–636.
Heslop-Harrison, J.S., Murata, M., Ogura, Y., Schwarzacher, T. and Motoyoshi, F. (1999) Polymorphisms and genomic organization of repetitive DNA from centromeric regions of Arabidopsis chromosomes. Plant Cell, 11, 31–42.
Heslop-Harrison, J.S. and Schwarzacher, T. (2011) Organisation of the plant genome in chromosomes. Plant J, 66, 18–33.
Higgins, J.D., Perry, R.M., Barakate, A., Ramsay, L., Waugh, R., Halpin, C., Armstrong, S.J. and Franklin, F.C. (2012) Spatio-temporal asymmetry of the meiotic program underlies the predominantly distal distribution of meiotic crossovers in barley. Plant Cell, 24, 4096–4109.
Higgins, J.D., Sanchez-Moran, E., Armstrong, S.J., Jones, G.H. and Franklin, F.C. (2005) The Arabidopsis synaptonemal complex protein ZYP1 is required for chromosome synapsis and normal fidelity of crossing over. Genes Dev, 19, 2488–2500.

Sepsi et al. 2016. Meiotic chromosome dynamics in wheat. Plant Journal. Page 14.
Holm, P.B. (1986) Chromosome pairing and chiasma formation in allohexaploid wheat, Triticum aestivum analyzed by spreading of meiotic nuclei. Carlsberg Res. Commun, 51, 239–294.
Holm, P.B. (1988) Chromosome pairing and synaptonemal complex formation in hexaploid wheat, monosomic for chromosome 5B. Carlsberg Res Commun, 53, 57–89.
Houben, A., Demidov, D., Gernand, D., Meister, A., Leach, C.R. and Schubert, I. (2003) Methylation of histone H3 in euchromatin of plant chromosomes depends on basic nuclear DNA content. Plant J, 33, 967–973.
Houben, A. and Schubert, I. (2003) DNA and proteins of plant centromeres. Curr Opin Plant Biol, 6, 554–560.
Ines, O. Da and White, C.I. (2015) Centromere Associations in Meiotic Chromosome Pairing. Annu Rev Genet, 49, 95–114.
Jenkins, G. (1983) Chromosome pairing in triticum aestivum cv. Chinese Spring. Carlsberg Res Commun, 48, 255–283.
Jiang, J., Birchler, J., Parrott, W. and Kelly Dawe, R. (2003) A molecular view of plant centromeres. Trends Plant Sci, 8, 570–575.
Jin, Q., Trelles-Sticken, E., Scherthan, H. and Loidl, J. (1998) Yeast nuclei display prominent centromere clustering that is reduced in nondividing cells and in meiotic prophase. J Cell Biol, 141, 21–29.
Karpen, G.H., Le, M.H. and Le, H. (1996) Centric heterochromatin and the efficiency of achiasmate disjunction in Drosophila female meiosis. Science, 273, 118–122.
Kemp, B., Boumil, R.M., Stewart, M.N. and Dawson, D.S. (2004) A role for centromere pairing in meiotic chromosome segregation. Genes Dev, 18, 1946–1951.
Khoo, K.H., Able, A.J. and Able, J.A. (2012) The isolation and characterisation of the wheat molecular ZIPper I homologue, TaZYP1. BMC Res Notes, 5, 106.
Kihara, H. (1930) Genomanalyse bei Triticum und Aegilops. Cytologia, 1, 263–284. Kironmai, K.M., Muniyappa, K., Friedman, D.B., Hollingsworth, N.M. and Byers, B.
(1998) DNA-binding activities of Hop1 protein, a synaptonemal complex component from Saccharomyces cerevisiae. Mol Cell Biol, 18, 1424–1435.
Kleckner, N., Zickler, D., Jones, G.H., Dekker, J. Padmore, R., Henle, J., and Hutchinson, J. (2004) A mechanical basis for chromosome function. Proc Natl Acad Sci U S A 101:12592–12597.
Lermontova, I., Sandmann, M., Mascher, M., Schmit, A.C. and Chabouté, M.E. (2015) Centromeric chromatin and its dynamics in plants. Plant J, 83, 4–17.
Li, B., Choulet, F., Heng, Y., Weiwei Hao, Paux, E., Liu, Z., Yue, W., Jin, W., Feuillet, C. and Zhang, X. (2013) Wheat centromeric retrotransposons: the new ones take a major role in centromeric structure. Plant J, 73, 952–965.
Lohmiller, L.D., De Muyt, A., Howard, B., Offenberg, H.H., Heyting, C., Grelon, M and Anderson L.K. (2008) Cytological analysis of MRE11 protein during early meiotic prophase I in Arabidopsis and tomato. Chromosoma 117:277–288.
Maestra, B., Hans de Jong, J., Shepherd, K. and Naranjo, T. (2002) Chromosome arrangement and behaviour of two rye homologous telosomes at the onset of meiosis in disomic wheat-5RL addition lines with and without the Ph1 locus. Chromosome Res, 10, 655–667.
Martinez-Perez, E., Shaw, P., Aragon-Alcaide, L. and Moore, G. (2003) Chromosomes form into seven groups in hexaploid and tetraploid wheat as a prelude to meiosis. Plant J, 36, 21–29.
Martinez-Perez, E., Shaw, P. and Moore, G. (2001) The Ph1 locus is needed to ensure specific somatic and meiotic centromere association. Nature, 411, 204–207.
Martínez-Pérez, E., Shaw, P., Reader, S., Aragón-Alcaide, L., Miller, T. and Moore, G. (1999) Homologous chromosome pairing in wheat. J Cell Sci, 112, 1761–1769.
Martinez-Perez, E., Shaw, P.J. and Moore, G. (2000) Polyploidy induces centromere association. J Cell Biol, 148, 233–238.
Meuwissen, R.L., Meerts, I., Hoovers, J.M., Leschot, N.J. and Heyting, C. (1997) Human synaptonemal complex protein 1 (SCP1): isolation and characterization of the cDNA and chromosomal localization of the gene. Genomics, 39, 377–384.
Mikhailova, E.I., Phillips, D., Sosnikhina, S.P., Lovtsyus, A.V., Jones, R.N. and Jenkins, G.

Sepsi et al. 2016. Meiotic chromosome dynamics in wheat. Plant Journal. Page 15.
(2006) Molecular assembly of meiotic proteins Asy1 and Zyp1 and pairing promiscuity in rye (Secale cereale L.) and its synaptic mutant sy10. Genetics, 174, 1247–1258.
Molnár, I., Vrána, J., Farkas, A., Kubaláková, M., Cseh, A., Molnár-Láng, M. and Doležel, J. (2015) Flow sorting of C-genome chromosomes from wild relatives of wheat Aegilops markgrafii, Ae. triuncialis and Ae. cylindrica, and their molecular organization. Ann Bot 116: 189–200.
Obeso, D. and Dawson, D.S. (2010) Temporal characterization of homology-independent centromere coupling in meiotic prophase. PLoS ONE, 5, e10336.
Page, S.L. and Hawley, R.S. (2004) The genetics and molecular biology of the synaptonemal complex. Annu Rev Cell Dev Biol, 20, 525–558.
Phillips, D., Nibau, C., Wnetrzak, J. and Jenkins, G. (2012) High resolution analysis of meiotic chromosome structure and behaviour in barley (Hordeum vulgare L.). PLoS One. doi: 10.1371/journal.pone.0039539
Prieto, P., Santos, A.P., Moore, G. and Shaw, P. (2004) Chromosomes associate premeiotically and in xylem vessel cells via their telomeres and centromeres in diploid rice (Oryza sativa). Chromosoma, 112, 300–307.
Rabl, C (1885) Über Zelltheilung. Morphologisches Jahrbuch, 10, 214 – 330. Riley, R. and Chapman, V. (1958) Genetic control of the cytologically diploid behaviour of
hexaploid wheat. Nature, 182, 713–715. Ronceret, A. and Pawlowski, W.P. (2010) Chromosome dynamics in meiotic prophase I in
plants. Cytogenet Genome Res, 129, 173–183. Schwarzacher, T. (2003) Meiosis, recombination and chromosomes: a review of gene isolation
and fluorescent in situ hybridization data in plants. J Exp Bot, 54, 11–23. Schwarzacher, T. (1997) Three stages of meiotic homologous chromosome pairing in wheat:
cognition, alignment and synapsis. Sex Plant Reprod, 10, 324–331. Schwarzacher, T., Anamthawat-Jónsson, K., Harrison, G.E., Islam, A. K. M. R., Jia, J. Z.,
King, I. P., Leitch, A. R., Miller, T. E., Reader, S. M., Rogers, W. J., Shi, M. and Heslop-Harrison, J. S. (1992) Genomic in situ hybridization to identify alien chromosomes and chromosome segments in wheat. Theor Appl Genet, 84, 778–786.
Schwarzacher, T. and Heslop-Harrison, J.S. (1991) In situ hybridization to plant telomeres using synthetic oligomers. Genome, 34, 317–323.
Sears, E.R. (1939) Cytogenetic studies with polyploid species of wheat. I. Chromosomal aberrations in the progeny of a haploid of Triticum vulgare . Genetics, 24, 509–523.
Sears, E.R. (1976) Genetic control of chromosome pairing in wheat. Annu Rev Genet, 10, 31–51.
Tsubouchi, T. and Roeder, G.S. (2005) A synaptonemal complex protein promotes homology-independent centromere coupling. Science, 308, 870–873.
Verdaasdonk, J.S. and Bloom, K. (2011) Centromeres: unique chromatin structures that drive chromosome segregation. Nat Rev Mol Cell Biol, 12, 320–332.
Wen, R., Moore, G. and Shaw, P.J. (2012) Centromeres cluster de novo at the beginning of meiosis in Brachypodium distachyon. PLoS ONE, 7, e44681.
Zhang, H. and Dawe, R.K. (2012) Total centromere size and genome size are strongly correlated in ten grass species. Chromosome Res, 20, 403–412
Zickler, D. and Kleckner, N. (1999) Meiotic chromosomes: integrating structure and function. Annu Rev Genet, 33, 603-754.
Zickler, D. and Kleckner, N. (1998) The leptotene-zygotene transition of meiosis. Annu Rev Genet, 32, 619–697.

Sepsi et al. 2016. Meiotic chromosome dynamics in wheat. Plant Journal. Page 16.
Table 1: Centromere and telomere associations during meiosis as chromosomes pair and synapse
1) This study 2) This study and Rabl 1885; Holm 1986 3) Armstrong et al., 2002, Mikhailova et al., 2006, Higgins et al., 2012 4) Bennett et al., 1973; Holm 1986
Mid-Late interphase
Leptotene Early Zygotene Mid to late Zygotene
Pachytene Early Diplotene Late Diplotene
CENH3 -centromeres1)
Coupling
(15-33 foci)
Clustering
(4-16 foci)
Clustering resolves gradually and centromeres disperse
Synapsis starts Complete centromere synapsis (ZYP1 visible within CENH3 signal)
Desynapsis
Chromatin Orientation - centromeres and telomeres2)
Rabl orientation Centromeres at one pole
Telomeres at the opposite end of the nucleus
Centromere clusters at one pole
Bouquet forms with telomeres moving together
Rabl orientation relaxes
Bouquet is fully formed and telomeres are clustered
Centromeres are dispersed
Bouquet with telomeres clustered
Bouquet relaxes and telomeres disperse
No specific orientation
ASY1/ZYP1 locations indicating SC progression3)
Dispersed punctate foci of ASY1
Stretches of Asy1 localized to chromatin threads
ASY1 continuous along lateral elements
ZYP1 elongated foci at subtelomeres indicating initiation of synapsis. Punctate interstitial ZYP1 foci
Longer stretches of ZYP1 indicating elongation of interstitial synapsis
Synapsis complete, ZYP1 along whole length of homologues
Interstitial regions desynapse and ASY1 threads separate, ZYP1 breaks down
Desynapsis almost complete, ASY1 threats separate, ZYP1 disappeared
Number and position of nucleoli4)
Two to four smaller nucleoli enclosed within the nucleus
One – three (but mainly two) centrally to peripherally located nucleoli
One peripherally located big nucleolus
One peripherally located big nucleolus
Positive ASY1 staining1)
The nucleolus disappear at the end of diplotene

Sepsi et al. 2016. Meiotic chromosome dynamics in wheat. Plant Journal. Page 17.
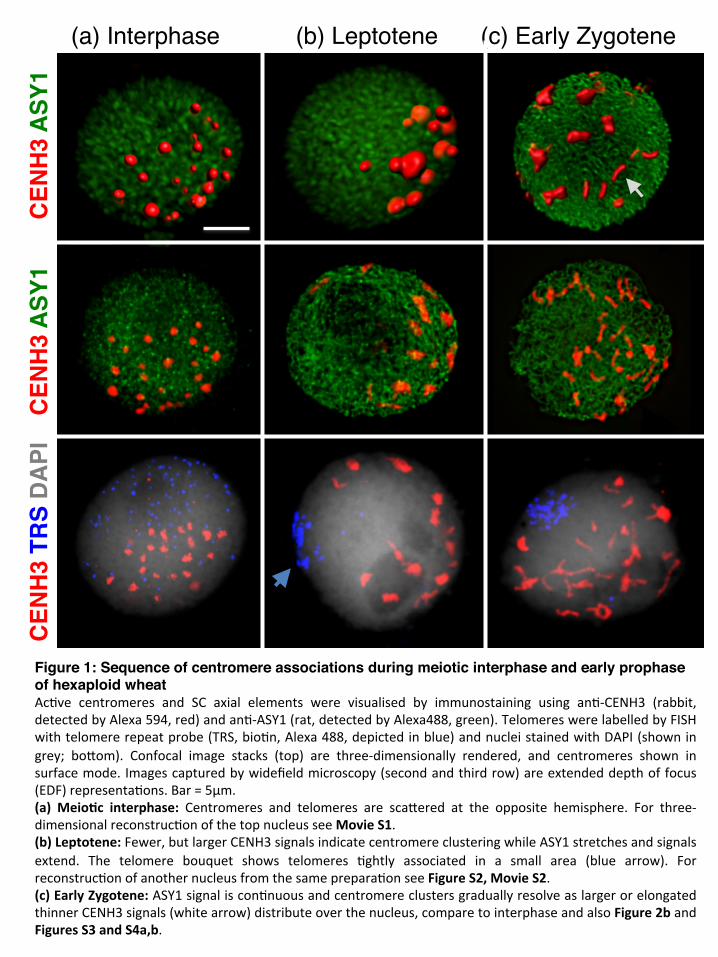
(a) Interphase (b) Leptotene (c) Early ZygoteneC
ENH
3 TR
S D
API
C
ENH
3 A
SY1
CEN
H3
ASY
1
Figure 1: Sequence of centromere associations during meiotic interphase and early prophase of hexaploid wheatAc#ve centromeres and SC axial elements were visualised by immunostaining using an#-CENH3 (rabbit,detectedbyAlexa594,red)andan#-ASY1(rat,detectedbyAlexa488,green).TelomereswerelabelledbyFISHwithtelomererepeatprobe(TRS,bio#n,Alexa488,depictedinblue)andnucleistainedwithDAPI(showningrey; boQom). Confocal image stacks (top) are three-dimensionally rendered, and centromeres shown insurfacemode.Imagescapturedbywidefieldmicroscopy(secondandthirdrow)areextendeddepthoffocus(EDF)representa#ons.Bar=5µm.(a)Meio(c interphase: Centromeres and telomeres are scaQered at the opposite hemisphere. For three-dimensionalreconstruc#onofthetopnucleusseeMovieS1.(b)Leptotene:Fewer,butlargerCENH3signalsindicatecentromereclusteringwhileASY1stretchesandsignalsextend. The telomere bouquet shows telomeres #ghtly associated in a small area (blue arrow). Forreconstruc#onofanothernucleusfromthesameprepara#onseeFigureS2,MovieS2.(c)EarlyZygotene:ASY1signaliscon#nuousandcentromereclustersgraduallyresolveaslargerorelongatedthinnerCENH3signals(whitearrow)distributeoverthenucleus,comparetointerphaseandalsoFigure2bandFiguresS3andS4a,b.
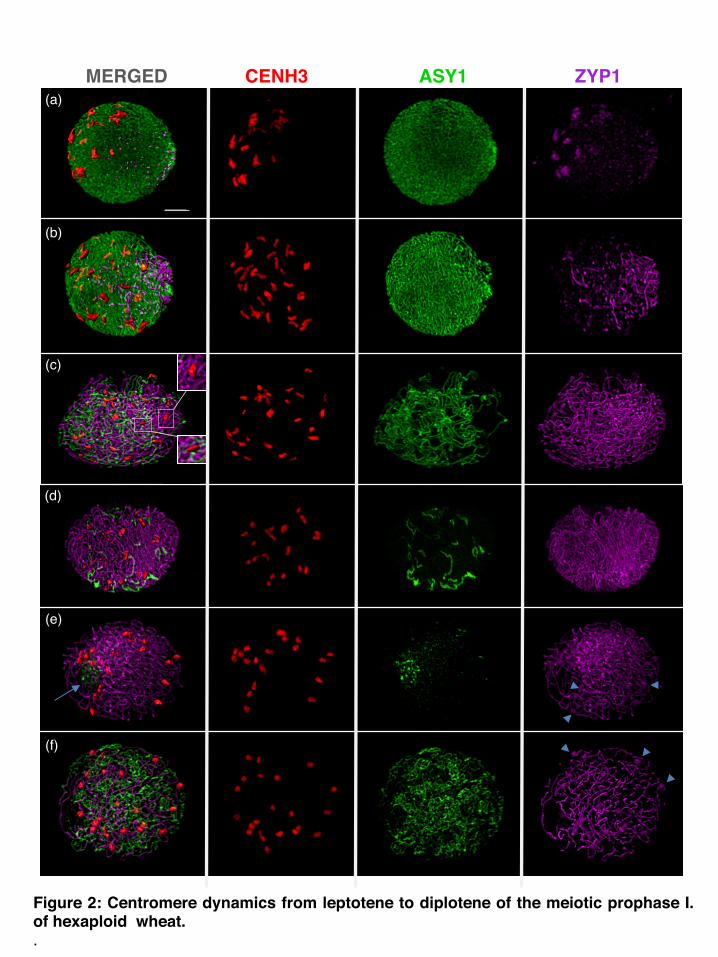
Figure 2: Centromere dynamics from leptotene to diplotene of the meiotic prophase I. of hexaploid wheat..
(a)
(b)
(c)
(d)
(e)
MERGED CENH3 ASY1 ZYP1
(f)

Figure 2: Centromere dynamics from leptotene to diplotene of the meiotic prophase I of hexaploid wheat. Triple immunstaining to visualise centromeres, and SC axial and central elements using anti- CENH3 (rabbit, detected by Alexa 594, red), anti-ASY1 (rat, detected by Alexa 488, green) and anti-ZYP1 (guinea pig, detected by Alexa 647, far-red shown in purple). Confocal image stacks are three-dimensionally reconstructed and centromeres are surface rendered on the left/merged panel. ZYP1 is surface rendered on (a) and (b) merged images. Bar = 5µm. An interpretive diagramme and additional high magnification images are shown in Figure 4.(a) Leptotene-zygotene transition: Polarized centromere clusters start to resolve coincident with continuous ASY-1 signal and the emergence of the first punctate ZYP1 foci at the opposite telomere pole. Dispersed ZYP1 signal is visible at the centromeres coinciding but not exactly the same as the CENH3 signal.(b) Early to mid zygotene: Centromeres lose their polarisation and are more dispersed (see also nucleus in Figure 1c) while short stretches of ZYP1 signal are visible at the telomere pole of the nucleus (see three dimensional reconstructed nucleus of Figure S3 and Movie S3). Punctate ZYP1 signal throughout the nucleus indicates interstitial initiation of synapsis.(c) Mid-zygotene: Centromeres are scattered within the nucleus, and chromosome arms gradually synapse as indicated by extended ZYP1 and reduced ASY1 signal. Unsynapsed centromeres col-localise with ASY1 (top inset) while synapsed centromeres col-localise with ZYP1 threads (bottom inset, and Figure 4e). (d) Late zygotene: Centromeres and exhibit an elongated signal together with ZYP1. Some interstitial regions show ASY1 signal indicating that synapsis is not complete. (e) Pachytene: ZYP1 forms extensive linear signal harbouring all centromeres (see Figure 4f, g). No ASY1 signal col-localises to the chromatin threads but with the nucleolus (arrow). Some diffuse ZYP1 signal is visible at centromeres (arrow heads; see Figure S6).(f) Diplotene: Desynapsis is initiated as ZYP1 disperses fractionally, and a fizzy ASY1 signal reappears. Centromeres col-localise with ZYP1 threads, indicating centromeric synapsis persists (see Figure 4h, i).
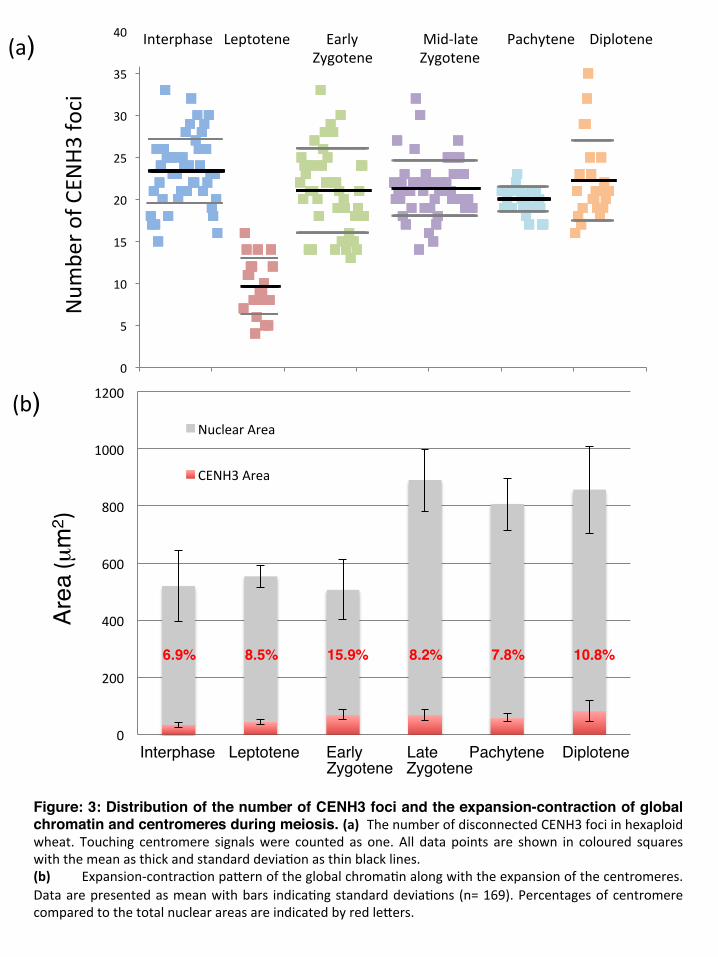
0
5
10
15
20
25
30
35
40 InterphaseLeptoteneEarlyMid-latePachyteneDiploteneZygoteneZygotene
Num
bero
fCEN
H3fo
ci
Figure: 3: Distribution of the number of CENH3 foci and the expansion-contraction of global chromatin and centromeres during meiosis. (a) ThenumberofdisconnectedCENH3fociinhexaploidwheat. Touching centromere signalswere counted as one.All data points are shown in coloured squareswiththemeanasthickandstandarddevia#onasthinblacklines.(b) Expansion-contrac#onpaQernoftheglobalchroma#nalongwiththeexpansionofthecentromeres.Dataarepresentedasmeanwithbars indica#ngstandarddevia#ons (n=169).PercentagesofcentromerecomparedtothetotalnuclearareasareindicatedbyredleQers.
0
200
400
600
800
1000
1200
NuclearArea
CENH3Area
6.9% 8.5% 15.9% 8.2% 7.8% 10.8%
Area
(µm
2 )
Interphase Leptotene Early Late Pachytene DiploteneZygotene Zygotene
(a)
(b)

(b) (d) (f) (h)
(c) (e) (g) (i)
Figure 4: Progression of centromere and arm synapsis from zygotene to diplotene of th(a) Cartoon diagram of the chromosome and centromere pairing(b–i) Single plane snapshots of confocal image stacks from meiocytes labelled by triple-immunostaining with anti-CENH3 (rabbit, detected by Alexa 594, red), anti-ASY1 (rat, detected by Alexa488, green) and anti-ZYP1 (guinea pig, detected by Alexa 647, far-red shown in purple). ZYP1 is not shown in (d) and CENH3 is not visualised in (g). Bar=5µmEarly of zygotene: Centromere clusters resolve individually (b) coincident with the initiation of subterminal synapsis at the opposite pole (also Figure 1c and 2a,b) seen as interconnections of lateral elements (ASY1) by the central element (ZYP1, orange arrow on (c). Interstitial regions show only punctate ZYP1 signal (c, white arrow). Centromeres harboured ASY1 strands, and are thus unpaired (b). Zygotene: Centromeres synapse following from interstitial synapsis (white arrow in d) and form larger associations (d; orange arrow). By late Zygotene unsynapsed lateral elements are mainly involved in interlocks (e; arrows ). Pachytene: Synapsis is complete at chromosome arms and centromeres that show compact CENH3 signals associated with ZYP1 (f, g; also Figure 2e).Diplotene: Desynapsis initiates at the interstitial regions while centromeres are still synapsed (h; white arrow). ZYP1 shows fragmental breakdown and the separating homologues are seen as fuzzy ASY1 signal (also Figure 2f.) Desynapsis is initiated at multiple points on the chromosome arms (i; arrows ). e meiotic prophase I. of hexaploid wheat.
MIDZYGOTENE PACHYTENE DIPLOTENE
Homologous chromosomes Centromeres ASY1 ZYP1
EARLY ZYGOTENE
(a)
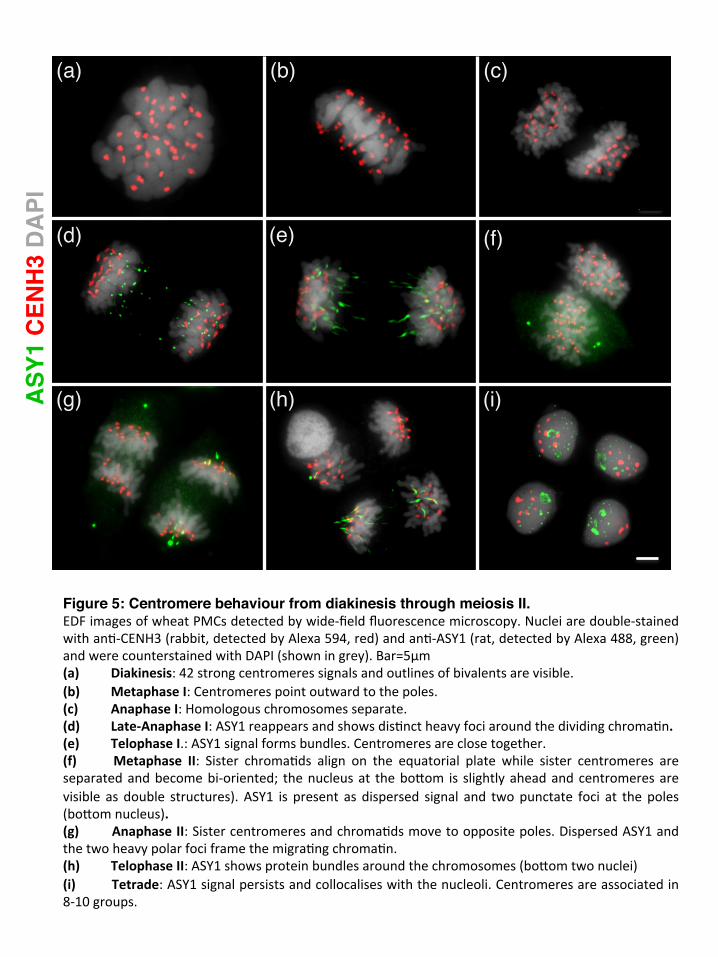
(a) (b) (c)
(d) (e) (f)
(g) (h) (i)ASY
1 C
ENH
3 D
API
Figure 5: Centromere behaviour from diakinesis through meiosis II.EDFimagesofwheatPMCsdetectedbywide-fieldfluorescencemicroscopy.Nucleiaredouble-stainedwithan#-CENH3(rabbit,detectedbyAlexa594,red)andan#-ASY1(rat,detectedbyAlexa488,green)andwerecounterstainedwithDAPI(showningrey).Bar=5µm(a) Diakinesis:42strongcentromeressignalsandoutlinesofbivalentsarevisible.(b) MetaphaseI:Centromerespointoutwardtothepoles.(c) AnaphaseI:Homologouschromosomesseparate.(d) Late-AnaphaseI:ASY1reappearsandshowsdis#nctheavyfociaroundthedividingchroma#n.(e) TelophaseI.:ASY1signalformsbundles.Centromeresareclosetogether.(f) Metaphase II: Sister chroma#ds align on the equatorial plate while sister centromeres areseparatedandbecomebi-oriented; thenucleusat theboQom is slightlyaheadandcentromeresarevisible as double structures). ASY1 is present as dispersed signal and twopunctate foci at thepoles(boQomnucleus).(g) AnaphaseII:Sistercentromeresandchroma#dsmovetooppositepoles.DispersedASY1andthetwoheavypolarfociframethemigra#ngchroma#n.(h) TelophaseII:ASY1showsproteinbundlesaroundthechromosomes(boQomtwonuclei)(i) Tetrade:ASY1signalpersistsandcollocaliseswiththenucleoli.Centromeresareassociatedin8-10groups.
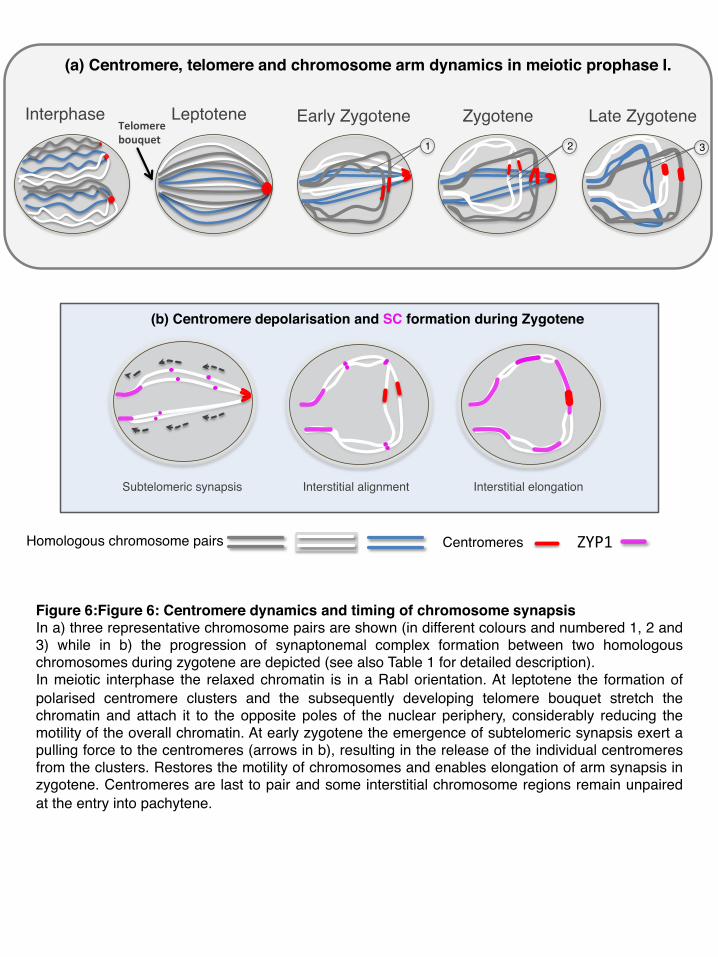
Figure 6:Figure 6: Centromere dynamics and timing of chromosome synapsis In a) three representative chromosome pairs are shown (in different colours and numbered 1, 2 and 3) while in b) the progression of synaptonemal complex formation between two homologous chromosomes during zygotene are depicted (see also Table 1 for detailed description).In meiotic interphase the relaxed chromatin is in a Rabl orientation. At leptotene the formation of polarised centromere clusters and the subsequently developing telomere bouquet stretch the chromatin and attach it to the opposite poles of the nuclear periphery, considerably reducing the motility of the overall chromatin. At early zygotene the emergence of subtelomeric synapsis exert a pulling force to the centromeres (arrows in b), resulting in the release of the individual centromeres from the clusters. Restores the motility of chromosomes and enables elongation of arm synapsis in zygotene. Centromeres are last to pair and some interstitial chromosome regions remain unpaired at the entry into pachytene.
(b) Centromere depolarisation and SC formation during Zygotene
Interphase Leptotene Zygotene Late ZygoteneTelomerebouquet
Homologous chromosome pairs Centromeres ZYP1
Early Zygotene 1 2 3
Subtelomeric synapsis Interstitial alignment Interstitial elongation
(a) Centromere, telomere and chromosome arm dynamics in meiotic prophase I.





























