Embed Size (px)
Citation preview

Central Projection of CalcitoninGene-Related Peptide (CGRP)- andSubstance P (SP)-Immunoreactive
Trigeminal Primary Neurons in the Rat
TOMOSADA SUGIMOTO,1* YOSHIAKI FUJIYOSHI,2 CHUN XIAO,1 YI-FEN HE,1
AND HIROYUKI ICHIKAWA1
1Department of Oral Anatomy II, Okayama University Dental School, 2-5-1 Shikata-cho,Okayama 700, Japan
2Department of Orthodontics, Okayama University Dental School, 2-5-1 Shikata-cho,Okayama 700, Japan
ABSTRACTSubstance P (SP) is implicated in transmission of primary afferent nociceptive signals. In
primary neurons, SP is colocalized with calcitonin gene-related peptide (CGRP), which isanother neuropeptide marker for small to medium primary neurons. CGRP coreleased withSP augments the postsynaptic effect of SP and thereby modulates the nociceptive transmis-sion. This study demonstrates the distribution of CGRP-like immunoreactivity (-ir) and SP-irin the lower brainstem of normal rats and after trigeminal rhizotomy or tractotomy at thelevel of subnucleus interpolaris (Vi). By comparing the results obtained from normal anddeafferented rats, we analyzed the central projection of trigeminal primary nociceptors. TheCGRP-immunoreactive (-ir) trigeminal primaries projected to the entire rostrocaudal extentof the spinal trigeminal nucleus, the principal nucleus (PrV), the paratrigeminal nucleus(paraV), and the lateral subnucleus of solitary tract nucleus (STN) on the ipsilateral side. Thetrigeminal primaries projecting to the spinal trigeminal nucleus, paraV and STN alsocontained SP-ir. The ipsilateral trigeminal primaries were the exclusive source of CGRP-irterminals in the PrV, the Vi and the dorsomedial nucleus within the subnucleus oralis (Vo).The medullary dorsal horn (MDH) and the lateral edge of Vo received convergent CGRP-irprojection from the ipsilateral trigeminal primaries and other neurons. The glossopharyngealand vagal primaries are candidates for the source of CGRP-ir projection to the Vo and theMDH, while the dorsal root axons supply the MDH with CGRP-ir terminals. In addition,contralateral primary neurons crossing the midline appear to contain CGRP and to terminatein the MDH. J. Comp. Neurol. 378:425–442, 1997. r 1997 Wiley-Liss, Inc.
Indexing terms: trigeminal nuclei; rhizotomy; tractotomy; nociception; orofacial pain
The central projection of primary afferent neurons hasbeen extensively investigated in both the spinal andtrigeminal nervous systems. With the advance in immuno-histochemistry, various neuropeptides have been found inthe spinal primary neurons. Because these neuropeptidesare synthesized in the cell body and are transported alongthe axon to the central terminal arbors, the distribution ofthese neuropeptides within the central nervous systemreflects the central projection of primary neurons whichcontain them. Moreover, the primary neurons can beclassified into several subpopulations on the basis of theneuropeptide content. Some of these subpopulations ex-hibit distinct morphological characteristics and may beassigned to specific functions. Therefore, a detailed study
of the central projection of primary neurons, with differentneuropeptides, will add much to the existing knowledgeabout the central connection and function of primaryneurons.Calcitonin gene-related peptide (CGRP), first reported
as an alternative expression product of the gene encodingcalcitonin (Amara et al., 1982, 1985; Rosenfeld et al., 1983,1984), is widely distributed in the central and peripheral
*Correspondence to: Dr. T. Sugimoto, SecondDepartment of OralAnatomy,Okayama University Dental School, 2-5-1 Shikata-cho, Okayama 700,Japan.Received 2 July 1996; Revised 9 October 1996; Accepted 22 October 1996
THE JOURNAL OF COMPARATIVE NEUROLOGY 378:425–442 (1997)
r 1997 WILEY-LISS, INC.

sensory nervous systems (Gibson et al., 1984; Skofitschand Jacobowitz, 1985a; Colin and Kruger, 1986; Peterfre-und and Vale, 1986; Carlton et al., 1987; Kuwayama et al.,1987; Silverman and Kruger, 1987, 1989; Chung et al.,1988; Harmon et al., 1988; Kruger et al., 1988a, b; Carltonand Hayes, 1989; Traub et al., 1989, 1990; Hokfelt, 1991;Quartu et al., 1992; Henry et al., 1996). The CGRP-likeimmunoreactivity (CGRP-ir) is found in a large proportionof small to medium-sized primary neuronal cell bodies(Mason et al., 1984; Skofitsch and Jacobowitz, 1985b;Goodman and Iversen, 1986; McCulloch et al., 1986; Ju etal., 1987) but is rare in autonomic ganglia (Rosenfeld et al.,1983; Landis and Fredieu, 1986). In peripheral tissues,CGRP-ir is detected in sensory axons that supply vascula-ture and epithelia with free nerve endings (Gibbins et al.,1985; Lee et al., 1985b; Lundberg et al., 1985; Rodrigo etal., 1985; Landis and Fredieu, 1986). Most CGRP-immunoreactive (CGRP-ir) peripheral axons are eitherunmyelimated or finelymyelinated (Yamamoto and Senba,1990) and sensitive to neonatal capsaicin (Gibbins et al.,1985; Rodrigo et al., 1985; Skofitsch and Jacobowitz,1985b; Hammond and Ruda, 1991). In the spinal cord,CGRP-ir is particularly intense in laminae I and II, andlaminae V and X also contain CGRP-ir (Rosenfeld et al.,1983; Gibson et al., 1984). Further, in vitro inhibition ofthe degradation of substance P (SP) by CGRP (LeGreves etal., 1985) and augmentation of C-fiber-mediated spinalreflex by intrathecally applied CGRP (Xu et al., 1990)suggested its neuromodulator role in the processing ofnociceptive information. Together, with a fact that CGRP-ircolocalizes with candidates for markers for primary noci-ceptors such as SP (Wiesenfeld-Hallin et al., 1984; Gibbinset al., 1985; Lee et al., 1985a, b; Lundberg et al., 1985;Landis and Fredieu, 1986), the above characteristics ofCGRP-ir suggested that at least part of CGRP-ir primaryneurons are implicated in nociception. It should be noted,however, that SP, but not CGRP, intrathecally applied tothe spinal cord evokes pain-related behavior (Wiesenfeld-Hallin et al., 1984; Gamse and Saria, 1986). Therefore, aquestion remains for the nociceptive role of CGRP-irprimary neurons without coexpression of SP.The purpose of this study was to analyze the central
projection of CGRP-ir and SP-ir trigeminal primary neu-rons to the lower brainstem (the brainstem sensory trigemi-nal nuclear complex-BSTC and surrounding structures) ofrats. Because nonprimary neurons may also express thepeptides, it is necessary to ascertain that the immunoreac-tivities observed in the normal brainstem are of theprimary neuronal origin. We therefore performed thetrigeminal rhizotomy to destroy all of the trigeminalprimary afferent axons on one side. A similar study hasbeen reported for the cat, but not for the rat (Tashiro et al.,1991; Henry et al., 1996). In a few rats, we transected thespinal trigeminal tract (TrV), instead of the root, at thelevel of the subnucleus interpolaris (Vi) to destroy thenontrigeminal as well as trigeminal peptidergic axons thatmay descend in the TrV which project to the caudalmedulla.
MATERIALS AND METHODS
A total of 13 male adult Sprague-Dawley rats (bodyweight 300–350 g) were used. They were deeply anesthe-
tized with ether and transvascularly perfused with 50 mlof saline followed by 500 ml of a fixative containing 4%formaldehyde (prepared fresh from paraformaldehyde)and 0.1 M sodium phosphate buffer (pH 7.4). The brain-stemwas excised, further fixed in a fresh volume of fixativeovernight at room temperature, cryoprotected by immers-ing in 20% sucrose in a phosphate-buffered saline at 4°Covernight, and cryosectioned at 50 µm in thickness. Twoalternate series of frontal sections were obtained fromeach rat and stained for CGRP-ir and SP-ir by an ABC(avidin-biotin-horseradish peroxidase complex) method.Sections were incubated with rabbit anti-CGRP serum(1:30,000, Cambridge Research Biochemicals, Wilming-ton, DE) or rabbit anti-SP serum (1:50,000, Peninsula,USA) for 3 days at 4°C followed by incubation withbiotinylated anti-rabbit IgG and ABC complex (VectastainABC kit, Vector Laboratories, Burlingame, CA). Antigen-antibody reaction products were visualized by a nickelammonium sulfate-intensified diaminobenzidine (DAB)histochemistry. The sections were usually incubated in a(regular) DAB medium containing 0.02% DAB, 0.0005%H2O2 and 0.08% nickel ammonium sulfate in Tris-bufferedsaline (pH 7.3). Some sections were incubated with ahalf-strengthmedium that contained 0.01%DAB, 0.0003%H2O2 and 0.08% nickel ammonium sulfate. The specificityof the primary antisera used has been described elsewhere(Ichikawa et al., 1990).In addition to five normal rats that were examined
without trigeminal primary deafferentation, some ratsunderwent a unilateral trigeminal rhizotomy 2 days(n 5 1), 3 days (n 5 1), 1 week (n 5 3), or 2 weeks (n 5 1)prior to perfusion fixation. They were anesthetized byintraperitoneal injection with a mixture of ethyl carba-mate (650 mg/kg body weight) and pentobarbital sodium(20 mg/kg body weight). The anesthetics were supple-mented arbitrarily for maintenance of the deep anestheticlevel. The rat was attached to a stereotaxic frame andcraniotomized to expose the superolateral aspect of thecerebral cortex. The cortex was dislocated medially toexpose the trigeminal ganglion and the sensory and themotor roots were cut with iridectomy scissors just anteriorto the petrosal ridge. Following the scheduled survivalperiods, the rats were reanesthetized with ether andperfusion-fixed for histological examination as describedabove. Completeness of the transection was confirmed bypostmortem examination by using an operation micro-scope.Two additional rats underwent either a TrV tractotomy
alone or a combined TrV tractotomy and upper cervicalhemicordotomy ipsilateral to the tractotomy. One weeklater, the rats were killed and the medulla and the spinalcord were processed for immunohistochemistry as de-scribed above. The exact location of the lesion was con-firmed in histological preparations. In the rat with thetractotomy alone, the lesion included the entire TrV andthe BSTC around the junction of the medullary dorsalhorn (MDH) and the Vi. In the other rat, the rostral lesionincluded the entire TrV and the adjacent BSTC at themid-Vi level, while the caudal one included the cervicaldorsal horn (CDH) and the overlying Lissauer’s tract atthe junction of the first and second cervical segments. Thedorsal rootlet was not observed entering the spinal cordrostral to the site of cordotomy.
426 T. SUGIMOTO ET AL.

RESULTS
When incubated with a regular-strength DAB medium,extremely dense DAB precipitates standing for CGRP-ircovered the superficial dorsal horn and the overlying TrVcontralateral to the rhizotomy and of intact rats. In suchpreparations, even the border between the TrV and theneuropil could not be determined. Therefore, the followingmorphological description and Figure 1Aabout CGRP-ir inthese areas (normal and contralateral to the rhizotomy)were derived mainly from the sections stained with ahalf-strength DAB medium. The remainder of descriptionand Figures, including those for the TrV and the dorsalhorn ipsilateral to the rhizotomy and tractotomy, are allbased on the observations obtained from the sectionsstained with a regular DAB medium. Difference in stain-ing intensity was little for SP-ir between the two differentstrength of DABmedia.
CGRP-ir
CGRP-ir profiles observed in our preparations were cellbodies, proximal dendrites, axons and axon terminals. Inthe white matter, the CGRP-ir axon had a smooth contourwith a relatively uniform diameter, and branching andvaricosity were rare. In the gray matter, slightly wavyaxons with smooth contour were occasionally seen travel-ing in small bundles. Another type of profile frequentlyseen in the gray matter as presumed terminal arbors ofaxons. The terminal arbors repeated branching within ashort distance and bore varicosities. Therefore, we coulddistinguish terminals from axons en passage with somecertainty unless extremely dense stain obscured the mor-phological details.The effect of rhizotomy first appeared in axon terminals
at 2 or 3 days following the rhizotomy. Some of the affectedterminal arbors exhibited a sign of fragmentation; i.e.,strands interconnecting varicosities became thinner ordisappeared and, consequently, the granular appearancewas exaggerated. At these postoperative intervals, how-ever, morphological sign of degeneration was not detectedin the parent axons traveling in the TrV. By 1 weekpostoperative, CGRP-ir axons and terminals were greatlyreduced in the ipsilateral TrV and BSTC. In the ventralpart of MDH lamina IIo, only coarse CGRP-ir granuleswere sparsely scattered. Such CGRP-ir granules stillpersisted at 2 weeks postoperative, and a further progres-sion of degeneration during the second postoperative weekwas barely discernible.Although our results suggested that a small fraction of
CGRP-ir trigeminal primary neurons project to the contra-lateral MDH (see below), reduction of CGRP-ir as a resultof contralateral rhizotomy was not detected in any part ofthe examined brainstem, probably because the contralat-eral/ipsilateral ratio of primary projection was very small.Similarly, there were areas, such as the solitary tractnucleus (STN) and the CDH, that appeared to receiveconvergent input from trigeminal and nontrigeminal pri-mary neurons. In such areas, the reduction in CGRP-irwas only a relative one, and the effect of ipsilateralrhizotomy could be best substantiated by comparing withthe contralateral side of the same rat rather than withintact rats.CGRP-ir in the upper CDH and MDH. The TrV
caudal to the obex contained extremely intense CGRP-ir.Neither the individual axon nor the border between the
TrV and the underlying MDH was visible in sectionsstained with a regular-stength DAB medium (Fig. 1E).When stained with a half-strength medium, CGRP-iraxons were seen distributed throughout the entire dorso-ventral extent of TrV and more concentrated in the exter-nal part of the TrV (Fig. 1A). The rhizotomy of 1 or 2 weeksstanding drastically reduced the CGRP-ir in the ipsilateralTrV to such extent that the morphological details could beeasily made out after staining with a regular-strengthDABmedium (Fig. 1B). The dorsomedial part of TrV at therostral MDH level retained relatively many CGRP-iraxons. Contrarily, deprivation of CGRP-ir in the middleand ventral parts of the TrV was nearly complete at thesimilar levels (compare Fig. 1B with C). Progressivelycaudal, the number of persisting CGRP-ir axons increasedin the middle and ventral parts, while that in the dorsome-dial part decreased. Following TrV tractotomy (with andwithout cordotomy), the CGRP-ir axons in both the dorso-medial part and the rest of TrV almost completely disap-peared at the rostral MDH levels (Fig. 2A).In the upper CDH (caudal to the pyramidal decussation,
Fig. 7A) and MDH (Fig. 7B–D), very dense aggregations ofDAB precipitates filled laminae I and IIo (Fig. 1A). How-ever, the morphological detail was obscure even afterstaining with a half-strength DAB medium. For the samereason, the laminar border between I and II could not bedetermined with confidence. In lamina IIi, moderatelydense reaction was observed. Because of the lower density,morphological detail could bemade out more easily than inlaminae I and IIo. Many of the CGRP-ir profiles in laminaIIi were terminal arbors. In addition, single ir axons orbundles of a few penetrated lamina IIi and entered laminaIII (Fig. 1A). These axons sometimes exhibited varicoseappearance within laminae III and IV but their trajectorywas rather straight without branching. CGRP-ir otherthan these radially oriented axons was rare in laminae IIIand IV. Upon approaching the laminae IV/V border, thebundle of CGRP-ir axons broke up and individual axonsbegan to arborize. The branches extended along the lami-nar border and formed moderately dense terminal arborsin lamina V. In frontal sections, such CGRP-ir terminalarbors formed a few patchy clumps separated by neuropilfree from CGRP-ir (Fig. 1A).The above laminar distribution of CGRP-ir was basically
similar in the CDH andMDH. However, the CGRP-ir axonterminals in lamina V were fewer in the rostral MDH (Fig.7B–D). Some CGRP-ir terminals could be seen in thesubtrigeminal region of the ventral medullary reticularformation (Fig. 7B).CGRP-ir was greatly reduced in the upper CDH and
MDH across laminae I through V ipsilateral to the rhi-zotomy by 1 week postoperative. The normal pattern ofCGRP-ir was lost from the rostral half of the MDH exceptfor the dorsomedial edge. The CGRP-ir almost completelydisappeared from laminae I and IIi, and extremely denseCGRP-ir in lamina IIo was replaced with sparse, coarsegranular ir that may stand for degeneration debris (Fig.1B, C). The ir axon terminals in lamina V also disappearedfrom the middle and rostral MDH (Fig. 1B). At thedorsomedial edge of the MDH, however, a relatively nor-mal pattern of CGRP-ir was preserved; i.e., laminae I andIIo exhibited definitely reduced but still intense CGRP-irthat obscured the fine structure (Fig. 1B). The accentua-tion of residual CGRP-ir in the dorsomedial part persistedthroughout the entire rostrocaudal extent of the MDH, but
CGRP- AND SP-IR IN THE RAT TRIGEMINAL NUCLEI 427

Fig. 1. Calcitonin gene-related peptide-like immunoreactivity(CGRP-ir) in the midmedullary dorsal horn (MDH; A–C) and parame-dian areas at the levels of the pyramidal decussation (D, E). A and Eare contralateral to the 1-week rhizotomy; B and C are from the samesection of MDH ipsilateral to the 1-week rhizotomy; D is from anunrhizotomized rat. The section shown in A was stained with ahalf-strength diaminobenzidine (DAB)medium.A:CGRP-ir is densestin laminae I and IIo (asterisk) and moderate in lamina IIi (doubleasterisk). The network of CGRP-immunoreactive (-ir) terminal arbors(arrowheads) at the IV/V border is connected to fine axons penetratingmore superficial laminae.B:Although marked reduction by rhizotomycan be seen in the superficial laminae, relatively intense CGRP-irpersists in the dorsal part of the spinal trigeminal tract (TrV) and atthe dorsomedial part of the MDH laminae I and II (between arrows).The CGRP-ir terminal at the IV/V border has almost disappeared. C:In a more ventrolateral area of the MDH, only sparse CGRP-ir axonsin the TrV and granules in lamina IIo (arrow) remain following therhizotomy (orientation: dorsolateral to the top and ventrolateral to the
left).D, E:At the levels of pyramidal decussation, CGRP-ir axons thatleave the dorsomedial edge of the MDH reach the midline (brokenlines) just ventral to the gracile fascicle (GF). Some of these axons formterminals in the median accessory nucleus (D), while others cross themidline (E) and reach the dorsomedial edge of contralateral dorsalhorn. Following incubation with a regular-strength DAB medium,extremely intense CGRP-ir obscures the morphological details of theTrV and superficial MDH (asterisk in E). Scale bars 5 200 µm forA, B,E; 50 µm for C, D.
Fig. 2. CGRP-ir in the mid-(A) and caudal MDH (B) ipsilateral to acombined TrV tractotomy and cervical hemicordotomy of 1 weekstanding. Depletion of CGRP-ir in the dorsal part of the TrV and thedorsomedial part of the MDH (between arrows) is more complete thanafter the rhizotomy (compare Figs. 2A with 1B). The dorsomedial(between arrows) as well as more lateral parts of caudal MDH containsome fibrous CGRP-ir in laminae I and IIo (B). Scale bars 5 200 µm.
428 T. SUGIMOTO ET AL.

was progressively reduced caudally. In the caudal half ofthe MDH, apparently healthy axons and terminals gradu-ally increased in progressively caudal sections.In the rats with the TrV tractotomy, the reduction of
CGRP-ir in the rostral half of the MDHwas more completethan after the trigeminal rhizotomy; i.e., the dorsomedialpart of the MDH contained only sparse granular CGRP-irbut terminal arbors were rarely seen (Fig. 2A). The caudalMDH and the rostral CDH, however, contained somefibrous CGRP-ir. Some of these fibers persisted even afterthe cordotomy (Fig. 2B), suggesting that they were derivedfrom axons that crossed the midline.Besides the above CGRP-ir axons terminating within
the dorsal horn, some CGRP-ir axons were found to leavethe dorsal horn and reach the midline (Fig. 1D, E). Theseaxons formed bundles at themedial edge of the dorsal hornat and slightly caudal to the levels of pyramidal decussa-tion and traveled medially. They occasionally gave rise toterminal arbors in themedullarymedian accessory nucleus(Figs. 1D, 7B, C) or traveled across the midline toward themedial edge of the contralateral dorsal horn at the samerostrocaudal level (Fig. 1E). Following the rhizotomy,tractotomy and cordotomy, at least some CGRP-ir axonstermininating in the median accessory nucleus and cross-ing the midline at the level of pyramidal decussationpersisted (Fig. 1E for an example after the rhizotomy).CGRP-ir in the Vi and paratrigeminal nucleus
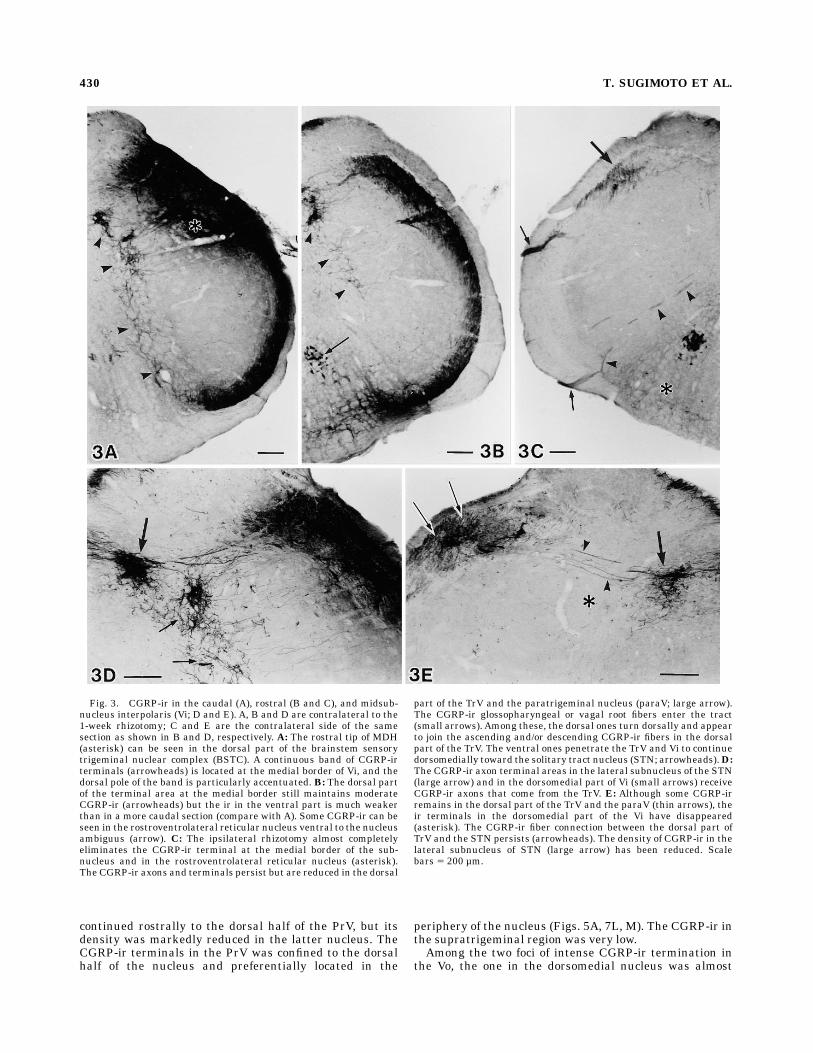
(paraV). At the Vi levels the TrV contained CGRP-ir,which was gradually reduced in intensity rostrad (Fig. 3A,B). Following the rhizotomy, the CGRP-ir was completelydepleted in the ventral half of the TrV but the dorsal partof the TrV retainedmany CGRP-ir axons (Fig. 3C, E). Suchresidual CGRP-ir was most abundant at the periobexlevels. The glossopharyngeal and vagal rootlets also con-tained many CGRP-ir axons and entered the brainstem atthe rostral Vi level (see Fig. 3C). Such CGRP-ir glossopha-ryngeal and vagal axons formed small bundles, penetratedthe inferior cerebellar peduncle and entered the TrV. Thebundles, which entered the dorsal half of the TrV, turneddorsally and appeared to either ascend or descend in thedorsal part of the TrV. More ventral bundles penetratedthe TrV and entered the Vi.At the periobex level, the MDH was gradually replaced
with the Vi (Fig. 7D, E). In frontal sections, the Vi firstappeared at the ventral pole of the BSTC and graduallyreplaced the MDH in progressively rostal sections. In thistransitional zone, the ventral part of the MDH was sepa-rated by the increasing bulk of Vi neuropil from the TrV.Atthe periobex levels, the dorsal parts of laminae I/II weredirectly connected to the paraV that was also filled withextremely dense CGRP-ir (Fig. 7E).The Vi including the dorsomedial nucleus was almost
devoid of CGRP-ir terminals except for its medial edge(Fig. 3A, B). Terminal arbors occupied a narrow areaextending between the medial edge of the subnucleus andthe adjacent bulbar reticular formation (Figs. 3A, 7F, G).Like for the terminals in lamina V of the dorsal horn,parent axons of these terminals traveled as single axons orbundles of a few axons through the overlying subnucleus(Fig. 3D). In the caudal part of the Vi, the terminal areaextended along the entire dorsoventral extent of themedial nuclear border and was densest at the dorsal pole.Rostrally, the above terminal area shrunk from the ventralpart, but the moderately dense terminals at the medialborder of the dorsomedial nucleus persisted and continued
to the terminal area in the same nucleus at the level ofsubnucleus oralis (Vo) (see below). The rostroventrolateralreticular nucleus that lay ventral to the nucleus ambiguusalso received moderately dense CGRP-ir terminal arborsderived from the TrV (Figs. 3B, 7G, H). The stain wasdensest at the caudal edge of the facial nucleus. As soon asthe facial nucleus appeared in the section, the CGRP-irterminal area in the reticular formation shifted dorsallyand then diminished.The CGRP-ir terminal arbors at the medial edge of the
Vi and their parent axons traversing the subnucleus werealmost completely eliminated by the trigeminal rhizotomy(Fig. 3C, E). CGRP-ir axon bundles that penetrated theventral part of the subnucleus toward the reticular forma-tion was not affected (Fig. 3C). The CGRP-ir terminals inthe rostroventrolateral reticular nucleus were also greatlyreduced (compare Fig. 3C with B).In addition to the above, a few thin bundles of CGRP-ir
axons were observed in each section of the rostral Vi. Thesebundles existed in the intact Vi but became clearly visibleafter removal of the dense CGRP-ir derived from theipsilateral trigeminal primary neurons by the rhizotomy.Among these, the bundles that traversed the dorsal part ofthe subnucleus appeared to bridge between the dorsal partof the TrV and the STN (Fig. 3D, E). On the other hand,ventral bundles penetrated the full thickness of the Vi andcontinued dorsomedially into the reticular formation. Atleast part of such ventral bundles were continuous fromthe glossopharyngeal or vagal root fibers that entered thebrainstem at the same rostrocaudal level (Fig. 3C for aview after the rhizotomy). The ventral bundles appeareddirected toward the STN but their final destination couldnot be confirmed.The paraV embedded in the TrV adjacent to the dorsal
part of Vi was normally filled with intense CGRP-ir (Fig.7E–G). Following the ipsilateral rhizotomy, relatively in-tense CGRP-ir in the paraV persisted at mid- and caudalVi levels. Although some CGRP-ir persisted in the morerostral paraV, the staining intensity was reduced (compareFig. 3C with E).CGRP-ir in the Vo and principal sensory trigeminal
nucleus(PrV). The TrV at the level of Vo and thetrigeminal sensory root contained many CGRP-ir axons(Figs. 4A, 5A). The general staining intensity was lowerthan at more caudal levels, and individual axons wereeasily distinguished after staining with a regular-strengthDAB medium. The ir axons were more abundant in theexternal part of the TrV, while such topographic preferenceof distribution was not noted in the root. Following therhizotomy, the CGRP-ir axons completely disappearedfrom the TrV and the root (Figs. 4B, 5B) except at thecaudal Vo level, where sparse CGRP-ir axons were re-tained in the dorsal part of the TrV.The ventral half of the normal Vo was free fromCGRP-ir.
On the other hand, the dorsal half of the Vo including thedorsomedial nucleus contained rich CGRP-ir terminalarbors (Figs. 4A, 7I–K). Two separate areas of the terminalstain were distinguished: the entire dorsomedial nucleusthat contained moderately dense CGRP-ir terminal ar-bors, and a narrow area subjacent to the TrV filled withdenser terminals and axons. The neuropil filling the gapbetween these two terminal areas did not contain richterminals but relatively thick bundles of CGRP-ir axonsthat bridged between the dorsomedial nucleus and theTrV. TheCGRP-ir terminal area in the dorsomedial nucleus
CGRP- AND SP-IR IN THE RAT TRIGEMINAL NUCLEI 429
continued rostrally to the dorsal half of the PrV, but itsdensity was markedly reduced in the latter nucleus. TheCGRP-ir terminals in the PrV was confined to the dorsalhalf of the nucleus and preferentially located in the
periphery of the nucleus (Figs. 5A, 7L, M). The CGRP-ir inthe supratrigeminal region was very low.Among the two foci of intense CGRP-ir termination in
the Vo, the one in the dorsomedial nucleus was almost
Fig. 3. CGRP-ir in the caudal (A), rostral (B and C), and midsub-nucleus interpolaris (Vi; D and E). A, B and D are contralateral to the1-week rhizotomy; C and E are the contralateral side of the samesection as shown in B and D, respectively. A: The rostral tip of MDH(asterisk) can be seen in the dorsal part of the brainstem sensorytrigeminal nuclear complex (BSTC). A continuous band of CGRP-irterminals (arrowheads) is located at the medial border of Vi, and thedorsal pole of the band is particularly accentuated. B: The dorsal partof the terminal area at the medial border still maintains moderateCGRP-ir (arrowheads) but the ir in the ventral part is much weakerthan in a more caudal section (compare with A). Some CGRP-ir can beseen in the rostroventrolateral reticular nucleus ventral to the nucleusambiguus (arrow). C: The ipsilateral rhizotomy almost completelyeliminates the CGRP-ir terminal at the medial border of the sub-nucleus and in the rostroventrolateral reticular nucleus (asterisk).The CGRP-ir axons and terminals persist but are reduced in the dorsal
part of the TrV and the paratrigeminal nucleus (paraV; large arrow).The CGRP-ir glossopharyngeal or vagal root fibers enter the tract(small arrows). Among these, the dorsal ones turn dorsally and appearto join the ascending and/or descending CGRP-ir fibers in the dorsalpart of the TrV. The ventral ones penetrate the TrV and Vi to continuedorsomedially toward the solitary tract nucleus (STN; arrowheads).D:The CGRP-ir axon terminal areas in the lateral subnucleus of the STN(large arrow) and in the dorsomedial part of Vi (small arrows) receiveCGRP-ir axons that come from the TrV. E: Although some CGRP-irremains in the dorsal part of the TrV and the paraV (thin arrows), their terminals in the dorsomedial part of the Vi have disappeared(asterisk). The CGRP-ir fiber connection between the dorsal part ofTrV and the STN persists (arrowheads). The density of CGRP-ir in thelateral subnucleus of STN (large arrow) has been reduced. Scalebars 5 200 µm.
430 T. SUGIMOTO ET AL.
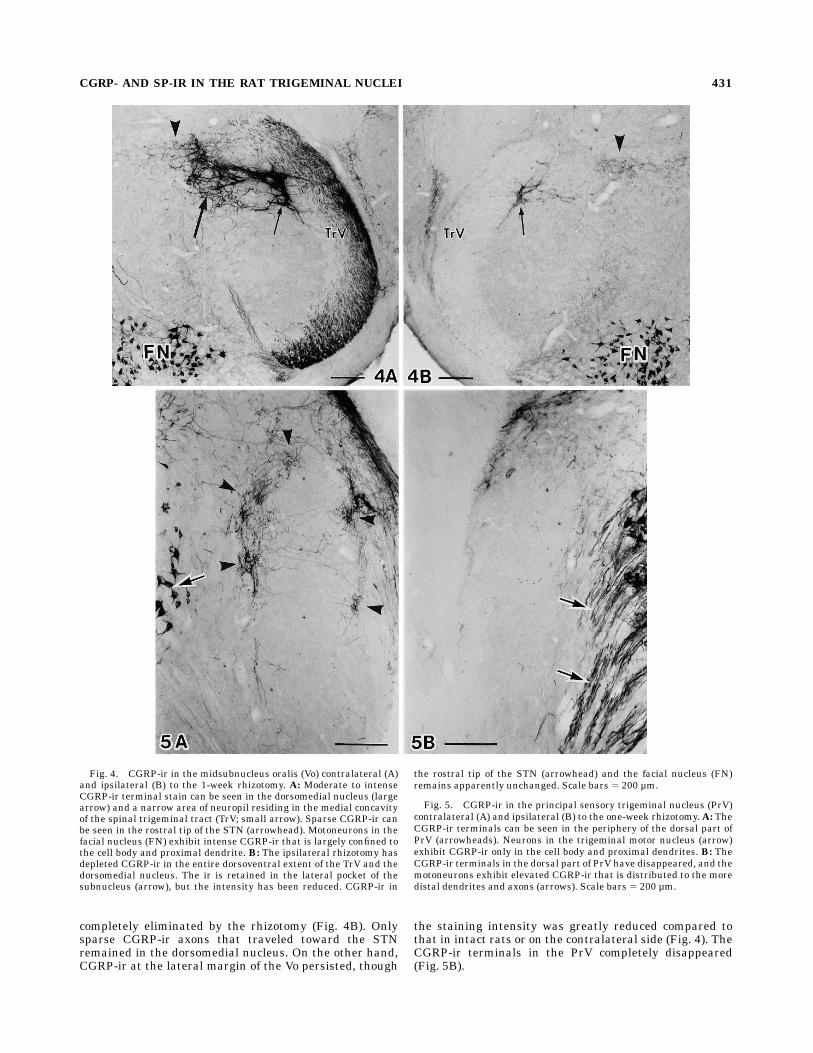
completely eliminated by the rhizotomy (Fig. 4B). Onlysparse CGRP-ir axons that traveled toward the STNremained in the dorsomedial nucleus. On the other hand,CGRP-ir at the lateral margin of the Vo persisted, though
the staining intensity was greatly reduced compared tothat in intact rats or on the contralateral side (Fig. 4). TheCGRP-ir terminals in the PrV completely disappeared(Fig. 5B).
Fig. 4. CGRP-ir in the midsubnucleus oralis (Vo) contralateral (A)and ipsilateral (B) to the 1-week rhizotomy. A: Moderate to intenseCGRP-ir terminal stain can be seen in the dorsomedial nucleus (largearrow) and a narrow area of neuropil residing in the medial concavityof the spinal trigeminal tract (TrV; small arrow). Sparse CGRP-ir canbe seen in the rostral tip of the STN (arrowhead). Motoneurons in thefacial nucleus (FN) exhibit intense CGRP-ir that is largely confined tothe cell body and proximal dendrite. B: The ipsilateral rhizotomy hasdepleted CGRP-ir in the entire dorsoventral extent of the TrV and thedorsomedial nucleus. The ir is retained in the lateral pocket of thesubnucleus (arrow), but the intensity has been reduced. CGRP-ir in
the rostral tip of the STN (arrowhead) and the facial nucleus (FN)remains apparently unchanged. Scale bars 5 200 µm.
Fig. 5. CGRP-ir in the principal sensory trigeminal nucleus (PrV)contralateral (A) and ipsilateral (B) to the one-week rhizotomy. A: TheCGRP-ir terminals can be seen in the periphery of the dorsal part ofPrV (arrowheads). Neurons in the trigeminal motor nucleus (arrow)exhibit CGRP-ir only in the cell body and proximal dendrites. B: TheCGRP-ir terminals in the dorsal part of PrV have disappeared, and themotoneurons exhibit elevated CGRP-ir that is distributed to the moredistal dendrites and axons (arrows). Scale bars 5 200 µm.
CGRP- AND SP-IR IN THE RAT TRIGEMINAL NUCLEI 431
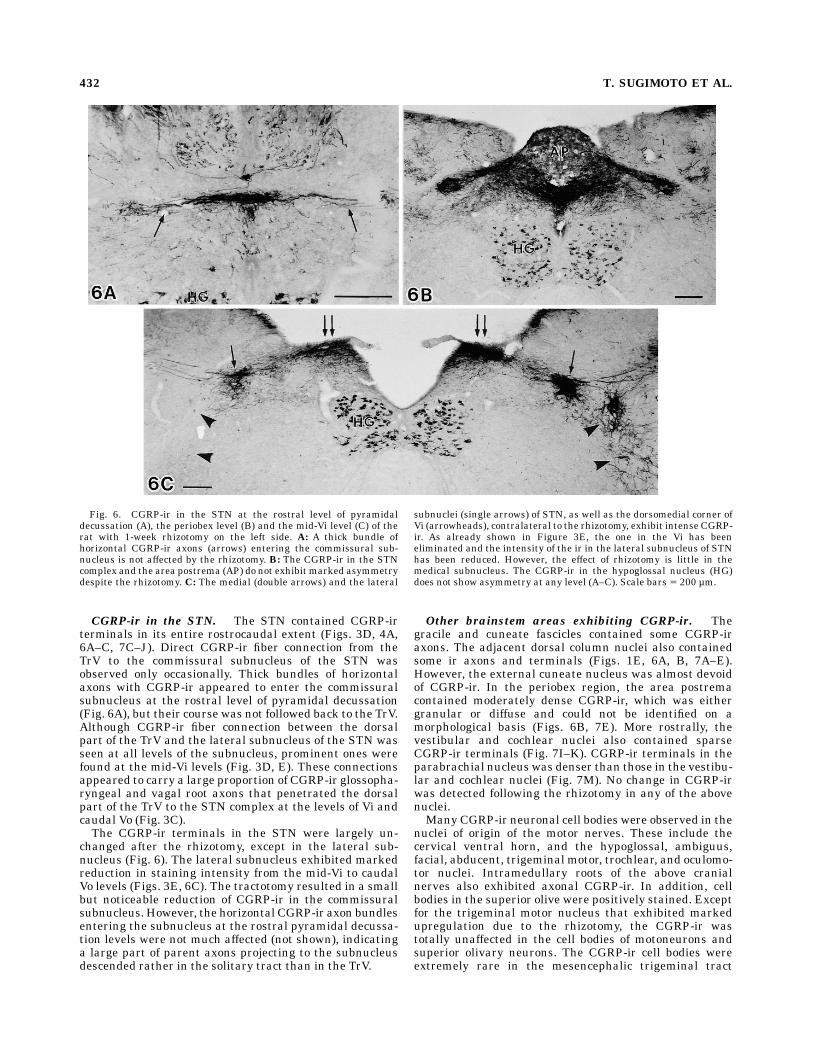
CGRP-ir in the STN. The STN contained CGRP-irterminals in its entire rostrocaudal extent (Figs. 3D, 4A,6A–C, 7C–J). Direct CGRP-ir fiber connection from theTrV to the commissural subnucleus of the STN wasobserved only occasionally. Thick bundles of horizontalaxons with CGRP-ir appeared to enter the commissuralsubnucleus at the rostral level of pyramidal decussation(Fig. 6A), but their course was not followed back to the TrV.Although CGRP-ir fiber connection between the dorsalpart of the TrV and the lateral subnucleus of the STN wasseen at all levels of the subnucleus, prominent ones werefound at the mid-Vi levels (Fig. 3D, E). These connectionsappeared to carry a large proportion of CGRP-ir glossopha-ryngeal and vagal root axons that penetrated the dorsalpart of the TrV to the STN complex at the levels of Vi andcaudal Vo (Fig. 3C).The CGRP-ir terminals in the STN were largely un-
changed after the rhizotomy, except in the lateral sub-nucleus (Fig. 6). The lateral subnucleus exhibited markedreduction in staining intensity from the mid-Vi to caudalVo levels (Figs. 3E, 6C). The tractotomy resulted in a smallbut noticeable reduction of CGRP-ir in the commissuralsubnucleus. However, the horizontal CGRP-ir axon bundlesentering the subnucleus at the rostral pyramidal decussa-tion levels were not much affected (not shown), indicatinga large part of parent axons projecting to the subnucleusdescended rather in the solitary tract than in the TrV.
Other brainstem areas exhibiting CGRP-ir. Thegracile and cuneate fascicles contained some CGRP-iraxons. The adjacent dorsal column nuclei also containedsome ir axons and terminals (Figs. 1E, 6A, B, 7A–E).However, the external cuneate nucleus was almost devoidof CGRP-ir. In the periobex region, the area postremacontained moderately dense CGRP-ir, which was eithergranular or diffuse and could not be identified on amorphological basis (Figs. 6B, 7E). More rostrally, thevestibular and cochlear nuclei also contained sparseCGRP-ir terminals (Fig. 7I–K). CGRP-ir terminals in theparabrachial nucleus was denser than those in the vestibu-lar and cochlear nuclei (Fig. 7M). No change in CGRP-irwas detected following the rhizotomy in any of the abovenuclei.Many CGRP-ir neuronal cell bodies were observed in the
nuclei of origin of the motor nerves. These include thecervical ventral horn, and the hypoglossal, ambiguus,facial, abducent, trigeminal motor, trochlear, and oculomo-tor nuclei. Intramedullary roots of the above cranialnerves also exhibited axonal CGRP-ir. In addition, cellbodies in the superior olive were positively stained. Exceptfor the trigeminal motor nucleus that exhibited markedupregulation due to the rhizotomy, the CGRP-ir wastotally unaffected in the cell bodies of motoneurons andsuperior olivary neurons. The CGRP-ir cell bodies wereextremely rare in the mesencephalic trigeminal tract
Fig. 6. CGRP-ir in the STN at the rostral level of pyramidaldecussation (A), the periobex level (B) and the mid-Vi level (C) of therat with 1-week rhizotomy on the left side. A: A thick bundle ofhorizontal CGRP-ir axons (arrows) entering the commissural sub-nucleus is not affected by the rhizotomy. B: The CGRP-ir in the STNcomplex and the area postrema (AP) do not exhibit marked asymmetrydespite the rhizotomy. C: The medial (double arrows) and the lateral
subnuclei (single arrows) of STN, as well as the dorsomedial corner ofVi (arrowheads), contralateral to the rhizotomy, exhibit intense CGRP-ir. As already shown in Figure 3E, the one in the Vi has beeneliminated and the intensity of the ir in the lateral subnucleus of STNhas been reduced. However, the effect of rhizotomy is little in themedical subnucleus. The CGRP-ir in the hypoglossal nucleus (HG)does not show asymmetry at any level (A–C). Scale bars 5 200 µm.
432 T. SUGIMOTO ET AL.

nucleus of normal rats. Following the rhizotomy, however,occasional cell bodies were seen with moderate to intenseCGRP-ir (see Yoshida et al., 1995 for a micrograph). SuchCGRP-ir mesencephalic trigeminal nuclear cells were rarecontralateral to the rhizotomy.
SP-ir
The SP-ir in the brainstem was finely granulated andthe ir profiles in many areas could not be identified on amorphological basis. Most of these SP-ir granules areconsidered to represent axons and axon terminals. NoSP-ir cell body was observed because colchicine treatment,which is necessary for visualization of SP-ir in the cellbody of many central neurons (Tessler et al., 1981), was notemployed in this study. Various amounts of granular SP-irwere found in many sensory and motor nuclei, and thereticular formationmedial to the BSTC. In the TrV and theBSTC, however, many SP-ir axons and terminal arborstook the form of solid fibrous stain or chain-like arrange-ment of SP-ir granules. Such morphologically identifiedSP-ir profiles in the BSTC were markedly reduced by 1week but not 3 days following the ipsilateral rhizotomy.However, reduction in general intensity of SP-ir inducedby the ipsilateral trigeminal rhizotomy was only a relativeone because many granular SP-ir remained apparentlyunchanged. Here again, SP-ir in all areas of the brainstemremained apparently unchanged following the contralat-eral trigeminal rhizotomy.As was the case for CGRP-ir, thechange during the second postoperative week was small;further reduction of SP-ir over that seen at 1 weekpostoperative or apparent recovery of it mediated byintrinsic central neurons, that reportedly occurred at alonger survival period (Tessler et al., 1980, 1981, 1984),was not noted. The following description concerns only theBSTC and the STN where asymmetry was caused by therhizotomy.SP-ir in the TrV. The distribution of SP-ir in the TrV
was similar to that of the CGRP-ir except that the stainingintensity was lower for the SP-ir. In the TrV, positive axonsappeared as chains of ir granules (Fig. 8D). The externalpart of the normal TrV contained moderate SP-ir through-out its entire rostrocaudal and dorsomedial extent (Figs.8A–D, 9A, 10A). Following the ipsilateral rhizotomy, SP-irwas totally depleted in the ventral half of the TrV rostral tothe obex (Figs. 9C, 10C), while the depletion in the dorsalhalf was incomplete caudal to the mid-Vo level (compareFigs. 10C, D with 8E–G, 9C, D). Progressively caudal fromthe obex level, the SP-ir increased in its density in both theventral and dorsal parts of the TrV (Fig. 8E–G) but wasweaker than on the contralateral unrhizotomized side.The glossopharyngeal and vagal axons also containedSP-ir (not shown).SP-ir in the upper CDH and MDH. Normal distribu-
tion of SP-ir within the CDH and MDHwas similar to thatof CGRP-ir. It was densest in laminae I and IIo, andweaker in lamina IIi (Fig. 8A–C). In these laminae, theintense staining obscured the morphological detail butsome torturous chains of ir granules probably standing forterminal arbors were observed (Fig. 8D). Similar terminalarbors were also seen in lamina V but were sparsercompared to the CGRP-ir ones in the same area (Fig. 8A,B). A few radially oriented SP-ir axons penetrated laminaeIII/IV, connecting the TrV and terminal arbors in lamina V.As was the case for CGRP-ir terminals, the SP-ir ones in
lamina V decreased in progressively rostral sections of theMDH.Following the rhizotomy, marked but incomplete deple-
tion of SP-ir was observed in laminae I and IIo ipsilaterally(compare Fig. 8A–C with E–G). In the caudal MDH, only aslight reduction in SP-ir was noted compared to thecontralateral side. More rostrally, the reduction was moreprofound and the morphologically identified SP-ir termi-nal arbors were greatly reduced (Fig. 8H). However, manySP-ir granules persisted even at the periobex level wherethe depletion was most prominent (Fig. 8G). The SP-iraxons in laminae III/IV and terminals in lamina V mostlydisappeared, leaving fine granular SP-ir (Fig. 8E,F)except at the caudal MDH and CDH levels, where at leastsome of these profiles survived the rhizotomy. The effectson SP-ir of TrV tractotomy and cordotomy were similar to,but less pronounced than those on CGRP-ir.In and around the commissural subnucleus of the STN
that contained dense granular SP-ir, parallel lines ofgranules probably standing for axons were found to travelhorizontally. These presumed SP-ir axons persisted follow-ing the rhizotomy, tractotomy and combined tractotomyand hemicordotomy (not shown).SP-ir in the Vi and paraV. The Vi was almost devoid
of SP-ir except at the dorsomedial border of the sub-nucleus, where the ir axons and terminal arbors formed acoarse network (Fig. 9A,B). Fine granular SP-ir wasscattered in this fibrous network. As was the case forCGRP-ir in the same area, the terminal arbors at thedorsomedial edge was separated by a narrow gap from theSTN that was filled with intense SP-ir. In the more ventralpart of the Vi, the SP-ir at the medial border was sparserandmostly granular. Such SP-ir could not be distinguishedfrom granular SP-ir that was seen in the parvicellularreticular nucleus just medial to the Vi. Occasional SP-iraxon bundles were seen traveling along the dorsal borderof the Vi toward the lateral subnucleus of the STN. The irterminal arbors at the dorsomedial edge of the Vi disap-peared after rhizotomy, but some granular ir persisted(Fig. 9C, D). Because of the moderate SP-ir in the adjacentparvicellular reticular nucleus, however, the effect of rhi-zotomy was not substantiated in the more ventral part ofthe medial border area. The rostroventrolateral reticularnucleus, as well as the more dorsal parvicellular reticularnucleus, contained many fine granular SP-ir, but the effectof rhizotomy was unclear.In the paraV, the normal pattern of distribution and the
reduction by the rhizotomy of SP-ir were essentially
Fig. 7. (overleaf). Schematic diagrams of the distribution of CGRP-irterminals in the normal BSTC and surrounding sensory nuclei. Thedark, moderate and light shades represent relative density of CGRP-irterminals in a descending order. Axons en passage and cell bodies arenot included. Numbers in parenthesis indicate distance in millimeterscaudal to the obex. Amb, ambiguus nucleus; AP, area postrema; AT,acoustic tubercle; CN, cuneate nucleus; ExC, external cuneate nucleus;FN, facial nucleus; FG, facial nerve genu; FR, facial nerve root; GF,gracile fascicle; GN, gracile nucleus; HG, hypoglossal nucleus; ICP,inferior cerebellar peduncle; IO, inferior olive; MDH, medullary dorsalhorn; MV, trigeminal motor nucleus; paraV, paratrigeminal nucleus;PrV, principal nucleus; PyrX, pyramidal decussation; rvl, rostroventro-lateral reticular formation; SCP, superior cerebellar peduncle; SO,superior olive; ST, solitary tract; TR, trigeminal root; TrV, spinaltrigeminal tract; III/IV, laminae III and IV of the spinal or medullarydorsal horn; Vi, subnucleus interpolaris; Vo, subnucleus oralis. Scalebar 5 1 mm.
CGRP- AND SP-IR IN THE RAT TRIGEMINAL NUCLEI 433
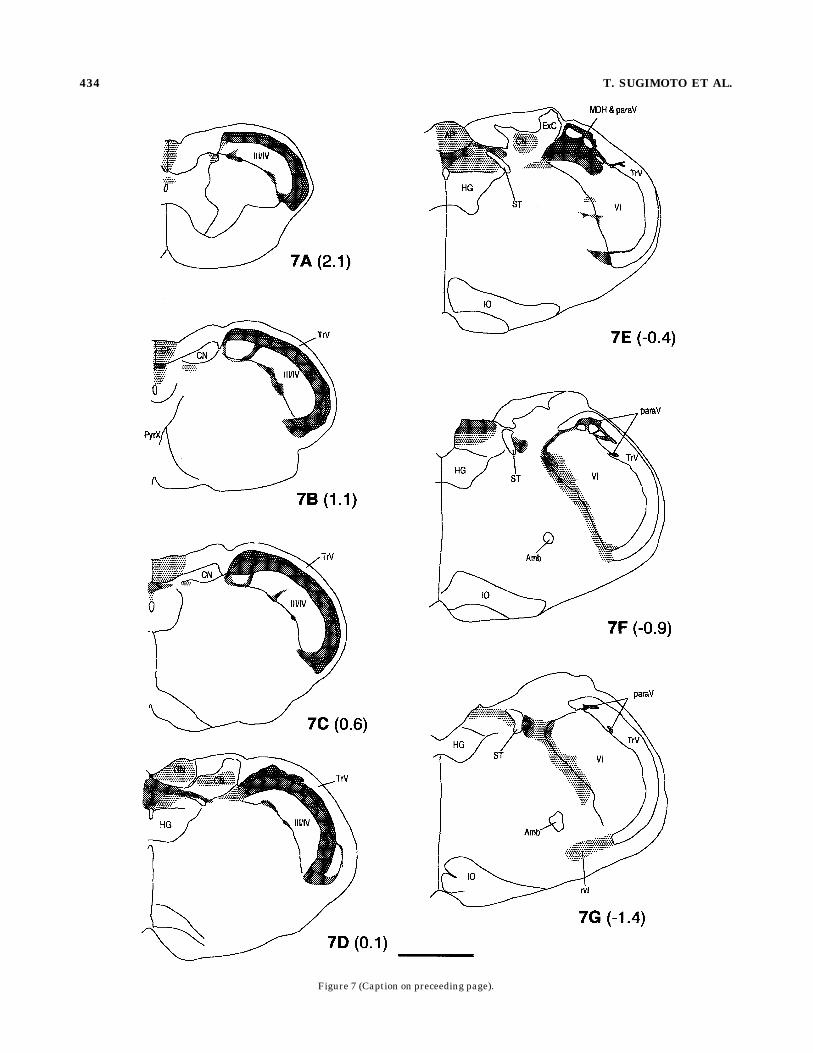
Figure 7 (Caption on preceeding page).
434 T. SUGIMOTO ET AL.
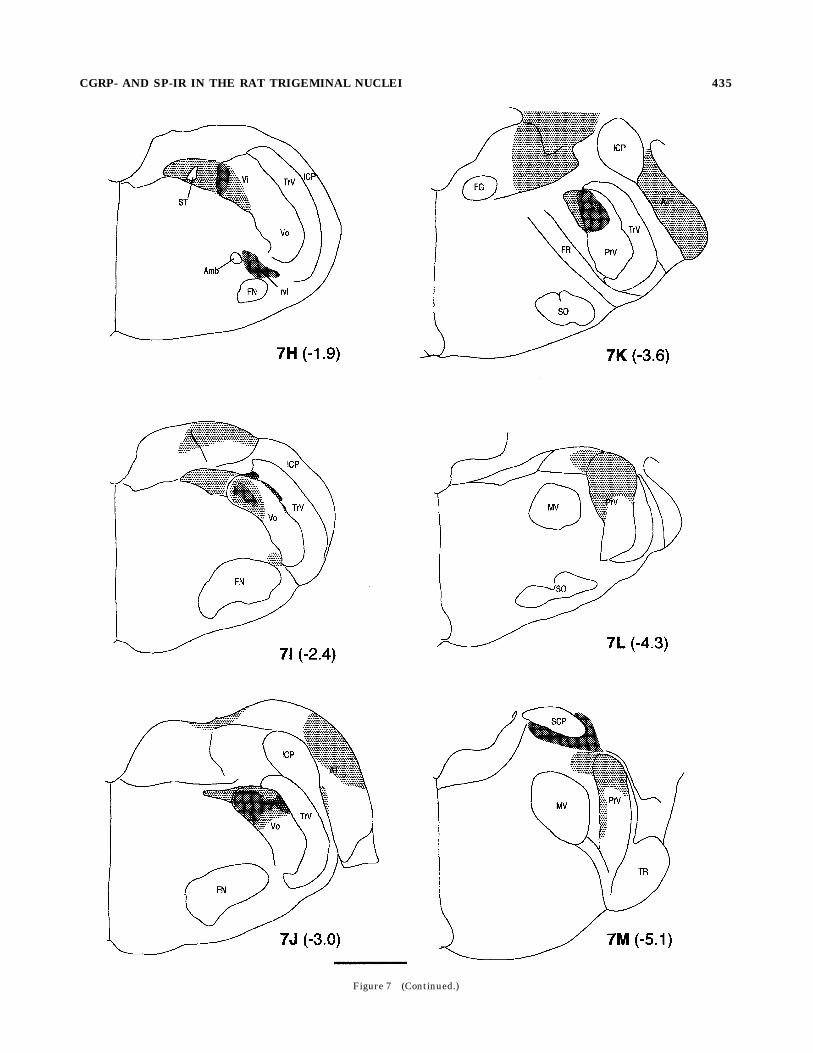
Figure 7 (Continued.)
CGRP- AND SP-IR IN THE RAT TRIGEMINAL NUCLEI 435
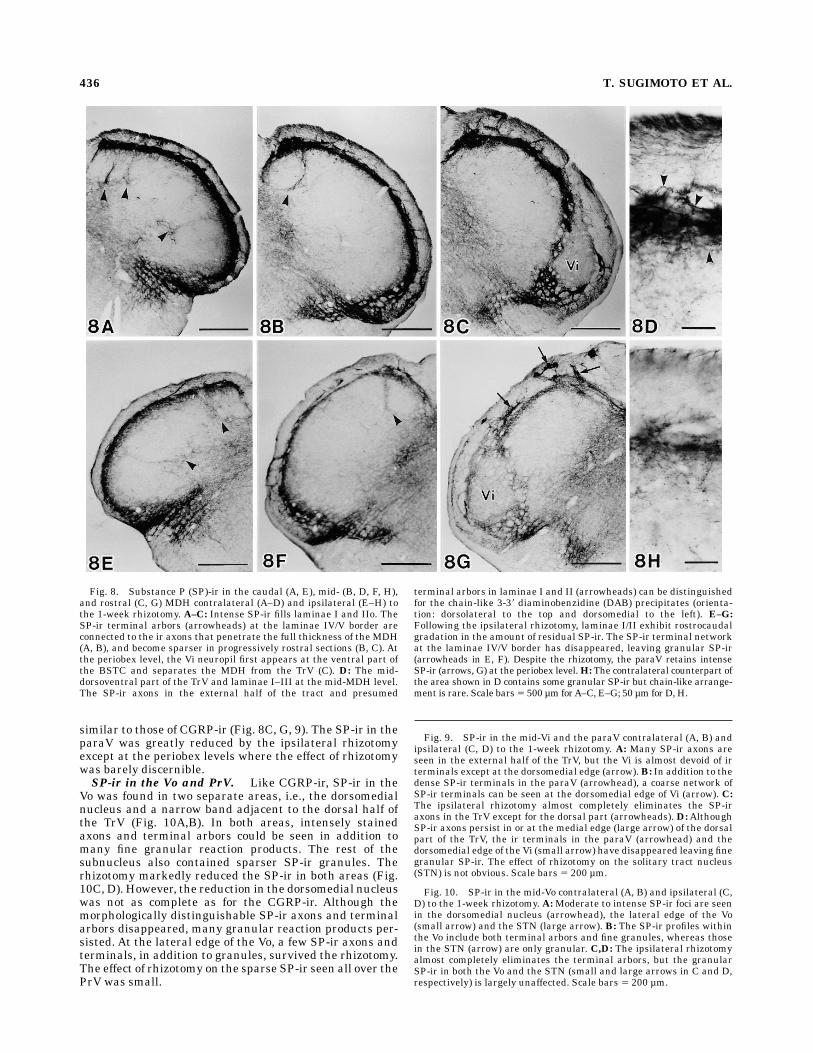
similar to those of CGRP-ir (Fig. 8C, G, 9). The SP-ir in theparaV was greatly reduced by the ipsilateral rhizotomyexcept at the periobex levels where the effect of rhizotomywas barely discernible.SP-ir in the Vo and PrV. Like CGRP-ir, SP-ir in the
Vo was found in two separate areas, i.e., the dorsomedialnucleus and a narrow band adjacent to the dorsal half ofthe TrV (Fig. 10A,B). In both areas, intensely stainedaxons and terminal arbors could be seen in addition tomany fine granular reaction products. The rest of thesubnucleus also contained sparser SP-ir granules. Therhizotomy markedly reduced the SP-ir in both areas (Fig.10C, D). However, the reduction in the dorsomedial nucleuswas not as complete as for the CGRP-ir. Although themorphologically distinguishable SP-ir axons and terminalarbors disappeared, many granular reaction products per-sisted. At the lateral edge of the Vo, a few SP-ir axons andterminals, in addition to granules, survived the rhizotomy.The effect of rhizotomy on the sparse SP-ir seen all over thePrV was small.
Fig. 8. Substance P (SP)-ir in the caudal (A, E), mid- (B, D, F, H),and rostral (C, G) MDH contralateral (A–D) and ipsilateral (E–H) tothe 1-week rhizotomy. A–C: Intense SP-ir fills laminae I and IIo. TheSP-ir terminal arbors (arrowheads) at the laminae IV/V border areconnected to the ir axons that penetrate the full thickness of the MDH(A, B), and become sparser in progressively rostral sections (B, C). Atthe periobex level, the Vi neuropil first appears at the ventral part ofthe BSTC and separates the MDH from the TrV (C). D: The mid-dorsoventral part of the TrV and laminae I–III at the mid-MDH level.The SP-ir axons in the external half of the tract and presumed
terminal arbors in laminae I and II (arrowheads) can be distinguishedfor the chain-like 3-38 diaminobenzidine (DAB) precipitates (orienta-tion: dorsolateral to the top and dorsomedial to the left). E–G:Following the ipsilateral rhizotomy, laminae I/II exhibit rostrocaudalgradation in the amount of residual SP-ir. The SP-ir terminal networkat the laminae IV/V border has disappeared, leaving granular SP-ir(arrowheads in E, F). Despite the rhizotomy, the paraV retains intenseSP-ir (arrows, G) at the periobex level.H: The contralateral counterpart ofthe area shown in D contains some granular SP-ir but chain-like arrange-ment is rare. Scale bars5 500 µm forA–C, E–G; 50 µm forD,H.
Fig. 9. SP-ir in the mid-Vi and the paraV contralateral (A, B) andipsilateral (C, D) to the 1-week rhizotomy. A: Many SP-ir axons areseen in the external half of the TrV, but the Vi is almost devoid of irterminals except at the dorsomedial edge (arrow).B: In addition to thedense SP-ir terminals in the paraV (arrowhead), a coarse network ofSP-ir terminals can be seen at the dorsomedial edge of Vi (arrow). C:The ipsilateral rhizotomy almost completely eliminates the SP-iraxons in the TrV except for the dorsal part (arrowheads). D:AlthoughSP-ir axons persist in or at the medial edge (large arrow) of the dorsalpart of the TrV, the ir terminals in the paraV (arrowhead) and thedorsomedial edge of the Vi (small arrow) have disappeared leaving finegranular SP-ir. The effect of rhizotomy on the solitary tract nucleus(STN) is not obvious. Scale bars 5 200 µm.
Fig. 10. SP-ir in the mid-Vo contralateral (A, B) and ipsilateral (C,D) to the 1-week rhizotomy. A:Moderate to intense SP-ir foci are seenin the dorsomedial nucleus (arrowhead), the lateral edge of the Vo(small arrow) and the STN (large arrow). B: The SP-ir profiles withinthe Vo include both terminal arbors and fine granules, whereas thosein the STN (arrow) are only granular. C,D: The ipsilateral rhizotomyalmost completely eliminates the terminal arbors, but the granularSP-ir in both the Vo and the STN (small and large arrows in C and D,respectively) is largely unaffected. Scale bars 5 200 µm.
436 T. SUGIMOTO ET AL.

Figures 9 and 10
CGRP- AND SP-IR IN THE RAT TRIGEMINAL NUCLEI 437

SP-ir in the STN. The STN contained dense granularSP-ir in all the three subnuclei. Presumed glossopharyn-geal and vagal root axons penetrating the TrV also exhib-ited SP-ir. The SP-ir axon bundles connecting the TrV andthe STN at the periobex level and around the junction ofthe Vo and Vi exhibited SP-ir. Marked reduction in stain-ing intensity due to rhizotomy was not discernible in anypart of the STN but the number of the ir axon bundlesconnecting the TrV and the nucleus was somewhat re-duced (e.g., Fig. 9D).
DISCUSSION
Immunohistochemical analyses have demonstratedCGRP-ir in the BSTC and surrounding structures in ratsand cats (Skofitsch and Jacobowitz, 1985a; Kruger et al.,1988a,b; Tashiro et al., 1991; Bennett-Clarke and Chiaia,1992; Stover et al., 1992; Henry et al., 1996). This studyusing the trigeminal rhizotomy provided an opportunity todistinguish CGRP-ir derived from trigeminal primaryafferent neurons from those from intrinsic central neuronsand from nontrigeminal primary neurons. Central axonsand terminals disconnected from their cell bodies areknown to undergo fragmentation (Ralston and Ralston,1979). This process appears to occur more rapidly in ratsthan in cats. A similar rhizotomy study in cats demon-strated that changes in CGRP-ir during a month followingthe rhizotomy are progressive, and that many CGRP-iraxons, whichmaintain relatively healthy appearance at 10days postoperative, ultimately disappear by 30 days (Henryet al., 1996). In this study, massive reduction in generalstaining intensity of the mid- to rostral MDH was almostcompleted during the first postoperative week, leavingsome apparently healthy axon terminals at the dorsome-dial edge and a few coarse granular CGRP-ir in lamina IIoof the more ventrolateral part. During the subsequentweek, the change in amount of such granules was barelydiscernible and the axons that survived the first weekremained apparently unchanged in amount, staining inten-sity and topographic distribution. These axon terminals atthe dorsomedial edge of MDH were derived from theglossopharyngeal and vagal axons, and were eliminated ina week following the tractotomy instead of rhizotomy (seebelow). Therefore, all the CGRP-ir axons and terminalsdisconnected from the cell body appear to complete frag-mentation in a week after the injury, leaving a minimalamount of debris. The exact reason for this species differ-ence is not known, but the shorter distance between thecell body and the terminal arbor may account for the morerapid fragmentation and elimination of the affected axonsin rats.Reduction of CGRP-ir following the rhizotomywas accom-
panied by a concomitant reduction in SP-ir. Many morpho-logically identified SP-ir terminal arbors in the Vi and Vo,and their parent axons in the TrV, disappeared during thefirst postoperative week. In addition, massive reduction ofthe SP-ir was observed in the paraV and laminae I and IIof the rostral MDH. Unlike for the CGRP-ir, however,complete elimination of SP-ir was not observed in any partof the examined brainstem, except the rostral part of theTrV. Multiple dorsal rhizotomies do not completely deprivethe superficial spinal dorsal horn of SP-ir, indicating thatintrinsic central neurons contribute to the SP-ir of thedorsal horn (Barber et al., 1979; Tessler et al., 1981). Suchnonprimary neurons, as well as nontrigeminal primaryneurons, might have contributed to the granular SP-ir
that persisted following the rhizotomy and tractotomy.Nevertheless, reduction of SP-ir in the BSTC indicatesthat the trigeminal primary neurons projecting to the Vo,Vi, paraV and MDH included SP-ir nociceptors.
Convergent primary afferent inputs to theBSTC
The presently demonstrated distribution of CGRP-irwithin the dorsal horn was basically similar to thatpreviously reported for rats and cats (Skofitsch and Jaco-bowitz, 1985a; Kruger et al., 1988b; Tashiro et al., 1991;Bennett-Clarke and Chiaia, 1992; Stover et al., 1992;Henry et al., 1996). Terminal labeling showed a clearlamination in laminae I to III; i.e., it was densest inlaminae I and IIo, moderate in lamina IIi, and almostnonexistent in lamina III. In addition, CGRP-ir terminalsformed patchy clumps in lamina V. Because the trigeminalrhizotomy caused a marked reduction of CGRP-ir andSP-ir, the MDH laminae I, II and V are considered to bemajor terminal fields of the trigeminal primary nocicep-tors. In the rat spinal dorsal horn, multiple dorsal rhizoto-mies induce nearly complete depletion of CGRP-ir inlaminae I, II and V (Chung et al., 1988), suggestingprimary neurons are the only source of CGRP-ir in theselaminae. If so, the CGRP-ir terminals that survived theipsilateral trigeminal rhizotomy are considered to havebeen derived from nontrigeminal primary neurons and/orcontralateral trigeminal primary neurons.The response of the dorsal horn to the trigeminal
rhizotomy was not uniform along either rostrocaudal ordorsoventral axes. The rostrocaudal gradation in amountof the residual CGRP-ir and SP-ir may suggest that one ofthe nontrigeminal sources of nociceptive primary projec-tion to the MDH is the cervical dorsal root fibers. Adensitometric analysis of CGRP-ir after the cervical Lissau-er’s tractotomy in the cat revealed that CGRP-ir dorsalroot axons ascend the TrV and project to the dorsomedialpart of the MDH (Stover et al., 1992). Another source ofCGRP-ir terminals in the dorsomedial part of the MDH isconsidered to be the ipsilateral glossopharyngeal andvagal nerve roots. The distribution within the TrV androstral MDH of degeneration argentophilia following com-bined glossopharyngeal and vagal rhizotomies (Torvik,1956) coincides with the distribution of the residualCGRP-ir and SP-ir after the trigeminal rhizotomy in thisstudy. Furthermore, the TrV tractotomy that was expectedto sever the glossopharyngeal and vagal axons descendingin the TrV, almost completely eliminated the morphologi-cally identified CGRP-ir terminals in the dorsomedial partof the MDH in this study.In addition to the above nontrigeminal primary axons
that descend or ascend in the ipsilateral TrV and project tothe dorsomedial part of the MDH, primary neuronalprojections from the contralateral side also appear tocontribute to the CGRP-ir in the MDH because the com-bined TrV tractotomy and upper cervical hemicordotomydid not completely eliminate CGRP-ir axons and terminalsin the caudal MDH. A tracer injection into the ganglion orthe peripheral nerve demonstrates contralateral projec-tion of trigeminal primary neurons of rats. Their axonscross the midline at the level of pyramidal decussation andreach the dorsomedial edge of the contralateral MDH(Jacquin et al., 1982, 1983; Takemura et al., 1987; Pfallerand Arvidsson, 1988; Marfurt and Rajchert, 1991). Thetrajectory of CGRP-ir axons in this study resembled that ofthe tracer-labeled axons in the previous studies, and we
438 T. SUGIMOTO ET AL.

assume that some of the contralaterally projecting trigemi-nal primary neurons contained CGRP. We cannot excludea possibility that glossopharyngeal, vagal or dorsal rootaxons also projected to the contralateral MDH.Intraoral structures and the nerves innervating them
are prone to various types of injuries. Damages due todental and periodontal diseases, oral surgery and traumaare examples of such injuries. In addition to the pain statedirectly caused by these injuries in their acute phases,chronic painful sensory disorders may result. In theseinstances, diffuse aberrant pain often spreads beyond thedamaged tissue or the receptive fields of injured nervesinto adjacent or remote facial and cervical structures, andeven into the arm (Kerr, 1979). Most nociceptive neuronsin the MDH have a relatively narrow mechanoreceptivefield in the oral mucosa or the facial skin. However, manyof them respond to electrical stimulation of afferentssupplying deep tissues out side the trigeminal territory(Sessle et al., 1986). Therefore, the MDH neurons thatnormally respond only to stimulation of the trigeminalreceptive fields have hidden extra trigeminal receptivefields in the neck and trunk. Experimental trigeminaldeafferentation is followed by increase in the rate ofspontaneous discharge, indicating a hyperactivity of thedeafferented MDH neurons (Anderson et al., 1971; Black,1974). In other instances, the partially deafferented MDHneurons begin to show a prolonged afterdischarge follow-ing stimulation of unaffected trigeminal receptive fields(Black, 1974). In such pathological conditions, the remain-ing convergent synaptic input, which is normally silent,may activate the deafferented dorsal horn neurons. Such‘‘unmasking’’ of previously silent input may result inhyperalgesia and referral and spread of pain followingperipheral injuries. Indeed, chronic partial deafferenta-tion by a single dorsal rhizotomy induces c-fos hyperinduc-ibility of spinal dorsal horn neurons in response to conver-gent primary nociceptive input through neighboring intactdorsal root fibers (Sugimoto et al., 1994). Extensive over-lap of CGRP- and SP-ir trigeminal, cervical, glossopharyn-geal and vagal primary projections in the MDH and upperCDH may provide a morphological substrate for switchingor expansion of nociceptive receptive fields of dorsal hornneurons under pathological conditions. Detailed discus-sion on this topic appeared elsewhere (Sessle, 1987; Stoveret al., 1992).Other regions of the BSTC that received convergent
CGRP- and SP-ir projection from other than the ipsilateraltrigeminal ganglion were the paraV and the lateral edge ofVo. A degeneration study demonstrated the glossopharyn-geal and/or vagal primary axons to project to the paraV.Although direct evidence is not available so far, theproximity of the area to the root entry zone may suggestthat the CGRP-ir and SP-ir in the lateral edge of Vo arepartially derived from the glossopharyngeal and/or vagalprimary neurons.
CGRP-ir primary afferent projection to therostral subdivisions of BSTC
The CGRP-ir axons have been known to project to theentire rostrocaudal extent of the BSTC (Kruger et al.,1988a,b; Tashiro et al., 1991; Bennett-Clarke and Chiaia,1992; Stover et al., 1992; Henry et al., 1996). Trigeminalrhizotomy studies in the cat demonstrated that CGRP-iraxon terminals in the Vi, Vo and PrV are almost exclu-sively of trigeminal origin (Tashiro et al., 1991; Henry etal., 1996). The present study is the first to demonstrate
that the CGRP-ir axon terminals in the Vi, Vo and PrV ofrats are also derived from the ipsilateral trigeminal pri-mary neurons with the exception of a part of the CGRP-irterminals at the lateral edge of Vo. Except for the PrVwhere the change was too small to be detected, therhizotomy caused concomitant reduction of SP-ir in theserostral subdivisions. A similar effect of the rhizotomy onthe rostral SP-ir was reported for the cat (Tashiro et al.,1991).The MDH (former trigeminal subnucleus caudalis) has
been long recognized as the trigeminal homologue of thespinal dorsal horn (Gobel et al., 1981; Dubner and Ben-nett, 1983 for review). The trigeminal tractotomy at thelevel of obex, which interrupts all the ipsilateral trigemi-nal primary projections to the MDH, brings about almostcomplete analgesia and thermal anesthesia of the skin inthe periphery of the face, leaving the sense of touch almostunaffected (Sjoqvist, 1938). This observation providedstrong evidence for the similarity between the MDH andspinal dorsal horn, and also suggested that the MDH wasthe sine qua non for the trigeminal pain perception.Careful reevaluation of the effect of trigeminal rhizotomyin human patients and laboratory species, however, demon-strated that the centrifacial and intraoral structures wererather insensitive to the analgesic effect of TrV tractotomy(Vyklicky et al., 1977; Ikegami and Kawamura, 1979;Young et al., 1981; Young, 1982; Young and Perryman,1984, 1986; Dallel et al., 1988). This suggests that aconsiderable proportion of the trigeminal primary nocicep-tive projection, especially from the intraoral structures, isdirected to the relay nuclei located in the rostral subdivi-sions of the BSTC. This idea is supported by the fact that alarge proportion of central projection of presumed primarynociceptors innvervating the tooth pulp is directed torostral subdivisions of the BSTC (Arvidsson and Gobel,1981; Westrum et al., 1981; Marfurt and Turner, 1984;Shigenaga et al., 1986b, 1989; Takemura et al., 1991;Sugimoto and Takemura, 1993).Combined retrograde labeling and immunohistochemis-
try studies in the rat revealed two major subpopulations oftrigeminal primary neurons innervating the tooth pulp.About 50% of the tooth pulp-innervating primary neuronsexpressed CGRP-ir, while 30% contained parvalbumin-ir(Fried et al., 1989; Ichikawa et al., 1995). Parvalbumin hasbeen recognized as a marker for large myelinated primaryneurons and coexists with carbonic anhydrase (Ichikawaet al., 1994). This enzyme is also expressed by 30% of thetooth pulp-innervating primary neurons of the rat (Sugi-moto et al., 1989, 1990). Although the tooth pulp-innervating primary neurons coexpress CGRP and car-bonic anhydrase, incidence of such coexpression is fairlylow (Ichikawa et al., 1995). Before this study, we supposedthat the parvalbumin-ir rather than CGRP-ir trigeminalprimary nociceptors transmitted the nociceptive informa-tion from the tooth pulp to the rostral BSTC, but it is morelikely that the tooth pulp-innervating neurons withCGRP-ir also contribute to the nociceptive transmission tothe rostral BSTC, especially the dorsomedial nucleus atthe Vo level. Because SP-ir neurons constituted only asmall fraction of the tooth pulp-innervating primary neu-rons (Fried et al., 1989), the SP-ir in the rostral BSTCmight have been also derived from trigeminal primarynociceptors other than those innervating the tooth pulp.It is noteworthy that the two densest foci of the CGRP-
and SP-ir trigeminal projection within the rostral subdivi-sions of BSTC were both located in the Vo. The medial one
CGRP- AND SP-IR IN THE RAT TRIGEMINAL NUCLEI 439

occupied the dorsomedial nucleus, while the lateral oneresided within the concavity of the TrV. The latter area hasnot been reported to receive CGRP-ir projection in the cat.The dorsomedial nucleus, first described by Åstrøm (1953),can be distinguished from the rest of the Vo by itscytoarchitecture and is highly developed in rodents. Thenucleus is known to receive trigeminal primary projectionfrom exclusively intraoral structures (Arvidsson and Go-bel, 1981; Marfurt, 1981; Jacquin et al., 1983; Marfurt andTurner, 1984; Shigenaga et al., 1986a,b, 1989; Takemuraet al., 1987, 1991). Prominent CGRP- and SP-ir projectionof exclusively trigeminal primary neurons as shown in thisstudy has added another characteristic that distinguishesthe rodent dorsomedial nucleus from the rest of the BSTC.Clinical experiences, as well as the knowledge accumu-lated through experimental studies, may suggest implica-tion of this nucleus in the mechanisms of pain perception(see below). On the other hand, the narrow area adjacentto the TrV is the terminal field of trigeminal primaryneurons innervating the hairy skin of the periphery of theface (Jacquin et al., 1983; Shigenaga et al., 1986a; Take-mura et al., 1987, 1991). This area of the Vo receivedconvergent CGRP- and SP-ir projection of trigeminal andnontrigeminal origins. Such convergence into this narrowarea may suggest that the CGRP- and SP-ir primaryprojection is involved in modulation of innocuous sensoryinformation rather than pain perception.In the cat, a much larger proportion of CGRP-ir trigemi-
nal primary projection is directed to the PrV than to theVo, while the PrV to Vo ratio was inverted in the rat. In theformer species, the PrV but not the Vo contains manyprojection neurons with ascending axons projecting to thecontralateral nucleus ventralis posteromedialis (VPM) ofthe thalamus (Shigenaga et al., 1983, 1986b). Therefore, itmay be that the principal relay nucleus involved in theintraoral nociception is the PrV in the cat. Unlike in thecat, the rat’s Vo contains a substantial number of neuronsthat project to the contralateral VPM, although their cellbodies are located mainly in the ventrolateral part of thesubnucleus (Luo and Dessem, 1995).
CGRP-ir trigeminal primary projectionto the STN
Using a degeneration technique, Torvik (1956) demon-strated primary afferent projections to the STN from theintermediate, trigeminal, glossopharyngeal and vagal nerveroots. The rostral pole of the nucleus received input mainlyfrom the intermediate nerve, while the rest of the nucleusreceived input from the glossopharyngeal and vagal nerves.In addition, the terminal field of trigeminal primaryneurons overlapped with the rostral part of glossopharyn-geal and vagal terminal fields. Such trigeminal projectionto the STN has been repeatedly confirmed in both cats andrats by contemporary tracing methods using such ashorseradish peroxidase (HRP; Jacquin et al., 1982, 1983;Pfaller and Arvidsson, 1988; Marfurt and Rajchert, 1991).Trigeminal primary projection to the STN is densest at thecaudal Vo level, and is mostly derived from the primaryneurons innervating the intraoral structures (Nomura etal., 1984; Takemura et al., 1987, 1991). Our data document-ing reduction of CGRP-ir in the STN and SP-ir axonsprojecting to the STN following the trigeminal rhizotomyindicate that such trigeminal primary neurons includenociceptors.It is not clear whether the STN is involved in pain
perception. The fact that the STN and the BSTC have
extensive reciprocal fiber connections suggested that thetrigeminal primary input is involved in somatovisceralreflex (Menetrey and Basbaum, 1987). Because the oralcavity constitutes the uppermost part of the alimentaryand respiratory tracts, direct primary input from the oralcavity through the trigeminal nerve to the STN may playan important role in the reflex control of such visceralfunctions.
CONCLUSIONS
Immunohistochemical staining for CGRP and SP wasperformed on the lower brainstem of normal rats and afterthe trigeminal rhizotomy or TrV tractotomy at the level ofVi. The results were mutually compared to elucidate thecentral projection of the trigeminal and other sensoryprimary nociceptors.The CGRP-ir trigeminal primary neurons projected to
the entire rostrocaudal extent of the ipsilateral BSTC(MDH, Vi, Vo and PrV), paraV and STN. A subpopulationof the trigeminal primary neurons projecting to the ipsilat-eral MDH, Vi, Vo, paraV and STN contained SP-ir.In theMDH, CGRP-ir wasmost intense in laminae I and
II, and weaker in lamina V. The CGRP-ir terminals in theMDH were mainly, but not exclusively, derived from theipsilateral trigeminal primary neurons. Extensive overlapof primary projection from the ipsilateral trigeminal, andglossopharyngeal and/or vagal primary neurons was evi-denced at the dorsomedial edge of laminae I and II of therostral MDH. In the caudal MDH, the convergent CGRP-irprimary projection appeared to have been derived from thecontralateral trigeminal primary neurons in addition tothe ipsilateral trigeminal and cervical primary neurons.Among the rostral subdivisions of the BSTC, the Vo
received the densest CGRP-ir projection from the ipsilat-eral trigeminal primary neurons. The terminals weredistributed in two separate areas of the dorsal part of theVo, i.e., the dorsomedial nucleus and the lateral edge of theVo. The dorsomedial nucleus received CGRP-ir projectionexclusively from the ipsilateral trigeminal primary neu-rons, while the lateral part received a convergent CGRP-irprojection from the ipsilateral trigeminal and presumedglossopharyngeal and/or vagal primary neurons.The CGRP-ir terminals in the Vi and PrV were fewer
than in the Vo, but appeared to have been derived exclu-sively from the ipsilateral trigeminal primary neurons. Inthe Vi, CGRP-ir was restricted to the medial edge of thesubnucleus and was more concentrated in the dorsal part.In the PrV, CGRP-ir terminals were located in the periph-ery of the dorsal half of the nucleus.The paraV received a convergent CGRP-ir from the
ipsilateral trigeminal and presumed glossopharyngeal/vagal primary neurons. The proportion of the trigeminalcontribution was greater for the rostral than for the caudalparaV. The STN also contained intense CGRP-ir through-out the nucleus, and the ipsilateral trigeminal projectioncontributed to such CGRP-ir in the lateral subnucleus ofthe STN at the levels of rostral Vi and caudal Vo.
LITERATURE CITED
Amara, S.G., V. Jones, M.G. Rosenfeld, E.S. Ong, and R.M. Evans (1982)Alternative RNA processing in calcitonin gene expression generatesmRNAs encoding different peptide products. Nature 298:240–144.
Amara, S.G., J.L. Arriza, S.E. Leff, L.W. Swanson, R.M. Evans, and M.G.Rosenfeld (1985) Expression in brain of a messenger RNA encoding a
440 T. SUGIMOTO ET AL.

novel neuropeptide homologous to calcitonin gene-related peptide.Science 229:1094–1097.
Anderson, L.S., R.G. Black, J. Abraham, and A.A. Ward (1971) Neuronalhyperactivity in experimental trigeminal deafferentation. J. Neuro-surg. 35:444–452.
Arvidsson, J., and S. Gobel (1981) An HRP study of the central projectionsof primary trigeminal neurons that innervate tooth pulp in the cat.Brain Res. 210:1–16.
Åstrøm, K.E. (1953) On the central course of afferent fibres in thetrigeminal, facial, glossopharyngeal and vagal nerves and their nucleiin the mouse. Acta Physiol. Scand. 29(Suppl. 106):209–320.
Barber, R.P., J.E. Vaughn, J.R. Slemmon, P.M. Salvaterra, E. Roberts, andS.E. Leeman (1979) The origin, distribution and synaptic relationshipsof substance P axons in rat spinal cord. J. Comp. Neurol. 184:331–352.
Bennett-Clarke, C.A., and N.L. Chiaia (1992) Normal development andeffects of neonatal infraorbital nerve damage upon the innervation ofthe trigeminal brainstem complex by primary afferent fibers containingcalcitonin gene-related peptide. J. Comp. Neurol. 324:282–294.
Black, R.G. (1974) A laboratory model for trigeminal neuralgia. Adv.Neurol. 4:651–659.
Carlton, S.M., and E.S. Hayes (1989) Dynorphin A (1-9) immunoreactivecell bodies, dendrites and terminals are postsynaptic to calcitoningene-related peptide primary afferent terminals in the monkey dorsalhorn. Brain Res. 504:124–128.
Carlton, S.M., D.L. McNeill, K. Chung, and R.E. Coggeshall (1987) A lightand electron microscopic level analysis of calcitonin gene-related pep-tide (CGRP) in the spinal cord of the primate: An immunohistochemicalstudy. Neurosci. Lett. 82: 145–150.
Chung, K., W.T. Lee, and S.M. Carlton (1988) The effects of dorsalrhizotomy and spinal isolation on calcitonin gene-related peptide-labeled terminals in the rat lumbar dorsal horn. Neurosci. Lett.90:27–32.
Colin, S., and L. Kruger (1986) Peptidergic nociceptive axon visualizationin whole-mount preparations of cornea and tympanic membrane in rat.Brain Res. 398:199–203.
Dallel, R., P. Raboisson, P.Auroy, andA. Woda (1988) The rostral part of thetrigeminal sensory complex is involved in orofacial nociception. BrainRes. 448:7–19.
Dubner, R., and G.J. Bennett (1983) Spinal and trigeminal mechanisms ofnociception. Ann. Rev. Neurosci. 6:381–418.
Fried, K., J. Arvidsson, B. Robertson, E. Brodin, and E. Theodorsson (1989)Combined retrograde tracing and enzyme/immunohistochemistry oftrigeminal ganglion cell bodies innervating tooth pulps in the rat.Neuroscience 33:101–109.
Gamse, R., and A. Saria (1986) Nociceptive behavior after intrathecalinjections of substance P, neurokinin A and calcitonin gene-relatedpeptide in mice. Neurosci. Lett. 70:143–147.
Gibbins, I.L., J.B. Furness, M. Costa, I. MacIntyre, C.J. Hillyard, and S.Girgis (1985) Co-localization of calcitonin gene-related peptide-likeimmunoreactivity with substance P in cutaneous, vascular and visceralsensory neurons of guinea pigs. Neurosci. Lett. 57:125–130.
Gibson, S.J., J.M. Polak, S.R. Bloom, I.M. Sabate, P.M. Muldery, M.A.Ghatei, G.P. McGregor, J.F.B. Morrison, J.S. Kelly, R.M. Evans, andM.G. Rosenfeld (1984) Calcitonin gene-related peptide immunoreactiv-ity in the spinal cord of man and of eight other species. J. Neurosci.4:3101–3111.
Gobel, S., S. Hockfield, and M.A. Ruda (1981) Anatomical similaritiesbetween medullary and spinal dorsal horns. In Y. Kawamura and R.Dubner (eds): Oral-Facial Sensory andMotor Functions. Tokyo: Quintes-sence, pp. 211–223.
Goodman, E.C., and L.L. Iversen (1986) Calcitonin gene-related peptide:Novel neuropeptide. Life Sci. 38:2169–2178.
Hammond, D.L., and M.A. Ruda (1991) Developmental alterations innociceptive threshold, immunoreactive calcitonin gene-related peptideand substance P, and fluoride-resistant acid phosphatase in neonatallycapsaicin-treated rats. J. Comp. Neurol. 312:436–450.
Harmon, P.A., K. Chung, R.P. Briner, K.N. Westlund, and S.M. Carlton(1988) Calcitonin gene-related peptide (CGRP) in the human spinalcord: A light and electron microscopic analysis. J. Comp. Neurol.269:371–380.
Henry, M.A., L.R. Johnson, N. Nousek-Goebl, and L.E. Westrum (1996)Light microscopic localization of calcitonin gene-related peptide in thenormal feline trigeminal system and following retrogasserian rhi-zotomy. J. Comp. Neurol. 365:526–540.
Hokfelt, T. (1991) Neuropeptides in perspective: The last ten years. Neuron7:867–879.
Ichikawa, H., S. Matsuo, S. Wakisaka, and M.Akai (1990) Fine structure ofcalcitonin gene-related peptide-immunoreactive nerve fibers in the rattemporomandibular joint. Arch. Oral Biol. 35:727–730.
Ichikawa, H., T. Deguchi, T. Nakago, D.M. Jacobowitz, and T. Sugimoto(1994) Parvalbumin, calretinin and carbonic anhydrase in the trigemi-nal and spinal primary neurons of the rat. Brain Res. 655:241–245.
Ichikawa, H., T. Deguchi, T. Nakago, D.M. Jacobowitz, and T. Sugimoto(1995) Parvalbumin- and calretinin-immunoreactive trigeminal neu-rons innervating the rat molar tooth pulps. Brain Res. 679:205–211.
Ikegami, S., and H. Kawamura (1979) Avoidance conditioning to tooth pulpstimulation in the cat after bulbar transection. Physiol. Behav. 23:593–596.
Jacquin, M.F., K. Semba, R.W. Rhoades, and M.D. Egger (1982) Trigeminalprimary afferents project bilaterally to dorsal horn and ipsilaterally tocerebellum, reticular formation, and cuneate, solitary, supratrigeminaland vagal nuclei. Brain Res. 246:285–291.
Jacquin, M.F., K. Semba, M.D. Egger, and R.W. Rhoades (1983) Organiza-tion of HRP-labeled trigeminal mandibular primary afferent neurons inthe rat. J. Comp. Neurol. 215:397–420.
Ju, G., T. Hokfelt, E. Brodin, J. Fahrenkrug, J.A. Fischer, R. Frey, R.P. Elde,and J.C. Brown (1987) Primary sensory neurons of the rat showingcalcitonin gene-related peptide immunoreactivity and their relation tosubstance P-, somatostatin-, galanin-, vasoactive intestinal peptide-and cholecystokinin-immunoreactive ganglion cells. Cell Tissue Res.247:417–431.
Kerr, F.W.L. (1979) Craniofacial neuralgias. In J.J. Bonica, J.C. Liebeskindand D.G. Albe-Fessard (eds): Advances in Pain Research and Therapy.NewYork: Raven Press, pp. 283–295.
Kruger, L., P.W. Mantyh, C. Sternini, N.C. Brecha, and C.R. Mantyh(1988a) Calcitonin gene-related peptide (CGRP) in the rat centralnervous system: Patterns of immunoreactivity and receptor bindingsites. Brain Res. 463:223–244.
Kruger, L., C. Sternini, N.C. Brecha, and P.W. Mantyh (1988b) Distributionof calcitonin gene-related peptide immunoreactivity in relation to therat central somatosensory projection. J. Comp. Neurol. 273:149–162.
Kuwayama, Y., G. Terenghi, J.M. Polak, J.Q. Trojanowski, and R.A. Stone(1987) A quantitative correlation of substance P-, calcitonin gene-related peptide- and cholecystokinin-like immunoreactivity with retro-gradely labeled trigeminal ganglion cells innervating the eye. BrainRes. 405:220–266.
Landis, S.C., and J.R. Fredieu (1986) Coexistence of calcitonin gene-relatedpeptide and vasoactive intestinal peptide in cholinergic sympatheticinnervation of rat sweat glands. Brain Res. 377:177–181.
LeGreves, P., F. Nyberg, L. Terenius, and T. Hokfelt (1985) Calcitoningene-related peptide is a potent inhibitor of substance P degradation.Eur. J. Pharmacol. 115:309–311.
Lee, Y., Y. Kawai, S. Shiosaka, K. Takami, H. Kiyama, C.J. Hillyard, S.Girgis, I. MacIntyre, P.C. Emson, and M. Tohyama (1985a) Coexistenceof calcitonin gene-related peptide and substance P- like peptides insingle cells on the trigeminal ganglion of the rat: Immunohistochemicalanalysis. Brain Res. 330:194–196.
Lee, Y., K. Takami, Y. Kawai, S. Girgis, C.J. Hillyard, I. MacIntyre, P.C.Emson, andM. Tohyama (1985b) Distribution of calcitonin gene-relatedpeptide in the rat peripheral nervous system with reference to itscoexistence with substance P. Neuroscience 15:1227–1237.
Lundberg, J.M., A. Granco-Cereceda, X. Hua, T. Hokfelt, and J.A. Fischer(1985) Co-existence of substance P and calcitonin gene-related peptidein sensory nerves in relation to cardiovascular and bronchoconstrictoreffects of capsaicin. Eur. J. Pharmacol. 108:315–319.
Luo, P., and D. Dessem (1995) Inputs from identified jaw-muscle spindleafferents to trigeminothalamic neurons in the rat: A double-labelingstudy using retrograde HRP and intracellular biotinamide. J. Comp.Neurol. 353:50–66.
Marfurt, C.F. (1981) The central projections of trigeminal primary afferentneurons in the cat as determined by the transganglionic transport ofhorseradish peroxidase. J. Comp. Neurol. 203:785–798.
Marfurt, C.F., and D.M. Rajchert (1991) Trigeminal primary afferentprojections to ‘‘non-trigeminal’’ areas of the rat central nervous system.J. Comp. Neurol. 303:489–511.
Marfurt, C.F., and D.F. Turner (1984) The central projections of tooth pulpafferent neurons in the rat as determined by the transganglionictransport of horesradish peroxidase. J. Comp. Neurol. 223:535–547.
Mason, R.T., R.A. Peterfreund, P.E. Sawchenko, A.Z. Corrigan, J.E. Rivier,and W.W. Vale (1984) Release of the predicted calcitonin gene-relatedpeptide from cultured rat trigeminal ganglion cells. Nature 308:653–655.
CGRP- AND SP-IR IN THE RAT TRIGEMINAL NUCLEI 441

McCulloch, J., R. Uddman, T.A. Kingman, and L. Edvinsson (1986)Calcitonin gene-related peptide: Functional role in cerebrovascularregulation. Proc. Natl. Acad. Sci. USA 83:5731–5735.
Menetrey, D., andA.I. Basbaum (1987) Spinal and trigeminal projections tothe nucleus of the solitary tract: A possible substrate for somatovisceraland viscerovisceral reflex activation. J. Comp. Neurol. 255:439–450.
Nomura, S., Y. Yasui, M. Takada, and N. Mizuno (1984) Trigeminal primaryafferent neurons projecting directly to the solitary tract nucleus in thecat: A transganglionic and retrograde horseradish peroxidase study.Neurosci. Lett. 50:257–262.
Peterfreund, R.A., and W.W. Vale (1986) Local anesthetics inhibit veratri-dine-induced secretion of calcitonin gene-related peptide (CGRP) fromcultured rat trigeminal ganglion cells. Brain Res. 380:159–161.
Pfaller, K., and J. Arvidsson (1988) Central distribution of trigeminal andupper cervical afferents in the rat studied by anterograde transport ofhorseradish peroxidase conjugated to wheat germ agglutinin. J. Comp.Neurol. 268:91–108.
Quartu, M., G. Diaz, A. Floris, M.L. Lai, J.V. Priestley, and M. Del Fiacco(1992) Calcitonin gene-related peptide in the human trigeminal sensorysystem at developmental and adult life stages: Immunohistochemistry,neuronalmorphology and coexistence with substance P.J. Chem. Neuro-anat. 5:143–157.
Ralston, H.J. III, and D.D. Ralston (1979) The distribution of dorsal rootaxons in laminae I, II and III of the macaque spinal cord: A quantitativeelectron microscope study. J. Comp. Neurol. 184:643–684.
Rodrigo, J., J.M. Polak, G. Terenghi, C. Cervantes, M.A. Ghatei, P.K.Mulderry, and S.R. Bloom (1985) Calcitonin gene-related peptide(CGRP)-immunoreactive sensory and motor nerves of the mammalianpalate. Histochemistry 82:67–74.
Rosenfeld, M.G., J.-J. Mermod, S.G. Amara, L.W. Swanson, R.E. Saw-chenko, J. Rivier, W.W. Vale, and R.M. Evans (1983) Production of anovel neuropeptide encoded by the calcitonin gene via tissue-specificRNAprocessing. Nature 304:129–135.
Rosenfeld, M.G., S.G. Amara, and R.M. Evans (1984) Alternative RNAprocessing: Determining neuronal phenotype. Science 225:1315–1320.
Sessle, B.J. (1987) The neurobiology of facial and dental pain: Presentknowledge, future directions. J. Dent. Res. 66: 962–981.
Sessle, B.J., J.W. Hu, N. Amano, and G. Zhong (1986) Convergence ofcutaneous, tooth pulp, visceral, neck and muscle afferents onto nocicep-tive and non-nociceptive neurones in trigeminal subnucleus caudalis(medullary dorsal horn) and its implication for referred pain. Pain 27:219–235.
Shigenaga, Y., Z. Nakatani, T. Nishimori, S. Suemune, R. Kuroda, and S.Matano (1983) The cells of origin of cat trigeminothalamic projections:Especially in the caudal medulla. Brain Res. 277:201–222.
Shigenaga, Y., T. Okamoto, T. Nishimori, S. Suemune, I.D. Nasution, I.C.Chen, K. Tsuru, A. Yoshida, K. Tabuchi, M. Hosoi, and H. Tsuru (1986a)Oral and facial representation in the trigeminal principle and rostralspinal nuclei of the cat. J. Comp. Neurol. 244:1–18.
Shigenaga, Y., S. Suemune, M. Nishimura, T. Nishimori, H. Sato, H.Ishidori, A. Yoshida, K. Tsuru, Y. Tsuiki, Y. Dateoka, I.D. Nasution, andM. Hosoi (1986b) Topographic representation of lower and upper teethwithin the trigeminal sensory nuclei of adult cat as demonstrated by thetransganglionic transport of horseradish peroxidase. J. Comp. Neurol.251:299–316.
Shigenaga, Y., M. Nishimura, S. Suemune, T. Nishimori, K. Doe, and H.Tsuru (1989) Somatotopic organization of tooth pulp primary afferentneurons in the cat. Brain Res. 477:66–89.
Silverman, J.D., and L. Kruger (1987) An interpretation of dental innerva-tion based upon the pattern of calcitonin gene-related peptide (CGRP)-immunoreactive thin sensory axons. Somatosens. Res. 5:157–175.
Silverman, J.D., and L. Kruger (1989) Calcitonin-gene-related-peptide-immunoreactive innervation of the rat head with special emphasis onspecialized sensory structures. J. Comp. Neurol. 280:303–330.
Sjoqvist, O. (1938) Studies on pain conduction in the trigeminal nerve. ActaPsychiatr. Scand., Suppl. 17:1–139.
Skofitsch, G., and D.M. Jacobowitz (1985a) Calcitonin gene-related peptide:Detailed immunohistochemical distribution in the central nervoussystem. Peptides 6:721–745.
Skofitsch, G., and D.M. Jacobowitz (1985b) Calcitonin gene-related peptidecoexists with substance P in capsaicin sensitive neurons and sensoryganglia of the rat. Peptides 6:747–754.
Stover, J.D., C.A. Schwab, andM.A.Matthews (1992) Selective deafferenta-tion of convergent inputs to trigeminal subnucleus caudalis: Effects oncalcitonin gene-related peptide distribution. Somatosens. Mot. Res.9:107–130.
Sugimoto, T., and M. Takemura (1993) Tooth pulp primary neurons: cellsize analysis, central projection, and carbonic anhydrase activity. BrainRes. Bull. 30:221–226.
Sugimoto, T., M. Takemura, H. Ichikawa, and M. Akai (1989) Carbonicanhydrase activity in the trigeminal primary afferent neuronal cellbodies with peripheral axons innervating the mandibular molar toothpulps of the rat. Brain Res. 505:354–357.
Sugimoto, T., M. Takemura, andN.Mukai (1990) Histochemical demonstra-tion of carbonic anhydrase activity within the mandibular molar andincisor tooth pulps of the rat. Brain Res. 529:245–254.
Sugimoto, T., A. Yoshida, K. Nishijima, and H. Ichikawa (1994) c-Fosinduction in the rat spinal dorsal horn partially deafferented by dorsalrhizotomy. Neurosci. Lett. 178:239–242.
Takemura, M., T. Sugimoto, and A. Sakai (1987) Topographic organizationof central terminal region of different sensory branches of the ratmandibular nerve. Exp. Neurol. 96:540–557.
Takemura, M., T. Sugimoto, and Y. Shigenaga (1991) Difference in centralprojection of primary afferents innervating facial and intraoral struc-tures in the rat. Exp. Neurol. 111:324–331.
Tashiro, T., O. Takahashi, T. Satoda, R. Matsushima, M. Uemura-Sumi,and N. Mizuno (1991) Distribution of axons showing calcitonin gene-related peptide- and/or substance P-like immunoreactivity in thesensory trigeminal nuclei of the cat. Neurosci. Res. 11:119–133.
Tessler, A., E. Glazer, R. Artymyshyn, M. Murray, and M.E. Goldberger(1980) Recovery of substance P in the cat spinal cord after unilaterallumbosacral deafferentation. Brain Res. 191:459–470.
Tessler, A., B.T. Himes, R. Artymyshyn, M. Murray, and M.E. Goldberger(1981) Spinal neurons mediate return of substance P following deaffer-entation of cat spinal cord. Brain Res. 230:263–281.
Tessler, A., B.T. Himes, K. Soper, M. Murray, and M.E. Goldberger (1984)Recovery of substance P but not somatostatin in the cat spinal cord afterunilateral lumbosacral dorsal rhizotomy: A quantitative study. BrainRes. 305:95–102.
Torvik, A. (1956) Afferent connections to the sensory trigeminal nuclei, thenucleus of the solitary tract and adjacent structures. J. Comp. Neurol.106:51–141.
Traub, R.J., A. Solodkin, and M.A. Ruda (1989) Calcitonin gene-relatedpeptide immunoreactivity in the cat lumbosacral spinal cord and theeffects of multiple dorsal rhizotomies. J. Comp. Neurol. 287:225–237.
Traub, R.J., B. Allen, A. Humphrey, and M.A. Ruda (1990) Analysis ofcalcitonin gene-related peptide-like immunoreactivity in the cat dorsalspinal cord and dorsal root ganglia provides evidence for a multiplesegmental projection of nociceptive C-fiber primary afferents. J. Comp.Neurol. 302:562–574.
Vyklicky, L., O. Keller, P. Jastreboff, L. Vyklicky, Jr., and S.M. Butkhuzi(1977) Spinal trigeminal tractotomy and nociceptive reactions evokedby tooth pulp stimulation in the cat. J. Physiol., Paris, 73:379–386.
Westrum, L.E., R.C. Canfield, and T.A. O’Connor (1981) Each canine toothprojects to all brain stem trigeminal nuclei in cat. Exp. Neurol.74:787–799.
Wiesenfeld-Hallin, Z., T. Hokfelt, J.M. Lundberg, W.G. Forssman, M.Reinecke, F.A. Tschopp, and J.A. Fischer (1984) Immunoreactive calci-tonin gene-related peptide and substance P coexist in sensory neuronsin spinal cord and interact in spinal behavioral responses of the rat.Neurosci. Lett. 52:199–204.
Xu, X.-J., Z. Wiesenfeld-Hallin, M.J. Villar, J. Fahrenkrug, and T. Hokfelt(1990) On the role of galanin, substance P and other neuropeptides inprimary sensory neurons of the rat: Studies on spinal reflex excitabilityand peripheral axotomy. Eur. J. Neurosci. 2:733–743.
Yamamoto, A.I., and E. Senba (1990) Cell types and axonal sizes ofcalcitonin gene-related peptide-containing primary sensory neurons ofthe rat. Brain Res. Bull. 24:759–764.
Yoshida, A., H. Ichikawa, and T. Sugimoto (1995) CGRP in peripherallyaxotomized mesencephalic trigeminal neurons of the rat. NeuroReport6:837–840.
Young, R.F. (1982) Effect of trigeminal tractotomy on dental sensation inhumans. J. Neurosurg. 56:812–818.
Young, R.F., and K.M. Perryman (1984) Pathways for orofacial painsensation in the trigeminal brain-stem nuclear complex of the Macaquemonkey. J. Neurosurg. 61:563–568.
Young, R.F., and K.M. Perryman (1986) Neural responses in rostralbrainstem nuclei of macaque monkeys after chronic trigeminal trac-totomy. J. Neurosurg. 65:508–516.
Young, R.F., T.D. Oleson, and K.M. Perryman (1981) Effect of trigeminaltractotomy on behavioral response to dental pulp stimulation in themonkey. J. Neurosurg. 55:420–430.
442 T. SUGIMOTO ET AL.





























