Embed Size (px)
Citation preview

CHAPTER – I
STATUS AND PREVALENCE OF
CRUSTACEA-BORNE TREMATODE
ZOONOSES IN THE REGION

2
INTRODUCTION
Zoonoses are the diseases which are naturally transmitted between animals
and humans. Parasitic zoonoses are well recognized and important public health
problems both in developing and developed countries. The burden of diseases caused
by food-borne pathogens (microbial, protozoal and macrobial) remains largely
unknown. Emerging microbial and protozoan zoonoses are major causes of diarrhoeal
disease in humans worldwide (Koopmans and Duizer, 2004; Smith et al., 2007). A
wide spectrum of helminthic infections is transmitted to humans by contamination of
food and water (Lloyd and Soulsby, 1998; Eckert et al., 2000; Muller, 2001; Vander
Hoek et al., 2003). The human behavioural patterns including habits, customs,
traditions and socio economic practices are closely related to such infections.
Globalization of the food supply, increased international travel, increase in the
population of highly susceptible persons, and change in culinary habits are some
factors associated with the increased diagnosis of food-borne parasitic diseases
worldwide (Dorny et al., 2009). Zoonotic parasitic diseases are transmitted to humans
either by ingesting environmentally robust transmissive stages (spores, cysts, ova,
larval and encysted stages) or by eating raw or undercooked `meat containing
infective tissue stages. Humans can be final, intermediate or paratenic (maintenance)
or accidental hosts. While the transmissive stages of some of these zoonoses can be
transmitted directly (e.g. by animal-human contact or through contact with
contaminated faeces, soil and garbage), they can also be transmitted through
contaminated water and food. A variety of infective tissue parasite stages are
responsible for transmitting meat- and fish-borne zoonoses. Eating raw, undercooked,
cured, smoked, salted, pickled or air-dried meat can increase the risk of contracting
food-borne parasite zoonoses, especially when the preservation treatment is
inadequate. Food-borne trematode infections, acquired through eating raw,
improperly cooked or processed freshwater fish, shell-fish, crabs, or unwashed or
inadequately washed vegetables have been well recognized as a public health problem
as evident from a study carried out under the “Southeast Asian Ministries of
Education Organization Regional Tropical Medicine and Public Health Project”
(SEAMEO-TROPMED) (Eduardo, 1991). Clonorchiasis, paragonimiasis, fascioliasis,
fasciolopsiasis and other intestinal trematodiases are the most important diseases

3
contracted, and strategies to control food-borne trematode infections have been
identified (WHO, 1995; Theresa et al., 2000). Many crustacea including crabs and
crayfishes themselves constitute an important food resource in hill tribes. Freshwater
crabs are also consumed for purported medicinal and tonic properties, including
treatment of stomach ailments and physical injuries (Dai, 1999; Darren et al., 2008).
Crabs also play an important role as the second intermediate host in the life cycle of
several trematode parasites and serve as a biological vector for transmission of their
infective stages to the final host. Besides, these crustaceans have also been reported as
potential vectors for cestode and acanthocephalan parasites from many parts of the
globe (Bratty et al., 1985; Thompson, 1985; Kuris and Gurney, 1997; Torchin et al.,
2001).
Crustacea-, snail- and water (aquatic vegetation) - borne trematodiases:
The lungflukes of the genus Paragonimus have been known as one of the most
important zoonotic parasites causing paragonimiasis in man, for which crabs serve as
the transmitting agents. Adult worms mature in capsules in the lungs and deposit ova,
which can be found on expectoration. The disease is often misdiagnosed as
tuberculosis due to overlapping clinical symptoms including chest pain, cough,
haemoptysis and confusing radiological findings on chest radiography (Toscano et al.,
1995). The parasite can migrate to several other vital tissues including the brain along
nerve sheaths (Kusner and King, 1993). Humans, dogs, cats and wild carnivores serve
as definitive hosts and are infected by eating undercooked crabs, crayfish or shrimp.
Crabs which are sometimes pickled in wine or brine, are a major source of
Paragonimus infection (Calum and Macpherson, 2005). More than 50 species are
known to infect different mammalian hosts throughout the world and approximately
15 species are known to infect humans (Bunnag and Harinasuta, 1985). The best-
known species is Paragonimus westermani (Kerbert 1878) Braun 1899 - a human
parasite that can undergo development in as many as 16 different snail species and 50
crustacean species (Blair et al., 1999b). Paragonimiasis, affecting more than 22
million people worldwide, is due to infection with P. westermani, P. miyazakii, P.
skrjabini and P. heterotremus in Asia, P. africanus and P. uterobilateralis in Africa,
and P. mexicanus and other species in Latin and South America. Countries with large

4
numbers of cases include Asia, Nigeria, Cameroon, Peru and Ecuador (Blair et al.,
1997, 2005).
In Indian context, Chandler and Read (1961) indicated Bengal, Assam and
some other parts of the country as endemic foci of human paragonimiasis. In India,
human paragonimiasis was first reported in Manipur by Singh et al. (1982), followed
by more reports in later years (Singh et al., 1986, 1993, 2004). The metacercarial
stages recovered from the freshwater crab, Potamiscus manipurensis, in
Churachandpur area were grown to adult stage in experimental dog hosts and based
on morphological criteria the fluke was confirmed to be Paragonimus hueit’ungensis
Chung, 1977- a species originally reported from China (Singh, 2002). Two more
species of Paragonimus have been reported as occurring in Manipur- P. skrjabini
from the same crab host (P. manipurensis) and P. heterotremus from Indochinamon
manipurensis (Singh et al., 2006, 2011).
Suspected foci of human paragonimiasis have also been reported from some
locales in Arunachal Pradesh (Narain et al., 2003). In Changlang district of Arunachal
Pradesh, during a community based survey it was reported that 95% of the local
inhabitants had the habit of consuming crabs and crayfishes in half cooked or in raw
form. To investigate the possible effect of human infection, stool and sputum samples
were also collected from suspected foci of infection; of these, two stool samples were
found positive for Paragonimus eggs. Two species of crustacean hosts, Barytelphusa
lugubris mansoniana and Sartoriana spinigera, were surveyed from the same locality,
but only the former were found positive for metacercarial infection; up to 1000
metacercariae were recovered from a single crab host. The species was identified as
P. heterotremus, the species implicated in human infection (Narain et al., 2003). In
subsequent studies based on surveys carried out in Kharshang and Miao regions of the
same district, the occurrence of P. westermani was also reported (Tandon et al., 2007;
Devi et al., 2010). In suspected endemic areas of the region, P. heterotremus is
reported to be implicated in human pleuropulmonary paragonimiasis (Devi et al.,
2007).
Apart from Paragonimus, many other trematodes, members of the family
Microphallidae in particular, utilize crabs as the intermediate host (Anantaraman and
Subramoniam, 1976; Heard and Overstreet, 1983; Pung et al., 2002). The family
Microphallidae Ward, 1901 indicates a large assemblage of small sized (usually <
1mm) digenean taxa (that represent more than 160 species under 28 genera arranged

5
in 10 subfamilies), which characteristically harbour the intestine of all groups of
vertebrates, mainly ducks (Martorelli et al., 2004) and mostly rodents among
mammals (Deblock, 1971, 2008). From amongst all these microphallids, only one,
viz. Spelotrema brevicaeca is reported to infect humans (Fried et al., 2004). The
infective metacercarial stages of microphallid flukes commonly occur in Crustacea
and are known to undergo extensive organogenesis in these intermediate hosts
(Caveny and Etges, 1971; Heard and Overstreet, 1983). Many workers have made
significant contributions to the studies of microphallid life histories (Cable and
Hunninen, 1940; Stunkard, 1957, 1958; James, 1968; Deblock, 2008). Most of the
adult forms have been recovered from aquatic birds, which get infected probably by
consumption of infected crustaceans. The taxon-rich digenean superfamily
Microphalloidea Ward, 1901 is an assemblage of 18 or more families, the taxonomic
history of which is rather complex and not free from debatable opinions regarding the
placement of various taxa; like most digenean trematodes microphallid taxa also
encompass several species/genera with little morphological differentiation (Bray et
al., 2008).
In addition to the above mentioned fluke infections, infective stages of several
other trematodes (eg. the fasciolid, amphistomid and echinostomid species), may
occur in the same acquatic environment in consonance with the crustacean
transmitters of trematodiases. The amphistome Gastrodiscoides and the echinostomid
fluke Artyfechinostomum are among the trematodes that have a zoonotic potential.
Gastrodiscoides hominis Lewis et McConnell, 1876 is commonly found in caecum
and colon regions of pig and human, where pig is a normal host species (Ahluwalia,
1960; Kumar, 1980; 1999; Mas-Coma et al., 2005), the infection in humans being a
rare occurence (Dada-Adegbola et al., 2004). The exact life cycle is unknown but
probably similar as in other species of Gastrodiscidae, involving aquatic vegetation as
the second intermediate environment that is used for the encystment of the
metacercarial infective stage (Zablotski, 1964; Dutt and Srivastava, 1972; Mas-Coma
et al., 2007). Gastrodiscoidiosis has symptoms similar to diarrhoea and, if untreated,
might kill the patient, mostly children (Kumar, 1980). G. hominis has a wide
distribution throughout India including the states of Assam, Bengal, Bihar, Uttar
Pradesh, Madhya Pradesh and Orissa (Shrivastav and Shah, 1970; Murty and Reddy,
1980). Buckley (1964) reported a high prevalence of G. hominis in humans, mostly
children (around 41%), in Kamrup District of Assam. In a later study, carried out in

6
Meghalaya, this parasite was shown to have a pattern of seasonal prevalence (Roy and
Tandon, 1992). Apart from India, G. hominis is widely distributed in countries like
Pakistan, Burma, Thailand, Vietnam, Philippines, China, Kazakhstan, and Russia
(Buckley, 1939; Ahuwalia, 1960; Kumar, 1980; Harinasuta et al., 1987; Yu and Mott,
1994; Ivanov and Semenova, 2000).
Artyfechinostomum sufrartyfex Lane, 1915 (synonyms- A. malayanum Leiper,
1901 Railliet, 1925; Euparyphium malayanum Leiper, 1911; Testifrondosa cristata
Bhalerao, 1924; Paryphostomum mehrai, Jain, 1957; Neoartyfechinostomum subhrai
Agarwal, 1963; Rao et Niphadker, 1963) is an echinostome intestinal fluke of pigs
that causes echinostomiasis. The first human infection of A. sufrartyfex was reported
in an 8-year old girl in Assam (Lane, 1915). Although echinostomiasis occurs world-
wide, most human infections have been reported from East and Southeast Asia.
Despite being rare, at least 19 species of echinostomes from 8 genera have been
reported in humans from China, Indonesia, Japan, Korea, Malaysia, Russia, Taiwan
and Thailand (Wanachiwanawin and Ungkanont, 2001). In India, the occurrence of A.
sufrartyfex has been reported from the states of Andhra Pradesh, Assam, Bihar, Tamil
Nadu, Uttar Pradesh and West Bengal (Beaver et al., 1984; Anonymous, 2005).
Another species of Artyfechinostomum, A. oraoni has been found to be of endemic
occurrence among the Oraon tribes of West Bengal (Bandopadhyay et al., 1989; Maji
et al., 1995). The human infection is acquired by ingestion of poorly cooked
freshwater fishes, prawns, crabs, molluscs and tadpoles (Adams and Motarjemi,
1999). Like most trematode eggs, identification of echinostome eggs as well as the
adult is difficult because of morphological similarity between biologically different
taxa and historical nomenclatural problems (Kostadinova and Gibson, 2000). Collar
spination, the first intermediate host (snail species) and geographical distribution are
the main diagnostic characters utilized for identification of the species; however,
sometimes these characters are also of limiting value (Morgan and Blair, 1995).
The present study was aimed at studying the trematode infections prevailing
among the edible crab species, the potential intermediate hosts for digenetic flukes in
selected localities in the region, Assam and Meghalaya in particular. For identifying
the species implicated in infection in the region, morphological criteria of the intra-
molluscan stages and metacercaria recovered were used including surface fine
topography. While the infective metacercaria stage is harboured in the crab host, the
preceding larval stages (sporocyst, redia, cercaria) develop in the snail (first

7
intermediate) hosts. Therefore, in order to ascertain the prevalence status of these
infections, snail hosts were also included in the survey study.
MATERIALS AND METHODS
1. Survey of intermediate hosts
With reference to the previously recorded cases, the survey was carried out in some
selected pockets of Arunachal Pradesh, Assam, Meghalaya, Nagaland and Tripura
wherein the native populations use crabs and snails as part of their food.
(i) Crabs: In rural markets of Northeastern India, crabs are commonly sold as a food
item. In rural localities and countryside of the region, eating of crabs is a common
food practice among the natives. Therefore, the collection and survey of crabs were
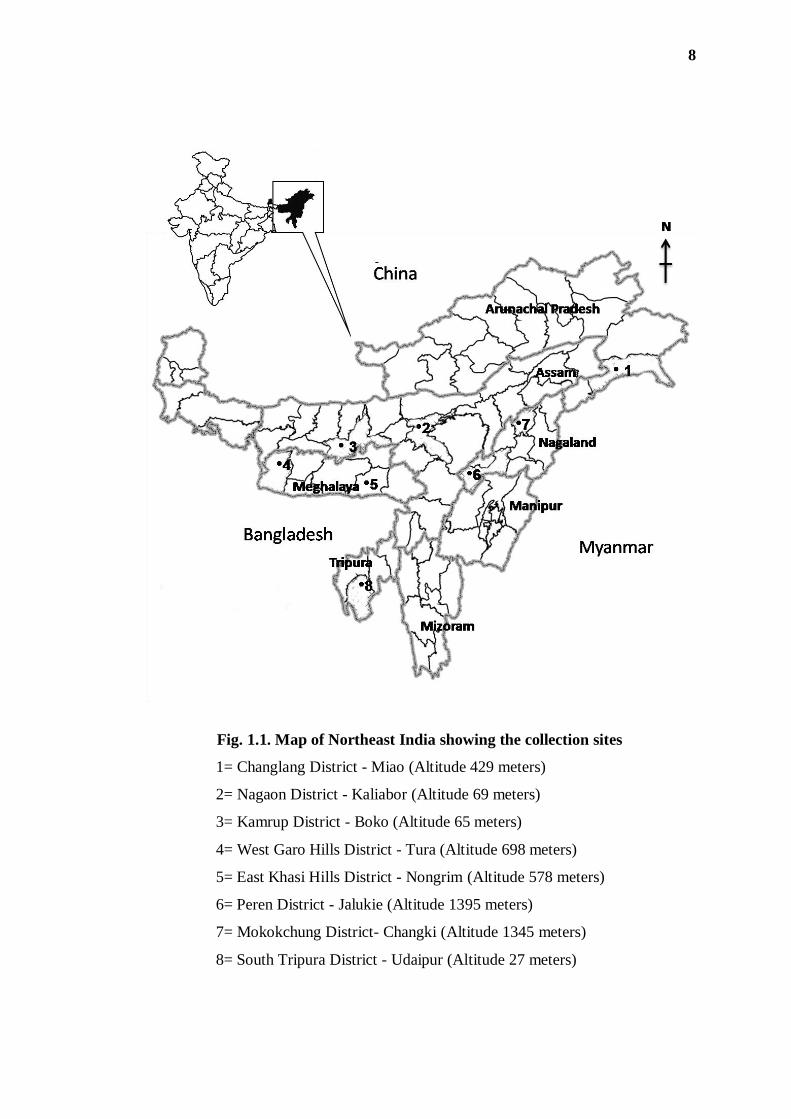
restricted to a selected few sites, which are shown in the map of the region (Fig. 1.1).
A total of 1548 crustacean hosts representing 3 species were examined for the
recovery of metacercariae, if any from them (Table 1.1). Naturally infected freshwater
crabs were periodically collected, mostly from mountain streams, paddy fields and
local markets in suspected foci of infection (Fig. 1.2, 1.3a-c). Further collections were
continued in only those locations where the crustaceans were found positive for
infection.
(ii) Snails: These gastropod molluscs, serving as the first intermediate host in the life
cycle of most trematodes, harbour in their soft tissues the pre metacercarial
developmental stages like the sporocyst and redia, with the latter giving rise to many
cercariae that emerge from the snail and invade the second intermediate host. Keeping
this fact in mind, snails were collected from all those aquatic bodies and localities
wherever the crab hosts were suspected to be positive for metacercarial infections
(Fig. 1.3d-g). Hundreds of snail specimens representing 4 genera with one species
each were examined for recovery of intra-molluscan stages of digenetic trematodes
(Table1.2).
8
Fig. 1.1. Map of Northeast India showing the collection sites
1= Changlang District - Miao (Altitude 429 meters)
2= Nagaon District - Kaliabor (Altitude 69 meters)
3= Kamrup District - Boko (Altitude 65 meters)
4= West Garo Hills District - Tura (Altitude 698 meters)
5= East Khasi Hills District - Nongrim (Altitude 578 meters)
6= Peren District - Jalukie (Altitude 1395 meters)
7= Mokokchung District- Changki (Altitude 1345 meters)
8= South Tripura District - Udaipur (Altitude 27 meters)

9
Fig. 1.2. A collection site in Garo Hills and crabs for sale in the rural market
area

10
Table 1.1. Surveyed areas for crab hosts for recovery of metacercarial stages
Sl. No Locality Species Status of infection
1 Arunachal Pradesh
Miao Barytelphusa lugubris mansoniana +
Sartoriana spinigera _
2 Assam
Nagaon B. l. mansoniana +
Kamrup Lobothelphusa fungosa _
3 Meghalaya
Tura B. l. mansoniana +
Nongrim B. l. mansoniana and _
S. spinigera _
4 Nagaland
Mokokchung B. l. mansoniana _
Peren B. l. mansoniana _
5 Tripura
Udaipur B. l. mansoniana _

11
Table 1.2. Surveyed areas for snail hosts for recovery of intramolluscan
trematode larval stages
Sl. No Locality Species Status of infection
1 Arunachal Pradesh
Miao Brotia costula +
2 Assam
Nagaon Bellamya bengalensis +
3 Meghalaya
Tura Paludomus loricatus +
B. bengalensis +
Melanoides tuberculata _
Nongrim M. tuberculata _

12
Fig. 1.3. Crab and molluscan species surveyed
a. Barytelphusa lugubris mansoniana (Henderson)
b. Lobothelphusa fungosa (Alcock)
c. Sartoriana spinigera (Wood-Manson)
d. Paludomus loricatus (Reeve)
e. Bellamya bengalensis (Kobelt)
f. Melanoides tuberculate (Muller)
g. Brotia costula (Rafinesque)

13
2. Recovery of the parasite material
(i) From Crabs: Metacercariae were isolated from the muscles of the crustacean host
by digestion technique. The crabs were cut into small pieces with the help of scissors
and digested by overnight incubation at 37oC in the artificial gastric juice
[Composition: HCl conc. (35 – 37%) 7-10 ml, distilled water – 1000 ml, pepsin
(1:10000) – 6 g].
The digested materials were filtered through mesh wire sieves and the
filterable sediments were washed repeatedly with tap water in order to get a clearer
supernatant. The sediment was examined for metacercariae under a dissecting
stereoscopic microscope.
(ii) From Snails: Snails to be examined were kept in a water-filled glass beaker and
exposed to sunlight for half-to-one hour prior to recovery of intra-molluscan stages.
Exposure to sunlight stimulates the shedding of cercariae from the infected snail into
the water. The shell of the mollusc specimen was broken open and its soft tissues
teased in a petri dish containing water. The contents were examined under a
stereoscopic microscope for recovery of sporocyst, redia or cercaria stages, which
were picked up and duly processed for further morphological studies.
3. Light microscopy (LM)
Freshly recovered metacercariae from crab hosts and intra-molluscan larval stages
were washed in 0.7% saline solution and narcotized with few drops of 70% ethyl
alcohol. The metacercariae were gently excysted using a fine needle, flattened
between a slide and a cover glass, fixed in 70% ethyl alcohol and stained with Borax
carmine and Mayer‟s carmellum stains, followed by dehydration through usual
dehydration media, i.e., ascending grades of ethyl alcohol, clearing in Methyl
benzoate and mounting in Canada balsam using standard protocols. For histological
study, serial sagittal sections of the fresh frozen material were also cut at a thickness
of 8-10 µm, using Leica CM 1850 cryotome.
The intra-molluscan larvae were also duly processed for whole mount
preparations following the standard protocol. All the prepared permanent slides were
observed and studied using the various optical systems available in the laboratory-

14
Wild M5APO stereo microscope, vision analyser, Zeiss image analyser, Leica DM
1000 image analysis system and Leitz Ortholux-2 research microscope.
Measurements of the specimens were taken using stage and ocular micrometers
and/or morphometric software in the image analyzer.
For identification of the parasite standard reference works were followed
(Yamaguti, 1971; Bray et al., 2008).
4. Scanning electron microscopy (SEM)
The isolated metacercariae were fixed in 2.5% glutaraldehyde in 0.1M sodium
cacodylate buffer for 6 hour at 4°C, washed in phosphate buffered saline and
dehydrated with ascending grades of acetone to pure dried acetone. The specimens
were then treated with Tetra methyl silane (TMS) in lieu of critical point drying
following Roy and Tandon (1991). The gold-coated specimens were observed using
LEO 435 VP scanning electron microscope at electron-accelerating voltages ranging
between 10 and 20 kV.
5. Analysis of prevalence data
The following parameters were used to analyze the data following Bush et al. (1997):
(i) Prevalence - the number of individuals in a population estimated to be infected
with a particular species of parasite (usually expressed in percentage).
(ii) Range - minimum to maximum number of individuals of a particular parasite
species in infected hosts in a sample.
(iii) Mean intensity - the average intensity, i.e., the total number of parasites of a
particular species found in a sample divided by the number of hosts infected; and
(iv) Abundance - the total number of individuals of a particular parasite species in a
sample of a particular host species divided by the total number of hosts (including both
infected and uninfected) of that species examined.

15
OBSERVATIONS/RESULTS
1. Larval trematode infection among crabs and molluscan hosts in
the suspected areas:
(i) In crab hosts: Of the three edible crab species surveyed from various localities in
Arunachal Pradesh, Assam, Meghalaya, Nagaland and Tripura only one, i.e.,
Barytelphusa lugubris mansoniana was found to be naturally infected with
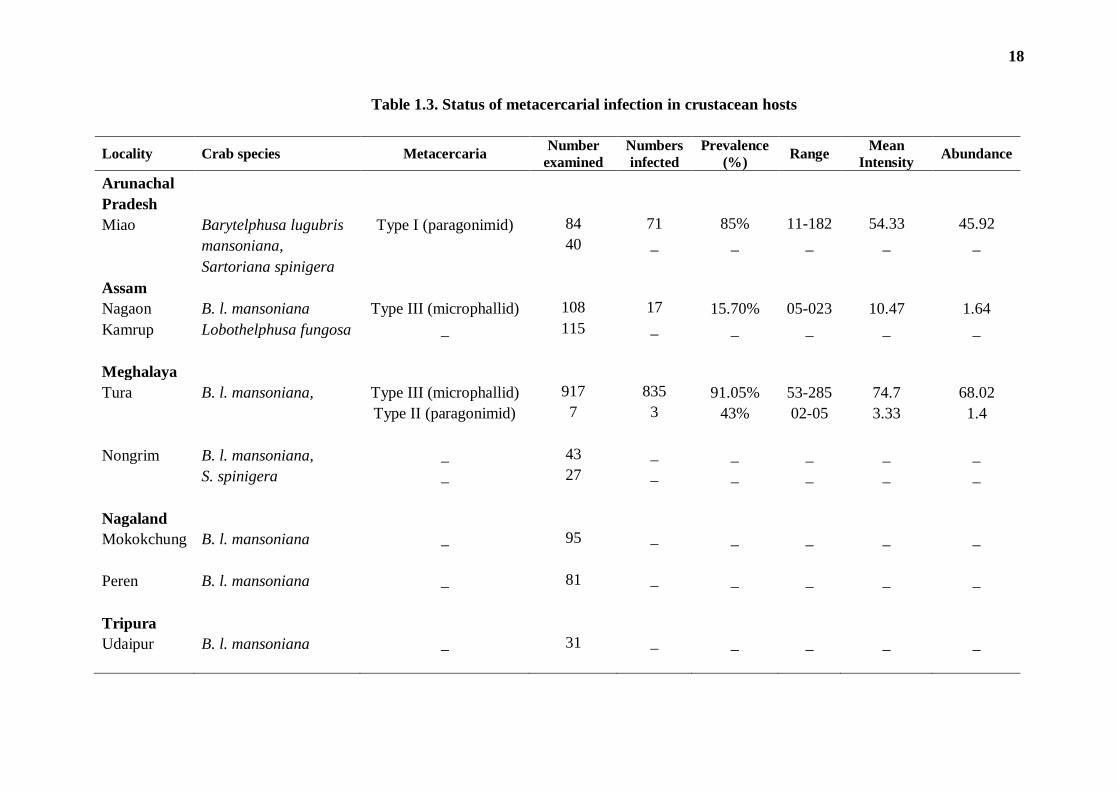
metacercarial forms. The distribution data of the parasites recovered are presented in
Table 1.3. The crab collected from only three localities, namely- Miao (in Changlang
District of Arunachal Pradesh), Tura (in West Garo Hills District of Meghalaya), and
Nagaon (in Assam) were found positive for metacercarial infection; B. l. mansoniana
from all other collection sites (in Nagaland and Tripura) did not harbour any infection.
The collection comprised metacercariae of three morphologically distinct
types. The type - I and type - II metacercariae both had characteristics of juvenile
Paragonimus flukes, whereas the type - III metacercaria showed a typical of
microphallid flukes. As a rare instance of occurrence, the type - II metacercaria was
recorded only once despite repeated surveys of crustacean hosts during the entire
period of the study. Though representing a paragonimid species, this metacercaria was
distinctly differentiable from the type - I metacercaria. The latter, also a paragonimid,
was recovered only from Barytelphusa crabs from the foci of infection in Arunachal
Pradesh. A high prevalence (85%) of the type - I metacercaria was recorded in the
study site. The crabs from the Tura region showed a very high prevalence (91.05%) of
infection of type - III metacercaria. These microphallid metacercariae were recovered
in high numbers (ranging between 53-285) per infected host and showed a high mean
intensity as well. In comparison, the crabs surveyed from Nagaon region also
harboured the similar infection, though with a low prevalence and low intensity of
infection.
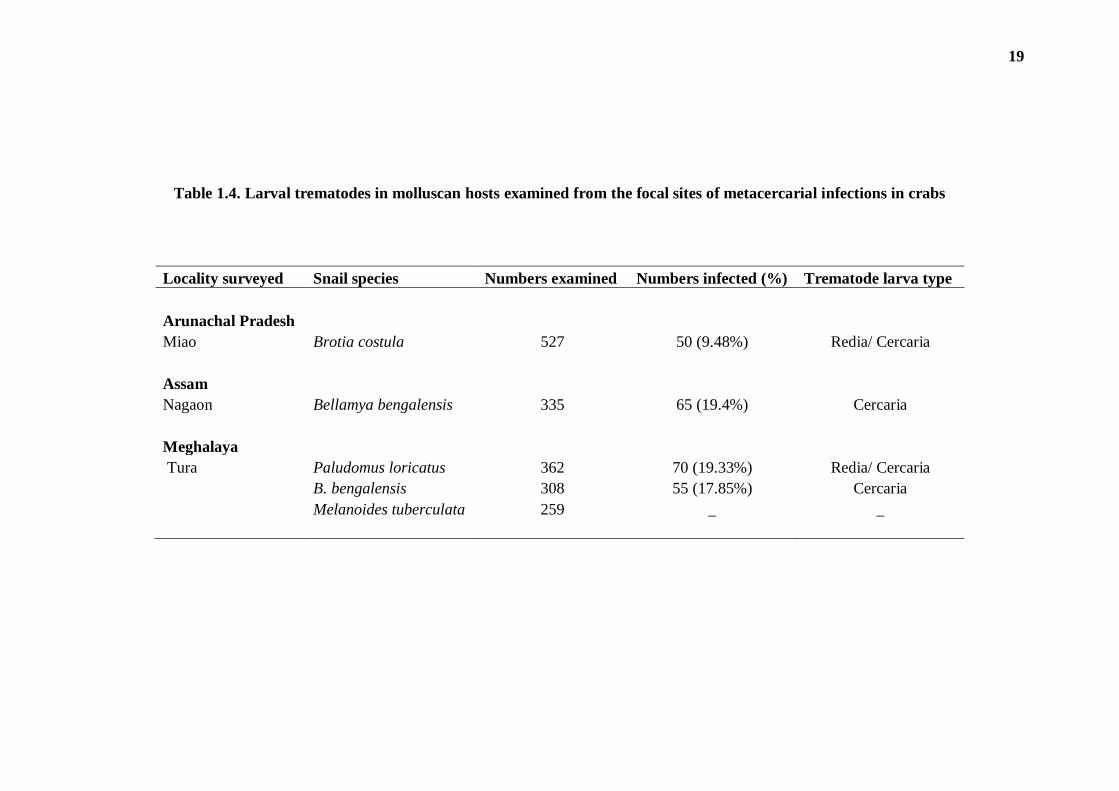
(ii) In snail hosts: With a view to recovering the possible infection of larval
trematodes from the molluscan host, snails were also screened only in those areas
where crabs showed positive for the metacercarial infection. The details of molluscan
hosts examined from the various localities and the status of infection among them is

16
depicted in Table 1.4. Of the four species of snails, prevalent in the study site, three,
viz. Paludomus loricatus, Bellamya bengalensis and Brotia costula were found
infected with trematode larvae; Melanoides tuberculate did not show any infection.
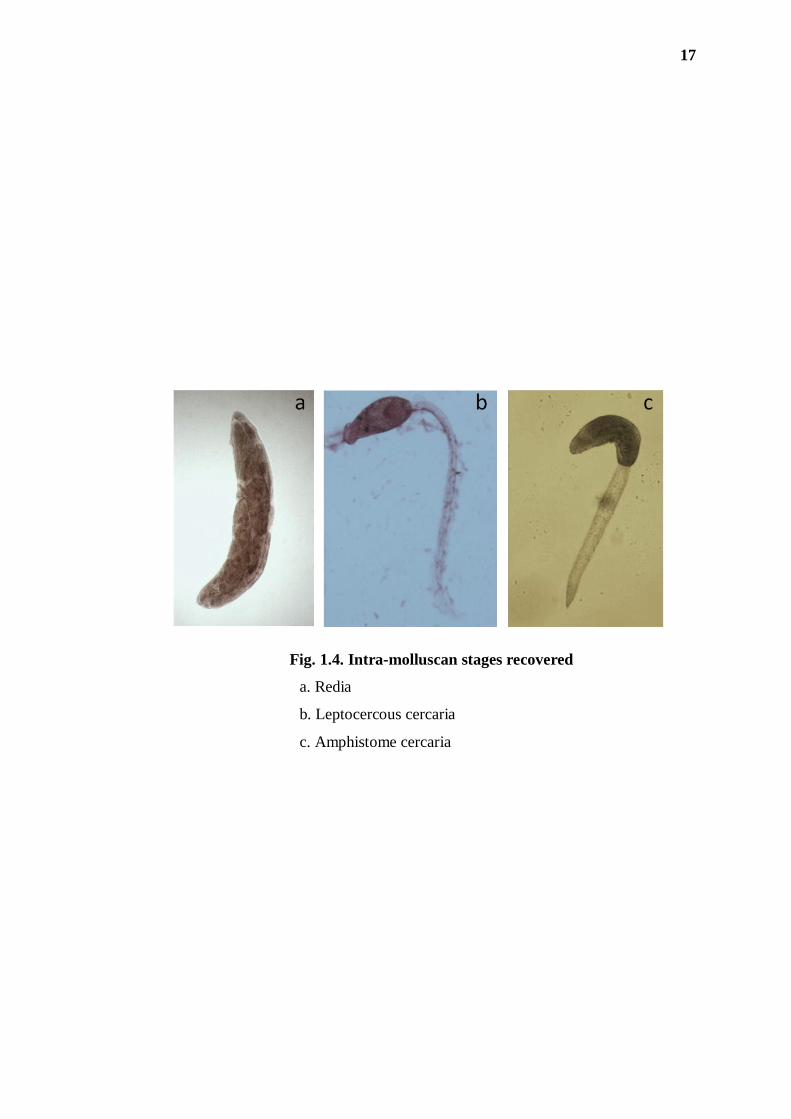
While P. loricatus from Meghalaya and B. costula from Arunachal Pradesh harboured
both redia and cercaria stages (Fig. 1.4a,b), B. bengalensis from Assam harboured
only cercaria (Fig. 1.4c). In morphological identification, the cercarial stages were
revealed to be of two types – one a distome leptocercous (with long slender tail) and
opthalmate cercaria having a pair of eye spots (Fig. 1.4b), and the other an
amphistome cercaria having its ventral sucker located at the base of the fore body
(Fig. 1.4c).
17
Fig. 1.4. Intra-molluscan stages recovered
a. Redia
b. Leptocercous cercaria
c. Amphistome cercaria
18
Table 1.3. Status of metacercarial infection in crustacean hosts
Locality Crab species Metacercaria Number
examined
Numbers
infected
Prevalence
(%) Range
Mean
Intensity Abundance
Arunachal Pradesh
Miao Barytelphusa lugubris Type I (paragonimid) 84 71 85% 11-182 54.33 45.92
mansoniana,
40 _ _ _ _ _
Sartoriana spinigera
Assam
Nagaon B. l. mansoniana Type III (microphallid) 108 17 15.70% 05-023 10.47 1.64
Kamrup Lobothelphusa fungosa _ 115 _ _ _ _ _
Meghalaya
Tura B. l. mansoniana, Type III (microphallid) 917 835 91.05% 53-285 74.7 68.02
Type II (paragonimid) 7 3 43% 02-05 3.33 1.4
Nongrim B. l. mansoniana, _ 43 _ _ _ _ _
S. spinigera _ 27 _ _ _ _ _
Nagaland
Mokokchung B. l. mansoniana _ 95 _ _ _ _ _
Peren B. l. mansoniana _ 81 _ _ _ _ _
Tripura
Udaipur B. l. mansoniana _ 31 _ _ _ _ _
19
Table 1.4. Larval trematodes in molluscan hosts examined from the focal sites of metacercarial infections in crabs
Locality surveyed Snail species Numbers examined Numbers infected (%) Trematode larva type
Arunachal Pradesh Miao Brotia costula 527 50 (9.48%) Redia/ Cercaria
Assam
Nagaon Bellamya bengalensis 335 65 (19.4%) Cercaria
Meghalaya
Tura Paludomus loricatus 362 70 (19.33%) Redia/ Cercaria
B. bengalensis 308 55 (17.85%) Cercaria
Melanoides tuberculata 259 _ _

20
2. Morphology of metacercariae
(i) Metacercaria type - I (paragonimid metacercaria - I):
The metacercarial cyst is oval in shape with a smooth surface. The body
surface of the excysted metacercaria is covered with numerous pointed tegumentary
spines. The tegument in the circum-oral region has a dense aggregation of small
spines that are arranged in several circular rows. On the rim of the oral sucker and
adjacent area, a few dome-shaped papillae are present in a random fashion. The
tegument of the ventral sucker and its surrounding region and that of the general body
surface also has a dense spination. The excysted metacercaria is elongated, 0.815 mm
x 0.492 mm in size, the ventral sucker is situated somewhat pre-equatorially, larger
than the oral sucker; the intestinal caeca are long and extend up to the posterior end of
the body; the conspicuous excretory bladder extends medially in the intercaecal space.
The metacercarial juvenile form is identified as Paragonimus westermani (Fig. 1.5a-
c).
(ii) Metacercaria type - II (paragonimid metacercaria - II):
From the crab species B. l. mansoniana,, another paragonimid type of
metacercaria was also recovered, though in very low number (2-5). These
metacercariae are much smaller in size than the type – I. The metacercarial cysts are
oval in shape, measuring 0.058 mm in length and up to 0.048 mm in maximum width;
the metacercarial cyst has a single layered thin wall. The excysted metacercaria is
described below (Fig. 1.6, a-f).
Description (based on 5 metacercariae): The excysted metacercaria has an
elongated body, with an average length of 0.09 mm and 0.034 mm in width. The fluke
inside fills the entire space within the cyst. The ventral sucker in the juvenile fluke is
situated pre-equatorially and is almost equal to or slightly larger than the oral sucker.
The intestinal caeca are long and extend up to the posterior extremity of the body. The
excretory bladder is medially extended between the caeca till the posterior end.
These characters are in conformity with the morphology of paragonimid
metacercaria stages. Therefore, the metacercarial type-II form represents the genus
Paragonimus.
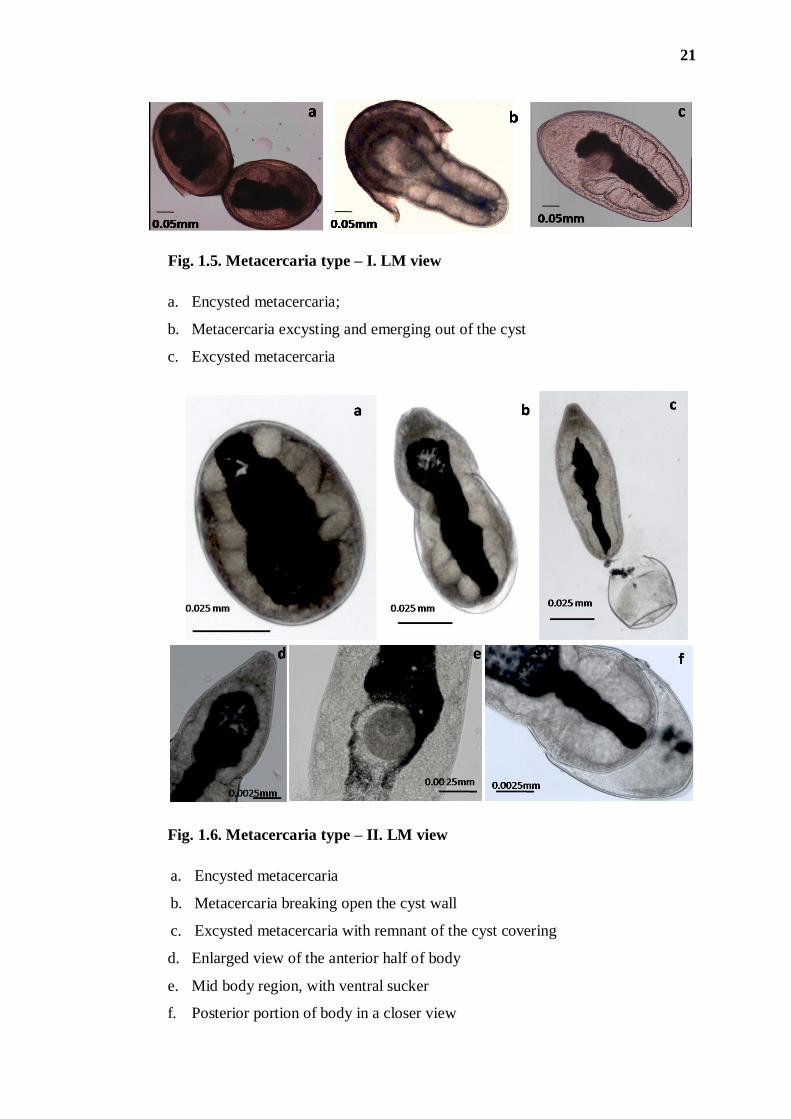
21
Fig. 1.5. Metacercaria type – I. LM view
a. Encysted metacercaria;
b. Metacercaria excysting and emerging out of the cyst
c. Excysted metacercaria
Fig. 1.6. Metacercaria type – II. LM view
a. Encysted metacercaria
b. Metacercaria breaking open the cyst wall
c. Excysted metacercaria with remnant of the cyst covering
d. Enlarged view of the anterior half of body
e. Mid body region, with ventral sucker
f. Posterior portion of body in a closer view

22
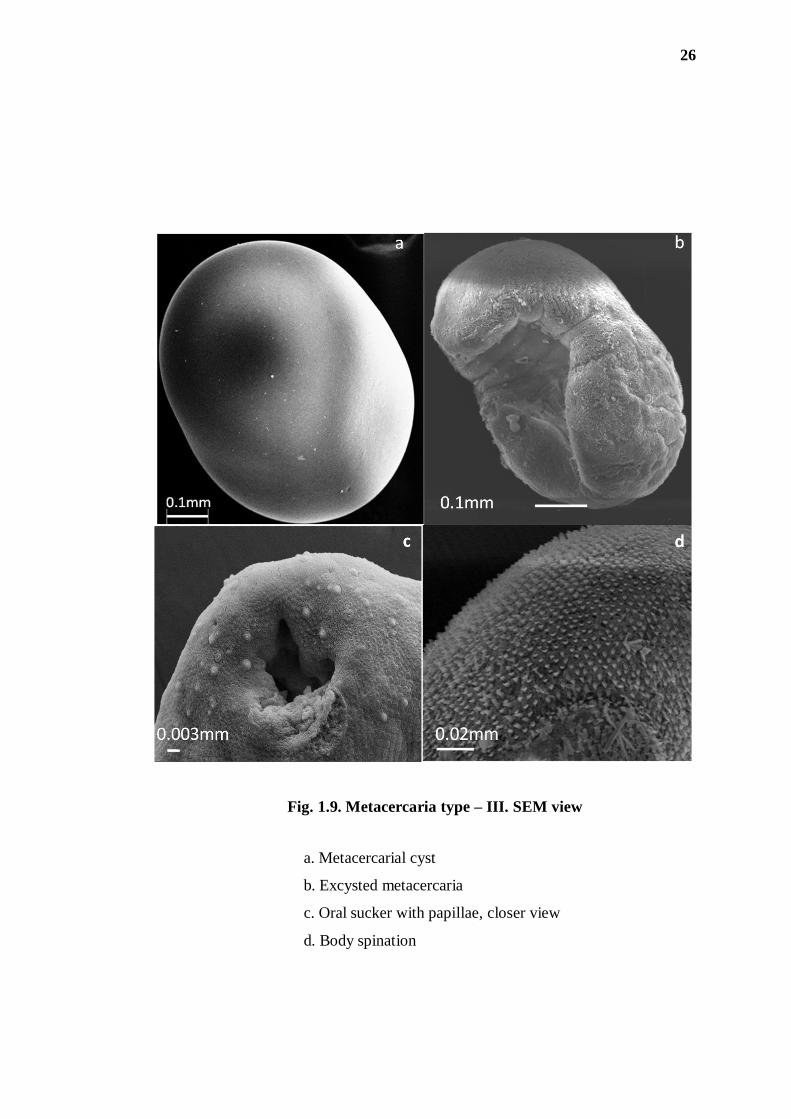
(iii) Metacercaria type - III ( = microphallid metacercaria):
The collection comprised a large number of type - III metacercarial cysts (53-
285) recovered from the infected crabs Barytelphusa lugubris mansoniana.
The cysts are elliptical in shape and have a prominent thick wall, which is
composed of two layers, the outer layer being thick but transparent. The excysted
metacercaria is described below.
Family Microphallidae Ward, 1901
Subfamily Microphallinae Ward, 1901
Genus Microphallus Ward, 1901
Description (based on 10 excysted specimens; Figs. 1.7, 1.8): Most of the
adult characters prominent; body pyriform in shape, minute in size (1.035 mm in
length, 0.655 mm in maximum width). Oral sucker subterminal; ventral sucker single
(not two), well developed, post equatorial in position. Prepharynx conspicuous;
pharynx small, short, muscular; oesophagus long, bifurcating in mid body region;
intestinal caeca short, ending in region at level of ventral sucker, pretesticular. Testes
symmetrical, located on either side of ventral sucker; cirrus pouch present, curved in
shape; cirrus (= phallus) well developed, conical, occupying entire genital atrium.
Genital atrium opening near by ventral sucker. Ovary located on right side of ventral
sucker or slightly overlapping right testis, oviduct emerging from mid posterior part
of ovary; uterus forming ascending and descending loops between testes and
excretory vesicle, containing numerous eggs. Vitelleria commencing from level of
testes, extending up to excretory vesicle; vitelline glandular cells arranged in two
groups- one with eight lobes on right, other with seven on left. Excretory vesicle „V‟
shaped, excretory pore terminal.
The morphometric measurements (mm) of the body and its organs are
provided in Table 1.5.
The SEM observations revealed the fine surface topography of the
metacercaria (Fig. 1.9). The metacercarial cyst has a smooth surface. The excysted
metacercaria is oval in shape and the entire body surface is covered with numerous
spines, the latter having digitated projections directed posteriorly; numerous papillae
abound in the circum-oral sucker region, where they appear to be randomly scattered
and not showing any pattern.
23
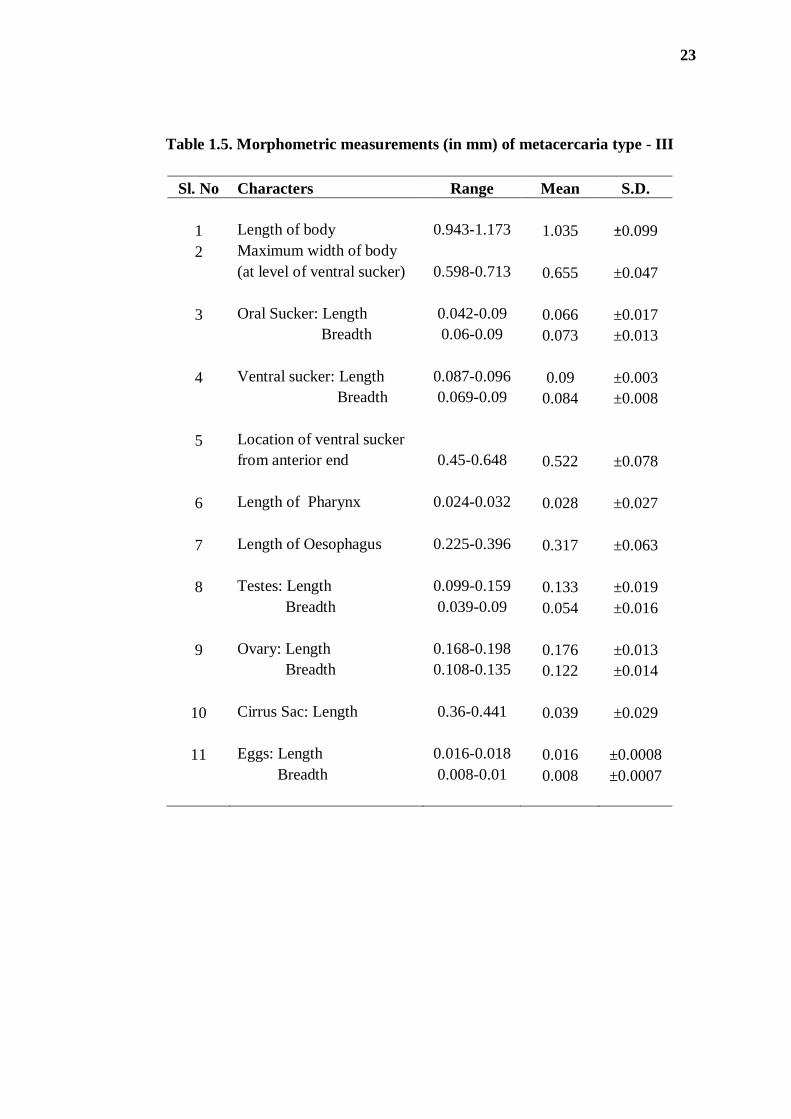
Table 1.5. Morphometric measurements (in mm) of metacercaria type - III
Sl. No Characters Range Mean S.D.
1 Length of body 0.943-1.173 1.035 ±0.099
2 Maximum width of body
(at level of ventral sucker) 0.598-0.713 0.655 ±0.047
3 Oral Sucker: Length 0.042-0.09 0.066 ±0.017
Breadth 0.06-0.09 0.073 ±0.013
4 Ventral sucker: Length 0.087-0.096 0.09 ±0.003
Breadth 0.069-0.09 0.084 ±0.008
5 Location of ventral sucker
from anterior end 0.45-0.648 0.522 ±0.078
6 Length of Pharynx 0.024-0.032 0.028 ±0.027
7 Length of Oesophagus 0.225-0.396 0.317 ±0.063
8 Testes: Length 0.099-0.159 0.133 ±0.019
Breadth 0.039-0.09 0.054 ±0.016
9 Ovary: Length 0.168-0.198 0.176 ±0.013
Breadth 0.108-0.135 0.122 ±0.014
10 Cirrus Sac: Length 0.36-0.441 0.039 ±0.029
11 Eggs: Length 0.016-0.018 0.016 ±0.0008
Breadth 0.008-0.01 0.008 ±0.0007
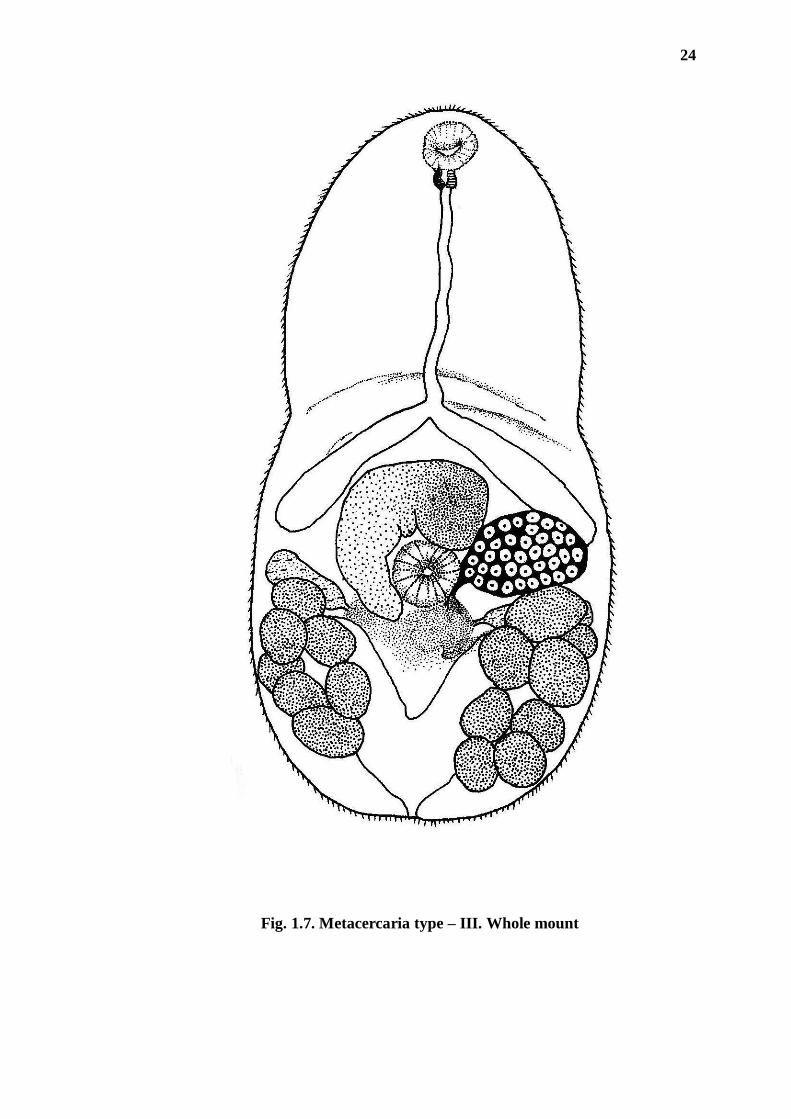
24
Fig. 1.7. Metacercaria type – III. Whole mount
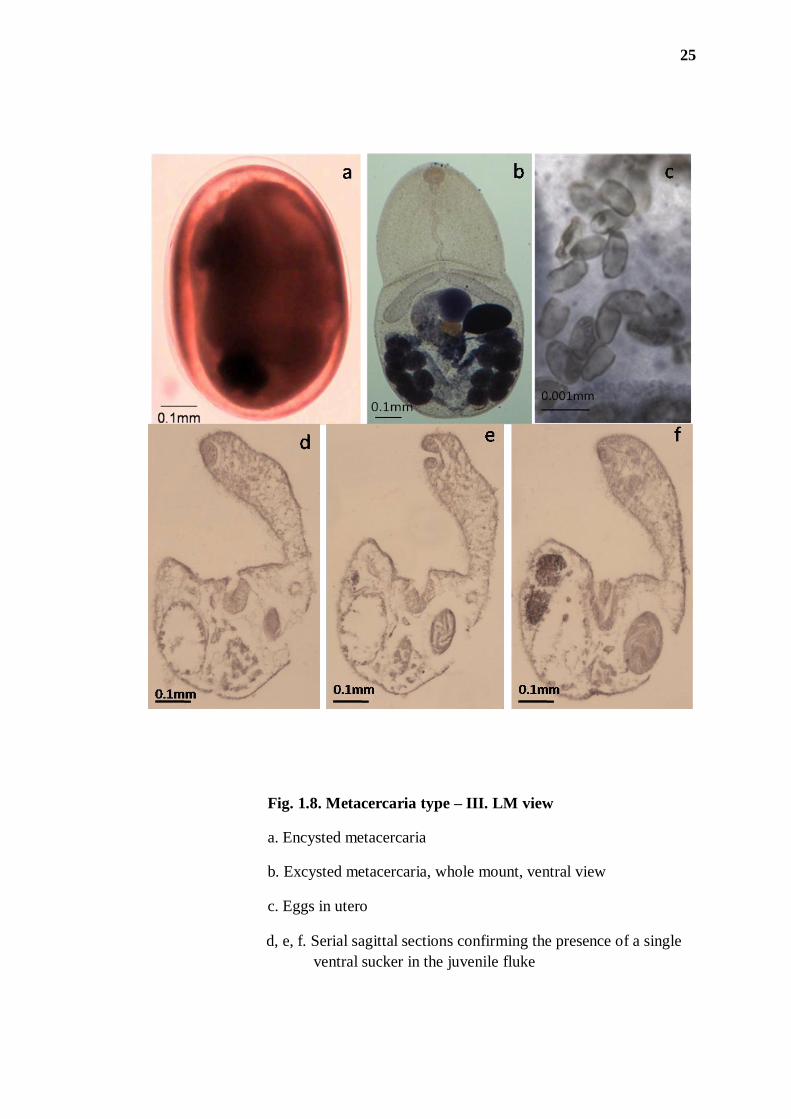
25
Fig. 1.8. Metacercaria type – III. LM view
a. Encysted metacercaria
b. Excysted metacercaria, whole mount, ventral view
c. Eggs in utero
d, e, f. Serial sagittal sections confirming the presence of a single
ventral sucker in the juvenile fluke
26
Fig. 1.9. Metacercaria type – III. SEM view
a. Metacercarial cyst
b. Excysted metacercaria
c. Oral sucker with papillae, closer view
d. Body spination

27
DISCUSSION
1. Prevalence status of trematode larval stages
During the field surveys carried out in different suspected foci of lungfluke
infection, the edible crab, Barytelphusa lugubris mansoniana was found to be
dominantly prevalent in the State of Arunachal Pradesh and Meghalaya, whereas in
Assam, in addition to this species, Lobotelphusa fungosa and Sartoriana spinigera
also abound. S. spinigera is available in Arunachal Pradesh as well. Of these three
crab species surveyed, only Barytelphusa crabs were found to be infected with three
types of metacercariae. In case of the study area in Arunachal Pradesh, the
Barytelphusa crab species were found to harbour metacercariae that were quite
different from those recovered from foci in Meghalaya and Assam and a high
prevalence (85%) of these metacercariae (type - I) was recorded from this
mountainous region. The metacercaria type - II, which in morphological characters
revealed conformity to paragonimid flukes, were recovered in very low numbers, just
2-5 per crab host. It was noticed that Barytelphusa crabs from Tura locality were
bearing yet another type of metacercarial infection, i.e., type - III was 91.05% and the
intensity high in crabs collected from Meghalaya; up to 285 metacercariae were
recovered from a single crab host. In Assam, however the intensity of infection was
rather low (15.70%, with maximum 23 metacercariae recovered from a single host).
The type - III forms revealed the characteristics of microphallid flukes.
In previous studies carried out with respect to Paragonimus infection in crabs,
Potamiscus manipurensis was reported to harbor metacercariae of Paragonimus spp
in Senapati District of Manipur (Singh and Singh, 1997). Later, Narain et al. (2003),
Tandon et al. (2007) and Devi et al. (2010) also reported the prevalence of
Paragonimus infection in B. lugubris crabs from Arunachal Pradesh and Meghalaya
respectively. Approximately 52% of children in Arunachal Pradesh (age ≤15 years)
were serologically found positive, of which 40% were expectorating Paragonimus
eggs in their sputum (Devi et al., 2010).

28
2. Morphology
Significant work has been done on metacercarial morphology of several
species of Paragonimus, namely P. skrjabini, P. iloktsuensis, P. ohirai, P.
pulmonalis, P. westermani, P. miyazaki, P. mexicanus, P. heterotremus (He et al.,
1982; Aji et al., 1984; Li et al., 1987; Higo and Ishi, 1987; Tongu et al., 1987;
Sugiyama et al., 1990; Jiang and Xia, 1993; Sugiyama et al., 2001) and microphallid
forms, viz. Microphallus abortivus from Ireland (Saville and Irwin, 1991), M.
sabanensis from Venezuela (Diaz et al., 2004); M. breviatus from Iceland (Kirill et
al., 2007); tegumental ultrastructure of Gynaecotyla squatarolae, Microphallus
koreana and Probolocoryphe uca has also been described (Guk et al., 2008; Lim et
al., 2008; Wafa et al., 2010).
On the basis of morphological criteria, the metacercarial type - I was
identifiable as Paragonimus westermani. The infection of P. westermani, a potential
zoonosis, has been reported to be dominantly prevalent in Barytelphusa spp. in
Arunachal Pradesh (Tandon et al., 2007). Though Devi et al. (2010) have reported the
occurrence of P. westermani in crabs of Meghalaya region, in the present study, this
species of lungfluke was not encountered from foci in even once during the entire
period of study.
With regard to the type - II metacercarial form, the main characters observed
were small, oval shaped metacercarial cyst having a thin and single layered cyst wall;
in the juvenile fluke the ventral sucker is located at the mid body level and
conspicuous intestinal caeca extend up to the posterior end of the body. These
characters are in conformity with the metacercarial stage of Paragonimus species
(Waikagul et al., 1989). In earlier studies from the Northeastern states, several species
of Paragonimus were recorded from different crab hosts; these include P.
hueit’ungensis and P. skrjabini from Potamiscus manipurensis; P. heterotremus from
Indochinamon manipurensis, all in Manipur (Singh et al., 2002, 2006, 2011); P.
heterotremus and P. westermani from B. l. mansoniana in Arunachal Pradesh (Narain
et al., 2003; Tandon et al., 2007); and P. westermani from B. l. mansoniana in
Meghalaya (Devi et al., 2010). But the present metacercarial form does not resemble
any of the above described species. On the basis of morphological characters the
present metacercaria (type - II) resembles Paragonimus macrorchis Chen, 1962, a
lungfluke species originally described from the crab host Potamon smithianus in

29
Thailand. However, to confirm the species status, the molecular taxonomic approach
is very much essential in this regard.
The type - III metacercaria in the present study bears the characters, namely
spiny body surface, ventral sucker at the middle third of the body, prepharynx distinct,
pharynx well developed, oesophagus moderately long, caeca short (may or may not
reach the level of testes); testes symmetrical, equatorial or at the posterior half of the
body, cirrus pouch more or less curved, ovary submedian and lateral to ventral sucker,
uterine coils confined to hind body, eggs small and excretory vesicle „V‟ shaped. In
having these features of morphology, particularly the precocious sexual development
while still in larval stage, the present form belongs to family Microphallidae Ward,
1901 [syn. Maritrematidae Nicoll, 1907]. As per Deblock (2008), there are nine
subfamilies under the family, which includes Microphallinae Ward, 1901;
Levinseniellinae Stiles & Hassall, 1901; Maritrematinae Nicoll, 1907;
Gynaecotylinae Guschanskaya, 1952; Pseudolevinseniellinae Tsai, 1955;
Basantisiinae Yamaguti, 1958; Sphairiotrematinae Deblock & Ky, 1966;
Androcotylinae Deblock & Heard, 1970; and Endocotylinae Deblock, 1971. After
considering all the characters, following the classification of Yamaguti (1971) and
Deblock (2008) the larval trematode form described herein comes under the
subfamily Microphallinae and the tribe Microphallini. There are six genera under the
tribe; these are: Microphallus (Microphallus) Ward, 1901 [Syns. Spelotrema
Jagerskiold, 1901; Monocaecum Stafford, 1903; Paraheterophyes Afanassief, 1941;
Carneophallus Cable & Kuns, 1951; Pseudocarneophallus Yamaguti, 1971;
Bulbovitellus Yamaguti, 1971; Feminacopula Ke, Liang & Yu, 1987; Microphallus
(Spelophallus) Jagerskiold, 1908]; Megalophallus Cable, Connor & Balling, 1960;
Atriophallophorus Deblock & Rose, 1964; Megalophalloides Ching & Ibanez, 1976;
Megistospermaticus Deblock & Canaris, 1992; and Rhyncostophallus Deblock &
Canaris, 1997. The present form comes close to the genus Microphallus in several
diagnostic characters viz. body linguiform, flattened; ventral sucker post equatorial,
without superficial veil; oesophagus medium sized; caeca short, divergent at the level
of ventral sucker; testes ovoid, post-ovarian and symmetrical; cirrus (phallus) atrial,
muscular and fleshy; genital atrium simple, enveloping the phallus closely; ovary
directly in front of right testes; uterus post caecal and coiled around the testis;
vitellaria formed of two relatively large clusters, each comprising 7-8 follicles,
vitelline ducts medium-sized, arched; and excretory vesicle short, post testicular, V-

30
shaped. In SEM observations the metacercarial cysts under the present study revealed
a smooth surface contour, whereas the excysted juvenile fluke has a spiny tegument
and numerous papillae in the region surrounding the oral sucker. Surface fine
topography of encysted and newly excysted metacercariae has been described in
respect of Microphallus abortivus, Cercaria sevillana, M. primas etc. in which a
spiny tegument and circum oral sensory papillae are the features of common
occurrence (Saville and Irwin, 1991; Pina et al., 2007, 2011).
There are only a few reports on the occurrence of Microphallidae flukes in
vertebrates in India. So far only a few microphallid taxa have been described and they
include: Levinseniella indica, Basantisia ramai and Pseudospeloterma indicum from
birds (Lal, 1936; Pande, 1938; Murhar, 1960; Bharadwaj, 1962); Mehraformes
jabalpurensis and Microphallus indicus from reptiles (Bharadwaj, 1963; Mukherjee &
Ghosh, 1967); Megalatriotrema hispidum from the common frog (Rao, 1969); and
Spelotrema narii from intestine of jackal (Rao, 1965). In addition to these,
microphallid metacercarial stages have also been reported from sand crabs and
brackish-water prawns near the South eastern coasts of the Indian Sub-continent
(Anantaraman and Subramoniam, 1976; Jayasree et al., 2001). A comparison of the
morphological features of the various microphallid species described so far from
crustacean hosts in India reveals a close similarity of the present metacercarial stage
with Microphallus indicus Mukherjee & Ghosh, 1967 which was originally described
from a reptilian host. Yamaguti (1972) included M. indicus under a new genus
Allomicrophallus providing a new combination [Allomicrophallus indicus (Mukherjee
& Ghosh, 1967)], differentiating it from Microphallus on the basis of characters like
the intestinal caeca being better developed, long ejaculatory duct and the male papilla
being non protrudable. However, while reviewing the family Microphallidae, Deblock
(2008) considered Allomicrophallus Yamaguti, 1972 (with Microphallus indicus
Mukherjee & Ghosh, 1967 as its type species) and Bengaliniella Deblock, Mukherjee
& Ghosh, 1970 (with Microphallus dicaecus Mukherjee & Ghosh, 1967 as its type
species) synonyms of Megalatriotrema Rao, 1969. According Deblock et al. (1970),
the latter genus has already been considered a senior synonym of Bengaliniella and
Allomicrophallus because of the synonymy of the type species M. hispidum Rao, 1969
with M. dicaecus and with M. indicus. However, pending a restudy of the material of
Megalatriotrema hispidum, in our opinion the species Microphallus indicus should be
retained in the genus Microphallus as originally described by Mukherjee and Ghosh

31
(1967). In view of foregoing, the metacercarial type – III under the present study is
identified herewith as Microphallus indicus Mukherjee & Ghosh, 1967.
The pre-metacercarial larval stages of most digenean flukes undergo
development in snail, the first intermediate host; the morphological features of the
cercaria larva in most cases are group-specific (Smith, 1994). For example, members
of the family Paragonimidae characteristically have microcercous xiphidocercariae,
which typically have a spiny body surface, short stumpy tail, prominent penetration
glands and an oral stylet. Likewise, the fork-tailed or furcocercous cercariae are
typical of strigeid and schistosome flukes. On the basis of morphological features of
the tail and position of the ventral sucker, the cercarial types recovered from snails
were identifiable as distome leptocercous type and amphistome type. Since these
cercariae were not microcercous xiphidocercaria type, it may be assumed that
Paragonims infection does not commonly prevail in the study area and that the larval
stages recovered may be representing other trematode species instead.
The survey results suggest that while the mountainous crabs in colder, high
altitude foci of Arunachal Pradesh predominantly harbour metacercariae of
paragonimid flukes, P. westermani in particular, the same crustacean species in
relatively warmer localities of Meghalaya and Assam seemingly do not sustain the
infective stages of lungfluke. Instead, in these areas with a tropical climate, it is the
microphalid infections, and not the paragonimid, which emerged as the highly
prevalent infection. Further study in this direction is warranted so as to establish the
influence of climate variables on the epidemiology of Crustacea-borne parasitic
infections.
*****





























