Embed Size (px)
Citation preview

Cmw
YS
a
KGlPIAAC
I
baScitllrDhdD
snttT
0d
Phytomedicine 18 (2011) 748– 755
Contents lists available at ScienceDirect
Phytomedicine
jou rn al hom epage: www.elsev ier .de /phymed
haracterization, molecular cloning, and in silico analysis of a novelannose-binding lectin from Polygonatum odoratum (Mill.)ith anti-HSV-II and apoptosis-inducing activities
un Yang1, Huai-long Xu1, Zi-ting Zhang, Jun-jie Liu, Wen-wen Li, Hua Ming, Jin-ku Bao ∗
chool of Life Sciences & State Key Laboratory of Oral Diseases, Sichuan University, Chengdu 610064, China
r t i c l e i n f o
eywords:alanthus nivalis agglutinin (GNA)-related
ectinolygonatum odoratum lectin (POL)n silico
a b s t r a c t
Polygonatum odoratum lectin (POL), a novel mannose-binding lectin with anti-viral and apoptosis-inducing activities, was isolated from rhizomes of Polygonatum odoratum (Mill.) Druce. POL was ahomo-tetramer with molecular weight of 11953.623 Da per subunits as determined by gel filtration,SDS-PAGE and mass spectrometry. Based on its N-terminal 29-amino acid sequence the full-length cDNA
nti-HSV-IIpoptosisaspase
sequence of POL was cloned. Subsequent phylogenetic analysis and molecular modeling revealed thatPOL belonged to the Galanthus nivalis agglutinin (GNA)-related lectin family, which acquired uniquemannose-binding specificity. The hemagglutinating activities of POL were metal ion-independent, andwere stable within certain range of pH and temperature alterations. Moreover, POL showed remarkableanti-HSV-II activity towards Vero cells, cytotoxicity towards human melanoma A375 cells and inducedapoptosis in a caspase-dependent manner.
© 2010 Elsevier GmbH. All rights reserved.
ntroduction
Plant lectins are proteins of non-immune origin that reversiblyind carbohydrate, agglutinate cells or precipitate polysaccharidesnd glycoconjugates (Goldstein et al. 1980; Sharon and Lis 1989;haron 2007). Recent advances in biochemical study, molecularloning, and structural analysis of lectins have categorized themnto seven families of structurally and evolutionarily related pro-eins, namely the legume lectins, the monocot mannose-bindingectins (also termed GNA-related lectins), the chitin-bindingectins, type II ribosome inactivating proteins (RIPs II), the jacalinelated lectins as well as the Cucurbitaceae phloem lectins (Vanamme et al. 1998, 2007). Among them, GNA-related lectins
ave been exclusively isolated from a subgroup of monocotyle-onous plants and consist of identical or similar subunits (Vanamme et al. 2000). Since the first GNA-related lectin namelyAbbreviations: POL, Polygonatum odoratum lectin; SDS-PAGE, sodium dodecylulfate-polyacrylamide gel electrophoresis; MS, mass spectrometry; GNA, Galanthusivalis agglutinin; HSV-II, Herpes Simplex Virus type II; RIPs II, type II ribosome inac-ivating proteins (RIPs II); MALDI-TOF, matrix-assisted laser desorption/ionizationime-of-flight; LDH, lactate dehydrogenase; PCL, Polygonatum cyrtonema lectin; TDL,yhonium divaricatum lectin.∗ Corresponding author. Tel.: +86 28 85415171; fax: +86 28 85415171.
E-mail addresses: [email protected], [email protected] (J.-k. Bao).1 These two authors contributed equally to this work.
944-7113/$ – see front matter © 2010 Elsevier GmbH. All rights reserved.oi:10.1016/j.phymed.2010.11.001
GNA was isolated from snowdrop (Galanthus nivalis) bulbs in 1987(Van Damme et al., 1987), more and more GNA-related lectinshave been purified and characterized in details with respect totheir biochemical properties, carbohydrate-binding specificitiesand various biological activities such as anti-viral, anti-tumor andanti-fungal activities (An et al. 2006; Tian et al. 2008; Li et al.2009; Liu et al. 2008a, 2009a,b,c, 2010a,b,c; Van Damme et al.1998).
Polygonatum odoratum (Mill.) Druce, a typical representative ofthe Liliaceae family, was an important traditional Chinese herbalmedicine from which a variety of biologically active compoundshave been identified (Huang 1999). However, little was knownabout the bioactive proteins of Polygonatum odoratum (Mill.) Druce.In this study, we purified a novel mannose-binding lectin fromrhizomes of Polygonatum odoratum (Mill.) Druce named as Polyg-onatum odoratum lectin (POL). Firstly, its N-terminal amino acidsequence was determined, thus leading to the molecular cloningof POL by 3′ and 5′ RACE. Subsequent sequence alignment analysisdemonstrated that POL belonged to the GNA-related lectin fam-ily, supported by the phylogenetic analysis of GNA-related lectinsrevealing the evolutional relationship between POL and the rest ofthis family. In addition, molecular modeling and molecular docking
of POL provided us more information regarding the carbohydrate-binding specificity of this lectin. We further characterized somesignificant biological properties of POL, in terms of its sugar-bindingspecificity, anti-viral and apoptosis-inducing activities.
edicin
M
C
fi
Mpddo3(3(RmW(aSmk
a
P
cgawl0wca
G
re(tdtoB
As
pM
tLt1f
Y. Yang et al. / Phytom
aterials and methods
hemicals and reagents
Rhizomes of Polygonatum odoratum (Mill.) Druce were collectedrom Pengzhou (Sichuan, China). Samples for RNA extraction werenstantly frozen in liquid nitrogen and stored at −70 ◦C before use.d-Mannose, Man-�(1,2)-Man, Man-�(1,3)-Man, Man-�(1,6)-
an, Man-�(1,3:1,6)-mannotriose, methyl �-d-mannopyranoside,-nitrophenyl �-d-mannopyranoside, d-fructose, d-glucose,-lactose, maltose, d-fucose, d-galactose, sucrose, N-acetyl--galactosamine, N-acetyl-d-glucosamine, thyroglobulin,vomucoid, acyclovir, dimethylsulfoxide (DMSO), isopropanol,-(4,5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazoliumbromideMTT), z-VAD-fmk (pan-caspase inhibitor), z-DEVD-fmk (caspase-
inhibitor), z-IETD-fmk (caspase-8 inhibitor) and z-LEHD-fmkcaspase-9 inhibitor) were purchased from Sigma (St. Louis, USA).abbit blood cells were obtained from local markets, and humanelanoma A375 cells were provided by Medical Sciences Center ofest China, Sichuan University. RPMI 1640 and fetal bovine serum
FBS) were purchased from GIBCO (New York, USA). CM Sepharosend Sephacyl S-100 were purchased from Pharmacia (Uppsala,weden). Standard molecular weight markers and gel filtrationarkers, pMD18-T vector, RNA Extraction Kit and 3′- and 5′-RACE
it were purchased from Takara Biotechnology, Dalian, China.All other chemicals used in this study were of the highest purity
vailable.
urification of Polygonatum odoratum lectin
The POL was purified as previously described with some modifi-ations (Liu et al. 2004; Chen et al. 2009). Briefly, plant samples wererinded and soaked overnight in 40 mM, NaAc-HAc buffer (pH 4.6)t 4 ◦C. After centrifugation (20,000 × g, 15 min), the supernatantas applied to a column of CM Sepharose (1.6 cm × 20 cm) equi-
ibrated with 40 mM, NaAc-HAc buffer (pH 4.6) and eluted with–0.5 M NaCl gradient. Fractions with hemagglutinating activityere pooled and concentrated to 3 mg/ml and loaded on a Sepha-
yl S-100 column equilibrated with 0.02 M PBS (pH 7.0). The elutedctive fractions were the purified POL.
el electrophoresis and mass spectrometry analysis
Protein analysis and molecular mass determination were car-ied out firstly with sodium dodecyl sulfate-polyacrylamide gellectrophoresis (SDS-PAGE) according to Laemmli and Favre1973). The native molecular mass was estimated by gel filtra-ion chromatography on the Sephacryl S-200 column as previouslyescribed (Andrews 1964). Profiles of matrix-assisted laser desorp-ion/ionization time-of-flight (MALDI-TOF) mass spectrum werebtained using a Voyager-RP mass spectrometer (PerSeptiveiosystems) (Woo et al. 2001).
ssays for hemagglutination activity and carbohydrate-bindingpecificity
Hemagglutinating activity of POL was performed as describedreviously with some modifications (Chen et al. 2009; Oda andinami 1986).To determine the sugar-binding specificity of POL, sugars were
ested for their abilities to inhibit POL-induced hemagglutination.
ectin concentration, just beyond the end point titre of hemagglu-ination, was chosen 30 �l of assayed lectin and equal volume of00 mM sugar or 2 mg/ml of glycoprotein were incubated at 37 ◦Cor 1 h.e 18 (2011) 748– 755 749
Effect of temperature, pH and metal ion on POL hemagglutination
The temperature and pH stabilities were tested as previouslydescribed for the Tricholoma mongolicum lectin (Lv et al. 2009).De-metallization of the purified POL was performed according toPaulová et al. (1971). The hemagglutination activities of POL in pre-and post-demetallized samples were compared.
Analysis of N-terminal amino acid sequence
The purified POL was subjected to SDS-PAGE and electro-transferred to a polyvinylidene difluoride (PVDF) membrane(BIO-RAD) followed by staining with Coomassie brilliant blue R-250. The band corresponding to the size of POL was excised and theN-terminal amino acid sequence of purified POL was determinedby using a Hewlett-Packard HP G1000A Edman degradation unitand a HP 1000 HPLC System (Lam et al. 1998).
RNA isolation and molecular cloning of POL gene
Total RNA was extracted using RNA Extraction Kit (TakaraBiotechnology, Dalian, China) and reversely transcribed with cDNAsynthesis primer AP (5′-GGCCACGCGTCGACTAGTAC (T) 21-3′)and primer 1 (5′-NAARACNCARTGYYTNCAYGCNTG-3′), which wasdesigned according to the N-terminal amino acid sequence of POL.3′-RACE was performed essentially according to the manufacturer’sinstructions.
Polymerase chain reaction (PCR) was carried out as follows: 30 sat 95 ◦C, 30 s at 50 ◦C, and 30 s at 72 ◦C, 32 cycles. The PCR prod-uct was cloned into pMD18-T vector (TaKaRa, Dalian, China) andsequenced. The 5′ end of POL cDNA was amplified with 5′-RACEusing specific primers designed based on the 3′ end sequence asdescribed (Matz et al. 1999).
The specific primers for 5′-RACE were: Primer2: 5′-CACCGCTGGTCCGCACAAT-3′; Primer 3: 5′-CGGGAAGGAGGACCATTA-3′; Primer 4: 5′-CAGAACGATGGGAA-CCTC-3′. The PCR reaction was 95 ◦C, 30 s; 60 ◦C, 30 s; 72 ◦C, 30 s.The obtained PCR product was sequenced as described above.
With the nucleotide sequence of the 3′- and 5′-RACE prod-ucts, the amplification of full-length cDNA of POL was acquiredusing Primer 5: 5′-ATGGCAGCTAGTAGTCCAAT-3′ and Primer 6: 5′-TTACTTAAGCAATGCTCTA-3′.
In silico analysis of POL
DNA full-length sequence was analyzed by SignalP 3.0 server(Bendtsen et al. 2004; Emanuelsson et al. 2007) and PROSITEtools (http://www.expasy.org/prosite/) (De Castro et al. 2006).Sequences of the GNA-related lectins were acquired by searchingNCBI with BLASTp (Stephen et al. 1990) as well as from curatedliterature. Alignment of selected sequences was carried out usingprogram CLUSTAL W (version 1.81) (Altschul et al. 1990; Thompsonet al. 1994). Corresponding phylogenetic tree was constructedwith Molecular Evolutionary Genetics Analysis (MEGA 4) software(Kumar et al. 2004; Tamura et al. 2007). The molecular modelingof POL was achieved with MODELLER9v7 (Sali and Blundell 1993)using the crystal structure of GNA [PDB code: 1jpc] as the template.Docking experiments were carried out with DOCK6 (Morris et al.1998).
Assay for anti-viral activity
Cell cultureVero cells were grown in RPMI medium (GIBCO) supplemented
with 10% FCS (GIBCO) and 100 �g/ml penicillin (Chongqing Yaoyou

7 edicine 18 (2011) 748– 755
PP
E
owavmo
A
obtttcvuT
Am
C
R1te
C
pwp2d
A
TtdkpC
C
2icmmwn
S
i
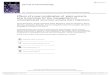
Fig. 1. Molecular mass determination of POL. (A) Tricine-SDS-PAGE analysis. Lane1, molecular weight standards (kDa): phosphorylase b (94 kDa) bovine serum albu-
50 Y. Yang et al. / Phytom
harmaceutical Co., Ltd) and 100 �g/ml Streptomycin (Shanghai 4th
harmaceutical Co., Ltd).
xamination of cytotoxicityVero cells were seeded onto 96-well plate with a concentration
f 5 × 104 cells per ml and a volume of 20 �l per well. POL samplesere serially diluted with PBS and aliquots of each dilution were
dsorbed on Vero cell monolayer. At the end of the adsorption theirus inoculum was removed and cells were replenished with freshedium. After incubation at 37 ◦C with 5% CO2 for 72 h, cells were
bserved and examined under electron microscopy.
nti-viral activity in infected cellsVero cells were seeded onto 96-well plate with a concentration
f 5 × 104 cells per ml and a volume of 20 �l per well. After incu-ation at 37 ◦C with 5% CO2 for 1 h, cells were then washed threeimes with PBS. Different concentrations of testing samples werehen added to culture well in 2-fold dilution. Maximum concentra-ion of POL (0.5 mg/ml) was used. Acyclovir was used as a positiveontrol for HSV assay. After incubation at 37 ◦C with 5% CO2 for 72 h,irus replication in infected cells was assayed. The amount of virussed in each experiment was based on infected target cells of 100CID50 of HSV.
ssay of in vitro apoptosis-inducing capability in humanelanoma A375 cells
ell cultureThe human melanoma A375 cells, routinely cultured in
PMI-1640 medium containing 10% fetal bovine serum (FBS),00 units/ml streptomycin and 0.03% l-glutamine, were main-ained at 37 ◦C with 5% CO2 in a humidified atmosphere (Chengt al. 2009a; Liu et al. 2009c).
ell viability assayA375 cells were dispensed in 96-well flat bottom microtiter
lates at a density of 5 × 104 cells/ml. After 24 h incubation, cellsere treated with different concentrations of POL for various timeeriods. Cell viability was measured by the 3-(4,5-dimetrylthiazol--yl)-2,5-diphenyltetrazolium bromide (MTT) assay as previouslyescribed (Mosmann 1983).
poptosis assayA375 cells were cultured with or without 25 �g/ml POL for 24 h.
he ultrastructure of apoptotic cell was observed under an elec-ron microscopy (Hitachi 7000, Japan) (Cheng et al. 2010). Lactateehydrogenase (LDH) activity was assessed using a standardizedinetic determination kit (Zhongsheng, LDH kit, Beijing, China) asreviously described with some modifications (Charrier et al. 2002;heng et al. 2008a,b, 2009b).
aspase assayCell lysate from 1 × 106 cells was incubated at 37 ◦C for 2 h with
00 �M z-DEVD-fmk (caspase-3 inhibitor), z-IETD-fmk (caspase-8nhibitor), z-LEHD-fmk (caspase-9 inhibitor) and z-VAD-fmk (pan-aspase inhibitor). The activities of caspase-3, -8 and -9 wereeasured with a colorimetric assay kit (Biovision) according to theanufacturer’s instructions. 405 nm adsorption of assayed sampleas recorded with a microplate reader (Bio-Tek Instruments) andormalized as fold increase as compared to DMSO-treated cells.
tatistical analysis of the data
All data were confirmed with at least three independent exper-ments. These data were represented as Means ± S.D. Statistical
min (67 kDa), ovalbumin (43 kDa), carbonicanhydrase (30 kDa), trypsin inhibitor(20.1 kDa), and lactalbumin (14.4 kDa); lane 2, POL on Sephacryl S-100. (B) Molecularmass determination by MALDI-TOF.
significance were determined by Student’s t-test. P < 0.05 was con-sidered statistical significant.
Results
Purification of Polygonatum odoratum lectin
Crude extracts from rhizomes of polygonatum odoratum (Mill.)Druce were first applied to a CM-Sepharose column and eluted with0–0.5 M NaCl gradient in 40 mM, NaAc–HAc buffer (pH 4.6) andpeak fractions with hemagglutinating activity were collected. Thenthe pooled active fractions were concentrated and loaded on thecolumn of Sephacyl S-100. The peak fraction showing hemaggluti-nating activity was purified Polygonatum odoratum lectin (POL).
Molecular mass of POL
This purified lectin exhibited a single band on SDS-PAGEunder non-reducing and reducing conditions, corresponding to amolecular mass of approximately 12 kDa (Fig. 1A). Consistently,MALDI-TOF mass spectrum showed that the molecular mass of POLwas 11953.623 Da (Fig. 1B). However, the molecular mass of POLmeasured by gel filtration predicted a protein with 48 kDa (datanot shown), suggesting that the purified lectin was a tetramer offour 11.9 kDa subunits that were linked by non-covalent bonds.
Hemagglutinating activity and carbohydrates-binding specificityof POL
POL agglutinated rabbit erythrocytes at a minimal concentrationof 3.75 �g/ml, while exhibited no agglutination towards humanblood groups (A, AB, B and O) erythrocytes. In order to deter-mine the carbohydrate-binding specificity of POL, the residualagglutinating activity of POL was assayed in the presence of a spec-trum of carbohydrates. The inhibitory concentrations of sugars andglycoproteins are shown in Table 1. Among these sugars and glyco-proteins only mannoses and thyroglobulin were shown to stronglyinhibit the hemagglutinating activity of POL.
Effect of temperature, pH and metal ion on POL hemaggultinatingactivity
Hemagglutinating activity assay of POL upon heat treatmentover a wide range of temperatures indicated that its activity wasthermally stable from 20 to 60 ◦C. However, the activity started

Y. Yang et al. / Phytomedicine 18 (2011) 748– 755 751
Table 1Hemagglutinating activity inhibition by sugars.
Saccharide/glycoprotein Concentration required for 50%inhibition (mM)
d-Fructose 150 mMd-Mannose 90 mMMe �-d-Man 120 mMMan-˛(1,2)-Man 20 mMMan-˛(1,3)-Man 10 mMMan-˛(1,6)-Man 5 mMMan-˛(1,3:1,6)-mannotriose 2.5 mMThyroglobulin 50 mM
Tsm
t2hpEEsm
F
rDaarafnpiAarmtaa
P
sG
Fig. 2. Full-length cDNA sequence and deduced amino acid sequence of POL. ThecDNA full-length sequence consists of 172 amino acids and the mature proteincontains 110 amino acids. The start codon and the stop codon were underlinedin italic type, and the arrows indicate cleavage sites (including the signal peptides
Fmp
here was no inhibition by d-glucose, d-lactose, maltose, d-fucose, d-galactose,ucrose, N-acetyl-d-galactosamine, N-acetyl-d-glucosamine, p-nitrophenyl ˇ-d-an up to 200 mM; ovomucoid up to 4 mg/ml.
o decline rapidly afterwards, and was completely abolished after0 min exposure at 90 ◦C and 100 ◦C. Moreover, POL maintained itsemagglutinating activity within pH 3–9, while was inactivated atH above 9 or below 3. Furthermore, after incubation with 50 mMDTA for 12 h with continuous shaking and removal of excessiveDTA by exhaustive dialysis, POL’s total hemagglutinating activitytill remained, revealing that this activity was independent ofetal ions.
ull-length cDNA cloning of POL
Using automated Edman degradation, 29 amino acidesidues at its N-terminus were sequenced as follows:NSPHRLRKTQCLHACRSTIFIMQEDCNL. Therefore, a degener-te primer 1 was synthesized according to the underlined aminocid sequence. Sequencing of 3′ RACE product with this primerevealed a 506 bp fragment possessing high similarity with lili-ceous mannose-binding lectins. Based on the sequence of thisragment, a 296 bp fragment of 5′ end of POL was obtained byested PCR. With the nucleotide sequences of the 3′ and 5′ RACEroducts, the full-length cDNA of POL was amplified with 674 bp
n length, encoding a protein with 172 amino acids (Fig. 2).ccording to previous studies in predicting lectin signal peptidend C-terminal cleavage site, POL contained a 31-amino acidesidue signal peptide that was most likely to be cleaved from theature lectin between amino acid residues A31 and D32; whereas
he cleavage site at the C-terminus was predicted between V141nd G142. Therefore, the mature POL possibly contained only 110mino acid residues.
hylogenetic analysis of POL
Sequence alignment analysis revealed that the POL proteinequence exhibited considerable high similarities to other typicalNA-related lectins (Fig. 3A), indicating that POL was probably a
ig. 3. Multiple alignment and phylogenetic analysis of POL. The sequences were aligneethods within MEGA 4 (B). These lectins possess the similar conserved motif of ‘QXDX
hylogenetic evolutionary tree revealed that POL has close evolutional relationship with
and C-terminal cleavage peptides). In addition, The three mannose-binding motifs‘QXDXNXVXY’ were marked in red. (For interpretation of the references to color inthis figure legend, the reader is referred to the web version of the article.)
typical member of the GNA-related lectin family. The amino acidsequences of GNA-related lectins shared significant similarity withGNA, including the conserved motif of ‘QXDXNXVXY’, which isessential in the mannose recognition (Van Damme et al. 2007).However, in the evolutional process, several sites in the three con-served motifs of GNA were probably mutated, thus it would resultin the changes of carbohydrate affinity. To determine the evo-lutionary relationships of POL with other GNA-related lectins, aphylogenetic tree was constructed using available sequences ofGNA-related lectins. The dendrogram consisted of three big clusters(Fig. 3B). Cluster a and b contained lectins mainly from species ofAmaryllidaceae and Orchidaceae, while the third cluster (cluster c)comprised the Liliaceae lectins including O. japonicus, P. cyrtonema,P. multiflorum and P. odoratum, suggesting that POL belonged to theGNA-related lectin family.
Molecular modeling of POL
Previously structural analysis of GNA domains suggested thatthey were composed of three repeating subdomains, and each sub-
domain had one mannose-binding sites with five conserved aminoacid residues (QXDXNXVXY), together conferring the unique speci-ficity to mannose-binding through a network of four hydrogenbonds interconnecting the hydroxyls C2–OH, C3–OH and C4–OH ofd by Clustal W program (A) and the tree was generated by neighbor-joining (NJ)NXVXY’ and their secondary structures are mainly composed by beta-sheets. Theother GNA-related lectins.

752 Y. Yang et al. / Phytomedicine 18 (2011) 748– 755
Fig. 4. The three-dimensional structure and docking experiments of POL (A) andGNA (B). The tertiary structure of POL was constructed taking GNA as a template:beta-sheets were indicated by patches. Turns and loops were indicated by lines andthe amino acids constituting the mannose-binding sites. Stereo view of docking intottt
mefitoTcfGaipsmPc
A
c
Table 2Anti-HSV-II activity of POL.
No. POL concentration(�g/ml)
Vero cells
40 20 10 5 2.5 1.25 HSV-II
1 – + + +++ +++ ++++ ++++ –2 – + ++ ++ ++++ ++++ ++++ –3 – – + ++ +++ ++++ ++++ –
“–”: no cell pathological changes; “+”: 25% cell pathological changes;“++”: 50% cell
FiA
he sugar-binding sites (I, II, and III) of GNA and POL were used to better understandhe sugar-binding specificities. Dashed lines corresponded to hydrogen bonds andheir distances were labeled.
annose (Hester et al. 1995; Hester and Wright 1996; Sauerbornt al. 1990). In our pursuit to understand the mannose-bind speci-city of POL, molecular modeling of POL was carried out based onhe X-ray coordinates of GNA. It was noteworthy that beta-strandsccurred predominantly in the structure of POL as shown in Fig. 4A.he overall folding of POL was typically derived from beta-strandsonnected by turns and loops, creating a very tight structural scaf-old. It was very similar with the three-dimensional structures ofNA (Fig. 4B), Polygonatum cyrtonema lectin (PCL) (Liu et al. 2008a)nd Ophiopogon japonicus lectin (OJL) (Zhang et al. 2010), suggest-ng that POL was a quite conserved protein in terms of its tertiaryrotein structure. Interestingly, although both POL and PCL pos-essed three sugar-binding sites, PCL exhibited only one conservedotif of ‘QXDXNXVXY’ instead of three in POL. This suggested that
OL might have different biological activities due to its distinctarbohydrate-binding abilities as compared to PCL.
nti-HSV-II activity of POL
The TCID50 (tissue culture infection dose) of HSV-II towards Veroells, determined by cytopathogenic effect (CPE), was 10−4 in the
ig. 5. POL inhibits A375 cells growth and LDH activity-based assay. (A) The A375 cells
nhibition ratios were measured by the MTT assay (n = 3, Means ± S.D.). (B) The cellular m375 cells were treated with 25 �g/ml POL for 12, 24, 36, and 48 h, and apoptotic and nec
pathological changes; “+++”: 75% cell pathological changes;“++++”: 100% cell patho-logical changes.
virus titration assay. Potential activity of POL in HSV-II inhibitionwas examined in HSV-II infected Vero cells. Although cell blankcontrol cultures and positive medicine control cultures showed nopathological changes after 72 h, remarkable pathological changeswere observed in negative medicine control cultures. However,POL displayed a dosage-dependent inhibition of HSV-II-inducedcell death by 33.3% and 16.7% observed at the concentrations of20 �g/ml and 10 �g/ml, respectively. And an average of 58.3% inhi-bition was observed at 2.5 �g/ml POL (Table 2). The 50% effectiveconcentration (EC50) obtained was comparatively low at approxi-mate 5–10 �g/ml.
POL induces apoptosis in A375 cells
POL caused remarkable inhibition of A375 cell growth in a time-and dosage-dependent manner. A375 cells treated with 25 �g/mlPOL for 24 h led to almost 50% inhibition in cell growth (Fig. 5A). Tocharacterize the POL-induced inhibition of A375 cell growth, mor-phological changes in cells after POL treatment were observed. Inthe presence of 25 �g/ml POL for 24 h, remarkable morphologicalalterations with apoptotic feature were observed under electronmicroscope (Fig. 5B). Furthermore, apoptosis was further evalu-ated by the measurement of LDH activity. In Fig. 5C, the number ofapoptotic cells was much more than that of necrotic cells, which
suggested that the major cause of POL-induced A375 cell deathwas apoptosis. Altogether, these results suggested that POL couldinduce apoptosis in A375 cells.were treated with various doses of POL for 12, 24, 36, and 48 h, respectively. Theorphology was observed without or with POL under electron microscopy. (C) Therotic ratio were measured by LDH activity-based assay (n = 3, Means ± S.D.).

Y. Yang et al. / Phytomedicin
Fig. 6. POL induces apoptosis in a caspase-dependent manner. (A) The A375 cellswere treated with POL for 24 h, with or without 200 �M z-DEVD-fmk, z-IETD-fmk,z-LEHD-fmk and z-VAD-fmk pretreatment at 37 ◦C for 2 h, and then cell growthinhibition growth inhibition was determined by the MTT assay (n = 3, Means ± S.D.).(tc
P
c(VsccFiPmb
D
vramcSltohSsfi
TT
B) After treatment with 25 �g/ml POL for 12, 24 or 36 h, the cytosolic fraction ofhe cells was analyzed for changes in the activities of caspase-3, -8 and -9 usingolorimetric assay (n = 3, Means ± S.D.).
OL induces A375 cell death in a caspase-dependent manner
To further evaluate the requirement of caspase in POL-inducedell death, the z-DEVD-fmk (caspase-3 inhibitor), z-IETD-fmkcaspase-8 inhibitor), z-LEHD-fmk (caspase-9 inhibitor) and z-AD-fmk (pan-caspase inhibitor) were applied as mentioned inection “Materials and methods”. After 24 h incubation with A375ells under POL treatment, these inhibitors were able to almostompletely inhibit the lectin-induced cell apoptosis as shown inig. 6A. Moreover, we observed the activities of caspase-3, -8 and -9n a time-dependent manner (Fig. 6B). These results suggested thatOL might induce A375 cells apoptosis via a caspase-dependentechanism. All these above-mentioned bioactivities of POL have
een summarized in Table 3.
iscussion
In this study, a novel mannose-binding lectin (POL) with anti-iral and anti-neoplastic activities has been isolated from thehizome of Polygonatum odoratum (Mill.) Druce. Electrophoreticnalysis, mass spectrometry, gel-filtration chromatography andolecular cloning indicated that POL was a homotetramer protein
omposed of four 11.9 kDa subunits, linked by non-covalent bonds.ince our study suggested POL as a new member of the GNA-relatedectin family, this result was not surprising considering the facthat most of the GNA-related lectins existed as dimmers, tetramers,r even octamers containing subunits of around 11.5–14 kDa andad no disulphide linkage between subunits (Peumans et al. 1986;
meets et al. 1994; Van Damme et al. 1992). And, subsequentequence alignment and phylogenetic analysis of POL further veri-ed that POL belonged to this family of lectins.able 3he important physicochemical and biological activities of POL.
Lectin name Polygonatum odoratum lectin
CharacteristicsAbbreviations POLPlant source Polygonatum odoratum (Mill.)
Druce, rhizomeMolecular weight of monomer Approximately 12 kDaMajor carbohydrate-binding specificity MannoseHemagglutinating activity ≥3.75 �g/ml
Anti-tumor activityHuman melanoma A375 cells Apoptosis (IC50: 25 �g/ml)Murine fibrosarcoma (L929) cells Apoptosis (IC50: 25 �g/ml)
Other biological activitiesVero cells infected with HSV-II Anti-HSV-II (EC50: about
5–10 �g/ml)
e 18 (2011) 748– 755 753
The majority of GNA-related lectins have hemagglutinatingactivity and carbohydrate affinity, and as a member of this familyPOL only agglutinated rabbit erythrocytes at a low concentra-tion of 3.75 �g/ml. Whereas, other GNA-related lectins like PCL,GNA and OJL exhibit hemagglutinating activity towards rab-bit erythrocytes with the minimal concentration at 0.98 �g/ml,1.8 �g/ml and 15.6 �g/ml, respectively (Van Damme et al. 1987;Tian et al. 2008; Zhang et al. 2010). The different hemaggluti-nating activity among various lectin members might result fromdistinct carbohydrate-binding properties of GNA-related lectins.Since among the carbohydrate we have tested so far, we can clas-sify this mannose specific lectin into ‘single-domain GNA-relatedlectins’ as all single-domain GNA-related lectins have an exclusivespecificity towards mannose and oligomannosides (Van Dammeet al. 2007).
Moreover, molecular modeling demonstrated that the three-dimensional structure of POL closely resembled that of otherGNA-related lectins, such as PCL (An et al. 2006; Liu et al. 2008a,2009e), OJL (Zhang et al. 2010) and GNA (Fig. 4B). However, thestructural difference was still obvious as PCL mainly existed as adimer (Ding et al. 2008) while POL reported here was a homote-tramer. These structural analyses of GNA-related lectin family areconsistent with the results of from our phylogenetic analysis whichsuggested that lectins from Liliaceae have taxonomically relatedrelationships. Furthermore, the analysis of the three subdomains ofPOL by molecular docking indicated that all of the three mannose-binding sites were conserved, which possibly explained the resultsof our carbohydrates-binding specificity tests that POL only pos-sessed mannose-binding activity. Whereas, with one mutatedcarbohydrate-binding site and two intact mannose-binding motifs,PCL showed affinities towards both mannose and sialic acid affini-ties.
In addition, many carbohydrate-specific lectins with plant ori-gin have shown remarkable anti-viral activity (Keyaerts et al. 2007).However, due to the diversified carbohydrate-binding specifici-ties, anti-viral activities of plant lectins vary considerably. Ourpresent finding showed that POL inhibited the cytopathic effect ofHSV-II with EC50 at 5–10 �g/ml. When compared with other twomannose-binding GNA-related lectins OJL and Tyhonium divarica-tum lectin (TDL) with anti-HSV-II EC50 of 3.93 �g/ml (Tian et al.2008) and 3.054 �g/ml (Luo et al. 2007), respectively, the anti-HSV-II effect of POL was slightly weaker than that of OJL and TDL.
To date numerous investigations have verified that plant lectinsdisplay anti-neoplastic activity (De Mejía and Prisecaru 2005;Liu et al. 2008b, 2009c,d,f; Yao et al. 2010; Li et al. 2010). POLalso showed remarkable inhibition of human melanoma A375 cellgrowth by inducing apoptotic cell death in a caspase-dependentmanner. Our previous study reported that POL induced L929 cellapoptosis with the IC50 at 25 �g/ml via a mitochondrial pathwaythrough activation of caspase and death-receptor (Liu et al. 2009f).In this study POL exerted almost the same growth inhibitory effecttowards A375 cells when compared with L929 cells. Besides this,another recent study in human melanoma A375 cells has demon-strated that PCL induces both apoptosis and autophagy througha mitochondria-mediated ROS-p38-p53 pathway with IC50 was15 �g/ml (Liu et al. 2009b,c). Comparing with PCL, POL showed rel-ative weaker cytotoxic activity towards A375 cells, and it mightresult from the distinct carbohydrate-binding specificity betweenthese two lectins. Similar with anti-HSV-II mechanism, a specula-tion has been proposed that POL might exert apoptosis-inducingfunction by binding with various cancer cell surface receptors in amannose-dependent manner (Lei and Chang 2009).
In summary, we purified POL from the rhizomes of polygonatumodoratum (Mill.) Druce and for the first time characterized some bio-logical characteristics, especially its anti-viral and anti-neoplasticactivities. Due to the accumulating evidence, our findings that POL

7 edicin
eotbcti
A
tcTraOT
R
A
A
A
B
C
C
C
C
C
C
C
D
D
D
E
G
H
H
H
K
54 Y. Yang et al. / Phytom
xhibited remarkable inhibition against Vero and A375 cells, wouldpen up a new exploration for natural lectins not only as a poten-ial therapeutic and preventative strategy against HSV-II infection,ut also a potential anti-neoplastic agent. Moreover, the molecularloning of POL gene would provide valuable information for fur-her understanding of the structure-function relationship throughts expression and mutagenesis.
cknowledgments
We are grateful to Dr. Ying Gu, Hao Peng and Chun-yang Li forheir critical reviews on this manuscript. We also thank Cheng-heng Zhou and Qi-jia Yu for their good suggestions on this work.his work was supported in part by grants from the National Natu-al Science Foundation of China (General Programs: No. 30670469nd No. 30970643) and Director Fund of State Key Laboratory ofral Diseases (Sichuan University) and the National Key Science &echnology Projects Fund (2009ZX8009-072B).
eferences
ltschul, S.F., Gish, W., Miller, W., Myers, E.W., Lipman, D.J., 1990. Basic local align-ment search tool. J. Mol. Biol. 215, 403–410.
n, J., Liu, J.Z., Wu, C.F., Li, J., Dai, L., Van Damme, E., Balzarini, J., De Clercq, E., Chen,F., Bao, J.K., 2006. Anti-HIV I/II activity and molecular cloning of a novel man-nose/sialic acid-binding lectin from rhizome of Polygonatum cyrtonema Hua.Acta Biochim. Biophys. Sin. (Shanghai) 38, 20–78.
ndrews, P., 1964. Estimation of molecular weights of proteins by Sephadex gel-filtration. Biochem. J. 91, 222–233.
endtsen, J.D., Nielsen, H., von Heijne, G., Brunak, S., 2004. Improved prediction ofsignal peptides: SignalP 3.0. J. Mol. Biol. 340, 783–795.
harrier, L., Jarry, A., Toquer, C., Bou-Hanna, C., Chedorge, M., Denis, M., Vallette,G., Laboisse, C.L., 2002. Growth phase-dependent expression of ICAD-L/DFF45modulates the pattern of apoptosis in human colonic cancer cells. Cancer Res.62, 2169–2174.
hen, J., Liu, B., Ji, N., Zhou, J., Bian, H.J., Li, C.Y., Chen, F., Bao, J.K., 2009. A novel sialicacid-specific lectin from Phaseolus coccineus seeds with potent antineoplasticand antifungal activities. Phytomedicine 16, 352–360.
heng, Y., Li, H.J., Ren, X.C., Niu, T.K., Hait, W.N., Yang, J.M., 2010. Cytoprotectiveeffect of the elongation factor-2 kinase-mediated autophagy in breast cancercells subjected to growth factor inhibition. PLoS One 5, e9715.
heng, Y., Qiu, F., Huang, J., Tashiro, S., Onodera, S., Ikejima, T., 2008a. Apoptosis-suppressing and autophagy-promoting effects of calpain on oridonin-inducedL929 cell death. Arch. Biochem. Biophys. 475, 148–155.
heng, Y., Qiu, F., Tashiro, S., Onodera, S., Ikejima, T., 2008b. ERK and JNK mediateTNFalpha-induced p53 activation in apoptotic and autophagic L929 cell death.Biochem. Biophys. Res. Commun. 376, 483–488.
heng, Y., Qiu, F., Ye, Y.C., Guo, Z.M., Tashiro, S., Onodera, S., Ikejima, T., 2009a.Autophagy inhibits reactive oxygen species-mediated apoptosis via activat-ing p38-nuclear factor-kappa B survival pathways in oridonin-treated murinefibrosarcoma L929 cells. FEBS J. 276, 1291–1306.
heng, Y., Qiu, F., Ye, Y.C., Tashiro, S., Onodera, S., Ikejima, T., 2009b. Oridonininduces G2/M arrest and apoptosis via activating ERK–p53 apoptotic pathwayand inhibiting PTK–Ras–Raf–JNK survival pathway in murine fibrosarcoma L929cells. Arch. Biochem. Biophys. 490, 70–75.
e Castro, E., Sigrist, C.J., Gattiker, A., Bulliard, V., Langendijk-Genevaux, P.S.,Gasteiger, E., Bairoch, A., Hulo, N., 2006. ScanProsite: detection of PROSITE sig-nature matches and ProRule-associated functional and structural residues inproteins. Nucleic Acids Res. 34 (Web Server issue), W362–W365.
e Mejía, E.G., Prisecaru, V.I., 2005. Lectins as bioactive plant proteins: a potentialin cancer treatment. Crit. Rev. Food Sci. Nutr. 45, 425–445.
ing, J.J., Bao, J.K., Zhu, D.Y., Zhang, Y., Wang, D.C., 2008. Crystallization and pre-liminary X-ray diffraction analysis of a novel mannose-binding lectin withantiretroviral properties from Polygonatum cyrtonema Hua. Prot. Pept. Lett. 15,411–414.
manuelsson, O., Brunak, S., von Heijne, G., Nielsen, H., 2007. Locating proteins inthe cell using TargetP, SignalP and related tools. Nat. Protoc. 2, 953–971.
oldstein, I.J., Hughes, R.C., Monsigny, M., Osawa, T., Sharon, N., 1980. What shouldbe called a lectin? Nature 285, 66.
ester, G., Kaku, H., Goldstein, I.J., Wright, C.S., 1995. Structure of mannose-specificsnowdrop (Galanthus nivalis) lectin is representative of a new plant lectin family.Nat. Struct. Biol. 2, 472–479.
ester, G., Wright, C.S., 1996. The mannose-specific bulb lectin from Galanthus nivalis(snowdrop) binds mono- and dimannosides at distinct sites. Structure analysisof refined complexes at 2.3 A and 3.0 A resolution. J. Mol. Biol. 262, 516–531.
uang, T.K., 1999. Handbook of Composition and Pharmacological Action ofCommonly-used Traditional Chinese Medicine. Chin. Med. Sci. Pub. Co., Shang-hai, pp. 664–667.
eyaerts, E., Vijgen, L., Pannecouque, C., Van Damme, E.J., Peumans, W., Egberink,H., Balzarini, J., Van Ranst, M., 2007. Plant lectins are potent inhibitors of coron-
e 18 (2011) 748– 755
aviruses by interfering with two targets in the viral replication cycle. AntiviralRes. 75, 179–187.
Kumar, S., Tamura, K., Nei, M., 2004. MEGA3: integrated software for MolecularEvolutionary Genetics Analysis and sequence alignment. Brief. Bioinform. 5,150–163.
Matz, M., Shagin, D., Bogdanova, E., Britanova, O., Lukyanov, S., Diatchenko, L.,Chenchik, A., 1999. Amplification of cDNA ends based on template-switchingeffect and step-out PCR. Nucleic Acids Res. 27, 1558–1560.
Laemmli, U.K., Favre, M., 1973. Maturation of the head of bacteriophage T4. I. DNApackaging events. J. Mol. Biol. 80, 575–599.
Lam, S.S., Wang, H., Ng, T.B., 1998. Purification and characterization of novel ribo-some inactivating proteins, alpha- and beta-pisavins, from seeds of the gardenpea Pisum sativum. Biochem. Biophys. Res. Commun. 253, 135–142.
Lei, H.Y., Chang, C.P., 2009. Lectin of Concanavalin A as an anti-hepatoma therapeuticagent. J. Biomed. Sci. 16, 10.
Li, C.Y., Meng, L., Liu, B., Bao, J.K., 2009. Galanthus nivalis agglutinin (GNA)-relatedlectins: traditional proteins. Burgeoning drugs? Curr. Chem. Biol. 3, 324–333.
Li, C.Y., Xu, H.L., Liu, B., Bao, J.K., 2010. Concanavalin A, from an old protein to novelcandidate anti-neoplastic drug. Curr. Mol. Pharmacol. 3, 37–42.
Liu, B., Bian, H.J., Bao, J.K., 2010a. Plant lectins: potential antineoplastic drugs frombench to clinic. Cancer Lett. 287, 1–12.
Liu, B., Cheng, Y., Liu, Q., Bao, J.L., Yang, J.M., 2010b. Autophagic pathways as newtargets for cancer drug development. Acta Pharmacol. Sin. 31, 1154–1164.
Liu, B., Wu, J.M., Li, J., Liu, J.J., Li, W.W., Li, C.Y., Xu, H.L., Bao, J.K., 2010c. Polygo-natum cyrtonema lectin induces murine fibrosarcoma L929 cell apoptosis andautophagy via blocking Ras-Raf and PI3K-Akt signaling pathways. Biochimie,doi:10.1016/j.biochi.2010.08.009.
Liu, B., Cheng, Y., Bian, H.J., Bao, J.K., 2009a. Molecular mechanisms of Polygonatumcyrtonema lectin-induced apoptosis and autophagy in cancer cells. Autophagy5, 253–255.
Liu, B., Cheng, Y., Zhang, B., Bian, H.J., Bao, J.K., 2009b. Polygonatum cyrtonema lectininduces apoptosis and autophagy in human melanoma A375 cells through amitochondria-mediated ROS-p38-p53 pathway. Cancer Lett. 275, 54–60.
Liu, B., Li, C.Y., Bian, H.J., Min, M.W., Chen, L.F., Bao, J.K., 2009c. Antiproliferative activ-ity and apoptosis-inducing mechanism of Concanavalin A on human melanomaA375 cells. Arch. Biochem. Biophys. 482, 1–6.
Liu, B., Min, M.W., Bao, J.K., 2009d. Induction of apoptosis by concanavalin A and itsmolecular mechanisms in cancer cells. Autophagy 5, 432–433.
Liu, B., Peng, H., Yao, Q., Li, J., Van Damme, E.J., Balzarini, J., Bao, J.K., 2009e. Bioinfor-matics analyses of the mannose-binding lectins from Polygonatum cyrtonema,Ophiopogon japonicus and Liparis noversa with antiproliferative and apoptosis-inducing activities. Phytomedicine 16, 601–608.
Liu, B., Xu, X.C., Cheng, Y., Huang, J., Liu, Y.H., Liu, Z., Min, M.W., Bian, H.J., Chen, J.,Bao, J.K., 2008a. Apoptosis-inducing effect and structural basis of Polygonatumcyrtonema lectin and chemical modification properties on its mannose-bindingsites. BMB Rep. 41, 369–375.
Liu, B., Zhang, B., Min, M.W., Bian, H.J., Chen, L.F., Liu, Q., Bao, J.K., 2009f. Inductionof apoptosis by Polygonatum odoratum lectin and its molecular mechanisms inmurine fibrosarcoma L929 cells. Biochem. Biophys. Acta 1790, 840–844.
Liu, Q., Wang, H., Ng, T.B., 2004. Isolation and characterization of a novel lectin fromthe wild mushroom Xerocomus spadiceus. Peptides 25, 7–10.
Liu, Z., Liu, B., Zhang, Z.T., Zhou, T.T., Bian, H.J., Min, M.W., Liu, Y.H., Chen, J., Bao, J.K.,2008b. A mannose-binding lectin from Sophora flavescens induces apoptosis inHeLa cells. Phytomedicine 15, 867–875.
Luo, Y.T., Xu, X.C., Liu, J.W., Li, J., Sun, Y.S., Liu, Z., Liu, J.Z., Van Damme, E., Balzarini,J., Bao, J.K., 2007. A novel mannose-binding tuber lectin from Typhonium divar-icatum (L.) Decne (family Araceae) with antiviral activity against HSV-II andanti-proliferative effect on human cancer cell lines. J. Biochem. Mol. Biol. 40,358–367.
Lv, H., Kong, Y., Yao, Q., Zhang, B., Leng, F.W., Bian, H.J., Balzarini, J., Van Damme, E.,Bao, J.K., 2009. Nebrodeolysin, a novel hemolytic protein from mushroom Pleuro-tus nebrodensis with apoptosis-inducing and anti-HIV-1 effects. Phytomedicine16, 198–205.
Morris, G.M., Goodsell, D.S., Halliday, R.S., Huey, R., Hart, W.E., Belew, R.K., Olson, A.J.,1998. Automated docking using a lamarckian genetic algorithm and an empiricalbinding free energy function. J. Comput. Chem. 19, 1639–1662.
Mosmann, T., 1983. Rapid colorimetric assay for cellular growth and survival:application to proliferation and cytotoxicity assays. J. Immunol. Methods 65,55–63.
Oda, Y., Minami, K., 1986. Isolation and characterization of a lectin from tulip bulbs.Tulipa gesneriana. Eur. J. Biochem. 159, 239–245.
Paulová, M., Entlicher, G., Tichá, M., Kostír, J.V., Kocourek, J., 1971. Studieson phytohemagglutinins. VII. Effect of Mn2+ and Ca2+ on hemagglutinationand polysaccharide precipitation by phytohemagglutinin of Pisum sativum l.Biochem. Biophys. Acta 237, 513–518.
Peumans, W.J., Allen, A.K., Cammue, B.P., 1986. A new lectin from meadow saffron(Colchicium autumnale). Plant. Physiol. 82, 1036–1039.
Sali, A., Blundell, T.L., 1993. Comparative protein modelling by satisfaction of spatialrestraints. J. Mol. Biol. 234, 779–815.
Sauerborn, M.K., Wright, L.M., Reynolds, C.D., Grossmann, J.G., Rizkallah, P.J., 1990.
Insights into carbohydrate recognition by Narcissus pseudonarcissus lectin: thecrystal structure at 2 A resolution in complex with alpha1-3 mannobiose. J. Mol.Biol. 290, 185–199.Sharon, N., 2007. Lectins: carbohydrate-specific reagents and biological recognitionmolecules. J. Biol. Chem. 282, 2753–2764.

edicin
SS
S
T
T
T
V
V
apoptosis-inducing activities. Process. Biochem. 45, 1477–1485.
Y. Yang et al. / Phytom
haron, N., Lis, H., 1989. Lectins as cell recognition molecules. Science 246, 227–234.meets, K., Van Damme, E.J., Peumans, W.J., 1994. Comparative study of the post-
translational processing of the mannose-binding lectins in the bulbs of garlic(Allium sativum L.) and ramsons (Allium ursinum L.). Glycoconj. J. 11, 309–320.
tephen, F.A., Warren, G., Webb, M., Eugene, W.M., David, J.L., 1990. Basic localalignment search tool. J. Mol. Biol. 215, 403–410.
amura, K., Dudley, J., Nei, M., Kumar, S., 2007. MEGA4: Molecular EvolutionaryGenetics Analysis (MEGA) software version 4.0. Mol. Biol. Evol. 24, 1596–1599.
hompson, J.D., Higgins, D.G., Gibson, T.J., 1994. CLUSTAL W: improving the sensi-tivity of progressive multiple sequence alignment through sequence weighting,position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 22,4673–4680.
ian, Q., Wang, W., Miao, C., Peng, H., Liu, B., Leng, F.W., Dai, L., Chen, F., Bao, J.K., 2008.Purification, characterization and molecular cloning of a novel mannose-bindinglectin from rhizomes of Ophiopogon japonicus with antiviral and antifungal activ-ities. Plant Sci. 175, 877–884.
an Damme, E.J., Allen, A.K., Peumans, W.J., 1987. Isolation and characterization of
a lectin with exclusive specificity toward mannose from snowdrop (Galanthusnivalis) bulbs. FEBS Lett. 215, 140–144.an Damme, E.J., Astoul, C.H., Barre, A., Rougé, P., Peumans, W.J., 2000. Cloningand characterization of a monocot mannose-binding lectin from Crocus vernus(family Iridaceae). Eur. J. Biochem. 267, 5067–5077.
e 18 (2011) 748– 755 755
Van Damme, E.J., Nakamura-Tsuruta, S., Smith, D.F., Ongenaert, M., Winter, H.C.,Rougé, P., Goldstein, I.J., Mo, H., Kominami, J., Culerrier, R., Barre, A., Hirabayashi,J., Peumans, W.J., 2007. Phylogenetic and specificity studies of two-domain GNA-related lectins: generation of mutispecificity through domain duplication anddevergent evolution. Biochem. J. 404, 51–61.
Van Damme, E.J., Peumans, W.J., Barre, A., Rougé, P., 1998. Plant lectins: a compositeof several distinct families of structurally and evolutionary related proteins withdiverse biological roles. Crit. Rev. Plant Sci. 17, 645–662.
Van Damme, E.J., Smeets, K., Torrekens, S., van Leuven, F., Goldstein, I.J., Peumans,W.J., 1992. The closely related homomeric and heterodimeric mannose-bindinglectins from garlic are encoded by one-domain and two-domain lectin genes,respectively. Eur. J. Biochem. 206, 413–420.
Woo, B.H., Lee, J.T., Na, D.H., Lee, K.C., 2001. Sepharose-unbinding ricin E as a sourcefor ricin A chain immunotoxin. J. Immunol. Methods 249, 91–98.
Yao, Q., Wu, C.F., Luo, P., Xiang, X.C., Liu, J.J., Mou, L., Bao, J.K., 2010. A new chitin-binding lectin from rhizome of Setcreasea purpurea with antifungal, antiviral and
Zhang, Z.T., Peng, H., Li, C.Y., Liu, J.J., Zhou, T.T., Yan, Y.F., Li, Y., Bao, J.K., 2010. Polyg-onatum cyrtonema lectin induces murine fibrosarcoma L929 cell apoptosis via acaspase-dependent pathway as compared to Ophiopogon japonicus lectin. Phy-tomedicine, doi:10.1016/j.phymed.2010.05.013.