Embed Size (px)
Citation preview

APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Nov. 1994, P. 4066-40710099-2240/94/$04.00+0Copyright © 1994, American Society for Microbiology
Characterization of Acinetobacter Type Strains and Isolates Obtainedfrom Wastewater Treatment Plants by PCR Fingerprinting
M. WIEDMANN-AL-AHMAD,' H.-V. TICHY,2 AND G. SCHONl*Institut fiir Biologie II/Mikrobiologie, D-79104 Freiburg,' and TUV-Sudwest,
Fachgruppe Biologische Sicherheit, D-79108 Freiburg,2 Germany
Received 17 May 1994/Accepted 6 September 1994
Acinetobacter type strains and isolates from wastewater treatment plants were differentiated by PCRfingerprinting. On the first level, PCR fingerprinting with two tRNA-gene specific primers (T5B and T3A) was
used for the identification of species (genospecies 1 to 17). On the second level, a single arbitrary primer (DAF4) was employed for strain differentiation. Upon comparison ofAcinetobacter type strains with 28 sewage sludgeisolates, 2 could be classified as belonging to A. johnsonii, 8 isolates could be classified as A. lwojfii, 8 could beclassified as A. baumannii, and 9 isolates were very closely related to the Acinetobacter species A. junii; only 1isolate could not be classified as one of the Acinetobacter type strains. The PCR fingerprinting method was
found to be a reproducible and fast method for differentiation and identification of Acinetobacter isolates.Because of some resulting discrepancies compared with previously described identification schemes, e.g.,DNA-DNA hybridization methods, the original identification experiments should be repeated and the resultsshould be reassessed.
Members of the genus Acinetobacter are a subject of intenseresearch because these ubiquitous organisms are gaining in-creasing importance in medicine (15), biotechnology (20, 21),and the environment (24). On that account, the fast andaccurate identification of Acinetobacter isolates is importantnot only for epidemiological, but also for ecological, research.The genus Acinetobacter as originally proposed by Brisou
and Prevot 1954 (10) comprises a heterogeneous collection ofbacteria which differ phenotypically and genotypically. Becausethey share many similarities in morphology and physiologywith members of the genera Moraxella, Branhamella, andNeisseria (13, 14), currently unresolved difficulties in identify-ing and classifying Acinetobacter strains exist.A number of recent studies have attempted to set up reliable
identification schemes based on DNA homology (7-9, 25), butuntil now there have still been some inconsistencies in theseidentification schemes. Moreover, they are not complete be-cause mostly clinical isolates were used in the development ofthese schemes. Soddell et al. (23) showed that none of thepublished phenotypically based identification systems for Acin-etobacter isolates performed well with nonclinical isolatesobtained from wastewater treatment pilot plants.
Several methods, such as biochemical assays, serologicalreactions, and the comparison of cell envelope protein patterns(1) or outer membrane protein patterns (16) as well asantibiotic resistance patterns, were not sufficient for the iden-tification of all species or strains within the genus Acineto-bacter.The aim of this study, therefore, was to use the recently
developed PCR fingerprinting technology for the differentia-tion of closely related strains of Acinetobacter isolated fromsewage plants or other biotopes and to obtain informationabout their relatedness to different Acinetobacter genospecies.
* Corresponding author. Mailing address: Institut fur Biologie II/Mikrobiologie, Schanzlestrasse 1, D-79104 Freiburg, Germany. Fax:49-761-203-2688.
MATERLALS AND METHODS
Bacterial strains and culture conditions. Acinetobacter bau-mannii LMD 85.20 (ATCC 15308), A. baumannii LMD 71.43(ATCC 13809), A. baumannii LMD 82.27 (ATCC 23220), A.baumannii LMD 82.33, A. baumannii LMD 82.47 (ATCC15150), A. baumannii LMD 85.46, A. calcoaceticus LMD 22.16(ATCC 23055T), A. haemolyticus LMD 85.26 (ATCC 17906T),A. johnsonii LMD 80.1 (ATCC 9036), A. junii LMD 58.1, andA. Iwoffli LMD 83.25 (ATCC 15309T, = A. iwoffli H) were
obtained from the Type Culture Collection, Kluyver Labora-tory of Biotechnology, University of Technology, Delft, TheNetherlands.A. baumannii CIP 70.34 (ATCC 19606T), Acinetobacter sp. 3
CIP 70.29 (ATCC 19004), Acinetobacter sp. 6 CIP A 165(ATCC 17979), Acinetobacter sp. 10 CIP 70.12 (ATCC 17924),Acinetobacter sp. 11 CIP 63.46 (ATCC 11171), Acinetobactersp. 13 CIP 64.2 (ATCC 17905), Acinetobacter sp. 14, Acineto-bacter sp. 15, Acinetobacter sp. 16 CIP 70.18 (ATCC 17988),Acinetobacter sp. 17, and A. radioresistens were obtained fromthe Collection de l'Institut Pasteur, Paris, France.A. baumannii (ATCC 1041T), A. anitratus, A. iwoffli (=A.
iwoffli II), A. Iwoffli (=A. Iwoffli I), and Acinetobacter sp. 9(=Acinetobacter sp. 9 A) were kindly provided by G. Auling,University of Hannover, Hannover, Germany; other Acineto-bacter sp. 9 strains were kindly provided by M. Wagner, Munich,Germany (=Acinetobacter sp. 9 M) and by P. Kampfer, Berlin,Germany (=Acinetobacter sp. 9 B), and strain 210 A was
provided by J. W. van Groenestijn, Wageningen, The Nether-lands, and by EAWAG/ETH, Kastanienbaum, Switzerland.The strains Fo-3, Br-2, and La 1 to La 25 were isolated from
wastewater treatment plants in Forchheim, near Freiburg (Fo);Braunschweig (Br); and Langen (La), Germany. The isolatesLa 1 to La 25 were obtained by N. Jardin, University ofDarmstadt, Darmstadt, Germany. All isolates and the strainsA. Iwoffli (I and II), A. anitratus, and strain 210 A wereidentified phenotypically and by their polyamine pattern; a
high diaminopropane content is characteristic for the Acineto-bacter species (2, 18).The strains Kingella kingae (ATCC 23330) and Branhamella
catarrhalis (ATCC 25239) were obtained from the American
4066
Vol. 60, No. 11
on Septem
ber 20, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

PCR FINGERPRINTING OF ACINETOBACTER SPP. 4067
Type Culture Collection, Rockville, Md., and Moraxella bovi(DSM 6328), Psychrobacter immobilis (DSM 7229), and Morax-ella nonliquefaciens (DSM 6327) were obtained from theDeutsche Sammlung von Mikroorganismen und ZellkulturenGmbH, Braunschweig, Germany.The two Moraxella phenylpyruvica strains (Moraxella phe-
nylpyruvica I and II) were kindly provided by K. Pelz, Institutfur Medizinische Mikrobiologie und Hygiene, Freiburg, Ger-many.The Acinetobacter strains were grown at 20°C in FPA
medium at a pH of 7.0. FPA medium contains (per liter) 0.1 gof peptone, 0.2 g of casein peptone, 0.1 g of yeast extract, 1.0g of sodium acetate, 1.0 ml of 50% lactate, 0.5 g of MgSO4, 0.5g of (NH4)2SO4, and 0.2 g of CaCl2. After autoclaving, 1 ml of0.01 M autoclaved phosphate buffer (K2HPO4-KH2PO4 [pH7.0]) was added.
All other strains were grown at 30°C on blood agar plates.PCR fingerprinting and oligonucleotide primers. Acineto-
bacter strains were classified by rapid methods based on PCR(22). For the purpose of species identification, a pair of tRNAgene-specific consensus primers was used (27).
In order to compare strains within Acinetobacter species, weused random amplified polymorphic DNA (RAPD) finger-printing with a single arbitrarily chosen primer (26, 29).The primers T3A (5'GGGGGTTCGAATTCCCGCCGGC
CCCA3') and T5B (5'AATGCTCTACCAACTGAACT3')used for tRNA consensus fingerprinting (27) and the primerDAF 4 (5'CGGCAGCGCC3') for the RAPD fingerprintswere custom synthesized by Pharmacia Biotech, Freiburg,Germany. All primers were 5' end labelled with fluorescein.PCR amplification. Reaction mixtures (50 RI) were pre-
pared with 1.25 U of Taq polymerase (Pharmacia Biotech), 1xPCR buffer (Pharmacia Biotech), 0.2 mM (each) dinucleosidetriphosphate (Pharmacia Biotech), 1 ,uM primer (or primers),and 2 ,u of crude cell lysates used as a source of templateDNA. To prepare a crude lysate, one bacterial colony wasresuspended in 50 ,ul of lysis buffer (1% Tween 20, lx PCRbuffer). The suspension was heated for 5 min at 95°C, and thencell debris was sedimented at 16,000 x g for 5 min in amicrocentrifuge. The reaction mixture was overlaid with lightmineral oil and cycled in a thermocycler (Landgraf Varius V45; Landgraf Laborgerate, Langenhagen, Germany) as follows.The temperature program for the tDNA PCR was 120 s at94°C; 45 cycles of 40 s at 94°C, 40 s at 50°C, and 120 s at 70°C;and 180 s at 70°C. The temperature program for the RAPDPCR was 120 s at 94°C, 45 cycles of 40 s at 94°C and 40 s at45°C, and 180 s at 70°C. The amplification products wereseparated by horizontal electrophoresis in 5% acrylamide-7 Murea-1 x Tris-borate-EDTA gels and visualized by silver stain-ing (3).ALF (Automated Laser Fluorescent) DNA sequencer. Am-
plification products were analyzed with the automated se-quencer ALF (Pharmacia Biotech). DNA fragments wereseparated by electrophoresis through 5% acrylamide-7 M ureain 0.6x Tris-borate-EDTA gels. For sample preparation, 2 p.lof the PCR products was mixed with 4 p.1 of stop solution (5 mgof blue dextran per ml in formamide) and incubated for 2 minat 95°C before being loaded onto the gel. The conditions ofseparation were 1,800 V, 45 mA, and 45 W for 240 min at 40°C.
RESULTS
PCR fingerprinting with tRNA consensus primers. tRNAgenes occur in multiple copies dispersed throughout the ge-nome and contain shared sequence motifs. The consequence isthat products resulting from a PCR with primers that contain
A1 2 3 4 5 6 7 8 9 10 11 12 13
I I vI Z
B1 2 3 4 5 6 7 8 9 1011
FIG. 1. tDNA PCR fingerprints of different Acinetobacter strains.PCR was performed with the primers T5B and T3A. (A) Lane 2,A. anitratus; lane 3, A. baumannii ATCC 1041T; lane 4, A. bauman-nii ATCC 15308; lane 5, A. baumannii ATCC 19606; lane 6, A. bau-mannii LMD 85.46; lane 7, A. baumannii LMD 82.33; lane 8, A.baumannii ATCC 23220; lane 9, A. baumannii ATCC 13809; lane 10,A. baumannii ATCC 15150; lane 11, A. calcoaceticus ATCC 23055;lane 12, A. haemolyticus ATCC 17906; lanes 1 and 13, size marker<>X174 restriction fragment (RF) DNA digested with HaeIII (1,358 bp,1,078 bp, 872 bp, 603 bp, 310 bp, 281 bp, 271 bp, and 234 bp). (B) Lane2, Acinetobacter sp. 3; lane 3, Acinetobacter sp. 6; lane 4, Acinetobactersp. 10; lane 5, Acinetobacter sp. 11; lane 6, Acinetobacter sp. 13; lane 7,Acinetobacter sp. 14; lane 8, Acinetobacter sp. 15; lane 9, Acinetobactersp. 16; lane 10, Acinetobacter sp. 17; lane 11, A. radioresistens; lane 1,size marker 4X174 RF DNA digested with HaeIII (see panel A).
consensus tRNA sequences (tDNA-PCR) are identical ornearly identical within a species (27). Fingerprints on thespecies level obtained by using the tRNA consensus primersT5B and T3A (27) showed different band patterns for all of the17 Acinetobacter species investigated (Fig. 1 and 2), except forthe band patterns ofA. johnsonii and of the Acinetobacter sp. 9strains. The banding patterns of the three Acinetobacter sp. 9strains were identical with the banding pattern ofA. johnsonii,thus suggesting their classification as a species very closelyrelated to A. johnsonii (Fig. 2B).
A1 2 3 4 5 6 7
B C
1 2 3 4 S 6 7
FE--vr .
!~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~
* 107j-1- _-:6
4~~~~~~t*
FIG. 2. tDNA PCR fingerprints of different Acinetobacter strainsand members of the genera very closely related to the genus Acineto-bacter. PCR was performed with the primers TSB and T3A. (A) Lane2, A. junii; lane 3, strain Fo-3; lane 4, A. johnsonii; lane 5, A. Iwoffli I;lane 6, A. iwoffli II; lane 7, A. Iwoffli H; lane 1, size marker +X174 RFDNA digested with HaeIII (1,358 bp, 1,078 bp, 872 bp, 603 bp, 310 bp,281 bp, 271 bp, and 234 bp). (B) Lane 2, strain Br-2; lane 3, Moraxellabovi; lane 4, Psychrobacter immobilis; lane 6, strain 210 A; lane 7, A.johnsonii; lane 8, strain Fo-3; lane 9, Acinetobacter sp. 9 M; lane 10,Acinetobacter sp. 9 B; lane 11, Acinetobacter sp. 9 A; lanes 1, 5, and 12,size marker +X174 RF DNA digested with HaeIII (see panel A). (C)Lane 2, Branhamella catarrhalis; lane 3, Kingella kingae; lane 4,Moraxella phenylpyruvica I; lane 5, Moraxella phenylpyruvica II; lane 7,Moraxella nonliquefaciens; lanes 1 and 6, size marker +X174 RF DNAdigested with HaeIII (see panel A).
VOL. 60, 1994
on Septem
ber 20, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from
4068 WIEDMANN-AL-AHMAD ET AL.
The pattern obtained with the strains A. baumannii ATCC13809 and A. baumannii ATCC 15150 was different from theother sevenA. baumannii patterns, but it was identical with thepattern of A. calcoaceticus, thus suggesting that these strainsshould be reclassified as A. calcoaceticus (Fig. 1A).The pattern obtained with strain I of A. iwoffli, classified by
the analysis of phenotypical properties, was different from theother two A. Iwoffli patterns. However, it was nearly identicalwith the pattern obtained from the A. junii type strain, thussuggesting that the strain is more closely related toA. junii thanto A. iwoffii (Fig. 2A).The analysis of banding patterns obtained by tRNA consen-
sus fingerprinting showed that the strain ofAcinetobacter sp. 16and the strain of Acinetobacter sp. 15 and also the strains ofAcinetobacter sp. 14 and Acinetobacter sp. 17 are very closelyrelated (Fig. 1B).
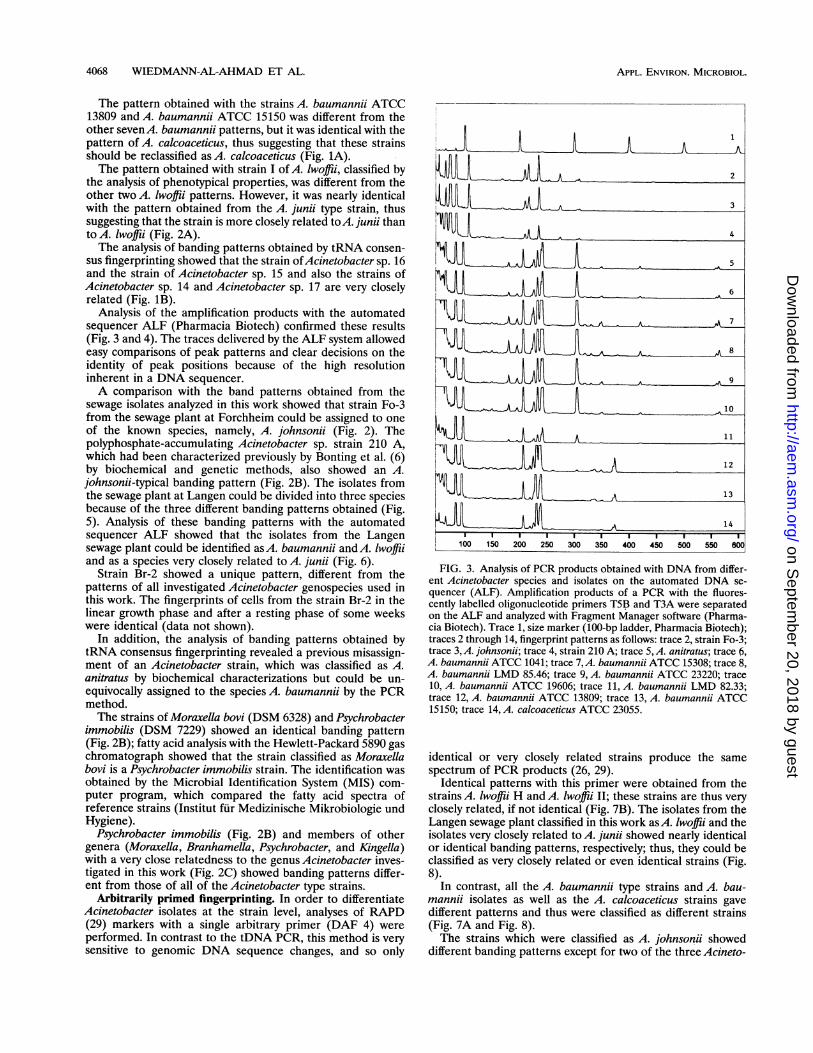
Analysis of the amplification products with the automatedsequencer ALF (Pharmacia Biotech) confirmed these results(Fig. 3 and 4). The traces delivered by the ALF system allowedeasy comparisons of peak patterns and clear decisions on theidentity of peak positions because of the high resolutioninherent in a DNA sequencer.A comparison with the band patterns obtained from the
sewage isolates analyzed in this work showed that strain Fo-3from the sewage plant at Forchheim could be assigned to oneof the known species, namely, A. johnsonii (Fig. 2). Thepolyphosphate-accumulating Acinetobacter sp. strain 210 A,which had been characterized previously by Bonting et al. (6)by biochemical and genetic methods, also showed an A.johnsonii-typical banding pattern (Fig. 2B). The isolates fromthe sewage plant at Langen could be divided into three speciesbecause of the three different banding patterns obtained (Fig.5). Analysis of these banding patterns with the automatedsequencer ALF showed that the isolates from the Langensewage plant could be identified as A. baumannii and A. Iwoffiiand as a species very closely related to A. junii (Fig. 6).
Strain Br-2 showed a unique pattern, different from thepatterns of all investigated Acinetobacter genospecies used inthis work. The fingerprints of cells from the strain Br-2 in thelinear growth phase and after a resting phase of some weekswere identical (data not shown).
In addition, the analysis of banding patterns obtained bytRNA consensus fingerprinting revealed a previous misassign-ment of an Acinetobacter strain, which was classified as A.anitratus by biochemical characterizations but could be un-equivocally assigned to the species A. baumannii by the PCRmethod.The strains of Moraxella bovi (DSM 6328) and Psychrobacter
immobilis (DSM 7229) showed an identical banding pattern(Fig. 2B); fatty acid analysis with the Hewlett-Packard 5890 gaschromatograph showed that the strain classified as Moraxellabovi is a Psychrobacter immobilis strain. The identification wasobtained by the Microbial Identification System (MIS) com-puter program, which compared the fatty acid spectra ofreference strains (Institut fuir Medizinische Mikrobiologie undHygiene).
Psychrobacter immobilis (Fig. 2B) and members of othergenera (Moraxella, Branhamella, Psychrobacter, and Kingella)with a very close relatedness to the genus Acinetobacter inves-tigated in this work (Fig. 2C) showed banding patterns differ-ent from those of all of the Acinetobacter type strains.
Arbitrarily primed fingerprinting. In order to differentiateAcinetobacter isolates at the strain level, analyses of RAPD(29) markers with a single arbitrary primer (DAF 4) wereperformed. In contrast to the tDNA PCR, this method is verysensitive to genomic DNA sequence changes, and so only
I 100 150 200 250 300 350 400 450 500 550
FIG. 3. Analysis of PCR products obtained with DNA from differ-ent Acinetobacter species and isolates on the automated DNA se-quencer (ALF). Amplification products of a PCR with the fluores-cently labelled oligonucleotide primers T5B and T3A were separatedon the ALF and analyzed with Fragment Manager software (Pharma-cia Biotech). Trace 1, size marker (100-bp ladder, Pharmacia Biotech);traces 2 through 14, fingerprint patterns as follows: trace 2, strain Fo-3;trace 3, A. johnsonii; trace 4, strain 210 A; trace 5, A. anitratus; trace 6,A. baumannii ATCC 1041; trace 7,A. baumannii ATCC 15308; trace 8,A. baumannii LMD 85.46; trace 9, A. baumannii ATCC 23220; trace10, A. baumannii ATCC 19606; trace 11, A. baumannii LMD 82.33;trace 12, A. baumannii ATCC 13809; trace 13, A. baumannii ATCC15150; trace 14, A. calcoaceticus ATCC 23055.
identical or very closely related strains produce the samespectrum of PCR products (26, 29).
Identical patterns with this primer were obtained from thestrains A. Iwoffli H and A. Iwoffli II; these strains are thus veryclosely related, if not identical (Fig. 7B). The isolates from theLangen sewage plant classified in this work asA. iwoffli and theisolates very closely related to A. junii showed nearly identicalor identical banding patterns, respectively; thus, they could beclassified as very closely related or even identicai strains (Fig.8).
In contrast, all the A. baumannii type strains and A. bau-mannii isolates as well as the A. calcoaceticus strains gavedifferent patterns and thus were classified as different strains(Fig. 7A and Fig. 8).The strains which were classified as A. johnsonii showed
different banding patterns except for two of the three Acineto-
APPL. ENVIRON. MICROBIOL.
on Septem
ber 20, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

PCR FINGERPRINTING OF ACINETOBACTER SPP. 4069
FIG. 4. Analysis of PCR products obtained with DNA from differ-ent Acinetobacter species on the automated DNA sequencer (ALF).Amplification products of a PCR with the fluorescently labelledoligonucleotide primers T5B and T3A were separated on the ALF andanalyzed with Fragment Manager software (Pharmacia Biotech).Trace 1, size marker (100-bp ladder, Pharmacia Biotech); trace 2,Acinetobacter sp. 14; trace 3, Acinetobacter sp. 17; trace 4, Acinetobactersp. 15; trace 5, Acinetobacter sp. 16.
bacter sp. 9 strains; Acinetobacter sp. 9 M and Acinetobacter sp.9 B are thus possibly identical or very closely related (Fig. 7B).
Again, the results obtained with the automated system wereconsistent with results obtained by silver staining of polyacryl-amide gels (data not shown), thus showing that fingerprint
M M M N N N M1 2 3 4 5 6 7 8 9 10 12 13 14 15 16 17 18 19 20 21 22 23 24
IIa44..o=*.
i~~~~~~~~~~~~~~;I:.I : +
FIG. 5. tDNA PCR fingerprints of Acinetobacter isolates from thewastewater treatment plant in Langen, Germany. PCR was performedwith the primers T5B and T3A. Lanes 1, 7, 13, 24, and 34 (=M), sizemarker 4XX174 RF DNA digested with HaeIII (1,358 bp, 1,078 bp, 872bp, 603 bp, 310 bp, 281 bp, 271 bp, 234 bp); lane 33 (=P), Escherichiacoli TG 1 (positive control); lanes 19, 22, and 23 (=N), negativecontrol; all other lanes, the banding patterns of the isolates La 1 to La25 from the sewage plant. The isolates show three different bandingpatterns (e.g., lane 2 = La 1, lane 3 = La 2, and lane 4 = La 3);consequently, the isolates can be divided into three species (see Fig. 6).
FIG. 6. PCR products analyzed on the automated DNA sequenzer(ALF). After a PCR with the primers T5B and T3A, amplificationproducts obtained from Acinetobacter isolates of the Langen wastewa-ter treatment plant (Germany) were separated and analyzed with theALF system. Trace 1, size marker (100-bp ladder, Pharmacia Biotech);traces 2 to 13, fingerprint patterns as follows: trace 2, A. junii; trace 3,La 2; trace 4, La 5; trace 5, La 10; trace 6,A. Iwoffii; trace 7, La 1; trace8, La 4; trace 9, La 20; trace 10,A. baumannii; trace 11, La 3; trace 12,La 7; trace 13, La 18.
analyses can be performed with the ALF sequencer, makinguse of its inherent benefits (e.g., the superior resolution andthe automated electronic storage of data).
DISCUSSION
In the past few years, it became clear that phenotypicdifferentiation is not sufficient for the exact determination ofspecies or strains of Acinetobacter and some other genera. Inmany cases, an exact taxonomic classification cannot be ob-tained even if complex determination systems are employed. Inparticular, difficulties arise with nonclinical isolates ofAcineto-bacter (23). If the PCR fingerprinting method is compared withthe classical methods, it becomes clear that not only does thisnew method allow a significant differentiation from relatedgenera (e.g., Moraxella, Branhamella, Psychrobacter, and Kin-gella) and an unequivocal taxonomic classification, but also it ismore convenient and can be carried out with much less cellmaterial. The results are obtained very fast compared withother nucleic acid techniques because it is not necessary to
VOL. 60, 1994
on Septem
ber 20, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

4070 WIEDMANN-AL-AHMAD ET AL.
1 2 3 4 5 6 7 8 9101112
_R:., - ow
- m
FIG. 7. RAPD fingerprinting with the primer DAF 4. Patternsobtained from different Acinetobacter species and isolates are shown.(A) Lane 2, A. anitratus; lane 3, A. baumannii ATCC 1041; lane 4, A.baumannii ATCC 15308; lane 5,A. baumannii ATCC 19606; lane 6,A.baumannii LMD 85.46; lane 7, A. baumannii LMD 82.33; lane 8, A.baumannii ATCC 23220; lane 9, A. baumannii ATCC 13809; lane 10,A. baumannii ATCC 15150; lane 11, A. calcoaceticus ATCC 23055;lanes 1 and 12, size marker 4X174 RF DNA digested with HaeIII(1,358 bp, 1,078 bp, 872 bp, 603 bp, 310 bp, 281 bp, 271 bp, 234 bp). (B)Lane 2, strain 210 A; lane 3, A. johnsonii; lane 4, strain Fo-3; lane 5,Acinetobacter sp. 9 M; lane 6, Acinetobacter sp. 9 B; lane 7, Acineto-bacter sp. 9 A; lane 8,A. Iwoffli II; lane 9,A. Iwoffli H; lane 10, Moraxellabovi; lane 11, Psychrobacter immobilis; lanes 1 and 12, size marker(X174 RF DNA digested with HaeIII (see panel A).
purify the DNA beforehand. Since it was found that identicalband patterns are obtained both after a simple cell lysis andwith DNA extracted from the cells, crude lysates were usedthroughout this study. The influence of growth stage of cul-tures was found to be negligible; the fingerprints of cells fromthe strain strain Br-2 in the linear growth phase and after aresting phase were identical. Minor differences in band inten-sities were sometimes seen on silver-stained gels, suggestingdifferent patterns (e.g., Fig. 1, lanes 3 and 4). However, theALF traces demonstrated their high similarity (e.g., Fig. 3,traces 6 and 7).The PCR fingerprinting method, which has been established
in recent years as a universal method for classifying a numberof bacterial species and other organisms (19, 26, 28, 29), wasfound to be very reliable. Prerequisites for reproducible results
FIG. 8. RAPD fingerprinting with the primer DAF 4. Patternswere obtained from Acinetobacter isolates from the wastewater treat-ment plant in Langen, Germany. Lanes 2 to 9, isolates which wereidentified as A. baumannii after tDNA fingerprinting; lanes 10 to 18,isolates very closely related to the species A. junii; lanes 19 to 26,isolates which were identified as A. Iwoffli; lanes 1 and 27, size marker4X174 RF DNA digested with HaeIII (1,358 bp, 1,078 bp, 872 bp, 603bp, 310 bp, 281 bp, 271 bp, 234 bp).
are, in our experience, however, completely identical reactionconditions, e.g., Mg2+, primer, and enzyme concentrations aswell as the temperature and number of PCR cycles.A comparison of Acinetobacter type strains with isolates
from wastewater treatment plants revealed that only 2 of 28strains could be classified as A. johnsonii. These two displayeddifferent (strain-specific) RAPD patterns. Interestingly, bothstrains were capable of storing large amounts of polyphos-phate. In other investigations of Acinetobacter strains fromtreatment plants with biological phosphate removal, polyphos-phate-accumulating strains of A. johnsonii (genospecies 7)were isolated (4, 5, 11, 17, 23). However, other polyphosphate-storing Acinetobacter strains, e.g., Br-2, could not be classifiedby their fingerprint patterns as one of the previously describedgenospecies. It is possible that A. johnsonii is the dominantspecies under certain conditions in wastewater treatmentplants with biological phosphorus removal. On the other hand,our results with the isolates La 1 to La 25 and Br-2 show thatAcinetobacter species other than A. johnsonii can also occur ina very large population: A. Iwoffii, strains very closely related tothe species A. junii, and, interestingly, also A. baumannii.Normally, A. baumannii strains have been found to be afrequent source of hospital-acquired infections (12).
Because of the difficulties encountered in these investiga-tions and in investigations of other authors using genetic traitsto obtain a complete taxonomic classification of nonclinicalisolates, the species division of the genus Acinetobacter, espe-cially of strains from nonclinical biotopes, is still provisional,and this genus probably requires further species division. Theprecise characterization of a large number of Acinetobacterisolates by PCR fingerprinting should improve our knowledgeof the ecology of the different Acinetobacter species.
ACKNOWLEDGMENTSWe thank B. Jager for excellent technical assistance; P. J. M. Bouvet,
J. W. van Groeenstijn, A. Schonborn, P. Kampfer, G. Auling, andN. Jardin for delivery of some Acinetobacter strains; and K. Pelz forsupplying Moraxella strains.
This work was supported in part by a grant to G. Schon from theBMFT (Bundesministerium fur Forschung und Technologie, contractno. 02 WA 91350).
REFERENCES1. Alexander, M., F. Ismail, P. J. H. Jackman, and W. C. Nobel. 1984.
Fingerprinting Acinetobacter strains from clinical sources by nu-merical analysis of electrophoretic patterns. J. Med. Microbiol.18:55-68.
2. Auling, G., F. Pilz, H.-J. Busse, S. Karrasch, M. Streichan, and G.Schon. 1991. Analysis of the polyphosphate-accumulating micro-flora in phosphorus-eliminating, anaerobic-aerobic activatedsludge systems by using diaminopropane as a biomarker for rapidestimation of Acinetobacter spp. Appl. Environ. Microbiol. 57:3585-3592.
3. Bassam, B. J., G. Caetano-Anolles, and P. M. Gresshoff. 1991. Fastand sensitive silver staining of DNA in polyacrylamide gels. Anal.Biochem. 196:80-83.
4. Bayly, R. C., A. Duncan, J. W. May, M. Schermbri, A. Smertjis, G.Vasiliadis, and W. G. C. Raper. 1991. Microbiological and geneticaspects of the synthesis of polyphosphate by species of Acineto-bacter. Water Sci. Technol. 23:747-757.
5. Beacham, A. M., R. J. Seviour, K. C. Lindrea, and I. Livingston.1990. Genospecies diversity of Acinetobacter isolates obtainedfrom a biological nutrient removal pilot plant of a modified UCTconfiguration. Water Res. 24:23-29.
6. Bonting, C. F. C., B. M. F. Willemsen, W. Akkermans-van Vliet,P. J. M. Bouvet, G. J. J. Kortsee, and A. J. B. Zehnder. 1992.Additional characteristics of the polyphosphate-accumulatingAcinetobacter strain 210 A and its identification as Acinetobacterjohnsonii. FEMS Microbiol. Ecol. 102:57-64.
A B1 2 3 4 5 6 7 8 9101112
--_-.- -.-w!m4,
APPL. ENvIRON. MICROBIOL.
on Septem
ber 20, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

PCR FINGERPRINTING OF ACINETOBACTER SPP. 4071
7. Bouvet, P. J. M., and P. A. D. Grimont. 1986. Taxonomy of thegenusAcinetobacter with the recognition ofAcinetobacter bauman-nii sp. nov., Acinetobacter haemolyticus sp. nov., Acinetobacterjohnsonii sp. nov., and Acinetobacter junii sp. nov. and emendeddescriptions of Acinetobacter calcoaceticus and AcinetobacterIwoffli. Int. J. Syst. Bacteriol. 36:228-240.
8. Bouvet, P. J. M., and P. A. D. Grimont. 1987. Identification andbiotyping of clinical isolates of Acinetobacter. Ann. Inst. Pasteur(Microbiol.) 138:569-578.
9. Bouvet, P. J. M., and S. Jeanjean. 1989. Delineation of newproteolytic genomic species in the genus Acinetobacter. Res.Microbiol. 140:291-299.
10. Brisou, J., and A. R. Prevot. 1954. Etudes des systematiquebacterienne. X. Revision des especes reunies dans le genreAchromobacter. Ann. Inst. Pasteur 86:722-728.
11. Duncan, A., G. E. Vasiliadis, R. C. Bayly, and J. W. May. 1988.Genospecies ofAcinetobacter isolated from activated sludge show-ing enhanced removal of phosphate during pilot-scale treatment ofsewage. Biotechnol. Lett. 10:831-836.
12. Graser, Y., L. Klare, E. Halle, R. Gantenberg, P. Buchholz, H. D.Jacobi, W. Presber, and G. Schonian. 1993. Epidemiological studyof an Acinetobacter baumannii outbreak by using the polymerasechain reaction fingerprinting. J. Clin. Microbiol. 31:2417-2420.
13. Henriksen, S. D. 1973. Moraxella, Acinetobacter, and the Mimeae.Bacteriol. Rev. 37:522-561.
14. Henriksen, S. D. 1976. Moraxella, Neisseria, Branhamella, andAcinetobacter. Annu. Rev. Microbiol. 30:63-83.
15. Hoffmann, S., C. E. Mabeck, and R. Vejsgaard. 1982. Bacteriuriacaused by Acinetobacter calcoaceticus biovars in a normal popula-tion and in general practice. J. Clin. Microbiol. 16:443-451.
16. Ino, T., and Y. Nishimura. 1989. Taxonomic studies of Acineto-bacter species based on outer membrane protein patterns. J. Gen.Appl. Microbiol. 35:213-224.
17. Kampfer, P., K. Bark, H.-J. Busse, G. Auling, and W. Dott. 1992.Numerical and chemotaxonomy of polyphosphate-accumulatingAcinetobacter strains with high polyphosphatic:AMP phospho-transferase (PPAT) activity. Syst. Appl. Microbiol. 15:409-419.
18. Karrasch, S., and G. Schon. 1991. Enhanced phosphate uptake by
activated sludge under anoxic growth conditions. Bioforum 14:94.19. Martin, G. B., J. G. K. Williams, and S. D. Tanksley. 1991. Rapid
identification of markers linked to a Pseudomonas resistance genein tomato by using random primers and near-isogenic lines. Proc.Natl. Acad. Sci. USA 88:2336-2340.
20. Pines, O., and D. Gutnick. 1986. Role for emulsan in growth ofAcinetobacter calcoaceticus RAG-1 on crude oil. Appl. Environ.Microbiol. 51:661-663.
21. Rosenberg, E., C. Rubinovitz, A. Gottlieb, S. Rosenhak, and E. Z.Ron. 1988. Production of biodispersan by Acinetobacter calcoace-ticus A2. Appl. Environ. Microbiol. 54:317-322.
22. Saiki, R. K., S. J. Scharf, F. Faloona, K. B. Mullis, G. T. Horn,H. A. Erlich, and N. Arnheim. 1985. Enzymatic amplification ofbeta-globin genomic sequences and restriction site analysis fordiagnosis of sickle cell anemia. Science 230:1350-1354.
23. Soddell, J. A., A. M. Beacham, and R. J. Seviour. 1993. Phenotypicidentification of non-clinical isolates of Acinetobacter species. J.Appl. Microbiol. 74:210-214.
24. Streichan, M., G. Schon, and J. R. Golecki. 1990. Polyphosphate-accumulating bacteria from sewage plants with different processesfor biological phosphorus removal. FEMS Microbiol. Ecol. 73:113-124.
25. Tjernberg, I., and J. Ursing. 1989. Clinical strains ofAcinetobacterclassified by DNA-DNA hybridisation. Acta Pathol. Microbiol.Immunol. Scand. 97:595-605.
26. Welsh, J., and M. McClelland. 1990. Fingerprinting genomes usingPCR with arbitrary primers. Nucleic Acids Res. 18:7213-7218.
27. Welsh, J., and M. McClelland. 1991. Genomic fingerprints pro-duced by PCR with consensus tRNA gene primers. Nucleic AcidsRes. 19:861-866.
28. Welsh, J., C. Peterson, and M. McClelland. 1991. Polymorphismsgenerated by arbitrarily primed PCR in the mouse: application tostrain identification and genetic mapping. Nucleic Acids Res.20:303-306.
29. Williams, J. G. K., A. R. Kubelik, K. J. Livak, J. A. Rafalski, andS. V. Tingey. 1990. DNA polymorphisms amplified by arbitraryprimers are useful as genetic markers. Nucleic Acids Res. 18:6531-6535.
VOL. 60, 1994
on Septem
ber 20, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from





























