Embed Size (px)
Citation preview

Aus dem Pathologischen Institut
der Friedrich-Alexander-Universität Erlangen-Nürnberg
Direktor: Prof. Dr. med. Arndt Hartmann
Charakterisierung des kardiovaskulären und renalen Phänotyps von BK β1 knockout Mäusen
Inaugural-Dissertation
zur Erlangung der Doktorwürde
der
Medizinischen Fakultät
der
Friedrich–Alexander-Universität
Erlangen-Nürnberg
vorgelegt von
Matthias Nußelt
aus
Dinkelsbühl

Gedruckt mit Erlaubnis der Medizinischen Fakultät der Friedrich-Alexander-Universität
Erlangen-Nürnberg
Dekan: Prof. Dr. med. Dr. h.c. J. Schüttler
Referent: Prof. Dr. med. K. Amann
Korreferent: Prof. Dr. med. K.F. Hilgers
Tag der mündlichen Prüfung: 06. Oktober 2010

Meinen Eltern in Dankbarkeit gewidmet.

4
Inhaltsverzeichnis1 Zusammenfassung ...............................................................................7
1.1 Hintergrund und Ziele..........................................................................7
1.2 Methoden...............................................................................................7
1.3 Ergebnisse und Beobachtungen...........................................................7
1.4 Schlussfolgerungen...............................................................................8
2. Summary..............................................................................................82.1 Background...........................................................................................8
2.2 Methods.................................................................................................9
2.3 Results....................................................................................................9
2.4 Conclusions............................................................................................9
3 Einleitung............................................................................................10
4 Material und Methodik......................................................................144.1 Versuchstiere ......................................................................................14
4.2 Haltung und Fütterung.......................................................................15
4.3 Perfusionsfixation ..............................................................................15
4.4 Gewebeaufarbeitung ..........................................................................16
4.5 Technik der Paraffinschnitte.............................................................164.5.1 Paraffinschnitte Niere...............................................................................16
4.5.2 Paraffinschnitte Herz und Aorta...............................................................17
4.6 Analyse.................................................................................................174.6.1 Analyse der Herzen und der Aorta...........................................................17
4.6.1.1 Analyse der Herzen....................................................................................17
4.6.1.2 Analyse der Aorten.....................................................................................18
4.6.2 Analyse der Nieren...................................................................................19
4.6.2.1 Schädigungsindices ...................................................................................19
4.6.2.1.1 Glomeruloskleroseindex (GSI)................................................................19
4.6.2.1.2 Tubulointerstitieller Index (TSI)..............................................................20
4.6.2.1.3 Vaskulärer Index (VSI)...........................................................................20
4.6.2.2 Glomerulusgeometrie ................................................................................21
4.6.2.2.1 Morphometrie und Stereologie................................................................21
4.6.2.2.2 Auswertung.............................................................................................22

5
4.6.2.2.2.1 Volumenfraktion von Nierenmark und Nierenrinde.............................23
4.6.2.2.2.2 Gesamtvolumen Niere und absolutes Volumen Nierenrinde ...............23
4.6.2.2.2.3 Glomeruli pro Fläche Rinde.................................................................24
4.6.2.2.2.4 Volumendichte der Glomeruli..............................................................24
4.6.2.2.2.5 Numerische Dichte der Glomeruli in der Rinde....................................25
4.6.2.2.2.6 Gesamtzahl und Gesamtvolumen der Glomeruli einer Niere................25
4.6.2.2.2.7 Mittleres glomeruläres Volumen .........................................................26
4.7 Statistik................................................................................................26
5 Ergebnisse...........................................................................................275.1 Allgemeine Parameter........................................................................27
5.1.1 Körpergewicht .........................................................................................27
5.1.2 Herzgewicht (HG) und relatives Herzgewicht (rHG)..............................27
5.1.3 Nierengewicht (NG) und relatives mittleres Nierengewicht (rmNG) .....28
5.2 Morphometrie.....................................................................................295.2.1 Morphometrie Herz – Wanddicken .........................................................29
rechter Ventrikel (RV), linker Ventrikel (LV) und Septum..............................29
5.2.2 Morphometrie intramyokardialer Arterien (IMA) ..................................30
5.2.3 Morphometrie Aorta - Wanddicke...........................................................30
5.3 Niere.....................................................................................................315.3.1 Semiquantitative Untersuchungen – renale Schädigungsindices.............31
5.3.2 Prozentuale Verteilung Nierenrinde und Nierenmark .............................32
5.3.3 Gesamtvolumen Niere (VN) und absolutes Volumen der Nierenrinde
(VR)...................................................................................................................32
5.3.4 Anzahl der Glomeruli pro Volumen (Nv)................................................33
5.3.5 Anzahl der Glomeruli einer Niere (NGlom)............................................33
5.3.6 Mittleres glomeruläres Volumen (VGlom ).............................................34
5.4 Übersicht der Parameter und Vergleiche zwischen den
untersuchten Gruppen.............................................................................345.4.1 Kardiovaskulärer Phänotyp......................................................................35
5.4.2 Renaler Phänotyp......................................................................................36
6 Diskussion...........................................................................................376.1 Interpretation des Phänotyps.............................................................39
6.1.1 Knockout vs. wildtyp................................................................................39

6
6.1.1.1 Kardialer Phänotyp.....................................................................................39
6.1.1.2 Vaskulärer Phänotyp..................................................................................40
6.1.1.3 Renaler Phänotyp.......................................................................................40
6.1.2 Junge vs. alte Tiere...................................................................................42
6.1.2.1 Kardialer Phänotyp.....................................................................................42
6.1.2.2 Vaskulärer Phänotyp..................................................................................43
6.1.2.3 Renaler Phänotyp.......................................................................................43
6.1.3 Alte Tiere gleichen Genotyps männlich vs. weiblich...............................44
6.1.3.1 Kardiovaskulärer Phänotyp........................................................................45
6.1.3.2. Renaler Phänotyp......................................................................................45
6.1.4 Schlussfolgerungen kardiovaskulärer Phänotyp.......................................46
6.1.5 Schlussfolgerungen renaler Phänotyp......................................................46
6.2 Zusammenfassung..............................................................................47
7 Literatur..............................................................................................48
8 Abkürzungsverzeichnis......................................................................53
9 Danksagung.........................................................................................55
10 Lebenslauf.........................................................................................56

7
1 Zusammenfassung
1.1 Hintergrund und Ziele
BK Kanäle regulieren das Membranpotential glatter Muskelzellen und spielen bei der
Regulation des Gefäßtonus eine wichtige Rolle. Die Gendeletion der β1-Untereinheit
des BK Kanals könnte zu erhöhtem Blutdruck und der Entstehung einer chronischen
Hypertonie von BK β1 knockout Mäusen führen. Ziel dieser Arbeit war es, den
kardiovaskulären und renalen Phänotyp von BK β1 knockout und wildtyp Mäusen
unterschiedlichen Alters zu untersuchen. Wir überprüften die Hypothese, dass die
Deletion der β1-Untereinheit des BK Kanals in der Maus zu einem charakteristischen
kardiovaskulären und oder renalen Phänotyp führt, der auf die fehlende Expression der
BK β1-Untereinheit während der Embryonalentwicklung zurückzuführen ist.
1.2 Methoden
Die Effekte der Deletion der BK β1-Untereinheit bei Mäusen wurde anhand des BK β1
Tiermodells studiert. Es wurden 6 Gruppen von Tieren untersucht, die sich jeweils aus
Tieren verschiedenen Alters, Geschlechts und Genotyps zusammensetzten. Nach
retrograder Perfusionsfixation wurden den Versuchstieren das Herz, die Aorta sowie
beide Nieren entnommen. Neben Organgewichten, morphometrischen Daten von Herz,
intramyokardialen Arterien und Aorta wurden spezielle renale Schädigungsindices
bestimmt. Durch stereologische Methoden war es zusätzlich möglich, aus
histologischen Schnitten der Nieren Informationen über deren dreidimensionale
Struktur zu gewinnen.
1.3 Ergebnisse und Beobachtungen
Signifikante Genotyp abhängige frühe Veränderungen mehrerer Parameter konnten wir
nicht nachweisen. Entgegen unseren Erwartungen konnten wir auch keine signifikante
Veränderung des kardialen Phänotyps im Sinne einer Herzhypertrophie nachweisen, wie
dies bereits für dieses Tiermodell beschrieben wurde. Der vaskuläre Phänotyp der Aorta
zeigte - hier wie erwartet - signifikante zeitabhängige Veränderungen bei den knockout
Tieren. Unsere Beobachtungen sprechen gegen einen deutlichen Einfluss der

8
Gendeletion auf den renalen Phänotyp und für eine eher zeitabhängige Veränderung
renaler Parameter. So können wir zum einen eine späte Schädigung männlicher
knockout Tiere beschreiben, zum anderen aber eine späte geschlechtsabhängige stärkere
Schädigung männlicher Tiere beiden Genotyps belegen, die den Einfluss der
Gendeletion wieder in Frage stellt und auch hier eine zeitabhängige Veränderung
vermuten lässt.
1.4 Schlussfolgerungen
Zusammenfassend sprechen unsere Ergebnisse gegen bereits früh bestehende,
signifikante kardiovaskuläre und renale Veränderungen bei BK β1 knockout Tieren und
damit gegen unsere anfangs formulierte Hypothese einer frühen embryonalen
Schädigung durch Gendeletion. Wir können lediglich für den vaskulären Phänotyp
späte, zeitabhängige Veränderungen bei den knockout Tieren beschreiben. Einen
sicheren Einfluss des Geschlechts auf den Phänotyp können wir mit unseren
Ergebnissen nicht belegen. Inwieweit geschlechtsabhängige Veränderungen des
Phänotyps stattfinden, sollte durch Einbeziehen weiblicher Tiere unterschiedlichen
Alters und Genotyps in folgenden Arbeiten genauer geprüft werden.
2. Summary
2.1 Background
BK channels regulate the membrane potential of arterial smooth muscle cells and play
an important role in regulating vascular resistance. Disruption of the gene encoding the
BK β1-subunit in mice may cause an elevation in blood pressure and lead to chronic
hypertension. This study aimed to determine the cardiovascular and renal phenotype in
mice of different age and BK β1 genotype. Our hypothesis was that the deletion of the
BK β1-subunit in mice leads to a characteristic cardiovascular and or renal phenotype,
caused by the missing expression of BK β1-subunits during embryonic development.

9
2.2 Methods
We used BK β1 knockout and control mice of both sex and different age.
We examined the heart, intramyocardial arteries, aortic rings and kidneys of each
animal. Body and organ weights, morphometric data of heart, intramyocardial arteries
und aorta were measured. Special renal indices were evaluated and stereologic methods
were used to obtain quantitative information about the renal three-dimensional structure.
2.3 Results
We could not verify significant early genotype-dependent variation of several
parameters. Contrary to expectations we also could not detect significant changes in
cardial phenotype in terms of cardiac hypertrophy, as previously described. Expectably,
the vascular phenotype of knockout animals showed significant time-dependent
changes. Our observations suggest that the gene deletion shows no clear effect on the
renal phenotype and we propose that changes in renal parameters are most likely time-
dependent. We can show both late impairment in male knockout animals and late
gender-dependent higher impairment in male animals of both genotype. This questions
the effect of gene deletion and argues for a time-dependent change as well.
2.4 Conclusions
In summary these results indicate that there are no early significant changes in
cardiovascular and renal phenotype in BK β1 knockout animals. This argues against our
primary hypothesis of an early embryonic impairment based on gene deletion.
Exclusively the vascular phenotype shows clearly time-dependent late changes. Our
results support the concept that there is no significant gender impact to the phenotype.
Further studies should determine the nature of gender-dependent changes in phenotype
by examining both male and female animals of different age and genotype.

10
3 Einleitung
Die arterielle Hypertonie ist eine häufig vorkommende, chronische Dysregulation des
arteriellen Blutdrucks und stellt einen wesentlichen Risikofaktor bei der Entstehung von
kardiovaskulären Erkrankungen wie Apoplex, Herzinfarkt und renalen Dysfunktionen
dar.[5] Die simple Frage, was genau zu einer pathologischen Erhöhung der
Blutdruckwerte führt, ist nur sehr schwer zu beantworten. In mehr als 90% aller Fälle
spricht man von einer essentiellen Hypertonie, deren Ursache man nicht kennt.[11]
Faktoren wie Alter, Geschlecht, Body-Mass-Index und verschiedene Diät-Varianten
wurden schon früh als Determinanten einer Blutdruckerhöhung erkannt und untersucht
[17], aber auch der genetische Einfluss auf den Blutdruck ist seit Jahrzehnten bekannt.
So belegen Zwillingsuntersuchungen aus den 70er Jahren bereits eindeutig einen
genetische Einfluss.[2,10] Bei der Entstehung einer Hypertonie sind ursächlich meist
eine Vielzahl an genetischen Faktoren sowie multiple Umwelteinflüsse und deren
Interaktionen zu nennen.[32] Nach Marteau et al. [20] tragen genetische Ursachen zur
Variation des Blutdrucks zu etwa 30 – 50 % bei. Physiologische Systeme, deren
genetische Variation bei der Regulation des Blutdrucks eine Rolle spielen, sind
mannigfaltig. Herzzeitvolumen und peripherer Gefäßwiderstand, Barorezeptoren,
natriuretische Peptide, das Renin-Angiotensin-Aldosteron-System, das Kinin-
Kallikrein-System, Adrenorezeptoren sowie Botenstoffe des Endothels wie NO oder
Endothelin wurden neben vielen anderen Variablen einzeln untersucht.[17] Da der
Blutruck aber nur im lebenden Organismus - in dem all diese Systeme komplex
interagieren - gemessen werden kann, gestaltet es sich schwierig, einzelne
Genvariationen dieser Systeme und deren Einfluss auf den Langzeitblutdruck
nachzuvollziehen.[21]
Lifton et al. [17] stellen 2001 die These auf, dass die bekannten genetischen Ursachen
einer Hypertonie immer in Änderungen der renalen Regulation des Salzhaushalts
münden und benennen die Niere als wichtigstes Organ bei der Entstehung einer
Hypertonie:
„Given the diversity of physiologic systems that can influence blood pressure, it is
striking that all Mendelian forms of hypertension and hypotension solved to date
converge on a final common pathway, altering blood pressure by changing net renal salt
balance. These findings clearly establish the role of altered salt homeostasis in blood

11
pressure variation in humans and underscore the key role of the kidney in the longterm
determination of blood pressure.“
Crowley et al. [6] konnten jedoch durch Kreuztransplantationen an Mäusen zeigen, dass
- unabhängig von der Nierenfunkion – nicht renale Gewebe, darunter vor allem die
glatte Gefäßmuskulatur (vascular smooth muscle cells; VSMC), einen bedeutenden
Einfluss auf die Regulation des Blutdrucks haben. Mendelsohn [21] formuliert
daraufhin die Hypothese, dass allein Veränderungen der VSMC langfristig direkt zu der
Entstehung einer Hypertonie führen können.
Mittlerweile gibt es mehrere Modelle transgener Tiere, bei denen die Deletion eines
Gens, das die Regulation des Gefäßwiderstands beeinflusst, langfristig zu einer
chronischen Erhöhung des Blutdrucks dieser Tiere führt.[21]
So beschreiben Brenner et al. [4] bereits 2000 das BK β1 Tiermodell, bei dem ein
Defekt des BK Kanals der Zellmembran glatter Gefäßmuskelzellen eine
Regulationsstörung der VSMCs verursacht und auch bei diesen Tieren zu einer
chronischen Hypertonie führt. Ein wesentlicher Schritt in der Entstehung der
Hypertonie ist dabei die Erhöhung des periphereren Gefäßwiderstands über einen
gesteigerten Gefäßtonus. Ein Einstrom von Kalziumionen über spannungsabhängige L-
Typ Kalziumkanäle (VDCC) der Zellmembran führt in VSMC zu einem globalen
intrazellulären Anstieg der Kalziumkonzentration mit Membrandepolarisation und
daraus resultierender Muskelzell- und Vasokonstriktion. Der globale Ca2 + Influx führt
weiterhin zu einem lokalen Kalzium-Efflux aus intrazellulären Speichern, so auch aus
dem sarkoplasmatischen Retikulum (SR).[23] Dieser Kalzium vermittelte
Kalziumstrom geschieht lokal an der Membran des SR via Ryanodin Rezeptoren (RyR).
Die entstehenden Kalziumströme werden Kalzium-Sparks genannt, welche nahe
gelegene BK Kanäle der Zellmembran aktivieren. Dies führt zu einem Kalium Efflux
am BK Kanal – sogenannte „spontaneous transient outward K+ currents“, STOCS
genannt - mit folgender Hyperpolarisation der Zellmemban und Muskelzell- und
Vasodilatation (Grafik 1). Der arterielle Gefäßtonus ist somit das Ergebnis zweier
entgegen wirkender kalziumabhängiger Prozesse.[4,17] Diese Ca2+ gesteuerte negative
Rückkoppelung via Sparks und STOCS scheint sehr wichtig für die Regulation des
Blutdrucks zu sein, denn wird der BK-Kanal durch Pharmaka blockiert oder mittels
Gendeletion in seiner Funktion gestört, so kommt es zu einer vermehrten Depolarisation
und damit zu einer gesteigerten Gefäßkontraktion in vivo. [3,16]

12
Grafik 1: Schematische Darstellung der Kalzium gesteuerten negativen RückkoppelungBK: BK Kanal; VDCC: spannungsabhängiger L-Typ Kalziumkanal; RyR: Ryanodin Rezeptor; SR: sarkoplasmatischen Retikulum; Erläuterung im Text.
BK Kanäle sind heterodimere Proteine, welche aus porenformenden α- Untereinheiten
und zusätzlichen β-Untereinheiten bestehen. Jede der bisher entdeckten β-
Untereinheiten ist gewebespezifisch und reguliert bestimmte Eigenschaften der BK
Kanäle. BK β1 knockout Mäuse zeigen eine reduzierte Kalziumsensitivität der BK
Kanäle, eine gestörte Spark/STOCS Koppelung bei erhaltenen Sparks, einen erhöhten
arteriellen Gefäßtonus und eine milde chronische Hypertonie im Vergleich zu wildtyp
Tieren.[4,26]
Das BK β1 knockout Tiermodell ist insofern sehr interessant, als die β1-Untereinheit,
deren Fehlen für die chronische Erhöhung des Blutdrucks verantwortlich zu sein
scheint, bis 2004 nur in der Zellmembran der VSMC nachweisbar war.[23,31] Nach
Mendelsohn [21] ist deshalb das BK β1 Tiermodell ein mögliches Beispiel einer allein
durch Veränderungen der VSMC begründeten, genetischen Hypertonieform.
Allerdings wurden bis heute keine ausgiebigen histologischen Untersuchungen von
kardiovaskulären und oder renalen Zellen und Geweben an BK β1 knockout Tieren
durchgeführt. So könnte eine Deletion der β1-Untereinheit des BK Kanals schon
während der Embryonalentwicklung der knockout Tiere zu Organschäden führen, die

13
dann ihrerseits – unabhängig vom Gefäßtonus - die Entstehung einer Hypertonie
verursachten.
In der hier vorgelegten Arbeit soll die Hypothese geprüft werden, dass die Deletion der
β1-Untereinheit des BK Kanals in der Maus zu einem charakteristischen
kardiovaskulären und oder renalen Phänotyp führt, der auf die fehlende Expression der
BK β1-Untereinheit während der Embryonalentwicklung zurückzuführen ist.
Zur Überprüfung der Hypothese wurden folgende Fragestellungen formuliert:
1. Finden sich Veränderungen an Herz, Aorta und Niere bei homozygoten BK β1
knockout Mäusen im Vergleich zu wildtyp Tieren ?
2. Gibt es Unterschiede im Zeitverlauf ?
3. Finden sich geschlechtsspezifische Unterschiede ?
Ein Vergleich von jungen männlichen Tieren sollte zunächst einmal die Beurteilung von
eventuell früh bestehenden Organschädigungen ermöglichen.
Der Vergleich von jungen männlichen und alten Tieren beiden Geschlechts sollte dann
die Möglichkeit bieten, die Entwicklung im Zeitverlauf geschlechtsspezifisch zu
beurteilen.

14
4 Material und Methodik
4.1 Versuchstiere
Will man die Hypothese prüfen, ob ein Signalmolekül wie der BK Kanal eine wichtige
Rolle in einem biologischen Prozess spielt, so kann man das Gen, welches für die
Expression dieser Struktur verantwortlich ist, eliminieren und daraufhin die
funktionellen Konsequenzen evaluieren. Dieses „gene targeting“ wurde in den 90er
Jahren zu einer weit verbreiteten Methode bei der Entwicklung von knockout Tieren.
[25] Mittels eigens dafür gezüchteten knockout Tieren erforschten mehrere Gruppen die
Auswirkungen einer fehlenden BK β1-Untereinheit in vivo bei Mäusen. Auch Plüger et
al. [26] generierten via „gen targeting“ transgene BK β1 knockout Mäuse, wobei sie 129
(R1) embryonale Stammzellen verwendeten und deren DNS nach erfolgreicher Deletion
der gewünschten Allele in Blastozyten von C57BL/6J Mäusen implantierten. Diese
Zellen wurden schließlich CBAxC57BL/6J Weibchen eingesetzt. Weibliche und
männliche Nachkommen unterschiedlichen Genotyps wurden so gepaart, das
homozygote knockout und wildtyp Tiere entstanden. Zur Kontrolle des Genotyps
wurden nach Terminierung der Tiere jeweils nochmals Southern Blot Analysen der
DNS aus Mäuseschwänzen nach gängigen Standardprotokollen durchgeführt.[26]
Als Versuchstiere dienten uns Tiere dieses BK β1 Stamms aus der Versuchstierhaltung
des Universitätsklinikum Hamburg-Eppendorf (Prof. Pongs, Prof. Ehmke).
Es wurden 6 Gruppen von Tieren untersucht, die sich jeweils aus Tieren verschiedenen
Alters, Geschlechts und Gentyps zusammensetzten. In den Gruppen 1 und 2 wurden
junge männliche, in den Gruppen 3 und 4 alte männliche und in den Gruppen 5 und 6
alte weibliche Tiere untersucht.
Gruppe Geschlecht Genotyp Anzahl Tiere Alter in Wochen1 ko 7 102 wt 7 73 ko 7 68,9 ± 9,54 wt 6 73,2 ± 10,75 ko 7 88 ± 15,46 wt 5 96,6 ± 20,8
♂ jung♂ jung♂ alt♂ alt♀ alt♀ alt

15
4.2 Haltung und Fütterung
Die Tiere aller sechs Gruppen wurden unter identischen Bedingungen gehalten. Alle
Tiere waren im selben Raum in mehreren Käfigen untergebracht. In diesem Raum
herrschte eine kontrollierte Umgebung mit konstanter Raumtemperatur von 22 ° Celsius
und einer relativen Luftfeuchtigkeit von 55%. Durch künstliches Licht wurde zwischen
6:00 Uhr und 18:00 Uhr ein Tag-Nachtrhythmus aufrecht erhalten.
Über den Versuchszeitraum hatten alle Tiere freien Zugang zu Trinkwasser und
Standardfutter Altromin C 1324 (Fa. Altromin, Lange/Lippe, Deutschland) mit 20%
Protein und 0,9% Ca2 +-Gehalt.
4.3 Perfusionsfixation
Das Experiment wurde durch retrograde Perfusionsfixation beendet. Dabei wurden die
Tiere zunächst mit Hilfe einer leichten Äthernarkose narkotisiert. Anschließend wurden
sie durch die intraperitoneale Applikation von 0,25 ml einer Stammlösung
(Zusammensetzung: 0,02 ml Ketaminhydrochlorid, 0,08 ml Medetomidinhydrochlorid
und 0,9 ml PBS) narkotisiert. Nach der Gewichtsbestimmung der Tiere wurden sie nach
erreichen einer tiefen Narkose mittels vier Kanülen auf einer Styroporplatte an den
Extremitäten in Rückenlage fixiert. Im Anschluss erfolgte eine mediane Laparotomie
mittels Scherenschnitt, eine Thorakotomie parasternal links und die Punktion des linken
Ventrikels mittels einer Kanüle. So konnte über eine Infusion Dextran 40
(Rheomacrodex) unter Zusatz von 0,5% Procainhydochlorid (Novocain) intrakardial
eingeleitet und das Gefäßsystem zwei Minuten gespült werden. Dextran 40 soll hierbei
einer thrombotischen Gefäßverlegung entgegenwirken und als onkotisch wirksame
Lösung ein interstitielles Ödem verhindern. Procain führt dabei am Gefäßsystem zu
einer Dilatation und am Herzen schließlich zum Herzstillstand.
Nach durchgeführter Spülung erfolgte eine zweite mit dreiprozentigem Glutaraldehyd in
0,2 molarer Phosphatpufferlöung (Sörensenpufferlösung) unter einem Perfusionsdruck
von 110 mmHg für zwölf Minuten zur Fixation der Organe.

16
4.4 Gewebeaufarbeitung
Bei allen Tieren wurden nach Fixation der Organe das Herz und die Aorta sowie beide
Nieren entnommen. Nach Messung der Nierengewichte wurden jeweils beide Nieren in
Glutaraldehyd 3% und 0,2 molarer Phosphatpufferlösung eingelagert. Die Aorta wurde
durch einen vertikal zum Gefäßlumen verlaufenden Schnitt tief abdominell quer
durchtrennt. Danach wurde das Gefäß direkt am Eintritt in den linken Ventrikel
abgesetzt. Nach Messung des Herzgewichtes wurden das Herz und die Aorta sodann in
10 % gepuffertem Formalin fixiert.
Da vor der Perfusion das Körpergewicht (KG) der Tiere bestimmt worden war, konnte
man unter Einbeziehung der ermittelten Nierengewichte (NG) und Herzgewichte (HG)
das jeweilige relative Gewicht dieser Organe (relatives mittleres Nierengewicht: rmNG;
relatives Herzgewicht: rHG) mittels folgenden Formeln bestimmen:
mNG = (NGlinks + NGrechts) / 2 [mg]
rmNG = mNG/KG x 100 [%]
rHG = HG/KG x 100 [%]
Da bei der späteren Verarbeitung jeweils nochmals die Organgewichte bestimmt
wurden, liegen diese Werte auch für die fixierten und gereinigten Organe vor.
4.5 Technik der Paraffinschnitte
4.5.1 Paraffinschnitte Niere
Senkrecht zu ihrer Interpolarachse wurden die Nieren in fünf 1,5 – 2 mm dicke
Scheiben geschnitten. Durch waschen in Sörensenpuffer und anschliessender
Dehydrierung in aufsteigender Ethanolreihe (30–100%-iger Ethanol), wurden die
gewonnenen Nierenscheiben für die Einbettung vorbereitet. Die Einbettung erfolgte in
Paraplast Tissue-tek II-Einbettkassetten, die mit Paraffin ausgegossen wurden. Mittels
eines Rotationsmikrotoms (RM25 von Leica) wurden im Anschluß 1 µm dicke

17
Paraffinschnitte hergestellt. Die hierdurch erhaltenen Schnitte wurden schließlich PAS
(Periocid-Acid-Schiff-Reaktion) gefärbt.
4.5.2 Paraffinschnitte Herz und Aorta
Die Herzen wurden horizontal so gedrittelt, dass die entstehenden Teilstücke eine
nahezu gleich große Dicke aufwiesen. Sowohl die Aorten als auch die 3 Herzanteile
wurden 24 Stunden gewässert und anschliessend durch eine aufsteigende Alkoholreihe
(30-100 % Ethanol) dehydriert. Es erfolgte die Einbettung mittels Paraplast in Tissue-
tek II Einbettkassetten, die mit Paraffin ausgegossen wurden. Im Anschluß wurden mit
Hilfe des Rotationsmikrotoms 1 μm dicke Paraffinschnitte angefertigt. Zur Auswertung
morphometrischer Parameter erfolgte abschließend die Färbung der Präparate mit
Hämatoxylin und Eosin.
4.6 Analyse
4.6.1 Analyse der Herzen und der Aorta
Die morphometrische Auswertung der Herzmaße, der intramyokardialen Arterien und
der Aorta erfolgte mit Hilfe eines halbautomatischen Bildanalyse-Systems (Analysis
PRO, SIS, Münster, Deutschland), wie vorausgehend detailliert beschrieben.[22] Alle
Messungen und Auswertungen erfolgten geblindet, d.h. der Auswertende hatte keinerlei
Kenntnisse über die Gruppenzugehörigkeit der Präparate.
4.6.1.1 Analyse der Herzen
Zur Beurteilung der Herzen wurden folgende Parameter untersucht:
Linker Ventrikel, LV (μm): Dicke des linken Ventrikels
Septum, S (μm): Dicke des Septums
Rechter Ventrikel, RV (μm): Dicke des rechten Ventrikels
Die Messung der Parameter erfolgte mit Hilfe eines halbautomatischen
Bildanalysesystems (SIS, Münster, Deutschland) bei 4-facher (linker Ventrikel und

18
Septum), bzw. 20-facher (rechter Ventrikel) Vergrößerung. Aus jeweils 10 gleichmäßig
verteilten Messungen pro Tier wurden die Durchschnittswerte berechnet.
Zur Beurteilung der intramyokardialen Gefäße wurden folgende Parameter gemessen
und berechnet:
Wanddicke, WD (μm, berechnet): Dicke der Gefäßwand
Wand-Lumen-Ratio, WLR (μm/μm): Quotient aus Wanddicke und
Lumendiameter
In die Messungen wurden pro Tier zwischen 6 und 10 intramyokardiale Arterien
einbezogen. Alle Messungen erfolgten mittels Analysis-System bei 400-facher
Vergrößerung. Die Ermittlungen der Werte wurden so durchgeführt, dass bei den meist
nicht senkrecht angeschnittenen Gefäßen jeweils der kürzeste messbare
Lumendurchmesser und - in direkt darauf liegender Projektion - der kleinste
Gesamtdurchmesser markiert und gemessen wurde. Dann wurde der
Lumendurchmesser vom Gesamtdurchmesser subtrahiert und das Ergebnis durch zwei
dividiert und so die Wanddicke des Gefäßes bestimmt. Dieses Vorgehen diente der
Minimierung möglicher Fehlerquellen (Schnittwinkelartefarkte, Messfehler bei sehr
kleinen Messstrecken), so dass die Messwerte den wahren Werten der Gefäße am
nahsten kommen. Die Wand-Lumen-Ratio ergab sich aus dem Quotienten aus einfacher
Wanddicke und Lumendiameter.
4.6.1.2 Analyse der Aorten
Zur Beurteilung der Aorten wurden folgende Parameter untersucht:
Wanddicke, WD (μm, berechnet): Dicke der Media
Die Wanddicke der Aorta wurde nicht wie bei den Arteriolen berechnet, sondern als
Mittelwert aus 10 willkürlichen Messungen der Mediadicke der thorakalen Aorta
ermittelt.

19
4.6.2 Analyse der Nieren
4.6.2.1 Schädigungsindices
4.6.2.1.1 Glomeruloskleroseindex (GSI)
Eine Methode zur Beurteilung einer glomerulären Schädigung ist die Bestimmung der
Proliferation des Mesangiums und die Vermehrung der mesangialen Matrix. Mit Hilfe
eines Lichtmikroskopes wurden die PAS gefärbten Paraffinschnitte in 400 facher
Vergrößerung mäanderförmig durchfahren, wobei pro Präparat 50 Glomeruli beurteilt
wurden. Dabei wurde die Schädigung gemäß der Methode von El Nahas et al. [9] in
folgende Stadien eingeteilt:
Stadium histologische Veränderungen Anteil der Veränderung
des Konvolutes 0 normales Glomerulum 0 % 1 mesangiale Verdickung mit und ohne
Proliferation von Mesangiumzellen; keine
Kapillarbeteiligung
< 25 %
2 mesangiale Proliferation mit partieller
Gefäßwandbeteiligung, segmentale Sklerose
25 bis 50 %
3 große Teile der Kapillaren mit Obliteration
oder Narbenformation, diffuse Sklerose
50 bis 75 %
4 totale Obliteration der Kapillaren mit und
ohne Kapillarthrombose, globale Sklerose
mit Kapillarkollaps
> 75 %
Der Glomeruloskleroseindex ließ sich folgendermaßen berechnen:
GSI = ( 0 x n0 + 1 x n1 + 2 x n2 + 3 x n3 + 4 x n4 ) / n0 + n1 + n2 + n3 + n4
( n0, n1, n2, n3, n4 : Anzahl der Glomeruli mit Stadium 0 bis 4 )
20
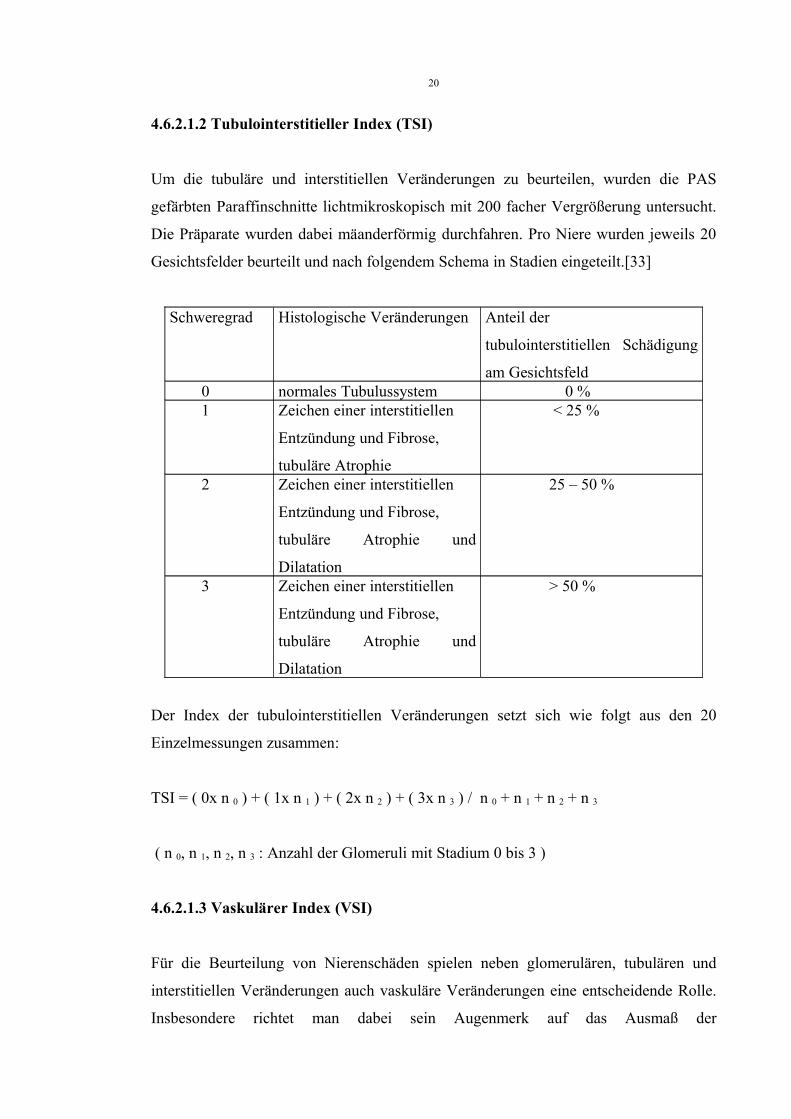
4.6.2.1.2 Tubulointerstitieller Index (TSI)
Um die tubuläre und interstitiellen Veränderungen zu beurteilen, wurden die PAS
gefärbten Paraffinschnitte lichtmikroskopisch mit 200 facher Vergrößerung untersucht.
Die Präparate wurden dabei mäanderförmig durchfahren. Pro Niere wurden jeweils 20
Gesichtsfelder beurteilt und nach folgendem Schema in Stadien eingeteilt.[33]
Schweregrad Histologische Veränderungen Anteil der
tubulointerstitiellen Schädigung
am Gesichtsfeld 0 normales Tubulussystem 0 % 1 Zeichen einer interstitiellen
Entzündung und Fibrose,
tubuläre Atrophie
< 25 %
2 Zeichen einer interstitiellen
Entzündung und Fibrose,
tubuläre Atrophie und
Dilatation
25 – 50 %
3 Zeichen einer interstitiellen
Entzündung und Fibrose,
tubuläre Atrophie und
Dilatation
> 50 %
Der Index der tubulointerstitiellen Veränderungen setzt sich wie folgt aus den 20
Einzelmessungen zusammen:
TSI = ( 0x n 0 ) + ( 1x n 1 ) + ( 2x n 2 ) + ( 3x n 3 ) / n 0 + n 1 + n 2 + n 3
( n 0, n 1, n 2, n 3 : Anzahl der Glomeruli mit Stadium 0 bis 3 )
4.6.2.1.3 Vaskulärer Index (VSI)
Für die Beurteilung von Nierenschäden spielen neben glomerulären, tubulären und
interstitiellen Veränderungen auch vaskuläre Veränderungen eine entscheidende Rolle.
Insbesondere richtet man dabei sein Augenmerk auf das Ausmaß der

21
Gefäßwandverdickung und der fibrinoiden Nekrose der afferenten Arteriolen,
interlobulären Gefäße und kleinen Arterien. Dazu wurden 20 Gesichtsfelder pro Niere
in 200 facher Vergrößerung mit dem Lichtmikroskop untersucht und in folgende
Stadien eingeteilt.[33]
Stadium Histologische Veränderungen Gefäßwandverdickung0 Normale Gefäße 0 %1 Geringe Gefäßwandverdickung < 25 %2 Moderate Gefäßwandverdickung 25-50 %3 Schwere Gefäßwandverdickung >50 %4 Fibrinoide Nekrose der Gefäße
Der Index der Gefäßveränderungen wurde aus dem Mittelwert der Einzelbestimmungen
berechnet:
VSI = ( 0x n 0 ) + ( 1x n 1 ) + ( 2x n 2 ) + ( 3x n 3 ) + ( 4x n 4 ) / n 0 + n 1 + n 2 + n 3 + n 4
( n 0, n 1, n2, n 3, n 4 : Anzahl der Glomeruli mit Stadium 0 bis 4 )
4.6.2.2 Glomerulusgeometrie
4.6.2.2.1 Morphometrie und Stereologie
Morphometrie und Stereologie sind Begriffe aus der Naturwissenschaft und umfassen
Verfahren zur Beschreibung räumlicher Anordnungen von Strukturen.[8] Zur
quantitativen Erfassung von Strukturelementen und Partikeln benutzt man
morphometrische Methoden.[24] Durch stereologische Methoden kann man aus
zweidimensionalen Abbildungen Informationen über dreidimensionale Strukturen
gewinnen.[35,36] Ermöglicht wird dies durch geometrische Wahrscheinlichkeiten, nach
denen sich eine dreidimensionale Struktur in ihren zufällig getroffenen
zweidimensionalen Abschnitten abbildet. Bezogen auf histologische Schnittpräparate ist
es dadurch möglich, Aussagen über die räumliche Struktur des untersuchten Gewebes
treffen zu können.
Delesse [7] beschreibt ein Verfahren, um das Volumen einer Struktur im histologischen
Schnitt zu bestimmen. Danach ist die volumetrische Zusammensetzung eines Organs

22
direkt aus der Flächenzusammensetzung der Schnittflächen abzuleiten. Um nun den
Volumenanteil einer Struktur zu bestimmen wird das sogenannte Punkte-Zähl-
Verfahren genutzt.
Hierfür wurde ein Mikroskop genutzt, dessen 10er Okular mit einer Integrationsplatte
ausgestattet war, auf dem sich ein quadratisches Gitter mit symmetrisch verteilten
Testpunkten befand. Diese Punkte projizierten sich direkt via Okular auf den jeweiligen
histologischen Schnitt. Die Formel: VV = AA besagt, dass die volumetrische
Zusammensetzung eines Gewebes im histologischen Schnitt unabhängig von seiner
räumlichen Orientierung direkt in der Flächenzusammensetzung der Schnittflächen
repräsentiert ist. Der Volumenanteil VV eines Objekts ist gleich dem Flächenanteils AA.
Des Weiteren ist die Anzahl der Punkte, die sich zufällig auf eine zu messende Struktur
projiziert, auch gleich dessen Flächenanteil: PP = AA . Dabei steht PP für die Anzahl der
gezählten Punkte, die auf das zu messende Gewebe fallen im Verhältnis zur Gesamtzahl
der Punkte des Punktegitters. Daraus ergibt sich dann: VV = PP , was bedeutet, dass der
Volumenanteil VV dem Anteil der Trefferpunkte, bezogen auf die Gesamtpunktzahl des
Messgitters entspricht.
Voraussetzung für die genannte Methode der Volumenbestimmung ist die
Unabhängigkeit des Delesse `schen Prinzips von der räumlichen Orientierung der
betreffenden Objekte. Nach Weibel [35,36] ist diese Voraussetzung erfüllt.
Die Fläche des Punktegitters wurde in den jeweiligen Vergrößerungen mittels eines
kalibrierten Objektmikrometers bestimmt und so die Flächen des Messgitters berechnet.
Dazu wurde dessen bekannter Maßstab mit der Skala des Okulargitters zur Deckung
gebracht und die Punktabstände bzw. die Seitenlängen des Quadrats ausgemessen.
4.6.2.2.2 Auswertung
Alle Messungen wurden von einer Person ohne Kenntnis der Gruppenzugehörigkeit der
zu untersuchenden histologischen Präparate durchgeführt.
Die Untersuchungen der Nierenschnitte wurden mit einem Lichtmikroskop (Fa.
Olympus BH- 2) mit 10er Okular und je nach zu bestimmendem Parameter mit einem
10er, 20er, 40er oder 100er Objektiv (mit Immersionsöl) durchgeführt.
Die Beurteilung der Nieren wurden an PAS gefärbten Paraffinschnitten
lichtmikroskopisch (Objektive 10, 20, 40) vorgenommen. Für die Untersuchungen

23
wurde die beschriebene Punkte- Zähl- Methode angewendet und dabei folgende Größen
bestimmt:
Punkte auf Nierenmark
Punkte auf Nierenrinde
Punkte auf Glomeruli
Anzahl Glomerli
4.6.2.2.2.1 Volumenfraktion von Nierenmark und Nierenrinde
In 100 facher Vergrößerung wurde mit Hilfe des beschriebenen Punkte- Zähl-
Verfahrens die Volumenfraktion von Nierenrinde (VVR) und Nierenmark (VVM)
bestimmt.
Dabei galten folgende Formeln:
Punkte auf Nierenrinde
VVR = ——————————— x 100 % [ % ]
Punkte auf Niere
Punkte auf Nierenmark
VVM = ——————————— x 100 % [ % ]
Punkte auf Niere
4.6.2.2.2.2 Gesamtvolumen Niere und absolutes Volumen Nierenrinde
Mit folgender Formel wurde aus dem gemessenen Nierengewicht m das
Gesamtvolumen der jeweiligen Niere VN berechnet:
VN = m / rho x 1000 [ mm 3]
rho : Dichte der Niere, empirischer Wert 1,04 g/ cm 3
m : Nierengewicht in g

24
Mit Hilfe des Gesamtnierenvolumens konnte nun das absoluten Volumen der
Nierenrinde VR berechnet werden:
VR = VVR x VN / 100 [ mm 3 ]
4.6.2.2.2.3 Glomeruli pro Fläche Rinde
In 100 facher Vergrößerung konnte über eine Bestimmung der Fläche der
Integrationsplatte mit Hilfe einer Eichskala die Nierenrindenfläche (AR) und damit dann
die Anzahl der Glomeruli pro Fläche Rinde (NA) berechnet werden. Die Fläche der
Integrationsfläche (10 x 10 Punkte) betrug dabei 0,7832 mm 2 . Somit ergaben sich
folgende Formeln:
AR = Punkte Rinde x 0,7832 / 100 [ mm 2 ]
Zahl der Glomeruli
NA = —————————— [1/ mm2 ]
AR x ( 1,08 ) 2
( 1,08 ) 2 entspricht dabei dem Schrumpfungsindex der Niere durch die Organfixation.
[24]
4.6.2.2.2.4 Volumendichte der Glomeruli
Die Volumendichte der Glomeruli in der Nierenrinde (VVGlom) wurde nach folgender
Formel berechnet:
Punkte Glomeruli
VVGlom = ————————— x 100 % [ % ]
Punkte Nierenrinde

25
4.6.2.2.2.5 Numerische Dichte der Glomeruli in der Rinde
Aus der Anzahl der Glomeruli pro Fläche Rinde (NA) und der Volumendichte der
Glomeruli (VVGlom ) ergab sich dann die Anzahl der Glomeruli pro Volumen (Nv) :
k
Nv = —— x NA 3/2 / (VVGlom x 0,01 ) 0,5 [ 1/ mm 3 ]
ß
k entspricht dem sogenannten Größenkoeffizienten der untersuchten Teilchen und hat
abhängig von der Form und Volumendichte den Wert 1,04. Der Koeffizient ß beschreibt
als Formfaktor die Gestalt der gemessenen Objekte und beträgt für die annähernd
sphärische Form der Glomeruli 1,38.[1] Denn nach Weibel und Gomez [34] gilt: Das
Verhältnis zwischen der Teilchenzahl in der Volumeneinheit und der Querschnittszahl
in der Fläche einer angeschnittenen Oberfläche ist unabhängig von einer
Größenänderung der Teilchen, vorausgesetzt, ihre Gestalt ist geometrisch ähnlich, was
für Glomeruli angenommen werden kann.
4.6.2.2.2.6 Gesamtzahl und Gesamtvolumen der Glomeruli einer Niere
Mit nachfolgender Formel ließ sich dann die absolute Anzahl der Glomeruli einer Niere
(NGlom) berechnen:
NGlom = Nv x VR
Für das Gesamtvolumen der Glomeruli einer Niere (VNGlom) galt dann entsprechend:
VNGlom = VR x VVGlom / 100 [ mm3 ]

26
4.6.2.2.2.7 Mittleres glomeruläres Volumen
Aus den oben dargestellten Formeln ließ sich dann das mittlere glomeruläre Volumen
(VGlom) folgendermaßen berechnen:
VGlom = VNGlom / NGlom x 106 [µm 3 ]
4.7 Statistik
Die statistische Auswertung erfolgte mittels SPSS, Version 10. Dabei wurden alle Daten
zunächst auf Normalverteilung der Werte und Homogenität der Varianzen geprüft.
Da es sich bei unseren Daten um unabhängige Variablen handelte, wurden
normalverteilte, homogene Stichproben mittels einfaktorieller ANOVA und
anschliessender Testung durch Bonferroni auf Unterschiede untersucht.
Normalverteilte, nicht homogene Stichproben wurden zunächst dem Test nach Brown
Forsythe unterzogen und schliesslich mittels Tamhane untersucht. Signifikanz wurde
dann angenommen, wenn die Irrtumswahrscheinlichkeit p kleiner als 0,05 war.

27
5 Ergebnisse
Alle Parameter werden als Mittelwerte mit Standardabweichungen der einzelnen
Gruppen angegeben.
5.1 Allgemeine Parameter
5.1.1 Körpergewicht
Tabelle 1Gruppe Genotyp Körpergewicht [g]
1 ♂ jung ko 26,9 ± 4,0
2 ♂ jung wt 23,8 ± 1,5
3 ♂ alt ko 29,9 ± 2,8
4 ♂ alt wt 31,2 ± 2,8
5 ♀ alt ko 31,5 ± 3,3
6 ♀ alt wt 30,3 ± 2,3ANOVA: p<0,01Bonferroni: 2 vs. 4 p<0,01
Bei den Körpergewichten zeigt sich ein signifikant niedrigerer Wert der jungen wildtyp
Tiere gegenüber den älteren männlichen wildtyp Tieren.
5.1.2 Herzgewicht (HG) und relatives Herzgewicht (rHG)
Tabelle 2Gruppe Genotyp HG [mg]
1 ♂ jung ko 162,4 ± 17,8
2 ♂ jung wt 129,7 ± 11,4
3 ♂ alt ko 185,6 ± 11,7
4 ♂ alt wt 187,5 ± 19,2
5 ♀ alt ko 169,0 ± 20,6
6 ♀ alt wt 161,8 ± 19,1ANOVA: p<0,01, Bonferroni:2 vs. 1 p<0,05; 2 vs. 4 p<0,001;
28
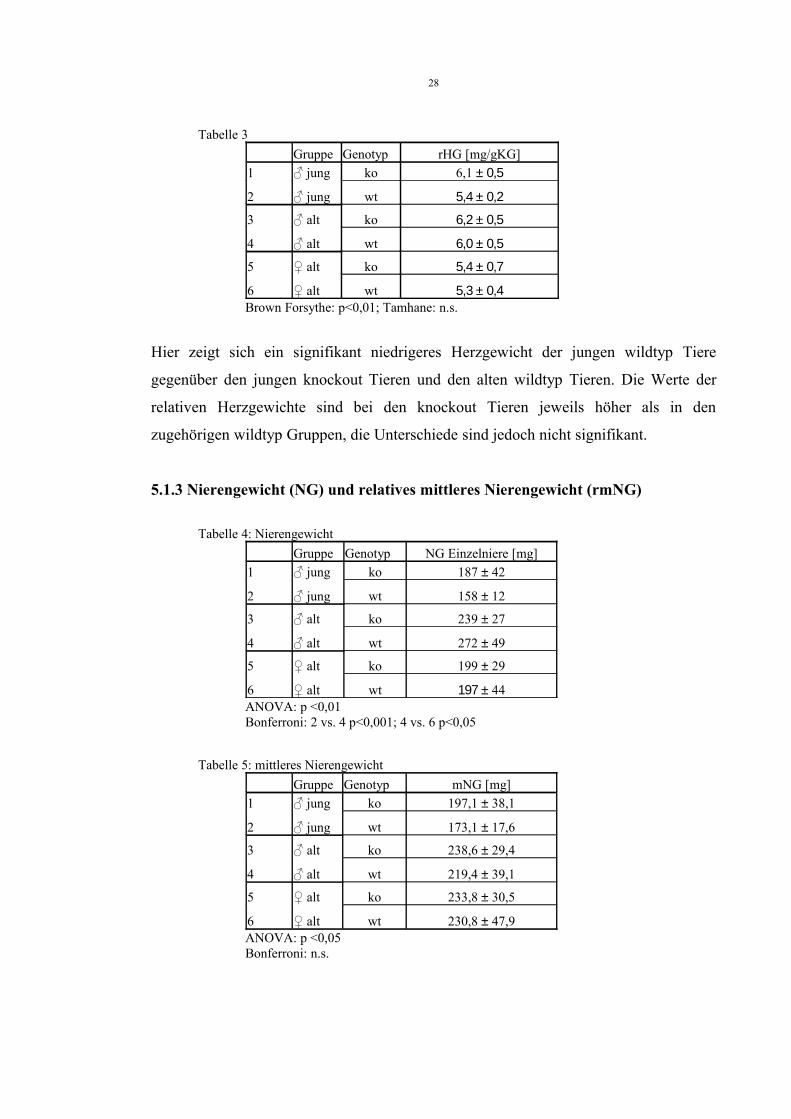
Tabelle 3Gruppe Genotyp rHG [mg/gKG]
1 ♂ jung ko 6,1 ± 0,5
2 ♂ jung wt 5,4 ± 0,2
3 ♂ alt ko 6,2 ± 0,5
4 ♂ alt wt 6,0 ± 0,5
5 ♀ alt ko 5,4 ± 0,7
6 ♀ alt wt 5,3 ± 0,4Brown Forsythe: p<0,01; Tamhane: n.s.
Hier zeigt sich ein signifikant niedrigeres Herzgewicht der jungen wildtyp Tiere
gegenüber den jungen knockout Tieren und den alten wildtyp Tieren. Die Werte der
relativen Herzgewichte sind bei den knockout Tieren jeweils höher als in den
zugehörigen wildtyp Gruppen, die Unterschiede sind jedoch nicht signifikant.
5.1.3 Nierengewicht (NG) und relatives mittleres Nierengewicht (rmNG)
Tabelle 4: NierengewichtGruppe Genotyp NG Einzelniere [mg]
1 ♂ jung ko 187 ± 42
2 ♂ jung wt 158 ± 12
3 ♂ alt ko 239 ± 27
4 ♂ alt wt 272 ± 49
5 ♀ alt ko 199 ± 29
6 ♀ alt wt 197 ± 44ANOVA: p <0,01Bonferroni: 2 vs. 4 p<0,001; 4 vs. 6 p<0,05
Tabelle 5: mittleres NierengewichtGruppe Genotyp mNG [mg]
1 ♂ jung ko 197,1 ± 38,1
2 ♂ jung wt 173,1 ± 17,6
3 ♂ alt ko 238,6 ± 29,4
4 ♂ alt wt 219,4 ± 39,1
5 ♀ alt ko 233,8 ± 30,5
6 ♀ alt wt 230,8 ± 47,9ANOVA: p <0,05Bonferroni: n.s.
29
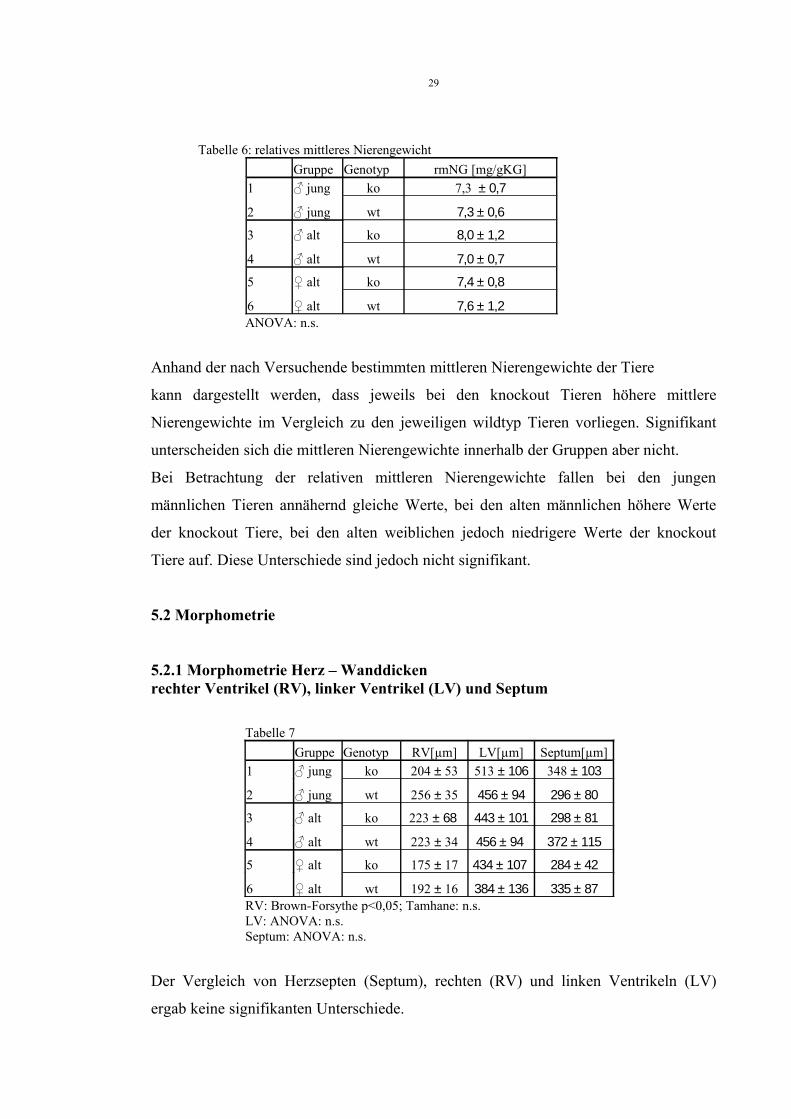
Tabelle 6: relatives mittleres NierengewichtGruppe Genotyp rmNG [mg/gKG]
1 ♂ jung ko 7,3 ± 0,7
2 ♂ jung wt 7,3 ± 0,6
3 ♂ alt ko 8,0 ± 1,2
4 ♂ alt wt 7,0 ± 0,7
5 ♀ alt ko 7,4 ± 0,8
6 ♀ alt wt 7,6 ± 1,2ANOVA: n.s.
Anhand der nach Versuchende bestimmten mittleren Nierengewichte der Tiere
kann dargestellt werden, dass jeweils bei den knockout Tieren höhere mittlere
Nierengewichte im Vergleich zu den jeweiligen wildtyp Tieren vorliegen. Signifikant
unterscheiden sich die mittleren Nierengewichte innerhalb der Gruppen aber nicht.
Bei Betrachtung der relativen mittleren Nierengewichte fallen bei den jungen
männlichen Tieren annähernd gleiche Werte, bei den alten männlichen höhere Werte
der knockout Tiere, bei den alten weiblichen jedoch niedrigere Werte der knockout
Tiere auf. Diese Unterschiede sind jedoch nicht signifikant.
5.2 Morphometrie
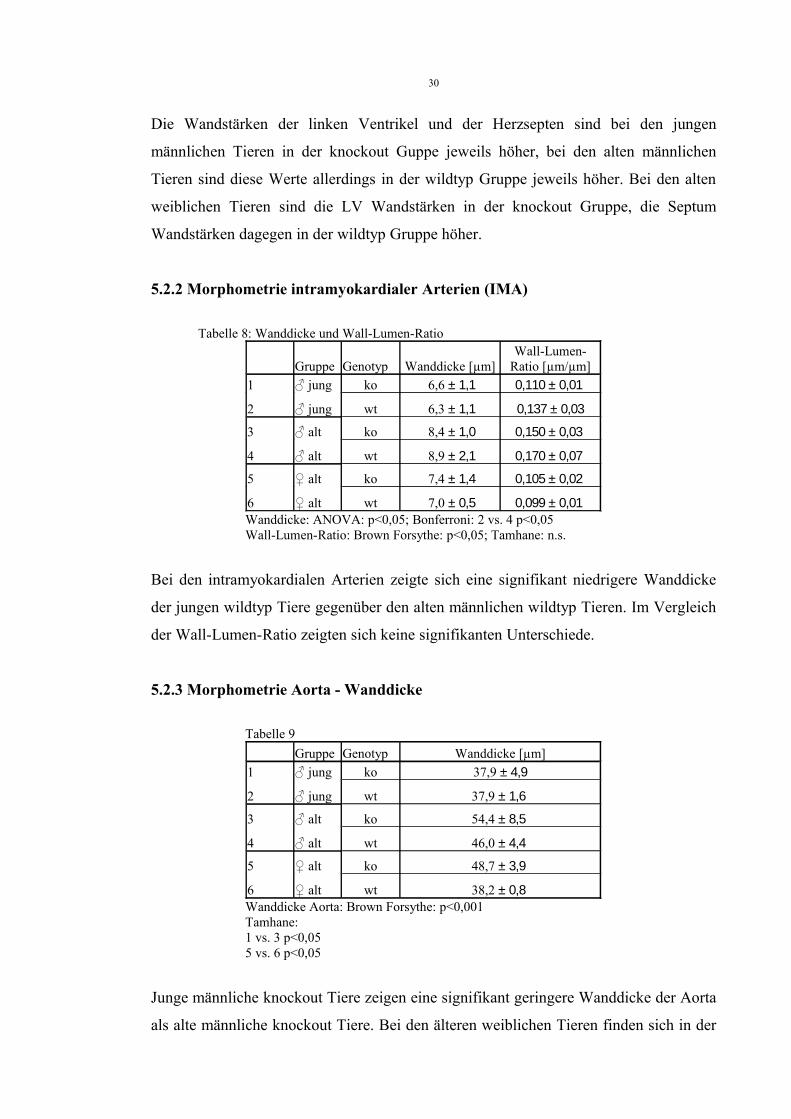
5.2.1 Morphometrie Herz – Wanddicken rechter Ventrikel (RV), linker Ventrikel (LV) und Septum
Tabelle 7Gruppe Genotyp RV[µm] LV[µm] Septum[µm]
1 ♂ jung ko 204 ± 53 513 ± 106 348 ± 103
2 ♂ jung wt 256 ± 35 456 ± 94 296 ± 80
3 ♂ alt ko 223 ± 68 443 ± 101 298 ± 81
4 ♂ alt wt 223 ± 34 456 ± 94 372 ± 115
5 ♀ alt ko 175 ± 17 434 ± 107 284 ± 42
6 ♀ alt wt 192 ± 16 384 ± 136 335 ± 87RV: Brown-Forsythe p<0,05; Tamhane: n.s.LV: ANOVA: n.s.Septum: ANOVA: n.s.
Der Vergleich von Herzsepten (Septum), rechten (RV) und linken Ventrikeln (LV)
ergab keine signifikanten Unterschiede.
30
Die Wandstärken der linken Ventrikel und der Herzsepten sind bei den jungen
männlichen Tieren in der knockout Guppe jeweils höher, bei den alten männlichen
Tieren sind diese Werte allerdings in der wildtyp Gruppe jeweils höher. Bei den alten
weiblichen Tieren sind die LV Wandstärken in der knockout Gruppe, die Septum
Wandstärken dagegen in der wildtyp Gruppe höher.
5.2.2 Morphometrie intramyokardialer Arterien (IMA)
Tabelle 8: Wanddicke und Wall-Lumen-Ratio
Gruppe Genotyp Wanddicke [µm]Wall-Lumen-
Ratio [µm/µm]1 ♂ jung ko 6,6 ± 1,1 0,110 ± 0,01
2 ♂ jung wt 6,3 ± 1,1 0,137 ± 0,03
3 ♂ alt ko 8,4 ± 1,0 0,150 ± 0,03
4 ♂ alt wt 8,9 ± 2,1 0,170 ± 0,07
5 ♀ alt ko 7,4 ± 1,4 0,105 ± 0,02
6 ♀ alt wt 7,0 ± 0,5 0,099 ± 0,01 Wanddicke: ANOVA: p<0,05; Bonferroni: 2 vs. 4 p<0,05Wall-Lumen-Ratio: Brown Forsythe: p<0,05; Tamhane: n.s.
Bei den intramyokardialen Arterien zeigte sich eine signifikant niedrigere Wanddicke
der jungen wildtyp Tiere gegenüber den alten männlichen wildtyp Tieren. Im Vergleich
der Wall-Lumen-Ratio zeigten sich keine signifikanten Unterschiede.
5.2.3 Morphometrie Aorta - Wanddicke
Tabelle 9Gruppe Genotyp Wanddicke [µm]
1 ♂ jung ko 37,9 ± 4,9
2 ♂ jung wt 37,9 ± 1,6
3 ♂ alt ko 54,4 ± 8,5
4 ♂ alt wt 46,0 ± 4,4
5 ♀ alt ko 48,7 ± 3,9
6 ♀ alt wt 38,2 ± 0,8 Wanddicke Aorta: Brown Forsythe: p<0,001Tamhane:1 vs. 3 p<0,055 vs. 6 p<0,05
Junge männliche knockout Tiere zeigen eine signifikant geringere Wanddicke der Aorta
als alte männliche knockout Tiere. Bei den älteren weiblichen Tieren finden sich in der

31
knockout Gruppe ebenfalls signifikant höhere Werte im Vergleich zu den wildtyp
Tieren. Bei den älteren männlichen Tieren sind die Werte der knockout Tiere ebenfalls
höher als bei den wildtyp Tieren, die Werte der jungen Tiere sind allerdings annähernd
gleich und diese Unterschiede sind zudem nicht signifikant.
5.3 Niere
5.3.1 Semiquantitative Untersuchungen – renale Schädigungsindices
Tabelle 10Gruppe Genotyp GSI TSI VSI
1 ♂ jung ko 0,38 ± 0,13 0,76 ± 0,27 0,11 ± 0,09
2 ♂ jung wt 0,22 ± 0,08 0,33 ± 0,18 0,17 ± 0,05
3 ♂ alt ko 0,37 ± 0,08 0,85 ± 0,36 0,21 ± 0,1
4 ♂ alt wt 0,25 ± 0,07 0,39 ± 0,14 0,18 ± 0,06
5 ♀ alt ko 0,49 ± 0,16 0,68 ± 0,33 0,29 ± 0,13
6 ♀ alt wt 0,38 ± 0,19 0,32 ± 0,15 0,18 ± 0,09 GSI: Brown Forsythe: p<0,05; Tamhane: n.s.TSI: ANOVA: p< 0,05; Bonferroni: 1 vs. 2 p<0,05 und 3 vs. 4 Tendenz p=0,055VSI: ANOVA: p< 0,05; Bonferroni: n.s.
Ein Vergleich des Glomeruloskleroseindex (GSI) zur Beurteilung einer glomerulären
Schädigung ergibt zwar jeweils höhere Werte der knockout Tiere im Vergleich zu den
wildtyp Tieren, aber aufgrund der Standardabweichung keine signifikanten
Unterschiede.
Die tubulären und interstitiellen Veränderungen (TSI) sind bei den jungen männlichen
knockout Tieren signifikant höher als bei den altersgleichen wildtyp Tieren. Bei den
älteren männlichen knockout Tieren zeigen sich tendenziell höhere Werte im Vergleich
zur wildtyp Gruppe und auch bei den älteren weiblichen Tiere sind die Werte der
knockout Gruppe höher als die der wildtyp Tiere.
Bei den alten Tieren beiden Geschlechts sind die VSI Werte der knockout Tiere jeweils
höher als die der wildtyp Tiere, nur bei den jungen männlichen Tieren sind die Werte
der wildtyp Tiere höher, diese Unterschiede sind jedoch nicht signifikant.
32
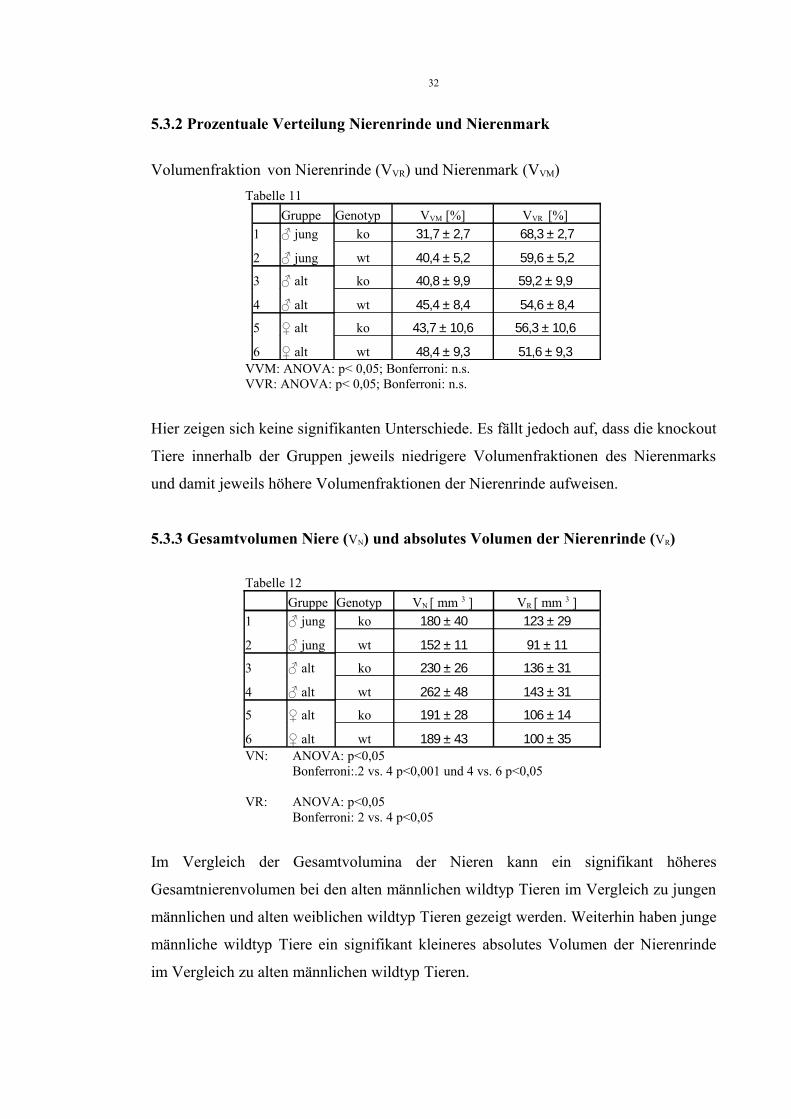
5.3.2 Prozentuale Verteilung Nierenrinde und Nierenmark
Volumenfraktion von Nierenrinde (VVR) und Nierenmark (VVM) Tabelle 11
Gruppe Genotyp VVM [%] VVR [%] 1 ♂ jung ko 31,7 ± 2,7 68,3 ± 2,7
2 ♂ jung wt 40,4 ± 5,2 59,6 ± 5,2
3 ♂ alt ko 40,8 ± 9,9 59,2 ± 9,9
4 ♂ alt wt 45,4 ± 8,4 54,6 ± 8,4
5 ♀ alt ko 43,7 ± 10,6 56,3 ± 10,6
6 ♀ alt wt 48,4 ± 9,3 51,6 ± 9,3 VVM: ANOVA: p< 0,05; Bonferroni: n.s.VVR: ANOVA: p< 0,05; Bonferroni: n.s.
Hier zeigen sich keine signifikanten Unterschiede. Es fällt jedoch auf, dass die knockout
Tiere innerhalb der Gruppen jeweils niedrigere Volumenfraktionen des Nierenmarks
und damit jeweils höhere Volumenfraktionen der Nierenrinde aufweisen.
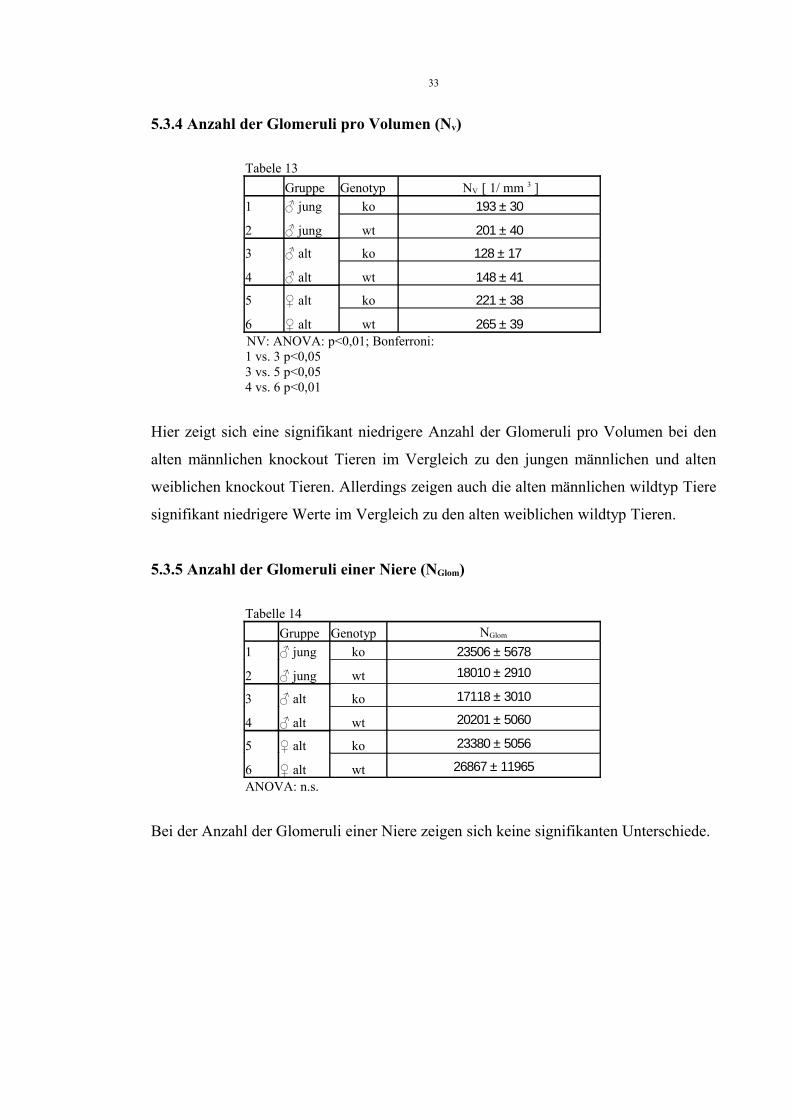
5.3.3 Gesamtvolumen Niere (VN) und absolutes Volumen der Nierenrinde (VR)
Tabelle 12Gruppe Genotyp VN [ mm 3 ] VR [ mm 3 ]
1 ♂ jung ko 180 ± 40 123 ± 29
2 ♂ jung wt 152 ± 11 91 ± 11
3 ♂ alt ko 230 ± 26 136 ± 31
4 ♂ alt wt 262 ± 48 143 ± 31
5 ♀ alt ko 191 ± 28 106 ± 14
6 ♀ alt wt 189 ± 43 100 ± 35VN: ANOVA: p<0,05
Bonferroni:.2 vs. 4 p<0,001 und 4 vs. 6 p<0,05
VR: ANOVA: p<0,05 Bonferroni: 2 vs. 4 p<0,05
Im Vergleich der Gesamtvolumina der Nieren kann ein signifikant höheres
Gesamtnierenvolumen bei den alten männlichen wildtyp Tieren im Vergleich zu jungen
männlichen und alten weiblichen wildtyp Tieren gezeigt werden. Weiterhin haben junge
männliche wildtyp Tiere ein signifikant kleineres absolutes Volumen der Nierenrinde
im Vergleich zu alten männlichen wildtyp Tieren.
33
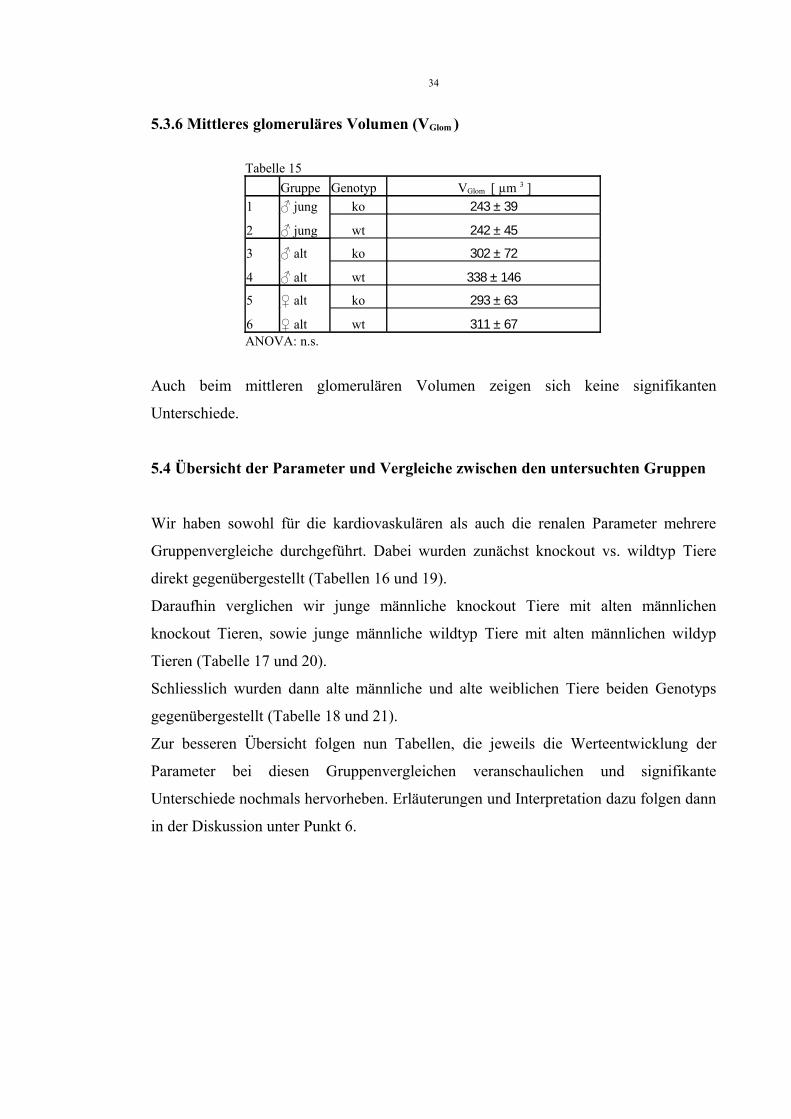
5.3.4 Anzahl der Glomeruli pro Volumen (Nv)
Tabele 13Gruppe Genotyp NV [ 1/ mm 3 ]
1 ♂ jung ko 193 ± 30
2 ♂ jung wt 201 ± 40
3 ♂ alt ko 128 ± 17
4 ♂ alt wt 148 ± 41
5 ♀ alt ko 221 ± 38
6 ♀ alt wt 265 ± 39NV: ANOVA: p<0,01; Bonferroni:1 vs. 3 p<0,053 vs. 5 p<0,054 vs. 6 p<0,01
Hier zeigt sich eine signifikant niedrigere Anzahl der Glomeruli pro Volumen bei den
alten männlichen knockout Tieren im Vergleich zu den jungen männlichen und alten
weiblichen knockout Tieren. Allerdings zeigen auch die alten männlichen wildtyp Tiere
signifikant niedrigere Werte im Vergleich zu den alten weiblichen wildtyp Tieren.
5.3.5 Anzahl der Glomeruli einer Niere (NGlom)
Tabelle 14Gruppe Genotyp NGlom
1 ♂ jung ko 23506 ± 5678
2 ♂ jung wt 18010 ± 2910
3 ♂ alt ko 17118 ± 3010
4 ♂ alt wt 20201 ± 5060
5 ♀ alt ko 23380 ± 5056
6 ♀ alt wt 26867 ± 11965ANOVA: n.s.
Bei der Anzahl der Glomeruli einer Niere zeigen sich keine signifikanten Unterschiede.
34
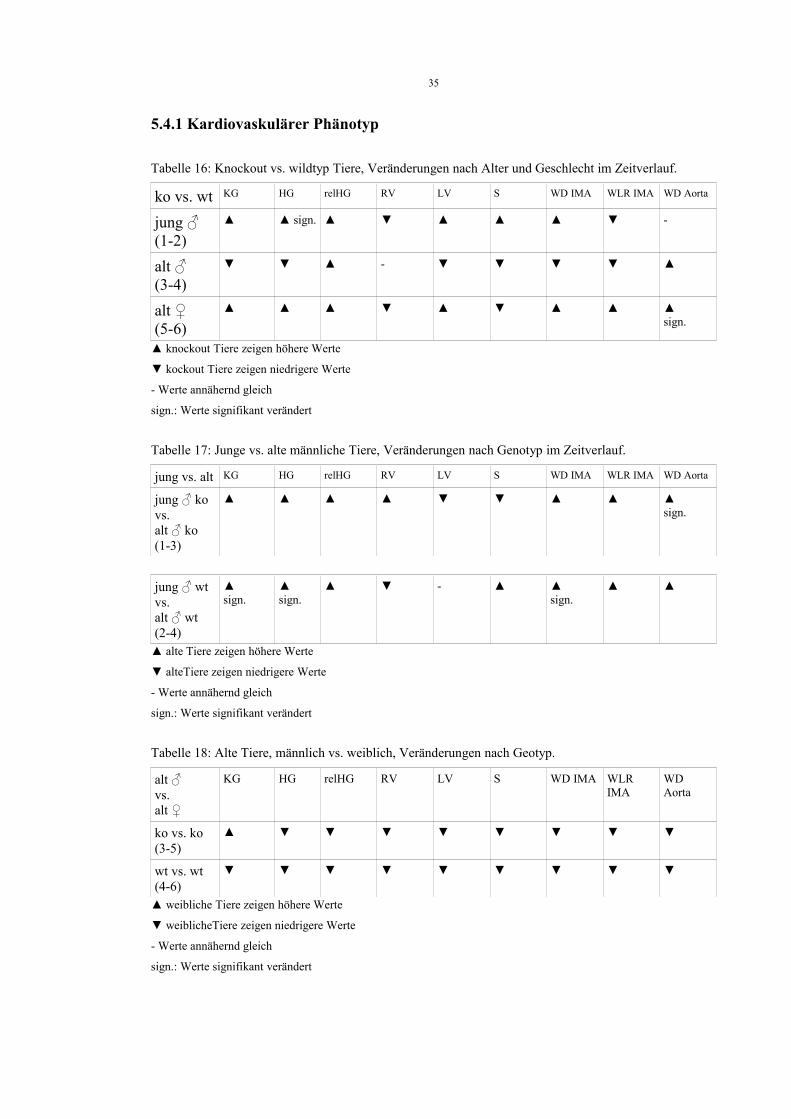
5.3.6 Mittleres glomeruläres Volumen (VGlom )
Tabelle 15Gruppe Genotyp VGlom [ µm 3 ]
1 ♂ jung ko 243 ± 39
2 ♂ jung wt 242 ± 45
3 ♂ alt ko 302 ± 72
4 ♂ alt wt 338 ± 146
5 ♀ alt ko 293 ± 63
6 ♀ alt wt 311 ± 67ANOVA: n.s.
Auch beim mittleren glomerulären Volumen zeigen sich keine signifikanten
Unterschiede.
5.4 Übersicht der Parameter und Vergleiche zwischen den untersuchten Gruppen
Wir haben sowohl für die kardiovaskulären als auch die renalen Parameter mehrere
Gruppenvergleiche durchgeführt. Dabei wurden zunächst knockout vs. wildtyp Tiere
direkt gegenübergestellt (Tabellen 16 und 19).
Daraufhin verglichen wir junge männliche knockout Tiere mit alten männlichen
knockout Tieren, sowie junge männliche wildtyp Tiere mit alten männlichen wildyp
Tieren (Tabelle 17 und 20).
Schliesslich wurden dann alte männliche und alte weiblichen Tiere beiden Genotyps
gegenübergestellt (Tabelle 18 und 21).
Zur besseren Übersicht folgen nun Tabellen, die jeweils die Werteentwicklung der
Parameter bei diesen Gruppenvergleichen veranschaulichen und signifikante
Unterschiede nochmals hervorheben. Erläuterungen und Interpretation dazu folgen dann
in der Diskussion unter Punkt 6.
35
5.4.1 Kardiovaskulärer Phänotyp
Tabelle 16: Knockout vs. wildtyp Tiere, Veränderungen nach Alter und Geschlecht im Zeitverlauf.
ko vs. wt KG HG relHG RV LV S WD IMA WLR IMA WD Aorta
jung ♂(1-2)
▲ ▲ sign. ▲ ▼ ▲ ▲ ▲ ▼ -
alt ♂(3-4)
▼ ▼ ▲ - ▼ ▼ ▼ ▼ ▲
alt ♀(5-6)
▲ ▲ ▲ ▼ ▲ ▼ ▲ ▲ ▲sign.
▲ knockout Tiere zeigen höhere Werte
▼ kockout Tiere zeigen niedrigere Werte
- Werte annähernd gleich
sign.: Werte signifikant verändert
Tabelle 17: Junge vs. alte männliche Tiere, Veränderungen nach Genotyp im Zeitverlauf.
jung vs. alt KG HG relHG RV LV S WD IMA WLR IMA WD Aorta
jung ♂ ko vs. alt ♂ ko (1-3)
▲ ▲ ▲ ▲ ▼ ▼ ▲ ▲ ▲ sign.
jung ♂ wt vs. alt ♂ wt (2-4)
▲sign.
▲sign.
▲ ▼ - ▲ ▲sign.
▲ ▲
▲ alte Tiere zeigen höhere Werte
▼ alteTiere zeigen niedrigere Werte
- Werte annähernd gleich
sign.: Werte signifikant verändert
Tabelle 18: Alte Tiere, männlich vs. weiblich, Veränderungen nach Geotyp.
alt ♂ vs. alt ♀
KG HG relHG RV LV S WD IMA WLR IMA
WD Aorta
ko vs. ko(3-5)
▲ ▼ ▼ ▼ ▼ ▼ ▼ ▼ ▼
wt vs. wt(4-6)
▼ ▼ ▼ ▼ ▼ ▼ ▼ ▼ ▼
▲ weibliche Tiere zeigen höhere Werte
▼ weiblicheTiere zeigen niedrigere Werte
- Werte annähernd gleich
sign.: Werte signifikant verändert
36
5.4.2 Renaler Phänotyp
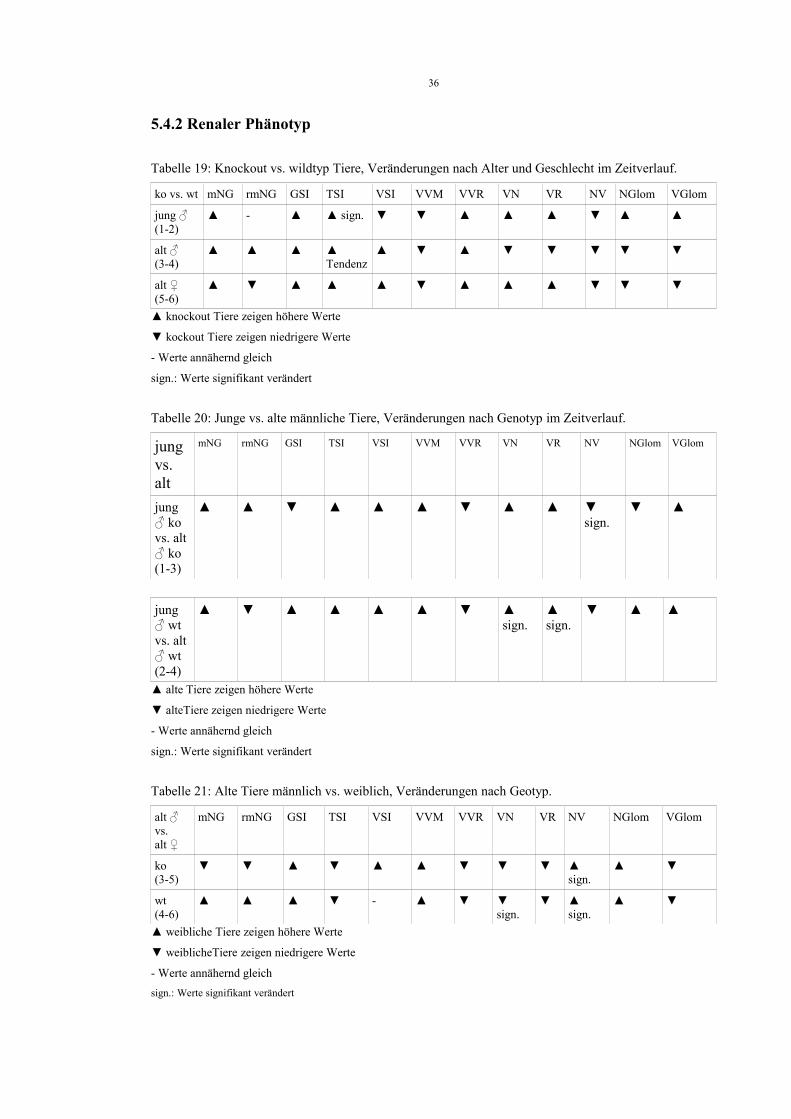
Tabelle 19: Knockout vs. wildtyp Tiere, Veränderungen nach Alter und Geschlecht im Zeitverlauf.
ko vs. wt mNG rmNG GSI TSI VSI VVM VVR VN VR NV NGlom VGlom
jung ♂(1-2)
▲ - ▲ ▲ sign. ▼ ▼ ▲ ▲ ▲ ▼ ▲ ▲
alt ♂(3-4)
▲ ▲ ▲ ▲Tendenz
▲ ▼ ▲ ▼ ▼ ▼ ▼ ▼
alt ♀(5-6)
▲ ▼ ▲ ▲ ▲ ▼ ▲ ▲ ▲ ▼ ▼ ▼
▲ knockout Tiere zeigen höhere Werte
▼ kockout Tiere zeigen niedrigere Werte
- Werte annähernd gleich
sign.: Werte signifikant verändert
Tabelle 20: Junge vs. alte männliche Tiere, Veränderungen nach Genotyp im Zeitverlauf.
jung vs. alt
mNG rmNG GSI TSI VSI VVM VVR VN VR NV NGlom VGlom
jung ♂ ko vs. alt ♂ ko (1-3)
▲ ▲ ▼ ▲ ▲ ▲ ▼ ▲ ▲ ▼ sign.
▼ ▲
jung ♂ wt vs. alt ♂ wt (2-4)
▲ ▼ ▲ ▲ ▲ ▲ ▼ ▲ sign.
▲ sign.
▼ ▲ ▲
▲ alte Tiere zeigen höhere Werte
▼ alteTiere zeigen niedrigere Werte
- Werte annähernd gleich
sign.: Werte signifikant verändert
Tabelle 21: Alte Tiere männlich vs. weiblich, Veränderungen nach Geotyp.
alt ♂ vs. alt ♀
mNG rmNG GSI TSI VSI VVM VVR VN VR NV NGlom VGlom
ko(3-5)
▼ ▼ ▲ ▼ ▲ ▲ ▼ ▼ ▼ ▲sign.
▲ ▼
wt(4-6)
▲ ▲ ▲ ▼ - ▲ ▼ ▼sign.
▼ ▲sign.
▲ ▼
▲ weibliche Tiere zeigen höhere Werte
▼ weiblicheTiere zeigen niedrigere Werte
- Werte annähernd gleichsign.: Werte signifikant verändert

37
6 Diskussion
Der BK Kanal der Zellmembran glatter Gefäßmuskelzellen (VSMCs) wurde bereits
1992 von Brayden et al. [3] als wichtige Determinante des Vasotonus und damit des
arteriellen Blutdrucks beschrieben. BK Kanäle sind mit BK β-Untereinheiten besetzt,
welche die porenbildende BK α-Einheit gewebespezifisch in deren Eigenschaften
beeinflussen. Die BK β1-Untereinheit, kodiert durch das KCNMB1 Gen [4], in VSMCs
trägt dazu bei, intrazelluläre Kalziumströme und auswärts gerichtete Kaliumströme der
Zellmembran zu koppeln und ist daher maßgeblich an der Regulation der Kontraktilität
dieser Zellen beteiligt. Damit vereinbar zeigen Mäuse mit einer Deletion dieser BK β1-
Untereinheit typische Veränderungen der Eigenschaften von BK Kanälen und
entwickeln eine milde, aber signifikante arterielle Hypertonie.
Brenner et al. [4] konnten 2000 eine reduzierte Kalziumsensitivität der BK β1 knockout
Kanäle und damit eine deutliche Störung der physiologischen negativen Rückkopplung
der Vasokonstriktion in vitro nachweisen. Weiterhin fanden sie bei BK β1 knockout
Tieren eine deutlich vermehrte Kontraktion zerebraler Arterien und in Untersuchungen
in vivo zeigten sie dann, dass BK β1 knockout Mäuse eine milde arterielle Hypertonie
entwickeln, die etwa 14 – 20 mmHg über den Werten von 129svj wildtyp Tieren liegt.
Daraus schlossen sie, dass BK β1 knockout Tiere über einen gesteigerten arteriellen
Gefäßtonus eine arterielle Hypertonie entwickeln.[4,18,28] Letztlich bewerteten sie das
BK β1 Tiermodell als außergewöhnliche Möglichkeit, bei bekanntem, klar definiertem
molekularen Defekt, die sekundären Effekte einer Hypertonie zu erforschen. Sie
beschreiben beispielsweise ein im Vergleich zur Kontrollgruppe signifikant höheres
relatives Herzgewicht von BK β1 knockout Tieren als Herzhypertrophie im Rahmen der
milden Hypertonie der BK β1 knockout Tiere.[4]
Plüger et al. [26] zeigten 2000 bei BK β1 knockout Tieren eine gestörte intrazelluläre
Signalübermittlung in VSMCs, eine gesteigerte Kontraktilität thorakaler
Aortenpräparate und ebenfalls eine chronische Erhöhung des Blutdrucks dieser Tiere.
Löhn et al. [18] beschreiben 2001 ebenfalls eine gestörte intrazelluläre
Signalübermitlung bei BK β1 knockout Tieren sowie eine chronische Hypertonie im
Tiermodell, allerdings lassen sie die Gründe für die Entstehung des Hypertonus offen.

38
Brenner et al. [4] und Plüger et al. [26] heben jeweils das gewebespezfische
Vorkommen der BK β Untereinheiten hervor und betonen, dass BK β1-Untereinheiten
überwiegend in VSMCs vorkommen. Auch Nelson et al. [23] gehen davon aus, dass die
β1-Untereinheit der BK Kanäle exklusiv in glatten Muskelzellen vorkommt und die
Hypertonie bei BK β1 knockout Tieren rein vaskulären Ursprungs ist.
Pluznick et. al. [28] beschrieben 2003 zunächst, dass die BK β1-Untereinheit des BK
Kanals überwiegend in glatten Gefäßmuskelzellen vorzufinden ist, dort die
Kalziumsensitivität der Zellen moduliert und damit für die Regulation des Gefäßtonus
eine wichtige Rolle spielt. Da damals aber schon bekannt war, dass BK α-Einheiten in
verschiedenen Regionen der Niere vorkommen, untersuchten sie bereits renale
Auswirkungen einer BK β1 Gendeletion und fanden signifikante Unterschiede der
glomerulären Filtrationsrate, sowie der Natrium- und Kaliumausscheidung unter akuter
Volumenbelastung.
Anhand weiterer Studien konnten Pluznick et al. und Grimm et al. [12,27,28,29]
belegen, dass eben auch andere Gewebe, darunter auch Strukturen der Niere, β1-
Untereinheiten enthalten. Aus dem Nachweis signifikanter Veränderungen von
glomerulärer Filtrationsrate und der Kaliumausscheidung unter Volumenbelastung
schlossen sie, dass ein renal bedingter Aldosteronismus der BK β1 knockout Tiere die
Hauptursache des Hypertonus dieses Tiermodells ist.[12] Damit wurde erstmals die
These der rein vasogen bedingten Hypertonie dieser Tiere in Frage gestellt. Zur Klärung
der Ursache einer Erhöhung des Langzeitblutdrucks im Tiermodell sind daher nun vor
allem renale, hormonelle und myogene Einflussfaktoren zu bedenken. Nachdem man
lang vom gewebespezfischen Vorkommen der BK β1-Untereinheit in VSMCs ausging
und damit die glatte Gefäßmuskulatur als Hauptdeterminante untersuchte, ist nun die
Nieren- und Nebennierenfunktion im Fokus der Forschung.
Bis heute wurden allerdings nur wenige Daten zu Veränderungen der histologischen
Struktur von Herz, Gefäßen und renalem Gewebe veröffentlicht.[4,26,27]
Brenner et al. [4] beschrieben strukturelle Veränderungen am Herzen als
Endorganschaden der milden Hypertonie homozygoter BK β1 knockout Tiere. Auch
beim Menschen führt ein länger bestehender, essentieller Bluthochdruck langfristig zu
einer Herzvergrößerung mit linksventrikulärer Hypertrophie. [37] Eine Gendeletion bei
BK β1 knockout Tieren könnte aber per se den histologischen Phänotyp dieser Tiere
verändern.

39
Die vorliegende Arbeit sollte klären, ob bei BK β1 knockout Tieren im Vergleich zur
gesunden Kontrollpopulation ein veränderter kardialer und oder renaler Phänotyp
nachzuweisen ist und in welcher Weise das Alter und das Geschlecht der Tiere den
Phänotyp beeinflusst. Zur Überprüfung der anfangs formulierten Hypothese wurden
Herzen, Gefäße und Nieren bei homozygoten BK β1 knockout Mäusen
unterschiedlichen Alters und Geschlechts im Vergleich zu wildtyp Tieren untersucht.
6.1 Interpretation des Phänotyps
6.1.1 Knockout vs. wildtyp
Um Auswirkungen der BK β1 Gendeletion beschreiben zu können verglichen wir
jeweils
(i) junge männliche knockout Tiere mit jungen männlichen wildtyp Tieren,
(ii) alte männliche knockout Tiere mit alten männlichen wildtyp Tieren und
(iii) alte weibliche knockout Tiere mit alten weiblichen wildtyp Tieren.
6.1.1.1 Kardialer Phänotyp
Wäre nach Brenner et al. [4] eine kardiale Hypertrophie und damit eine signifikante
Erhöhung der relativen Herzgewichte zumindest der alten knockout Tiere als (Spät)-
Folge des Hypertonus zu erwarten, so zeigten unsere Ergebnisse keine signifikanten
Unterschiede, lediglich die absoluten Werte der relativen Herzgewichte waren jeweils in
allen knockout Gruppen höher. Zur genaueren Beurteilung haben wir neben den
relativen Herzgewichten auch die Werte der Wandstärken der linken Ventrikel
verglichen. Aber auch diese zeigten keine signifikanten Unterschiede im Sinne einer
Herzhypertrophie der knockout Tiere. Dies ist um so erstaunlicher, als die knockout
Tiere in Brenners Arbeit [4] alle männlich und ca. 15 Wochen alt, die alten Tieren in
unserer Arbeit dagegen zwischen 68 und 96 Wochen und damit deutlich älter waren. Im
Gegensatz zu Brenner et al.[4], die nur junge männliche Tiere in ihre Messungen
miteinbezogen, können wir zwar eine Zunahme der relativen Herzgewichte auch bei
alten knockout Tieren beiden Geschlechts beobachten, aber eben keine signifikante
Hypertrophie beschreiben. Die linksventrikuläre Wandstärke der alten männlichen
knockout Tiere war sogar nicht signifikant niedriger als in der wildtyp Gruppe. So
können wir mit unseren Zahlen weder die Ergebnisse von Brenner bestätigen, noch

40
einen signifikanten alters- oder geschlechtsabhängigen Unterschied beschreiben.
Sausbier et al. [30] konnten bei 16 bis 24 Wochen alten BK α knockout Tieren ebenfalls
keine signifikante kardiale Hypertrophie nachweisen, obwohl deren knockout Tiere eine
signifikante Hypertonie entwickelten. Die Autoren begründeten ihre Ergebnisse mit
geringeren Blutdruckwerten ihrer Versuchstiere im Vergleich zu Brenners BK β1
knockout Tieren. Blutdruckmessungen wurden an unseren Tieren nicht durchgeführt.
Um den kardialen Phänotyp noch genauer zu beurteilen, wurden neben relativen
Herzgewichten und Wandstärken der linken Ventrikel die Wandstärken und die Wall-
Lumen-Ratio von intramyokardialen Arterien gemessen. Auch hier zeigten sich aber
keine signifikanten Unterschiede. Da der Genotyp unserer Versuchstiere dem der Tiere
entspricht, die Plüger et al. [26] untersuchten, nehmen wir an, dass der Blutdruck in
unseren knockout Gruppen ebenfalls signifikant erhöht war. Unklar bleibt die
Werteentwicklung der älteren knockout Tiere, bei denen keine signifikante Veränderung
des kardialen Phänotyps nachzuweisen war. Unsere Ergebnissen sprechen aber
zumindest gegen eine bereits früh bestehende, signifikante Veränderung des kardialen
Phänotyps und damit gegen unsere anfangs formulierte Hypothese.
6.1.1.2 Vaskulärer Phänotyp
Die Wandstärken der Aorta der jungen männlichen Tiere unterschieden sich kaum,
sowohl alte männliche als auch alte weibliche knockout Tiere hatten jeweils dickere
Wandstärken als die zugehörige Kontrollgruppe, die Werte der alten weiblichen
knockout Tiere waren sogar signifikant höher. Diese Zunahme der Aortenwandstärken
bei älteren knockout Tieren beiden Geschlechts deuten auf eine zeitabhängige vaskuläre
Veränderung der knockout Tiere hin, die sich sowohl mit der von Plüger et al.[26]
gefundenen erhöhten Kontraktilität thorakaler Aortenringe bei BK β1 knockout Tieren
als auch mit der milden Hypertonie dieser Tiere erklären ließe. Der nicht signifikante
Unterschied bei den alten männlichen knockout Tieren könnte mit dem geringeren
mittleren Alter im Vergleich zu den alten weiblichen Tieren begründet werden.
6.1.1.3 Renaler Phänotyp
Die Schädigungsindices GSI, TSI und VSI ergaben bei allen älteren knockout Tieren
jeweils höhere Werte als bei den zugehörigen wildtyp Tieren. Niedrigere VSI Werte der

41
jungen knockout Tiere bei gleichzeitig jeweils höheren VSI Werten der älteren
knockout Tiere wären ebenfalls mit einer zeitabhängigen renovaskulären Schädigung
vereinbar, die wir so auch bei den Aortenwandstärken beschreiben konnten; die
Unterschiede sind hier jedoch statistisch nicht signifikant. Eine zeitabhängige
renovaskuläre Schädigung spräche auch gegen die Ergebnisse von Magnusson et al.
[19], die dem BK Kanal keine wichtige Rolle bei der Steuerung des renalen Vasotonus
zuschreiben. Sie könnte aber mit der sich entwickelnden Hypertonie dieser Tiere in
Verbindung gebracht werden.
Signifikant höhere TSI Werte der jungen knockout Tiere und tendentiell höhere Werte
der alten männlichen knockout Tiere sowie nicht signifikant höhere TSI Werte der alten
weiblichen knockout Tiere könnten als (bereits früh beginnende) tubuläre Veränderung
gedeutet werden. Es stellt sich jedoch die Frage, warum sich die Signifikanz der
Veränderung bei den jungen nicht bei den älteren Tiere fortsetzt. Zu bedenken ist, dass
die Werte der erhobenen Schädigungsindices alle unter 1,0 lagen. Bei theoretisch
möglichen Werten von 0 bis 4 für GSI und VSI und 0-3 für TSI spricht dies allein schon
für sehr geringe Unterschiede und stellt die meist nicht signifikanten Veränderungen
damit bezüglich deren Aussagekraft zumindest teilweise in Fage.
Auffällig ist die Werteentwicklung der älteren Tiere für die absolute Anzahl der
Glomeruli einer Niere (NGlom) und das mittlere glomeruläre Volumen (VGlom). Diese
Werte waren bei den jungen männlichen knockout Tieren höher, bei den alten Tieren
beiden Geschlechts aber jeweils in der knockout Gruppe niedriger. Die geringere
Anzahl der Glomeruli pro Niere bei den alten knockout Tieren könnte als zeitabhängige
Schädigung zu werten sein. Gleichzeitig war aber das mittlere glomeruläre Volmen
VGlom jeweils bei den alten knockout Tieren erniedrigt. Dies entspricht nicht der
Erwartung einer sich entwickelnden glomerulären Schädigung, bei der VGlom höhere
Werte ergeben müsste.
Die absoluten Werte für GSI, Volumenfraktionen der Nierenrinde (VVR) und das
mittlere Nierengewicht waren bei allen knockout Tieren jeweils höher, die Anzahl der
Glomeruli pro Volumen (Nv) jeweils niedriger als bei den wildtyp Tieren. Diese Zahlen
belegen bei fehlender Signfikanz nur sehr geringe Veränderungen des renalen
Phänotyps, sprechen aber zumindest gegen eine signifikante frühe und
geschlechtsabhängige Veränderung des renalen Phänotyps.
Die Werte für das Gesamtvolumen Niere (VN) und das absolute Volumen der
Nierenrinde (VR) zeigten bei den alten Tieren eine unterschiedliche Entwicklung. Bei

42
den alten männlichen knockout Tieren waren die absoluten Zahlen erniedrigt, bei den
alten weiblichen knockout Tieren erhöht (jeweils im Vergleich zu den wildtyp
Gruppen). Da die Werteentwicklung bei den männlichen Tieren nicht mit einem
höheren mittleren Alter der männlichen Tiere erklärt werden kann, könnte man dies –
bei fehlender Signifikanz - als mögliche geschlechtsabhängige Veränderung des renalen
Phänotyps deuten, (vergleiche dazu auch Punkt 6.1.2.3 und 6.1.3.2).
6.1.2 Junge vs. alte Tiere
Um die Entwicklung der Tiere im Zeitverlauf genauer beurteilen zu können, verglichen
wir
(i) junge männliche knockout Tiere mit alten männlichen knockout Tieren,und
(ii) junge männliche wildtyp Tiere mit alten männlichen wildtyp Tieren.
6.1.2.1 Kardialer Phänotyp
Die Wanddicke der intramyokardialen Arterien war bei den älteren wildtyp Tieren
signifikant größer, woraus man eine rein zeitabhängige Veränderung des kardialen
Phänotyps bei den männlichen wildtyp Tieren ableiten könnte. Dies passt nicht zu den
Ergebnissen der Abeitsgruppe um Brenner [4,31,14], die zeigen konnte, dass keine
Korrelation zwischen zunehmenden Alter und den relativen Herzgewichten von
gesunden Tieren besteht. Die Werte der knockout Tiere zeigten hier keinen
signifikanten Unterschied, bei einem rein zeitäbhängigen Faktor wären auch hier
signifikante Unterschiede zu erwarten.
Im Vergleich der knockout Tiere fallen ein höheres relatives Herzgewicht, sowie höhere
Werte bei den Wandstärken der rechten Ventrikel bei den älteren Tieren auf - was die
These der zeitabhängigen Veränderungen unterstreichen würde. Allerdings waren die
Wandstärken der linken Ventrikel und die der Herzsepten bei den alten Tieren niedriger.
Dies spricht gegen die These einer im Zeitverlauf zunehmenden, späten Veränderung
des kardialen Phänotyps, zu bedenken ist aber die fehlende Signifikanz all dieser
Unterschiede.
Abgesehen von den Wandstärken der rechten Ventrikel und der Herzsepten waren alle
erhobenen Parameter in knockout und wildtyp Gruppen gleich gerichtet verändert. So

43
zeigten sich zum Beispiel sowohl in der knockout als auch der wildtyp Gruppe jeweils
höhere Werte der relativen Herzgewichte bei den älteren Tieren.
Eine zeitabhängige Veränderung des kardialen Phänotyps in beiden Gruppen kann
weder durch signifikante Unterschiede belegt, noch ausgeschlossen werden. In der
Summe sprechen unsere Ergebnisse aber zumindest gegen einen deutlichen Einfluss des
Genotyps auf zeitabhängige Veränderungen des kardialen Phänotyps männlicher Tiere.
6.1.2.2 Vaskulärer Phänotyp
Die Werte der Aortenwandstärken weisen auch bei diesem Vergleich auf eine
zeitabhängige Schädigung/Veränderung hin, da sowohl alte knockout als auch alte
wildtyp Tiere jeweils höhere Aortenwandstärken aufwiesen verglichen mit jungen
männlichen Tieren gleichen Genotyps. Signifikant unterschieden sich jedoch nur die
knockout Tiere voneinander. Dies unterstreicht die unter 6.1.1.2 genannte These, dass
zeitabhängige vaskuläre Veränderung bei BK β1 knockout Tieren stattfinden. Dies
spricht eher gegen eine bereits früh embryonal bestehende vaskuläre Schädigung durch
die Gendeletion. Zukünftige Arbeiten könnten die Ätiologie der Wandstärkenzunahme
klären. In diesem Zusammenhang kommen die von Plüger et al.[26] gefundene erhöhte
Kontraktilität thorakaler Aortenringe und die sich entwickelnde Hypertonie dieser Tiere
in Frage.
6.1.2.3 Renaler Phänotyp
Die relativen mittleren Nierengewichte der alten knockout Tiere waren höher, die der
alten wildtyp Tiere niedriger, bei der Anzahl der Glomeruli einer Niere (NGlom) und dem
GSI Wert verhielt es sich genau umgekehrt. Abgesehen von diesen drei Parametern war
die Werteentwicklung aber sowohl in wildtyp als auch knockout Gruppe gleich.
Bei höheren Werten der älteren knockout als auch wildtyp Tiere kann eine rein
zeitabhängige, genotypunabhängige Veränderung nicht ausgeschlossen weden,
beziehungsweise anders ausgedrückt, keine Veränderung beschrieben werden, die
eindeuig durch Gendeletion erklärbar ist.
Diese Aussage gilt für VSI, TSI, das Gesamtvolumen Niere (VN), das absolute
Volumen der Nierenrinde (VR) und das mittlere glomeruläre Volumen (VGlom).

44
Die höheren VSI Werte der älteren knockout als auch wildtyp Tiere deuten - wie für die
knockout Tiere bereits in 6.1.1.3 beschrieben – ebenfalls auf eine zeitabhängige
Veränderung hin.
Das Gesamtvolumen der Niere (VN) und das absolutes Volumen der Nierenrinde (VR )
zeigten beim Vergleich wildtyp vs. knockout (siehe 6.1.1.3) eine geschlechtsabhängige
Werteentwicklung mit höheren Werten bei den weiblichen, niedrigeren bei den
männlichen Tieren. Hier vergleichen wir nur männliche Tiere gleichen Genotyps, aber
unterschiedlichen Alters. Nach den Ergebnissen unter 6.1.1.3 wären niedrigere Werte
zumindest der älteren knockout Tiere zu erwarten. Tatsächlich fanden wir aber höhere
Werte in dieser Gruppe. Ganz unerwartet fanden wir sogar signifikant höhere Werte für
VN und VR bei den alten wildtyp Tieren.
Damit lässt sich der in 6.1.1.3. vermutete geschlechtsabhängige Unterschied der
knockout Tiere nicht bestärken, wir müssen uns vielmehr fragen, wie die signifikant
höheren Werten der alten wildtyp Tiere im Vergleich zu den jungen wildtyp Tieren zu
erklären sein könnten. Leider fehlt uns hier ein Vergleich von jungen weiblichen Tieren
mit alten weiblichen Tieren.
Das mittlere glomeruläre Volumen (VGlom) ist bei den älteren Tieren in absoluten Zahlen
erhöht, was im Vergleich wildtyp vs knockout - wie in 6.1.1.3.beschrieben - für die
alten knockout Tiere nicht der Fall war und so nicht in das Bild einer sich
entwickelnden Nierenschädigung passt.
Bei niedrigeren Werten der älteren knockout als auch wildtyp Tiere kann eine rein
zeitabhängige, genotypunabhängige Veränderung ebenfalls nicht ausgeschlossen weden.
Diese Aussage gilt für die Glomeruli pro Volumen, die alten knockout Tiere zeigten
hier allerdings signifikant niedrigere Werte, was bei nicht signifikant niedrigeren
Werten der alten wildtyp Tiere dann eben doch für eine Genotyp- und zeitabhängige
renale Schädigung sprechen könnte.
6.1.3 Alte Tiere gleichen Genotyps männlich vs. weiblich
Um die Entwicklung der Tiere im Zeitverlauf abhängig vom Geschlecht beurteilen zu
können, verglichen wir
(i) alte männliche knockout Tiere mit alten weiblichen knockout Tieren, und
(ii) alte männliche wildtyp Tiere mit alten weiblichen wildtyp Tieren.

45
6.1.3.1 Kardiovaskulärer Phänotyp
Es fanden sich bei den männlichen Tieren für alle erhobenen kardiovaskulären
Parameter jeweils höhere absolute Werte im Vergleich zu den weiblichen Tieren, alle
Veränderungen zeigten keine Signifikanz und waren für knockout und wildtyp Tiere
jeweils gleich gerichtet verändert.
Hier können wir eine stärkere Schädigung der alten männlichen Tiere beiden Genotyps
beschreiben, bei fehlender Signifikanz ist eine Interpretation allerdings schwierig.
Gerade beim vaskulären Phänotyp waren die alten weiblichen knockout Tiere die
signifikant stärker geschädigte Gruppe (6.1.1.2). Ein Vergleich von jungen männlichen
knockout Tieren mit alten weiblichen knockout Tieren zeigte ebenfalls signifikant
höhere Aortenwandstärken bei den alten weiblichen Tieren.
Zur Klärung des Geschlechtseinflusses wäre es sinnvoll, in zukünftigen
Untersuchungen junge weibliche Tiere einzubeziehen und diese mit alten weiblichen
Tieren zu vergleichen, wie wir das für männliche Tiere unter 6.1.2 getan haben.
6.1.3.2. Renaler Phänotyp
Bei diesem Vergleich zeigten alle Werte ausser den Nierengewichten und VSI bei den
alten Weibchen in wildtyp und knockout Gruppen gleichartige Werteentwicklungen.
Auffällig sind hier die signifikant höheren Glomeruli pro Volumen (Nv) Werte alter
weiblicher wildtyp und knockout Tiere.
Die Anzahl der Glomeruli pro Volumen (Nv) war bei den weiblichen Tieren signifikant
größer, was für einen stärkere zeitabhängige Schädigung der männlichen Tiere spricht,
zumal eine höhere Anzahl der Glomeruli einer Niere (NGlom) und ein niedrigeres
mittleres glomeruläres Volumen (VGlom) der weiblichen Tiere - wenn auch nicht
signifikant - ebenfalls in diese Richtung deuten.
TSI ist bei den weiblichen Tieren kleiner, was für eine zeitabhängige stärkere
Schädigung der männlichen Tiere spricht, dies passt auch zu den Ergebnissen beim
Vergleich jung vs. alt. Der GSI ist bei den weiblichen Tieren jeweils größer, was für
eine zeitabhängige stärkere Schädigung der weiblichen Tiere sprechen könnte. Offenbar
ergibt sich bezüglich der renalen Schädigungsparameter der alten Tiere im
Geschlechtsvergleich kein klares Muster.

46
6.1.4 Schlussfolgerungen kardiovaskulärer Phänotyp
Nach den Ausführungen von Brenner et al. [4] und Sausbier et al. [30] hätten wir bei
deutlich älteren Tieren in unserem Versuchsaufbau eine signifikante Veränderung des
kardialen Phänotyps im Sinne einer Herzhypertrophie erwartet. Unklar bleibt, warum
wir keine signifikanten Veränderung der relativen Herzgewichte und der
linksventrikulären Wandstärken nachweisen konnten. Eindeutig genotypabhängige
Veränderungen des kardialen Phänotyps im Zeitverlauf konnten wir nicht nachweisen.
Bei unserem Vergleich junge vs. alte Tiere fanden wir lediglich unerwartet signifikant
höhere Wandstärken der intramyokardialen Arterien bei älteren männlichen wildtyp
Tieren, nicht jedoch bei den knockout Tieren.
Der vaskuläre Phänotyp zeigt – wie erwartet - signifikante zeitabhängige
Veränderungen bei den alten weiblichen knockout Tieren. Dies unterstreicht die
Beobachtungen von Plüger et al. [26], die bereits eine gesteigerte Kontraktilität von
Aortenpräparaten nachweisen konnten. Im Zeitverlauf zeigten auch alte männliche
knockout Tiere signifikant höhere Werte als junge knockout Tiere. Wie es scheint, sind
die Aorten (im Vergleich zu den kardialen Parametern) entweder vulnerabler, bezogen
auf die Auswirkungen einer sich entwickelnden Hypertonie, oder aber durch die
Gendeletion per se im Zeitverlauf stärker betroffen. Einen sicheren Einfluss des
Geschlechts auf den vaskulären Phänotyp können wir mit unseren Ergebnissen nicht
belegen.
6.1.5 Schlussfolgerungen renaler Phänotyp
Im direkten Vergleich knockout vs. wildtyp konnten wir nur signifikant erhöhte TSI
Werte der jungen männlichen knockout Tiere beobachten. Diese Entwicklung war bei
den alten Tieren beiden Geschlechts aber schon nicht mehr signifikant verändert und
kann so nur schwer als bereits früh bestehende tubuläre Schädigung gedeutet werden.
Alle anderen renalen Parameter waren nicht signifikant verändert und zeigten zudem
auch widersprüchliche Tendenzen, so dass auch eine späte Veränderung des renalen
Phänotyps nicht sicher beschrieben werden kann. Die Werteentwicklung für VSI könnte
in Richtung zeitabhängige renovaskuläre Schädigung bei knockout Tieren gedeutet
werden, die für das Gesamtvolumen Niere (VN) und das absolute Volumen der
Nierenrinde (VR) deutet auf eine geschlechtsabhängige Veränderung hin. Insgesamt

47
sprechen die Ergebnisse aber gegen bereits frühe Veränderungen des renalen Phänotyps
und damit auch hier gegen unsere anfangs formulierte Hypothese.
Bis auf wenige Parameter zeigen sowohl wildtyp als auch knockout Tiere im Vergleich
junge vs. alte Tiere gleichgerichtete Werteentwicklungen. Auffällig sind signifikant
höhere Werte bei den alten wildtyp Tieren für das Gesamtvolumen Niere (VN) und das
absolute Volumen der Nierenrinde (VR). Leider haben wir den Vergleich alt vs. jung nur
für männliche Tiere erhoben und keine Blutdruckmessungen durchgeführt, trotzdem
sprechen unsere Beobachtungen gegen einen deutlichen Einfluss der Gendeletion auf
den renalen Phänotyp und für eine eher zeitabhängige Veränderung unabhängig vom
Genotyp. Lediglich für die Anzahl der Glomeruli pro Volumen (Nv) passt die sinifikante
Werteentwicklung zum Bild einer genotyp- und zeitabhängigen renalen Schädigung
männlicher Tiere. Gegen die Genabhängigkeit und für die Geschlechts- und
zeitabhängige Veränderung spricht, dass die Anzahl der Glomeruli pro Volumen (Nv)
schliesslich beim Vergleich alter männlicher vs. alter weiblicher Tiere bei den
weiblichen Tieren beiden Genotyps signifikant größer war. Dies spricht für eine stärkere
rein zeitabhängige Schädigung der männlichen Tiere, zumal eine höhere Anzahl der
Glomeruli einer Niere (NGlom) und ein niedrigeres mittleres glomeruläres Volumen
(VGlom) der weiblichen Tiere - wenn auch nicht signifikant verändert - ebenfalls in diese
Richtung deuten. Die teils signifikante Werteentwicklung der Anzahl der Glomeruli pro
Volumen (Nv) ist also nicht eindeutig, wir können zum einen eine späte Schädigung
männlicher knockout Tiere beschreiben, zum anderen aber eine späte
geschlechtsabhängige stärkere Schädigung männlicher Tiere beiden Genotyps belegen,
die den Einfluss der Gendeletion wieder in Frage stellt.
6.2 Zusammenfassung
Zusammenfassend sprechen unsere Ergebnisse gegen eine bereits früh bestehende,
signifikante Veränderung des kardiovaskulären und renalen Phänotyps bei BK β1
knockout Tieren und damit gegen unsere anfangs formulierte Hypothese einer frühen
embryonalen Schädigung der kardiovaskulären und renalen Systeme durch Gendeletion.
Inwieweit geschlechtsabhängige Veränderungen des Phänotyps stattfinden, sollte durch
Einbeziehen weiblicher Tiere unterschiedlichen Alters und Genotyps in folgenden
Arbeiten genauer geprüft werden.

48
7 Literatur
1. Bendtsen TF, Nyengaard JR (1989) Unbiased estimation of particle number
using sections--an historical perspective with special reference to the stereology
of glomeruli. J Microsc 153:93-102.
2. Biron P, Mongeau JG, Bertrand D (1976) Familial aggregation of blood pressure
in 558 adopted children. Can Med Assoc J 115: 773–774.
3. Brayden JE, Nelson MT (1992) Regulation of arterial tone by activation of
calcium-dependent potassium channels. Science 256:532-535.
4. Brenner R, Peréz GJ, Bonev AD, Eckman DM, Kosek JC, Wiler SW, Patterson
AJ, Nelson MT, Aldrich RW (2000) Vasoregulation by the β1 subunit of the
calcium-activated potassium channel. Nature 407:870–876.
5. Bulpitt CJ, Beevers DG, Butler A, Coles EC; Hunt D; Munro-Faure AD,
Newson RB, O'Riordan PW, Petrie JC, Rajagopalan B (1986) The survival of
treated hypertensive patients and their causes of death: a report from the DHSS
hypertensive care computing project (DHCCP). J Hypertens 4:93–99
6. Crowley SD, Gurley SB, Oliverio MI, Pazmino AK, Griffiths R, Flannery PJ,
Spurney RF, Kim HS, Smithies O, Le TH, Coffman TM (2005) Distinct roles
for the kidney and systemic tissues in blood pressure regulation by the renin-
angiotensin system. J Clin Invest 115:1092–1099.
7. Delesse MA (1847) Procédé méchanique pour determiner la composition des
roches. Compt Rend Acad Sci 25:544-545.
8. Elias H, Hyde DM (1980) An elementary introduction to stereology
(quantitative microscopy). Am J Anat 159:412-446.

49
9. El Nahas AM, Bassett AH, Cope GH, LE Carpentier JE (1991) Role of growth
hormone in the development of experimental renal scarring. Kidney Int 40:29-
34.
10. Feinleib M, Garrison RJ, Fabsitz R, Christian JC, Hrubec Z, Borhani NO,
Kannel WB, Rosenman R, Schwartz JT, Wagner JO (1977) The NHLBI twin
study of cardiovascular disease risk factors: methodology and summary of
results. Am. J. Epidemiol 106:284–285.
11. Fernández-Fernández JM, Tomás M, Vázquez E, Orio P, Latorre R, Sentí M,
Marrugat J, Valverde MA (2004) Gain-of-function mutation in the KCNMB1
potassium channel subunit is associated with low prevalence of diastolic
hypertension. J Clin Invest 113: 1032–1039.
12. Grimm PR, Irsik DL, Settles DC, Holtzclaw JD, Sansom SC (2009)
Hypertension of Kcnmb1-/- is linked to deficient K secretion and aldosteronism.
Proc Natl acad sci USA 106:11800-11805.
13. Grimm PR, Sansom SC (2007) BK channels in the kidney. Curr Opin Nephrol
Hypertens 16:430–436.
14. Han J, Hosokawa M, Umezawa M, Yagi H, Matsushita T, Higuchi K, Takeda T
(1998) Age-related changes in blood pressure in the senescence-accelerated
mouse (SAM): aged SAMP1 mice manifest hypertensive vascular disease. Lab
Anim Sci 48:256-263.
15. Knot HJ, Nelson MT (1998) Regulation of arterial diameter and wall [Ca2 +] in
cerebral arteries of rat by membrane potential and intravascular pressure. J
Physiol (Lond). 1998;508:199 –209.
16. Knot HJ, Standen NB, Nelson MT (1998) Ryanodine receptors regulate arterial
diameter and wall [Ca2+] in cerebral arteries of rat via Ca2+-dependent K+
channels. J Physiol 508:211-221.

50
17. Lifton RP, Ghavari AG, Geller DS (2001) Molecular Mechanisms of Human
Hypertension. Cell 104:545–556.
18. Löhn M, Lauterbach B, Haller H, Pongs O, Luft FC, Gollasch M (2001) β1-
subunit of BK channels regulates arterial wall [Ca2+] and diameter in mouse
cerebral arteries. J Appl Physiol 91:1350–1354.
19. Magnusson L, Sorensen CM, Braunstein TH, Holstein-Rathlou NH,
Salomonsson M. (2007) Renovascular BKCa channels are not activated in vivo
under resting conditions and during agonist stimulation. Am J Physiol Regul
Integr Comp Physiol 292:R345–353.
20. Marteau JB, Zaiou M, Siest G, Visvikis-Siest S (2005) Genetic determinants of
blood pressure regulation. J Hypertens 23:2127–2143.
21. Mendelsohn ME (2005) In hypertension, the kidney is not always the heart of
the matter. J Clin Invest 115:840–844.
22. Nabokov AV, Amann K, Wessels S, Munter K, Wagner J, Ritz E (1999)
Endothelin receptor antagonists influence cardiovascular morphology in uremic
rats. Kidney Int. 55:512-519.
23. Nelson MT, Bonev AD (2004) The β1 subunit of the Ca2+-sensitive K+ channel
protects against hypertension. J Clin Invest 113:955-957.
24. Oberholzer M (1983) Schrumpfung und Dehnungsartefarkte. Morphometrie in
der klinischen Pathologie. Springer, Berlin Heidelberg New York.
25. Patterson AJ, Henrie-Olson J, Brenner R (2002 )Vasoregulation at the molecular
level: a role for the beta1 subunit of the calcium-activated potassium (BK)
channel. Trends Cardiovasc Med 12:78-82.

51
26. Plüger S, Faulhaber J, Fürstenau M, Löhn M, Waldschütz R, Gollasch M, Haller
H, Luft FC, Ehmke H, Pongs O (2000) Mice with disrupted BK channel beta1
subunit gene feature abnormal Ca2+ spark/STOC coupling and elevated blood
pressure. Circ Res 87:E53–E60.
27. Pluznick JL, Sansom SC (2006) BK channels in the kidney: role in K(+)
secretion and localization of molecular components. Am J Physiol Renal Physiol
291:F517–529.
28. Pluznick JL, Wei P, Carmines PK, Sansom SC (2003) Renal fluid and
electrolyte handling in BKCa-beta1 -/- mice. Am J Physiol Renal Physiol
284:F1274–1279.
29. Pluznick JL, Wei P, Grimm R, Sansom SC (2005) BK β1 subunit:
immunolocalization in the mammalian connecting tubule and its role in the
kaliuretic response to volume expansion. Am J Physiol Renal Physiol 288:F846-
854.
30. Sausbier M, Arntz C, Bucurenciu I, Zhao H, Zhou XB, Sausbier U, Feil S,
Kamm S, Essin K, Sailer CA, Abdullah U, Krippeit-Drews P, Feil R, Hofmann
F, Knaus HG, Kenyon C, Shipston MJ, Storm JF, Neuhuber W, Korth M,
Schubert R, Gollasch M, Ruth P (2005) Elevated blood pressure linked to
primary hyperaldosteronism and impaired vasodilation in BK channel-deficient
mice. Circulation 112:60-68.
31. Simpson LO (1980) A mouse model of spontaneous renal hypertension. blood
pressure, heart weight, kidney weight and proteinuria relationships in NZB x
OUW F1 hybrid female mice. Pathology 12:347-357.
32. Touyz RM (2000) Molecular and cellular mechanisms regulating vascular
function and structure - implications in the pathogenesis of hypertension. Can J
Cardiol 16:1137–1146.

52
33. Vèniant M, Heudes D, Clozel JP, Bruneval P, Ménard J (1994) Calzium
blockade versus ACE inhibition in clipped and unclipped kidneys of 2K-1C rats.
Kidney Int 46:421-429.
34. Weibel ER, Gomez DM (1962) A principle for counting tissue structures on
random sections. Appl Physiol 17:343.
35. Weibel ER (1979) Stereological methods, Vol1. Practical methods for
Biological Morphometry. Academic Press, London.
36. Weibel ER (1979) Stereological methods, Vol2. Practical methods for
Biological Morphometry. Academic Press, London.
37. Wilson PW (1999) From hypertension to heart failure: what have we learned?
Clin Cardio 22 Suppl 5:V1-10.

53
8 Abkürzungsverzeichnis
AA Flächenanteil
AR Nierenrindenfläche
Ca2 + Kalziumion
DNS Desoxyribonukleinsäure
GSI Glomeruloskleroseindex
HG Herzgewicht
IMA Intramyokardiale Arterien
KG Körpergewicht
ko knockout
LV linker Ventrikel
mNG mittleres Nierengewicht
NG Nierengewicht
NO Stickstoffmonoxid
NA Anzahl der Glomeruli pro Fläche Nierenrinde
NGlom Anzahl der Glomeruli einer Niere
Nv Anzahl der Glomeruli pro Volumen
n.s. nicht signifikant
p Irrtumswahrscheinlichkeit
PP Anzahl der gezählten Punkte
PAS Periocid-Acid-Schiff-Reaktion
PBS Phosphate-Buffered Saline
relHG relatives Herzgewicht
rmNG relatives mittleres Nierengewicht
RyR Ryanodin Rezeptor
RV rechter Ventrikel
S Septum (Herzseptum)
sign. signifikant
SR Sarkoplasmatischen Retikulum
STOCS spontaneous transient outward K+ currents
TSI Tubulointerstitieller Index
VDCC voltage dependant calcium channel, spannungsabhängiger
Kalziumkanal

54
VSI Vaskulärer Index
VSMC vascular smooth muscle, glatte Gefäßmuskulatur
VGlom Mittleres glomeruläres Volumen
VN Gesamtvolumen der Niere
VNGlom Gesamtvolumen der Glomeruli einer Niere
VR Absolutes Nierenrindenvolumen
VV Volumenanteil
VVGlom Volumendichte der Glomeruli in der Nierenrinde
VVM Volumenfraktion Nierenmark
VVR Volumenfraktion Nierenrinde
WD Wanddicke
WLR Wand-Lumen-Ratio
wt wildtyp

55
9 Danksagung
Mein Dank gilt Herrn Professor Dr. med. Arndt Hartmann, Direktor des Pathologischen
Institutes für die Überlassung des Themas der Arbeit und die Möglichkeit, diese in
seiner Abteilung durchführen zu dürfen.
Ganz besonders bedanken möchte ich mich bei meiner persönlichen Betreuerin, Frau
Professor Dr. med. Kestin Amann, die mich mit beispielloser Geduld, motivierender
Energie und hervorragenden Sachverstand schließlich doch zum Abschluss dieser
Arbeit geführt hat.
Großer Dank gilt meinem Komilliton und Freund Herrn Sergej Kronewald sowie den
medizinisch-technischen Laborassistentinnen und Assistenten, allen voran Frau Monika
Klewer und Herrn Stefan Söllner, für ihren unermüdlichen Beistand bei den
Laborarbeiten.
Mein inniger Dank gilt meiner lieben Frau Stephanie für ihre emotionale Unterstützung
und dauerhafte verständnisvolle Motivation.
Diese Arbeit möchte ich meiner Mutter Karin und meinem Vater Karl widmen, ohne
deren Liebe, Fürsorge und Weitsicht meine akademische Laufbahn nicht möglich
gewesen wäre.

56
10 Lebenslauf
Persönliche Daten
Name: Nußelt
Vorname: Matthias
Geburtsdatum und -ort: 09.01.1977 Dinkelsbühl
Konfession: evangelisch
Staatsangehörigkeit: deutsch
Familienstand: verheiratet
Schulbildung
1983 – 1987 Grundschule Dinkelsbühl
1978 – 1988 Hauptschule Dinkelsbühl
1988 – 1997 Gymnasium Dinkelsbühl
1997 Abschluss: Allgemeine Hochschulreife
Zivildienst
07/1997 – 07/1998 Kreiskrankenhaus Feuchtwangen
Hochschulbildung
1999 – 2005 Studium der Humanmedizin an der Friedrich-
Alexander-Universität Erlangen-Nürnberg
2001 Ärztliche Vorprüfung
2002 Erster Abschnnitt der Ärztlichen Prüfung
2004 Zweiter Abschnitt der Ärztlichen Prüfung
2005 Dritter Abschnitt der Ärztlichen Prüfung
29.04.2005 Approbation als Arzt
Berufliche Weiterbildung
Seit Juli 2005 Facharztweiterbildung Anästhesiologie am Klinikum Fürth.










![Bbstracts. - jnnp.bmj.com · ABSTRACTS [2] Studies in the pathological anatomyof ' Landry's paralysis' (Studienzur pathologischen Anatomie der 'Landryschen Paralyse ').-GRUNE- WALD](https://img.pdfslide.net/doc/110x75/5eabc93d3788374713509997/bbstracts-jnnpbmjcom-abstracts-2-studies-in-the-pathological-anatomyof-.jpg)


















