Embed Size (px)
Citation preview

1
1
Cholesterol metabolismand protein prenylation
Handout #3
2
Objectives• Recognize the different types of sterols• Understand cholesterol nutrition and the concept
of feedback inhibition• Know the functions of the isoprenoid biosynthetic
pathway• Know how cholesterol is synthesized• Understand the role of HMGCoA reductase and
how the statin drugs affect this enzyme• Understand the mechanism that regulates SREBP
and how cholesterol regulates this process• Understand how SREBP regulates cholesterol
synthesis• Understand the mechanism and function of protein
prenylation and the function of caveolae
99:163 - Handout #3 Lecturer - Weeks

2
3
KEY WORDS - Cholesterolmetabolism
sitosterol ergosterolSRE SREBPSCAP InsigHMG CoA reductase mevalonatestatin isopentenylpyrophosphatefarnesyl pyrophosphate geranylgeranylpyrophosphateCAAX box farnesylationgeranylgeranylation caveolaecaveolin squalene lanosterol 7-dehydrocholesterolΔ7-reductase
4
STRUCTURE OFCHOLESTEROL AND STEROLS
27 Carbons4 rings; A, B and C are 6-membered, D is 5-membered –
2 methyl groups – A/B (C19) and C/D (C18)One double bond in B-ring at C5; OH-group at C38 C saturated hydrocarbon chain attached to D-ring at C17
99:163 - Handout #3 Lecturer - Weeks

3
5
Non-animal sterols
Plant sterols are poorly absorbed
from the diet by humans
Ergosterol (yeast) Sitosterol (plants)
Sitosterolemia: this is a genetic deficiency that allowsexcess absorption of plant sterols- cause is amutation in an ABCG transporter leading to defectivesterol excretion.
6
Sources of Cholesterol
• Diet- cholesterol comes from foods derivedfrom animals
• Synthesis- Cholesterol is not an essentialnutrient, there is plenty of synthetic capacityto take care of nutritional needs. Howevertypically about half of the cholesterol is fromthe diet.
• The liver is the main site of synthesis butmany tissues can produce cholesterol. Thebrain seems to synthesize all of itscholesterol.
99:163 - Handout #3 Lecturer - Weeks

4
7
Cholesterol productionregulated
• Cholesterol serves as a feedbackinhibitor of the committed step incholesterol synthesis- the reactioncarried out by HMG-CoA reductase. Asdietary cholesterol is increased,endogenous synthesis of cholesterol inthe liver is decreased.
8
CHOLESTEROL --Modulates membranefluidity, precursor of hormones
1. Condensation of 3 CH3COO- to form mevalonate 2. Synthesis of isopentenyl pyrophosphate, activated isoprene unit (key building block).3. Condensation of 6 isopentenyl pyrophospate to form squalene.4. Squalene cyclizes, tetracyclic product converted into cholesterol.
Liver:synthesis
99:163 - Handout #3 Lecturer - Weeks

5
9
MEVALONATE SYNTHESIS: committed step enzyme: HMG-CoA reductase
Stage 1
HMG CoA synthase-
HMG CoA reductase-
10
MEVALONATE SYNTHESIS: committed step enzyme: HMG-CoA reductase
Stage 1
HMG CoA synthase-
HMG CoA reductase-
99:163 - Handout #3 Lecturer - Weeks

6
11
Major intermediates inCholesterol synthesis
C2 → C6 → [C5] → C30 → C27
Note: isoprene starting Point for many natural products (terpenes)Vitamins D, A, K, E
12
Stage 2:Squalene (C30) from 6 x C5
condensation reactions:C5 → C10 → C15 → C30
Geranyl - C10farnesyl - C15squalene - C30
using isopentenylpyrophosphate
99:163 - Handout #3 Lecturer - Weeks

7
13
STAGE 3 CYCLIZATION
Conversion to cholesterol :removal of 3 methylgroups, reduction of C=Cby NADPH, migration ofother double bond.
14
Lanosterol to Cholesterol
6 genetic diseases –mutations of 6 genesSterol substrate of themutated enzymeaccumulatesSevere birth defects –malformations,neurological defects,mental retardation
D7 Reductase deficiency– Smith-Lemli-OpitzSyndrome
99:163 - Handout #3 Lecturer - Weeks

8
15
Synthesized inliver; transportedby lipoproteins orstored in liver
16Champe et al.
99:163 - Handout #3 Lecturer - Weeks
9
17
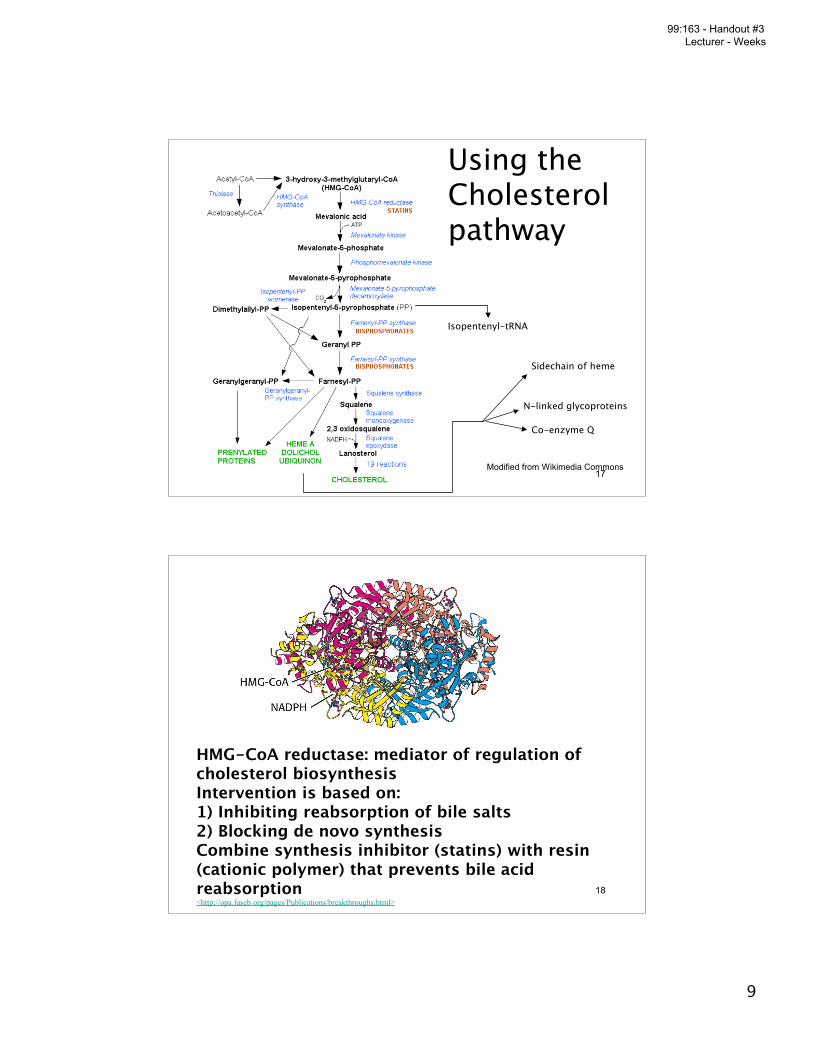
Using theCholesterolpathway
Isopentenyl-tRNA
Sidechain of heme
N-linked glycoproteins
Co-enzyme Q
Modified from Wikimedia Commons
18
HMG-CoA reductase: mediator of regulation ofcholesterol biosynthesisIntervention is based on:1) Inhibiting reabsorption of bile salts2) Blocking de novo synthesisCombine synthesis inhibitor (statins) with resin(cationic polymer) that prevents bile acidreabsorption<http://opa.faseb.org/pages/Publications/breakthroughs.html>
99:163 - Handout #3 Lecturer - Weeks

10
19
Cholesterol level management by HMG-CoAinhibitors: statins: competitive inhibitors
Lipitor
20
Regulation of Cholesterol SynthesisHMG-CoA reductase: rate limiting step
Regulation by:1) Transcriptional control. Sterol regulatory element-binding protein (SREBP) that binds to a Sterolregulatory element (SRE) upstream of the HMG-CoAreductase gene.2) Protein modification ( phosphorylation) - ReductaseKinase and HMG-CoA reductase phosphatase. Active inunmodified state.3). Hormonal regulation of protein modification-insulin cascade leads to greater activity, glucagon lessactivity.4). Rate of translation of mRNA (↓ by Chl & mevalonate)somewhat controversial.5). Degradation of the reductase (senses ↑ sterols)
99:163 - Handout #3 Lecturer - Weeks

11
21
MEVALONATE SYNTHESIS: committed step enzyme: HMG-CoA reductase
Stage 1
HMG CoA synthase-
HMG CoA reductase-
22
MEVALONATE SYNTHESIS: committed step enzyme: HMG-CoA reductase
Stage 1
HMG CoAsynthase-Thereare two differentforms of thisenzyme- one inthe cytosol andone in themitochondria .They are encodedby different genes
HMG CoA reductase-
99:163 - Handout #3 Lecturer - Weeks

12
23
Mitochondrial shuttling
Lehninger Biochemistry 5th ed.
24
Side effects of Statins - muscle pain and weakness
1.SLCO1B1 variants and Statin-induced Myopathy- A Genomewide studyNEJM August 21, 2008.Background: Lowering low-density lipoprotein cholesterol with statintherapy results in substantial reductions in cardiovascular events, andlarger reductions in cholesterol may produce larger benefits. In rare cases,myopathy occurs in association with statin therapy, especially when thestatins are administered at higher doses and with certain othermedications.Conclusions: We have identified common variants in SLCO1B1 that arestrongly associated with an increased risk of statin-induced myopathy.Genotyping these variants may help to achieve the benefits of statintherapy more safely and effectively.Note: SLCO1 encodes a transporting peptide that regulates statin uptake inthe liver. Muscle weakness side effect frequency increases with dose-article suggests that with normal doses incidence is 1/10,000.2. The muscle-specific ubiquitin ligase atrogin-1/MAFbx mediates statin-induced muscle toxicity.JCI December 2007. In patients with statininduced muscle pain atrogin is increased.
99:163 - Handout #3 Lecturer - Weeks

13
25
Regulation of HMG-CoA reductase
26
Linking transcription to cholesterol levels
99:163 - Handout #3 Lecturer - Weeks

14
27
Linking transcription to cholesterol levels
http://www4.utsouthwestern.edu/moleculargenetics/pages/brown/lab.html
There are at least 3 SREBPs-Two made by SREBP1 ( a and c) and SREBP2SREBP1a and SREBP2 are involved in cholesterolmetabolismSREBP1c in fatty acid synthesis
28
Linking transcription to cholesterol levels
http://www4.utsouthwestern.edu/moleculargenetics/pages/brown/lab.html
99:163 - Handout #3 Lecturer - Weeks

15
29
Linking transcription to cholesterol levels
http://www4.utsouthwestern.edu/moleculargenetics/pages/brown/lab.html
30
Linking transcription to cholesterol levels
http://www4.utsouthwestern.edu/moleculargenetics/pages/brown/lab.html
99:163 - Handout #3 Lecturer - Weeks

16
31
Linking transcription to cholesterol levels
http://www4.utsouthwestern.edu/moleculargenetics/pages/brown/lab.html
32
Linking transcription to cholesterol levels
http://www4.utsouthwestern.edu/moleculargenetics/pages/brown/lab.html
99:163 - Handout #3 Lecturer - Weeks

17
33
Linking transcription to cholesterol levels
http://www4.utsouthwestern.edu/moleculargenetics/pages/brown/lab.html
ER- endoplasmic reticulumInsig- ER retention protein- interact with SCAP preventing transit to golgi if sterol levels are sufficient, when
cholesterol levels fall in themembrane then the SCAP-SREPBcomplex can move to the gogi
SCAP- SREBP cleavage activating proteinS1P - Site 1 protease that makes the first
cut on SREBPS2P- Site 2 protease that make the second
cut on SREBP
34
Insig-1 and Insig-2
Insig-1 is binds to SCAP/SREBP1a and 2 complexes as longas there is adequate cholesterol in the ER membrane. Italso binds to the sterol sensing region of HMGCoAreductase if cholesterol levels are high enough, stimulatingthe ubiquitin mediated degradation of HMG CoA reductase.Insig-1 may also interact with NP-C.Protein and Patched, the receptor for Hedgehog. Insig-2associates with SCAP/SREBP1c has similar properties andparticular variants has been linked to obesity.
99:163 - Handout #3 Lecturer - Weeks

18
35
Phosphorylationand hormonalcontrol.
Figure and explanation come from: www.med.unibs.it/~marchesi/cholest.html
Hormones such as glucagon andepinephrine negatively affectcholesterol biosynthesis by increasingthe activity of the inhibitor ofphosphoprotein phosphatase-1, PPI-1.Conversely, insulin stimulates theremoval of phosphates and, thereby,activates HMG-CoA reductase activity.Additional regulation of HMG-CoAreductase occurs through an inhibitionof synthesis of the enzyme byelevation in intracellular cholesterollevels.
36
Isoprenylation of proteins
The protein signal is a CAAX box- refers tothe amino acid sequence cys-aliphatic-aliphatic-X where the “X” amino acid specificethe prenyl group. The prenyl groupsattached this way are geranylgeranyl andfarnesyl. The cys is important because it isthe attachment site. Proteins that areisoprenelated include the Rab family of Gproteins and the Ras oncogene.
99:163 - Handout #3 Lecturer - Weeks

19
37
Nature Reviews Cancer 5, 930-942 (December 2005)
a | Rho, which has the carboxy-terminalcysteine-aliphatic-aliphatic-X (CAAX) motif,is combined with FPP at the cysteine residueby farnesyl transferase. b | The CAAXprotease, Ras-converting enzyme 1 (RCE1),then cleaves off the AAX tripeptide from thecarboxy terminus of the protein at theendoplasmic reticulum membrane. c | Theisoprenylcysteine carboxyl methyltransferase(ICMT) enzyme then adds a carboxy methylgroup from S-adenosylmethionine (SAM) tothe prenylated cysteine residue. Rho, with itsnewly-attached hydrophobic anchor, thenmoves to the appropriate cellular membrane,remaining in the cytoplasmic compartment.
Isoprenylation of proteins
Isoprenylation of proteins is important incompartmentalization. Often isoprenylatedproteins have a second Cys that ispalmitoylated. Giving three hydrophobicreasons to associate with a membrane.
38
Cholesterol in the membrane
99:163 - Handout #3 Lecturer - Weeks

20
39
Membrane Lipids: Amphipathic, StructuralStructure determines “packing”, fluidity
PhospholipidsGlycolipidsCholesterol
40
Signaling molecules aredisproportionately in lipid rafts
99:163 - Handout #3 Lecturer - Weeks

21
41
The composition of lipid and protein in membranesis neither uniform nor random.
42
The association ofcaveolin with areas ofthe membrane lead tothe formation of pitlike structures calledcaveoli.
99:163 - Handout #3 Lecturer - Weeks

22
43
Summary
-Cholesterol is obtained from either the diet or bybiosynthesis from acetyl CoA.
-Foods containing animal fat are rich in cholesterol.
-Dietary cholesterol is absorbed well, but plant sterolsare poorly absorbed and very little plant sterol normally ispresent in the plasma or tissues.
-Cholesterol is one of the products of the isoprenoidbiosynthetic pathway. This pathway also synthesizesother important compounds necessary for glycoproteinsynthesis, tRNA function, mitochondrial electrontransport, and protein prenylation.
44
Summary
-The isoprenoid pathway enzyme that is regulated bycholesterol availability is HMG CoA reductase. It is therate-limiting step in cholesterol synthesis and the site offeedback inhibition, the process whereby cholesterolsynthesis is regulated by cholesterol availability.
-The statin drugs inhibit HMG CoA reductase.-Transcriptional regulation of the HMG CoA reductasegene is mediated by SREBP, a protein bound to theendoplasmic reticulum (ER) that is activated in responseto a decrease in the cell cholesterol content.
-Insig-1 is the cholesterol-sensing protein in the ER thatbinds to SCAP when there is enough cholesterol in theER membrane. It keeps the SREBP-SCAP complexbound to the ER.
99:163 - Handout #3 Lecturer - Weeks

23
45
Summary-When the ER cholesterol content decreases, Insig-1releases SCAP and the SREBP-SCAP complextranslocates to the Golgi where SREBP is cleaved by twoproteases. This releases the transcription factor domainof SREBP so it can enter the nucleus, bind to SRE in thepromoter of sterol responsive gene and thereby initiatethe expression of these genes.-Cholesterol synthesis proceeds from mevalonatethrough farnesyl pyrophosphate, squalene, lanosterol,and either 7-dehydrocholesterol or desmosterol tocholesterol.−Proteins can be prenylated, some with a farnesyl groupand others with a geranylgeranyl group. Thecomposition of the X in the CAAX box determines whichprenyl group is added. Prenylation targets these proteinsto specific domains in cell membranes.
46
Summary
キ Membranes contain cholesterol. It is embedded inthe lipid bilayer between phospholipids. The OH-group isoriented outward in the same region as the phospholipidhead groups, and the rings and hydrocarbon chainproject towards the center of the lipid bilayer and interactwith the phospholipid fatty acyl chains.キ Most of the membrane cholesterol in a cell ispresent in the plasma membrane.キ Caveolae are a specialized form of lipid rafts inmembranes that contain the protein caveolin. They arerich in cholesterol and sphingolipids. Prenylated proteinsare targeted to caveolae and are components of signaltransduction complexes.キ Caveolae also regulate intracellular cholesterolcontent, either redistributing cholesterol to the cell interioror facilitating its excretion from the cell.
99:163 - Handout #3 Lecturer - Weeks

24
47
CLINICAL CORRELATIONSHypercholesterolemia
This is a very common condition in which the plasma cholesterol concentration measured after anovernight period of fasting is elevated.
Plasma cholesterol is a component of plasma lipoproteins. Therefore, hypercholesterolemia is anelevation in lipoproteins that contain cholesterol. There are several forms ofhypercholesterolemia, depending on which plasma lipoprotein is elevated. A gene defect thatproduces an elevation in low density lipoproteins (LDL) is the most severe form.Hypercholesterolemia is a very complicated topic, and we will consider it in much more detailwhen we cover plasma lipoproteins.
Hypercholesterolemia is a serious risk factor for atherosclerosis, a disease of the arterial wall inwhich cholesterol accumulates and triggers an inflammatory reaction. Therefore,hypercholesterolemia requires treatment as soon as it is detected even though in can beasymptomatic for many years.
The cause of the hypercholesterolemia must be characterized before treatment is started in orderto know what type of treatment to prescribe. For example, one of the cholesterol-carryinglipoproteins is HDL (high density lipoproteins). HDL actually protects against atherosclerosis and,of course, cholesterol elevations due to HDL must not be treated.
The most serious cases of hypercholesterolemia are genetic in origin. They usually cause thehighest plasma cholesterol elevations and almost always require drug therapy. However, manycases in developed countries are diet induced. The diet in these cases contains too much saturatedfat and cholesterol, and the feedback inhibition of cholesterol biosynthesis is not enough toprevent the amount accumulating in the plasma from increasing. Other people who are eatingsimilar diets do not have hypercholesterolemia. Therefore, individuals who have diet-inducedhypercholesterolemia must have genetic differences from those of us who have similar diets butdo not develop cholesterol elevations. A number of candidate genes for diet-inducedhypercholesterolemia have been detected, but the cause has not yet been fully delineated. Theplasma cholesterol in the diet-induced form usually is only moderately elevated and often can beadequately controlled by dietary restriction, without the need for cholesterol-lowering drugs.
48
SitosterolemiaOnly trace amounts of plant sterols normally are absorbed from the diet, and the
amounts in the plasma and tissues are extremely small. In a rare genetic disease calledsitosterolemia, large amounts of dietary plant sterols are absorbed and accumulate in theplasma and tissues. Dr. William Connor discovered the disease in 1974 when he was a facultymember in Internal Medicine at the University of Iowa. Two sisters were referred to himbecause they had xanthomas. Xanthomas are cholesterol accumulations in the tendon sheaths,usually due to severe hypercholesterolemia. However, the sisters did not havehypercholesterolemia, at least according to the guidelines used in 1974.
Using gas-liquid chromatography, Dr. Connor discovered that the sisters had highlevels of plant sterols in their plasma and tissues. He correctly concluded that this was due to agenetic abnormality, and his further studies demonstrated that the mutation caused a muchhigher absorption of plant sterols from the diet.
Sitosterolemia is inherited as a recessive trait. We now know that the moleculardefect is caused by a mutation of an ABCG transporter, a membrane ATP cassette bindingprotein that utilizes the energy released by ATP hydrolysis to “pump” sterols out of theintestinal enterocytes so they are not absorbed and do not build up in the plasma. Normally,the ABCG transporters pump out almost all the plant sterols that are taken up from the diet bythe intestinal enterocytes, and only a very small amount remains in the body. Because the ABCGpumps are genetically defective in sitosterolemia, the absorbed plant sterols cannot beadequately excreted and accumulate in the plasma and tissues. There are many different formsof plant sterols, and in addition to sitosterol, other plant sterols also accumulate. They includecampesterol, stigmasterol and avenosterol. Like sitosterol, these plant sterols differ fromcholesterol only in the structure of the hydrocarbon chain attached to the steroid nucleus.
Sitosterolemia produces atherosclerosis and often leads to death from coronary heartdisease at middle age or even earlier. The atherosclerosis is due primarily to accumulation inthe arteries of cholesterol, not plant sterols, even though the plasma cholesterol is only slightlyto moderately elevated. The explanation is still uncertain. A likely possibility is that ABCGtransporters function to excrete excess cholesterol in addition to plant sterols. When thesetransporters are genetically defective, cholesterol cannot be adequately excreted from thetissues, and enough apparently accumulates in the arteries to produce atherosclerosis.
99:163 - Handout #3 Lecturer - Weeks

25
49
CLINICAL CORRELATIONS
Smith-Lemli-Opitz syndromeThe Smith-Lemli-Opitz syndrome occurs in 1/20,000 live births in North AmericanCaucasians and has a carrier frequency of 2 %. Therefore, it is a fairly common geneticdisease. It causes severe developmental abnormalities, including microcephaly, pituitaryagenesis, midline facial anomalies, and defects in many internal organs. The disease alsoproduces neurological dysfunction and severe mental retardation. Interestingly, Dr. Opitzwas a faculty member in Neurology at the University of Iowa.
The cause of the Smith-Lemli-Opitz syndrome is a mutation in the sterol D7-reductasegene. The sterol D7-reductase operates in the isoprenoid synthesis pathway betweenlanosterol and cholesterol. Because of this genetic defect, cholesterol synthesis cannotoccur. Instead, there is an accumulation of two sterols that are substrates for this enzyme,7-dehydrocholesterol and cholest-5, 7, 24-trien-3b-ol .
A mother carrying a Smith-Lemli-Opitz syndrome fetus provides adequate amounts ofplasma cholesterol to the fetal circulation through the placenta, but this apparently doesnot compensate for the inability of the fetal tissues to synthesize cholesterol. It isuncertain whether the accumulation of 7-dehydrocholesterol and cholest-5, 7, 24-trien-3b-ol, or the failure to synthesize cholesterol, causes the Smith-Lemli-Opitz syndromeabnormalities.
Attempts have been made to treat Smith-Lemli-Opitz syndrome patients by feeding themvery large amounts of cholesterol. The rationale for this approach is two-fold. One is toreplace the cholesterol that cannot be synthesized. The other is to down-regulate HMG CoAreductase through feedback inhibition and thereby reduce the production of 7-dehydrocholesterol and cholest-5, 7, 24-trien-3b-ol. While there are a few reportsindicating some improvement, the results have not been overly successful. This probably isdue to the fact that most of the severe abnormalities are imprinted in utero, and beginningtreatment after the child is born is far too late.
99:163 - Handout #3 Lecturer - Weeks