Embed Size (px)
Citation preview

Plant Ecology 131: 233–239, 1997. 233c 1997 Kluwer Academic Publishers. Printed in Belgium.
Clonal plasticity in response to nutrient availability in the pseudoannualherb, Trientalis europaea L.
Ming Dong1;2, Heinjo J. During1 & Marinus J. A. Werger1
1Department of Plant Ecology and Evolutionary Biology, Utrecht University, PO Box 800.84, 3508 TB Utrecht,The Netherlands; 2Institute of Botany, The Chinese Academy of Sciences, Beijing, 100093, The People’s Republicof China
Received 2 October 1996; accepted in revised form 4 March 1997
Key words: Clonal plant, Foraging, Hibernacle, Rhizome, Size-number trade-off
Abstract
Trientalis europaea plants were subjected to four levels of nutrient availability to test two hypotheses: (1) Clonalmorphology of pseudoannuals with short-lived rhizomes is responsive to nutrient availability in terms of spacerlength (a negative correlation expected) and branching intensity (a positive correlation expected), and (2) the sizeof the hibernacles of T. europaea shows a positive correlation with nutrient availability. The results support thefirst hypothesis, since the primary spacers of T. europaea were longer at lower nutrient levels while the branchingintensity of the primary rhizomes increased at increasing nutrient supply. The second hypothesis was not confirmed,however; the plants produced fewer, but bigger hibernacles at lower levels of nutrient availability. The ecologicalsignificance of the results is discussed in the context of foraging for essential resources and habitat-related effectsof hibernacle size on survival and establishment chances of daughter ramets.
Introduction
Many clonal plants are able to respond to variation inenvironmental conditions by altering their clonal mor-phology in terms of spacer length (sensu Bell 1984) andbranching (Hutchings & de Kroon 1994) as well as bymodifying their clonal reproduction in terms of size andnumber of newly produced ramets (Slade & Hutchings1987; Hutchings & Slade 1988). This plasticity maybe adaptive and may thus contribute to the spreading ofthe risk of genet extinction (Eriksson & Jerling 1990)as well as to foraging for essential resources in hetero-geneous environments (Hutchings & de Kroon 1994;Dong & de Kroon 1994).
Clonal plasticity and type of clonal growth (e.g.,stoloniferous and/or rhizomatous) may differ betweenspecies (Leakey 1981; de Kroon & Hutchings 1995),and they may to some extent be correlated to eachother (Dong & Pierdominici 1995; Dong et al. 1996).Based on the longevity of spacers (Bell 1984), twogroups of clonal plants can be distinguished: ‘Genet-splitters’ (Eriksson & Jerling 1990) with short-lived
spacers and genets with long-lived spacers (Pitelka& Ashmun 1985). Recent multi-species comparisons(Dong & Pierdominici 1995; Dong et al. 1996) showthat long-lived rhizomes are unresponsive to nutrientavailability in terms of their total lengths and inter-node lengths. This has been interpreted as an indica-tion that the rhizomes primarily serve as storage organsfor meristems and carbohydrates (Dong & de Kroon1994). However, this may not hold for rhizomatousgenet-splitters. Their rhizomes live so briefly (usuallynot longer than one growing season) that they may notbe able to act as storage organs for resources and/ormeristems. Instead, they probably function as organsof spatial placing of newly produced ramets (foragingsensu Hutchings & de Kroon 1994; Dong & de Kroon1994). Based on the model results of Sutherland &Stillman (1988) it may be hypothesized that the lengthof these short-lived rhizomes is negatively correlatedwith nutrient supply, while their branching intensitywill show a positive correlation.
Some ‘genet-splitters’ are pseudoannuals (Salis-bury 1942) whose ramets and spacers both live for
Gr.: 201002978 , PIPS Nr. 136405 BIO2KAP
*136405 veg10661.tex; 23/06/1997; 7:41; v.7; p.1
234
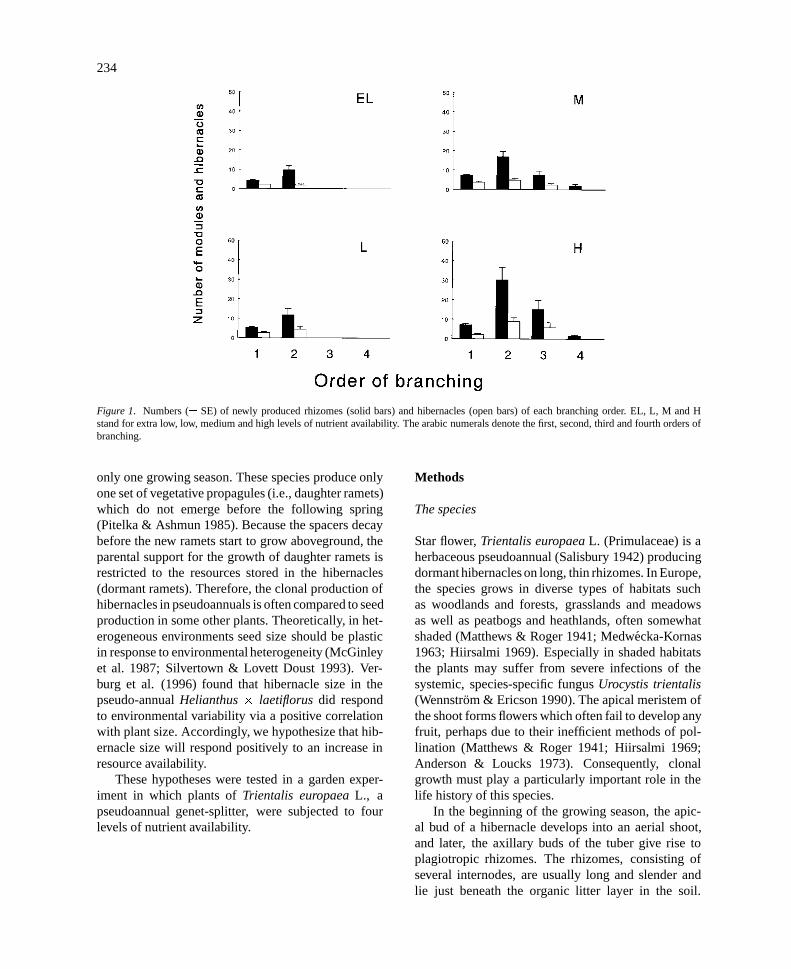
Figure 1. Numbers (� SE) of newly produced rhizomes (solid bars) and hibernacles (open bars) of each branching order. EL, L, M and Hstand for extra low, low, medium and high levels of nutrient availability. The arabic numerals denote the first, second, third and fourth orders ofbranching.
only one growing season. These species produce onlyone set of vegetative propagules (i.e., daughter ramets)which do not emerge before the following spring(Pitelka & Ashmun 1985). Because the spacers decaybefore the new ramets start to grow aboveground, theparental support for the growth of daughter ramets isrestricted to the resources stored in the hibernacles(dormant ramets). Therefore, the clonal production ofhibernacles in pseudoannuals is often compared to seedproduction in some other plants. Theoretically, in het-erogeneous environments seed size should be plasticin response to environmental heterogeneity (McGinleyet al. 1987; Silvertown & Lovett Doust 1993). Ver-burg et al. (1996) found that hibernacle size in thepseudo-annual Helianthus � laetiflorus did respondto environmental variability via a positive correlationwith plant size. Accordingly, we hypothesize that hib-ernacle size will respond positively to an increase inresource availability.
These hypotheses were tested in a garden exper-iment in which plants of Trientalis europaea L., apseudoannual genet-splitter, were subjected to fourlevels of nutrient availability.
Methods
The species
Star flower, Trientalis europaea L. (Primulaceae) is aherbaceous pseudoannual (Salisbury 1942) producingdormant hibernacles on long, thin rhizomes. In Europe,the species grows in diverse types of habitats suchas woodlands and forests, grasslands and meadowsas well as peatbogs and heathlands, often somewhatshaded (Matthews & Roger 1941; Medwecka-Kornas1963; Hiirsalmi 1969). Especially in shaded habitatsthe plants may suffer from severe infections of thesystemic, species-specific fungus Urocystis trientalis(Wennstrom & Ericson 1990). The apical meristem ofthe shoot forms flowers which often fail to develop anyfruit, perhaps due to their inefficient methods of pol-lination (Matthews & Roger 1941; Hiirsalmi 1969;Anderson & Loucks 1973). Consequently, clonalgrowth must play a particularly important role in thelife history of this species.
In the beginning of the growing season, the apic-al bud of a hibernacle develops into an aerial shoot,and later, the axillary buds of the tuber give rise toplagiotropic rhizomes. The rhizomes, consisting ofseveral internodes, are usually long and slender andlie just beneath the organic litter layer in the soil.
veg10661.tex; 23/06/1997; 7:41; v.7; p.2

235
Apical buds of the rhizomes may form orthotropichibernacles. At each rhizome node there is only oneaxillary bud, which may or may not develop into anew rhizome. This causes a sympodial frameworkof rhizomes of several branching orders. Within onegrowing season, an individual plant usually consistsof only one aboveground shoot, which is producedby the parent hibernacle and which produces a num-ber of daughter hibernacles which are interconnectedby string-like rhizomes (spacers). During the autumn,the parent shoot as well as the whole rhizome systemconnecting the daughter hibernacles die off (Matthews& Roger 1941), consequently leaving the newly pro-duced daughter hibernacles free in the soil. The daugh-ter hibernacles, which may have their own small rootsystems, usually hibernate and keep dormant until thenext growing season.
Experimental design
In February 1991, plant material (hibernacles) wasobtained from a small population in a pine forest under-storey in a part of the Botanical Garden near Utrecht,the Netherlands. The experiment was conducted insidea shade house at the Experimental Garden at De Uithof,Utrecht, the Netherlands. It ran from March 1 to August3, 1991. The shade cloth reduced photosynthetic act-ive radiation (PAR) to 40% of full daylight and didnot significantly alter the ratio of red to far-red light(R/FR). The hibernacles were grown in pots (50 cmin diameter and 30 cm high) filled with river sand andsubjected to four levels of nutrient availability. Thehigh nutrient level (H) was equivalent to 200 kg nitro-gen as NH4NO3, 60 kg phosphorus as Na2PO4
�2H2Oand 200 kg potassium as KCl per hectare and year. Themedium (M), low (L), and extra low (EL) levels were50%, 25% and 10% of the high level, respectively.The nutrient solutions used were similar in volumeand differed in the concentration of the nutrients. Thenutrient solutions were supplied twice a month. Dur-ing the experiment the plants were watered as needed.Sixteen replicate plants per treatment were employed.
Harvesting and analyses
The plants were harvested after five months of growth.Each plant was separated into individual modules(products of an apical meristem) according to theirbranching order. For each module, rhizome length, andnumber of internodes were measured. Afterwards, themodules were individually separated into rhizome,hib-
Figure 2. (A) Plant dry weight (� SE) and (B) percentage biomassallocation (� SE) to roots (solid portion), hibernacles (shaded por-tion), rhizomes producing hibernacles (hatched portion), rhizomeswithout hibernacles (cross-hatched portion) and leaves (open por-tion), in Trientalis europaea. For the same portion, bars sharing thesame letter are not significantly different at P = 0:05. Code ofnutrient levels as in Figure 1.
ernacles, roots, leaves, shoot stem and flower. The dryweights of different plant components were determinedafter drying at 75 �C for at least 48 h. From the meas-urements, the following three groups of traits werederived. (1) Plant growth and biomass allocation: plantdry weight as well as percentage biomass allocated todifferent plant components. (2) Clonal morphology:length, internode length and specific length of individu-al primary and secondary spacers (spacers are rhizomeswhich terminate in hibernacles; specific spacer lengthis the length of a spacer divided by the weight ofthe spacer) as well as branching intensity (number ofrhizome branches from a rhizome divided by num-ber of nodes of the rhizome). (3) Clonal reproduction:number of hibernacles per plant and dry weight perhibernacle.
veg10661.tex; 23/06/1997; 7:41; v.7; p.3

236
The data were statistically analyzed using one-wayANOVA with Tukey-grouping (SAS 1985). Beforethe analyses, the data have been subjected to log-transformation. Very few plants in the present exper-iment flowered and therefore flowers were excludedfrom the analysis.
Results
Plant growth and biomass allocation
At the end of the experiment, Trientalis europaeaplants under the four conditions had formed onlyone aboveground shoot from the initial hibernaclesand produced many new rhizomes and new hiber-nacles without aboveground shoots. Most of the newlyproduced hibernacles rooted. Of the newly producedrhizomes, some did not form hibernacles at theirends and the amount of that type of rhizomes seemedto increase with increasing nutrient availability (Fig-ure 1).
The plants accumulated more biomass in terms ofdry weight at the H nutrient level than at the EL andL levels (Figure 2A). Percentage biomass allocationto leaves was similar among plants at different levelsof nutrient supply. Allocation to roots tended to besmaller at higher levels of nutrient supply but differedsignificantly only between the plants at the H level andthe plants at the EL and L levels (Figure 2B). Therewas no significant difference in allocation to hiber-nacles among the treatments, although it tended to besmaller at higher levels of nutrient availability. Theplants allocated a greater proportion of biomass to pro-duce significantly more rhizomes without hibernaclesat higher levels of nutrient supply while the percent-age biomass allocation to rhizomes with hibernacleswas similar between the treatments. As a result, theplants tended to allocate a greater proportion of bio-mass to total rhizomes as the nutrient supply increased,although it was significantly different only between theplants growing at the M and H levels and those growingat the EL and L levels.
Clonal morphology
There was a tendency that Trientalis europaea plantsproduced longer individual primary spacers (Fig-ure 3A) with longer internodes (Figure 3B) at lowernutrient levels, although this was significantly differ-ent only between plants at the EL as against the H
nutrient levels. Specific rhizome length did not signi-ficantly respond to the treatments (Figure 3C). Branch-ing intensity of the primary rhizomes was lower at theEL and L than at the M and H nutrient levels (Fig-ure 3D). All investigated morphological traits of thesecondary rhizomes did not significantly respond tothe treatments (Figure 3).
Clonal reproduction
At a higher level of nutrient supply, the plants tendedto produce more hibernacles; differences were signi-ficant between the plants growing at the H level andthose growing at the EL and L levels of nutrient supply(Figure 4A). In contrast, the dry weight per hibernacletended to be less at higher levels of nutrient supply,with differences significant between the plants grow-ing at the EL level and those growing at the H nutrientlevels (Figure 4B).
Discussion
Clonal morphology
In the present experiment, particularly the branchingintensity of the primary rhizomes expressed high levelsof plasticity; those at the H nutrient level were 105%more intensely branched than those at the EL level(Figure 3D). Furthermore, primary spacers formed atthe H level were more than 50% shorter than thoseformed at the EL level. The traits investigated forthe secondary spacers did not significantly respondto the treatments, perhaps due to the development-al immaturity of the secondary and the higher orderrhizomes at harvest date (cf. Hutchings & de Kroon1994). These results are consistent with the expect-ation that both branching intensity and spacer lengthwould be responsive to the nutrient supply.
The behaviour expressed by T. europaea is consid-erably different from that found for many rhizomatousperennials whose rhizome lengths are usually unre-sponsive to nutrient availability (Dong & Pierdomin-ici 1995; de Kroon & Hutchings 1995; Dong et al.1996). Data available to date (de Kroon & Hutchings1995) indicate that among eleven rhizomatous peren-nials investigated only Brachypodium pinnatum andCarex bigelowii are responsive to nutrient availabil-ity (de Kroon & Knops 1990; Carlsson & Callaghan1990), but with much lower degrees of response, as
veg10661.tex; 23/06/1997; 7:41; v.7; p.4

237
Figure 3. (A) Individual spacer length (� SE), (B) spacer internode length (� SE), (C) specific spacer length (� SE) and (D) branchingintensity (� SE) of the first (solid bars) and second (open bars) orders of branches in Trientalis europaea. Code of nutrient levels as in Figure 1.In each panel and branching order, bars sharing the same letter are not significantly different at P = 0:05.
compared with that of T. europaea in the present exper-iment.
In many types of habitat, such as coniferous wood-lands, peatbogs and grasslands, where T. europaeaoccurs, nutrient availability is likely to be distributedin a heterogeneous manner (Grime 1979, 1994). Thestrong morphological responses expressed by T. euro-paea in the experiment provide evidence that this plantpossesses the potential to selectively place the newlyformed ramets in nutrient-rich patches in the habitats aspredicted by the model study of Sutherland & Stillman(1988). Such selective placement of newly producedramets may contribute to spreading of the risk of genetextinction (Eriksson & Jerling 1990) as well as to anincreased acquisition of patchily distributed nutrients(foraging, Hutchings & de Kroon 1994; Dong & deKroon 1994).
Clonal reproduction
At higher nutrient levels the plants produced more, butsmaller hibernacles. Thus, hibernacle size responds
plastically to variation in habitat quality, but the direc-tion of the response does not conform to our hypothesis.This is remarkable, since many other clonal plants pro-duce larger offspring ramets at higher resource levels(Slade & Hutchings 1987; Dong et al. 1996; Verburget al. 1996), but it does correspond to the assumptionsabout the effect of resource availability on seed sizein McGinley et al. (1987). Using a similar argumentas these authors did, we hypothesize that the minimalhibernacle size necessary for successful establishmentin the next year will be higher in resource-poor envir-onments. Then, a trade-off between number and sizeof hibernacles produced may favour the production ofmore, smaller hibernacles under resource-rich condi-tions. This, together with the shorter spacers under suchconditions, will lead to locally higher shoot densitiesof T. europaea in nutrient-rich patches. A higher shootdensity might also be advantageous through increasedattractiveness for pollinators because of a higher flowerdensity.
It is not known whether the changes in spacer lengthand hibernacle size were a response to the local condi-
veg10661.tex; 23/06/1997; 7:41; v.7; p.5

238
Figure 4. (A) Number of hibernacles per plant (� SE) and (B)dry weight per hibernacle (� SE) in Trientalis europaea. In eachpanel, bars sharing the same letter are not significantly different atP = 0:05. Code of nutrient levels as in Figure 1.
tions experienced by the growing rhizome apices, or tothe conditions experienced by the mother ramet. Dueto the uniform soil environment used in this study wecannot answer this question, but future experiments inthis direction would be rewarding.
Acknowledgements
The authors are very grateful to Hans de Kroon fordiscussion during the experiment and the data analysis.We would also like to thank Vijko Lukkien for guidingthe collection of the plant materials.
References
Anderson, R. C. & Loucks, O. L. 1973. Aspects of the ecology ofTrientalis borealis Raf. Ecology 54: 798–808.
Bell, A. D. 1984. Dynamic morphology: a contribution to plantpopulation ecology. Pp. 48–65. In: Dirzo, R. & Sarukhan, J. (eds),Perspectives on plant population ecology. Sinauer, Sunland.
Carlsson, B. A. & Callaghan, T. V. 1990. Programmed tiller differ-entiation, intraclonal density regulation and nutrient dynamics inCarex bigelowii. Oikos 58: 219–230.
de Kroon, H. & Hutchings, M. J. 1995. Morphological plasticityin clonal plants: The foraging concept reconsidered. Journal ofEcology 83: 143–152.
de Kroon, H. & Knops, J. 1990. Habitat exploration through mor-phological plasticity in two chalk grassland perennials. Oikos 59:39–49.
Dong, M. & de Kroon, H. 1994. Plasticities in morphology andbiomass allocation in Cynodon dactylon, a grass species formingstolons and rhizomes. Oikos 70: 99–106.
Dong, M., During, H. J. & Werger, M. J. A. 1996. Morphologicalresponses to nutrient availability in four clonal plants. Vegetatio123: 183–192.
Dong, M. & Pierdominici, M. G. 1995. Morphology and growthof stolons and rhizomes in three grass species, as affected bydifferent light supply. Vegetatio 116: 25–22.
Eriksson, O. & Jerling, L. 1990. Hierarchical selection and riskspreading in clonal plants. Pp. 79–94. In: van Groenendael, J.& de Kroon, H. (eds), Clonal growth in plants: regulation andfunction. SPB Academic Publishing, The Hague.
Grime, J. P. 1979. Plant strategies and vegetation processes. JohnWiley and Sons, Chichester.
Grime, J. P. 1994. The role of plasticity in exploiting environmentalheterogeneity. Pp. 1–20. In: Caldwell, M. & Pearcy, R. W. (eds),
Exploitation of environmental heterogeneity by plants. AcademicPress, San Diego.
Hiirsalmi, H. 1969. Trientalis europaea L. A study of the reproduct-ive biology, ecology and variation in Finland. Ann. Bot. Fennici6: 119–173.
Hutchings, M. J. & de Kroon, H. 1994. Foraging in plants: the roleof morphological plasticity in resource acquisition. Adv. Ecol.Res. 25: 159–238.
Hutchings, M. J. & Slade, A. J. 1988. Morphological plasticity,foraging and integration in clonal perennial herbs. Pp. 83–109.In: Davy, A. J., Hutchings, M. J. & Watkinson, A. R. (eds), Plantpopulation ecology. Symposia of the British Ecological Society28. Blackwell Scientific Publications, Oxford.
Leakey, R. R. B. 1981. Adaptive biology of vegetatively regeneratingweeds. Adv. Appl. Biol. 6: 57–90.
Matthews, J. R. & Roger, J. G. 1941. Variation in Trientalis europaeaL. J. Bot. 79: 80–83.
McGinley, M. A., Temme, D. H. & Geber, M. A. 1987. Parentalinvestment in offspring in variable environment: theoretical andempirical considerations. Am. Natur. 130: 370–389.
Medwecka-Kornas, A. 1963. Observations on the variability of Tri-entalis europaea L. in Finland, Norway and Poland. Berichte desGeobotanischen Institut ETH, Stiftung Rubel, 34: 28–37.
Pitelka, L. F. & Ashmun, J. W. 1985. Physiology and integration oframets in clonal plants. Pp. 399–435. In: Jackson, B. C., Buss,L. W. & Cook, R. E. (eds), Population biology and evolution ofclonal organisms. Yale University Press, New Haven.
Salisbury, E. J. 1942. The reproductive capacity of plants. Bell,London.
SAS, 1985. SAS/STAT guide for personal computers version 6 Edi-tion. SAS Institute Inc., Cary.
Silvertown, J. W. & Lovett Doust, J. 1993. Introduction to plantpopulation biology. Blackwell Scientific Publications, Oxford.
Slade, A. J. & Hutchings, M. J. 1987. The effects of nutrient availab-ility on foraging in the clonal herb Glechoma hederacea. J. Ecol.75: 95–112.
Sutherland, W. J. & Stillman, R. A. 1988. The foraging tactics ofplants. Oikos 52: 239–244.
veg10661.tex; 23/06/1997; 7:41; v.7; p.6

239
Verburg, R. W., Kwant, R. & Werger, M. J. A. 1996. Does plant sizeaffect vegetative reproduction in a pseudoannual? Vegetatio 125:185–192.
Wennstrom, A. & Ericson, L. 1990. The interaction between theclonal herb Trientalis europaea and the host specific smut fungusUrocystis trientalis. Oecologia 85: 238–240.
veg10661.tex; 23/06/1997; 7:41; v.7; p.7