Embed Size (px)
Citation preview

Vol. 254, No 5, Issue of March 10, pp. 1707-1715, 1979 Printed in U.S A.
Comparisons of Soybean Urease Isolated from Seed and Tissue Culture*
(Received for publication, May 10, 1978, and in revised form, September 15, 1978)
Joseph C. PolaccoS and Evelyn A. Havir
From the Departments of Genetics and Biochemistry, The Connecticut Agricultural Experiment Station, New Haven, Connecticut 06504
Urease was purified 500-fold to electrophoretic ho- mogeneity from ground, dry soybeans. Sodium dodecyl sulfate-acrylamide gel electrophoresis indicates a sub- unit size of 93,500 daltons which is identical with that of jack bean urease. In solutions of high ionic strength, there exists a single urease species (species 1) with a size of about 480,000 daltons based on agarose column chromatography and migration in acrylamide gels. In solutions of lower ionic strength (e.g. 3 nm phosphate), a new lighter species (species 2) predominates which has a size of about 280,000 daltons. Soybean and jack bean urease are serologically related but also contain unique antigenic determinants. The amino acid com- position profiles of soybean and jack bean urease show only small differences in the amounts of four amino acids. The urease activity purified from soybean cell suspension cultures was electrophoretically identical with the seed enzyme (both species 1 and species 2 being present in buffers of intermediate ionic strength, viz. 10 mM phosphate). Antibodies to soybean seed urease were purified by affinity chromatography. They were then separated from cross-reacting antibodies to jack bean urease by chromatography over Sepharose 4B containing covalently linked jack bean urease. The effluent antibodies no longer precipitated nor inhibited jack bean urease but inhibited ureases from soybean seed and soybean tissue culture to the same extent. In spite of this immunological evidence for a detailed sim- ilarity between soybean seed urease and urease par- tially purified from suspension culture, urease activity in crude extracts of suspension culture is heteroge- neous both with respect to size and affinity for mono- specific antiseed urease antibodies.
Urease is a seed protein common to most of the Legumi- nosae. Jack bean urease, the first enzyme to be isolated to crystalline purity (l), has a high methionine content (2, 3). Generally in legume seed proteins, this amino acid is present in limiting amounts for human and monogastric animal nutri- tion (4). If the methionine content of soybean urease is similar to that of jack bean, breeding for high seed urease levels may lead to increases in seed methionine levels.
In this report, we examine the similarity between soybean and jack bean urease and between the ureases of soybean seed and soybean tissue culture. Soybean seed urease was purified approximately 500-fold to electrophoretic homogeneity. Soy-
* This work was supported in part by Grant PCM 78-18501 from the National Science Foundation. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisement” in accord- ance with 18 USC. Section 1734 solely to indicate this fact.
$ To whom correspondence should be addressed.
bean and jack bean urease are serologically related and share a common subunit size as determined by SDS’-polyacryl- amide electrophoresis. Moreover, the amino acid profiles, including methionine content, are very similar. Urease from each source can exist in different interconvertible polymeric forms. By the criteria of electrophoretic mobility, gel filtration patterns, and inactivation by antibodies to seed urease, the purified urease of soybean seed and partially purified urease from suspension culture were identical. However, urease ac- tivity in crude extracts of suspension culture showed apparent size and immunological heterogeneity.
MATERIALS AND METHODS
Urease Assay and Definition of Unit-Urease activity was deter- mined by incubating appropriate volumes of enzyme preparations (5 to 100 ~1) in 1 ml of 0.5 M urea (ultrapure, Bethesda Research Laboratories) in 0.1 M Tris/maleate, 1 mM EDTA, 1 mM P-mercap- toethanol, pH 7.0 (Buffer C). At times of 15 s to 5 min, % volume of 2 N H2S04 was added. A O.l-ml aliquot of the acid-stopped reaction mix was added to 5.0 ml of Hz0 and followed by the addition of 0.1 ml of Nessler’s reagent (ammonia color reagent, Sigma). Ammonia was measured by the absorbance at 436 nm after allowing color to develop for 10 min. When crude seed extracts were assayed, it was necessary to, centrifuge down proteins from acid-stopped incubations before determining ammonia. Ammonium sulfate standards, prepared in Buffer C and mixed with ‘/z volume of 2 N H804, were usually assayed concurrently.
Column fractions were assayed most conveniently by incubating 5 d of column effluent with 0.2 ml of 0.5 M urea in Buffer C, after which 0.1 ml of 2 N H,SO, was added to stop the reaction. Five milliliters of HZ0 and 0.3 ml of Nessler’s reagent were added in succession and ammonia was determined as above. All assays were performed at 30°C and pH 7.0. A unit of urease hydrolyzes 1 pmol of urea/min at pH 7.0 and 30°C.
Purification of Soybean Urease-Except where noted, all steps were performed at 4°C. Two pounds of whole dry soybeans (Prize, Burpee Seed Co.) were ground in a Wiley Mill No. 1 (Arthur H. Thomas) and added to 6 liters of 0.1 M Tris/maleate, 30 mM sodium EDTA, 10 mM /I-mercaptoethanol, pH 7.6 (Buffer A). The slurry was stirred for 3 h and left to stand at 4°C overnight. It was then strained through two layers of cheesecloth and the extruded fluid was centri- fuged at 16,000 X g for 10 min. The supernatant was poured through two layers of cheesecloth to remove some of the lipid.
The resulting crude extract (4 liters) was heated in a 60°C waler bath for 1 h (1300 ml/2-liter glass beaker). Following cooling to about 3O”C, solid ammonium sulfate was added to 20% saturation. After centrifugation at 16,000 x g, the pellet was discarded and the super- natant made 55% saturated in ammonium sulfate. Protein was pel- leted at 16,000 X g for 10 min, resuspended in about 500 ml of 0.1 M
Tris/maleate, 1 mM EDTA, 10 IIIM /&mercaptoethanol, pH 7.0 (Buffer B), and dialyzed against four batches (4 liters) of Buffer B. Denatured protein was removed by centrifugation and % volume of acetone (-20°C) was slowly added to the supernatant with stirring. Precipi- tated protein was removed by centrifugation and % volume of acetone was added to the supernatant as described above. After centrifugation,
-
I The abbreviations used are: SDS, sodium dodecyl sulfate, CNBr, cyanogen bromide.
1707
by guest on July 15, 2018http://w
ww
.jbc.org/D
ownloaded from

1708 Urease in Soybean Seed and Cultured Cells
the pellet was resuspended in approximately 100 ml of 0.1 M Tris/ maleate, 1 IIIM EDTA, 1 mM ,&mercaptoethanol, pH 7.0 (Buffer C) and dialyzed against 1 liter of Buffer C overnight. Denatured protein was removed by centrifugation and the supernatant was diluted to approximately 300 ml with Buffer C and applied to a column (2 x 45 cm) of DEAE-cellulose (Sigma) prepared as described by Peterson and Sober (5) and equilibrated with Buffer C.
A linear gradient (total volume, 1100 ml) of 0.1 to 0.4 M Tris/ maleate was applied to the DEAE-cellulose column and 6.3-ml frac- tions were collected. Urease activity eluted between 515 and 725 ml at approximately 0.25 M Tris/maleate. Fractions containing at least 10 units/ml of urease were pooled and made 55% saturated in am- monium sulfate. Precipitated protein was pelleted and resuspended in 5 ml of 10 mM potassium phosphate, 1 mM EDTA, and 1 ITIM ,8-
mercaptoethanol, pH 7.0. After overnight dialysis against 2 liters of the same buffer, insoluble protein was removed by centrifugation and the supernatant was applied to a column (2 x 45 cm) of hydroxylap- atite (Hypatite-C, Clarkson Chemical Co.). Alternatively, pooled DEAE-cellulose fractions were dialyzed against 2 P-liter volumes of 10 mM potassium phosphate, 1 mM EDTA, 1 IIIM ,&mercaptoethanol, pH 7.0, and applied directly to the hydroxylapatite column. The enzyme was eluted with a linear gradient of 10 to 300 mM potassium phosphate (total volume, 800 ml; 4.3-ml fractions). Peak urease activ- ity eluted with 70 to 80 mM phosphate in different preparations. Fractions containing at least 20 units/ml and free of a late eluting major contaminant protein (present in amounts roughly equal to urease protein) were pooled and concentrated with 55%~ ammonium sulfate. The remaining urease-containing fractions were pooled, di- alyzed against 10 mM phosphate, reapplied to the hydroxylapatite column and the enzyme was eluted with a 10 to 250 mM phosphate gradient (800 ml). Fractions of specific activity 375, or over, were pooled. The urease recovered from this recycling through hydroxyl- apatite was roughly 25% of that obtained in the first pass through hydroxylapatite.
Hydroxylapatite fractions from both passes were concentrated by the addition of (NH,)SO, to 55% saturation and then applied (total volume, ~4 ml) to a column (4 x 45 cm) of agarose A-15m (Bio-Rad) equilibrated with 10 mM KPO,, 1 mM EDTA, 1 mM P-mercaptoetha- nol, pH 7.0. The peak fractions contained electrophoretically pure urease. Pooled fractions were stored at 4°C in the presence of 0.02% NaN.I. When necessary, enzyme was concentrated with ammonium sulfate or by dialysis against polyethylene glycol (Aquacide III, Cal- biochem).
The purification is greatly simplified when 1 pound of seed is extracted instead of 2. Volumes are halved; urease is eluted from DEAE-cellulose by a 0.1 to 0.3 M Tris/maleate gradient (instead of 0.1 to 0.4 M); only one pass through hydroxylapatite is performed and the final pass through agarose is unnecessary.
Soybean Cell Suspension Cultures-Callus and suspension cul- tures of soybean (Glycine max, Kanrich variety, Burpee Seed Co.) were induced from shoot tips of sterile IO-day-old etiolated seedlings. Excised shoot tips placed on agar surfaces of R3 medium (6) give “wet” friable callus within 30 days. Suspension cultures were induced by inoculating R3 liquid medium (40 ml in 125-ml flask) with approx- imately 2 g of friable callus. Upon shaking for several days at 90 cycles/min in the dark at 27”C, some flasks exhibited a marked increase in finely dispersed cells which probably arise both from disaggregation of callus and from cell growth. After roughly a quad- rupling of the cell population (about 2 weeks), cells from each flask were filtered through one layer of cheesecloth and used to reinoculate a single flask. The fntration and transfer schedule was continued until a rapidly growing finely dispersed suspension was obtained. For normal maintenance, 10 ml of a midlog culture were added to 65 ml of fresh R3 medium in 250-ml flasks every 5 to 7 days. The cell line used in these experiments has been in culture for about 26 months. Under optimum conditions, the cell division time is 32 h (7).
To induce maximum urease activity for enzyme extraction, soybean cells were cultured in nitrogen-free medium (with added 1 mM KCl). After 24 h, 25 mM urea, 10 mM potassium citrate (pH 6.0). and lo-’ IIIM NiSO, were added from 100 strength stock solutions. The nickel. citrate complex is added to increase urease levels (7). Cells were harvested 1 to 2 days later when there was approximately a doubling of the original cell mass. All flasks were assayed for contamination 24 h prior to or upon harvesting by streaking media and cells on potato dextrose agar (Difco) and on L broth (8) agar.
Preparation of Tissue Culture Urease-Soybean cells, cultured as described above in 2 to 4 l-liter flasks, were collected by suction on Miracloth, washed with distilled water, and taken up as a slurry in
water. Ten to fifteen grams (filtered fresh weight) of cells were added to Braun homogenizer (Melsungen, West Germany) bottles which also contained 20 g of glass beads and 2 ml of 1.0 M potassium phosphate, 0.1 M EDTA, 0.1 M fi-mercaptoethanol, pH 7.0. Water was added to fill each bottle completely (to avoid foaming on homogeni- zation). The contents of each bottle were homogenized for 7 min and cooled with 3- to 5-s bursts of liquid CO, every minute (final concen- tration of buffer is about 40 mM KPO+ 4 mM EDTA, and 4 mu ,/- mercaptoethanol).
The resultant slurry was centrifuged at 10,000 x g for 20 min and to the supernatant, solid ammonium sulfate was rapidly added with stirring to 55% saturation. After dissolution of the ammonium sulfate and standing for 45 min, precipitated protein was collected by cen- trifugation at 16,000 X g for 10 min, resuspended in 10 ml of 10 mM
KPOI, 1 mM EDTA, 1 mM P-mercaptoethanol, pH 7.0, and dialyzed against 1 liter of the same buffer with two changes of buffer. The dialyzed preparation was cleared by centrifugation at 40,000 x g for 10 min. Some preparations were heated at 60°C for 30 min with gentle swirling, followed by centrifugation to remove precipitated protein. The preparation was applied to a column of hydroxylapatite (de- scribed above, (1 x 45 cm)) which was equilibrated with 10 mrvr KPO,, 1 mM EDTA, 1 mM P-mercaptoethanol, pH 7.0. A 600.ml linear gradient of 10 to 300 mrw KPO,, 1 mM EDTA, 1 mM /I-mercaptoeth- anol, pH 7.0, was applied to the column. Fractions of 7.5 ml were collected and those containing maximal activity were pooled and made 55% saturated in ammonium sulfate. Protein was collected bv centrifugation, resuspended in 1 ml of 10 mrvr KPO,, 1 mM EDTA, 1 mM ,&mercaptoethanol, pH 7.0, and dialyzed against this buffer. The enzyme in a volume 52 ml was applied to a column (2 x 45 cm) of agarose A-15m equilibrated with the same buffer and 4.3.ml fractions were collected. Peak activity fractions were pooled and concentrated by precipitation in 55% saturated ammonium sulfate.
Polyacrylamide Slab Gel Electrophoresis-The apparatus for slab gel electrophoresis was from Hoefer Scientific Instruments. Gels (0.075 x 10 X 14 cm) of the appropriate acrylamide concentration were prepared according to the formulation of Gabriel (9). The pH of the gel buffer was 8.9 (Tris/HCl) and the pH of the electrode buffer, 8.3 (Tris/glycine). The electrophoresis period was approximately I h (or until the marker dye was I cm from the edge of the gel) at 15 mA (initial voltage 110 V). After electrophoresis, the gels were removed from between the glass plates and stained with Coomassie blue for protein or with the reagent described by Fishbein (10) for urease activity.
Gels containing SDS were prepared according to Swank and Munkres (11). The polymerization was done in two stages as recom- mended in the procedure provided by the manufacturer. The gel surrounding the Teflon comb did not contain SDS in order to obtain good polymerization. Proteins of known molecular weight which were used as standards were: paramyosin (prepared according to Johnson et al. (12)), bovine serum albumin, ovalbumin, chymotrypsinogen A (Sigma), and phenylalanine ammonia-lyase (maize, prepared accord- ing to Havir and Hanson (13)). Samples were prepared for SDS-gel electrophoresis by the method of Weber and Osborn (14).
Molecular Weight Determination by Gel Filtration on Agarose A 15m Column-A column (4 x 45 cm) of agarose A-15m (Bio-Rad, 200 to 400 mesh) was calibrated with the following standards: dextran blue, thyroglobulin (10 mg), phenylalanine ammonia-lyase from maize (250 milliunits, 4 mg of protein, assay of Havir and Hanson (13)), beef liver catalase (4 x lo5 units, 2 mg of protein, assay of Beers and Sizer (15)), and tritiated water (THO, 4.2 x 10” dpm). The flow rate was 1% ml/h. The percentage of retardation for each of the protein standards was calculated along with the percentage of retardation of urease from jack bean (500 units, -1.0 mg of protein), soybean seed (358 units, 0.6 mg of protein), and soybean tissue culture (see text). Each run with a sample of soybean enzyme also contained at least one standard as well as THO to check the calibration of the column.
Immunological Studies-Purified soybean and jack bean ureases were injected into female albino New Zealand rabbits using low, oft- repeated, and slowly escalating doses, since urease is reported to be toxic to rabbits when given intravenously (16). However, in later trials we found that subcutaneous injections given in Freund’s complete adjuvant are not toxic up to 0.5 mg (250 to 350 units) per initial dose. Four weeks after the initial dose, another 0.5 mg was injected and 40 to 75 ml of blood were withdrawn by cardiac puncture every 2 weeks thereafter. Booster injections were given periodically when titers fell. Blood was heated at 37°C for 1 h in 30-ml Corex centrifuge tubes. The clot was separated by centrifugation (5500 x g, 20 min) and the serum supernatant was mixed with an equal volume of glycerol and
by guest on July 15, 2018http://w
ww
.jbc.org/D
ownloaded from

Urease in Soybean Seed and Cultured Cells 1709
stored at -20°C. The glycerol/serum preparation was used directly in double diffusion plates.
Double diffusion precipitin reactions were observed in g-cm plastic Petri plates containing 15 ml of 0.15 M NaCl, 0.01 M KPO,, pH 7.0, 0.02% NaN3 in 1% Nobel agar. Antigen and antisera, usually 30 ~1, were added to wells prepared with a No. 2 cork borer and aspiration. Plates were maintained at 30°C.
Purified monospecific antibodies to soybean seed urease (see be- low) were used as a sensitive probe to compare urease activity purified from seed with the partially purified tissue culture urease. Urease from seeds of soybean or jack bean was diluted to 1 pg/ml and 75 ~1 were mixed with an equal volume of monospecific antiseed urease antibodies (100 pg/ml) in a final volume of 225 ~1 containing 10 IIIM
potassium phosphate, 0.15 M NaCl, 0.33 mM EDTA, pH 7.0. An equal amount of urease activity (7.5 to 11 milliunits) from tissue culture preparations was used in place of seed urease in some samples. Controls contained protein from a 40% ammonium sulfate fraction of preimmune rabbit serum in place of monospecific antibodies. The mixtures were incubated % h at 30°C in center wells (0.9 x 2.0 cm) in 50-ml flasks. Then, a 1.9-cm’ fluted piece of Whatman No. 3 MM falter paper impregnated with 50 ~1 of 9 M ethanolamine was placed in the flask outside the well and 75 ~1 of 2 M [?L!]urea (12,000 dpm/ pmol) in 0.2 M Tris/maleate, 2 IIIM EDTA, pH 7.0, was added to the center well. Flasks were closed with serum stoppers and incubated 45 min at 3O”C, after which 0.1 ml of 2 N H$04 was injected into the center wells by passing a 3-inch Hamilton syringe needle through the serum stopper. After 30 min, all H14CO:g was trapped on the filter squares which were removed and immersed in scintillation fluid. Radioactivity was determined after another 30 min in a Searle Delta 300 liquid scintillation counter. Radioactivity among the samples was quench-corrected by the external standard ratio method.
Purification of Monospecific Anti-urease Antibodies-Serum was made 40% saturated in ammonium sulfate and precipitated protein was collected by centrifugation, suspended in 10 mM KPO,, 0.15 M
NaCl, pH 7.0, and dialyzed exhaustively (until the dialysate gave a negative Nessler’s reaction) against this buffer. The prec@ation with 40% ammonium sulfate was repeated. This preparation was chromat- ographed on an affinity column containing soybean urease covalently linked to Sepharose. The preparation of the affinity resin is described below.
Urease, 50 mg in 40 ml of 0.1 M KPO,, 1 mM EDTA, and 1 mM ,f-
mercaptoethanol, pH 7.0, was added to 2 g of cyanogen bromide- activated Sepharose 4B (Sigma) and gently swirled overnight at 27°C. The resin was collected by gentle suction filtration on a glass fiber filter. The protein in the filtrate was 12% of the original protein. The resin was resuspended in 50 ml of 0.44 M hydroxylamine, 5 mM KPO.,, 0.075 M NaCl, and 0.5 mM ,fLmercaptoethanol, pH 7.0, and shaken for 2 h. The resin (7.ml bed volume) was then poured into a lo-ml disposable syringe and washed with 0.1 M KPO,, 1 ITIM EDTA, and 1 mM ,&mercaptoethanol until the effluent contained no material ab- sorbing at 280 nm.
About 620 mg of protein in the 40x1 ammonium sulfate serum fraction were poured over the Sepharose-urease column. There was no detectable anti-urease activity in any effluent fraction. The column was sequentially washed with 50 ml of 0.1 M sodium acetate, 1 M
NaCl, pH 5.5; 50 ml of 0.1 M NaHCO:], 1 M NaCl, pH 8.5, 50 ml of 2.5 M magnesium(acetate)Z, 0.05 M Tris, 2 mM EDTA, pH 7.5, and 50 ml of 10 mM KPO,, 0.15 M NaCl, pH 7.0. Bound antibodies were eluted with 3 M KSCN in 10 mM KPO,, 1 mM EDTA, pH 7.0. Generally, 25- ml aliquots of thiocyanate solution were poured over the column. Upon complete elution of bound antibodies, the column could be loaded and eluted several more times. Inexplicably, KSCN eluted some urease as well as antibodies, especially in the early washes. Upon dialysis against 10 IIIM KP04, 0.15 M NaCl, pH 7.0, the urease. antibody complex precipitates and the purified antibodies remain in the supernatant. There were fewer than 2.8 milliunits of urease associated with each milligram of pure antibodies. From the maximum antibody inhibition of urease activity, 40% (Table IV), it is estimated that there are no more than 10 ng of urease in every milligram of antibody (<O.OOl%). The column-purified antibodies increased in titer at least 30.fold, depending on the preparation.
Antibodies to soybean urease cross-react with jack bean urease (Fig. 8). To make the purified antibodies monospecific for soybean urease, 20 mg of jack bean urease were covalently bound to CNBr- activated Sepharose. The procedure was similar to that used for soybean urease except that there was virtually complete ligation of the enzyme to the resin. Pure urease antibodies (-0.8 mg/ml) were
were tested for precipitin reaction with jack bean urease. All fractions reactive to soybean urease, but nonreactive to jack bean urease, were pooled and recycled over the column three times. Antibody prepara- tions were stored in 10 mM KPO,, 0.15 M NaCl, 0.02%~ NaN.,, pH 7.0, at 5°C.
Other Analytical Techniques-In earlier phases of this work, pro- tein was assayed by the modification of the biuret procedure described elsewhere (17). This assay was subsequently supplanted by the Bio- Rad protein dye assay because of its much greater sensitivity and convenience. Protein values obtained from each procedure were vir- tually identical.
Phosphate was determined using a commercial kit (Sigma) or by a modification of the procedure of Murphy and Riley (18). Ten-micro- liter aliquots of lo-fold dilutions of hydroxylapatite effluent fractions were mixed with 1 ml of the molybdate/ascorbate reagent. After 2 min, 4 ml of Hz0 were added and the absorbance at 640 nm was determined after another 20 min.
Amino acid analysis was performed by Gary Davis in the laboratory of Paul Fletcher, Yale University. Protein (70 to 160 pg) was hydro- lyzed for 20 h in 6 N HCl at llO°C. Approximately 25 fig were chromatographed. Tryptophan is destroyed during hydrolysis and cysteine is greatly reduced. Peak areas on chromatograms were inte- grated and compositions were calculated with a Durrum D-500 com- puter program. Threonine and serine values were corrected for h,v- drolytic losses.
RESULTS
Purification of Soybean Seed U-ease to Electrophoretic Homogeneity
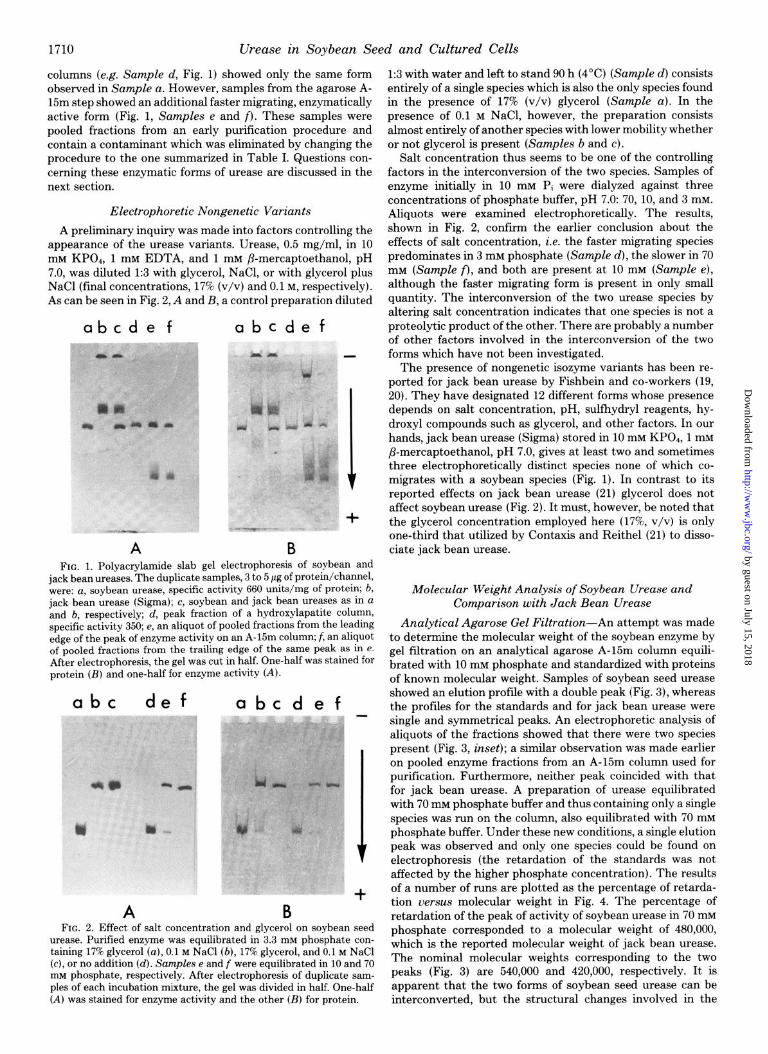
Urease was purified approximately 500-fold with a yield of 11 to 17%. Data from a purification are given in Table I. In the final purification step, m-ease elutes from agarose A-15m as a single peak. The electrophoretic examination of the enzyme after ammonium sulfate concentration is shown in Fig. 1, Sample a. The purified enzyme shows only one protein band which coincides with the one activity band. In contrast, purified jack bean urease (Fig. 1, Sample b) showed several protein and activity bands, none of which coincided with that of soybean urease. When the two enzymes were mixed and then electrophoresed, they separated completely (Fig. 1, Sam- ple c) and yielded no new electrophoretic variants.
During the purification of soybean urease from seed, it was noted that under certain conditions, urease existed as more than a single protein species. The electrophoresis of aliquots of peak fractions from DEAE-cellulose and hydroxylapatite
TABLE I
Purification of soybean seed urease
Procedure pure
Supernatant of 2 lbs ground and extracted seedh
Protein
wz 108,200
Units” Specific activity
unrts/mg
Fold
141,350 1.3 1 100
Heat 6O”C, 1 h, 20 to 55 saturated AmSO
17,960 99,600 5.5 4.2 70.5
33 to 50% acetone (v/v)
2,820 75,600 26.8
DEAE-cellulose, Tris/maleate gradient
218 56,800 261
20.6 53.5
201 40.2
Hydroxylapatite, 91.5 34,300 375 288 24.3 KPO, gradient
Agarose, A-15m 37.1 24.630 664 511 17.4
IL One unit will hydrolyze 1 pmol of urea in 1 min at 3O”C, pH 7.0. applied to the jack bean urease-Sepharose column. Effluent fractions ” Prize soybeans, Burpee Seed Co.
by guest on July 15, 2018http://w
ww
.jbc.org/D
ownloaded from
1710 U-ease in Soybean Seed and Cultured Cells
columns (e.g. Sample d, Fig. 1) showed only the same form observed in Sample a. However, samples from the agarose A- 15m step showed an additional faster migrating, enzymatically active form (Fig. 1, Samples e and f). These samples were pooled fractions from an early purification procedure and contain a contaminant which was eliminated by changing the procedure to the one summarized in Table I. Questions con- cerning these enzymatic forms of m-ease are discussed in the next section.
Electrophoretic Nongenetic Variants
A preliminary inquiry was made into factors controlling the appearance of the urease variants. Urease, 0.5 mg/ml, in 10 mM KPO*, 1 mM EDTA, and 1 mM P-mercaptoethanol, pH 7.0, was diluted 1:3 with glycerol, NaCl, or with glycerol plus NaCl (final concentrations, 17% (v/v) and 0.1 M, respectively). As can be seen in Fig. 2, A and B, a control preparation diluted
abcde f abcdef
A B FIG. 1. Polyacrylamide slab gel electrophoresis of soybean and
jack bean ureases. The duplicate samples, 3 to 5 pg of protein/channel, were: a, soybean urease, specific activity 660 units/mg of protein; b, jack bean urease (Sigma); c, soybean and jack bean ureases as in a and b, respectively; d, peak fraction of a hydroxylapatite column, specific activity 350; e, an aliquot of pooled fractions from the leading edge of the peak of enzyme activity on an A-15m column; f, an aliquot of pooled fractions from the trailing edge of the same peak as in e. After electrophoresis, the gel was cut in half. One-half was stained for protein (B) and one-half for enzyme activity (A).
abc def abcdef
B FIG. 2. Effect of salt concentration and glycerol on soybean seed
urease. Purified enzyme was equilibrated in 3.3 mM phosphate con- taining 17% alvcerol (a). 0.1 M NaCl (b). 17% zlvcerol. and 0.1 M NaCl (c), orno ad&ion (d). &n&es e and f were &&ilibrated in 10 and 70 mM phosphate, respectively. After electrophoresis of duplicate sam- ples of each incubation mixture, the gel was divided in half. One-half (A) was stained for enzyme activity and the other (B) for protein.
1:3 with water and left to stand 90 h (4’C) (Sample d) consists entirely of a single species which is also the only species found in the presence of 17% (v/v) glycerol (Sample a). In the presence of 0.1 M NaCl, however, the preparation consists almost entirely of another species with lower mobility whether or not glycerol is present (Samples b and c).
Salt concentration thus seems to be one of the controlling factors in the interconversion of the two species. Samples of enzyme initially in 10 mM P, were dialyzed against three concentrations of phosphate buffer, pH 7.0: 70, 10, and 3 mM.
Aliquots were examined electrophoretically. The results, shown in Fig. 2, confirm the earlier conclusion about the effects of salt concentration, i.e. the faster migrating species predominates in 3 mM phosphate (Sample d), the slower in 70 mM (Sample f), and both are present at 10 mM (Sample e), although the faster migrating form is present in only small quantity. The interconversion of the two urease species by altering salt concentration indicates that one species is not a proteolytic product of the other. There are probably a number of other factors involved in the interconversion of the two forms which have not been investigated.
The presence of nongenetic isozyme variants has been re- ported for jack bean urease by Fishbein and co-workers (19, 20). They have designated 12 different forms whose presence depends on salt concentration, pH, sulfhydryl reagents, hy- droxyl compounds such as glycerol, and other factors. In our hands, jack bean urease (Sigma) stored in 10 mM KPO*, 1 mM
/&mercaptoethanol, pH 7.0, gives at least two and sometimes three electrophoretically distinct species none of which co- migrates with a soybean species (Fig. 1). In contrast to its reported effects on jack bean urease (21) glycerol does not affect soybean urease (Fig. 2). It must, however, be noted that the glycerol concentration employed here (17%, v/v) is only one-third that utilized by Contaxis and Reithel (21) to disso- ciate jack bean urease.
Molecular Weight Analysis of Soybean Urease and Comparison with Jack Bean Urease
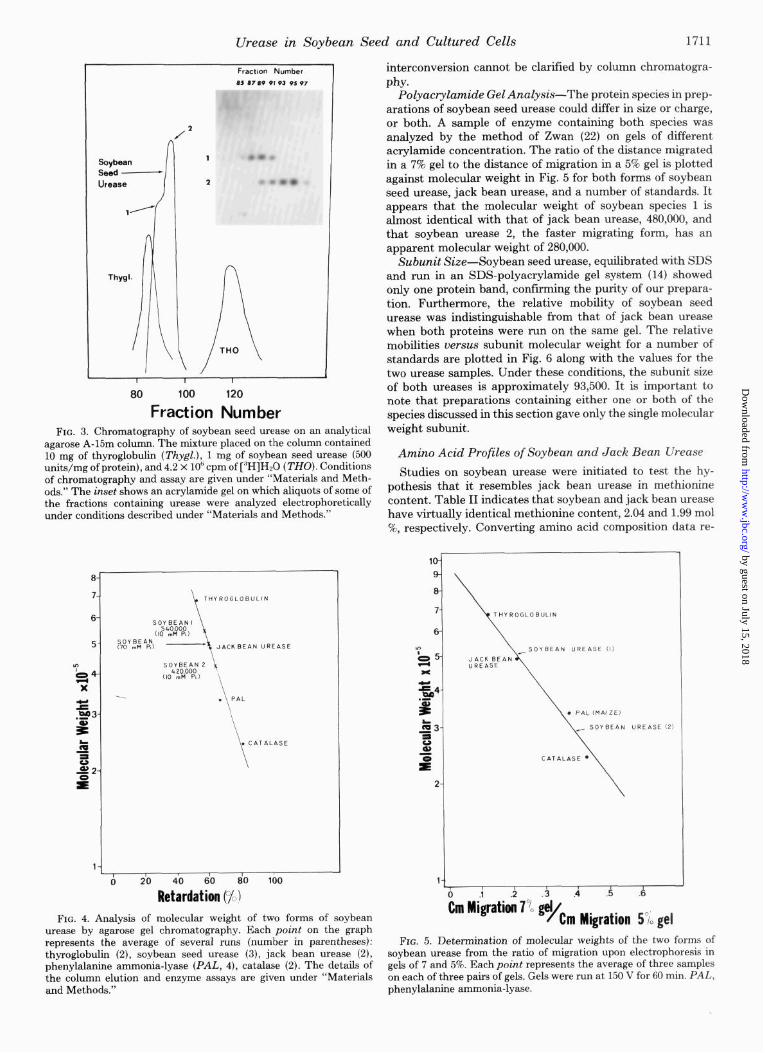
Analytical Agarose Gel Filtration-An attempt was made to determine the molecular weight of the soybean enzyme by gel filtration on an analytical agarose A-15m column equili- brated with 10 mM phosphate and standardized with proteins of known molecular weight. Samples of soybean seed urease showed an elution profile with a double peak (Fig. 3), whereas the profiles for the standards and for jack bean urease were single and symmetrical peaks. An electrophoretic analysis of aliquots of the fractions showed that there were two species present (Fig. 3, inset); a similar observation was made earlier on pooled enzyme fractions from an A-15m column used for purification. Furthermore, neither peak coincided with that for jack bean urease. A preparation of urease equilibrated with 70 mM phosphate buffer and thus containing only a single species was run on the column, also equilibrated with 70 mM phosphate buffer. Under these new conditions, a single elution peak was observed and only one species could be found on electrophoresis (the retardation of the standards was not affected by the higher phosphate concentration). The results of a number of runs are plotted as the percentage of retarda- tion uersus molecular weight in Fig. 4. The percentage of retardation of the peak of activity of soybean urease in 70 mM phosphate corresponded to a molecular weight of 480,000, which is the reported molecular weight of jack bean m-ease. The nominal molecular weights corresponding to the two peaks (Fig. 3) are 540,000 and 420,000, respectively. It is apparent that the two forms of soybean seed urease can be interconverted, but the structural changes involved in the
by guest on July 15, 2018http://w
ww
.jbc.org/D
ownloaded from
W-ease in Soybean Seed and Cultured Cells 1711
interconversion cannot be clarified by column chromatogra-
phy.
.z
1 . -
2 - . .
Thygl.
h
Polyacrylamide Gel Analysis--The protein species in prep- arations of soybean seed urease could differ in size or charge, or both. A sample of enzyme containing both species was analyzed by the method of Zwan (22) on gels of different acrylamide concentration. The ratio of the distance migrated in a 7% gel to the distance of migration in a 5% gel is plotted against molecular weight in Fig. 5 for both forms of soybean seed urease, jack bean urease, and a number of standards. It appears that the molecular weight of soybean species 1 is almost identical with that of jack bean urease, 480,000, and that soybean urease 2, the faster migrating form, has an apparent molecular weight of 280,000.
\ \
I I I
80 100 120
Fraction Number FIG. 3. Chromatography of soybean seed urease on an analytical
agarose A-15m column. The mixture placed on the column contained 10 mg of thyroglobulin (Z’hygl.), 1 mg of soybean seed urease (500 units/mg of protein), and 4.2 x lo6 cpm of [3H]Hz0 (Z’HO). Conditions of chromatography and assay are given under “Materials and Meth- ods.” The inset shows an acrylamide gel on which aliquots of some of the fractions containing urease were analyzed electrophoreticahy under conditions described under “Materials and Methods.”
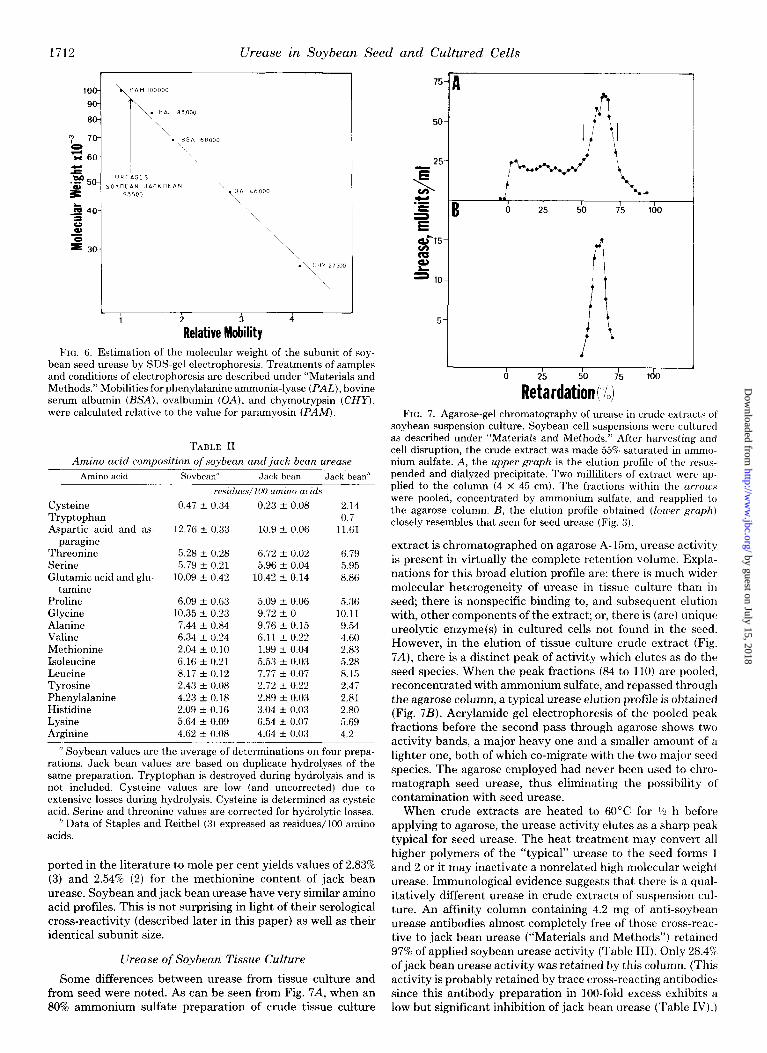
Subunit Size-Soybean seed urease, equilibrated with SDS and run in an SDS-polyacrylamide gel system (14) showed only one protein band, confiiing the purity of our prepara- tion. Furthermore, the relative mobility of soybean seed urease was indistinguishable from that of jack bean urease when both proteins were run on the same gel. The relative mobilities versus subunit molecular weight for a number of standards are plotted in Fig. 6 along with the values for the two urease samples. Under these conditions, the subunit size of both ureases is approximately 93,500. It is important to note that preparations containing either one or both of the species discussed in this section gave only the single molecular weight subunit.
Amino Acid Profiles of Soybean and Jack Bean Urease
Studies on soybean urease were initiated to test the hy- pothesis that it resembles jack bean urease in methionine content. Table II indicates that soybean and jack bean urease have virtually identical methionine content, 2.04 and 1.99 mol %, respectively. Converting amino acid composition data re-
8
7
6
1
\
0 20 40 60 80 100
Retardation (“/,I FIG. 4. Analysis of molecular weight of two forms of soybean
urease by agarose gel chromatography. Each point on the graph represents the average of several runs (number in parentheses): thyroglobulin (2), soybean seed urease (3), jack bean urease (2), phenylalanine ammonia-lyase (PAL, 4), catalase (2). The details of the column elution and enzyme assays are given under “Materials and Methods.”
10
9
8
6
I-
bll Migrati6n2 7’6 .6
52 gel FIG. 5. Determination of molecular weights of the two forms of
soybean urease from the ratio of migration upon electrophoresis in gels of 7 and 5%. Each point represents the average of three samples on each of three pairs of gels. Gels were run at 150 V for 60 min. PAL, phenylalanine ammonia-lyase.
by guest on July 15, 2018http://w
ww
.jbc.org/D
ownloaded from
Urease in Soybean Seed and Cultured Cells
10
9
8
i i Relative Mohty
4
FIG. 6. Estimation of the molecular weight of the subunit of soy- bean seed urease by SDS-gel electrophoresis. Treatments of samples and conditions of electrophoresis are described under “Materials and Methods.” Mobilities for phenylalanine ammonia-lyase (PAZ,), bovine serum albumin (BSA), ovalbumin (OA), and chymotrypsin (CHY), were calculated relative to the value for paramyosin (PAM).
TABLE II
Amino acid comDoaition of sovbean and jack bean urease Amino acid
Cysteine Tryptophan Aspartic acid and as-
paragine Threonine Serine Glutamic acid and glu-
tamine Proline Glycine Alanine Valine Methionine Isoleucine Leucine Tyrosine Phenylalanine Histidine Lysine Arginine
Soybean" Jack bean Jack bean*
resrduen/lOO ammo acids
0.47 +- 0.34 0.23 f 0.08 2.14 0.7
12.76 & 0.33 10.9 + 0.06 11.61
5.28 f 0.28 6.72 f 0.02 6.79 5.79 f 0.21 5.96 f 0.04 5.95
10.09 f 0.42 10.42 + 0.14 8.86
6.09 + 0.63 5.09 + 0.06 5.36 10.35 f 0.23 9.72 f 0 10.11
7.44 f 0.84 9.76 + 0.15 9.54 6.34 + 0.24 6.11 f 0.22 4.60 2.04 f 0.10 1.99 f 0.04 2.83 6.16 k 0.21 5.53 + 0.03 5.28 8.17 f 0.12 7.77 f 0.07 8.15 2.43 f 0.08 2.72 + 0.22 2.47 4.23 k 0.18 2.89 f 0.03 2.81 2.09 f 0.16 3.04 * 0.03 2.80 5.64 + 0.09 6.54 + 0.07 5.69 4.62 f 0.08 4.64 -c 0.03 4.2
’ Soybean values are the average of determinations on four prepa- rations. Jack bean values are based on duplicate hydrolyses of the same preparation. Tryptophan is destroyed during hydrolysis and is not included. Cysteine values are low (and uncorrected) due to extensive losses during hydrolysis. Cysteine is determined as cysteic acid. Serine and threonine values are corrected for hydrolytic losses.
’ Data of Staples and Reithel (3) expressed as residues/100 amino acids.
ported in the literature to mole per cent yields values of 2.83% (3) and 2.54% (2) for the methionine content of jack bean urease. Soybean and jack bean urease have very similar amino acid profiles. This is not surprising in light of their serological cross-reactivity (described later in this paper) as well as their identical subunit size.
Urease of Soybean Tissue Culture
Some differences between urease from tissue culture and from seed were noted. As can be seen from Fig. 7A, when an 80% ammonium sulfate preparation of crude tissue culture
i
I I I I 0 25 50 75 do
Retardation(%) FIG. 7. Agarose-gel chromatography of urease in crude extracts of
soybean suspension culture. Soybean cell suspensions were cultured as described under “Materials and Methods.” After harvesting and cell disruption, the crude extract was made 55% saturated in ammo- nium sulfate. A, the upper graph is the elution profile of the resus- pended and dialyzed precipitate. Two milliliters of extract were ap- plied to the column (4 x 45 cm). The fractions within the arrou’s were pooled, concentrated by ammonium sulfate, and reapplied to the agarose column. B, the elution profile obtained (Zouw .qra$) closely resembles that seen for seed m-ease (Fig. 3).
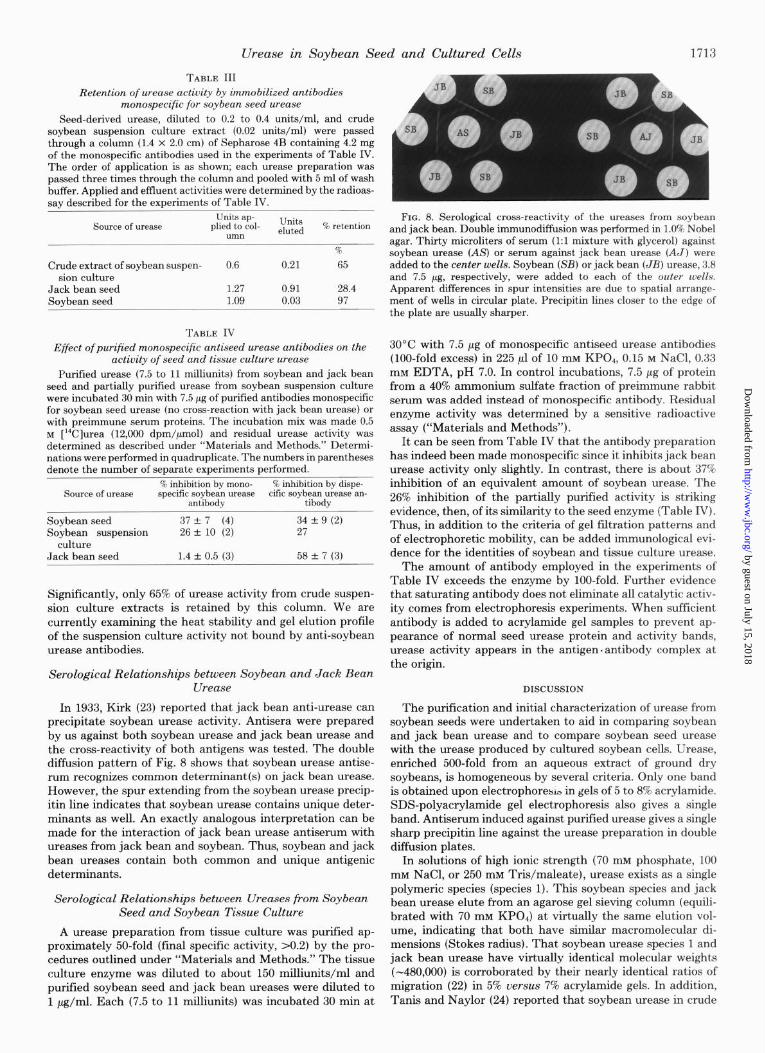
extract is chromatographed on agarose A-15m, urease activity is present in virtually the complete retention volume. Expla- nations for this broad elution profile are: there is much wider molecular heterogeneity of urease in tissue culture than in seed; there is nonspecific binding to, and subsequent elution with, other components of the extract; or, there is (are) unique ureolytic enzyme(s) in cultured cells not found in the seed. However, in the elution of tissue culture crude extract (Fig. 7A), there is a distinct peak of activity which elutes as do the seed species. When the peak fractions (84 to 110) are pooled, reconcentrated with ammonium sulfate, and repassed through the agarose column, a typical urease elution profile is obtained (Fig. 7B). Acrylamide gel electrophoresis of the pooled peak fractions before the second pass through agarose shows two activity bands, a major heavy one and a smaller amount of a lighter one, both of which co-migrate with the two major seed species. The agarose employed had never been used to chro- matograph seed urease, thus eliminating the possibility of contamination with seed urease.
When crude extracts are heated to 60°C for X h before applying to agarose, the urease activity elutes as a sharp peak typical for seed urease. The heat treatment may convert all higher polymers of the “typical” urease to the seed forms 1 and 2 or it may inactivate a nonrelated high molecular weight urease. Immunological evidence suggests that there is a qual- itatively different urease in crude extracts of suspension cul- ture. An affinity column containing 4.2 mg of anti-soybean urease antibodies almost completely free of those cross-reac- tive to jack bean urease (“Materials and Methods”) retained 97% of applied soybean urease activity (Table III). Only 28.4% of jack bean urease activity was retained by this column. (This activity is probably retained by trace cross-reacting antibodies since this antibody preparation in loo-fold excess exhibits a low but significant inhibition of jack bean urease (Table IV).)
by guest on July 15, 2018http://w
ww
.jbc.org/D
ownloaded from
Urease in Soybean Seed and Cultured Cells
TABLE III
Retention of w-ease activity by immobilized antibodies monospecific for soybean seed urease
Seed-derived urease, diluted to 0.2 to 0.4 units/ml, and crude soybean suspension culture extract (0.02 units/ml) were passed through a column (1.4 x 2.0 cm) of Sepharose 4B containing 4.2 mg of the monospecific antibodies used in the experiments of Table IV. The order of application is as shown; each urease preparation was passed three times through the column and pooled with 5 ml of wash buffer. Applied and effluent activities were determined by the radioas- say described for the experiments of Table IV.
Units ap- Source of urease plied to col- Units
eluted % retention “TIllI
Crude extract of soybean suspen- sion culture
Jack bean seed Soybean seed
0.6
1.27 1.09
% 0.21 65
0.91 28.4 0.03 97
TABLE IV
Effect ofpurified monospecific antiseed w-ease antibodies on the activity of seed and tissue culture w-ease
Purified urease (7.5 to 11 milliunits) from soybean and jack bean seed and partially purified urease from soybean suspension culture were incubated 30 min with 7.5 pg of purified antibodies monospecific for soybean seed urease (no cross-reaction with jack bean urease) or with preimmune serum proteins. The incubation mix was made 0.5 M [14C]urea (12,000 dpm/pmol) and residual urease activity was determined as described under “Materials and Methods.” Determi- nations were performed in quadruplicate. The numbers in parentheses denote the number of separate experiments performed.
% inhibition by mono- % inhibition by dispe- Source of urease specific soybean urease ciiic soybean urease an-
antibody tibody
Soybean seed 37 f 7 (4) 34 + 9 (2) Soybean suspension 26 + 10 (2) 27
culture Jack bean seed 1.4 f 0.5 (3) 58 + 7 (3)
Significantly, only 65% of urease activity from crude suspen- sion culture extracts is retained by this column. We are currently examining the heat stability and gel elution profile of the suspension culture activity not bound by anti-soybean urease antibodies.
Serological Relationships between Soybean and Jack Bean U-ease
In 1933, Kirk (23) reported that jack bean anti-urease can precipitate soybean urease activity. Antisera were prepared by us against both soybean urease and jack bean urease and the cross-reactivity of both antigens was tested. The double diffusion pattern of Fig. 8 shows that soybean urease antise- rum recognizes common determinant(s) on jack bean urease. However, the spur extending from the soybean urease precip- itin line indicates that soybean urease contains unique deter- minants as well. An exactly analogous interpretation can be made for the interaction of jack bean urease antiserum with ureases from jack bean and soybean. Thus, soybean and jack bean ureases contain both common and unique antigenic determinants.
Serological Relationships between W-eases from Soybean Seed and Soybean Tissue Culture
A urease preparation from tissue culture was purified ap- proximately 50-fold (final specific activity, >0.2) by the pro- cedures outlined under “Materials and Methods.” The tissue culture enzyme was diluted to about 150 milliunits/ml and purified soybean seed and jack bean ureases were diluted to 1 pg/ml. Each (7.5 to 11 milliunits) was incubated 30 min at
FIG. 8. Serological cross-reactivity of the ureases from soybean and jack bean. Double immunodiffusion was performed in 1.0% Nobel agar. Thirty microliters of serum (1:l mixture with glycerol) against soybean urease (AS) or serum against jack bean urease (AJ) were added to the center wells. Soybean (SB) or jack bean (JB) urease, 3.8 and 7.5 pg, respectively, were added to each of the outer wells. Apparent differences in spur intensities are due to spatial arrange- ment of wells in circular plate. Precipitin lines closer to the edge of the plate are usually sharper.
30°C with 7.5 pg of monospecific antiseed urease antibodies (loo-fold excess) in 225 ~1 of 10 mM KPO+ 0.15 M NaCl, 0.33 mM EDTA, pH 7.0. In control incubations, 7.5 pg of protein from a 40% ammonium sulfate fraction of preimmune rabbit serum was added instead of monospecific antibody. Residual enzyme activity was determined by a sensitive radioactive assay (“Materials and Methods”).
It can be seen from Table IV that the antibody preparation has indeed been made monospecific since it inhibits jack bean urease activity only slightly. In contrast, there is about 37% inhibition of an equivalent amount of soybean urease. The 26% inhibition of the partially purified activity is striking evidence, then, of its similarity to the seed enzyme (Table IV). Thus, in addition to the criteria of gel filtration patterns and of electrophoretic mobility, can be added immunological evi- dence for the identities of soybean and tissue culture urease.
The amount of antibody employed in the experiments of Table IV exceeds the enzyme by loo-fold. Further evidence that saturating antibody does not eliminate all catalytic activ- ity comes from electrophoresis experiments. When sufficient antibody is added to acrylamide gel samples to prevent ap- pearance of normal seed urease protein and activity bands, urease activity appears in the antigen. antibody complex at the origin.
DISCUSSION
The purification and initial characterization of urease from soybean seeds were undertaken to aid in comparing soybean and jack bean urease and to compare soybean seed urease with the urease produced by cultured soybean cells. Urease, enriched 500-fold from an aqueous extract of ground dry soybeans, is homogeneous by several criteria. Only one band is obtained upon electrophore& in gels of 5 to 8% acrylamide. SDS-polyacrylamide gel electrophoresis also gives a single band. Antiserum induced against purified urease gives a single sharp precipitin line against the urease preparation in double diffusion plates.
In solutions of high ionic strength (70 mM phosphate, 100 mM NaCl, or 250 mM Tris/maleate), urease exists as a single polymeric species (species 1). This soybean species and jack bean urease elute from an agarose gel sieving column (equili- brated with 70 mM KPOd at virtually the same elution vol- ume, indicating that both have similar macromolecular di- mensions (Stokes radius). That soybean urease species 1 and jack bean urease have virtually identical molecular weights (-480,000) is corroborated by their nearly identical ratios of migration (22) in 5% uersas 7% acrylamide gels. In addition, Tanis and Naylor (24) reported that soybean urease in crude
by guest on July 15, 2018http://w
ww
.jbc.org/D
ownloaded from

1714 Urease in Soybean Seed and Cultured Cells
preparations can exist in an 18.0 and 13.3 S form. This agrees well with the values of 18.3 and 11.8 S they reported for crystalline jack bean urease. Contaxis and Reithel(21) utilized sedimentation equilibrium measurements to derive molecular weights of 240,000 and 480,000 for the “monomer” and “dimer” species, respectively, of jack bean urease.
Analysis of gel electrophoresis migration rates for soybean species 2 indicates that it is a smaller multimer (Mr = 280,000) than species 1 (Fig. 5). Since they are interconvertible and since they yield the same single band in SDS-acrylamide gels, the simplest interpretation is that multimer species 1 splits to give two identical half-species 2. Interpretations based on nonequal splitting cannot explain the lack of appearance of a third species in the species 1 to species 2 conversion. Based on the SDS-derived value for the subunit size, 93,500, species 2 appears to be a trimer. However, the expected hexameric molecular weight of 560,000 for species 1 is somewhat larger than the 480,000 value derived from gel sieving (Fig. 4) and acrylamide gel migration ratios (Fig. 5). It seems likely that species 1 and 2 differ in shape; the molecular weight analyses are also complicated by lack of understanding of the basis for molecular weight determination by the Zwan (22) technique.
Amino acid analysis of our preparations of soybean and jack bean urease revealed similar methionine contents (2 mol 970). When the amino acid profile determined for jack bean urease by Staples and Reithel (3) is converted to mole per cent there is generally good agreement between their values and ours (Table II). Their analysis (3), unlike ours, includes accurate values for tryptophan and cysteine. However, the sum of these two is only 2.84 mol % so that a meaningful comparison of the mole per cent of the other amino acids can be made. Staples and Reithel (3) report that 2.83% of the residues are methio- nine. This figure agrees well with NH2-terminal and cyanogen bromide peptide analysis which indicates that there are 8 mol of methionine/30,400 g of enzyme (3). While we cannot explain the lower methionine levels (2%) found in our preparations of soybean and jack bean urease than that reported for jack bean, 2.83% (3), the analyses at least indicate comparable methionine levels in soybean and jack bean urease. Staples and Reithel (3) reported that 30,400 is the calculated molec- ular weight of jack bean urease which gives the best fit integer values for the molar composition of amino acid residues. Indeed, when urease was treated with performic acid, SDS- gel electrophoresis gave rise to a major band of M, = 32,000 (3). Using the method of Weber and Osborn (14), we obtained SDS-derived molecular weight values of 93,500 for soybean and jack bean urease, indicating that both enzymes contain inaccessible or unusually stable sulfhydryl bridges, or both.
Previous work from our laboratories has demonstrated that urea can serve as a nitrogen source for soybean callus and suspension cultures (7, 17, 25). Such cultured cells contain an ATP-independent ureolytic activity. This ureolytic activity reaches a peak soon after cells (within one to two generations) leave lag phase. Ammonia and methylamine appear to inter- fere with the urea-induced stimulation of urease activity in early logarithmic growth (17). We have consistently observed that ureolytic activity is virtually zero in stationary phase cells and in cells which have been deprived of nitrogen for 24 h. By contrast, two other putative nitrogen assimilatory enzymes, glutamic dehydrogenase and arginase, exhibit activity levels little changed in stationary phase (17) and after nitrogen- depletion (7), respectively. Glutamine synthetase activity be- haves somewhat like that of urease, dropping 80% after a 48- h culture in nitrogen-free medium and reaching maximal levels within 24 h of transfer to nitrogen-sufficient medium.’
’ J. C. Polacco, unpublished results.
The above studies on the regulation of urease activity in cultured soybean cells were undertaken to help devise selec- tions for urease overproducing mutants. Since urease is a seed protein in the intact plant, interesting parallels between its regulation in tissue culture and in planta may be elucidated. Tissue culture can then be used as a genetic system to select for cell lines which regenerate a plant exhibiting high seed urease levels. The results of the work presented here indicate the validity of the first basic assumption of this approach, namely that the soybean enzyme, like that of jack bean, has a high methionine content. The second basic assumption, that the cell culture and seed enzymes are identical, has at least partially been borne out. It has been demonstrated that cul- tured cells produce a urease indistinguishable from the seed enzyme on the basis of electrophoretic species, gel filtration, and immunoinhibition. However, it appears that another ureo- lytic enzyme(s) may be produced by cultured cells. While 100% of the ureolytic activity of crude cell extracts is retained by purified antibodies to soybean urease, only 65% is retained by urease antibodies which were separated from those which cross-react with jack bean urease. At present, it is not known whether the 35% of the activity which is immunologically inactive represents a unique enzyme or simply a processing variant (altered carbohydrate content, higher order multimer, etc.) of the “normal” urease. We are currently studying the gel sieving properties and subunit size of this “altered” urease activity.
There is strong evidence that both the seed and tissue culture enzymes are nickel metalloenzymes. We have observed that soybean tissue cultures will not produce any urease in “nickel-free” medium (containing the nickel chelator citrate). Nickel supplementation results in sharp increases in urease levels (7, 25). Preliminary analysis of our preparation of puri- fied soybean seed urease indicates that there are about 2 atoms of nickel/subunit molecule (93,500 daltons) .” This agrees well with the values of 2 nickel atoms/105,000 daltons of jack bean urease reported by Dixon et al. (26).
Acknowledgments-We gratefully acknowledge the expert techni- cal assistance of Carla Padula-Forte, Katherine Clark, and Nancy DePalma. We thank Louise Camera and J. Allan Dodds for introduc- ing some immunological techniques to us and R. B. Sparks, Jr. for helpful discussion.
1. 2.
3.
4.
5.
6.
7. a.
9. 10.
11.
12.
REFERENCES
Sumner, J. B. (1926) J. Biol. Chem. 69,435-441 Milton, J. M., and Taylor, I. E. P. (1969) Biochem. J. 113, 678-
680 Staples, S. J., and Reithel, F. J. (1976) Arch. Biochem. Biophys.
174,651-657 Bressani, R., and Elias, L. G. (1974) in New Protein Foooorls
Technology (Altschul, A. M., ed) Vol. lA, pp. 230-297, Aca- demic Press, New York
Peterson, E. A., and Sober, H. A. (1962) Methods Enzymol. 5,3- 27
Chaleff, R. S., and Carlson, P. S. (1975) in Modification of the Information Content of Plant Cells (Markham, R., Davies, 11. R., Hopwood, D. A., and Horne, R. W., eds) pp. 197-214, American Elsevier Publishing Co., New York
Polacco, J. C. (1977) PZant Physiol. (Bethesda) 59, 827-830 Miller, J. H. (1972) Experiments in Molecular Genetics, p. 433,
Cold Spring Harbor Laboratory, Cold Spring Harbor, N. Y. Gabriel, 0. (1971) Methods Enzymol. 22, 565-578 Fishbein, W. N. (1969) in Fifth International Symposium on
Chromatography and Electrophoresis, pp. 238-241, Ann Arbor Humphrey Science Press, Ann Arbor, Mich.
Swank, R. T., and Munkres, K. D. (1971) Anal. Biochem. 39,462- 477
Johnson, W. H., Kahn, J. S., and Szent-GyGrgyi, A. G. (1959) Science 130, 160-161
-
’ T. Grove and F. W. Sunderman, Jr., personal communication.
by guest on July 15, 2018http://w
ww
.jbc.org/D
ownloaded from

Urease in Soybean Seed and Cultured Cells 1715
13. Havir, E. A., and Hanson, K. (1973) Biochemistry 12,1583-1591 14. Weber, K., and Osborn, M. (1969) J. Biol. Chem. 244,4406-4412 15. Beers, R. F., Jr., and Sizer, I. W. (1952) J. Biol. Chem. 195, 133-
140 16. Kirk, J. S., and Sumner, J. B. (1931) J. Biol. Chem. 94,21-28 17. Polacco, J. C. (1976) Plant Physiol. (Bethesda) 58, 350-357 18. Murphy, J., and Riley, J. P. (1962) Anal. Chim. Acta 27,31-36 19. Fishbein, W. N., Nagarajan, K., and Scurzi, W. (1975) in Isozymes,
Molecular Structure (Markert, C. L., ed) Vol. 1, pp. 403-417, Academic Press, New York
20. Fishbein, W. N., Nagarajan, K., and Scurzi, W. (1976) Arch. Biochem. Biophys. 172,726-733
21. Contaxis, C. C., and Reithel, F. J. (1971) J. BioZ. Chem. 246,677- 685
22. Zwan, J. (1967) Anal. Biochem. 21, 155-168 23. Kirk, J. S. (1933) J. BioZ. Chem. 100,667-670 24. Tanis, R. J., and Naylor, A. W. (1968) Biochem. J. 108, 771-777 25. Polacco, J. C. (1977) Plant Sci. Lett. 10, 249-255 26. Dixon, N. E., Gazzola, C., Blakeley, R. L., and Zerner, B. (1975)
J. Am. Chem. Sot. 97,4131-4133
by guest on July 15, 2018http://w
ww
.jbc.org/D
ownloaded from

J C Polacco and E A HavirComparisons of soybean urease isolated from seed and tissue culture.
1979, 254:1707-1715.J. Biol. Chem.
http://www.jbc.org/content/254/5/1707.citation
Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/254/5/1707.citation.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on July 15, 2018http://w
ww
.jbc.org/D
ownloaded from





























