Embed Size (px)
Citation preview

of November 27, 2018.This information is current as
Antigen into B Cell FolliclesComplement-Dependent Transport of
Turley and Michael C. CarrollKuligowski, Lisa A. Pitcher, Søren E. Degn, Shannon J. Santiago F. Gonzalez, Veronika Lukacs-Kornek, Michael P.
http://www.jimmunol.org/content/185/5/2659doi: 10.4049/jimmunol.1000522
2010; 185:2659-2664; ;J Immunol
Referenceshttp://www.jimmunol.org/content/185/5/2659.full#ref-list-1
, 32 of which you can access for free at: cites 73 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved.Copyright © 2010 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on Novem
ber 27, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on N
ovember 27, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

Complement-Dependent Transport of Antigen into B CellFolliclesSantiago F. Gonzalez,*,† Veronika Lukacs-Kornek,†,‡ Michael P. Kuligowski,*,†
Lisa A. Pitcher,*,† Søren E. Degn,*,†,x Shannon J. Turley,†,‡ and Michael C. Carroll*,†,{
Since the original proposal by Fearon and Locksley(Fearon and Locksley. 1996. Science 272: 50–53) thatthe complement system linked innate and adaptiveimmunity, there has been a rapid expansion of studieson this topic. With the advance of intravital imaging, anumber of recent papers revealed an additional novelpathway in which complement C3 and its receptorsenhance humoral immunity through delivery of Ag tothe B cell compartment. In this review, we discuss thispathway and highlight several novel exceptions recentlyfound with a model influenza vaccine, such as mannose-binding lectin opsonization of influenza and uptake bymacrophages, and the capture of virus by dendritic cellsresiding in the medullary compartment of peripherallymph nodes. The Journal of Immunology, 2010, 185:2659–2664.
Peripheral lymphnodes (LNs) and the spleenmakeup thesecondary lymphoid organ tissue that provide a speci-alized environment for circulating B and T lymphocytes
to interact and encounter cognate Ag (1). Although T cellshome to the paracortical region of LNs, B cells traffic to thefollicles in search of Ag. This directed migration is dependenton chemokines produced by stromal cells in the respectivecompartments. Recent elegant intravital imaging of T andB cell trafficking within the peripheral LNs revealed a directedmigration along stromal “highways” (2, 3). Fibroblast reticularcells (FRCs) secrete the collagen-rich fibers that form the net-work within the paracortical region, and they secrete T cellchemokines CCL19 and CCL21. B cell migration within thefollicles is dependent on follicular dendritic cells (FDCs) den-dritic processes and a less-dense network of FRC fibers.Although the reticular network within LNs was characterizedmore than three decades ago (4), only more recently did itbecome apparent that it acts as a conduit for the delivery ofcytokines, chemokines, and small-protein Ags to the T (5–8)and B cell areas (9, 10). B cell conduits are structurally and
immunochemically similar to those in the T cell area. Theydiffer primarily by specificity of the chemokine secreted (i.e.,follicular FRCs secrete CXCL-13, whereas paracortical FRCssecrete CCL19 and CCL21). Although the outer diameter ofconduits is ∼1–2 mm, they are tightly packed with collagenfibers with a spacing of 5–8 nm, which acts as a molecular sieve(Fig. 1). Thus, only proteins less than ∼60 kDa enter theconduits. Whether conduit structures are altered to accommo-date larger Ags during infection is not clear.
Trafficking of lymph-borne Ags into B cell follicles
Small protein Ags gain direct access into the B cell follicles viagaps in the subcapsular sinus (SCS) floor (11) or through theFRC conduits (Fig. 2A) (9). The latter pathway provides adirected flow of small Ags to the FDCs for transient retention,or in the presence of Ab and complement, long-term bindingvia specific receptors. Although cognate B cells can access Agdraining via the conduits (9), their principal role is more likelydirecting the Ag to FDCs for stable retention. Although theseinitial experiments involved model Ags, such as lysozyme (9),or OVA (10), in the natural setting it seems likely that a majorsource of Ag is degraded products of pathogens that drain fromtissues via the lymphatics, as suggested by Jenkins and col-leagues (12).Lymph-borne particulate Ags, such as vesicular stomatitis
virus (13), and protein-coated beads (14) are rapidly taken upby macrophages that line the SCS (SSMs) (15). Interestingly,the particulate Ags are shuttled to the underlying surface wherethey are made available to cognate B cells. Similarly, largeprotein Ags injected s.c. into passively immunized mice alsoseem to bind rapidly to SSMs. However, in the latter example,capture by SSMs is complement dependent. Thus, formation ofimmune complexes (ICs) activates complement, resulting information of C3-coated ICs (C3-ICs) that enhance uptake viaCR3 (Mac-1) and FcRIIb on the SSMs (16). Subsequently,C3-ICs are relayed to the underlying B cell compartment wherethey are transferred to naive B cells (Fig. 2A) (17). FcRIIb isknown to recycle to the surface following internalization and
*The Immune Disease Institute and Program in Cellular and Molecular Medicine,Children’s Hospital, †Department of Pathology, and {Department of Pediatrics, Har-vard Medical School; ‡Department of Cancer Immunology and AIDS, Dana FarberCancer Institute, Boston, MA; and xDepartment of Medical Microbiology and Immu-nology, Arhus University, Arhus, Denmark
Received for publication April 12, 2010. Accepted for publication May 19, 2010.
This work was supported byNational Institutes of Health Grants 5 RO1 AI039246, 1 PO1AI078897, 5 RO1AI067706 (toM.C.C.), andRO1DK074500 (to S.J.T.); a Glaxo-SmithKline postdoctoral fellowship award (to M.K.); Marie Curie International Outgoing Fel-lowship for Career Development Award 220044 (to S.F.G.); and National Institutes ofHealth Transfusion Biology and Medicine Training Grant 5T32HL066987-09 (to L.A.P.).
Address correspondence and reprint requests to Dr. Michael C. Carroll, Immune DiseaseInstitute, 200 Longwood Avenue, WAB, Room 251, Boston, MA 02115. E-mail address:[email protected]
Abbreviations used in this paper: C3-IC, C3-coated immune complex; CR, cysteine-rich; DC, dendritic cell; FDC, follicular dendritic cell; FRC, fibroblast reticular cell; GC,germinal center; IC, immune complex; LN, lymph node; MF, macrophage; MBL,mannose-binding lectins; MM, medullary macrophage; MP-IVM, multiphoton intra-vital microscopy; MZ, marginal zone; SCS, subcapsular sinus; SIGN-R1, specific intra-cellular adhesion molecule-grabbing nonintegrin R1; SSM, subcapsular sinus-liningmacrophage.
Copyright� 2010 by TheAmerican Association of Immunologists, Inc. 0022-1767/10/$16.00
www.jimmunol.org/cgi/doi/10.4049/jimmunol.1000522
by guest on Novem
ber 27, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

not go through a lysosomal compartment (18). Therefore, it ispossible that C3-ICs are partially protected by this cycling proc-ess. It is not clear how C3-ICs are actually transferred to B cells;however, uptake on the naive B cells is CD21/35 dependentand highly efficient. Strikingly, .25% of naive B cells withinthe LN follicles take up C3-ICs within 8 h of s.c. injection ofAg into immune mice (9, 16).In naive mice in which Ab does not pre-exist, it is not clear
how large protein Ags within the lymph are captured by sinus-lining macrophages. The lymph, in general, is thought toinclude a similar repertoire of recognition proteins that activatecomplement as found in blood, such as mannose-binding lec-tins (MBL), ficolins, and pentraxin. Alternatively, protein Agsmight be directly taken up by scavenger or lectin receptors ex-pressed on the sinus-lining macrophages. Medullary macro-
phages (MMs), which are more similar to marginal zone(MZ) macrophages of the spleen, express specific intracellularadhesion molecule-grabbing nonintegrin R1 (SIGN-R1) andmannose receptor (MR), in addition to CR3. However, SSMsare more similar to the metallophilic macrophages of the MZand express metallophilic macrophages Ab-1 (17).
B cell transport of C3-ICs is CD21/35 dependent
In mice, complement receptors CD21 and CD35 are encodedat the Cr2 locus (19, 20). CD21 represents a splice product ofthe CD35 mRNA, whereas in humans they are encoded atseparate loci (21, 22). Murine CD21 and CD35 (CD21/35)are coexpressed on B cells, FDCs, and a subset of T cells.CD35 binds activated C3b and C4b and, like CD21, it bindsthe cleavage products iC3b, C3d,g, and C3d covalently
FIGURE 1. The conduit network, formed by collagen
fibers, is secreted by the FRCs and drains small Ags from
the SCS area of the LN to the B cell follicle. FDCs present
in the follicle are closely associated with the collagen fibers.
Right panel shows an electron micrograph of the SCS
area. The arrow indicates a conduit opening into the sinus
(S.F. Gonzalez, unpublished results). Scale bar, 2 mm.
MF, macrophage; SCS, subcapsular sinus.
FIGURE 2. A, Pathways for the cir-
culation of Ag in the LN. 1, Comple-
ment C3 opsonizes Ag in the presence of
Ab. C3-ICs are formed by the deposi-
tion of complement proteins and IgG
on the surface of the Ag. 2, The reten-tion of C3-ICs on the surface of the
SSMs is CR3 and FcRIIb dependent.
3, B cells transport C3-ICs from the sur-
face of the SSMs to the FDCs in a
CD21-dependent manner. 4, C3-ICs aretransferred to FDCs. 5a, Cognate B cells
acquire small Ag drained via FRC con-
duits directly or (5b) from the FDC sur-
face. B, Influenza virus uptake by SSMs
is MBL dependent.
2660 BRIEF REVIEWS: COMPLEMENT IN ADAPTIVE IMMUNITY
by guest on Novem
ber 27, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
attached to Ags (19, 23–25). They play a critical role inhumoral immunity because blocking with Abs (26, 27), asoluble receptor (28), or deletion of the receptors (29, 30)leads to impaired humoral immunity to model protein Ags,bacteria (31), and viruses (32, 33) (Fig. 3). On B cells,CD21/35 forms a coreceptor with CD19 and CD81, andcoligation with the BCR lowers the threshold of B cell acti-vation (34–36).A second major function of CD21/35 is retention of Ag on
FDCs. They represent the major Ag receptors on FDCs (37–39). Ag is also retained for long periods via FcRIIb; however,its expression is not constitutive but upregulated on activatedFDCs (40). FDCs are thought to be the major source of Agrequired for clonal selection of B cells during the germinalcenter (GC) reaction (41) and possibly for maintenance ofmemory (42–44). They retain Ag and secrete B cell growthfactors and the B cell attractant CXCL-13, as mentionedabove (45). Mice deficient in C3 or CD21/35 have fewerand smaller GCs, and B cells fail to survive (46, 47).There is growing evidence that B cells acquire Ag and are
activated most efficiently when it is attached to membrane (48–50). Therefore, retention of Ag and C3d on the FDC surfacecould enhance formation of the B cell synapse. Support for arole for FDCs as an important source of Ag comes from therecent studies of Cyster et al. (51); they demonstrated directcapture of Ag from FDC surface by cognate B cells usingmultiphoton intravital microscopy (MP-IVM). Importantly,they also identified a role for CD21/35 in efficient uptake ofmedium-affinity Ags. In their model system, MD4 B cells defi-cient in CD21/35 acquired less Ag from FDCs. One explan-ation for their results is that the coreceptor acts to enhanceBCR signaling in the synapse between the BCR and FDC.A third novel role for CD21/35 is transport of lymph-borne
C3-ICs into the B cell follicles and transfer to FDCs. As
discussed above, naive B cells acquire C3-ICs from SSMs liningthe SCS via their CD21/35 receptors. Whether uptake inducesa direct signal via CD21/35 or involves the coreceptor CD19and CD81 is not clear. In a recent study using mice in whichCD21/35 are uncoupled from CD19 (Cr2D/D), naive B cellsin the mutant line seemed to take up C3-ICs normally anddeliver them to FDCs (42). CD21/35 receptors include a cyto-plasmic domain so it is possible that uptake of C3-ICs inducesa signal by CD21, independent of CD19 (52), and triggersB cell migration to the FDCs. An alternative possibility isthat B cells within the follicles migrate constitutively to FDCsbased on a chemokine gradient in a manner similar to thatdescribed for MZ B cells (53); therefore, direct signaling byC3-ICs may not be essential. Thus, as B cells enter the periph-eral LNs via high endothelial cells, they cycle to the SCSarea, which is rich in marginal reticular cells that secreteB cell chemokines (54), and subsequently to FDCs, whichare also a source of chemokines (45).Earlier studies identified a role for C3 and CD21/35 in the
capture of ICs by MZ B cells and transport to FDCs withinB cell follicles (55–57). Using intravital imaging, Cyster andcolleagues (53) identified a similar pathway for capture ofC3-ICs in the spleen as reported for the LNs. They found thatMZ B cells continuously shuttled between the splenic MZand the B cell follicles by a CXCR5-chemokine-sphingosine1-phosphate receptor-dependent mechanism. Thus, naiveMZ B cells capture C3-coated Ags within the MZ sinusand transport them into the follicles where they are trans-ferred to FDCs. How C3-ICs are handed off to FDCs isnot clear; however, uptake on the FDCs is dependent onCD21/35 and FcRIIb. Because the latter receptors (FcRIIb)are not constitutively expressed but are upregulated followingFDC activation, complement receptors are critical for theinitial capture and retention of C3-ICs (40).
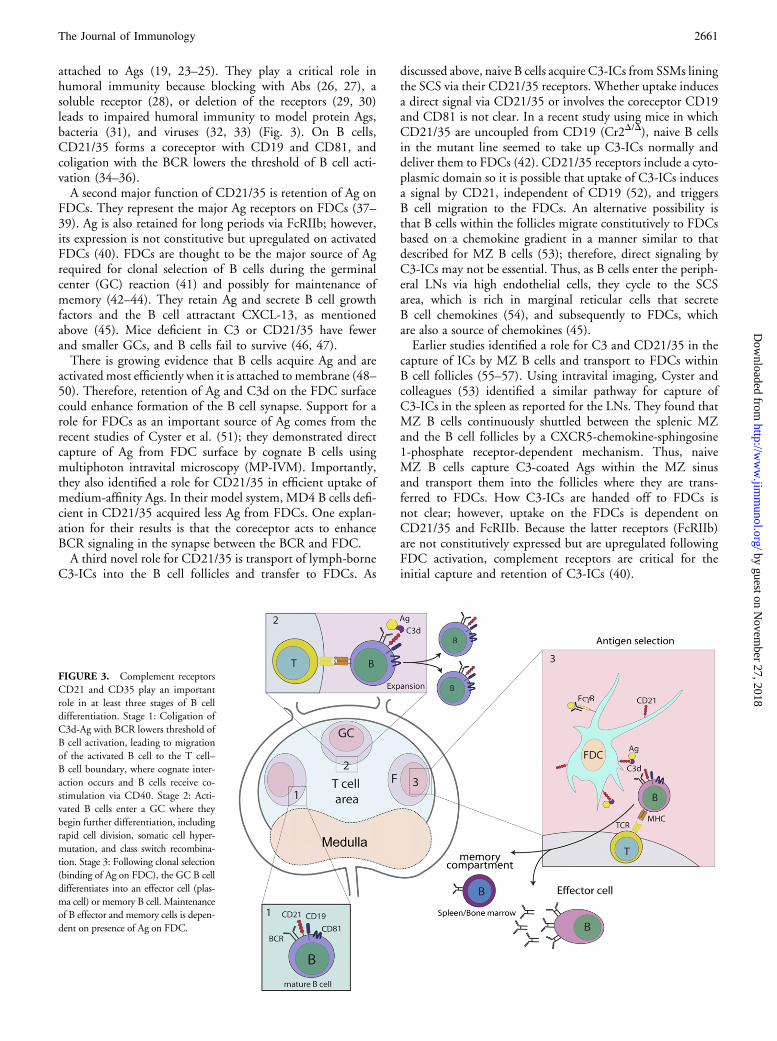
FIGURE 3. Complement receptors
CD21 and CD35 play an important
role in at least three stages of B cell
differentiation. Stage 1: Coligation of
C3d-Ag with BCR lowers threshold of
B cell activation, leading to migration
of the activated B cell to the T cell–
B cell boundary, where cognate inter-
action occurs and B cells receive co-
stimulation via CD40. Stage 2: Acti-
vated B cells enter a GC where they
begin further differentiation, including
rapid cell division, somatic cell hyper-
mutation, and class switch recombina-
tion. Stage 3: Following clonal selection
(binding of Ag on FDC), the GC B cell
differentiates into an effector cell (plas-
ma cell) or memory B cell. Maintenance
of B effector and memory cells is depen-
dent on presence of Ag on FDC.
The Journal of Immunology 2661
by guest on Novem
ber 27, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

Influenza virus is captured via a novel pathway
Influenza infection represents a major pathogen of humans; ingeneral it is controlled by annual vaccination and induction ofa protective humoral response (58–60). Complement C3 andCD21/35 are important components in humoral immunity toinfluenza, based on earlier studies (33, 61, 62). Although muchof the focus on the enhancing role of complement has been onthe B cell coreceptor, the C3-CD21/35 pathway is also importantfor transport and retention of Ag on FDCs, as discussed above.Recent studies identified a novel pathway by which viral Ag is
captured and possibly transported to the FDCs. Gonzalez et al.(63) used a UV-inactivated form of influenza virus (strain PR8)as a model vaccine. Using a fluorescent-tagged form of UV-PR8 injected in the footpad (s.c.) and imaging by MP-IVM,they identified rapid filling of the sinus and capture by SSMsand MMs. As discussed earlier, macrophages lining theseregions represent distinct populations, as distinguished byfunctional maturity and cell surface markers (17).The study by Gonzalez et al. (63) reported that labeled
UV-PR8 virus was rapidly bound by the SSMs and MMs,similar to that reported for vesicular stomatitis virus (13). Incontrast to the earlier studies, UV-PR8 was not retained on thesurface but was internalized. Binding and phagocytosis wasdependent on MBL, because SSMs in MBL2/2 mice failedto bind the virus. Opsonization by MBL is in agreement withearlier studies by Ezekowitz and colleagues (64), who reportedthat guinea pig mannose-binding lectin bound and neutralizedinfluenza. Interestingly, blocking of UV-PR8 uptake by SSMsalone or SSMs and MMs did not impair humoral immunity.This was somewhat surprising because earlier reports suggestedthat capture of particulate Ags by SSMs was important foractivation of cognate B cells. However, capture of UV-PR8by sinus-lining macrophages was important in limiting thespread of the virus to downstream LNs and the spleen. Thus,the role of macrophages lining the sinus of peripheral LNs isimportant in the efficient capture and spread of virus, but it isnot important for humoral immunity to inactive influenza.It is not clear howMBL-opsonizedUV-PR8 is phagocytosedby
SSMs. Because a known receptor for murine MBL is not clear itis possible that uptake is via a complement ligand, because thelectin pathway activates C2 and C4 or C3 directly via a bypasspathway, as described in human serum (Fig. 2B) (65).
Medullary dendritic cells bind influenza
In addition to capture of virus by sinus-lining macrophages,Gonzalez et al. (63) reported that a resident population of
dendritic cells (DCs) bound virus in themedullary region.Usingflow-cytometric assays, they identified a CD11c+ CD11bhi,SIGN-R1+ population that bound a significantly large amountof virus. Interestingly,∼50% of the PR8-bindingDCs expresseda receptor for the cysteine-rich (CR) domain of the MR basedon binding with an Ig Fc fusion protein with the CR domain(i.e., CR-Fc) (66). The receptor for the CR domain of MR isassociated with dendritic-like cells that accumulate within theB cell follicles following immunization and possibly representcells transporting Ag (67).Studies by Park and colleagues (68) identified murine SIGN-
R1 as the major receptor in the uptake of Streptococcus pneumo-niae by MZ macrophages. SIGN-R1 is structurally similar tohuman DC-SIGN and is a C-type lectin that binds glycans richin mannose, such as dextran and capsular polysaccharides ofpneumococcus (69, 70). It is the major receptor for S. pneumo-niae, because pretreatment of mice with a monoclonal Ab(clone 22D1) that downregulates macrophage expression ofSIGN-R1 led to an impaired humoral response and increasedmortality (68). Notably, binding of bacteria or purified poly-saccharide by SIGN-R1 induces receptor aggregation, activa-tion of C1q, and C3 deposition (68). Mice deficient in C3have impaired immunity to S. pneumoniae, and this is associ-ated with impaired uptake of bacteria on FDCs. Thus, oneinterpretation of the findings is that opsonization of bacteriawith C3 via SIGN-R1 is critical for uptake on FDCs and accessby follicular B cells.
SIGN-R1 is required for local humoral immunity to influenza
Using a similar approach as Park and colleagues (68), Gonzalezet al. (63) showed that treatment of mice with a mAb toSIGN-R1 that downregulates receptor expression significantlyreduced the uptake of UV-PR8 by resident DCs. Moreover,blocking of UV-viral uptake via SIGN-R1 in MBL2/2 micesubstantially reduced the capture of UV-PR8 in the drainingLN, and the local B cell response was reduced by 3-fold.However, in these experiments, the systemic response wasnot affected, suggesting that blocking of viral capture locallyled to efficient spreading of the virus and response in down-stream LNs and spleen. Further support for the importance ofthe resident DCs in viral capture and humoral immunitywas determined by the elimination of DCs systemically inCD11c-DTR bone marrow chimeric mice (71). In this model,CD11chi cells express the monkey diphtheria toxin receptor andare sensitive to ablation on treatment with diphtheria toxin.Thus, the CD11c DC-ablated chimeric mice had a significantly
FIGURE 4. Lymph-borne UV-inactive influenza (strain
PR8) fills the SCS of the LN, where it is partially taken
up by SSMs. Binding of virus to SSM is MBL dependent.
Virions are channeled to the LN medulla, where MMs and
resident DCs capture them in a SIGN-R1–dependent
manner. Model speculates that C1q is activated by SIGN-
R1, leading to C3 activation and deposition on PR8. Sub-
sequently, resident DCs transport C3-coated virus complex
to the follicles, where it is transferred to FDCs.
2662 BRIEF REVIEWS: COMPLEMENT IN ADAPTIVE IMMUNITY
by guest on Novem
ber 27, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

reduced CD4 T cell-dependent (IgG) and -independent (IgM)day-10 humoral response to UV-PR8. Because the primaryIgM response to inactive influenza under the conditions usedin the study was largely independent of CD4 T cells (72) (M.C.Carroll, unpublished observations), the results support a re-quirement for DCs in the transport of Ag to the B cell compart-ment. It is noted that under different conditions, in which miceare infected with live influenza virus, the primary response ispartly T cell dependent (73).Additional support for a role for resident DCs in the
transport of UV-PR8 into the B cell compartment came fromMP-IVM analysis of CD11c-EYFP mice injected with labeledinfluenza. Tracking of CD11chi cells before and after injectionof labeled virus revealed that only those cells that bound virusmoved in a nonrandom manner and that their net displace-ment and velocity were significantly increased relative toneighboring DCs that did not bind virus. Moreover, calcula-tion of the net vector of directionality (i.e., net overall direc-tion of migration) indicates that DCs that bind UV-PR8migrate toward the B cell follicles (63).It is tempting to speculate that capture of influenza by
SIGN-R1 on theDC surface leads to activation ofC1q and dep-osition of C3 on the inactive virus, as reported with S. pneumo-nia. Thus, it will be important in future studies to learnwhether LN-resident DCs deliver C3d-coupled inactive virusto the FDCs, much like that identified for naive B cells (Fig. 4).
ConclusionsIn this review, we discussed a novel role for complement in thetransport of soluble Ags, ICs, and viral Ags to the B cell com-partment. The finding that complement C3 and its receptors(i.e.,CD21/35 andCR3 [CD11b/18]) participate in the captureand transport of lymph-borne Ags to the FDCs provides anotherimportant mechanism by which complement enhances adaptiveimmunity. Moreover, the identification of a network of con-duits that directly funnel small Ags to the FDCs further eluci-dates howAgs aremade accessible to naive B cells.We also high-light a new pathway in which capture of inactive influenza viathe SIGN-R1 receptor signals the resident DCs to migrate to-ward the B cell compartment. It will be important, in future ex-periments, to determine whether C3 is activated and depositedon the virus following binding by SIGN-R1 and to track residentDCs with bound PR8 into the B cell follicles and determinewhether they hand-off viral Ag directly to FDCs (Fig. 4).
AcknowledgmentsWe thank Alex Gillmore for assistance with assembly of the manuscript and
figures and members of the laboratory for discussions on Ag trafficking.
DisclosuresThe authors have no financial conflicts of interest.
References1. von Andrian, U. H., and T. R. Mempel. 2003. Homing and cellular traffic in lymph
nodes. Nat. Rev. Immunol. 3: 867–878.2. Bajenoff, M., J. G. Egen, L. Y. Koo, J. P. Laugier, F. Brau, N. Glaichenhaus, and
R. N. Germain. 2006. Stromal cell networks regulate lymphocyte entry, migration,and territoriality in lymph nodes. Immunity 25: 989–1001.
3. Mempel, T. R., T. Junt, and U. H. von Andrian. 2006. Rulers over randomness:stroma cells guide lymphocyte migration in lymph nodes. Immunity 25: 867–869.
4. Anderson, A.O., andN.D. Anderson. 1975. Studies on the structure and permeabilityof the microvasculature in normal rat lymph nodes. Am. J. Pathol. 80: 387–418.
5. Gretz, J. E., A. O. Anderson, and S. Shaw. 1997. Cords, channels, corridors andconduits: critical architectural elements facilitating cell interactions in the lymphnode cortex. Immunol. Rev. 156: 11–24.
6. Gretz, J. E., C. C. Norbury, A. O. Anderson, A. E. Proudfoot, and S. Shaw. 2000.Lymph-borne chemokines and other low molecular weight molecules reach high endo-thelial venules via specialized conduits while a functional barrier limits access tothe lymphocyte microenvironments in lymph node cortex. J. Exp.Med. 192: 1425–1440.
7. Sixt, M., N. Kanazawa, M. Selg, T. Samson, G. Roos, D. P. Reinhardt, R. Pabst,M. B. Lutz, and L. Sorokin. 2005. The conduit system transports soluble antigensfrom the afferent lymph to resident dendritic cells in the T cell area of the lymphnode. Immunity 22: 19–29.
8. Anderson, A. O., and S. Shaw. 2005. Conduit for privileged communications in thelymph node. Immunity 22: 3–5.
9. Roozendaal, R., T. R. Mempel, L. A. Pitcher, S. F. Gonzalez, A. Verschoor,R. E. Mebius, U. H. von Andrian, and M. C. Carroll. 2009. Conduits mediatetransport of low-molecular-weight antigen to lymph node follicles. Immunity 30:264–276.
10. Bajenoff, M., and R. N. Germain. 2009. B-cell follicle development remodels theconduit system and allows soluble antigen delivery to follicular dendritic cells. Blood114: 4989–4997.
11. Pape, K. A., D. M. Catron, A. A. Itano, and M. K. Jenkins. 2007. The humoralimmune response is initiated in lymph nodes by B cells that acquire soluble antigendirectly in the follicles. Immunity 26: 491–502.
12. Catron, D. M., K. A. Pape, B. T. Fife, N. van Rooijen, and M. K. Jenkins. 2010. Aprotease-dependent mechanism for initiating T-dependent B cell responses to largeparticulate antigens. J. Immunol. 184: 3609–3617.
13. Junt, T., E. A. Moseman, M. Iannacone, S. Massberg, P. A. Lang, M. Boes, K. Fink,S. E. Henrickson, D. M. Shayakhmetov, N. C. Di Paolo, et al. 2007. Subcapsularsinus macrophages in lymph nodes clear lymph-borne viruses and present them toantiviral B cells. Nature 450: 110–114.
14. Carrasco, Y. R., and F. D. Batista. 2007. B cells acquire particulate antigen in amacrophage-rich area at the boundary between the follicle and the subcapsular sinusof the lymph node. Immunity 27: 160–171.
15. Martinez-Pomares, L., and S. Gordon. 2007. Antigen presentation the macrophageway. Cell 131: 641–643.
16. Phan, T. G., I. Grigorova, T. Okada, and J. G. Cyster. 2007. Subcapsular encounterand complement-dependent transport of immune complexes by lymph node B cells.Nat. Immunol. 8: 992–1000.
17. Phan, T. G., J. A. Green, E. E. Gray, Y. Xu, and J. G. Cyster. 2009. Immunecomplex relay by subcapsular sinus macrophages and noncognate B cells drivesantibody affinity maturation. Nat. Immunol. 10: 786–793.
18. Bergtold, A., D. D. Desai, A. Gavhane, and R. Clynes. 2005. Cell surface recycling ofinternalized antigen permits dendritic cell priming of B cells. Immunity 23: 503–514.
19. Molina, H., T. Kinoshita, K. Inoue, J.-C. Carel, and V. M. Holers. 1990. Amolecular and immunochemical characterization of mouse CR2. Evidence for asingle gene model of mouse complement receptors 1 and 2. J. Immunol. 145:2974–2983.
20. Molina, H., W. Wong, T. Kinoshita, C. Brenner, S. Foley, and V. M. Holers. 1992.Distinct receptor and regulatory properties of recombinant mouse complementreceptor 1 (CR1) and Crry, the two genetic homologues of human CR1. J. Exp.Med. 175: 121–129.
21. Carroll, M. C., E. M. Alicot, P. J. Katzman, L. B. Klickstein, J. A. Smith, andD. T. Fearon. 1988. Organization of the genes encoding complement receptors type1 and 2, decay-accelerating factor, and C4-binding protein in the RCA locus onhuman chromosome 1. J. Exp. Med. 167: 1271–1280.
22. Ahearn, J. M., and D. T. Fearon. 1989. Structure and function of the complementreceptors, CR1 (CD35) and CR2 (CD21). Adv. Immunol. 46: 183–219.
23. Kalli, K. R., and D. T. Fearon. 1994. Binding of C3b and C4b by the CR1-like sitein murine CR1. J. Immunol. 152: 2899–2903.
24. Isenman, D. E. 1983. The role of the thioester bond in C3 and C4 in the deter-mination of the conformational and functional states of the molecule. Ann. N. Y.Acad. Sci. 421: 277–290.
25. Law, S. K., A. W. Dodds, and R. R. Porter. 1984. A comparison of the properties oftwo classes, C4A and C4B, of the human complement component C4. EMBO J. 3:1819–1823.
26. Gustavsson, S., T. Kinoshita, and B. Heyman. 1995. Antibodies to murine com-plement receptor 1 and 2 can inhibit the antibody response in vivo withoutinhibiting T helper cell induction. J. Immunol. 154: 6524–6528.
27. Heyman, B., E. J. Wiersma, and T. Kinoshita. 1990. In vivo inhibition of theantibody response by a complement receptor-specific monoclonal antibody. J. Exp.Med. 172: 665–668.
28. Hebell, T., J. M. Ahearn, and D. T. Fearon. 1991. Suppression of the immuneresponse by a soluble complement receptor of B lymphocytes. Science 254: 102–105.
29. Ahearn, J. M., M. B. Fischer, D. Croix, S. Goerg, M. Ma, J. Xia, X. Zhou,R. G. Howard, T. L. Rothstein, and M. C. Carroll. 1996. Disruption of the Cr2locus results in a reduction in B-1a cells and in an impaired B cell responseto T-dependent antigen. Immunity 4: 251–262.
30. Molina, H., V. M. Holers, B. Li, Y. Fung, S. Mariathasan, J. Goellner, J. Strauss-Schoenberger, R. W. Karr, and D. D. Chaplin. 1996. Markedly impaired humoralimmune response in mice deficient in complement receptors 1 and 2. Proc. Natl.Acad. Sci. USA 93: 3357–3361.
31. Haas, K. M., M. Hasegawa, D. A. Steeber, J. C. Poe, M. D. Zabel, C. B. Bock,D. R. Karp, D. E. Briles, J. H. Weis, and T. F. Tedder. 2002. Complement receptorsCD21/35 link innate and protective immunity during Streptococcus pneumoniaeinfection by regulating IgG3 antibody responses. Immunity 17: 713–723.
32. Da Costa, X. J., M. A. Brockman, E. Alicot, M. Ma, M. B. Fischer, X. Zhou,D. M. Knipe, and M. C. Carroll. 1999. Humoral response to herpes simplex virus iscomplement-dependent. Proc. Natl. Acad. Sci. USA 96: 12708–12712.
The Journal of Immunology 2663
by guest on Novem
ber 27, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

33. Fernandez Gonzalez, S., J. P. Jayasekera, and M. C. Carroll. 2008. Complementand natural antibody are required in the long-term memory response to influenzavirus. Vaccine 26(Suppl. 8): I86–I93.
34. Bradbury, L. E., G. S. Kansas, S. Levy, R. L. Evans, and T. F. Tedder. 1992. TheCD19/CD21 signal transducing complex of human B lymphocytes includes thetarget of antiproliferative antibody-1 and Leu-13 molecules. J. Immunol. 149: 2841–2850.
35. Dempsey, P. W., M. E. Allison, S. Akkaraju, C. C. Goodnow, and D. T. Fearon.1996. C3d of complement as a molecular adjuvant: bridging innate and acquiredimmunity. Science 271: 348–350.
36. Carter, R. H., and D. T. Fearon. 1992. CD19: lowering the threshold for antigenreceptor stimulation of B lymphocytes. Science 256: 105–107.
37. Carroll, M. C. 2004. The complement system in B cell regulation. Mol. Immunol.41: 141–146.
38. Fang, Y., C. Xu, Y. X. Fu, V. M. Holers, and H. Molina. 1998. Expression ofcomplement receptors 1 and 2 on follicular dendritic cells is necessary for thegeneration of a strong antigen-specific IgG response. J. Immunol. 160: 5273–5279.
39. Brockman, M. A., A. Verschoor, J. Zhu, M. C. Carroll, and D. M. Knipe. 2006.Optimal long-term humoral responses to replication-defective herpes simplex virusrequire CD21/CD35 complement receptor expression on stromal cells. J. Virol. 80:7111–7117.
40. El Shikh, M. E., R. El Sayed, A. K. Szakal, and J. G. Tew. 2006. Folliculardendritic cell (FDC)-FcgammaRIIB engagement via immune complexes induces theactivated FDC phenotype associated with secondary follicle development. Eur. J.Immunol. 36: 2715–2724.
41. Rajewsky, K. 1996. Clonal selection and learning in the antibody system. Nature381: 751–758.
42. Barrington, R. A., T. J. Schneider, L. A. Pitcher, T. R. Mempel, M. Ma,N. S. Barteneva, and M. C. Carroll. 2009. Uncoupling CD21 and CD19 ofthe B-cell coreceptor. Proc. Natl. Acad. Sci. USA 106: 14490–14495.
43. Barrington, R. A., O. Pozdnyakova, M. R. Zafari, C. D. Benjamin, andM. C. Carroll. 2002. B lymphocyte memory: role of stromal cell complement andFcgammaRIIB receptors. J. Exp. Med. 196: 1189–1199.
44. Verschoor, A., M. A. Brockman, D. M. Knipe, and M. C. Carroll. 2001. Cuttingedge: myeloid complement C3 enhances the humoral response to peripheral viralinfection. J. Immunol. 167: 2446–2451.
45. Allen, C. D., and J. G. Cyster. 2008. Follicular dendritic cell networks of primaryfollicles and germinal centers: phenotype and function. Semin. Immunol. 20: 14–25.
46. Fischer, M. B., M. Ma, S. Goerg, X. Zhou, J. Xia, O. Finco, S. Han, G. Kelsoe,R. G. Howard, T. L. Rothstein, et al. 1996. Regulation of the B cell response toT-dependent antigens by classical pathway complement. J. Immunol. 157: 549–556.
47. Fischer, M. B., S. Goerg, L. Shen, A. P. Prodeus, C. C. Goodnow, G. Kelsoe, andM. C. Carroll. 1998. Dependence of germinal center B cells on expression ofCD21/CD35 for survival. Science 280: 582–585.
48. Batista, F. D., D. Iber, and M. S. Neuberger. 2001. B cells acquire antigen fromtarget cells after synapse formation. Nature 411: 489–494.
49. Carrasco, Y. R., S. J. Fleire, T. Cameron, M. L. Dustin, and F. D. Batista. 2004.LFA-1/ICAM-1 interaction lowers the threshold of B cell activation by facilitatingB cell adhesion and synapse formation. Immunity 20: 589–599.
50. Randall, K. L., T. Lambe, A. Johnson, B. Treanor, E. Kucharska, H. Domaschenz,B. Whittle, L. E. Tze, A. Enders, T. L. Crockford, et al. 2009. Dock8 mutationscripple B cell immunological synapses, germinal centers and long-lived antibodyproduction. Nat. Immunol. 10: 1283–1291.
51. Suzuki, K., I. Grigorova, T. G. Phan, L. M. Kelly, and J. G. Cyster. 2009. Visu-alizing B cell capture of cognate antigen from follicular dendritic cells. J. Exp. Med.206: 1485–1493.
52. Bouillie, S., M. Barel, and R. Frade. 1999. Signaling through the EBV/C3d receptor(CR2, CD21) in human B lymphocytes: activation of phosphatidylinositol 3-kinasevia a CD19-independent pathway. J. Immunol. 162: 136–143.
53. Cinamon, G., M. A. Zachariah, O. M. Lam, F. W. Foss Jr., and J. G. Cyster. 2008.Follicular shuttling of marginal zone B cells facilitates antigen transport. Nat.Immunol. 9: 54–62.
54. Katakai, T., H. Suto, M. Sugai, H. Gonda, A. Togawa, S. Suematsu, Y. Ebisuno,K. Katagiri, T. Kinashi, and A. Shimizu. 2008. Organizer-like reticular stromal celllayer common to adult secondary lymphoid organs. J. Immunol. 181: 6189–6200.
55. Ferguson, A. R., M. E. Youd, and R. B. Corley. 2004. Marginal zone B cellstransport and deposit IgM-containing immune complexes onto folliculardendritic cells. Int. Immunol. 16: 1411–1422.
56. Youd, M. E., A. R. Ferguson, and R. B. Corley. 2002. Synergistic roles of IgM andcomplement in antigen trapping and follicular localization. Eur. J. Immunol. 32:2328–2337.
57. Pozdnyakova, O., H. K. Guttormsen, F. N. Lalani, M. C. Carroll, andD. L. Kasper. 2003. Impaired antibody response to group B streptococcal type IIIcapsular polysaccharide in C3- and complement receptor 2-deficient mice. J. Immu-nol. 170: 84–90.
58. Gerhard, W., K. Mozdzanowska, M. Furchner, G. Washko, and K. Maiese. 1997.Role of the B-cell response in recovery of mice from primary influenza virus infec-tion. Immunol. Rev. 159: 95–103.
59. Palladino, G., K. Mozdzanowska, G. Washko, and W. Gerhard. 1995. Virus-neutralizing antibodies of immunoglobulin G (IgG) but not of IgM or IgA isotypescan cure influenza virus pneumonia in SCID mice. J. Virol. 69: 2075–2081.
60. Skehel, J. 2009. An overview of influenza haemagglutinin and neuraminidase.Biologicals 37: 177–178.
61. Ross, T. M., Y. Xu, R. A. Bright, and H. L. Robinson. 2000. C3d enhancement ofantibodies to hemagglutinin accelerates protection against influenza virus challenge.Nat. Immunol. 1: 127–131.
62. Watanabe, I., T. M. Ross, S. Tamura, T. Ichinohe, S. Ito, H. Takahashi, H. Sawa,J. Chiba, T. Kurata, T. Sata, and H. Hasegawa. 2003. Protection against influenzavirus infection by intranasal administration of C3d-fused hemagglutinin. Vaccine21: 4532–4538.
63. Gonzalez, S. F., V. Lukacs-Kornek, M. P. Kuligowski, L. A. Pitcher, S. E. Degn,Y.-A. Kim, M. Cloninger, L. Martinez-Pomares, S. Gordon, S. J. Turley, andM. C. Carroll. 2010. Capture of influenza by medullary dendritic cells viaSIGN-R1 is essential for humoral immunity in draining lymph nodes. Nat. Immu-nol. 11:427–434.
64. Anders, E. M., C. A. Hartley, P. C. Reading, and R. A. Ezekowitz. 1994. Com-plement-dependent neutralization of influenza virus by a serum mannose-bindinglectin. J. Gen. Virol. 75: 615–622.
65. Selander, B., U. Martensson, A. Weintraub, E. Holmstrom, M. Matsushita,S. Thiel, J. C. Jensenius, L. Truedsson, and A. G. Sjoholm. 2006. Mannan-bindinglectin activates C3 and the alternative complement pathway without involvement ofC2. J. Clin. Invest. 116: 1425–1434.
66. Martınez-Pomares, L., M. Kosco-Vilbois, E. Darley, P. Tree, S. Herren,J. Y. Bonnefoy, and S. Gordon. 1996. Fc chimeric protein containing thecysteine-rich domain of the murine mannose receptor binds to macrophages fromsplenic marginal zone and lymph node subcapsular sinus and to germinal centers. J.Exp. Med. 184: 1927–1937.
67. Berney, C., S. Herren, C. A. Power, S. Gordon, L. Martinez-Pomares, andM. H. Kosco-Vilbois. 1999. A member of the dendritic cell family that entersB cell follicles and stimulates primary antibody responses identified by a mannosereceptor fusion protein. J. Exp. Med. 190: 851–860.
68. Kang, Y. S., Y. Do, H. K. Lee, S. H. Park, C. Cheong, R. M. Lynch, J. M. Loeffler,R. M. Steinman, and C. G. Park. 2006. A dominant complement fixation pathwayfor pneumococcal polysaccharides initiated by SIGN-R1 interacting with C1q. Cell125: 47–58.
69. Geijtenbeek, T. B., P. C. Groot, M. A. Nolte, S. J. van Vliet, S. T. Gangaram-Panday, G. C. van Duijnhoven, G. Kraal, A. J. van Oosterhout, and Y. van Kooyk.2002. Marginal zone macrophages express a murine homologue of DC-SIGN thatcaptures blood-borne antigens in vivo. Blood 100: 2908–2916.
70. Kang, Y. S., S. Yamazaki, T. Iyoda, M. Pack, S. A. Bruening, J. Y. Kim,K. Takahara, K. Inaba, R. M. Steinman, and C. G. Park. 2003. SIGN-R1, anovel C-type lectin expressed by marginal zone macrophages in spleen, mediatesuptake of the polysaccharide dextran. Int. Immunol. 15: 177–186.
71. Jung, S., D. Unutmaz, P. Wong, G. Sano, K. De los Santos, T. Sparwasser, S. Wu,S. Vuthoori, K. Ko, F. Zavala, et al. 2002. In vivo depletion of CD11c+dendritic cells abrogates priming of CD8+ T cells by exogenous cell-associatedantigens. Immunity 17: 211–220.
72. Sha, Z., and R. W. Compans. 2000. Induction of CD4(+) T-cell-independentimmunoglobulin responses by inactivated influenza virus. J. Virol. 74: 4999–5005.
73. Lee, B. O., J. Rangel-Moreno, J. E. Moyron-Quiroz, L. Hartson, M. Makris,F. Sprague, F. E. Lund, and T. D. Randall. 2005. CD4 T cell-independent antibodyresponse promotes resolution of primary influenza infection and helps to preventreinfection. J. Immunol. 175: 5827–5838.
2664 BRIEF REVIEWS: COMPLEMENT IN ADAPTIVE IMMUNITY
by guest on Novem
ber 27, 2018http://w
ww
.jimm
unol.org/D
ownloaded from





























