Embed Size (px)
Citation preview

1251
Conservation Biology, Pages 1251–1260Volume 17, No. 5, October 2003
Conservation in Conflict: the Tale of Two Endangered Species
GARY W. ROEMER* AND ROBERT K. WAYNE†
*Department of Fishery and Wildlife Sciences, New Mexico State University, P.O. Box 30003, MSC 4901 Las Cruces, NM 88003-8003, U.S.A., email [email protected]†Department of Organismic Biology, Ecology, & Evolution (OBEE), University of California, 621 Charles E. Young Drive South, Los Angeles, CA 90095–1606, U.S.A.
Abstract:
With its umbrella of provisions, the U.S. Endangered Species Act ( ESA) provides critical protectionto threatened or endangered wildlife. It provides minimal guidance, however, on identifying taxa worthy ofconservation, lacks guidelines for resolving endangered species conflicts, and subsequent recovery programsoften focus on the species rather than the ecosystem. These deficiencies are exemplified by the recovery pro-gram for the San Clemente Loggerhead Shrike (
Lanius ludovicianus mearnsi
) and the recent proposed rule togrant federal protection to one of its predators, the island fox (
Urocyon littoralis
). Recovery actions that haveincluded euthanasia of foxes have likely contributed to a 40–60% decline in the population size of the SanClemente Island fox (
U. l. clementae
), a subspecies listed by the state of California as threatened. The U.S. Fishand Wildlife Service proposes to list four other subspecies of the island fox as endangered but excludes the SanClemente Island fox and the sixth and last subspecies, the San Nicolas Island fox (
U. l. dickeyi
), ignoring theirevolutionary distinctness and the recent decline in population size of
U. l. clementae.
Using published mor-phological and genetic information, we show that the shrike’s current taxonomic and legal status should bereevaluated. We also reexamine the current recovery program for the shrike and conclude that the implemen-tation of the ESA’s provisions to protect the shrike was species-centric. The shrike recovery program is primar-ily centered on two approaches: the release of captive-bred shrikes and control of native and non-native pred-ators. The predator control program has contributed to the endangerment of the distinct San Clemente Islandfox. Given that five of the six fox subspecies face extirpation, the proposed rule to list only four of the six as en-dangered is inadequate. This endangered-species conflict might have been avoided through a more balancedecosystem approach that considers the ecological role of all native taxa and strives to enhance habitats criticalto both the shrike and the fox.
Conservación en Conflicto: el Cuento de Dos Especies en Peligro
Resumen:
Con su normatividad general, el Acta de Especies en Peligro de E. U. A. (1973) (AEP) proporcionaprotección decisiva a vida silvestre amenazada o en peligro. Sin embargo, proporciona una orientaciónmínima para identificar taxones merecedores de conservación y aumenta las directrices para resolver conflic-tos con especies en peligro, y los subsecuentes programas de recuperación a menudo enfocan en la especie y noen el ecosistema. Estas deficiencias están ejemplificadas por el programa de recuperación para el alcaudón(
Lanius ludovicianus mearnsi
) y la recientemente propuesta ley para otorgar protección federal a uno de susdepredadores, el zorro (
Urocyon littoralis
). Las acciones de recuperación, que han incluido la eutanasia dezorros, probablemente han contribuido a la declinación del 40–60% de la población del zorro de San Cle-
mente (
Urocyon l. clementae
), una subespecie listada como amenazada por el estado de California. El Serviciode Pesca y Vida Silvestre de E. U. A. propone listar otras cuatro subespecies de
Urocyon littoralis
como en peli-gro pero excluye a
Urocyon l. clementae
y la sexta y última subespecie,
U. l. dickeyi,
ignorando su singular-idad evolutiva y la reciente declinación de la población de
Urocyon l. clementae.
Utilizando informaciónmorfológica y genética publicada, mostramos que el estatus taxonómico y legal de
Lanius ludovicianusmearnsi
deben ser reevaluados. También reexaminamos el actual programa de recuperación para
Lanius lu-
Paper submitted May 13, 2002; revised manuscript accepted January 15, 2003.

1252
Conservation in Conflict Roemer & Wayne
Conservation BiologyVolume 17, No. 5, October 2003
dovicianus mearnsi
y concluimos que la instrumentación de las disposiciones de AEP para proteger a la espe-cie son especie-céntricas. El programa de recuperación de
Lanius ludovicianus
mearnsi se centra principal-mente en dos tratamientos: la liberación de alcaudones criados en cautiverio y el control de depredadoresnativos y no nativos. El programa de control de depredadores ha contribuido a poner en peligro a
Urocyon l.clementae.
Dado que cinco de las seis subespecies de zorro encaran la extirpación, la propuesta ley de listarsolo cuatro de las seis como en peligro es inadecuada. Este conflicto de especies en peligro pudo haberse evi-tado con un enfoque ecosistémico más balanceado que considere el papel ecológico de todos los taxones nati-
vos y se esfuerce por incrementar los hábitats críticos tanto para el alcaudón como para los zorros.
Introduction
The U.S. Endangered Species Act (1973) is widely con-sidered the most significant wildlife protection legisla-tion ever enacted ( National Research Council 1995 ).The ESA mandates that the Secretary of the Interior de-termine whether a species is endangered or threatenedwith extinction “solely on the basis of the best scientificand commercial data available . . . ” (U.S. Code. 1999.16 U.S.C. Section 1533). Once a species is listed, it isprotected by an umbrella of prohibitions, including Sec-tion 7, which “prohibits any federal action that will jeop-ardize the future of any endangered species, includingany threat to designated critical habitat” ( National Re-search Council 1995). However, the act and subsequentlegal opinions provide little guidance on how to identifytaxa worthy of conservation or how to resolve endangered-species conflicts, such as when one endangered speciespreys on another (National Research Council 1995). Theactions taken to recover the critically endangered San Cle-mente Loggerhead Shrike (
Lanius ludovicianus mearnsi
)on San Clemente Island, California, and the subsequent in-adequacy of a proposed rule to list one of its predators, theisland fox (
Urocyon littoralis
), as endangered are exam-ples of how biased implementation of the provisions ofthe ESA can lead to an endangered-species conflict.
In the case of the shrike recovery program, a species-centered philosophy was adopted that resulted in theimplementation of a comprehensive predator-control pro-gram that contributed to a decline in the San Clemente Is-land fox (
U. l. clementae
), a subspecies listed as threat-ened by the state of California (California Department ofFish and Game 1987). Four other subspecies of the islandfox (
U. l. littoralis
,
U. l. santarosae
,
U. l. santacruzae
,and
U. l. catalinae
) are already critically endangered (Roe-mer 1999; Timm et al. 2000; Roemer et al. 2003), are thefocus of an ecosystem-centered conservation strategy( Coonan 2001; Roemer et al. 2001
b
, 2002, 2003), andwere recently recommended to be listed as federally en-dangered by the U.S. Fish and Wildlife Service ( USFWS;U.S. Department of the Interior 2001 ). However, theproposed rule ignored the recent decline in the SanClemente Island fox and excluded both this subspe-
cies and the sixth and final subspecies, the San NicolasIsland fox (
U. l. dickeyi
), from potential federal protec-tion. Both of the islands harboring the excluded subspe-cies are the jurisdiction of the U.S. Navy. We (1) com-pare the evolutionary distinctiveness of the San ClementeLoggerhead Shrike to that of the island fox, with specialreference to the San Clemente and San Nicolas islandfoxes, (2) contrast the current species-centric recoveryprogram for the shrike with the potential benefits of anecosystem approach, and (3) examine the proposed ruleto list only four of six subspecies of the island fox andcomment on its efficacy. We posit that the momentumof legal, institutional, and public support for conserva-tion of the shrike has led to an endangered-species con-flict that has contributed to the endangerment of the en-demic island fox.
Evolutionary Distinctiveness of the San Clemente Loggerhead Shrike
First described by Ridgway (1903), the San Clemente Log-gerhead Shrike was later considered an endemic formfound only on San Clemente Island, California (Miller1931). In his taxonomic study of the Loggerhead Shrike,Miller (1931) described 11 different subspecies. In addi-tion to
L. l. mearnsi
, he also described another islandform,
L. l. anthonyi
, a subspecies found on three otherChannel Islands (Santa Catalina, Santa Cruz, and SantaRosa), and two mainland forms,
L. l. gambeli
and
L. l.grinnelli
, both found in California. In his classification,Miller (1931) used a method that relied heavily on thewing cord–to–tail length ratio ( WC:TL). However, theWC:TL is a poor character for classifying shrike subspe-cies; its use led to the misclassification of up to 39% ofshrike specimens (Collister & Wicklum 1996). Recently,Patten and Campbell (2000) applied a more rigorous ap-proach. They used nine plumage and mensural charac-ters coupled with a discriminant function analysis to as-sess the subspecific status of
L. l. mearnsi
. Historic samplesof
L. l. mearnsi
collected between 1897 and 1939 were dis-tinguishable from other shrike subspecies and were most

Conservation BiologyVolume 17, No. 5, October 2003
Roemer & Wayne Conservation in Conflict
1253
closely allied with
L. l. anthonyi
, the only other islandform. However, shrike specimens obtained from the re-cent San Clemente Loggerhead Shrike recovery programwere indistinguishable from
L. l. anthonyi
, promptingPatten and Campbell (2000) to conclude that “Our mor-phological analyses indicate that Loggerhead Shrikes onSan Clemente Island are no longer assignable to the
mearnsi
subspecies. Instead, they appear to be hybridsor intergrades between
mearnsi
and
anthonyi
. . . .”Molecular genetic analyses of current and historic popu-
lations of southern California Loggerhead Shrikes also havebeen used to assess the distinctiveness of
L. l. mearnsi
( Mundy et al. 1997
a
, 1997
b
). Mundy et al. (1997
a
) foundfrequency differences in mtDNA haplotypes among popu-lations of the two island subspecies (
L. l. mearnsi
and
L. l.anthonyi
) and two mainland populations of
L. l. gambeli
.Four different mtDNA haplotypes are recognized, and, al-though maximum pairwise sequence divergence amongpopulations is low (1.25%), strong genetic substructure isevident. The
F
ST
values are high for island-versus-mainlandcomparisons (0.54–1.0) and between
L. l. mearnsi
and
L. l.anthonyi
(0.61), indicating a low level of female-biasedgene flow among shrike populations (Mundy et al. 1997
a
).
L. l. mearnsi
has two haplotypes, one identical to that inmainland birds and the other the sole haplotype among thenine birds assayed from
L. l. anthonyi.
This suggests the ex-istence of an island-specific haplotype, but there are nohaplotypes unique to
L. l. mearnsi
.Mundy et al. (1997
b
) completed their investigation ofthe phylogeographic relationships among southern Cali-fornia shrike populations by comparing contemporary( 1995 ) and historic ( 1915 ) samples of
L. l. mearnsi
with contemporary samples from mainland populationsof
L. l. gambeli
. The contemporary population of
L. l.mearnsi
is depauperate in genetic variation comparedwith the mainland populations, having lower allelic di-versity and an expected heterozygosity (
H
e
) averaging60% that of mainland populations. Significant differ-ences in allele frequency among microsatellite loci alsooccur between the historic and contemporary islandsamples. The authors concluded that genetic variationwas low in
L. l. mearnsi
prior to the most recent declineand that genetic drift is probably the main evolutionarymechanism responsible for the low levels of variation inboth the historic and contemporary samples of
L. l.mearnsi
. The contemporary
L. l. mearnsi
sample hastwo microsatellite alleles that are not in the historic sam-ple, but, as with the mtDNA haplotypes, there are no mi-crosatellite alleles unique to
L. l. mearnsi
. Interestingly,all the historic samples of
L. l. mearnsi
have a singlemtDNA haplotype (haplotype A), whereas the contem-porary sample has two haplotypes ( haplotypes A and B)( Mundy et al. 1997
b
). The authors concluded that thesedifferences are likely a result of introgression, or geneflow, that occurred between
L. l. mearnsi
and another
shrike population, most likely
L. l. anthonyi
, in the past80 years.
Thus, both morphological and genetic evidencesuggest that gene flow likely occurred between
L. l.mearnsi
and
L. l. anthonyi
( Mundy et al. 1997
b
; Patten& Campbell 2000). Although Mundy et al. (1997
a
, 1997
b
)concluded that
L. l. mearnsi
was sufficiently distinct fromneighboring subspecies to warrant current conservationefforts, Patten and Campbell (2000) suggest that a moreprudent conservation strategy is to protect all the ChannelIsland shrike populations rather than focus conservationefforts only on San Clemente Island.
Mainland shrikes migrate to San Clemente Island, andcaptive-reared
L. l. mearnsi
are found at mainland locales( Juola et al. 1997; Mundy et al. 1997
a
, 1997
b
). Thesemovements, along with the evidence for introgression,are troubling because they strongly suggest that
L. l.mearnsi
is hybridizing with other island and mainlandshrikes and may no longer be a distinct form. The ESA doesnot extend protection to hybrids except in special cases(e.g., Florida panther [
Puma concolor coryi
]; NationalResearch Council 1995), and, although a draft intercrosspolicy for the ESA has been written, it has not beenadopted (U.S. Department of the Interior & Departmentof Commerce 1996). If the San Clemente LoggerheadShrike is interbreeding with other shrike subspecies,there is little precedence for continuing an intensive re-covery program that affects other native species, includ-ing clearly defined and critically endangered endemictaxa.
Evolutionary Distinctiveness of the Island Fox
Island foxes are a unique dwarf form endemic to the sixlargest of the eight California Channel Islands ( Moore &Collins 1995). Several researchers have explored the col-onization history, evolution, and phylogeography of is-land foxes by using morphological and molecular geneticevidence (Gilbert et al. 1990; Collins 1991
a
, 1991
b
, 1993;Wayne et al. 1991; Goldstein et al. 1999; Roemer et al.2001
a
).Collins (1993) used 29 cranial and mandibular charac-
ters measured from 2207 island and gray fox (
U.cinereoargenteus
) specimens to correctly classify 91%of all island fox specimens to their island of origin. Wayneet al. (1991) used seven restriction enzymes to exploremtDNA variation in both island and gray foxes. These twospecies do not share mtDNA haplotypes ( Table 1 ),whereas island foxes have a unique restriction site forrestriction enzyme
Hha
, a synapomorphy that clustersall subspecies into a monophyletic group (Wayne et al.1991). The San Clemente Island fox has a mtDNA haplo-type shared only with San Miguel Island foxes, suggestingthat the San Clemente population was derived from them
1254
Conservation in Conflict Roemer & Wayne
Conservation BiologyVolume 17, No. 5, October 2003
( Wayne et al. 1991). The San Nicolas Island fox has aunique mtDNA haplotype not found in any other islandfox subspecies (Table 1).
The genetic difference between the island fox and grayfox revealed by the mtDNA analyses is mirrored by analy-ses of nuclear DNA (Gilbert et al. 1990; Wayne et al. 1991;Goldstein et al. 1999). Of 19 microsatellite loci that arepolymorphic across the two species, 120 alleles are foundin the gray fox, and 53 (44%) of these are unique to grayfoxes (Table 1). Similarly, of 86 alleles in island foxes, 19(22%) are unique to them ( Table 1). Nei’s (1978) aver-age unbiased genetic distance between the gray fox andisland fox is 0.76. Phylogenetic analyses of these data con-sistently cluster all island fox subspecies together (only 2of 183 fox genotypes were misassigned), with the grayfox as the ancestral population, supporting the findingfrom the mtDNA analysis that island foxes are mono-phyletic. Importantly, the San Clemente Island fox con-tains both unique minisatellite and microsatellite allelesnot found in other island fox populations, and the SanNicolas Island fox also contains unique microsatellite al-leles despite its low genetic variability (Gilbert et al. 1990;Goldstein et al. 1999). The San Clemente Island fox haseight unique microsatellite alleles, and the San Nicolas Is-land fox has two (Table 1). From the standpoint of ge-netic diversity, the San Clemente Island fox and the SanNicolas Island fox are well-defined and important sub-species contributing to the overall genetic diversity ofthe species. Further, the morphological and genetic evi-dence support both the current classification of the is-land fox as a separate species (Wilson & Reeder 1993) andthe subspecific status of each island fox population ( Hall
1981; Moore & Collins 1995). Finally, island foxes fromdifferent subspecific populations would be unable to hy-bridize unless transfer among islands was facilitated by hu-mans (Collins 1991
a
, 1991
b
).
Conservation of the San Clemente Loggerhead Shrike and Decline of the San Clemente Island Fox
The San Clemente Loggerhead Shrike is one of the mostendangered birds in the world.
L. l. mearnsi
was re-duced to an estimated 17 individuals in the wild in 1988before a recovery program was initiated in 1991 ( U.S.Department of the Interior 1977; Juola et al. 1997). Theprogram is centered on the island, funded by the U.S.Navy, and supported by both public and private organi-zations.
The shrike recovery program has been successful atraising and releasing shrikes and establishing breedingpairs, but the population has remained extremely small.In July 2000, the wild population was estimated to be 25individuals and has never exceeded 35 individuals in thehistory of the program ( Mader et al. 2000; Blackford etal. 2001). Breeding failures attributed to nest predationhave plagued the recovery program and led to a plan formitigating threats to shrike nestlings and fledglings ( U.S.Department of Agriculture 1998). Although diet studiessuggest that birds typically form a very small portion( 3–6%) of the diet of the island fox (Laughrin 1977; Crooks& Van Vuren 1995), San Clemente Island foxes have beenobserved preying on nestling shrikes (Cooper et al. 2001).Mitigation included live-capture and removal or euthana-sia of foxes and euthanasia of native species of raptorsand non-native nest predators. This plan was endorsedby the USFWS and initiated by the U.S. Department of Ag-riculture’s division responsible for animal damage con-trol. As a result, in 1999 at least 32 foxes were euthanizedor permanently removed from the island to zoological in-stitutions. Other native species were also killed, includ-ing 105 Common Ravens (
Corvus corax
), 48 AmericanKestrels (
Falco sparverius
), 27 Red-tailed Hawks (
Buteojamaicensis
), and nine Barn Owls (
Tyto alba
) (Elliot &Popper 1999). After an article appeared in the
Los Ange-les Times
describing the control program (Schoch 1999),the U.S. Navy stopped killing foxes.
During the late 1990s, the U.S. Navy experimented withother means of predator control. They used a commer-cially available shock collar system to exclude foxes from alimited area surrounding shrike nest sites (Cooper et al.2001). A wire broadcasts a signal to a battery-poweredcollar that then delivers a shock to a collared fox. Thesystem worked well at nest sites because foxes werecontinually shocked until they left the vicinity of thenest. To exclude foxes from a larger area, where shrikefledglings might land, the USFWS required that release
Table 1. The total number and number of unique mtDNA haplotypes and microsatellite alleles found in gray foxes (
Urocyon cinereoargenteus
) and island foxes (
U. littoralis
).*
mtDNA haplotypes
Microsatellite loci
totalno.
unique totalno.
unique
Gray fox 7 7 120 53Island fox 5 5 86 19Island fox subspecies
San Clemente Island fox,
U. l. clementae
1 0 39 8Santa Catalina Island fox,
U. l. catalinae
3 1 49 2San Nicolas Island fox,
U. l. dickeyi
1 1 19 2Santa Cruz Island fox,
U. l. santacruzae
2 0 44 2Santa Rosa Island fox,
U. l. santarosae 2 0 47 7San Miguel Island fox,
U. l. littoralis 2 0 31 2
*Mitochondrial DNA data are from Wayne et al. (1991), and micro-satellite data are from G.W.R. and R.K.W. (unpublished data).
Conservation BiologyVolume 17, No. 5, October 2003
Roemer & Wayne Conservation in Conflict 1255
sites and canyons containing shrike nests also be wired.Foxes readily transgressed these wired areas because theywould no longer experience the shock deterrent aftermoving a short distance away from the perimeter wire.Further, a “three strikes” rule was established: foxes thattransgressed a wire-protected area three times would beheld in captivity or euthanized. In 1999, 49 foxes wereheld temporarily in small pens (0.55 m2) in addition tothe 32 foxes that were permanently removed or eutha-nized. Of the foxes held captive, 20 females were sus-pected of having dependent pups in the wild (Cooper etal. 2001). Adult female San Clemente Island foxes weanan average of 1.25 (SE � 0.015) pups per reproductiveevent ( Roemer 1999). Thus, an estimated 25 pups areexpected to have starved to death as a result of confine-ment of their mothers. In 2000 no foxes were euthanized,but 71 foxes, about 12% of the estimated population,were held in captivity for an average of 35 days (range6–114 days) during the fox reproductive season (Cooperet al. 2001).
Shock-wired canyons where shrikes nest are prime foxhabitat, and the consistent removal of foxes from theseareas created population sinks into which foxes wouldpersistently disperse (Cooper et al. 2001). Resident anddispersing foxes were essentially removed from the breed-ing pool, either by being placed in captivity or by repeatedcapture and confinement in traps. In 2000 alone 13,347box traps were set to capture foxes and non-native preda-tors in an area encompassing approximately two-thirds ofthe island, or about 100 km2 (Cooper et al. 2001). Thesetraps were set over 279 nights (76% of the year) and re-sulted in captures of 180 foxes 1460 times; one fox wascaptured 66 times (Cooper et al. 2001). Capture success(defined as the number of foxes captured divided by thenumber of traps set) of foxes on San Clemente Island in2000 (10.9%) was considerably lower than capture suc-cess in 1998 (36.3%) (Cooper et al. 2001; Roemer et al.2001b).
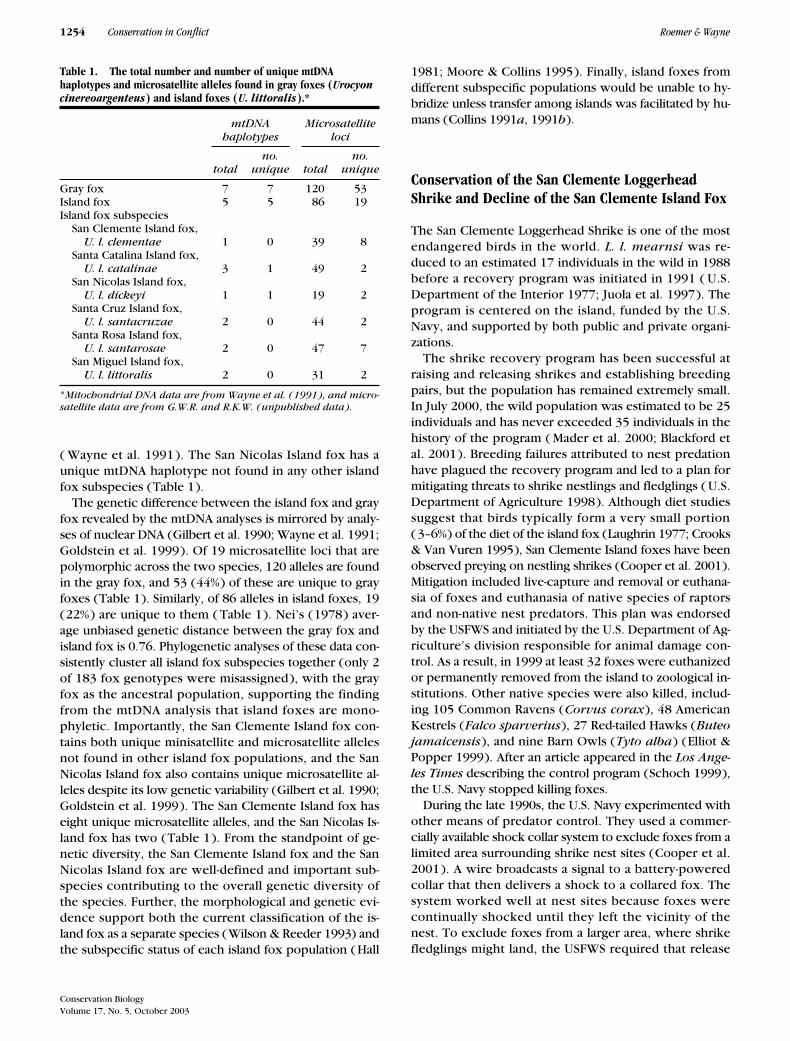
To assess the potential impact of the removal and con-finement strategies on the fox population, we constructeda Lefkovitch matrix based on 10 years of demographicdata (1988–1997) collected at three sampling sites on SanClemente Island (Roemer et al. 2000, 2003). This matrixyielded a population growth rate (� ) of 0.9566, or ap-proximately a 4.4% decline in annual population size.We used this rate of decline to estimate the trend in pop-ulation size from 1998 to 2002. We estimated that theSan Clemente Island fox population will be at approxi-mately 60% of its pre-1992 population size by 2002 ( Fig.1). A comparison of fox demographic data collected onSan Clemente Island suggests the decline may be evenmore significant. Average density, population size, andthe mean number of pups captured have declined bynearly 60% between 1988 and 2000 (Table 2). We sug-gest that the single-species philosophy that has charac-terized the San Clemente Loggerhead Shrike recovery
program has significantly affected the San Clemente Is-land fox and contributed to the endangerment of the is-land fox.
Status and Conservation of the Island Fox
Recently, the USFWS was petitioned by two nonprofit or-ganizations and subsequently proposed to list four sub-species of the island fox as endangered ( U.S. Departmentof the Interior 2001). These four subspecies, U. l. littora-lis, U. l. santarosae, U. l. santacruzae, and U. l. catali-nae, have declined drastically owing to hyperpredationby Golden Eagles (Aquila chrysaetos; Roemer et al. 2001b,2002) and infection with canine distemper virus (Timm etal. 2000). U. l. clementae and U. l. dickeyi were not in-
Table 2. Mean density (SE), estimated population size, and mean number of island fox pups captured on three demographic grids on San Clemente Island, California, between 1988 and 2000.
PeriodaDensity
( foxes/km2)Population
sizeb No. of pups
1988–1992 6.4 (0.41) 929 6.7 (0.68)1993–1997 5.3 (0.28) 771 4.3 (1.24)1999–2000 2.8 (0.71) 410 2.8 (1.01)aData from 1988 to 1997 are condensed from Roemer et al. (2000,2001b ), and data from 1999 and 2000 are from Schmidt et al.(2002). Trapping was not conducted in 1998.bPopulation size was simply estimated as the mean density multi-plied by island area (145 km2).
Figure 1. Estimated trend in island fox population size on San Clemente Island. Average population size (upper and lower 95% confidence interval, UCI and LCI, respectively) between 1988 and 1997 was esti-mated based on densities determined from a capture-recapture approach (Roemer et al. 1994, 2001b).Estimates from 1998 to 2002 (Est N) are based on a deterministic population model with vital rates de-rived from the previous 10 years of data (Roemer et al. 2000). In 1992, predator management to protect the shrike was initiated.

1256 Conservation in Conflict Roemer & Wayne
Conservation BiologyVolume 17, No. 5, October 2003
cluded in the proposed listing, despite a substantial popula-tion decline in U. l. clementae and evidence of morpho-logic and genetic distinction in both subspecies (Gilbertet al. 1990; Wayne et al 1991; Goldstein et al. 1999; Roe-mer et al. 2000, 2001b; U.S. Department of the Interior2001).
An independent assessment of the status of the islandfox argues that the entire species, not just select subspe-cies, be listed as endangered ( Roemer et al. 2003). Us-ing the World Conservation Union (IUCN) Red List cate-gories ( World Conservation Union 2001), Roemer et al(2003) recommended that the subspecies on San Migueland Santa Rosa islands, U. l. littoralis and U. l. santaro-sae, be listed as extinct in the wild. Extensive surveyshave found no evidence of wild foxes present on eitherSan Miguel or Santa Rosa islands, and there are, respec-tively, only 28 and 45 foxes currently in captivity (Coonan2001, 2002). The subspecies on Santa Cruz Island, U. l.santacruzae, and the subspecies on Santa Catalina Island,U. l. catalinae, are recommended to be listed as criticallyendangered. Each has been reduced in number by approx-imately 90% in the past 10 years, and each subspecies hasa population size of �250 adults, with all individuals in asingle subpopulation ( IUCN criteria A.1 & C.1) ( Roemer1999; Timm et al. 2000; Roemer et al. 2001b, 2002). Theestimated probability of extinction in the wild for U. l.santacruzae is 50% within 10 years (IUCN criterion E)( Roemer et al. 2001b ). The San Clemente Island fox,U. l. clementae, is recommended to be listed as endangeredbecause it has a population estimated at �2500 matureindividuals, has been reduced by 50% over the past 10years, and is declining, and all individuals are in a singlesubpopulation that occupies �500 km2 ( IUCN criteriaA.4, B.2b,c & C.2b) (Roemer 1999; Roemer et al. 2001b;Schmidt et al. 2002). The San Nicolas Island subspecies,U. l. dickeyi, is recommended to be listed as vulnerablebecause the population is estimated to number �1000individuals ( IUCN criterion D) ( Roemer et al. 1994; Roe-mer 2001). Furthermore, U. l. dickeyi is one of the mostgenetically uniform wild populations ever described (Gil-bert et al. 1990; Wayne et al. 1991; Goldstein et al. 1999),and past demographic studies suggest that it wentthrough a population crash in the 1970s, recovering inthe early to mid-1980s ( Laughrin 1980; Kovach & Dow1981, 1985).
In sum, the biological determination made in the pro-posed rule is inadequate because it excludes both U. l.clementae and U. l. dickeyi from potential federal pro-tection. These two subspecies are small populations thatare either declining or have been through an historicpopulation decline and thus are vulnerable to future fluc-tuations in population size. Both subspecies may be sig-nificantly affected by the introduction of a lethal caninedisease, such as the canine distemper epizootic that deci-mated the Santa Catalina Island fox (Timm et al. 2000).Both subspecies have unique genetic variation that con-
tributes to the overall genetic diversity of the species. Fur-ther, Section 3(6) of the ESA states that “The term ‘endan-gered species’ means any species which is in danger ofextinction throughout all or a significant portion of itsrange . . . ” (U. S. Code. 1999. 16 U.S.C. Section 1533). Fiveof six island fox subspecies are now in danger of extinc-tion, representing a combined land area equaling 93.5% ofthe species’ range. Clearly, the entire species should re-ceive federal protection.
Single-Species Approach versus an Ecosystem Approach to Conservation
Rational arbitration of the conservation conflict betweenthe fox and the shrike should not only rest upon athoughtful evaluation of the evolutionary history and adap-tive diversity of both taxa ( Moritz 1994; Crandall et al.2000; Fraser & Bernatchez 2001) but should also considerthe ecological significance of both species (Carroll et al.1996). For example, the loss of fox populations on thenorthern Channel Islands has had immediate community-level impacts. The decline in foxes has resulted in anincrease in the island spotted skunk (Spilogale gracilisamphiala) on Santa Cruz Island, owing to competitiverelease (Roemer et al. 2002), and an increase in deermouse (Peromyscus maniculatus) densities on San MiguelIsland, owing to relaxed predation (Coonan et al. 2000).The shrike’s effect on the community is unknown.
The current conservation conflict might have beenavoided if the ecological importance of a variety of spe-cies had been considered and if an ecosystem approachto conservation had been adopted. A primary problemin shrike conservation is the absence of appropriate hab-itats. Habitat degradation that reduces foraging perchesand increases the density and cover of herbaceous vege-tation, thereby decreasing the availability of prey, has beenidentified as a key cause of declining shrike populationsworldwide (Temple 1995). Habitats on San Clemente Is-land were degraded through historic ranching activitiesand overgrazing by feral herbivores (Scott & Morrison1990), and, although the U.S. Navy made considerableprogress toward arresting habitat loss by eradicating feralherbivores, habitat restoration following the eradication ef-forts has been limited. Furthermore, captive-reared shrikeswere also released without the necessary refugia frompredators (appropriate nesting substrate and escape cover)that existed previously when foxes and shrikes actuallycoexisted. Habitat enhancement is needed in the form ofshrub establishment to increase the number of perchesand improve escape cover and nesting substrate forshrikes.
The proliferation of exotic annual grasses following fe-ral herbivore removal (Klinger et al. 1994; Laughrin etal. 1994) may also have exacerbated the problem by re-
Conservation BiologyVolume 17, No. 5, October 2003
Roemer & Wayne Conservation in Conflict 1257
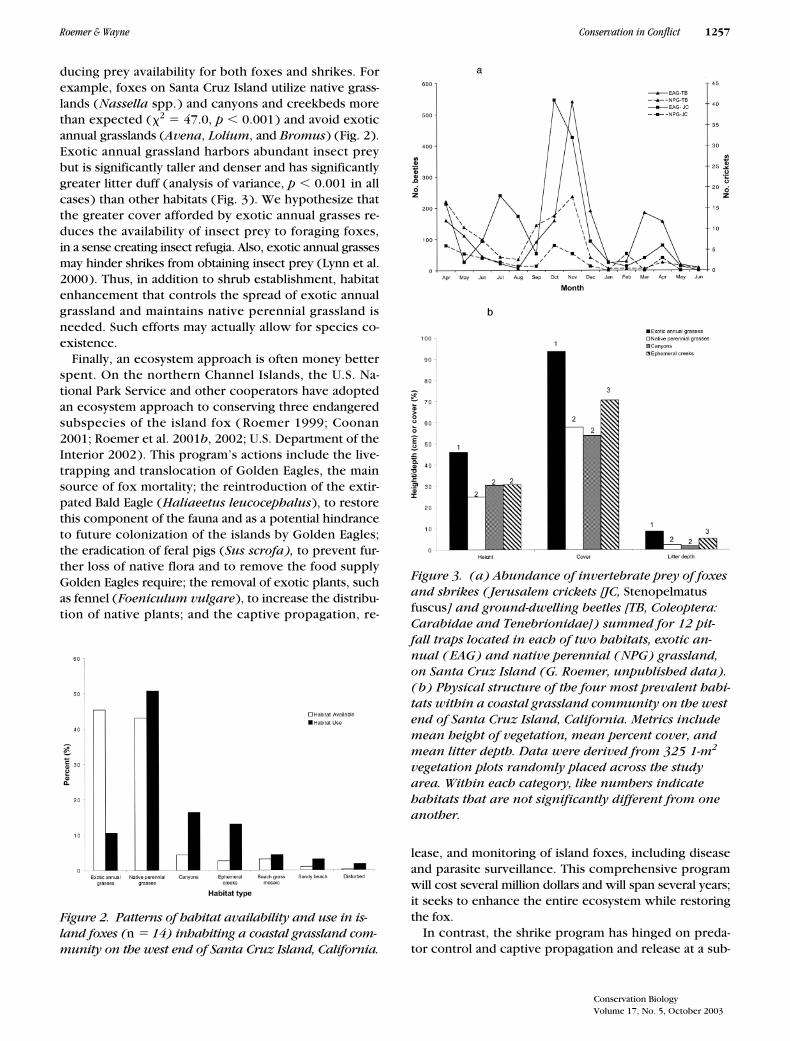
ducing prey availability for both foxes and shrikes. Forexample, foxes on Santa Cruz Island utilize native grass-lands (Nassella spp.) and canyons and creekbeds morethan expected (�2 � 47.0, p � 0.001) and avoid exoticannual grasslands (Avena, Lolium, and Bromus) (Fig. 2).Exotic annual grassland harbors abundant insect preybut is significantly taller and denser and has significantlygreater litter duff (analysis of variance, p � 0.001 in allcases) than other habitats (Fig. 3). We hypothesize thatthe greater cover afforded by exotic annual grasses re-duces the availability of insect prey to foraging foxes,in a sense creating insect refugia. Also, exotic annual grassesmay hinder shrikes from obtaining insect prey (Lynn et al.2000). Thus, in addition to shrub establishment, habitatenhancement that controls the spread of exotic annualgrassland and maintains native perennial grassland isneeded. Such efforts may actually allow for species co-existence.
Finally, an ecosystem approach is often money betterspent. On the northern Channel Islands, the U.S. Na-tional Park Service and other cooperators have adoptedan ecosystem approach to conserving three endangeredsubspecies of the island fox ( Roemer 1999; Coonan2001; Roemer et al. 2001b, 2002; U.S. Department of theInterior 2002). This program’s actions include the live-trapping and translocation of Golden Eagles, the mainsource of fox mortality; the reintroduction of the extir-pated Bald Eagle (Haliaeetus leucocephalus), to restorethis component of the fauna and as a potential hindranceto future colonization of the islands by Golden Eagles;the eradication of feral pigs (Sus scrofa), to prevent fur-ther loss of native flora and to remove the food supplyGolden Eagles require; the removal of exotic plants, suchas fennel (Foeniculum vulgare), to increase the distribu-tion of native plants; and the captive propagation, re-
lease, and monitoring of island foxes, including diseaseand parasite surveillance. This comprehensive programwill cost several million dollars and will span several years;it seeks to enhance the entire ecosystem while restoringthe fox.
In contrast, the shrike program has hinged on preda-tor control and captive propagation and release at a sub-
Figure 2. Patterns of habitat availability and use in is-land foxes (n � 14) inhabiting a coastal grassland com-munity on the west end of Santa Cruz Island, California.
Figure 3. (a) Abundance of invertebrate prey of foxes and shrikes ( Jerusalem crickets [JC, Stenopelmatus fuscus] and ground-dwelling beetles [TB, Coleoptera: Carabidae and Tenebrionidae]) summed for 12 pit-fall traps located in each of two habitats, exotic an-nual ( EAG) and native perennial ( NPG) grassland, on Santa Cruz Island (G. Roemer, unpublished data). ( b) Physical structure of the four most prevalent habi-tats within a coastal grassland community on the west end of Santa Cruz Island, California. Metrics include mean height of vegetation, mean percent cover, and mean litter depth. Data were derived from 325 1-m2 vegetation plots randomly placed across the study area. Within each category, like numbers indicate habitats that are not significantly different from one another.

1258 Conservation in Conflict Roemer & Wayne
Conservation BiologyVolume 17, No. 5, October 2003
stantial cost, $2.3 million spent in 1999 alone ( Morrisonet al. 1995; Schoch 1999; Cooper et al. 2001). Althoughpredator control is a necessary tool with which to con-trol or eradicate harmful non-native predators such as fe-ral cats (Felis catus), its negative effects on native taxacannot be ignored. Allocating funds toward habitat res-toration that would benefit the island ecosystem maybe one approach that could help alleviate the currentendangered-species conflict.
Conclusions and Recommendations
The U.S. ESA and its implementation have been criticizedon many grounds (Clark et al. 1994), some of which areexemplified by the current conflict between the shrikeand the fox. Biased implementation of the ESA has re-sulted in a species-centric approach to protecting the SanClemente Loggerhead Shrike, a listed subspecies of equiv-ocal evolutionary distinction and ecological importance.The subsequent recovery program has hinged upon ag-gressive control of native and non-native predators, whichis contributing to the decline of the San Clemente Islandfox, a distinct evolutionarily significant unit that plays animportant ecological role. The USFWS ignored the declinein the San Clemente Island fox and the importance ofthis subspecies and the San Nicolas Island fox to theoverall genetic diversity of the species, whereas the fourremaining island fox subspecies were being consideredfor federal protection. This lack of consideration may in-crease the risk of extinction for these subspecies and thespecies as a whole, especially if current practices to savethe San Clemente Loggerhead Shrike continue and theU.S. Navy is not subject to future Section 7 consultations.
Efforts to preserve the San Clemente Loggerhead Shrikeneed reconsideration. As has been suggested for an en-dangered subspecies of the California Gnatcatcher (Poliop-tilla californica californica; Cronin 1997), we recom-mend that both the San Clemente Loggerhead Shrike andthe other Channel Island form, L. l. anthonyi, be treated asmanagement units by instituting a cooperative agreementamong the various resource agencies and stakeholders.We recommend that all actions that negatively affect theSan Clemente Island fox be halted and that some of thefunds used for predator control be allocated toward hab-itat restoration. Finally, we recommend that all subspe-cies of the island fox receive federal protection and thatany action that may affect an island fox subspecies bebrought to the attention of all concerned parties, espe-cially to independent organizations that have no finan-cial or political interests on the islands ( Reading & Miller1994).
Given that the number of listed species or species thatare candidates for listing exceeds 1500 ( Hoekstra et al.2002), endangered-species conflicts will become com-monplace. Single-species management is a traditional so-
lution to conserving our natural heritage that might beimproved by a proactive ecosystem approach (Carroll etal. 1996). We have a better chance of conserving imper-iled species and the habitats on which they depend if en-dangered-species recovery programs weigh the value ofall populations to the ecosystem, consider the evolution-ary legacy and adaptive uniqueness of vulnerable taxa,and take into account the likelihood of a program’s suc-cess relative to its financial cost.
Acknowledgments
We thank the U.S. Navy for supplying some of the re-ports requested, and we thank the following individualsfor thoughtful comments on the manuscript: D. Cowley,C. J. Donlan, B. LaHaye, E. Main, C. Moritz, T. Smith, B. VanValkenburgh, M. Willson, and two anonymous reviewers.This work was supported by grants from the National Geo-graphic Society, the National Park Service, the Universityof California, Los Angeles, and the New Mexico Agricul-tural Experiment Station.
Literature Cited
Blackford, A. V., C. Couroux, H. Carlisle, J. Plissner, and N. Warnock.2001. Breeding report: 2000 population monitoring of the SanClemente Loggerhead Shrike on NALF, San Clemente Island, Cali-fornia. U.S. Navy, Natural Resources Management Branch, South-west Division, Naval Facilities Engineering Command, San Diego.
California Department of Fish and Game (CDFG). 1987. Five-year statusreport on the island fox (Urocyon littoralis). CDFG, Sacramento.
Carroll, R., C. Augspurger, A. Dobson, J. Franklin, G. Orians, W. Reid,R. Tracy, D. Wilcove, and J. Wilson. 1996. Strengthening the use ofscience in achieving the goals of the Endangered Species Act: an as-sessment by the Ecological Society of America. Ecological Applica-tions 6:1–11.
Clark, T. W., R. P. Reading, and A. L. Clarke. 1994. Endangered speciesrecovery, finding the lessons improving the process. Island Press,Washington, D.C.
Collins, P. W. 1991a. Interaction between island foxes (Urocyonlittoralis) and Indians on islands off the coast of Southern Califor-nia. I. Morphologic and archaeological evidence of human assisteddispersal. Journal of Ethnobiology 11(1):51–81.
Collins, P. W. 1991b. Interaction between island foxes (Urocyonlittoralis) and Native Americans on islands off the coast of South-ern California. II. Ethnographic, archaeological and historical evi-dence. Journal of Ethnobiology 11(2):205–229.
Collins, P. W. 1993. Taxonomic and biogeographic relationships of theisland fox (Urocyon littoralis) and gray fox (U. cinereoargenteus)from western North America. Pages 351–390 in F. G. Hochberg,editor. Third California islands symposium: recent advances in re-search on the California Islands. Santa Barbara Museum of NaturalHistory, Santa Barbara, California.
Collister, D. M., and D. Wicklum. 1996. Intraspecific variation in Log-gerhead Shrikes: sexual dimorphism and implication for subspeciesclassification. The Auk 113:221–223.
Coonan, T. J. 2001. Recovery plan for island foxes (Urocyon littoralis)on the northern Channel Islands. Park headquarters, Channel Is-lands National Park, Ventura, California.
Coonan, T. J. 2002. Findings of the island fox conservation working

Conservation BiologyVolume 17, No. 5, October 2003
Roemer & Wayne Conservation in Conflict 1259
group, Ventura, California. Park headquarters, Channel Islands Na-tional Park, Ventura, California.
Coonan, T. J., C. A. Schwemm, G. W. Roemer, and G. Austin. 2000.Population decline of island foxes (Urocyon littoralis) on SanMiguel Island. Pages 289–297 in D. R. Browne, K. L. Mitchell, andH. W. Chaney, editors. Proceedings of the fifth channel islandssymposium. U.S. Department of the Interior, Minerals ManagementService, Pacific Outer Continental Shelf Region, Camarillo, California.
Cooper, D. M., E. L. Kershmer, G. A. Schmidt, and D. K. Garcelon.2001. San Clemente Loggerhead Shrike predator research and man-agement program—2000. Institute for Wildlife Studies, Arcata, Cali-fornia. Final report. U. S. Navy, Natural Resources ManagementBranch, Southwest Division, Naval Facilities Engineering Command,San Diego, California.
Crandall, K. A., O. R. P. Bininda-Emonds, G. Mace, and R. K. Wayne.2000. Considering evolutionary processes in conservation biology.Trends in Ecology & Evolution 15:290–295.
Cronin, M. A. 1997. Systematics, taxonomy, and the Endangered Spe-cies Act: the example of the California gnatcatcher. Wildlife Soci-ety Bulletin 25:661–666.
Crooks, K. R., and D. Van Vuren. 1995. Resource utilization by two in-sular endemic mammalian carnivores, the island fox and islandspotted skunk. Oecologia 104:301–307.
Elliot, T., and B. Popper. 1999. Predator management report for theprotection of the San Clemente Loggerhead Shrike. U. S. Navy, Nat-ural Resources Management Branch, Southwest Division Naval Fa-cilities Engineering Command, San Diego.
Fraser, D. J., and L. Bernatchez. 2001. Adaptive evolutionary conserva-tion: towards a unified concept for defining conservation units.Molecular Ecology 10:2741–2752.
Gilbert, D. A., N. Lehman, S. J. O’Brien, and R. K. Wayne. 1990. Ge-netic fingerprinting reflects population differentiation in the Cali-fornia channel island fox. Nature 344:764–767.
Goldstein, D. B., G. W. Roemer, D. A. Smith, D. E. Reich, A. Bergman, andR. K. Wayne. 1999. The use of microsatellite variation to infer patternsof migration, population structure and demographic history: an evalu-ation of methods in a natural model system. Genetics 151:797–801.
Hall, E. R. 1981. The mammals of North America. 2nd edition. John Wiley& Sons, New York.
Hoekstra, J. M., J. A. Clark, W. F. Fagan, and P. D. Boersma. 2002. Acomprehensive view of Endangered Species Act recovery plans.Ecological Applications 12:630–640.
Juola, F. A., W. T. Everett, and C. E. Koehler. 1997. Final report: 1996population and habitat survey of the Loggerhead Shrike on NALFSan Clemente Island, California. U.S. Navy, Southwest Division, Na-val Facilities Engineering Command, San Diego.
Klinger, R. C., P. T. Schuyler, and J. D. Sterner. 1994. Vegetation re-sponse to the removal of feral sheep from Santa Cruz Island. Pages341–350 in W. L. Halvorsen and G. J. Maender, editors. The fourthCalifornia islands symposium: update on the status of resources.Santa Barbara Museum of Natural History, Santa Barbara, California.
Kovach, S. D., and R. J. Dow. 1981. Status and ecology of the islandfox on San Nicolas Island, 1980. Technical memorandum TM–81–28. Pacific Missile Defense Center, Point Mugu, California.
Kovach, S. D., and R. J. Dow. 1985. Island fox research on San NicolasIsland. Annual report. Department of the Navy, Pacific Missile TestCenter, San Diego.
Laughrin, L. 1977. The island fox: a field study of its behaviour andecology. Ph.D. dissertation. University of California, Santa Barbara.
Laughrin, L. 1980. Populations and status of the island fox. Pages 745–749 in D. M. Power, editor. The California islands: proceedings of amultidisciplinary symposium. Santa Barbara Museum of NaturalHistory, Santa Barbara, California.
Laughrin, L., M. Carroll, A. Bromfield, and J. Carroll. 1994. Trends invegetation changes with removal of feral animal grazing pressureson Santa Catalina Island. Pages 523–530 in W. L. Halvorsen andG. J. Maender, editors. The fourth California islands symposium:
update on the status of resources. Santa Barbara Museum of NaturalHistory, Santa Barbara, California.
Lynn, S., J. A. Martin, D. M. Cooper, K. M. Wakelee, G. A. Schmidt, andD. K. Garcelon. 2000. Research efforts to aid in the recovery of theSan Clemente Loggerhead Shrike—1999. U. S. Navy, Natural Re-sources Management Branch, Southwest Division, Naval FacilitiesEngineering Command, San Diego.
Mader, T., N. Warnock, and W. Ostheimer. 2000. Final report: 1999 popu-lation monitoring of the Loggerhead Shrike on NALF, San Clemente Is-land, California. U. S. Navy, Natural Resources Mangement Branch,Southwest Division Naval, Facilities Engineering Command, San Diego.
Miller, A. H. 1931. Systematic revision and natural history of the Amer-ican shrikes (Lanius). University of California Publications in Zool-ogy 38:11–242.
Moore, C. M., and P. W. Collins. 1995. Urocyon littoralis. No. 489. Mam-malian species. The American Society of Mammalogists, BrighamYoung University, Provo, Utah.
Moritz, C. 1994. Defining ‘evolutionarily significant units’ for conserva-tion. Trends in Ecology & Evolution 9:373–375.
Morrison, M. L., C. M. Kuehler, T. A. Scott, A. A. Lieberman, W. T.Everett, R. B. Phillips, C. E. Koehler, P. A. Aigner, C. Winchell, andT. Burr. 1995. San Clemente Loggerhead Shrike: recovery plan foran endangered species. Proceedings of the Western Foundation ofVertebrate Zoology 6:293–295.
Mundy, N. I., C. S. Winchell, and D. S. Woodruff. 1997a. Genetic dif-ferences between the endangered San Clemente Loggerhead ShrikeLanius ludovicianus mearnsi and two neighboring subspeciesdemonstrated by mtDNA control region and cytochrome b se-quence variation. Molecular Ecology 6:29–37.
Mundy, N. I., C. S. Winchell, T. Burr, and D. S. Woodruff. 1997b.Microsatellite variation and microevolution in the critically endan-gered San Clemente Island Loggerhead Shrike (Lanius ludovi-cianus mearnsi). Proceedings of the Royal Society London Series B264:869–875.
National Research Council. 1995. Science and the Endangered SpeciesAct. National Academy Press, Washington, D.C.
Nei, M. 1978. Estimation of average heterozygosity and genetic dis-tance from a small number of individuals. Genetics 89:583–590.
Patten, M. L., and K. F. Campbell. 2000. Typological thinking and theconservation of subspecies: the case of the San Clemente IslandLoggerhead Shrike. Diversity and Distributions 6:177–188.
Reading, R. P., and B. J. Miller. 1994. The black-footed ferret recoveryprogram: unmasking professional and organizational weaknesses.Pages 73–100 in T. W. Clark, R. P. Reading, and A. L. Clarke, edi-tors. Endangered species recovery: finding the lessons, improvingthe process. Island Press, Washington, D.C.
Ridgway, R. 1903. Descriptions of new genera of species and subspe-cies of American birds. Proceedings of the Biological Society ofWashington 16:105–112.
Roemer, G. W. 1999. The ecology and conservation of the island fox.Ph.D. dissertation. University of California, Los Angeles.
Roemer, G. W. 2001. Annual progress report 2000: summary of the de-mography of the San Nicolas Island fox (Urocyon littoralis dickeyi).Final report. U. S. Navy, Point Mugu Naval Air Station, Camarillo, Cal-ifornia.
Roemer, G. W., D. K. Garcelon, T. J. Coonan, and C. Schwemm. 1994.The use of capture-recapture methods for estimating, monitoring,and conserving island fox populations. Pages 387–400 in W. L. Hal-vorsen, and G. J. Maender, editors. The fourth California islandssymposium: update on the status of resources. Santa Barbara Mu-seum of Natural History, Santa Barbara, California.
Roemer, G. W., P. S. Miller, J. Laake, C. Wilcox, and T. J. Coonan.2000. Island fox demographic workshop report. Final report.Channel Islands National Park, Ventura, California.
Roemer, G. W., D. A. Smith, D. K. Garcelon, and R. K. Wayne. 2001a.The behavioural ecology of the island fox. Journal of Zoology 255:1–14.

1260 Conservation in Conflict Roemer & Wayne
Conservation BiologyVolume 17, No. 5, October 2003
Roemer, G. W., T. J. Coonan, D. K. Garcelon, J. Bascompte, and L.Laughrin. 2001b. Feral pigs facilitate hyperpredation by Golden Ea-gles and indirectly cause the decline of the island fox. Animal Con-servation 4:307–318.
Roemer, G. W., C. J. Donlan, and F. Courchamp. 2002. Golden eagles,feral pigs and insular carnivores: how exotic species turn nativepredators into prey. Proceedings of the National Academy of Sci-ences of the United States of America 99:791–796.
Roemer, G. W., T. J. Coonan, L. Munson, and R. K. Wayne. 2003. Canidaction plan for the island fox. In press in C. Sillero-Zubiri, J. R.Ginsberg, and D. W. Macdonald, editors. Foxes, wolves, jackals anddogs: status survey and conservation action plan. 2nd edition.World Conservation Union, Gland, Switzerland.
Schmidt, G. A., D. K. Garcelon, and J. Sloan. 2002. Fox monitoring andresearch in support of the San Clemente Loggerhead Shrike preda-tor control program on Naval Auxiliary Landing Field, San Clem-ente Island, California. Report of the Institute for Wildlife Studies,Arcata, California. U.S. Navy, Natural Resources MangementBranch, Southwest Division, Naval Facilities Engineering Com-mand, San Diego.
Schoch, D. 1999. Hard lessons in the tale of the fox and the shrike spe-cies on San Clemente Island, the Navy has been killing a rare preda-tor in a bid to save its nearly extinct prey. Los Angeles Times 17May:1.
Scott, T. A., and M. L. Morrison. 1990. Natural history and manage-ment of the San Clemente Loggerhead Shrike. Proceedings of theWestern Foundation of Vertebrate Zoology 4:23–60.
Temple, S. A. 1995. Priorities for shrike research and conservation.Proceedings of the Western Foundation of Vertebrate Zoology 6:296–298.
Timm, S. F., J. M. Stokely, T. B. Gehr, R. L. Peebles, and D. K. Garcelon.
2000. Investigation into the decline of island foxes on Santa Cat-alina Island. Institute for Wildlife Studies, Arcata, California.
U.S. Department of Agriculture Animal and Plant Inspection Service–Wild-life Services. 1998. Final environmental assessment: predator damagemanagement to protect the federally endangered San Clemente Log-gerhead Shrike on San Clemente Island. U.S. Navy, Southwest Divi-sion, Naval Facilities Engineering Command, San Diego, California.
U.S. Department of the Interior. 1977. Determination that seven Chan-nel Island animals and plants are either endangered species orthreatened species. Federal Register 42(155):40682–40685.
U.S. Department of the Interior. 2001. Endangered and threatenedwildlife and plants: listing the San Miguel Island fox, Santa Rosa Is-land fox, Santa Cruz Island fox and Santa Catalina Island fox as en-dangered (50 CFR Part 17). Federal Register 66(237):63654–63665.
U.S. Department of the Interior. 2002. Santa Cruz Island primary resto-ration plan. Final environmental impact statement. Park Headquar-ters, Channel Islands National Park, Ventura, California.
U.S. Department of the Interior and Department of Commerce. 1996.Proposed policy on the treatment of intercrosses and intercrossprogeny (the issue of “hybridization”); Proposed rule (50 CFR Part424). Federal Register 61(26):4709–4713.
Wayne, R. K., S. B. George, D. Gilbert, P. W. Collins, S. D. Kovach, D.Girman, and N. Lehman. 1991. A morphologic and genetic study ofthe island fox, Urocyon littoralis. Evolution 45:1849–1868.
Wilson, D. E., and D. M. Reeder. 1993. Mammal species of the world: ataxonomic and geographic reference. 2nd edition. Smithsonian In-stitution Press, Washington, D.C.
World Conservation Union ( IUCN). 2001. IUCN red list categories andcriteria. Version 3.1. Species Survival Commission, IUCN, Gland, Swit-zerland.





























