Embed Size (px)
Citation preview

Proc. Natl. Acad. Sci. USAVol. 92, pp. 11955-11959, December 1995Biochemistry
Crystal structure of the membrane-exposed domain from arespiratory quinol oxidase complex with an engineereddinuclear copper center
(cytochrome oxidase/cupredoxin fold)
MATTHIAS WILMANNS, PEKKA LAPPALAINEN, MARK KELLY, ELISABETH SAUER_ERIKSSON*, AND MATTI SARASTEEuropean Molecular Biology Laboratory, Meyerhofstrasse 1, Postfach 10.2209, D-69012 Heidelberg, Germany
Communicated by Helmut Beinert, University of Wisconsin, Madison, WI, September 18, 1995 (received for review August 31, 1995)
ABSTRACT Cytochrome oxidase is a membrane proteincomplex that catalyzes reduction of molecular oxygen to waterand utilizes the free energy of this reaction to generate atransmembrane proton gradient during respiration. The elec-tron entry site in subunit II is a mixed-valence dinuclearcopper center in enzymes that oxidize cytochrome c. Thiscenter has been lost during the evolution of the quinol-oxidizing branch of cytochrome oxidases but can be restoredby engineering. Herein we describe the crystal structures ofthe periplasmic fragment from the wild-type subunit II(CyoA) of Escherichia coli quinol oxidase at 2.5-A resolutionand of the mutant with the engineered dinuclear copper center(purple CyoA) at 2.3-A resolution. CyoA is folded as an11-stranded mostly antiparallel 13-sandwich followed by threea-helices. The dinuclear copper center is located at the loopsbetween strands 185-.16 and 189-1310. The two coppers are ata 2.5-A distance and symmetrically coordinated to the mainligands that are two bridging cysteines and two terminalhistidines. The residues that are distinct in cytochrome c andquinol oxidases are around the dinuclear copper center. Struc-tural comparison suggests a common ancestry for subunit II ofcytochrome oxidase and blue copper-binding proteins.
Two main branches of the cytochrome oxidase family inbacteria alternatively use cytochrome c or quinol as theelectron donor (1, 2). They typically contain three subunitsthat are homologous to the key components of the mitochon-drial enzyme. Subunit I is largely buried in the membrane andforms the core of the complex, harboring the heme-copperactive site. Subunit II has been predicted to be composed of anN-terminal transmembrane helical hairpin and a hydrophilicC-terminal domain that resides in the outer side of thecytoplasmic membrane (3). In cytochrome c oxidases, this do-main contains a copper center known as CUA that functions as theprimary acceptor for the electron from cytochrome c (4, 5).Subunit III is a membrane protein without redox centers (3).The CUA site is a single electron acceptor and donor. It has
been spectroscopically characterized as a dinuclear center witha mixed valence [Cu(1.5)-Cu(1.5)]S = 1/2 configuration (4,6, 7), where the coppers must be in very similar chemicalenvironments (8). The key amino acids involved in the bindingsite are found in the CCHM amino acid motif, which ispreceded by an invariant His in all subunit II sequences ofcytochrome c oxidases (Fig. 1). None of these residues areconserved in quinol oxidases lacking the metal center (9, 12,13). The "lost" CUA site has been reengineered to subunit IIof the Escherichia coli quinol oxidase (CyoA) by a simulta-neous substitution of six residues that results in a protein withpurple color (9) (hereafter referred to as purple CyoA). Wehave determined the crystal structure of CyoA and purple
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement" inaccordance with 18 U.S.C. §1734 solely to indicate this fact.
CyoA at high resolutiont and present a description of thestructure of the CUA site in atomic detail.
METHODS
Purification and Crystallization. The CyoA fragment waspurified and crystallized as described (9, 10, 13). Single crystalswith maximum dimensions of 0.5 x 0.5 x 0.15 mm3 wereobtained by the macroseeding method at 4°C. Fresh dropswere seeded after 3-5 h with previously washed crystal frag-ments. Because no useful heavy atom derivatives could beobtained with the wild-type CyoA crystals, residues Asn-158,Ser-167, Ser-174, Asn-198, Thr-202, Ser-207, Ser-209, Ser-211,Lys-219, Ser-241, Ser-246, Ser-258, or Ser-267 were replaced byCys residues by site-directed mutagenesis (Fig. 1). All mutantswere overexpressed and purified as the original fragment andshowed the same properties as the wild-type protein exceptK219C (reduced solubility) and S21 1C (dimerization underoxidized conditions). Crystals could be obtained from theN158C, S174C, S207C, S241C, S246C, and S267C mutants.Macroseeded crystals of N158C, S207C, S241C, and S267Chad sufficient size for x-ray analysis. The best crystals withmaximum dimensions of 0.8 x 0.8 x 0.2 mm3 were obtainedwith the S207C mutant at room temperature. The crystals ofall analyzed mutants were isomorphous to those of wild-typeCyoA (a = 47.6 + 0.2 A; b = 84.1 ± 0.5 A; c = 94.7 ± 0.7 A;space group, I222; one molecule per asymmetric unit) exceptfor about half of the crystals of the S207C mutant (a = 93.8 ±0.5 A; b = 84.1 + 0.5 A; c = 48.2 ± 0.2 A; space group, P21212;two molecules per asymmetric unit).The DNA coding for residues 116-296 of the original purple
CyoA was amplified by PCR and ligated to a modified pET3dvector (9). The protein was expressed and purified as described(10). Purple CyoA crystals grew as squared plates within 1month at 4°C to maximum dimensions of 0.5 x 0.5 x 0.2 mm3.Initial hanging drops were prepared by mixing 2 ,ul of proteinsolution (15-20 mg/ml) in 20 mM Tris HCl (pH 8.2) with 2 j,lof reservoir solution [20% (wt/vol) PEG (Mr = 6000)/180 mMacetic acid/20 mM sodium acetate, pH 4.8/50mM ammoniumsulfate/i mM NaN3]. The crystals have a deep purple coloralong the plane of the plate and appear colorless when viewedvertically to this plane. They grow in the space group P21212with cell dimensions a = 46.0 A, b = 98.2 A, and c = 39.9 A(based on the data set measured at 100 K) with one moleculeper asymmetric unit.Data Collection and Processing. Two wild-type (cyoA-I and
cyoA-II) and four heavy-atom derivative (S207C/EtHgP,
Abbreviations: CyoA, subunit II of quinol oxidase from Escherichiacoli; purple CyoA, CyoA with engineered CUA binding site.*Present address: Department of Molecular Biology, Uppsala Uni-versity, P.O. Box 590, S-75124 Uppsala, Sweden.tThe atomic coordinates and structure factors have been deposited inthe Protein Data Bank, Chemistry Department, Brookhaven Na-tional Laboratory, Upton, NY 11973 (codes iCYW and 1CYX).
11955
Dow
nloa
ded
by g
uest
on
Janu
ary
25, 2
020

11956 Biochemistry: Wilmanns et al.
120 130 140 150 160t I
cyoA THALEPSKPLAHDEKPITIEVV W FFIYPEQG. ...................... IATVNEIAFPANTPVYFcox2 LFRSQEM . ... PNDPDLVIKAIGHUYSYEYPNDGVAFDALMLEKEALADAGYSEDEYLLATDNPVVVPVGKKVLV
170 180 190 200 210 220 230 240I1* I * * * It
cyoA KVTSNSVNNSFFIPRLGSQIYAHAGMQTRLHLIANEPGTYDGISASYSGPGFSGMKFKAIATPDRAAFDQWVAKAKQcox2 QVTAl§I>WTIPAFAVK5tPGRIAQLWFSVDQEVGVYFGIsYMPIVVKAVSQE. KYEAWLAGAKE
c
250 260 270 280 290 300 310
CYOA SPNTMSDMAAFEKLAAPSEYNQVEYFSNVKPDLFADVINKFEAHGKSMDMTQPEGEHSAHEGMEGMDMSHAESAHcox2 EFAA
FIG. 1. Alignment of two representative sequences of the membrane-exposed part of CyoA and a cytochrome c oxidase (Paracoccus denitrificanscox2). The CyoA sequence covers the region used for crystallization of the wild-type CyoA (9, 10). The C terminus of Cox2 corresponds to themature protein after post-translational processing (11). The color coding of individual residues is based on a multiple alignment of CyoA andcytochrome c oxidases (3, 12). The numbering refers to the full-length CyoA sequence. Highlighted residues are invariant (orange), conserved(yellow), or specific for cytochrome c oxidases (purple) or quinol oxidases (blue). The residues that were substituted for engineering of the CUAsite in purple CyoA are indicated by asterisks (see ref. 9). The secondary structure elements of CyoA are schematically shown on top of the sequencealignment. The N and C termini are the limits of the polypeptide chain defined in the electron density. Arrows show }3-strands that can besuperimposed with the cupredoxin fold in green and remaining ,B-strands in yellow; red cylinders are a-helices. Loops that are involved in copper
binding are shown with purple lines, and remaining loops are in blue. The secondary structure elements are labeled.
S207C/HgCl2, S207C/PbEt3Ac, and S241C/EMTS) data setsof Cys mutants were used for the crystallographic structuredetermination of CyoA (where EtHgP is ethyl mercury phos-phate, PbEt3Ac is triethyl lead acetate, and EMTS is ethylmercury thiosalicylate). CyoA soaking conditions were withmercury compounds at a concentration of 1 mM for 1-2 daysand with PbEt3Ac at a concentration of 20 mM for 7 days. Thedata sets were recorded under the following conditions: Datasets cyoA-I, S207C/EtHgP, S207C/PbEt3Ac, S241C/EMTSwere from small Mar imaging plate (radius = 90 mm) mountedon a Siemens/MacScience model MX18 generator; data setcyoA-II was from a FAST area detector mounted on an Elliotmodel GX21 generator; data set S207C/HgCl2 was from a
small Mar imaging plate mounted on the XlI beam line at theEuropean Molecular Biology Laboratory/Deutsches Elek-tronen Synchrotron synchrotron facility (Hamburg); data setpurple-cyoA was from a big Mar imaging plate (radius = 150mM) mounted on an Enraf-Nonius model FR571 generator.Cu-Ka radiation was used for all data sets except for the dataobtained from the Xll beam line (A = 0.93 A). All data setsexcept purple-cyoA were recorded at room temperature.Purple CyoA crystals were washed with reservoir solution. Thewashing solution with the crystal was then mixed with 40%
PEG (Mr = 6000) in a 1:1 (vol/vol) ratio and immediatelytransferred into a small fiber loop and shock-frozen at 100 K,in an Oxford CryoSystems cryostream instrument. All datasets were reduced and processed with the XDS program
package (14). Overall statistics are given in Tables 1 and 2.Structure Determination and Refinement. The coordinates
of the major heavy atom sites of all used derivative data setswere determined from difference Patterson maps. Anomalousdifference Patterson maps of S207C/EtHgP and S207C/HgCI2unambiguously confirmed the heavy atom positions. Theanomalous differences of both derivative data sets were furtherused in phase determination. Heavy atom positions and oc-
cupancies were refined with the program MLPHARE (15). Theabsolute configuration was determined from difference Fou-rier maps in several combinations from SIRAS phases ofS207C/EtHgP. The phases were improved and extended to2.7-A resolution with the program SQUASH (16) by employingsolvent flattening and histogram matching. The resultingelectron density had sufficient quality for building a partialmodel using the program o (17). Partial model phases were
combined with MLPHARE phases and again improved with theprogram SQUASH. Several rounds of model building and phasecombination were carried out to complete =80% of the model
Table 1. X-ray structure determination: Data and MIRAS phasing statistics
Purple-cyoA-I cyoA-II S207C S207C S207C S241C cyoA
Heavy atom derivative EtHgP HgCl2 PbMe3Ac EMTSMeasured reflections, no. 20,398 12,996 14,154 18,002 4608 18,002 17,363Unique reflections, no. 5029 5943 4007 6140 3543 4363 7426Resolution, A 2.7 2.5 2.9 2.5 3.0 2.9 2.3Completeness, % 91.4 86.3 89.8 89.2 76.3 97.5 87.3Rmerge, % 6.1 7.0 7.8 7.5 15.6 14.5 8.4Ri,o, % 22.4 26.5 30.2 16.2Phasing power 2.30 1.22 1.26 0.87Rcullis, % 0.60 (0.92) 0.79 (0.96) 0.80 0.88
EtHgP, ethyl mercury phosphate; PbMe3Ac, trimethyl lead acetate; EMTS, ethyl mercury thiosalicylate. Rmerge = XhklYjlIhklj- (Ihkl)I/yhkl,j (Ihkl), using Friedel mates as independent observations. Ri,O = 7_k,IFjso - Fnatj/YhkIFnat. Phasing power is definedas (FH/lack of closure). Rcullis = (lack of closure)/(isomorphous difference) for acentric reflections, and RCullis(anom.) = (lackof closure)/(anomalous difference) (shown in parentheses).
Proc. Natl. Acad. Sci. USA 92 (1995)
N-
Dow
nloa
ded
by g
uest
on
Janu
ary
25, 2
020

Proc. Natl. Acad. Sci. USA 92 (1995) 11957
Table 2. Refinement statistics
cyoA Purple-cyoA
Residue range 125-283 125-282No. of nonhydrogen atoms 1285 1405No. of included water molecules 46 168Average temperature factors, A2Main chain atoms 28.6 18.5Side chain atoms 29.2 19.6Water molecules 32.7 31.6
R factor (Rfree) 19.2 (32.6) 19.6 (32.8)Rms deviationFrom ideal bond length, A 0.013 0.010From ideal bond angles, degrees 1.80 1.58From ideal planar groups, degrees 1.68 1.39
Residue position numbers are defined in Fig. 1. The temperaturefactors of the two copper ions are 15.6 and 17.5 2. R factor = 72hkhIFobs- FcaIcI/7hkIFobs for 90% of all observed data with intensities alobetween 6.0-A resolution and high-resolution limit (see text). The freeR factor was calculated from the remaining 10% of the observed datathat were not used during the refinement.
of the refined CyoA structure. The model was further im-proved by 10 cycles of X-PLOR molecular dynamics refinement(18) and interactive model building (17). The resulting modelhad an R factor of -22%.
This model was used as a template to determine the crystalstructure of purple CyoA by molecular replacement with theprogram AMORE (19). All amplitudes between 8.0- and 3.0-Aresolution of purple-cyoA data set were used. The integrationsphere radius was 17 A. The correct translation solution hadan R factor of 49.4% and correlation coefficient of 0.351 asdefined in the program AMORE. This solution was refined bythe FITTING function of AMORE to an R factor of 48.0% andcorrelation coefficient of 0.439. The model of purple CyoA wasrefined to 2.3-A resolution by a cyclic procedure of interactivemodel building with the program o (17), molecular dynamicsrefinement was with the program X-PLOR (18), and electrondensity improvement was with the program ARP (20). The CUAloop was completely rebuilt due to the strong conformationalchange, and its structure was verified by simulated annealingOMIT maps (18).Attempts were carried out to further extend the model toward
the N and C termini. Extensions were only accepted if both theR factor and free R factor decreased. The two highest differencepeaks were located at the expected site of the dinuclear center andwere interpreted as copper ions. Some of the included water
molecules might be located in areas of uninterpretable proteinelectron density. The refined purple CyoA model was in turn usedto guide further refinement of the CyoA structure against thecyoA-II data set at a 2.5-A resolution. The relatively high free Rfactors are probably due to uninterpretable electron density ofboth termini that can considerably contribute to scattering matterwithin the unit cell.
All other crystallographic computations were carried out byprograms of the CCP4 suite (21). Further statistics are sum-marized in Tables 1 and 2.
RESULTS AND DISCUSSIONOverall Fold. The three-dimensional structure of CyoA
consists of a large, mostly antiparallel ,B-sandwich with anoverall Greek key (3-barrel topology (Fig. 2). The hydrogenbonding pattern is, however, interrupted between strands/37-f38 and (34-f311. The C-terminal part contains three a-he-lices that mainly interact with the sheet f8-,35-f1-f2-133-(311. The turn in loop ,B9-,B10 (CUA loop) is followed by a shorthelix containing residues 215-218 (data not shown). The 14N-terminal and 33 C-terminal residues in both CyoA structuresare not defined in the electron density (Fig. 1). Based on themain-chain atoms, the three-dimensional structure of purpleCyoA superimposes with a 0.56-A- rms deviation on the CyoAstructure. The two structures are virtually identical with theexception of the CUA loop (Fig. 3). Upon metal binding, theengineered loop undergoes a strong conformational rear-rangement that is pivoted at residue Cys-211.
Dinuclear Copper Center. The CUA site in purple CyoA issymmetrical with respect to the two His and two Cys ligands(Fig. 3 D-F). The distance between the two coppers is 2.5 A.His-172 and His-215 function as terminal ligands of the twometals. The imidazole rings of the two His residues share acommon plane that is tilted by 150 with respect to the Cu-Cuaxis. The two thiolate groups of Cys-207 and Cys-211 act asbridging ligands between the coppers. All four Sw-Cu coordi-nation bonds are -2.2A long. Asymmetry is introduced by twofurther bonds between the 58 atom of Met-218 and Cu-1 andthe main-chain carbonyl group of Glu-209 and Cu-2. The twothiolate groups and the coordinating atoms of Glu-209 andMet-218 are in the trans position with respect to the Cu-Cuaxis. Each copper ion can be seen as coordinated by fourligands in a distorted tetrahedral configuration. It is probablethat the paramagnetic electron is delocalized via the thiolate
FIG. 2. Three-dimensional struc-ture of CyoA. (A) Topology diagramof the CyoA fold is shown with colorcodes and numbering of the secondarystructure elements. The residue num-bers of structural elements are alsogiven. The numbers of 13-sheet hydro-gen bonds are shown between pairs of,3-strands. Note that there are no reg-ular 1-sheet hydrogen bonds between,37-,B8 and 1311-134. The dinuclear cop-per center is indicated by a purpledumbbell. (B) Ribbon drawing (22) ofthe structure of purple CyoA with thecolor codes and numbers of secondarystructure elements are as in A.
Biochemistry: Wilmanns et al.
b
Dow
nloa
ded
by g
uest
on
Janu
ary
25, 2
020
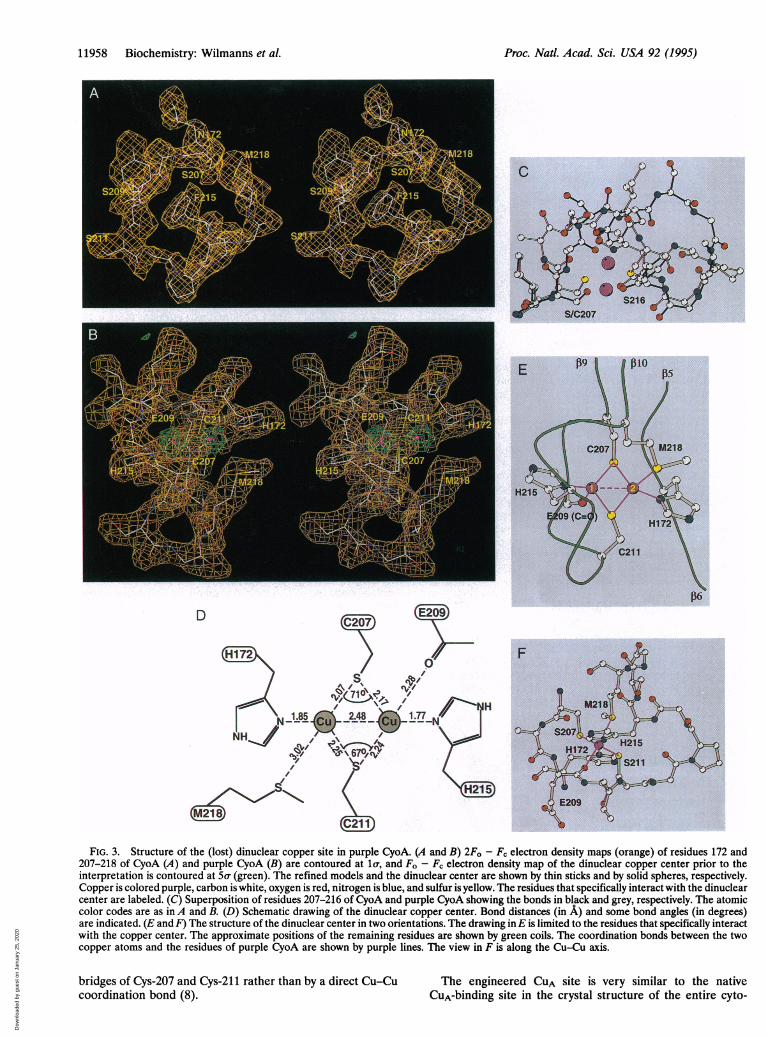
11958 Biochemistry: Wilmanns et al.
FIG. 3. Structure of the (lost) dinuclear copper site in purple CyoA. (A and B) 2Fo - FC electron density maps (orange) of residues 172 and207-218 of CyoA (A) and purple CyoA (B) are contoured at la(, and Fo - Fc electron density map of the dinuclear copper center prior to theinterpretation is contoured at 5o- (green). The refined models and the dinuclear center are shown by thin sticks and by solid spheres, respectively.Copper is colored purple, carbon is white, oxygen is red, nitrogen is blue, and sulfur is yellow. The residues that specifically interact with the dinuclearcenter are labeled. (C) Superposition of residues 207-216 of CyoA and purple CyoA showing the bonds in black and grey, respectively. The atomiccolor codes are as in A and B. (D) Schematic drawing of the dinuclear copper center. Bond distances (in A) and some bond angles (in degrees)are indicated. (E and F) The structure of the dinuclear center in two orientations. The drawing in E is limited to the residues that specifically interactwith the copper center. The approximate positions of the remaining residues are shown by green coils. The coordination bonds between the twocopper atoms and the residues of purple CyoA are shown by purple lines. The view in F is along the Cu-Cu axis.
bridges of Cys-207 and Cys-211 rather than by a direct Cu-Cu The engineered CUA site is very similar to the nativecoordination bond (8). CuA-binding site in the crystal structure of the entire cyto-
Proc. Natl. Acad. Sci. USA 92 (1995)
Dow
nloa
ded
by g
uest
on
Janu
ary
25, 2
020

Proc. Natl. Acad. Sci. USA 92 (1995) 11959
chrome c oxidase complex of Paracoccus denitrificans (23),supporting the idea that the quinol oxidases have lost this siteduring evolution (9). The x-ray structure confirms proposalsfor the ligation and electronic state of the two coppers basedon mutagenesis and different spectroscopic studies (4, 13). Inparticular, the structure satisfies theoretical considerationsthat imply symmetry within the dinuclear site (4, 8). Until now,it has not been possible to make a choice between twoalternative models, known as the "terminal" model with aCu-Cu metal bond and two Cys residues as terminal ligands(24) and the "bridging" model with two Cys residues asbridging ligands (8, 25). The bridging model has been con-firmed by our x-ray structure. However, the coppers are closeto each other at the distance accurately measured by extendedx-ray absorption fine structure (24, 25), confirming the datathat have led to the terminal model.Amino acids that are specific for cytochrome c or for quinol
oxidases or are conserved in both are highlighted in Figs. 1 and4. A cluster of aromatic residues below the CuA-binding siteforms a conserved aromatic channel along the axis of the(3-barrel. The highly conserved residue Trp-136 is located atthe port to the CuA-binding site and could be crucial todirecting electrons into the site. Its substitution affects thecatalytic activity of the E. coli quinol oxidase (R. Gennis,personal communication). All residues that are specific forcytochrome c oxidases and absent from the quinol oxidases arelocated either within the CuA-binding site or in its closevicinity. These include the carboxylic acid residues that havebeen suggested to form the cytochrome c binding site (26).Common Fold with Blue Copper Proteins. It has been
postulated (3, 9, 27) that blue copper proteins and subunit IIof cytochrome oxidase share the cupredoxin fold (28). Thethree-dimensional structure of CyoA confirms this hypothesis.Two additional (3-strands are, however, inserted between the
FIG. 4. Schematic drawing of the smoothed backbone of CyoAhighlighting conserved residues. The orientation is as in Fig. 2B. Thecolor codes of the conserved positions are as in Fig. 1.
canonical strands (31 and (32 of the cupredoxin fold (28); theseare (32 and ,B3 in CyoA (Fig. 2). The second (3-sheet is extendedby the strand (311. The presence of this (3-strand, however, isprobably not a general feature of subunit II of the cytochromeoxidase family since several sequences either terminate beforethis region or are highly divergent at their C termini (3). Thecommon fold in subunit II of the cytochrome oxidase familyand blue copper proteins suggests a common ancestry. Thisrelationship implies that cytochrome oxidase acquired adinuclear copper center from an ancestral mononuclear cop-per site for a functional reason.
We thank R. Gennis, H. Michel, and A. Thomson for discussionsand access to unpublished results; K. Djinovic and P. Tucker for criticalreading of the manuscript; J. van der Oost for participation in theinitial phase of this project; and D.-J. Slotboom for help with proteinpurification. This work has received financial support from the Euro-pean Union. We also acknowledge fellowships from the EuropeanMolecular Biology Organisation, the Academy of Finland, and theWellcome Trust.
1. Babcock, G. T. & Wikstrom, M. (1992) Nature (London) 356,301-309.
2. Garcia-Horsman, J. A., Barquera, B., Rumbley, J., Ma, J. &Gennis, R. B. (1994) J. Bacteriol. 176, 5587-5600.
3. Saraste, M. (1990) Q. Rev. Biophys. 23, 331-366.4. Malmstrom, B. G. & Aasa, R. (1993) FEBS Lett. 325, 49-52.5. Hill, B. C. (1994) J. Biol. Chem. 269, 2419-2425.6. Kroneck, P. M. H., Antholine, W. A., Riester, J. & Zumft, W. G.
(1988) FEBS Lett. 242, 70-74.7. Antholine, W. E., Kastrau, D. H. W., Steffens, G. C. M., Buse,
G., Zumft, W. G. & Kroneck, P. H. M. (1992) Eur. J. Biochem.209, 875-881.
8. Farrar, J. A., Lappalainen, P., Zumft, W. G., Saraste, M. &Thompson, A. J. (1995) Eur. J. Biochem. 232, 294-303.
9. Van der Oost, J., Lappalainen, P., Musacchio, A., Warne, A.,Lemieux, L., Rumbley, J., Gennis, R. B., Aasa, R., Pascher, T.,Malmstrom, B. G. & Saraste, M. (1992) EMBO J. 11, 3209-3217.
10. Van der Oost, J., Musacchio, A., Pauptit, R. A., Ceska, T. A.,Wierenga, R. K. & Saraste, M. (1993) J. Mol. Biol. 229, 794-796.
11. Steinbrucke, P., Steffens, G. C., Panskus, G., Buse, G. & Ludwig,B. (1987) Eur. J. Biochem. 167, 431-439.
12. Castresana, J., Lubben, M., Saraste, M. & Higgins, D. G. (1994)EMBO J. 13, 2516-2525.
13. Kelly, M., Lappalainen, P., Talbo, G., Haltia, T., Van der Oost,J. & Saraste, M. (1993) J. Biol. Chem. 268, 16781-16787.
14. Kabsch, W. (1988) Acta Crystallogr. A 21, 916-924.15. Otwinowski, Z. (1991) in Isomorphous Replacement and Anom-
alous Scattering, eds. Evans, P., Leslie, A. & Wolf, W. (DaresburyLab., Daresbury, U.K.), pp. 80-86.
16. Zhang, K. Y. J. & Main, P. (1990)Acta Crystallogr. A 46,377-381.17. Jones, T. A., Zou, J.-Y., Cowan, S. W. & Kjelgaard, M. (1991)
Acta Crystallogr. A 47, 110-119.18. Brunger, A. T., Kuriyan, J. & Karplus, M. (1987) Science 235,
458-460.19. Navaza, J. (1987) Acta Crystallogr. A 43, 645-653.20. Lamzin, V. S. & Wilson, K. S. (1993) Acta Crystallogr. D 49,
129-147.21. CCP4 (1994) Acta Crystallogr. D 50, 760-763.22. Kraulis, P. (1991) J. Appl. Crystallogr. 24, 946-950.23. Iwata, S., Ostermeier, C., Ludwig, B. & Michel, H. (1995) Nature
(London) 376, 660-669.24. Blackburn, N. J., Barr, M. E., Woodruff, W. H., van der Oost, J.
& de Vries, S. (1994) Biochemistry 33, 10401-10407.25. Henkel, G., Muller, A., Weissgraber, S., Buse, G., Soulimane, T.,
Steffens, G. C. M. & Nolting, H.-F. (1995)Angew. Chem. Int. Ed.Engl. 34, 1488-1492.
26.- Lappalainen, P., Watmough, N. J., Greenwood, C. & Saraste, M.(1995) Biochemistry 34, 5824-5830.
27. Ryden, L. G. & Hunt, L. T. (1993) J. Mol. Evol. 36, 41-66.28. Adman, E. T. (1991) Adv. Protein Chem. 42, 145-197.
Biochemistry: Wilmanns et al.
Dow
nloa
ded
by g
uest
on
Janu
ary
25, 2
020
















![EMTS Release 2.3 User Group MeetingEMTS User Group 3.12.13 Tuesday, March 26, 2013 Page 1 of 15 Slide 1 - EMTS Release 2.3 User Group Meeting [Scott Christian - EPA] Welcome to the](https://img.pdfslide.net/doc/110x75/6038bddc51e1d909ba72714b/emts-release-23-user-group-meeting-emts-user-group-31213-tuesday-march-26-2013.jpg)












