Embed Size (px)
Citation preview

Arch. Zootec. 59 (226): 233-244. 2010.Recibido: 6-2-08. Aceptado: 22-10-08.
DETECCIÓN MÚLTIPLE DE SNPs RELACIONADOS CONCRECIMIENTO Y CALIDAD DE CARNE EN PORCINO
MULTIPLEX DETECTION OF SNPs ASSOCIATED WITH GROWTHAND MEAT QUALITY IN PIGS
Padilla, J.A.1A, Portilla, F.J.1B, Salazar, J.1C, Parejo, J.C.1D, Martínez-Trancón, M.1E,Rabasco, A.1F, Sansinforiano, M.E.1G, Corral, J.M.2A, Izquierdo, M.2B
y Hernández-García, F.I.2C
1Departamento de Producción Animal y Ciencia de los Alimentos. Facultad de Veterinaria. Universidad deExtremadura. Campus Universitario s/n. 10071 Cáceres. España. [email protected]; [email protected];[email protected]; [email protected]; [email protected]; [email protected]; [email protected];2Centro de Investigación La Orden-Valdesequera. Junta de Extremadura. Apdo. 22. 06080 Badajoz.España. [email protected]; [email protected];[email protected]
PALABRAS CLAVE ADICIONALES
Minisecuenciación (primer extension). Loci H-FABP. Loci MC4R. Loci LEPR.
ADDITIONAL KEYWORDS
Minisequencing (primer extension). Loci H-FABP.Loci MC4R. Loci LEPR.
RESUMENSe describe un método basado en análisis por
primer extension para genotipar simultáneamente7 SNPs relacionados con el crecimiento y la cali-dad de la carne en porcino. Las muestras de ADNgenómico fueron obtenidas de 193 animales per-tenecientes a varias razas (54 Ibérico, 63 Duroc,47 Large White-Landrace x Large White) y a jabalí(29). A partir de las secuencias de los genes H-FABP, MC4R y LEPR, se diseñaron 4 parejas decebadores con el fin de amplificar las regiones delgenoma porcino que contienen esos 7 SNPs. Lareacción de minisecuenciación primer extension(ABI PRISM SNaPshot MultiplexTM kit. AppliedBiosystem) se realizó con cebadores diseñadosen las regiones flanqueantes de cada uno de losSNPs de interés. Los genotipos fueron identifica-dos por mini-secuenciación con ABIPrism 3130 yGeneMapper 3.7 (Applied Biosystems) pudiéndo-se así discriminar, en una sola reacción, losdiferentes alelos de los 7 SNPs de cada animal.Gracias a esta técnica han sido detectados lospolimorfismos T221C y C233T del gen LEPR(Genbank AF092422), que no eran reconociblespor enzimas de restricción. Este método permiteidentificar los animales con genotipo más desea-bles para la selección.
SUMMARYThis work describes a method based on pri-
mer extension analyses to genotype simul-taneously seven SNPs associated with growthand meat quality in pigs. Genomic DNA sampleswere obtained from 193 animals belonging toseveral porcine breeds (54 Iberian, 63 Duroc and47 Large White-Landrace x Large White) and 29wild board. Four pairs of primers have beendesigned from the sequences of the genes H-FABP, MC4R and LEPR and were used for themultiplex amplification of regions in the pig genomecontaining seven SNPs. Primer extension reaction(ABI PRISM SNaPshot Multiplex™ kit. AppliedBiosystems) was realized with primers designedin the flanking regions to each of the SNP ofinterest. The genotypes were identified byminisequencing with ABIPrism 3130 andGeneMapper 3.7. (Applied Biosystems) and thedifferent alleles of the seven SNPs werediscriminated in a sole reaction. In this work havebeen detected, for the first time, the polymorphismsT221C and C233T of the LEPR gene (GenbankAF092422), which are not recognizable byrestriction. This method allows identifying thecarrying animals with the best desirable genotypesfor selection.

Archivos de zootecnia vol. 59, núm. 226, p. 234.
PADILLA ET AL.
INTRODUCCIÓN
En el sector porcino se esta comenzandoa aplicar la selección asistida por marcado-res para mejorar la calidad y rendimiento delos animales (Van der Steen et al., 2005). LosSNPs (single nucleotide polymorphisms)son los marcadores moleculares que se es-tán utilizando en la evaluación de estoscaracteres. Aunque se han detectado milesde lugares polimórficos SNPs en porcino(USMARC, NCBI SNP database), sólo unospocos se han asociado con caracteres eco-nómicamente importantes. La identificaciónprecoz de los animales portadores de losSNPs de interés puede incrementar la preci-sión de la selección y, por tanto, la eficaciade los Programas de Mejora, pero la evalua-ción sólo será eficiente con un método rápi-do y fiable de detección de SNPs. Existe unaamplia gama de métodos y tecnologías parala detección de SNPs (AS-PCR, OLA liga-ción de oligonucleótidos, hibridación, etc.;Syvänen, 2001) que superan algunas difi-cultades del método clásico PCR-RFLP(Suda et al., 2003). La elección del métodoes crítica para responder a las necesidadesprácticas.
El método primer extension o mini-secuenciación (Syvänen, 1999) se basa enla detección de la extensión de un únicodideoxinucleótido (ddNTPs) marcadofluorescentemente a partir de un cebadorespecífico no marcado. El kit de reactivoscomercial SNaPshot (Applied Biosystems)permite la detección multiple de varios SNPs,pero es necesario diseñar los cebadores dePCR para la generación de los ampliconesque contengan los lugares polimórficos ylos cebadores específicos de extensión queinterrogan el lugar SNP.
Entre los genes con influencia sobre lacantidad y calidad de carne de porcino(Kollers et al., 2005) de los que se teníaevidencia de polimorfismos específicos, sehan elegido para el análisis múltipleSNaPshot los genes H-FABP, MC4R y LEPR,por la función que realizan (formación del
músculo, ingesta de alimentos, metabolis-mo y almacenamiento de grasa), por el cono-cimiento de su variabilidad, la longitud delfragmento de PCR específico y la posiciónde la mutación seleccionada, o porque nosean reconocibles por los métodos clásicosde restricción.
El gen H-FABP codifica una proteínarelacionada con el transporte intracelular deácidos grasos en el músculo esquelético ycon la regulación del metabolismo lipídico.Este gen fue ubicado en una región delcromosoma 6 (Gerbens et al., 1997, 2000)que contenía un QTL para caracteres degrasa (Ovilo et al., 2002a; Yue et al., 2003).Se han evaluado los efectos de tres SNPs:Las sustituciones nucleotídicas del intrón-2 C1489T y C1811G del gen H-FABP(GenBank Y16180) y la sustitución T1324Cde la región reguladora 5’UTR del gen(GenBank X98558). Gerbens et al. (1999) encerdos Duroc, encontraron una asociaciónsignificativa entre estos tres SNPs y el con-tenido en grasa intramuscular, grasa dorsaly peso corporal.
El receptor 4 de la melanocortina (MC4R)es una de las moléculas relacionadas con laregulación del comportamiento alimentarioy el peso corporal. El gen que codifica estaproteína se considera, por tanto, candidatopara caracteres de engrasamiento y creci-miento. En este gen, Kim et al. (2000) detec-taron una sustitución nucleotídica G1426Ano sinónima (Asp298Asn) en la secuenciadel gen (GenBank AB021664), que ha mos-trado una asociación significativa con elespesor del tocino dorsal, el crecimiento yel consumo de alimentos.
Los genes de la leptina (LEP) y su recep-tor (LEPR) influyen en la regulación delpeso corporal regulando el control de laingesta y la actividad metabólica celular. Sehan identificado dos lugares polimórficosen la secuencia del exón 4 del gen LEPR(GenBank AF092422): la sustitución T221C(Ovilo et al., 2002b), no reconocible porninguna de las enzimas de restricción dis-ponibles, causa un cambio Met/Thr en la

Archivos de zootecnia vol. 59, núm. 226, p. 235.
DETECCIÓN MULTIPLE DE SNPs EN PORCINO
secuencia del polipéptido codificado, y unasustitución dinucleotídica T232A y C233T,que puede crear cuatro codones: TCA(serina); ATA (isoleucina); TTA (leucina);ACA (treonina). En la sustitución TC→ ATsolamente el primer nucleótido (T232A) esreconocible por restricción con Tsp509I(AATT) (Mackowski et al., 2005). Esta mu-tación parece estar asociada con la topogra-fía de la grasa (Mackowski et al., 2005).
En este trabajo se aborda un métodofiable, basado en la técnica de minise-cuenciación, para el genotipado simultáneode los 7 SNPs, como base para la evaluaciónrutinaria del rendimiento cárnico en el gana-do porcino para sustituir a las técnicas desecuenciación y PCR-RFLP actualmenteutilizadas.
MATERIAL Y MÉTODOS
La detección de los 7 polimorfismos deuna sola base (SNPs) se realizó con la técnicade minisecuenciación o Primer extension(Syvänen, 1999). Se diseñaron los cebadoresde PCR utilizados para la obtención de losamplicones que contienen los SNPs. El di-seño de cebadores de PCR se realizó me-diante el programa Primer3 v0.3.0 (http://frodo.wi.mit.edu) de Rozen y Skaletsky(2000). Se utilizaron secuencias de 200 ba-ses aguas arriba y aguas abajo a partir de laposición de cada SNP, excluyendo 30 basesa ambos lados del SNP. Se utilizaron losparámetros (tm, longitud, % GC, etc.) esta-blecidos por defecto en el programa Pri-mer3. Mediante el programa OligoAnalizer3.0 de Integrated DNA Technologies (http://www.idtdna.com/analyzer/Applications/OligoAnalyzer, se obtuvieron los valoresde estabilidad termodinámica (energía librede Gibbs; ∆G°) de las interacciones detec-tadas entre cebadores (formación de hor-quillas, homodímeros o heterodímeros) quepueden dar lugar a errores en las reaccionesde PCR.
Los cebadores de extensión fueron dise-ñados aguas arriba o aguas abajo y adya-
centes al lugar polimórfico SNP, de formaque tuvieran una tm predicha de alrededorde 60ºC y evitando que formaran estructu-ras secundarias estables entre ellos. Loscebadores de extensión se examinaron fren-te al resto de los amplicones molde paraevitar una posible hibridación no específi-ca. Las longitudes de los cebadores semodificaron por la adición de colasdinucleotídicas (dAT) al extremo 5'.
Las reacciones de PCR se realizaron envolúmenes de 25 µl, conteniendo 100 ng deADN genómico, 1,5 mM de MgCl2, 0,2 mMdNTPs, 0,4 mM de cada cebador, 1 x Buffery 0,5 U de Taq polimerasa (Biotools). Lasamplificaciones se han llevado a cabo en untermociclador PTC 200 (MJ Research) condiferentes condiciones de amplificaciónsegún el fragmento, exceptuando las dife-rencias recogidas en la tabla I en relacióncon el número de ciclos y la temperatura dehibridación utilizada para cada loci. Losdiferentes fragmentos se amplificaron encondiciones uniformes: 94ºC/4 min; 30 ci-clos (94ºC/30 seg, 60ºC/30 seg, 72ºC/30 seg)y una extensión final de 72ºC/15 min.
Los productos de reacciones separadasse mezclaron a volúmenes iguales y se puri-ficaron a 37ºC durante 90 min con 5 U deFosfatasa alcalina (SAP) y 2 U de ExonucleasaI por cada 15 µl de reacción PCR, e incubandoa 37ºC durante 90 min. La acción enzimáticase desactivó a 80ºC durante 20 min.
El examen de los siete SNPs se realizó enuna única reacción utilizando el kit ABIPRISM® SNaPshot Multiplex™ (AppliedBiosystems; Forster City, CA, USA). Cadareacción de minisecuenciación SNaPshot(10 µl) contenía 3 µl de la mezcla de reaccióndel kit, 1 µl de una mezcla de los cebadoresinternos o de extensión a concentracionesequimolares, 3 µl de PCR molde (0,2 pmol) y3 µl de agua desionizada. Las condicionesdel termociclado para la reacción de exten-sión fueron 25 ciclos a 96ºC/10 seg y 60ºC/35 seg. El exceso de ddNTPs marcadosfluorescentemente fueron inactivados porla adición de 1 U de fosfatasa alcalina (SAP)

Archivos de zootecnia vol. 59, núm. 226, p. 236.
PADILLA ET AL.
e incubación a 37ºC durante 60 min, ydesactivándola después a 80ºC durante 20min.
Los productos resultantes se separaronmediante electroforesis capilar en unsecuenciador automático ABIPrism 3130,mezclando 14 µl de Hi-Di formamide (AppliedBiosystems), 0,4 µl de LIZ-120 como estándarinterno de tamaños (Applied Biosystems),y 0,9 µl de la reacción de extensión tratadacon SAP. La identificación de los genotiposse realizó mediante el software GeneMapperver. 3.7 (Applied Biosystems; Forster City,CA, USA).
La fiabilidad del método de genotipadose contrastó por secuenciación doble de 2animales de cada uno de los genotipos de-tectados, empleando secuenciación cíclica(BigDye Terminator v.3.1 Cycle sequencingKits. Applied Biosystems; Forster City, CA,USA) de los productos de los 4 genes.
Para la verificación del método de iden-tificación de los genotipos se analizaron untotal de 193 animales de diversas razasporcinas y de su pariente salvaje el jabalí(tabla VI). El ADN genómico de cada animalfue extraído utilizando el procedimiento pro-puesto por el fabricante del kit Ultra CleanTM
DNA Blood SpinTM (MoBio) a partir de san-gre completa (Duroc, Ibéricos y LWLD xLW) o de lisados hepáticos en el caso de losjabalíes. Las muestras de sangre de los ani-males Ibéricos y Duroc, procedían de losrebaños experimentales del Centro de Se-lección y Reproducción Animal (CENSYRA)de Badajoz. Las muestras de sangre de cer-dos comerciales se obtuvieron en los mata-deros de Cáceres y Arroyo de la Luz(Cáceres). Las muestras hepáticas proce-dían de jabalíes abatidos en las monterías delas campañas de 2006/07 en Extremadura.
Para cada población analizada se hanestimado las frecuencias alélicas medianteel programa GENEPOP 4.0 (Raymond yRousset, 1995).
RESULTADOS Y DISCUSIÓN
En la tabla I se presentan las secuenciasde los cebadores de PCR seleccionados apartir de las secuencias de los genes H-FABP (GenBank X98558), MC4R (GenBankAB021664) y LEPR (GenBank AF092422),sus tamaños, tm y el tamaño esperado delfragmento amplificado en cada reacción dePCR. Además se presenta el nº de ciclos y la
Tabla I. Cebadores de PCR utilizados para la obtención de los amplicones. (PCR primer pairsdesigned for obtention of amplicons).
Locus Secuencia 5'→3' pb tm Nº de Ta Tamañociclos ºC pb
F: ATTGCCTTCGGTGTGTTTGAG 21 62,3 5 63H-FABP AD* 816R: TCAGGAATGGGAGTTATTGG 20 57,5 25 60
F: CAGCCCAAGAGTGAGTTTCC 20 59,8H-FABP H 30 63 185R: AGGACCAGTCCCCTTTCCT 19 59,9
F: TTGATTGGGGTCTTTGTGGT 20 60,2MC4R 30 60 194R: TTGAAGGTTTTCCTCAGTTCTTG 23 59,9
F: CTCTTGCCTGCTGGA ATCTC 21 60,1LEPR 30 60 183R: CCTTCCCTGCAATGTTGTCT 20 60,1
*Cebadores diseñados por Gerbens et al., 1997.
Archivos de zootecnia vol. 59, núm. 226, p. 237.
DETECCIÓN MULTIPLE DE SNPs EN PORCINO
temperatura de hibridación utilizados en suamplificación. Para incrementar la especifi-cidad de la PCR y el éxito de tipificación demuestras degradadas, los tamaños de losamplicones resultantes de las PCR diseña-das fueron menores de 200 pb.
En la tabla II se presentan los valores deestabilidad termodinámica (∆G°) de las inte-racciones entre cebadores (formación de
horquillas, homodímeros o heterodímeros).Se seleccionaron las parejas de cebadorescon valores de estabilidad altos, es decir,con energía libre de Gibbs cercana a 0 (en unrango entre -7 y 0). En las parejas decebadores H-FABPAD (F y R) y LEPR (F yR) pueden producirse heterodímeros más omenos estables. Ninguna de las horquillasque pueden formar los cebadores seleccio-
Tabla II. Formación de dímeros y horquillas entre las parejas de cebadores de PCR. HMD=homodímero, HTD= heterodímero. (Formation of dimers and hairpins between PCR primer pairs.HMD= self-dimer; HTD= hetero-dimer).
Cebadores (∆Gº)* Nº de HTD Nº de H MD Horquillas**kcal/mol. HTD más estable* H MD más estable*
H-FABPAD foward -40,33 6 -3,61 1(0,54/16,7ºC)13 -7,81H-FABPAD reverse -37,15 7 -3,42 1(-0,77/37,3ºC)
H-FABP H foward -37,84 7 -3,14 1 (0,09/23,3 ºC)15 -4,64
H-FABP H reverse -38,06 7 -6,24 1(-1,59/42,7ºC)
MC4R foward -38,12 2 -1,57 1 (0,71/16ºC)13 -3,54
MC4R reverse -40,99 11 -4,67 1(-1,38/47,2ºC)
LEPR foward -37,93 7 -3,14 1(-1,99/55,6ºC)8 -7,04
LEPR reverse -38,1 5 -7,05 1(-0,16/27,6ºC)
*Estabilidad termodinámica definida por energía libre de Gibbs (∆Gº) en kcal/mol. Valores por defecto:concentración de Na + 0,25 µM; concentración de oligonucleótidos 0,25 µM.**Relación de valores: nº de estructuras terciarias (∆Gº de la más estable/temperatura de formaciónde dicha estructura). Valores por defecto: temperatura 25ºC; concentración de Mg2+ 0 mM; concentra-ción de Na + 0,25 µM; concentración de oligonucleótidos 0,25 µM; límite de plegamientos, 20.
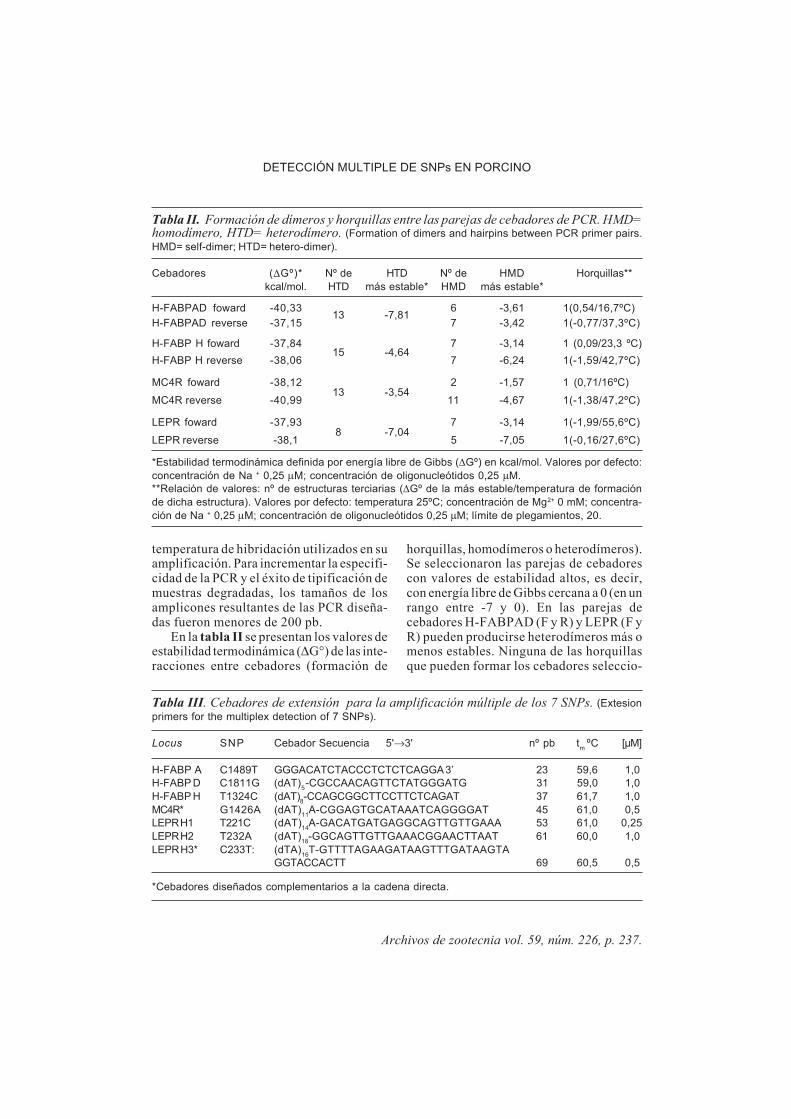
Tabla III. Cebadores de extensión para la amplificación múltiple de los 7 SNPs. (Extesionprimers for the multiplex detection of 7 SNPs).
Locus S N P Cebador Secuencia 5'→3' nº pb tm ºC [µM]
H-FABP A C1489T GGGACATCTACCCTCTCTCAGGA 3’ 23 59,6 1,0H-FABP D C1811G (dAT)5 -CGCCAACAGTTCTATGGGATG 31 59,0 1,0H-FABP H T1324C (dAT)8-CCAGCGGCTTCCTTCTCAGAT 37 61,7 1,0MC4R* G1426A (dAT)11A-CGGAGTGCATAAATCAGGGGAT 45 61,0 0,5LEPR H1 T221C (dAT)14A-GACATGATGAGGCAGTTGTTGAAA 53 61,0 0,25LEPR H2 T232A (dAT)18-GGCAGTTGTTGAAACGGAACTTAAT 61 60,0 1,0LEPR H3* C233T: (dTA)16T-GTTTTAGAAGATAAGTTTGATAAGTA
GGTACCACTT 69 60,5 0,5
*Cebadores diseñados complementarios a la cadena directa.

Archivos de zootecnia vol. 59, núm. 226, p. 238.
PADILLA ET AL.
nados implica un riesgo a considerar en lasreacciones de PCR.
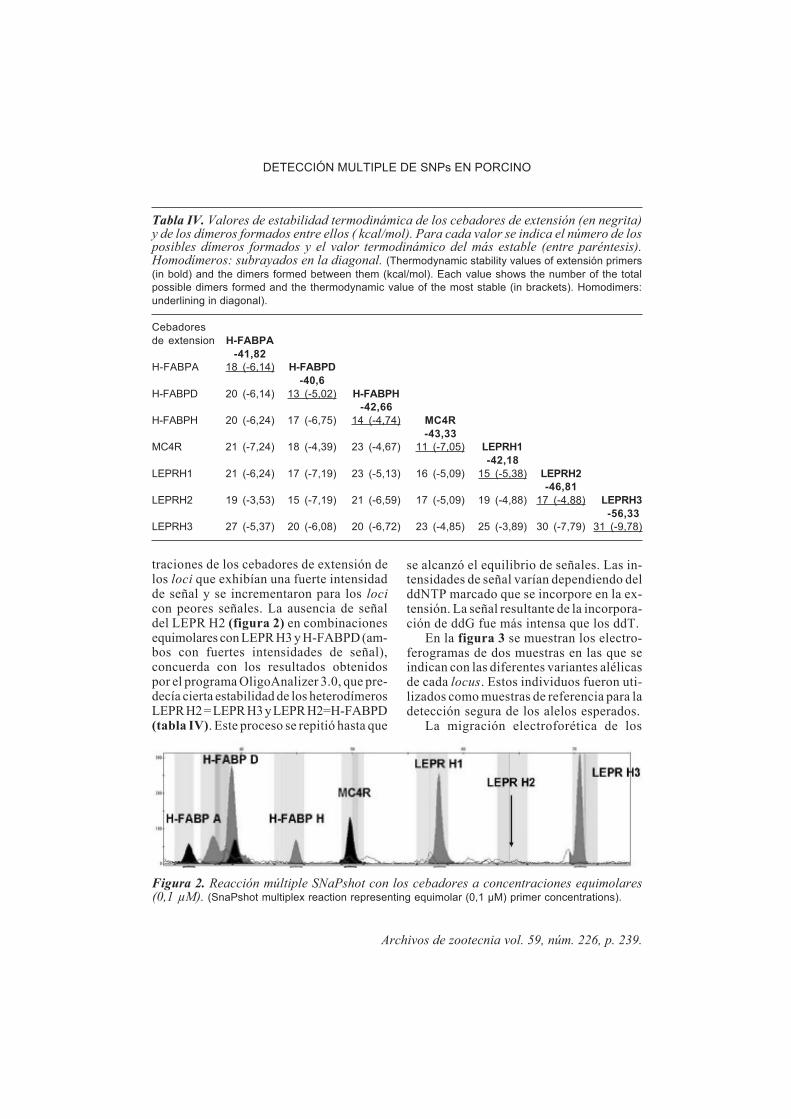
En la figura 1 se muestran los ampliconesPCR obtenidos una vez optimizadas la com-posición de la mezcla y las reacciones deamplificación. Para ello se modificaron, degrado en grado, las temperaturas de hibrida-ción (Ta), a partir de la tm media de cadapareja de cebadores (tabla I), hasta conse-guir la amplificación específica del fragmen-to deseado. Así, en el caso del amplicón H-FABP AD se aumentó la eficacia de la reac-ción forzando a los cebadores a hibridarprimero a 63ºC durante 5 ciclos y posterior-mente a 60ºC durante 25 ciclos. Las condi-ciones finales se indican en material y méto-dos.
En la tabla III se presentan los 7cebadores de extensión diseñados, cadauno de tamaño diferente por la adición alextremo 5' de colas poli-AT de diferente
tamaño, a fin de facilitar su resoluciónelectroforética.
Para identificar la sustitución dinucleo-tídica T232A y C233T del gen LEPR,reconocibles como H2 y H3 respectivamen-te, se diseñó el oligonucleótido de exten-sión LEPR H3 en la cadena complementariapara evitar competencias de hibridación enlugares adyacentes de la cadena. Así mis-mo, el cebador de extensión del gen MC4Rse diseñó complementario a la cadena direc-ta para adecuar la temperatura de fusión a loestablecido en el protocolo de minise-cuenciación múltiple.
En la formación de dímeros y horquillasintramoleculares en los cebadores de exten-sión, se detectan algunas potencialesinteracciones no deseables (tabla IV). Así,existe cierta estabilidad termodinámica en laposible formación de homodímeros en loscebadores LEPR H3 y MC4R, y de losheterodímeros: H-FABP A=MC4R; H-FABPD=LEPR H1; H-FABP D=LEPR H2 y LEPRH2=LEPR H3.
Para optimizar la reacción de extensiónmúltiple de los 7 SNPs fue necesario evaluarlos resultados del ensayo completo, com-probando los componentes de la reacciónde extensión, separación electroforética ytipificación de la señal fluorescente.
En la tabla III se indican las concentra-ciones óptimas para los cebadores de exten-sión. La cantidad de producto PCR genera-do no está correlacionado necesariamentecon la eficiencia o la intensidad de señalobservada en la reacción de extensión(Vallone et al., 2004). Sin embargo, las dife-rencias entre las intensidades fluorescentes(medidas en RFUs, relative fluorescenceunits) dependen de las concentraciones delos cebadores de extensión, de las interac-ciones entre ellos y de la eficacia de emisiónfluorescente de cada fluorocromo.
La optimización del equilibrio de las se-ñales fue un proceso iterativo a partir dereacciones de extensión con cebadores aconcentraciones equimolares (0,1 µM).Posteriormente, se redujeron las concen-
Figura 1. Fragmentos amplificados de cadauno de los genes. Gel de agarosa al 2% p/v.(Amplified fragmnents of each genes analyzed.2% (w/v) agarose gel).
Archivos de zootecnia vol. 59, núm. 226, p. 239.
DETECCIÓN MULTIPLE DE SNPs EN PORCINO
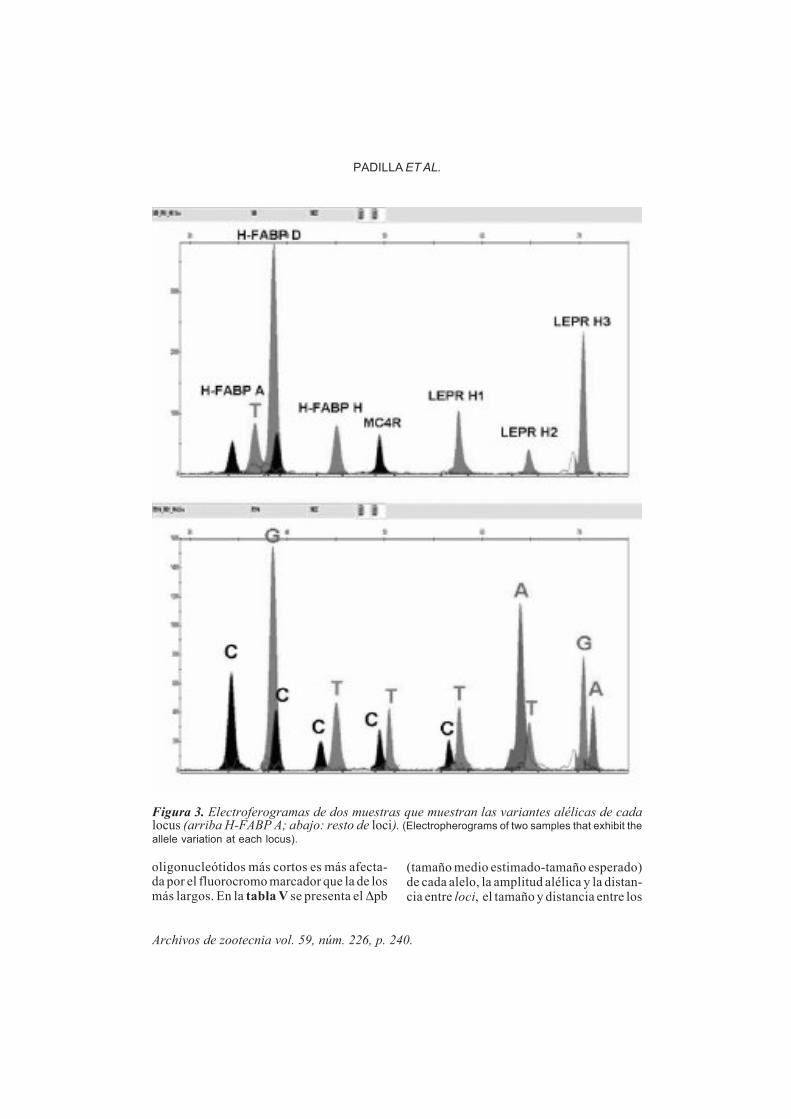
traciones de los cebadores de extensión delos loci que exhibían una fuerte intensidadde señal y se incrementaron para los locicon peores señales. La ausencia de señaldel LEPR H2 (figura 2) en combinacionesequimolares con LEPR H3 y H-FABPD (am-bos con fuertes intensidades de señal),concuerda con los resultados obtenidospor el programa OligoAnalizer 3.0, que pre-decía cierta estabilidad de los heterodímerosLEPR H2 = LEPR H3 y LEPR H2=H-FABPD(tabla IV). Este proceso se repitió hasta que
se alcanzó el equilibrio de señales. Las in-tensidades de señal varían dependiendo delddNTP marcado que se incorpore en la ex-tensión. La señal resultante de la incorpora-ción de ddG fue más intensa que los ddT.
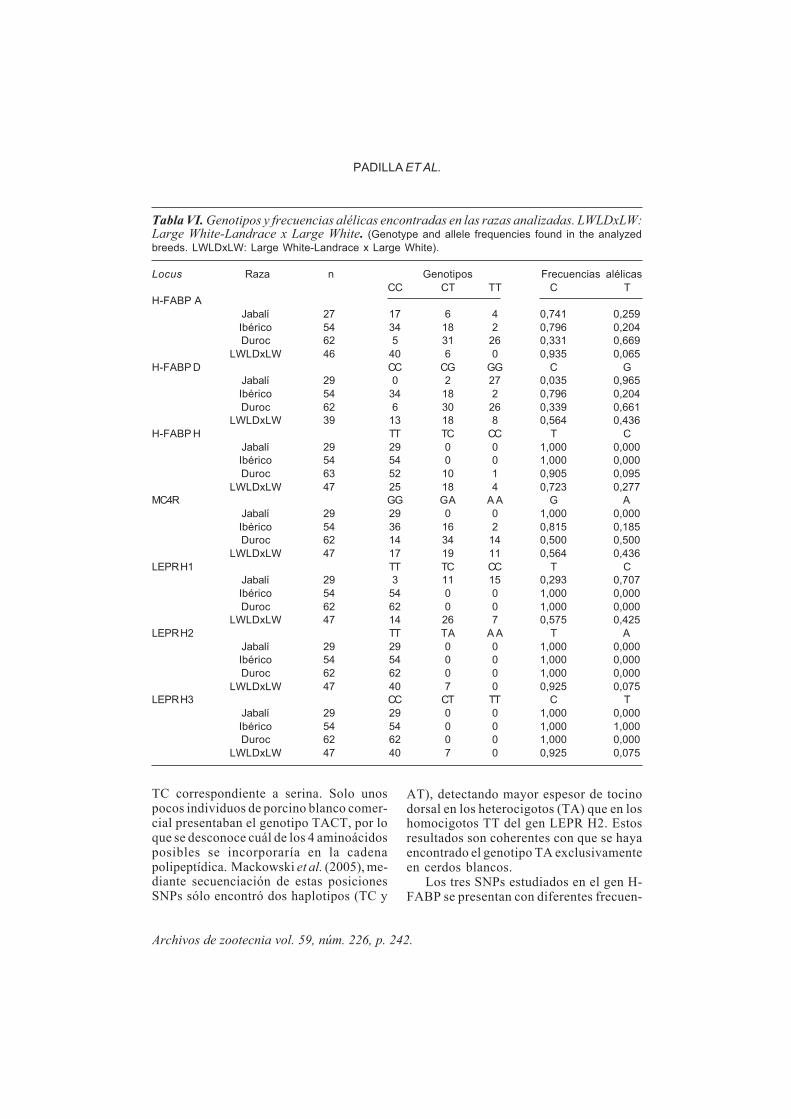
En la figura 3 se muestran los electro-ferogramas de dos muestras en las que seindican con las diferentes variantes alélicasde cada locus. Estos individuos fueron uti-lizados como muestras de referencia para ladetección segura de los alelos esperados.
La migración electroforética de los
Tabla IV. Valores de estabilidad termodinámica de los cebadores de extensión (en negrita)y de los dímeros formados entre ellos ( kcal/mol). Para cada valor se indica el número de losposibles dímeros formados y el valor termodinámico del más estable (entre paréntesis).Homodímeros: subrayados en la diagonal. (Thermodynamic stability values of extensión primers(in bold) and the dimers formed between them (kcal/mol). Each value shows the number of the totalpossible dimers formed and the thermodynamic value of the most stable (in brackets). Homodimers:underlining in diagonal).
Cebadores de extension H-FABPA
-41,82H-FABPA 18 (-6,14) H-FABPD
-40,6H-FABPD 20 (-6,14) 13 (-5,02) H-FABPH
-42,66H-FABPH 20 (-6,24) 17 (-6,75) 14 (-4,74) MC4R
-43,33MC4R 21 (-7,24) 18 (-4,39) 23 (-4,67) 11 (-7,05) LEPRH1
-42,18LEPRH1 21 (-6,24) 17 (-7,19) 23 (-5,13) 16 (-5,09) 15 (-5,38) LEPRH2
-46,81LEPRH2 19 (-3,53) 15 (-7,19) 21 (-6,59) 17 (-5,09) 19 (-4,88) 17 (-4,88) LEPRH3
-56,33LEPRH3 27 (-5,37) 20 (-6,08) 20 (-6,72) 23 (-4,85) 25 (-3,89) 30 (-7,79) 31 (-9,78)
Figura 2. Reacción múltiple SNaPshot con los cebadores a concentraciones equimolares(0,1 µM). (SnaPshot multiplex reaction representing equimolar (0,1 µM) primer concentrations).
Archivos de zootecnia vol. 59, núm. 226, p. 240.
PADILLA ET AL.
oligonucleótidos más cortos es más afecta-da por el fluorocromo marcador que la de losmás largos. En la tabla V se presenta el ∆pb
(tamaño medio estimado-tamaño esperado)de cada alelo, la amplitud alélica y la distan-cia entre loci, el tamaño y distancia entre los
Figura 3. Electroferogramas de dos muestras que muestran las variantes alélicas de cadalocus (arriba H-FABP A; abajo: resto de loci). (Electropherograms of two samples that exhibit theallele variation at each locus).

Archivos de zootecnia vol. 59, núm. 226, p. 241.
DETECCIÓN MULTIPLE DE SNPs EN PORCINO
productos de extensión estimados comopromedio en 30 individuos, con el polímeroPOP7. La precisión de las medidas de lostamaños para varios set de carreras fuemenor de 0,1 pb. La variación de los tamañospara los cebadores extendidos depende dela longitud y composición de la secuencia ydel tipo de marcador fluorescente unido(Vallone et al., 2004).
Los lugares polimórficos fueron ratifica-dos por secuenciación doble de dos anima-les de cada uno de los genotipos detecta-dos hallando plena coincidencia entre losgenotipos definidos mediante primerextension y las secuencias encontradas enlos animales secuenciados. Este métodopresenta, por tanto, una elevada precisión yfiabilidad. Con objeto de conocer su efica-cia en el genotipado poblacional, el métodose ha aplicado en la identificación de losgenotipos de 193 animales de diferentesrazas porcinas y de jabalíes. En la tabla VI semuestran los genotipos y las frecuenciasalélicas encontradas en las diferentes razasy especies analizadas para cada uno de losSNPs estudiados.
El alelo G del locus MC4R que se haasociado con menor espesor del tocinodorsal, menor tasa de crecimiento (medidacomo ganancia media diaria) y menor ingestade alimentos, se encuentra fijado en la po-
blación de jabalí y a frecuencias elevadas enla población de la raza Ibérica, que no hasido seleccionada para estos caracteres confines industriales. En las poblaciones deDuroc y cerdos blancos comerciales anali-zadas, los valores de las frecuencias de losalelos G/A son intermedias. Estos resulta-dos son similares a los obtenidos por Burgoset al. (2006), que analizaron la incidenciaalélica de este gen en las mismas razas.
No se tienen referencias bibliográficassobre la incidencia alélica, o su asociacióncon caracteres productivos, de la mutaciónnucleotídica T/C del LEPR H1, ya que es laprimera vez que ha sido estudiada en pobla-ciones animales. Esta mutación causa uncambio de Met a Thr (Mackowski et al.,2005) en el dominio extracelular de la se-cuencia del polipéptido codificado. En latabla VI, el alelo T se encuentra fijado en lasrazas Ibérica y Duroc por lo que en esaposición siempre se traduce a Met. Por elcontrario, en los jabalíes, el alelo C (Thr) sepresenta en altas frecuencias, que son inter-medias en los cerdos blancos analizados. Lasustitución dinucleotídica TC / AT del genLEPR (H2 y H3) puede crear cuatro codones:TCA (serina); ATA (isoleucina); TTA(leucina); ACA (treonina) en la proteínacodificada. En el jabalí y en las razas Ibéricay Duroc de porcino está fijado el haplotipo
Tabla V. Tamaño y distancia entre los productos de extension. (Sizing and spacing of primerextension products).
Locus Tamaño ∆pb ∆pb Amplitud Distanciaesperado pb alelo 1 alelo 2 alélica1 entre loci2
H-FABP A 23 10,1 12,2 2,1 C/T -H-FABP D 31 6,5 6,8 0,3 G/C 4,7H-FABP H 37 5,3 6,7 1,4 C/T 6,2MC4R 45 4 5,1 1,1 C/T 7,8LEPR H1 53 3,3 4,6 1,0 C/T 7,6LEPR H2 61 2,9 3,8 0,9 A/T 8,2LEPR H3* 69 1,4 2,4 1,0 G/A 8,5
∆pb (tamaño estimado-tamaño esperado).1Diferencia absoluta de tamaño entre los productos de extensión en un locus. 2Diferencia entre dos lociadyacentes= diferencia entre el alelo más grande del locus N y el más pequeño del locus N-1.
Archivos de zootecnia vol. 59, núm. 226, p. 242.
PADILLA ET AL.
TC correspondiente a serina. Solo unospocos individuos de porcino blanco comer-cial presentaban el genotipo TACT, por loque se desconoce cuál de los 4 aminoácidosposibles se incorporaría en la cadenapolipeptídica. Mackowski et al. (2005), me-diante secuenciación de estas posicionesSNPs sólo encontró dos haplotipos (TC y
AT), detectando mayor espesor de tocinodorsal en los heterocigotos (TA) que en loshomocigotos TT del gen LEPR H2. Estosresultados son coherentes con que se hayaencontrado el genotipo TA exclusivamenteen cerdos blancos.
Los tres SNPs estudiados en el gen H-FABP se presentan con diferentes frecuen-
Tabla VI. Genotipos y frecuencias alélicas encontradas en las razas analizadas. LWLDxLW:Large White-Landrace x Large White. (Genotype and allele frequencies found in the analyzedbreeds. LWLDxLW: Large White-Landrace x Large White).
Locus Raza n Genotipos Frecuencias alélicasCC CT TT C T
H-FABP AJabalí 27 17 6 4 0,741 0,259Ibérico 54 34 18 2 0,796 0,204Duroc 62 5 31 26 0,331 0,669
LWLDxLW 46 40 6 0 0,935 0,065H-FABP D CC CG GG C G
Jabalí 29 0 2 27 0,035 0,965Ibérico 54 34 18 2 0,796 0,204Duroc 62 6 30 26 0,339 0,661
LWLDxLW 39 13 18 8 0,564 0,436H-FABP H TT TC CC T C
Jabalí 29 29 0 0 1,000 0,000Ibérico 54 54 0 0 1,000 0,000Duroc 63 52 10 1 0,905 0,095
LWLDxLW 47 25 18 4 0,723 0,277MC4R GG GA A A G A
Jabalí 29 29 0 0 1,000 0,000Ibérico 54 36 16 2 0,815 0,185Duroc 62 14 34 14 0,500 0,500
LWLDxLW 47 17 19 11 0,564 0,436LEPR H1 TT TC CC T C
Jabalí 29 3 11 15 0,293 0,707Ibérico 54 54 0 0 1,000 0,000Duroc 62 62 0 0 1,000 0,000
LWLDxLW 47 14 26 7 0,575 0,425LEPR H2 TT TA A A T A
Jabalí 29 29 0 0 1,000 0,000Ibérico 54 54 0 0 1,000 0,000Duroc 62 62 0 0 1,000 0,000
LWLDxLW 47 40 7 0 0,925 0,075LEPR H3 CC CT TT C T
Jabalí 29 29 0 0 1,000 0,000Ibérico 54 54 0 0 1,000 1,000Duroc 62 62 0 0 1,000 0,000
LWLDxLW 47 40 7 0 0,925 0,075

Archivos de zootecnia vol. 59, núm. 226, p. 243.
DETECCIÓN MULTIPLE DE SNPs EN PORCINO
cias según la raza estudiada. El locus H-FABP A (C/T) es polimórfico en todas lasrazas excepto en el cruzamiento comercialLWLDxLW que casi exclusivamente poseeel alelo C, no habiéndose encontrado elgenotipo TT. Este resultado es similar a loencontrado por Nechtelberger et al. (2001)en las razas de porcino austriacas, Landrace,Piétrain y Large White y por Alfonso yArana (2004) en una línea seleccionada co-mercial originaria de esta última raza.Similarmente, el alelo C es también el másfrecuente en jabalí y en Ibérico, mientrasque en Duroc el más frecuente fue el alelo T(0,669). Alfonso y Arana (2004) en cerdospío negro del País Vasco, de alto desarrolloadiposo, encontraron el alelo T a frecuen-cias similares (0,72). La presencia de estealelo T se ha asociado con mayor contenidoen grasa intramuscular, mayor espesor deltocino dorsal y mayor peso corporal encerdos de la razas Duroc (Gerbens et al.,1999), y de otras razas chinas y europeas(Wei-Jun y Gong-She, 2006). Sin embargo,en Landrace ha sido el alelo C (Nechtelbergeret al., 2001) el que se ha relacionado conmayor ganancia media diaria e índice deconversión de alimentos. En el locus H-FABP D (C/G), el alelo G se presenta afrecuencias muy altas, casi fijado, en jabalí,siendo variable en el resto de razas analiza-das. Este alelo se ha asociado con un mayorcontenido en grasa intramuscular, mayorespesor del tocino dorsal y mayor pesocorporal en cerdos de la raza Duroc (Gerbenset al., 1999). En Large White este alelo se haasociado claramente con mayor gananciamedia diaria (Nechtelberger et al., 2001). En
el caso del locus H-FABP H (T/C), el alelo Testá fijado en jabalí y en porcino Ibérico, yse encuentra a frecuencia muy alta (0,9) enDuroc, las dos razas porcinas con mayorcontenido en grasa intramuscular. En estecaso el alelo T se ha asociado claramentecon un mayor contenido en grasa intramus-cular y mayor espesor del tocino dorsal encerdos de la raza Duroc (Gerbens et al.,1999). En Large White este alelo se ha aso-ciado con mayor ganancia media diaria ycolor de la carne (Nechtelberger et al., 2001).
CONCLUSIONES
Este estudio muestra que el métodoSNaPshot puede reemplazar fiablemente losmétodos de secuenciación y PCR-RFLP,particularmente en la detección de las muta-ciones del exón 4 del gen LEPR H1 (T221C)y H3 (C233T) que no son reconocibles porninguna endonucleasa de restricción. Unavez optimizado, el método de primerextension ha resultado ser rápido y fiable enla determinación del genotipo. En una solareacción se discriminan 7 SNPs relaciona-dos con la cantidad y calidad de la carne enporcino y puede servir para identificar yseleccionar los animales portadores de losgenotipos más deseables.
AGRADECIMIENTOS
A los Drs. Calero y Antúnez del(CENSYRA) de Badajoz por facilitarnosmuestras de porcino.
Este trabajo es parte del ProyectoPDT05B017. Junta de Extremadura. FEDER.
BIBLIOGRAFÍAAlfonso, L. and Arana, A. 2004. Characterisation
of some fatness candidate genes in BasqueBlack Pied and Large White pigs. Arch. Zootec.,53: 411-414.
Burgos, C., Carrodeguas, J.A., Moreno, C.,Altarriba, J., Tarrafeta, L., Barcelona, J.A. andLópez-Buesa, P. 2006. Allelic incidence inseveral pig breeds of a missense variant of pig
melanocortin-4 receptor (MC4R) gene associa-ted with carcass and productive traits; its rela-tion to IGF2 genotype. Meat Sci., 73: 144-150.
Gerbens, F., Rettenberger, G., Lenstra, J.A.,Veerkamp, J.H. and Te Pas, M.F.W. 1997.Characterization, chromosomal localization,and genetic variation of the porcine heartfatty acid-binding proteine gene. Mamm.

Archivos de zootecnia vol. 59, núm. 226, p. 244.
PADILLA ET AL.
Genome, 8: 328-332.Gerbens, F., Van Erp, A.J.M., Harders, F.L.,
Verburg, F.J., Meuwissen, T.H.E., Veerkamp,J.H. and Te Pas, M.F.W. 1999. Effect on geneticsvariants of the Heart fatty acid-binding proteingene on intramuscular fat and performancetraits in pig. J. Anim. Sci., 77: 846-852.
Gerbens, F., De Koning, D.J., Harders, F.L.,Meuwissen, T.H.E., Janss, L.L.G., Groenen,M.A.M., Veerkamp, J.H., Van Arendonk, J.A.M.and Te Pas, M.F.W. 2000. The effect of adipocyteand heart fatty acid binding protein genes onintramuscular fat and backfat content in Meishancrossbred pigs. J. Anim. Sci., 78: 552-559.
Kim, K.S., Larsen, N., Short, T., Plastow, G. andRothschild, M.F. 2000. A missense variant of theprocine melanocortin-4 receptor (MC4R) geneis associated with fatness, growth, and feedintake traits. Mamm. Genome, 11: 131-135.
Kollers, S., Mégy, K. and Rocha, D. 2005. Analysisof public single nucleotide polymorphisms incommercial pig populations. Anim. Genet., 36:426-431.
Mackowski, M., Szymoniak, K., Szydlowski, M.,Kamyczek, M., Eckert, R., Rozycki, M. andSwitonski, M. 2005. Missense mutations in exon4 of the porcine LEPR gene encoding extracellulardomain and their association with fatness traits.Anim. Genet., 36: 135-137.
Nechtelberger, D., Pires, V., Solkner, J., Stur, I.,Brem, G., Mueller, M. and Mueler, S. 2001.Intramuscular fat content and genetic variantsat fatty acid-binding loci in Austrian pigs. J.Anim. Sci., 79: 2798-2804.
Ovilo, C., Oliver, A., Noguera, J.L., Clop, A., Barra-gán, C., Varona, L., Rodríguez, C., Toro, M.,Sánchez, A., Pérez-Enciso, M. and Silió, L.2002a. Test for positional candidate genes forbody composition on pig chromosome 6. Genet.Sel. Evol., 34: 465-479.
Ovilo, C., Melendez, B. and Benitez, J. 2002b.Study of polymorphism of leptin receptor genein Iberian and Landrace pigs. In: XXVIIIInternational Conference on Animal Genetics,11-15 August 2002. Goettingen, paper D097
(Ed. by B. Beening y J-N Meyer). Georg-August-University of Goettingen. Germany. pp. 123.
Raymond, M. and Rousset, F. 1995. GENEPOP(version 1.2): population genetics software forexact tests and ecumenicism. J. Heredity, 86:248-249.
Rozen, S. and Skaletsky, H.J. 2000. Primer3 on theWWW for general users and for biologistprogrammers. In: Krawetz, S., Misener, S.(eds.). Bioinformatics Methods and Protocols:Methods in Molecular Biology. Humana Press.Totowa, NJ. pp. 365-386.
Suda, T., Katoh, M., Hiratsuka, M., Fujiwara, M.,Irizawa, Y. and Oshimura, M. 2003. Use of realtime RT-PCR for the detection of allelic expre-sión o fan imprinted gene. Int. J. Mol. Med., 12:243-246.
Syvänen, A.C. 1999. From gels to chips:“minisequencing” primer extension analysis ofpoint mutations and single nucleotidepolymorphisms. Hum. Mutat., 13: 1-10.
Syvänen, A.C. 2001. Accesing genetic variation:genotyping single nucleotide polymorphisms.Nat. Rev. Genet., 2: 930-942.
Vallone, P.M., Just, R.S., Coble, M.D., Butler, J.M.and Parsons, T.J. 2004. A multiplex allele-specific primer extension assay for forensicallyinformative SNPs distributed throughout themitochondrial genome. Int. J. Legal. Med., 118:147-157.
Van der Steen, H.A.M., Prall, G.F.W. and Plastow,G.S. 2005. Application of genomics to the porkindustry. J. Anim. Sci., 83: E1-E8.
Wei-Jun, P. and Gong-She, Y. 2006. Relationshipbetween genetic variations of 5'-upstream andthe second intron region of H-FABP gene andIMF content in eight pig breeds and wild pig.Chinese J. Agricul. Biotech., 3: 101-107.
Yue, G., Stratil, A., Kopecny, M., Schroffelova, D.,Schroffel Jr., J., Hojny, J., Cepica, S., Davoli, R.,Zambonelli, P., Brunsch, C., Sternstein, I., Moser,G., Bartenschlager, H., Reiner, G. andGeldermann, H. 2003. Linkage and QTL mappingfor Sus scrofa chromosome 6. J. Anim. BreedGenet., 120: 45-55.










![김동환 2009암학회워크샵.ppt [호환 모드]•Basic concepts of SNPs •Applications of SNPs into cancer research ... 21 SNPs a/w antileukemic drug disposition-> 63 SNPs a/w](https://img.pdfslide.net/doc/110x75/601ede878cebc154024e5352/ee-2009oeoefppt-eeoe-abasic-concepts-of-snps-aapplications.jpg)

















