Embed Size (px)
Citation preview

Development of Grossensee (Holstein, Germany): variations in trophic status fromthe analysis of subfossil microfauna
Jutta GUntherOrdinariat fur Bodenkunde, Universitiit Hamburg, Von-Melle-Park 10, D-2000 Hamburg 13, FederalRepublic of Germany
Keywords: paleolimnology, lake development, trophic status, Chironomidae, Cladocera, Ostracoda
Abstract
Microfossil analysis was carried out on 90 samples of a sediment core from Grossensee. The diversity ofsubfossil Cladocera, Chironomidae, Chaoborus, and Ostracoda reflects at least five different stages ofdevelopment. Three extended cycles correspond to different climatic periods in the first 10 000 years of thelake's history, while during the last 2 000 years, two shorter cycles can be identified which correlate withproofs or even historical events for human activity in the catchment area. During this time the profundal faunachanged from stenoxibiontic to euryoxibiontic species. In the planktonic fauna a succession of threeEubosmina types, B. longispina, B. coregoni kessleri, andB. coregoni coregoni, was observed. The chydoridand ostracod assembly of the littoral showed no clear change.
Introduction
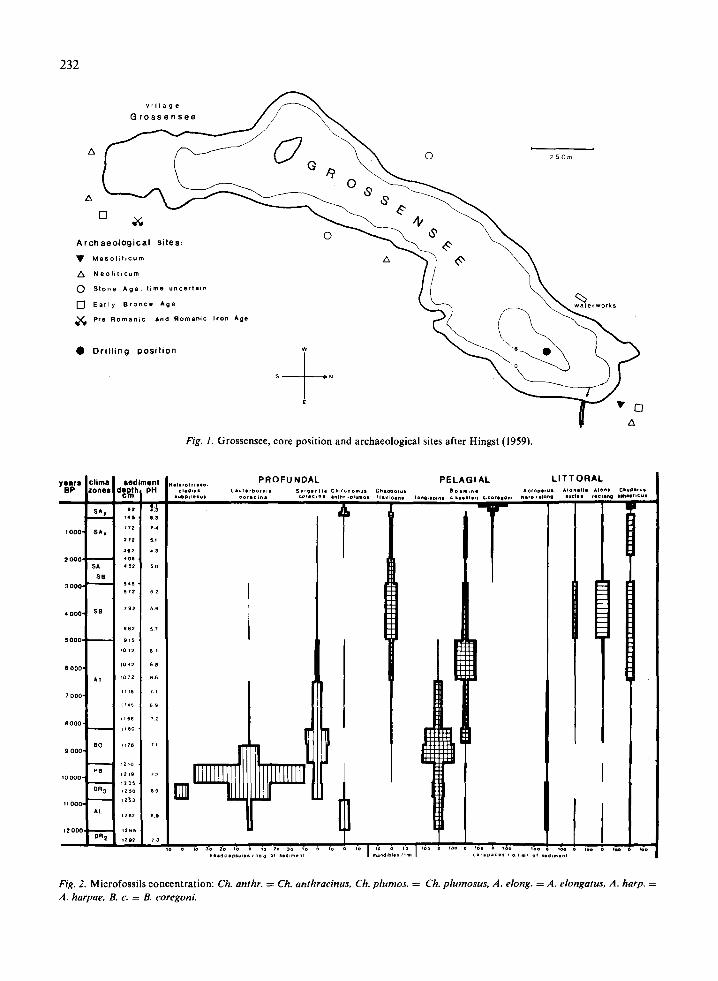
Grossensee (Lake Grossen) is situated east ofHamburg (Fig. 1). It has an area of 75 ha, lies 38 mabove sea level and has an average water depth of12 m. The lake is wind-exposed and stretches overca. 2.3 km from SSW to NNE. A small island marksthe border line between the shallow southern andthe deep northern part. According to pollen analysis(Averdieck 1983), sedimentation began after thelast glaciation, about 12 000 BP.
Materials and methods
The investigated sediment core was taken fromthe deepest part of the lake, at 15.65 m. A modifiedLivingstone corer was used. The core was 12.93 mlong and consisted of different types of gyttja. Thelowest layers were rich in carbonates, clay and silt.From a depth of 9.50 m (ca. 5 000 BP) to the surfaceit consisted of lime-free gyttja. Coarse sands andcalcareous material form the base of the core.
Microfossil analysis is based on the identificationof the head capsules of chironomids, mandibles ofChaoborusflavicans, head shields or carapaces ofcladocera, and valves of ostracods.
Chironomid analysis was performed on 52 sam-ples of I ml volume and 38 samples of ca. 10 gsediment. For cladocera analysis 35 samples wereselected, 1-7 for each climate period. Each samplewas equivalent to 0.03 ml of sediment. Ostracodswere collected from 38 larger samples.
The samples were prepared according to standardmethods (Frey 1959; Hofman 1971; Lffler 1978).For identification papers of Diebel & Pietrzeniuk(1975), F6lssner (1972), Hofmann (1971) and Klie(1938) were also used.
Results
I found 1778 head capsules of 44 different chiro-nomid species. Each sample of I ml contained anaverage of 8 head capsules (2-25).From Allerod(AL) until early Boreal (BO) period (12 000-9 000
Hydrobiologia 103, 231-234 (1983).© Dr W. Junk Publishers, The Hague. Printed in the Netherlands.
232
vllag e
Grossensee
A
A
C
Arch
V Me
A Ne
0 St
[ Ea
a Pr
O
O
a
Dr
S N
Fig. 1. Grossensee, core position and archaeological sites after Hingst (1959).
PROFUNDALLutfrborni Srenlil Chironomul COholorus
c1Otin& curlecna anthr DpUmO. Il-liln
[
PELAGIAL LITTORALBolmin. ACrOer0u5 l Aionell AIOn ChydoP1I- ....... ... A-1 ...--- - -. ----- I ... iii. ~ ~ . w _
[
r
I
jo o 20 20 o io05 o to o to o o t o i ooIhelcapsles /iog DI sIe. menf mnandble Iml
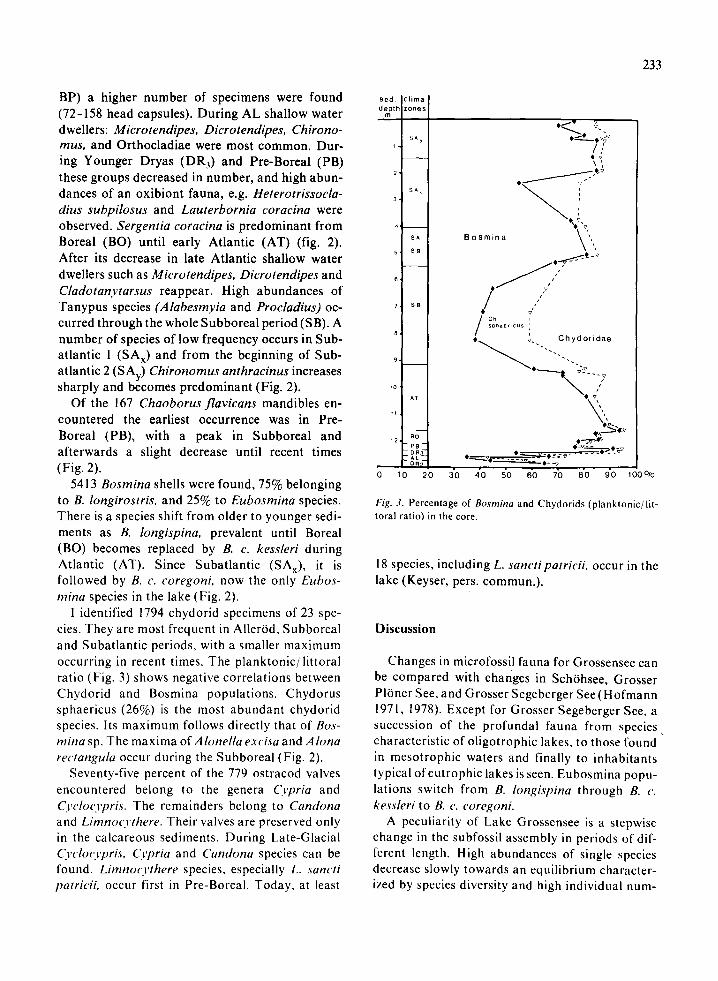
Fig. 2. Microfossils concentration: Ch. anthr. = Ch. anthracinus, Ch. plumos. = Ch. plumosus, A. elong. = A. elongatus, A. harp. =A. harpae. B. c. = B. coregoni.
waterworks
sedimentpH
Htr110 lriSo.
.ubpilol.
yearsBP
1000-
2 000
3000,
4 000
5000.
6 000
7 000'
8000.
9 000
10 000-
11 000-
12000
climecones
SA,
SA6
So
Sc
SB
AT
B0
PB
OR3
AL
DR,
1:5013.3
.:.4
5.I
43
50
52
S I
56
61
6.8
71
7 2
71
69
52
14
wT;
I72
°5a6o
512
102
882
9t5
10 12
1042
1072
1115
1160
1171
1210
1219
~355
1250I ,1
12 3
1262
125
1292
- | | I I -
|- -. - ·-
i -l - -j I
D
- . - - -ln1 · .r ......l
dst
l
1 f
IJ- -- - - - ·- ·- i - ~ . i , n~ -
I FAA A | AA .:_I0O 1O 0 100 I14 1J 0 100C-l-i- I 1 -dilrnl
233
BP) a higher number of specimens were found(72-158 head capsules). During AL shallow waterdwellers: Microtendipes, Dicrotendipes, Chirono-mus, and Orthocladiae were most common. Dur-ing Younger Dryas (DR3 ) and Pre-Boreal (PB)these groups decreased in number, and high abun-dances of an oxibiont fauna, e.g. Heterotrissocla-dius subpilosus and Lauterbornia coracina wereobserved. Sergentia coracina is predominant fromBoreal (BO) until early Atlantic (AT) (fig. 2).After its decrease in late Atlantic shallow waterdwellers such as Microtendipes, Dicrotendipes andCladotanytarsus reappear. High abundances ofTanypus species (Alabesmyia and Procladius) oc-curred through the whole Subboreal period (SB). Anumber of species of low frequency occurs in Sub-atlantic 1 (SAx) and from the beginning of Sub-atlantic 2 (SAy) Chironomus anthracinus increasessharply and becomes predominant (Fig. 2).
Of the 167 Chaoborus flavicans mandibles en-countered the earliest occurrence was in Pre-Boreal (PB), with a peak in Subboreal andafterwards a slight decrease until recent times(Fig. 2).
5413 Bosmina shells were found, 75% belongingto B. longirostris, and 25% to Eubosmina species.There is a species shift from older to younger sedi-ments as B. longispina, prevalent until Boreal(BO) becomes replaced by B. c. kessleri duringAtlantic (AT). Since Subatlantic (SAX), it isfollowed by B. c. coregoni, now the only Eubos-mina species in the lake (Fig. 2).
I identified 1794 chydorid specimens of 23 spe-cies. They are most frequent in Aller6d, Subborealand Subatlantic periods, with a smaller maximumoccurring in recent times. The planktonic/littoralratio (Fig. 3) shows negative correlations betweenChydorid and Bosmina populations. Chydorussphaericus (26%) is the most abundant chydoridspecies. Its maximum follows directly that of Bos-mina sp. The maxima of A lonella exc isa and A lonarectangula occur during the Subboreal (Fig. 2).
Seventy-five percent of the 779 ostracod valvesencountered belong to the genera C pria andCy'clocipris. The remainders belong to Candonaand Limnocy there. Their valves are preserved onlyin the calcareous sediments. During Late-GlacialCyclocpris, Cpria and Candona species can befound. Limnocythere species, especially L. sanctipatricii, occur first in Pre-Boreal. Today, at least
seddepthm
2-
3.
4
5'
6,
8.
12.
clima
zones
SA
SA
sa
SB
AT
BO
PB--oR
OR7 _- __
0 10 20 30 40 50 60 70 80 90 1 00 /o
Fig. 3. Percentage of Bosmina and Chydorids (planktonic/lit-toral ratio) in the core.
18 species, including L. sanctipatricii, occur in thelake (Keyser, pers. commun.).
Discussion
Changes in microfossil fauna for Grossensee canbe compared with changes in Schohsee, GrosserPl1ner See, and Grosser Segeberger See (H ofmann1971, 1978). Except for Grosser Segeberger See, asuccession of the profundal fauna from speciescharacteristic of oligotrophic lakes, to those foundin mesotrophic waters and finally to inhabitantstypical of eutrophic lakes is seen. Eubosmina popu-lations switch from B. longispina through B. c.kessleri to B. c. coregoni.
A peculiarity of Lake Grossensee is a stepwisechange in the subfossil assembly in periods of dif-ferent length. High abundances of single speciesdecrease slowly towards an equilibrium character-ized by species diversity and high individual num-
I I~~~~ v
_ -~~~~~ _
I
· · ·_^

234
bers. Except for Allerod near the end of each periodthere is an increase in Bosmina abundances. There-after the population decreases rapidly and Ch.sphaericus spreads (Fig. 3), a precursor of a newbiocoenosis and not necessarily indicative of moreeutrophic status. Five cycles can be observed.
I. Initial status of water. Between Older Dryasand Aller6d an eutrophic shallow lake comes intoexistence.
2. Together with the decrease of temperature inYounger Dryas species diversity diminishes. Apopulation of species characteristic for lakes (e.g.L. sancti patricii) has developed up to the end ofAtlantic (Fig. 2). A high planktonic/ littoral ratio isseen (Fig. 3). In the beginning the profundal faunaconsists of highly oxibiontic species but it is fol-lowed successively by species with less oxygen de-pendence. In the end of Atlantic species diversityand high biomass of the whole microfauna reflect aclimax state of a lake with high biological activity.The breakdown of this community at about 6 000BP (Fig. 2) represents the end of the oligotrophicphase. The gyttja becomes lime-free at this point.
3. The Subboreal sediment accumulation rate istwice that of Atlantic, perhaps due to water leveloscillations or catchment disturbance by humanactivity (Fig. ). Only a few species present in Atlan-tic assembly remain. Of the species which emerge inSubboreal most thrive in more dystrophic condi-tions (e.g. Chaoborus, Alonella excisa, Alona rec-tangula (Fig. 2). The changes in chironomid popu-lation as well as the sharp decline of Bosmina inrelation to the Chydorid assembly support the as-sumption that large lowland areas are influencingthe metabolism of the lake (Ohle 1972). Ranking ina trophic scheme is difficult but increasing eutroph-ication up to early Subatlantic (SB/SA, about3 000 BP) is clearly taking place.
4. Archaeological findings of early Iron Age(about 2000 BP) indicate human activity. Theplanktonic/littoral ratio diminishes as does thespecies diversity of chironomids. The new cycleresembles the previous one, but due to human dis-turbance it comes to an end before climax. Theseobservations correlate with a land clearing and set-tling period about 1240 AD.
5. In the early part of Subatlantic 2 (SAy) thenumber of chironomid species decreases to half ofthe previous value. In the newly emerging assemblyChironomus anthracinus becomes established and
dominates in the profundal zone. B. c. coregoni isthe only Eubosmina. The lake changes to a Chi-ronomus anthracinus type (Thienemann 1954).Phases of stability and disturbance occur moreoften than in times before human impact. The spe-cies assembly, however, is stable and has notchanged during SAy.
Acknowledgements
I am indebted to Deutsche Forschungsgemein-schaft for financial support. I wish to thank Profes-sor Dr. W. Ohle (P16n), who drew my attention tothe problem and Professor Dr. W. Scharpenseel(Hamburg), who gave me the sediment samples andenabled me to carry out the study. I want to thankDr. W. Hofmann for his scientific help, and alsoV6ge and Voge who allowed me to use their data ofwater depth for Fig. 1. I should also like to thankmy husband for his assistance.
ReferencesAverdieck, F.-R., 1983. Palynological investigations of the sed-
iments of ten lakes in Eastern Holstein, North Germany.Hydrobiologia, this volume.
Diebel, K. & Pietrzeniuk, E., 1975. Mittel- und jungpleisto-zane Ostracodenfauna des Raumes Potsdam-Brandenburg-Rathenau in stratigraphischer und kologischer Sicht. Z.geol. Wiss. Berlin 3:1197-1233.
Flbssner, D., 1972. Kiemen- und Blattfiisser, Branchipoda,Fischlause Brachiura. In: Dahl, M. & Peus, F. (eds.). DieTierwelt Deutschlands 60: 1-499.
Frey, D. G., 1959. The taxonomic and phylogenetic significanceof the head pores of the Chydoridae (Cladocera). Int. Revueges. Hydrobiologia 44: 27-50.
Hingst, H., 1959. Vorgeschichte des Kreises Storman. Wach-holtz, Neumuiinster.
Hofmann, W., 1971. Die postglaziale Entwicklung der Chirono-miden und Chaoborus Fauna (Dipt.) des Schohsees. Arch.Hydrobiol. Suppl. 40: 1-74.
Hofmann, W., 1971. Zur Taxonomie und Palokologie subfos-siler Chironomiden (Dipt.) in Seesedimenten. Arch. Hydro-biol. Beih. Ergebn. Limnol. 6: 1-50.
Hofmann, W., 1978. Analysis of animal microfossils fromGrosser Segeberger See (F.R.G.). Arch. Hydrobiol. 82:316 346.
Klie, W., 1938. Ostracoda, Muschelkrebse. In: Die TierweltDeutschlands. Dahl, Jena 34: 1-230.
Loffler, H., 1967. Ostracoda. In: llies, J. (ed.). LimnofaunaEuropaea. Enke, Stuttgart: 162-172.
Ohle, W., 1972. Die Sedimente des Grossen Ploner Sees alsDokumente der Zivilisation. Jb. Heimatkunde Kreis Plon. 2:7-27.
Thienemann, A., 1954. Chironomus. Die Binnengewasser 20:1-834.





























