Embed Size (px)
Citation preview

DISCUSSION
Self-incompatibility mechanism in angIOsperms IS widespread and has
received considerable attention in recent years (Takayama and Isogai, 2005; McClure
and Franklin-Tong, 2006). Hamelia patens of the family Rubiaceae is grown chiefly
for its attractive flowers throughout the year. The plant is propagated mainly by
vegetative means due to failure of seed-set. In order to study the reproductve failure,
detailed analyses ware carried out.
According to Brewbaker (1959) and Vuilleumier (1967), there are six families
VIZ. Primulaceae, Saxifragaceae, Rubiaceae, Oleaceae, Apocynaceae and
Polygonaceae displaying both homomorphic and heteromorphic systems of self
incompatibility. The family Rubiaceae shows diversity in sexual systems. Florally
homomorphic and heteromorphic as well as sporophytic and gametophytic
incompatibility systems have been reported (Bir Bahadur, 1968; Bawa and Beach,
1983; Faivre, 2002). In Rubiaceae, self-incompatibility studies have been done in
Coffea arabica and C. canephora. In C. canephora, the self-incompatibility system is
of the gametophytic type and is found controlled by a single locus with multiple
alleles (Berthaud, 1980). H patens is homomorphic because it does not show any
morphological differences in the floral characters.
Fertilization in flowering plants requires remarkable cellular coordination to
carry sperm cells to the ovules through stylar tissues. The study of factors that control
the reproductive process in higher plants necessitates a thorough knowledge of the
pollen and gynoecium. While considerable information is available regarding the
pollen and the ovary, not much is known about the structure of stigma (Maheswari,

1950; Hulskamp et aI., 1997; Spielman et aI., 1997; Kuzoff et aI., 1999,2001; Soltis
and Hufford, 2002; Yang et aI., 2003).
Anther has been a matter of interest for investigators not only as the male
reproductive organ of the flower but also as a system to facilitate breeding
manipulations in crop plants. In H patens, the fully developed microsporangium
consists of a mass of haploid pollen enclosed within the sporangial wall. Mature
anther is bithecous and tetrasporangiate. The development of anther corresponds to
the reduced type. The outermost layer is epidermis. The layer of cells immediately
beneath the epidermis is endothecium. The cells of the endothecium became radially
elongated and decorated with fibrous bands that run upward from the inner tangential
walls, ending near the outer wall of each cell as an incomplete ring. Fibrous
thickening of endothecium was reported by Periasamy and Kandasamy (1981) in
Annona squamosa, Strittmatter and Galati (2001) in Mysotis azorica and M laxa and
Oliveira and Mariath (2001) in Anacardium occidentale. The tapetum is a specialized
layer of cells whose development is very much in harmony with the events occurring
in the sporogenous cells. At maturity, the tapetal cells collapse. According to
Raghavan (1999) tapetal cells attain the maximum development when the microspores
are in the tetrad stage, after which the cells collapse. Functional as well as molecular
aspects of tapetum have been studied by Mariani et al. (1990) and Staiger et al.
(1994). In H patens anther dehiscence occur between 11-12 pm. Zavada (1984)
revealed that in angiosperm pollen, the reticulate perforate exine sculpturing and
imperforate and microperforate exine sculpturings are associated with the
gametophytic self-incompatibility. However, perforate-matted exine sculpturing was
noticed in H patens, but the plant showed many characters of gametophytic
incompatibility.
78

The relationship between stigma characteristics and the self-incompatibility
was studied by Heslop-Harrison et ai. (l975a). Stigmas not only discriminate between
different pollen grain but also promote out crossing or self-fertilization by the
coordinated timing of their maturation with pollen release. Angiosperm stigmas is
classified into two broad categories, wet and dry, depending on whether or not they
possess a surface secretion (Heslop-Harrison and Shivanna, 1977; Heslop-Harrison,
1981). Stigma of H patens is wet and papillate with sticky exudate. Rupture of cuticle
layers during development of wet stigma is associated with exudate production
originating from epidermal and adjacent cell layers that accumulates in the
intercellular spaces of the stigmatic tissue and below the cuticle-pellicle layer of
epidermal cells. The cuticle-pellicle layer is tom away with continued secretion of
exudates (Konar and Linskens, 1966b; Dumas et aI., 1978). Degeneration of stigmatic
surface cells was accompanied by the profuse accumulation of surface exudate in
older flowers. Copious stigmatic secretions could enhance pollen hydration,
germination and tube growth. The papillae are modified epidermal cells that cover the
stigma and are responsible for most of the secretions on the stigma surface (Heslop
Harrison and Shivanna, 1977; Considine and Knox, 1979; Owens and Horsefield,
1982). In H patens, from the early bud stage (4 days before anthesis) onwards, the
stigma surface appears as an irregular mass due to the presence of sticky exudate.
Taking into account all the features described by Heslop-Harrison and Shivanna
(1977), it seems fairly justifiable to categorize the stigma of H patens under group III
of wet type.
The stigma is reported to be receptive at the time of anthesis in many tree
crops such as peach, apricot, sweet cherry, apple and kiwi (Sanzol and Herrero, 2001).
Herrero (1983) reported that stigma of pear was not receptive at anthesis and pollen
79

germination increased after four days. Likewise, stigma in apricot was not mature at
the balloon stage, but attained higher receptivity in two and four days after anthesis
(Egea et aI., 1991). Contrary to this, H patens stigma is receptive from the very early
stages of bud development.
According to Heslop-Harrison and Shivanna (1977), receptivity in wet
stigma often coincides with the accumulation of stigmatic secretion. In vivo pollen
germination revealed that even in the IV stage of the bud development, 45% pollen
germination was noticed. Stigma receptivity of H patens was also confirmed by the
activity of esterase enzyme.
Stigmatic secretions play a key role in pollen capture and adhesion (Dumas
and Gaude, 1983; Dumas et aI., 1984). In solid-styled sporophytic self-incompatible
species, the pollen tube was arrested on the stigma. In yellow passion fruit, two
inhibition sites ofpollen-tube growth, in stigma and style, were identified (Rego et aI.,
2000). In the case of dry stigma, the pellicle acts as the first site of molecular contact
for the pollen grains (Mattsson et aI., 1974; Heslop-Harrison, 1978; Dumas et aI.,
1984). The pellicle serves two functions, one is to confine the exudate to the stigma
and the other is to localize and expose pollen recognition molecules in a prominant
and accessible position.
In H patens, the style is solid with a multi-layered stigmoid tissue. The
stigmoid tissue has been named differently by different authors: conducting tissue
(Hanf, 1935) and transmitting tissue (Vasil and Johri, 1964). Most dicotyledons have
a closed style in which the pollen tubes grow through the transmitting tissue, whereas
their longitudinal walls are separated by the intercellular matrix (1M). Chemical
analysis has shown that the 1M contains free sugars, polysaccharides, free amino
80

acids, proteins, glycoproteins, proteoglycans and phenolic compounds, usually
thought to function in pollen tube nutrition, recognition, and guidance (Cheung,
1996b). According to Maheswari (1950), the transmitting tissues serve an easy
passage for the pollen grains and provides nutrients for the growth of pollen tubes
towards the ovary. In H patens, the transmitting tissue is made up of elongated,
parenchymatous cells connected end to end, which is rich in cytoplasm. According to
Garg and Bhatnagar (1988), in Withania, the transmitting tissue is in continuity with
the placental epidermis. The transmitting tissue provides continuity, through or over
which pollen tubes grow en route the ovary (Cresti et. aI., 1979; Tilton and Homer,
1980; Clifford and Owens 1990). In Acer saccarum, the path from stigma to
micropyle is open (stylar canal) but pollen tube path is a continuous tract of mucilage
from the stigma surface through the solid style to the ovule micropyle (Webb and
Williams, 1988; Peck and Lersten, 1991). In soybean stigma, the cells in the stylar
transmitting tissues are secretory, producing exudate, detectable in the intracellular
spaces (Tilton et. aI., 1984). It is quite obvious that the pollen tubes while growing
through the intracellular spaces initially will have to exert pressure on the spaces to
widen. The normal occurrence of longitudinal separation of transmitting tissues
ensured that an axial array of channels filled with exudate, through which the pollen
tube can grow.
In Prunus avium and in Petunia hybrida the stylar vascular elements are found
around the transmitting tissue (Konar and Linskens, 1966a; Uwate and Lin, 1981). In
Ornithogalum, ground tissue consists of elongated parenchyma cells and vascular
tissues (Tilton and Homer, 1980).Vascular element in H patens are seen outside the
centrally placed transmitting tissue as distinct patches.
81

The ovary bears ovules on a specialized ridge on the ovary wall termed
placentum. The ovule generally develops by periclinal divisions of cells of either L2
or L3 or both these layers of the placenta (Raghavan ,1999). The mature ovule is
raised on a stalk and consists of a mass of homogenous cells of the nucellus, protected
by one or two multilayered covering, the integument. The degree of curvature of the
ovule and the number of integuments covering the nucellus add to the variation in the
morphology of embryosac developed on the placenta. Extensive studies have been
carried out on the structural details of the ovule and embryosac (Maheswari, 1950,
1963; Tilton, 1981a, b; Tilton and Lersten, 1981; Cresti et aI., 1992). The ovary in H
patens is pentacarpellary, syncarpus, pentalocular with centrally placed axile
placentum.
In H patens, meiotic chromosome analysis was carried out in order to analyse
the possibilities of any meiotic abnormalities that may lead to the sterile pollen grains.
However microsporogenesis was normal and showed 12 bivalents at metaphase-l
which is first in the species. Futher stages of meiosis were found normal and pollen
sterility was only 10%.
Mature angiosperm pollen grains contain vegetative and generative nuclei,
complete with cell wall and plasma membrane (Raghavan,1999). This arrangement is
accomplished soon after meiosis. Subsequently, the generative cell undergoes a
second mitosis to form two sperm cells required for double fertilization.
"Tricellular" pollen completes this division before it is released from the anther,
whereas "bicellular" pollen undergoes this division only later, within the elongating
pollen tube. Brewbaker (1957) pointed out a correlation between incompatibility
systems and the number of nuclei in the pollen. In general, species showing
gametophytic and sporophytic incompatibility systems have two or three nuclei
82

respectively. Most dry stigmatic SI systems were historically associated with
tricellular pollen grains and bicellular pollen grains with wet stigmas and stylar SI.
Stigmatic SI in association with bicellular pollen and a dry or wet stigma was
considered anomalous (Brewbaker, 1957; Heslop-Harrison, 1975; de Nettancourt,
1977). Recognition and rejection of bicellular pollen on wet stigma was reported in
Rubiaceae (Brewbaker, 1957; de Nettancourt, 1977, 1997; Franklin et aI., 1995). Wet
stigmas with binucleate pollen grains were observed in H patens. Since SI in all taxa
with bicellular pollen has been demonstrated to be under gametophytic control,
regardless of whether recognition and rejection takes place on a dry/wet stigma or in
the style, it is assumed that the SI in Rubiaceae is under gametophytic control.
Pollen viability is considered as an important parameter of pollen quality
(Dafui and Firmage, 2000). In physical method, pollen viability is generally tested by
FCR which has been suggested by Heslop-Harrison and Heslop-Harrison (1970). FeR
test was found satisfactory for a range of pollen species (Shivanna and Heslop
Harrison, 1981; Heslop-Harrison et aI., 1984; Jain and Shivanna, 1988). Most of the
pollen grains are metabolically quiescent and highly desiccated, ranging from 15 to
35% water content, when released from anthers (Heslop-Harrison, 1979; Buitink et
aI., 2000). Shivanna and Heslop-Harrison (1981) reported good correlation between
FCR and in vitro germination. In H. patens, 90% of pollen grains were intensely
stained with FDA showing its viability status.
In biological method, germinability is used to test the viability of pollen
grains. In vitro pollen germination method is rapid, reasonably simple and the most
commonly used for assessing pollen viability. Pollen germination generally requires a
carbohydrate source. Sucrose is the most commonly used carbohydrate source in
culture medium which serves the maintenance of osmotic pressure of the medium
83

and as carbon source for metabolism which is essential for pollen germination (Johri
and Shivanna, 1985). The optimum sugar concentration required for the pollen
gennination differs considerably with species (Johri and Vasil, 1961). Besides the
carbohydrate source, boron and calcium also play an important role in pollen
gennination and tube growth (Brewbaker and Kwack, 1963; Kwack, 1967; Balatkova
et aI., 1980; Steer and Steer. 1989). A culture solution containing sucrose and
minerals suggested by Brewbaker and Kwack (1963) is widely used for in vitro pollen
gennination. By manipulating individual components, it has been possible to achieve
satisfactory in vitro germination in a large number of taxa with binucleate pollen. In
the present study, Brewbaker and Kwack's medium supplemented with 25% sucrose
showed optimum germination of 72.5%. Since the pollen sterility is less (10%) and
pollen germination is more, pollen sterility cannot be attributed as the reason for lack
of seed- set in this plant. In many species in vitro pollen germination showed positive
correlation with fruit and seed-set (Janssen and Hermsen, 1976; Akihama et aI.,
1978). But in H patens, a negative correlation was found between pollen germination
and seed-set.
5.1 Pollen-pistil interactions
During pollination, numerous cell to cell interaction events occur between the
cells of the sporophyte and the male gametophyte. The timing of pollination is
critical, because the stigmatic surface of the pistil is only receptive to pollen for a
relatively short period. Pollination outside this window of female receptivity will
result in reduced seed set or no seed set (Herrero, 2003). The recognition events
appear extremely complex and are not fully understood (Clarke and Gleeson, 1981;
de Nettancourt 1984; Dumas et aI., 1984; Knox, 1984). The influence of the stigma
and style on pollen germination and pollen tube growth was studied by Heslop-
84

Harrison (1983). According to him, the pollen tube growth terminates when pollen
falls on the pistil of the same genotype. Self-incompatible stigma rejects self-pollen
by inhibiting pollen hydration, germination and tube invasion into the stigma. These
processes have been reviewed extensively (Nasrallah, 2000; Silva and Goring, 2001;
Edlund et aI., 2004).
Most reliable test for pollen viability, stigma receptivity and seed setting is the
in vivo germination method. Aniline blue staining method is commonly used to study
in vivo germination. The fluorescence microscopic method has been used extensively
for tracing pollen tube growth and behaviour in pistil in a wide range of species (Kho
and Baer, 1968; Stout, 1972; Remming et aI., 1978; Vishnyakova.1991; Sniezko and
Winiarezyk, 1995). In H patens, 70-75% pollen germination was noticed on stigma
surface of open and self-pollinated flowers. But the pollen tubes never penetrated into
the stigma and they showed various abnormalities like curling of the tubes, swelling
of tube tip and irregular callus deposition followed by pollen tube growth inhibition. It
clearly indicates stigmatic surface recognition which leads to the inhibition of pollen
tube growth.
In general, the pollen tubes develop and deposit a considerable amount of
callose along their wall during elongation, which is a pre-programmed step in the
development of pollen tubes (Shivanna and lohri, 1985). In the germinating pollen of
angiosperms, callose plugs are formed in the pollen tube as it grows. According to
Hepler et al. (2001) callose plug formation helps to maintain a constant amount of
cytoplasm containing the germ units in the pollen tube tip region without division of
the tube cell, although it has been indicated that callose plug synthesis is not
dependent on the movement of germ units into the tube. The formation and function
of callose plug have been studied by Laitiainen et ai. (2002). Although callose was
85

originally believed to play a role in prevention of pollen tube penetration into the
stigmatic papilla, evidence indicated that callose is unlikely to play an essential part of
self recognition and rejection in other species (Singh and Paolillo, 1990; Franklin
et aI., 1995; Sulaman et aI., 1997; Radhamany, 2002). In H patens, incompatible
pollen tube growth was arrested on the surface of stigma and showed various
abnormalities like callose deposition at irregular intervals and swollen tube tips.
However, deposition of callose cannot be considered as the cause of pollen tube
inhibition as it is observed in the case of compatible pollination also.
Quite often, callose is synthesized in the cell as a response to some stress
conditions. According to Vishnyakova (1991), strong anomalous callose occlusions
occur in pollen grains and pollen tubes as a result of their incompatible interactions
with the pistil. Such non-programmed callose occlusions, as a rule, are linked with
metabolic changes. Hence in H patens, there was an incompatible interaction
between pollen tube and components on the surface of the stigma after self-pollination
and cause metabolic changes and irregular callose deposition, ultimately leading to
pollen tube growth inhibition.
According to Knox et al. (1976), the receptors on the stigma surface consist of
numerous components, some of which are involved in pollen germination and others
help in the entry of tubes. Previous reports suggested that phenolic compounds in the
surface deposits were involved in pollen tube growth inhibition (Martin, 1969; Martin
and Ruberte, 1972). Various enzymes were reported to play an important role in
pollen tube interaction on the stigma surface (Knox and Heslop-Harrison, 1970;
Heslop-Harrison and Heslop-Harrison, 1975a; de Nettancourt, 1977; Heslop-Harrison,
1977; Heslop-Harrison and Shivanna, 1977). Pollen- pistil interaction was reported to
be based on the interaction of protein of the stigmatic surface and the pollen wall
86

(Shivanna, 1978). The rejection response in stigma can be interpreted as an evidence
for a mechanism controlling normal tube growth of the pollen which is responding to
molecular signals from the pistil (Trognitz, 1995). According to Taylor and Hepler
(1997) pollen tube growth requires highly active and polarized secretion at the tube
apex. Hydrolysis of pectin in the stigma cell wall is also necessary for pollen tube
growth (Kim et aI., 1996; Wu et aI., 1996). According to Hiscock et ai. (2002a),
cutinase inhibitors significantly reduce the ability of pollen tube to penetrate the
stigma in Brassica. A pollen specific Brassica polygalacturonase have been detected
at the tip of the pollen tube as they enter the stigmatic papillar cell walls (Dearnaley
and Daggard, 2001). In H patens, as it has wet stigma, the compounds on the surface
deposits may playa role in the inhibition ofpollen tube growth.
5.2 Methods to overcome self-incompatibility
Various methods such as temperature treatment, bud pollination and excised
style pollination were employed to eliminate the incompatibility barriers in H patens.
Extreme conditions such as end-of season and high temperature were reported
to reduce incompatibility. The effect of temperature in incompatibility response was
studied by Lewis (1942) and Radhamany (2002). Leffel (1963) showed plants grown
at relatively high temperature produced more seeds after self-pollination than plants
of the same clones at low temperature. Temperature has been found to affect the
incompatibility response of Oenothera rhombipetala, 0. organensis, Lilium
longiflorum and Trifolium (Bali, 1963; Hecht, 1964; Ascher and Peloquin, 1970).
Townsend (1968) found that style exposed to 32°C for one or two days changed self
incompatibility in alsike clover (Trifolium hybridum ) to self-compatibility. Exposure
of the pistil above 34°C at the time of pollen germination caused decrease in the
87

germination percentage and the rate of pollen tube growth in this plant. Two species
of Primula as well as Prunus avium and Pyrus malus also showed retarding effect of
high temperature on t~ growth of incompatible pollen tube (Lewis, 1942;
Modlibowska, 1945). In H patens, treatment of style with extreme temperature was
tried. However exposure to 45, 50 and 55°C for 3 minutes caused reduction in the
pollen tube growth. Low temperature treatment at 20°C caused 65.9 % pollen
germination on the surface of stigma where as very low temperature (4°C) showed
only 8.2% pollen germination. In gametophytically controlled systems, overcoming
self-incompatibility by heat treatment seems to be caused by the denaturation of
metabolites (Hecht, 1960). Most probable molecules for the metabolite mediated heat
treatment are proteins including glycoprotein. The absence of fruit-set at high
temperature is not considered as outcome of a single malfunctioning factor, but
triggering a series of factors simultaneously impaired by high temperature. Lower
temperature (12°C) and higher humidity (90%) positively affected bud pollination, in
Brassica oleracea L. var. capitata (Zur et aI., 2003).
Excised style culture technique might be helpful not only in studying the
interaction between style and pollen tubes during the tube growth but also in detecting
the number of tubes at a distance from the stigma. The application of pollen to the cut
surface of the style after removing the stigma in parents with strong incompatibility
was effective in overcoming cross-incompatibility (Gardella, 1950). Stigma and style
grafts have been employed to overcome incompatibility barriers in Petunia and
Oenothera (Hecht, 1960, 1964; Herrero and Dickinson, 1979). Style shortening and
stigma regrafting have been successfully applied in Oenothera (Gardella, 1950).
Pollination on decapitated stigma has been used successfully in Solanum
(Swaminathan, 1955). In the present study, application of the pollen to the cut surface
88

of the stigma and style has been found not successful to overcome incompatibility
barriers.
Eventhough many techniques have been standardized to overcome self-·
incompatibility, bud pollination and delayed pollinations were found to be the
simplest and most commonly used methods (Shivanna, 1982; Shivanna and Johri,
1985). It could be utilized in species with gametophytic as well as sporophytic
incompatibility. Bud pollination was found to be very successful in Brassica,
Raphanus and Brunelsia (Haruta, 1966; Radhamany, 2002). In 1964, Linskens
suggested that the principle of self-compatibility following bud pollination is based on
the fact that the incompatibility substances are absent or are not fully effective in
developing styles. According to Shivanna (1978), bud self-compatibility is due to the
stigma factors inhibiting the pollen in incompatible combinations which are not
present or are inactive in buds of the age accepting self pollen. In the present study
bud pollination was not successful.
5.3 Molecular characterization by RAPD
Random amplified polymorphic DNA markers have been used to analyze the
genetic diversity at low taxonomic levels (Mehrnia et aI., 2005; Rajasekar et aI., 2006;
Das et aI., 2007). RAPD is simple and easy to perform and can be applied to a large
number of genotypes without adding much to the cost of experiment (Srivastava et al.,
2007). It has been successfully employed in the evaluation of genetic relationship in
various plant species (Gunter et al.,1996; Gielis et aI., 1997; Nair et aI., 1999; Nayak
et aI., 2003; Rout, 2006)
The self-incompatibility recognition systems of angiosperms are a classical
object of interest to geneticists interested in the spectacular polymorphism found in
89

natural populations, and cell biologists interested in recognition functions. The
maintenance of multiallelic polymorphism has been thoroughly investigated by
classical evolutionary and population genetic approaches (Lawrence and Franklin
Tong, 1994; Lawrence et aI., 1994; Vekemans and Slatkin, 1994; Schierup, 1998). In
all the species studied with homomorphic SI systems, (gametophytic as well as
sporophytic) the S-loci have spectacular numbers of S-allele within the populations
(Lane and Lawrence, 1993). It has long been understood that, if two different loci are
involved, the self-incompatibility genes must recombine rarely or not at all, since
recombination would generate genotype having the pollen type of one allele with the
pistil specificity of a different allele, and these would presumably be self-compatible.
Outcrossing species tend to have higher levels of variability within populations but
smaller degrees of differentiation among populations than selfing species (Hamrick
and Godt, 1990; Schoen and Brown, 1991).
Recently, biochemical and molecular tools have determined the S-genotype of
cultivars in various species. PCR based S-allele typing system could be useful for
determining pollen compatibility groups of commercially important new cultivars to
elucidate their incompatibility relationships. The molecular typing system of
genotypes based on PCR is useful and has been used as a rapid method for indicating
new S-allele and incompatibility groups in sweet cherry in the absence of pollination
tests (Choi et aI., 2002).
For RAPD marker analysis, 14 different accessions of H patens collected
from various parts of Kerala have been used. Field observations showed that
morphological characters are quiet consistent within 13 accessions, but accession
No. 3 shared some unique morphological features and also form compatible
combination when used as either male or female parent. For the RAPD analysis,
90

10 among 20 pnmers used were selected from the clear, reproducible, and
polymorphic banding profile. Genotypic relationship based on RAPD phenotypes of
the 14 accessions were analysed and constructed a UPGMA phenogram. The topology
of the phenogram shows expected grouping with relation to the morphological and
compatibility difference found among the accessions.
RAPD and sdf-incompatibility (SI) typing techniques were applied to find out
the genetic integrity of the two varieties of Raphanus sativus (Kwak et aI., 2009).
Bulk segregant analysis (BSA) and RAPD were employed to identify molecular
markers linked to a self-incompatibility gene in self-incompatible and self-compatible
near-isogenic lines of non-heading chinese cabbage (Brassica campestris L. ssp.
chinensis Makino var. communis), (Shi and Hou, 2004). PCR analysis with primers
designed on the conserved sequences of sweet cherry S-RNases has been used to
characterize the S-genotype of 71 sweet cherry cultivars, including 26 cultivars whose
S-allele constitution had not been previously described (Wunsch and Hormaza, 2004).
A total of 17 pollen incompatibility groups in sweet cherry (Prunus avium L.) were
identified among 46 accessions by PCR based S-allele typing analysis and by
controlled test pollinations (Choi et aI., 2002). cDNA sequences corresponding to
five self-incompatibility alleles (S-alleles) of the apple cv 'Golden Delicious' have
been reported by Janssens et aI. (1995). Based on the nucleotide sequences of these
S-allele cDNAs, a molecular technique has been developed for the diagnosis of the
five different S-alleles in apple cultivars.
Tamura et aI. (2000) developed a method for identifying S-RNase using the
cDNA sequence, a PCR-based identification system using genomic DNA. PCR
amplification of genomic DNA followed by restriction analysis is used for the
identification of S alleles involved in self-incompatibility in Brassica oleracea (Brace
91

et aI., 1993). In H patens, RAPD analysis of 14 accessions revealed two different
groups with variation in the specific bands, clearly evident that allelic variation may
exist in Acc. No.3. In the present analysis Acc. No.3 is distinctly seperated from the
rest. It has same reproductive significance in the sense that it is compatible when used
as either male or female parent. So it can be concluded that Acc. No. 3 with this
reproductive significance may have a different genotype.
5.4 Infraspecific pollination
As various methods used have failed to overcome incompatibility in
H patens, infraspecific pollinations carried out between Acc. No.1 and 3 were found
to be successful and resulted in viable seed. The production of viable seeds may be
considered as the ultimate criterion for reproductive success (Wiens et aI., 1987).
In the case of cross-pollinations, large number of pollen grains adhered and
germinated on the surface of the stigma. These germinated pollen tubes penetrated
into the style and reached the ovary and effected fertilization. This was confirmed by
pollen-tube behaviour and by the developmental anatomy of ovary after pollination.
Field pollination of H patens showed significant differences in ovule development
between self and cross-pollinations. A significantly greater proportion of megaspores
in the ovules failed to develop after self-pollination. Developmental anatomy of ovary
after cross pollination showed normal cell differentiation. To decide whether certain
cross combination is compatible or not, fruit set and combinations of fruit and seed
set have been used as indicators (Cipar et aI., 1964; Grun and Aubertin, 1966;
Abdalla, 1970; Olsder and Hernisen, 1976).
In the present study, significant difference was noticed in the pollen
germination and tube growth pattern during self and cross- pollinations. So it is
92
assumed that self-pollination induced biochemical changes in the stigma/style tissues
that may affect the metabolic status of the tissue, acts as a reproductive barrier in
H patens. However, cross- pollination did not affect the metabolic activity and
normal functioning of the stigma/style tissues which in turn lead to compatible
pollination. In H patens, infraspecific pollination was found to be effective in
producing seeds. Continued development of ovary after compatible and incompatible
pollination also clearly indicated the success of infraspecific pollination in H patens.
5.5 Histochemistry and enzyme cytochemistry
Pollination initiates many biochemical changes in the pistil. In order to find
out the pollination induced changes in the primary metabolites and key enzymes in
the pistil of H patens, histochemical and cytochemical localizations were done and
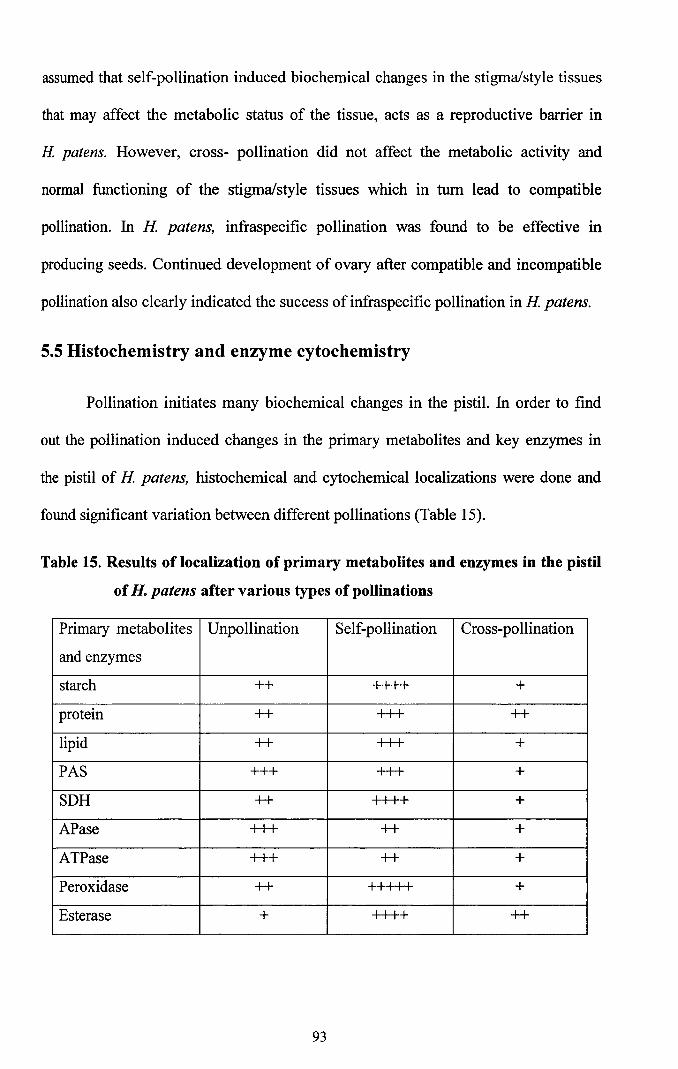
found significant variation between different pollinations (Table 15).
Table 15. Results of localization of primary metabolites and enzymes in the pistil
of H. patens after various types of pollinations
Primary metabolites Unpollination Self-pollination Cross-pollination
and enzymes
starch ++ ++++ +
protein ++ +++ ++
lipid ++ +++ +
PAS +++ +++ +
SDH ++ ++++ +
APase +++ ++ +
ATPase +++ ++ +
Peroxidase ++ I I I I I +
Esterase + ++++ ++
93

Accumulation of insoluble starch and polysaccharide was indicated as
reserves, which acts as the main source of energy for cellular metabolism. In
unpollinated pistils, meagre deposition of starch was noticed. In self-pollinated pistils
of H. patens, intense deposition of starch has been found at secretory zone as well as
in ground tissues. Starch is an energy reserve, and its level decrease when the pollen
tube growth is heterotrophic. In Petunia hybrida, the starch level in the transmitting
tissue drops heavily after pollination (Herrero and Dickinson, 1979). In the normal
course of compatible pollination, carbohydrates produced in the pistil are utilized by
pollen tubes for their growth. Activity of polysaccharides and starch in pollinated
stigma is higher than the unpollinated ones, which indicates that these reserves are
accumulated during pollination and subsequently mobilized for meeting the energy
requirement. As a result, there is reduction in the amount of carbohydrate in the pistil
with an increase in age (Yamada, 1965; Rosen and Thomas, 1970). In H. patens,
intense deposition of starch was seen after self-pollination and it may be due to
immobilizition of starch. Since the pollen tube growth was inhibited on the surface of
the stigma, the reserved food materials may not be used for tube growth and thus their
accumulation. In compatible pollination, starch was utilized for normal pollen tube
growth and therefore transmitting tissue was found to be free of its deposition.
Presence of protein was identified by using mercuric-bromophenol blue.
Protein deposition was less in unpollinated stigma, whereas meagre deposition was
seen in papillar region, ground tissues and transmitting tissues. Proteins playa major
role in the recognition and rejection phenomena on the stigma surface (Nasarallah and
Wallace, 1967). Mattsson et al. (1974) suggested that the proteinaceous pellicle
overlaying the stigmatic papillae may contain the molecules responsible for
recognition of pollen grains. Numerous investigations have demonstrated that proteins
94

(Arabinogalactan-protein) are involved in the process of sexual reproduction of
angiosperms and play multiple roles, by providing surface adhesion, nutrients and
directional cues or signals for pollen tubes (Sommer-Knudsen et aI., 1997; Cheung
and WU,1999; Majewska-Sawka and Nothnagel, 2000; Qin and Zhao, 2004; Qin
et aI., 2007) . They may also participate in gametal recognition, embryo development
and differentiation (Hu et aI., 2006). In H patens, the protein component present on
the stigmatic surface may have a role in pollen adhesion. After pollination, proteins
are localized on the transmitting zone and ground tissue which indicated the role of
proteins in directional signal for pollen tubes.
Lipids are essential factors needed for pollen tube to penetrate the stigma and
directing pollen-tube growth by controlling the flow of water to pollen in species with
wet stigmas, while pollen coat is functionally equivalent to the species with dry
stigma (Wolters-Arts et aI., 1998). Lipids probably regulate the availability of water
to the pollen, and prevent desiccation of stigma (Martin, 1970). The greasy exudates
of the stigma included chiefly lipid compounds which can be separated into eight
principal fatty acids (Konar and Linskens, 1966b). Lipids also playa major role in the
nutrition of the pollen grains (Kuruvila and Shah, 1988; Wolters-Arts et aI., 1998). In
the present study, intense accumulation of lipids was found in the extreme tip of the
stigma and on either sides of vascular tissues. In the stigma of Withania somnifera,
stigmatic exudate is rich in lipids (Philip, 1989). Vasil and JoOO (1964) found that in
Aegle and Pavonia, the stigma secretion showed the maximum concentration of lipids
at the time of pollination. In the present study, presence of lipid was mainly localized
in the region where stigmatic exudate is seen. The deposition of lipids was found to be
maximum on the day of anthesis and reduced after pollination. So it is assumed that
95

lipid is a major components of the exudate and plays a key role in the adhesion and
hydration ofpollen grains.
In H patens, self- and unpollinated stigmas stained for PAS showed uniform
distribution of PAS positive bodies. When compared to the self- pollinated stigma,
meagre activity was observed in cross-pollinated pistils. In this case, activity of PAS
was found to be less in the secretory zone and transmitting region. PAS existed in
several forms, viz, soluble sugars, insoluble cell wall, polysaccharides (cellulose,
pectin etc) and even storage starch polysaccharides contribute to the cellular
energetics and to the production of energy rich triphosphates. Polysaccharides are
necessary for pollen tube growth (Ciampolini and Cresti, 1998). During this process
of pollen-pistil interaction, pistil plays an essential role in the production of
compounds necessary for recognition, guidance, protection and nourishment of the
pollen tubes (Kao and Mc Cubbin, 1996). The presence of storage starch indicated
potential energy which would be utilized during different growth and differentiation
processes in plants. In H patens, the reserve food materials were not utilized for
pollen tube growth in self-pollination. Hence PAS was accumulated on the surface of
stigma.
Enzyme cytochemistry has developed with great speed and is now being
applied to many diverse areas of plant biology. There were a few studies concerning
changes in the activities of enzymes in stigma-style tissues following compatible and
incompatible pollination (Bredemeijer and Blaas, 1975; Dhaliwal and Malik, 1985).
Most of these were conducted in species characterized by gametophytic self
incompatibility (Schlosser, 1961; Linskens et aI., 1969; Bredemeijer, 1974, 1979).
Since compatible and incompatible pollinations are affected by alterations in the
metabolic status of stigma and stylar tissues, it would be imperative to study the
96

changes in the enzyme activities of stigma and stylar tissues following compatible and
incompatible pollinations (Dhaliwal and Malik, 1985). Since the enzymes control
biochemical reactions, and their synthesis is controlled by specific gene(s), any
change in the activity of an enzyme would reflect in the pattern of gene expression
and corresponding metabolic events in the cell. Hence, the enzymes can be used as
tools to study the problem of self-incompatibility at the biochemical level. In the
present investigation, changes in the activities of peroxidase, esterase, ATPase, APase
and SDH were studied.
The activity of succinic dehydrogenase is very intense in the papillar region
and vascular tissue after self-pollination. Before pollination, the activity of this
enzyme was meagre. SDH has been associated with mitochondria and was involved in
transfer of electrons to oxygen. This enzyme was also found to be involved in the
process of cellular respiration leading to ATP synthesis (Pearse, 1972; Malik, 1977).
The activity of this enzyme increased with a corresponding increase in the age of
stigma, which shows that the rate of metabolism was high in the stigma matrix. In
Catharanthus roseus and Withania somnifera, activity of SDH was moderate in
younger buds, and in mature buds intense activity was noticed (Philip, 1989). Intense
activity of this enzyme is also noticed in the vascular tissue regions of plants (Shah et
aI., 1980). The activity of SDH is more intense in the sub epidermal regions and
stigmatic papillae of Brunfelsia undulata (Radhamany, 2002). From the present study
it is assumed that pollination may induce excess ATP production, which may lead to
an increase in SDH activity in H patens after pollination.
Among the hydrolases, the most pronounced activity was that of acid
phosphstase. The hydrolytic enzymes are lysosomal in nature (Gahan and Mc Lean,
1969). These enzymes have a role in hydrolyzing the stored reserves like protein,
97

sugar and starch. The products of hydrolysis of these reserves are usually utilized for
the metabolic activities of the cell (Chaffey and Harris, 1985). Acid phosphatase
(APase) are limited with the differentiation and maturity of stigma. According to
Simola and Sopanen (1970) APase was connected with lipid synthesis. In H patens,
papillar, epidennal, sub epidennal and upper few layers of ground tissue showed
intense activity. The central core of transmitting tissue was devoid of its activity.
APase activity was maximum in unpollinated stigma and after pollination stigma
showed slight decrease in the activity. In H patens, reduced activity of APase is
related to the reduction in the lipid synthesis as well as sugar transfonnation as a
result of pollen tube growth inhibition on the surface of stigma.
ATPase is an enzyme primarily involved in the breakdown of ATP for the
release of high energy present on the phosphate bond of the molecule required for the
various metabolic and developmental activities in plants (Gahan, 1981). In H patens,
intense activity of ATPase was seen in the papillar region which is evidenced by the
lead sulfide deposition, depicting its activity, and was confined to the cell membrane/
wall regions. The activity of this enzyme was restricted to the cytoplasm and the
reaction was diffuse. The cytoplasmic activity may indicate its role in the metabolic
process in tenns of high protein synthesis, while wall bound activity signifies its role
in the intercellular communication and transport required for the cellular
differentiation and subsequent development. In Brassica campestris, ATPase was
differentially distributed in a zonate pattern in the vegetative apices, showed a
heterogeneous localization with the highest activity in the central zone and the
meristematic zone. At the early transition stage of development, ATPase activity in
the peripheral zone increased slightly and at the transitional stage it was not localized
within the peripheral zone. In the present study, ATPase activity was less in pollinated
98

pistils. This observation agrees with Gahan (1984), and that high activity of this
enzyme during morphogenesis could be the indication of cell differentiation. In the
present study, pollen tube growth inhibition on the stigmatic surface causes reduction
in the activity of enzyme. So the activity of ATPase also reduced after self
pollination.
Angiosperm stigmas have long been known to exhibit high levels of
peroxidase activity when they are mature and most receptive to pollen but the
biological function within the stigma is not known. Bredemeijer and Blaas (1980)
reported that pollination and pollen tube growth caused an increase in total peroxidase
activity. Peroxidases generally catalyse the breakdown of hydrogen peroxide to yield
highly oxidizing intermediates, which oxidize a variety of organic and inorganic
reducing substrates. Understanding peroxidase function in plants has proved
problematic because of their broad range of substrate preferences and lack of tissue
specificity. Peroxidases should now be considered as potentially important
components of signal transduction pathways in plants. Early isozyme studies in
Nicotiana suggested a potential role in pollination processes, particularly SI (Pandey,
1967b). These studies showed that stylar peroxidase activity was higher after
incompatible pollinations compared with compatible pollinations. Unpollinated
pistils, however, showed stylar peroxidase activities comparable to those of selfed
(incompatible) pistils, and hence Bredemeijer (1974) inferred that in the compatible
style, peroxidase activity was somehow inactivated. Closer inspection indicated that
only the peroxidase activity associated with the extra cellular secretion of the stylar
transmitting tissues were reduced following compatible pollinations (Bredemeijer and
Blaas, 1975). Later Carraro et al. (1986) suggested that peroxidase is responsible for
the inhibition of incompatible pollen in Petunia and Nicotiana. However, in
99

Nicotiana, Lycopersicon and Petunia a peroxidase gene is linked to each of their
respective S loci (Hoopen et aI., 1998), but these S-linked peroxidase loci have never
been characterized. The first plant peroxidase gene expressed exclusively in stigmas
was recently identified (McInnis et aI., 2005). Importantly, this peroxidase, SSP
(stigma-specific peroxidase), is expressed specifically in the specialized secretory
cells (papillae) of the stigma epidermis and is developmentally regulated. Expression
of SSP in flower buds is developmentally regulated, with maximum level of
expression coinciding with anthesis; when stigmas are most receptive to pollen and
expressed self-incompatibility (Bredemeijer, 1974). Increase in peroxidase content of
the transmitting tissue following self-pollination was reported by many authors
(Bredemeijer and Blaas, 1975; Bredemeijer, 1976). The results indicated the presence
of peroxidases on the cell wall of unpollinated and self-pollinated styles support the
hypothesis of Bredemeijer (1977) that only the fraction of extracellular peroxidases is
responsible for pollen tube growth inhibition. Compatible pollination does not cause
the increase in the number of cells showing peroxidase activity, but does cause the
disappearance of the peroxidase activity observed in unpollinated samples. According
to Galen and Plowright (1987) in Pedicularis canadensis and Clintonia borealis the
stigma peroxidase activity is a reliable indicator of receptivity. In H patens, self
pollination causes an increase in the activity of peroxidase in the pistil and reduced
activity during cross-pollination, which support the hypothesis that only a fraction of
extra cellular peroxidase is responsible for pollen tube inhibition.
Studies on the esterases have utilized the high enzymatic activity in stigma as
indicators to assess the stigmatic receptivity (Lavithis and Bhalla 1995: Tandon et aI.,
2001). Bhattacharya and MandaI (1997, 2003) correlated the esterase activity and
stigma receptivity. The wet type stigma secretes exudates containing lipids, phenolic
100

compounds, carbohydrates, proteins, phosphatases, lectins and aminoacids including
esterases (Baker et aI., 1974; Vasil, 1974). According to Chaffey and Haris (1985)
this enzyme hydrolyse the stored reserves like starch, protein and sugars thereby
initiating them for the metabolism of cells in morphogenetic events. In H patens, the
activity of esterase increases after self-pollination so as to facilitate the growing
pollen tube. But pollen tube growth inhibition on the stigmatic surface may cause
accumulation of this compound.
In H patens, pollination induced many metabolic changes in stigma; most of
these changes are related to pollen tube growth. Many of the metabolites were
accumulated after self-pollination because pollen tube growth was inhibited on the
surface of the stigma.
5.6 Electrophoresis
Although the functional role of S-proteins in the self-incompatibility
interactions has not been demonstrated in vivo, many lines of evidences strongly
suggested their involvement in the recognition and rejection of self-pollen. The
association between S-glycoproteins and self-incompatibility was further supported
by Herrero and Dickinson (1980) in Petunia by locating these proteins at the site of
pollen tube arrest and their increasing expression during development of flower. Since
infraspecific pollination has been found to be successful in H patens, the protein
composition of the stigma homogenate and leachate of unpollinated, self- and cross-
pollinated (compatible) flowers have been analyzed electrophoretically and revealed
the presence of specific protein bands. Between self- and cross-pollinated pistil,
protein bands were found to be common with in arrange of 22 to 95 kDa but showed
quantitative difference.
101

Protein profile of stigma leachate showed five bands at 23, 30, 36, 70 and 99
kDa regions. These bands were also found in the protein profile of stigma
homogenate. As like in the stigma homogenate, protein profile of leachate showed
quantitative difference in bands between self- and cross-pollinated pistils, and band at
36 kDa region was absent in unpollinated stigma leachate. Protein band at 36 kDa
region has been found to be present in stigma leachate as well as homogenate while it
is absent in unpollinated pistil. In H patens tube growth inhibition of self-pollen takes
place on the surface of the stigma as well as in the in vitro assay medium. So the
factor(s) responsible for incompatible interaction is present in leachate as well as
homogenate after self-pollination. So it is assumed that protein band at 36 kDa may
be responsible for incompatible interaction in H patens.
Proteins and glycoproteins involved in the manifestation of self
incompatibility are proven to be associated with particular S-alleles (Nasrallah et
al.,1985b; Anderson et al.,1986; Cornish et aI., 1987, 1988). Immunological and
electrophoretic data indicated that S- allele specific protein present in the stigma of
Brassica caused strong self-incompatibility (Nasrallah and Wallace, 1967; Nasrallah
et aI., 1970; Sedgley, 1974). The protein composition of the stigma secretion and
stigma cell extracts of different species of plants have been analyzed
electrophoretically with or without SDS (Miki-Hiroshige et aI., 1987; Reger, 1989). In
Lycopersicon peruvianum ,the SI associated proteins have been shown to exhibit a
high degree of polymorphism in the molecular weight range of23-30 kDa (Bematzky,
1993; Lee et aI., 1994). McCubbin et al. (1997) obtained several transgenic plants of
Petunia with an intermediate SI system; their styles produce much less S-protein,
whereas self-compatible style lacks S-proteins. S-allele associated proteins with
RNase activity was detected from the Japanese pears (Sassa et aI., 1992; 1993;
102

Hiratsuka et aI., 1995; Ishimizu et aI., 1996). According to Zhang and Hiratsuka
(1999), S-proteins in the style may be one of the determinants of SI in Japanese pear
cultivar.
Bredemeijer and Blaas (1981) was the first to establish the correlation between
the genetic phenomenon of S-allele-specific pollen rejection and specific protein
products in the style. Anderson et ai. (1986) cloned the first S-protein in a
gametophytic SI system, Similar S-proteins were also cloned in Nicotiana,
Lycopersicon, Petunia and Solanum and found a basic glycoprotein of about 30 kDa
was extremely abundant in the pollen transmitting tract matrix (Anderson et aI., 1989;
Ai et aI., 1990,1992; Clark et aI., 1990; Kheyr-Pour et aI., 1990; Xu et aI., 1990; Tsai
et aI., 1992).
In buck wheat, protein profile of stigmatic leachate showed quantitative
difference at 56 kDa region in incompatible and compatible pistil (Miljus-Djukic
et aI., 2004). From the previous reports and present study, it is assumed that a protein
band at 36 kDa region may be involved in the self-incompatibility mechanism in
Hpatens.
5.7 S-RNase activity
The most phylogenetically widespread gametophytic incompatibility relies on
ribonucleases called S-RNases (Igic and Kohn, 2001). S-RNases are basic
glycoproteins of about 30 kDa that are secreted into the extra cellular matrix of the
stigma, transmitting tract, and the inner epidermis of the ovary (Anderson et al,
1986,1989; Cornish et al., 1987; McClure et aI., 1993). According to Jahnen et ai.
(1989), S-allele specific pollen rejection requires extremely high level of S- RNase in
the extracellular matrix. It has also been shown that the S-RNase activity is required
103

for pollen rejection (Huang et aI., 1994; Karunanandaa et aI., 1994). Thus, S-RNases
are thought to function as highly specific cytotoxin that inhibits the growth of
incompatible pollen.
Hamelia patens showed many of the characters of gametophytic
incompatibility system. After self-pollination, pollen tube growth inhibition takes
place on the surface of the stigma and electrophoresis of stigma protein from
incompatible and compatible stigma found variation in the amount of protein at
36 kDa region. In self-pollinated stigma, the intensity of specific protein band was
found more. To determine activity of RNase, diffusion plate assay was done and
found its activity on the surface of the stigma. When compared, cross-pollinated
stigma showed less activity than self-pollinated one, where as in the style or the cut
end of the style lack RNase activity.
RNase activity has been reported III S-proteins extracted from styles of
Nicotiana a/ata and Pyrus serotina (McClure et ai. 1989; Gray et ai. 1991; Sassa et
al., 1992). In P.serotina, significantly more RNase activity was found in styles of self
incompatible varieties than in those of a self-compatible mutant. In N a/ata, the
S-RNase, which represented a high percentage of the stylar proteins, caused
degradation of self-pollen mRNA and rRNA, leading to the inhibition of pollen tube
growth (McClure et aI., 1989). Proteins with sequences similar to those of S-RNase
have been found in styles of self-compatible Arabidopsis thaliana (Taylor et al.,
1991) and Petunia hybrida (Clark et aI., 1990) may be involved in a wide range of
cell-cell recognition processes. Gray et ai. (1991) used gold-immunolabeling to show
that stylar RNase penetrates into self-pollen tubes. In self-compatible Lycopersicon,
absence or decreased S-RNase activity was accompanied by loss of production of the
stylar protein HT (Kondo et aI., 2002b), which is necessary for a functional SI
104

response (McClure et aI., 1999). Low stylar RNase production was reported to be
associated with SC in both Nicotiana (Golz et aI., 1998) and Lycopersicon (Kowyama
et al., 1994; Royo et aI., 1994; Kondo et aI., 2002a). Less activity of RNase in
compatible pistil (cross) may be the reason for compatible interaction in H patens.
In H patens, activity of RNase gradually increases in the stigma from five
days before anthesis to the day of anthesis. Eventhough the RNase activity in stigma
five days before anthesis was very low, the buds are not compatible with mature
pollen. In Witheringia solanacea, buds 2 days prior to anthesis produced stylar
RNase at levels equivalent to that of mature flowers, yet bud pollinations still had
reduced SI, (Jahnen et aI., 1989; McClure et aI., 1989). The electrophoretic analysis of
S-RNase proteins of W solanacea showed a marked increase between small and large
buds rather than between large buds and open flowers.
The selective inhibitory action of stigma leachate on the growth of self and
cross pollen tube in vitro and the indispensability of RNase activity in inhibition of
pollen tube growth were studied by in vitro assay. In H patens, stigma leachate
selectively inhibits growth of incompatible pollen than cross-pollen. When pollen
germination was done in stigmatic leachate alone or in Brewbakers medium
supplemented with stigmatic leachate, pollen germination and tube growth of self
pollen were inhibited when compared to the control. In the case of cross pollen,
germination and tube growth were not significantly inhibited. Germination of
incompatible pollen was inhibited by the stigma leachate whereas that of compatible
pollen was not affected by leachate. RNase isolated from self-incompatible apple was
also found to inhibit pollen tube growth in vitro (Katoh et aI., 2002). In Nicotiana
alata, the S-protein was also cloned and sequenced and was found to share homology
with fungal RNase. The isolated S-protein showed RNase activity and inhibitory
105

activity when added to the in vitro pollen germination medium (McClure et al., 1989;
Haring et aI., 1990; Gray et aI., 1991).
To clarify the relationship between RNase activity and pollen growth
inhibition, Rnase A from Bovine serum was added instead of stigma leachate. When
Bovine serum was added alone or in combination with stigma leachate to the culture
medium, significant reduction in the pollen germination and tube growth was
observed in the case of self-pollen. But presence of RNase A did not affect the
germination and growth of cross-pollen. From the present study it may also be
concluded that the stigma leachate has inhibitory activity similar to that of RNase A.
According to McCubbin and Kao (2000), if the S-RNase arrests the pollen
tube growth of self-pollen in the pistil as well as in the in vitro assay, the S-RNase
should be modified or controlled by any molecules in the cross-pollen tube to
inactivate the inhibitory activity. One hypothesis is that the S-RNase is taken up by
the incompatible pollen tube as it grows through the extracellular matrix. Once taken
into the pollen tube, rRNA genes are not transcribed into the pollen, resulting in
pollen tube death (Mascarenhas, 1990, 1993; McClure et aI., 1990). In this case, the
specificity of the interaction would be controlled either by the uptake of the S-RNase
into the pollen tube or by the presence of an inhibitor that would act in an allele
specific manner to prevent RNA degradation in compatible pollen tube.
To examine the role of RNase activity in pollen tube inhibition, chemicals
such as zinc sulfate and cupric sulfate, having the property to inactivate stigmatic
RNase, were incorporated in the in vitro pollen germination medium. When these
RNase inhibitors were added to the medium, the rate of pollen germination and tube
length of self pollen significantly increases. In H patens, ZnS04 dramatically reduced
106

the inhibitory activity of RNase on pollen tube growth whereas CUS04 cause further
disturbed the pollen tube gennination. The inhibitory activity of ZnS04 is depended
on its concentration. One mM ZnS04 showed maximum inhibition of RNase activity
and high rate of self pollen gennination. Above this concentration, ZnS04 disturbed
the germination and growth of pollen. According to Huang et al. (1994) RNase
activity of stigmatic protein is essential for their function in rejecting self-pollen. The
result may suggest that stigma leachate of H patens is having RNase activity and it is
essential for inhibition of self-pollen gennination and tube growth. In cross-pollen the
RNase may modify or less activity of RNase may leads to compatibility. In the
present study, the inhibition of S-RNases by ZnS04 affect the ability to reject self
pollen in H patens.
RNase activity III the stigma of H patens is developmentally regulated.
During anthesis, the stigma actively secretes RNase, which inhibits pollen
germination and pollen tube growth in vitro and in vivo. The ability of RNA, as a
competitive inhibitor, to reduce the effect of stigmatic diffusate on pollen tube growth
further indicate that the inhibition is derived directly from the stigmatic RNase.
Analyses concluded that strong RNase based gametophytic incompatibility system is
operating in H patens. Due to strong incompatibility sexual reproduction is beset and
seed-set is absent in H patens.
107