Embed Size (px)
Citation preview

A O S 2000
3
Dissection and cotransplantationof large pieces of RPE and neuralretina; effect of protease K on thedevelopmentRajesh Kumar Sharma
Department of Ophthalmology, University Hospital of Lund, Lund, Sweden
ABSTRACT.Purpose: This study attempts to cotransplant large pieces of the RPE and neu-ral retina in the subretinal space of rabbits by using protease K for dissectionof the donor tissue, and to investigate the effect of dissection technique on thedevelopment of the grafts.Methods: Eyes from 15-day-old pigmented rabbit embryos were partly digestedby protease K to assist dissection of sclera and the choroid from RPE andneural retina. Large pieces of RPE and the neural retina thus obtained werecotransplanted into the eyes of adult albino rabbits who were allowed to survivefor up to 63 days. The transplants were examined under light microscope.Results: It was possible to transplant large sheets of RPE and neural retinatogether. Both the RPE and the neural retina survived after cotransplantation.Retinal pigment epithelium survived in layers, but at places formed clusters. Incotransplants neural retina formed rosettes, developed gliosis, and photorecep-tors failed to develop outer segments, possibly due to the action of protease K.Conclusion: Proteases seem to be injurious for the development of the neuralretina.
Key words: cotransplantation – neural retina – retinal pigment epithelium – retinal transplan-tation – retinal degeneration – protease K.
Acta Ophthalmol. Scand. 2000: 78: 3–8Copyright c Acta Ophthalmol Scand 2000. ISSN 1395-3907
Retinal degenerations have thus farbeen considered untreatable. In the
last few years much attention has beenpaid to research aimed at improving themanagement of these disorders (Shar-ma & Ehinger 1999). Retinal transplan-tation is an emerging concept for thetreatment of retinal degeneration. Trans-plantation of both the retinal pigmentepithelium (RPE) and the neural retinaare being separately developed (Algvereet al. 1994; Kaplan et al. 1997; Sharmaet al. 1995).
Transplantation of the RPE cells hasbeen shown to rescue the photorecep-tors from degeneration in Royal Collegeof Surgeon (RCS) rats (Lopez et al.1989; Li & Turner 1988). The photore-
ceptors in this strain of rats degeneratedue to an inherited defect in the RPE(LaVail et al. 1974; Young & Bok 1969).Freshly harvested neonatal donor cellshave a long-term rescuing effect (Sheed-lo et al. 1993, 1992). Another approachto treat retinal degeneration is to recon-struct the degenerated neural retina bytransplanting retinal neurons. Neuralretina can be transplanted in variousphysical forms such as suspension, layerof photoreceptors, fragmented pieces orlarge sheets of whole retinas (Sharma etal. 1997; Silverman & Hughes 1989;Gouras et al. 1992; Turner & Blair1986; Kondo et al. 1984). After trans-plantation of the embryonic retina, thegrafts proliferate, survive, and differen-
tiate (Sharma 1999a, 1999b, 1999c;Sharma & Ehinger 1997a; Bergstrom etal. 1992). The RPE is important for thedevelopment of the photoreceptor cells(Layer & Willbold l993; Hollyfield &Witkovsky 1974). It has a favorable ef-fect on the development of the retinalneurons; especially the development ofthe outer segments in co-cultures ofRPE and neuroretinal explants (Spar-row et al. 1990; Caffe et al. 1989).Moreover, certain clinical situationsmay demand replacement of both RPEand neural retina. Neural retina andRPE have been cotransplanted earlier infragmented form (Seiler et al. 1995), buttransplantation of large pieces of neuralretina has certain advantages in termsof lamination (Ghosh et al. 1998; Shar-ma et al. 1997). It is therefore desirableto evolve techniques for co-trans-planting large sheets of RPE and theneural retinas together. In rabbits it isdifficult to dissect the RPE and the neu-ral retina together without causingmechanical damage to the donor tissue.Even harvesting sheets of RPE are as-sociated with technical difficulties suchas damage to the cells and secondarycontractions, and therefore efforts arebeing made to develop techniques toharvest the RPE from donor eyes fortransplantation (Tezel et al. 1997; Ho etal. 1997). Knowledge regarding the ad-vantages and disadvantages of differenttechniques is important in the searchfor an ideal method for dissection. Inthis study the donor tissue was obtainedby using protease K. Feasibility oftransplanting large sheets of RPE andneural retina thus obtained, and the ef-fect of the dissection technique on thedevelopment of the grafts was studied.

A O S 2000
4
Materials and MethodsTransplantation
Ordinary mixed strain pigmented rabbitsfrom embryonic day (E) 15 were used asdonors. Embryos were obtained by Caes-arean section after sacrificing the preg-nant female rabbits. They were kept atπ4æC in Ames’ solution having the fol-lowing composition (mM): NaCl 120,KCl 3.6, MgS04 1.2, CaCl2 1.2, NaHCO3
23, NaH2PO4 0.1, Na2HPO4 0.4 and glu-cose 10. Eyes from these embryos wereenucleated and treated with 0.5% pro-tease K (SigmaA P-656; 19 units/mg ofprotein) for 210 seconds at π4æC. In or-der to have maximum effect of the en-zyme on the sclera and minimum on theretina, intact eyes were treated with theprotease without dissecting apart the an-terior and the posterior segments. Theenzyme action was stopped by rinsing theeyes with Ames’ solution several times. Inorder to optimize the enzyme action, sev-eral pilot experiments were performed inwhich the time of action of the enzymewas varied. Protease greatly facilitatedthe dissection of the sclera and the uveawithout disturbing the retinal pigmentepithelium. Under a dissecting micro-scope the sclera and the choroid were sep-arated from the inner coats of the eyecontaining the RPE and the neural ret-ina. Finally, the anterior segment of theeye was dissected out, giving a cupshapedpiece of RPE with adhering neuroretina.These retinas were kept in the Ames’solution at π4æC until transplanted (al-ways less then 3 hours from dissection).
A special instrument was used for co-transplanting the pigmented epitheliumand the neuroretina. The details of the in-strument have been published earlier(Sharma et al. 1997). In brief, the instru-ment comprised an 18 gauge injectionneedle cannula which contained a thin-walled flat polyethylene tube (about1.8¿0.6 mm outer and 1.6¿0.4 mm innerdimensions). This polyethylene tube andthe cannula were mounted on a speciallydesigned instrument that could push thetube out of the cannula in a controlledfashion. Further, the plastic tube was at-tached to a precision microsyringe. Do-nor tissue was sucked into the polyethy-lene tube with the help of the microsy-ringe. In this process tissue consisting ofRPE and neural-retina enters the tube asa single piece.
The instrument containing 3 to 4 cupshaped pieces of the donor tissue (about1.6 to 2 mm in diameter) was introduced
into the right eye of the anaesthetizedadult albino rabbits through a scleral in-cision about 5–6 mm behind the limbus.The instrument was advanced transvitre-ally until it reached the predeterminedtransplantation site at the posterior poleof the host eye. On reaching the retina,the polyethylene tube was pushed out ofthe cannula, and introduced into the sub-retinal space by doing a retinotomy withthe tip of the tube itself. The donor tissuewas then deposited in the subretinal spaceby pushing the piston of the microsyringeattached to the cannula. After thesurgery, animals were allowed to wake upand were transferred to their normalcages with a 12/12 hour light/dark cycles.No antibiotics or immunosuppressiveswere given postoperatively. All animalswere sacrificed at a pre-decided time (nΩ15, rangeΩ7 to 63 days). The experimentswere conducted according to the rules setforth in the ARVO resolution on animalexperimentation.
Fixation and stainingEyes were enucleated upon sacrificing theanimals, and fixed in 4% formaldehydefor 24 hours. After fixation, the eyes werehemisected and the part carrying thetransplant was excised. The tissue waswashed with Sorensen’s phosphate buffer
Fig. 1. Photomicrograph of an RPE-neuroretinal cotransplant from E 15 pigmented rabbit retinainto the choroid of an albino rabbit 41 days after transplantation. The figure shows a survivingmonolayer of transplanted pigmented epithelium (RPE-Tr). The neural retina (NR-Tr) also sur-vived but developed rosettes (R). Plastic embedding, methylene blue staining. Ch-HΩhost choroid,OS-HΩhost photoreceptor outer segments, ONL-HΩhost outer nuclear layer.
(0.1M pH 7.2) and then with the samebuffer containing 5%, 10% and 20% su-crose before sectioning on a cryostat (12mm). Four cotransplantation specimenswere fixed in 2.0% glutaraldehyde in 0.1Mphosphate buffer for 24 hours and post-fixed with 1% osmium tetroxide. Thisgives better morphological preservationand also the possibility of future ultra-structural studies, but it does not permitimmunohistochemical analyses. After de-hydration with increasing concentrationsof ethyl alcohol and propylene oxide,pieces of the posterior pole of the eye car-rying the transplant were embedded inplastic and 2 mm thick sections were cuton a microtome. The sections werestained with methylene blue (plastic) orhematoxylin and eosin (cryostat) andexamined under the light microscope.
Observations and ResultsDissection of the donor tissue andtransplantationTreatment with protease K resulted influffing of the sclera and the choroid ofthe donor eyes. The time of action of pro-tease K is very crucial for optimum dis-section. If the time allowed was short andthe eyes were underdigested, it was diffi-

A O S 2000
5
Fig. 2. Photomicrograph of an RPE-neuroretinal cotransplant from E 15 pigmented rabbit retinainto the subretinal space of an albino rabbit, 53 days after transplantation. The figure shows asurviving monolayer of transplanted pigmented epithelium (RPE-Tr). The neural retina (NR-Tr)also survived but became disorganized. The host retina (H) covering the transplant was alsothinned out. Plastic section, methylene blue staining.
Fig. 3. Photomicrograph of an RPE-neuroretinal cotransplant from E 15 pigmented rabbit retinainto the subretinal space of an albino rabbit, 48 days after transplantation. The figure showssurviving pigmented epithelium transplant forming clusters (RPE-Tr with small arrows) and atother places forming a layer (RPE-Tr with large arrows). The neuroretinal cells (NR-Tr) developrosettes (R) and at places plexiform layers also survive (PL-Tr). The host retina overlying thetransplant is degenerated (H). Plastic embedding, methylene blue staining. Ch-HΩhost choroid.
cult to remove the choroid and sclerafrom the RPE and the neural retina. Onthe other hand, if the enzyme was allowedto act for a longer time, the pigment epi-thelium became frail and was easilyripped off from the neural retina duringthe dissection. During the pilot study itwas found that the time of action of theprotease for optimum dissection was fair-ly consistent for various specimens. Thetransplantation process was simple, but itwas not always possible to place thetissue in the subretinal space. On one oc-casion the cannula pierced Bruch’s mem-brane and the donor tissue was trans-planted into the choroid rather than intothe subretinal space (Fig. 1). On two oc-casions, only a part of the transplant wasin the subretinal space, the other partbeing at the site of retinotomy and thevitreal cavity.
Development of the graftDonor pigment epithelium and the neu-ral retina could be observed at all thetransplantation sites. Since the cotrans-plantation was done in albino rabbits, thepigmented cells were presumed to belongto the graft. It is possible for the hostRPE to phagocytize the pigments re-leased by the transplanted RPE, but thelocation of the pigmented cells in relationto the transplanted neural retina sug-gested that indeed these belonged to thegraft. The pigmented cells often formedlarge sheets in the grafts at all the trans-plantation sites (Fig. 1 & 2), but at someplaces the RPE cells formed small clus-ters.
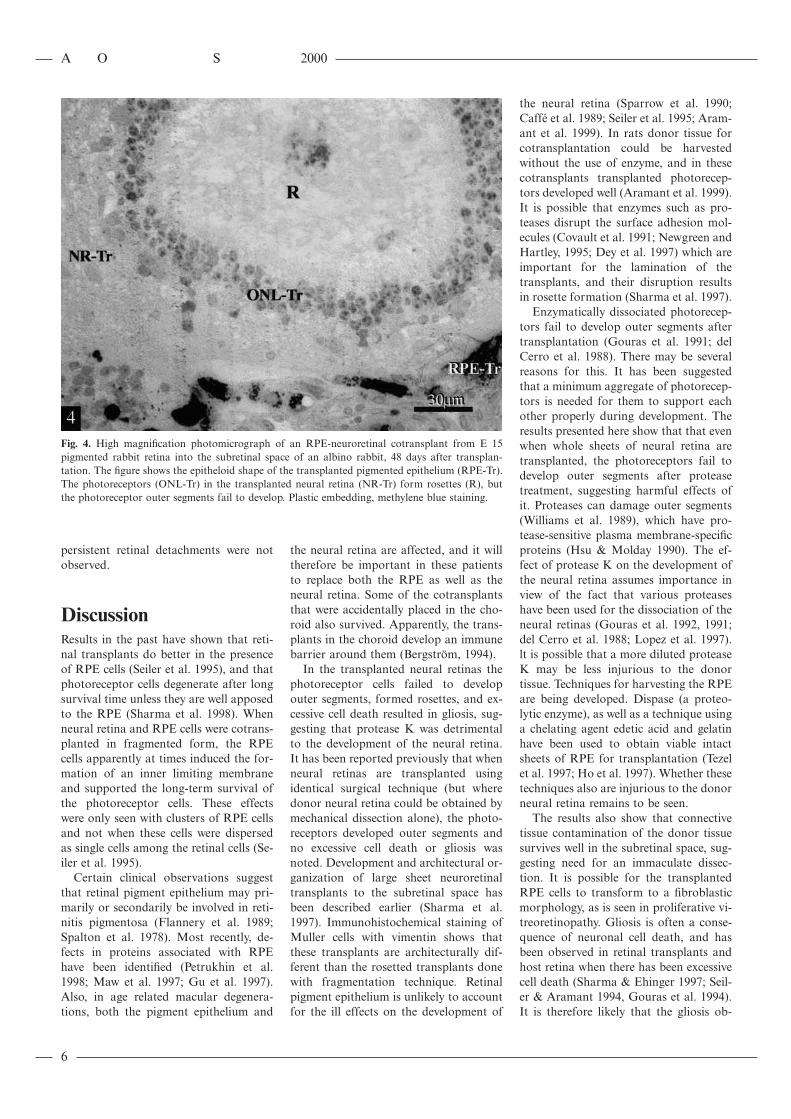
Contrary to expectations, neural retinain cotransplants developed rosettes (Fig.3). The neural retinal transplants wereoften found apposed to the graft RPEwhen the latter formed a sheet. Clustersor isolated RPE cells could be foundwithin the neural retinal transplants.Photoreceptor cells were identified bytheir darkly stained perikarya and theirlocation in the transplants. The photore-ceptor cells were not fully developed asthey lacked the photoreceptor outer seg-ments and the outer limiting membrane(Fig. 4). In some other transplants theneural retina was disorganized. In these,the neuroretinal grafts lacked the rosettedorganization and contained many glialcells, as judged by the overall mor-phology and the pallor of the cytoplasmand the nuclei.
Rarely, few elongated cells identified asconnective tissue could be seen. Compli-cations such as vitreoretinopathies or
A O S 2000
6
Fig. 4. High magnification photomicrograph of an RPE-neuroretinal cotransplant from E 15pigmented rabbit retina into the subretinal space of an albino rabbit, 48 days after transplan-tation. The figure shows the epitheloid shape of the transplanted pigmented epithelium (RPE-Tr).The photoreceptors (ONL-Tr) in the transplanted neural retina (NR-Tr) form rosettes (R), butthe photoreceptor outer segments fail to develop. Plastic embedding, methylene blue staining.
persistent retinal detachments were notobserved.
DiscussionResults in the past have shown that reti-nal transplants do better in the presenceof RPE cells (Seiler et al. 1995), and thatphotoreceptor cells degenerate after longsurvival time unless they are well apposedto the RPE (Sharma et al. 1998). Whenneural retina and RPE cells were cotrans-planted in fragmented form, the RPEcells apparently at times induced the for-mation of an inner limiting membraneand supported the long-term survival ofthe photoreceptor cells. These effectswere only seen with clusters of RPE cellsand not when these cells were dispersedas single cells among the retinal cells (Se-iler et al. 1995).
Certain clinical observations suggestthat retinal pigment epithelium may pri-marily or secondarily be involved in reti-nitis pigmentosa (Flannery et al. 1989;Spalton et al. 1978). Most recently, de-fects in proteins associated with RPEhave been identified (Petrukhin et al.1998; Maw et al. 1997; Gu et al. 1997).Also, in age related macular degenera-tions, both the pigment epithelium and
the neural retina are affected, and it willtherefore be important in these patientsto replace both the RPE as well as theneural retina. Some of the cotransplantsthat were accidentally placed in the cho-roid also survived. Apparently, the trans-plants in the choroid develop an immunebarrier around them (Bergstrom, 1994).
In the transplanted neural retinas thephotoreceptor cells failed to developouter segments, formed rosettes, and ex-cessive cell death resulted in gliosis, sug-gesting that protease K was detrimentalto the development of the neural retina.It has been reported previously that whenneural retinas are transplanted usingidentical surgical technique (but wheredonor neural retina could be obtained bymechanical dissection alone), the photo-receptors developed outer segments andno excessive cell death or gliosis wasnoted. Development and architectural or-ganization of large sheet neuroretinaltransplants to the subretinal space hasbeen described earlier (Sharma et al.1997). Immunohistochemical staining ofMuller cells with vimentin shows thatthese transplants are architecturally dif-ferent than the rosetted transplants donewith fragmentation technique. Retinalpigment epithelium is unlikely to accountfor the ill effects on the development of
the neural retina (Sparrow et al. 1990;Caffe et al. 1989; Seiler et al. 1995; Aram-ant et al. 1999). In rats donor tissue forcotransplantation could be harvestedwithout the use of enzyme, and in thesecotransplants transplanted photorecep-tors developed well (Aramant et al. 1999).It is possible that enzymes such as pro-teases disrupt the surface adhesion mol-ecules (Covault et al. 1991; Newgreen andHartley, 1995; Dey et al. 1997) which areimportant for the lamination of thetransplants, and their disruption resultsin rosette formation (Sharma et al. 1997).
Enzymatically dissociated photorecep-tors fail to develop outer segments aftertransplantation (Gouras et al. 1991; delCerro et al. 1988). There may be severalreasons for this. It has been suggestedthat a minimum aggregate of photorecep-tors is needed for them to support eachother properly during development. Theresults presented here show that that evenwhen whole sheets of neural retina aretransplanted, the photoreceptors fail todevelop outer segments after proteasetreatment, suggesting harmful effects ofit. Proteases can damage outer segments(Williams et al. 1989), which have pro-tease-sensitive plasma membrane-specificproteins (Hsu & Molday 1990). The ef-fect of protease K on the development ofthe neural retina assumes importance inview of the fact that various proteaseshave been used for the dissociation of theneural retinas (Gouras et al. 1992, 1991;del Cerro et al. 1988; Lopez et al. 1997).lt is possible that a more diluted proteaseK may be less injurious to the donortissue. Techniques for harvesting the RPEare being developed. Dispase (a proteo-lytic enzyme), as well as a technique usinga chelating agent edetic acid and gelatinhave been used to obtain viable intactsheets of RPE for transplantation (Tezelet al. 1997; Ho et al. 1997). Whether thesetechniques also are injurious to the donorneural retina remains to be seen.
The results also show that connectivetissue contamination of the donor tissuesurvives well in the subretinal space, sug-gesting need for an immaculate dissec-tion. It is possible for the transplantedRPE cells to transform to a fibroblasticmorphology, as is seen in proliferative vi-treoretinopathy. Gliosis is often a conse-quence of neuronal cell death, and hasbeen observed in retinal transplants andhost retina when there has been excessivecell death (Sharma & Ehinger 1997; Seil-er & Aramant 1994, Gouras et al. 1994).It is therefore likely that the gliosis ob-

A O S 2000
7
served in some transplants may have re-sulted due to the excessive neuronal deathbecause of injurious effects of protease.
In conclusion, this study reports thatdissection of the donor tissue is the mainlimitation in RPE/neuroretinal co-trans-plantation in rabbits, but once these areobtained the donor tissue can be trans-planted into the subretinal space. Pro-tease K was apparently injurious to theneural tissue.
AcknowledgementsThis work was supported by the Swedish Medi-cal Research Council (project 14X-2321), theFoundation Fighting Blindness, Kronprinses-san Margaretas Arbetsnamnd, the SwedishSociety for Medical Research, EU BioMed 2program, Knut and Alice Wallenberg Founda-tion and the Medical Faculty of the Universityof Lund.
ReferencesAlgvere P, Berglin L, Gouras P & Sheng Y
(1994): Transplantation of fetal retinal pig-ment epithelium in age-related macular de-generation with subfoveal neovascularis-ation. Graefe’s Arch Clin Exp Ophthalmol232: 707–716.
Aramant RB, Seiler MS & Ball SL (1999): In-tact sheet fetal cografts of RPE with retinato adult RCS rat retina replace both photo-receptors and RPE. Invest Ophthalmol VisSci 40: 1557–1564.
Bergstrom A, Ehinger B, Wilke K, Zucker CL,Adolph AR, Aramant R & Seiler M (1992):Transplantation of embryonic retina to thesubretinal space in rabbits. Exp Eye Res 55:29–37.
Bergstrom A (1994): Embryonic rabbit retinaltransplants survive and differentiate in thechoroid. Exp Eye Res 59: 281–289.
Caffe AR, Visser H, Jansen HG & Sanyal S(1989): Histotypic differentiation of neo-natal mouse retina in organ culture. CurrEye Res 8: 1083–1092.
Covault J, Liu QY & el Deeb S (1991): Cal-cium-activated proteolysis of intracellulardomains in the cell adhesion moleculesNCAM and N-cadherin. Brain Res MolBrain Res 11: 11–16.
del Cerro M, Notter MF, Wiegand SJ, JiangLQ & del Cerro C (1988): Intraretinal trans-plantation of fluorescently labeled retinalcell suspensions. Neurosci Lett 92: 21–26.
Dey PM, Polunas MA, Philbert MA & ReuhlKR (1997): Altered expression of polysialyl-ated NCAM in mouse hippocampus follow-ing trimethyltin administration. Neuro-toxicology 18: 633–643.
Flannery JG, Farber DB, Bird AC & Bok D(1989): Degenerative changes in a retinaaffected with autosomal dominant retinitis
pigmentosa. Invest Ophthalmol Vis Sci 30:191–211.
Ghosh F, Arner K & Ehinger B (1998): Trans-plant of full-thickness embryonic rabbit ret-ina using pars plane vitrectomy. Retina 18:136–142.
Gouras P, Du J, Gelanze M, Kwun R, Kjeld-bye H & Lopez R (1991): Transplantation ofphotoreceptors labeled with tritiated thymi-dine into RCS rats. Invest Ophthalmol VisSci 32: 1704–1707.
Gouras P, Du J, Kjeldbye H, Yamamoto S &Zack DJ (1992): Reconstruction of degener-ate rd mouse retina by transplantation oftransgenic photoreceptors. Invest Ophthal-mol Vis Sci 33: 2579–2586.
Gouras P, Du J, Kjeldbye H, Yamamoto S &Zack DJ (1994): Long-term photoreceptortransplants in dystrophic and normal mouseretina. Invest Ophthalmol Vis Sci 35: 3145–3153.
Gu S, Thompson DA, Srikumari CRS, LorenzB, Finckh U, Nicoletti A, Murthy KR,Rathmann M, Kumaramanickavel G,Denton MJ & Gal A (1997): Mutations inRPE65 cause autosomal recessive child-hood-onset severe retinal dystrophy. NatGenet 17: 194.
Ho TC, Del Priore LV & Kaplan HJ (1997):Tissue culture of retinal pigment epitheliumfollowing isolation with a gelatin matrixtechnique. Exp Eye Res 64: 133–139.
Hollyfield JG & Witkovsky P (1974): Pig-mented retinal epithelium involvement inphotoreceptor development and function. JExp Zool 189: 357–378.
Hsu SC & Molday RS (1990): Glyceraldehyde-3-phosphate dehydrogenase is a major pro-tein associated with the plasma membraneof retinal photoreceptor outer segments. JBiol Chem 265: 13308–13313.
Kaplan HJ, Tezel TH, Berger AS, Wolf ML &Del Priore LV (1997): Human photoreceptortransplantation in retinitis pigmentosa. Asafety study. Arch Ophthalmol 115: 1168–1172.
Kondo H, Takahashi H & Takahashi Y (1984):Immunohistochemical study of S-100 pro-tein in the postnatal development of Mullercells and astrocytes in the rat retina. CellTissue Res 238: 503–508.
LaVail MM, Sidman M, Rausin R & SidmanRL (1974): Discrimination of light intensityny rats with inherited retinal degeneration.Vision Res 14: 693–702.
Layer PG & Willbold E (1993): Histogenesis ofthe avian retina in reaggregation culture:from dissociated cells to laminar neuronalnetworks. Int Rev Cytol 146: 1–47.
Li L & Turner JE (1988): Transplantation ofretinal pigment epithelial cells to immatureand adult hosts: short- and long-term sur-vival characteristics. Exp Eye Res 47: 771–785.
Lopez R, Gouras P, Kjeldbye H, Sullivan B,Reppucci V, Brittis M, Wapner F & Golu-boff E (1989): Transplanted retinal pigmentepithelium modifies the retinal degeneration
in the RCS rat. Invest Ophthalmol Vis Sci30: 586–588.
Lopez R, Lopez Gallardo M, Medina JI,Ramos M, Ramirez G & Prada C (1997):A Streptomyces fradiae protease dissociatesstructurally preserved neurons and glial cellsfrom the embryonic and adult central ner-vous system of vertebrates. J NeurosciMethods 73: 9–16.
Maw MA, Kennedy B, Knight A, Bridge R,Roth KE, Mani EJ, Mukkadan JK, Nancar-row D, Crabb JW & Denton MJ (1997):Mutation of the gene encoding cellular retin-aldehyde binding protein in autosomal reces-sive retinitis pigmentosa. Nat Genet 17: 198.
Newgreen DF & Hartley L (1995): Extracellu-lar matrix and adhesive molecules in theearly development of the gut and its inner-vation in normal and spotting lethal rat em-bryos. Acta Anat Basel 154: 243–260.
Petrukhin K, Koisti MJ, Bakall B, Li W, XieG, Marknell T, Sandgren O, Forsman K,Holmgren G, Andreasson S, Vujic M,Bergen AA, McGarty Dugan V, Figueroa D,Austin CP, Metzker ML, Caskey CT & Wad-elius C (1998): Identification of the gene re-sponsible for Best macular dystrophy. NatGenet 19: 241–247.
Seiler M, Aramant R & Bergstrom A (1995):Co-transplantation of embryonic retina andretinal pigment epithelial cells to rabbit ret-ina: the importance of retinal pigment epi-thelial cells for retinal differentiation. CurrEye Res 14: 199–207.
Seiler M & Aramant RB (1994): Photoreceptorand glial markers in human embryonic ret-ina and in human embryonic retinal trans-plants to rat retina. Brain Res Dev Brain Res80: 81–95.
Sharma RK, Bergstrom A & Ehinger B (1995):Retinal cell transplants. Prog Retinal EyeRes 15: 197–230.
Sharma RK, Bergstrom A & Ehinger B (1997):Influence of technique and transplantationsite on rosette formation in rabbit retinaltransplants. Acta Ophthalmol Scand 75: 3–10.
Sharma RK, Bergstrom A, Zucker CL, Ad-olph AR & Ehinger B (1998): Long-termrabbit retinal cell transplants. Exp Eye Res67: S.234 (Abstract).
Sharma RK 1999a): Development of glycine-accumulating neurons in transplanted ret-inas. Ophthalmologica (in press).
Sharma RK (1999b): Retinal Transplants:Growth Differentiation Integration Organ-ization and Survival. 1-169 (Ph.D. disser-tation University of Lund, Sweden).
Sharma RK (1999c): Developmental express-ion of GABAA receptors inretinal trans-plants. Ophthalmic Res (In press).
Sharma RK & Ehinger B (1997): Cell prolifer-ation in retinal transplants. Cell Transplant6: 141–148.
Sharma RK & Ehinger B (1999): Managementof hereditary retinal degenerations; presentstatus and future directions. Surv Ophthal-mol 43: 427–444.

A O S 2000
8
Sheedlo HJ, Li L, Gaur VP, Young RW, SeatonAD, Stovall SV, Jaynes CD & Turner JE(1992): Photoreceptor rescue in the dys-trophic retina by transplantation of retinalpigment epithelium. Int Rev Cytol 138: 1–49.
Sheedlo HJ, Li L & Turner JE (1993): Effectsof RPE age and culture conditions on sup-port of photoreceptor cell survival in trans-planted RCS dystrophic rats. Exp Eye Res57: 753–761.
Silverman MS & Hughes SE (1989): Trans-plantation of photoreceptors to light-dam-aged retina. Invest Ophthalmol Vis Sci 30:1684–1690.
Spalton DJ, Bird AC & Cleary PE (1978): Reti-nitis pigmentosa and retinal oedema. Br JOphthalmol 62: 174–182.
Sparrow JR, Hicks D & Barnstable CJ (1990):
Cell commitment and differentiation in ex-plants of embryonic rat neural retina. Com-parison with the developmental potential ofdissociated retina. Brain Res Dev Brain Res51: 69–84.
Tezel TH, Del Priore LV & Kaplan HJ (1997):Harvest and storage of adult human retinalpigment epithelial sheets. Curr Eye Res 16:802–809.
Turner JE & Blair JR (1986): Newborn rat reti-nal cells transplanted into a retinal lesionsite in adult host eyes. Brain Res (Dev BrainRes) 391(26): 91–104.
Williams DS, Shuster TA, Moldrawski MR,Blest AD & Farber DB (1989): Isolation ofrod outer segments on Percoll gradients: ef-fect of specific protease inhibition. Exp EyeRes 49: 439–444.
Young RW & Bok D (1969): Participation of
the retinal pigment epithelium in the rodouter segment renewal process. J Cell Biol42: 392–403.
Received on May 3rd, 1999.Accepted on September 11th, 1999.
Corresponding author:
Dr. Rajesh Kumar Sharma, M.D. Ph.D.Department of OphthalmologyUniversity Hospital of LundS-221 85 LundSwedenPhone: π46 46 172789.Fax: π46 46 172721.e-mail: rajesh–kumar.sharma/oft.lu.se





























