Embed Size (px)
Citation preview

Chapter 3 Distribution of Arctica islandica in the Irish Sea
3-1
Chapter 3
Distribution of Arctica islandica in the Irish Sea
3.1 Distribution of Arctica islandica in the North Atlantic shelf seas
The extensive distribution of Arctica islandica populations in the shelf seas that surround
the north Atlantic is a key aspect of its potential as a climate proxy in these climatically
important regions. The strength and position of the north Atlantic meridional overturning
circulation is a critical element of the forcings and feedbacks that drive the global climate
(Bard, 2002; Sutton and Hodson, 2005; Eiríksson et al., 2006; Hald et al., 2007), and the
detailed insight into historical changes in regional hydrography that is necessary for an
adequate understanding of present day climate change necessitates the identification of
high-resolution marine proxies of precisely the kind provided by A. islandica. Water mass
movements affect shell growth in A. islandica through their influence on food supply and
bottom water temperature (Witbaard, 1996; Witbaard et al., 1997a; Witbaard et al., 2003).
There is also a hydrographic signal in the age of the radiocarbon reservoir, the history of
which can be monitored at high resolution by carrying out radiocarbon analyses on
absolutely-dated shells (see Chapter 4; Butler et al., 2009). Figure 3-1 (adapted from
(Dahlgren et al., 2000)) shows that the sites of living populations and fossil shell of A.
islandica from throughout the Quaternary are within the regions influenced by the North
Atlantic Current and the North Atlantic Polar Front.
If the spatial distribution of A. islandica populations is sufficiently coherent, it may be
possible to build regional networks of chronologies, allowing the common environmental
signal expressed in synchronous shell growth to be monitored and interpreted in space and
in time. Equivalent networks of tree-ring chronologies have been constructed, very

Chapter 3 Distribution of Arctica islandica in the Irish Sea
3-2
extensively for the Northern Hemisphere and on a smaller scale in parts of Southern
Hemisphere (Briffa, 2000), but the explanatory power of these is largely limited to the
atmospheric and terrestrial systems. The value of an integrated proxy history of the marine
environment of the shelf seas surrounding the North Atlantic to research into Holocene
climate change cannot be overstated. The detailed assessment of local and regional
distributions of A. islandica populations will provide an initial insight into possible sites of
absolutely dated chronologies that are anchored with live-collected specimens, and the
distribution of fossil shell highlights the potential for long absolutely-dated chronologies and
floating chronologies with radiometrically constrained dates.
Figure 3-1 Present-day distribution (shaded grey), recently extinct populations (<9,000
years BP, wavy lines) and other extinct populations (from 10,000 years BP back to the
Pleistocene, indicated by dots) of Arctica islandica around the north Atlantic. The study
area is indicated by the white rectangle. Figure adapted from Dahlgren et al (2000).

Chapter 3 Distribution of Arctica islandica in the Irish Sea
3-3
3.2 Existing records of Arctica islandica in the Irish Sea1
An essential prerequisite for the collection of shells for use in an Arctica islandica
chronology was the identification of sites where live clams and fossil shells might be
acquired in sufficient quantities. Existing records of living A. islandica in the Irish Sea were
obtained from various heterogeneous sources (Table 3-1). The primary node for access to
these records was the National Biodiversity Network (http://data.nbn.org.uk/), although
some data (CEDaR, County Down and Isle of Man scallop surveys and the Belfast Lough
monitoring programme) were obtained through personal contact.
Records compiled in this way are likely to be patchy and incomplete. They represent a
combination of thorough but local surveys, wide-ranging and systematic regional surveys
which are generally not targeted at the macrobenthos, and other opportunistic reports.
Because of the focus of the kind of surveys that are likely to recover A. islandica (eg
scientific scallop surveys or habitat surveys connected with conservation areas), they are
likely to be biased towards coastal populations. In contrast, Witbaard and Bergman (2003)
were able to combine data from a number of systematic surveys into a comprehensive
gridded analysis of population densities in all parts of the North Sea. Systematic sampling
of this kind has not been carried out in the Irish Sea. From the point of view of the goal of
this project, a further disadvantage of reliance on existing records was that they generally
recorded only live animals. Although it could reasonably be assumed that fossil shell was
likely to be found within and around existing live populations, nothing was known about the
taphonomy or chronological age range of fossil shell at any particular site and hence it was
not initially possible to assess the potential of any site as the focus of a long shell
chronology.
3.3 Targeted surveys
In order to investigate in detail the distribution of live clams and fossil shells a series of
surveys using a bespoke dredge was carried out targeting sites where A. islandica was
known to occur. The initial site selection was based on existing records. Three dedicated
1 The Irish Sea is defined as the area between latitudes 52ºN and 55ºN (equivalent to ICES Division VIIa).

Chapter 3 Distribution of Arctica islandica in the Irish Sea
3-4
Data provider Dataset Surveys Region Date Marine Biological Association (MBA)
Marine Life Information Network (MarLIN) - Data Archive for Seabed Species and Habitats (DASSH)
CEFAS
UK & Ireland
Marine Biological Association (MBA)
Marine Life Survey Data
Joint Nature Conservation Committee (JNCC)
Marine Data Conservation Review (MNCR)
MNCR UK inshore (< 3nm) & intertidal
1987-1998
Marine Conservation Society (MCS)
Seasearch Marine Surveys
Seasearch Marine Surveys. Dive surveys
Inshore 2003 - present
Countryside Council for Wales (CCW)
Benthic Marine data from CCW Technical Support (Research & Monitoring) Contracts
Wales inshore (< 3nm) & intertidal
1993 - present
Conchological Society of Great Britain & Ireland
Mollusc (marine) data for Great Britain and Ireland
Marine census Great Britain & Ireland
1876 - present
National Museums, Northern Ireland Centre for Environmental Data and Recording (CEDaR)
Irish Sea
Countryside Council for Wales (CCW)
Pembrokeshire Marine Species Atlas
Pembrokeshire 1899-1998
National Museum of Wales Biomor Surveys / Macrofaunal survey of Welsh sandbanks
Irish Sea 1989 - 2001
Environment Agency Macrobenthic monitoring near Milford Haven
1996-1997
Countryside Council for Wales (CCW)
Tremadog Bay sublittoral sediment survey / HABMAP survey of southern Irish Sea
2005
Dept of Agriculture and Rural Development, Northern Ireland (DARDNI)
County Down scallop surveys
County Down coast
2001 and 2005
Port Erin Marine Laboratory, University of Liverpool / Isle of Man Government
Isle of Man scallop surveys
Isle of Man waters
2000-2004
Environment and Heritage Service, Northern Ireland
Pollution monitoring in Belfast Lough
Belfast Lough
Table 3-1. Sources of Arctica islandica observations

Chapter 3 Distribution of Arctica islandica in the Irish Sea
3-5
cruises of the RV Prince Madog to different parts of the Irish Sea were undertaken, and the
customized “Arctica” dredge (Figure 2-2) was used to collect living animals and fossil shells.
Subsequently some of the same stations (in particular the target site for the chronology to
the west of the Isle of Man and the site of a particularly dense population in Belfast Lough)
were resampled and it was possible to estimate the distributions around these stations in
greater detail. The details of the cruises, and the numbers of live and dead A. islandica
shells collected, are shown in Appendix C.
3.4 Distribution of Arctica islandica in the Irish Sea
The spread of observations of live A. islandica in the Irish Sea, including pre-existing
records and results from the targeted surveys, is shown in Figure 3-2. Sites sampled with
the “Arctica” dredge during targeted cruises are shown with red circles; the size of the
circles shows the relative density as numbers of live specimens collected per km trawled.
The actual numbers collected per km are shown in Appendix C and on the detailed maps of
local areas (Figures 3-4 to 3-10). Because the efficiency of the fishing method is not
known, this gives no indication of the true density of the populations. The positions of other
records of live specimens are shown with black stars. The brown and yellow shaded areas
indicate sediments which are respectively too fine (muddy) or too coarse (gravelly) to
support A. islandica. The unshaded areas indicate various grades of muddy sand, sand
and sandy mud, all of which may support A. islandica. The contour lines show the 50- and
100-metre isobaths.
The greatest density of A. islandica records occurs in coastal areas. This does not
necessarily demonstrate that the densest populations of A. islandica are in these areas,
since opportunistic sightings and surveys conducted in relation to marine conservation
areas are likely to be biased to inshore waters. Nevertheless, there are extensive offshore
areas where the sediment is too fine or too coarse to support this species (Figure 3-2), and
consequently it seems unlikely that the Irish Sea hosts any significant offshore populations
of comparable density to those that have been identified in parts of the North Sea (Witbaard
and Bergman, 2003) and off the north American coast (Lewis et al., 2001). In targeted
dredging of offshore sites, the most concentrated populations have been found to the north

Chapter 3 Distribution of Arctica islandica in the Irish Sea
3-6
Figure 3-2 Recorded observations of Arctica islandica in the Irish Sea. Filled red circles show
sites where live specimens were collected with the bespoke “Arctica” dredge. The size of the
circle indicates the relative density. Stars show other records of live Arctica islandica. Yellow
shading indicates gravelly sediment, brown indicates muddy sediment (both unsuitable for A.
islandica). 50- and 100-metre isobaths are also shown. Geological and bathymetric
information adapted from Tappin et al (1994) and Jackson et al (1995)

Chapter 3 Distribution of Arctica islandica in the Irish Sea
3-7
and west of the Isle of Man, but these are sparse by comparison. In coastal areas, A.
islandica populations are concentrated in the northern part of the eastern Irish coast
(Belfast Lough, Strangford Lough, Carlingford Lough and Dublin Bay), the southwest coast
of the Isle of Man, and the west coast of Wales (Caernarfon Bay, central and northern parts
of Cardigan Bay and the west coast of Pembrokeshire). Records of A. islandica from the
Irish coast south of Dublin and the Scottish, English and north Wales coasts around the
eastern Irish Sea are sparse, an indication, given the bias towards coastal reports, that it is
in fact rare in these areas.
It has been shown (Chapter 5; Butler et al., submitted) that food supply is probably not a
limiting factor in the growth of A. islandica in waters to the west of the Isle of Man. Since
these waters are reasonably typical of the main body of Irish Sea water, and since nutrient
concentrations in coastal areas are generally higher than they are in offshore waters
(Gowen et al., 2002; Gowen and Stewart, 2005), it is acceptable, at least as a first
approximation, to extend this conclusion to all parts of the Irish Sea. Consequently, the
principle controls on the distribution of A. islandica can be reduced to hydrography (which
controls the distribution and retention of larvae), sediment type (which determines the
possibility of settlement), and seawater temperature (which affects the survival of larvae). If
sediment type and temperature are assumed to be constant with time, then populations of
A. islandica will be supported wherever the local hydrography supplies a larval retention
mechanism or reliably imports larvae from elsewhere. A retention mechanism can be
assumed to operate where outflow to the Irish Sea is topographically restricted, as it is in
Carlingford Lough, Strangford Lough and Larne Lough, and A. islandica has indeed been
recorded in all these semi-enclosed sea loughs (figures 3-4 and 3-5). Other estuaries and
embayments which support A. islandica but which are exposed to the main body of the Irish
Sea (such as Belfast Lough, Caernarfon Bay and the north and central parts of Cardigan
Bay) would require topographically steered gyres or a suitable inward residual flow pattern
to ensure the persistence of the populations over time.
Populations in the open waters of the Irish Sea appear to be maintained by large-scale
circulation patterns associated with tidal mixing fronts. Where a summer thermocline
develops above a cold, saline pool of water left over from the previous winter, strong near-
bed density gradients develop, driving a cyclonic circulation system. In the Irish Sea, such
systems have been described for the area west of the Isle of Man and north of Dublin

Chapter 3 Distribution of Arctica islandica in the Irish Sea
3-8
(Horsburgh et al., 2000), and for the part of the Celtic Sea front that lies above the Celtic
Deep (Brown et al., 2003). Populations of A. islandica have been found associated with
both of these systems (see sections 3.4.2 and 3.4.7).
The surveys carried out during this research found no evidence of recent settlement in any
A. islandica population south and east of the Isle of Man. Whereas some small clams (<50
mm) were recovered from Belfast Lough and from northwest of the Isle of Man (station D11;
see Figure 3-5), these appeared (on the basis of counts of external growth increments) to
have settled no more than four or five years earlier, and only one live specimen smaller
than 70mm was found at any other site (Red Wharf Bay, Anglesey). The increase in
seawater temperatures in the Irish Sea during the A. islandica reproduction season has
been significant in recent decades, with integrated mean bottom temperatures for the
months from August to November showing a stepwise increase from 12.8º C to 13.1º C in
1987 and again to 14.1º C in 1997 (Figure 3-2). The upper limits to optimal temperatures
for various stages of larval development may vary regionally, but research on populations
from the North American coast has indicated that survival to the veliger stage is adversely
affected at temperatures above 15º C and that survival through metamorphosis is adversely
affected above 13º C (Landers, 1976; Lutz et al., 1982; Mann and Wolf, 1983). If
comparable limiting temperatures operate in the Irish Sea communities, it is likely that the
ability of the larvae to progress successfully to settlement has been compromised during
the past twenty years. The temperatures shown in Figure 3-2 were measured at the Cypris
station off the southwest of the Isle of Man (Figure 4-1). The temperature gradients in the
Irish Sea are positive to the south and east in relation to Cypris and negative only to the
north. The observation that only a few (<10) small clams were recovered, and these only
from north of the Isle of Man and from Belfast Lough is consistent with the hypothesis that
successful larval production, survival and settlement in A. islandica populations south and
east of the Isle of Man has declined or even ceased because of increasing seawater
temperature. A similar absence of juvenile A. islandica in areas where there were large
numbers of mature clams was noted in the English Channel during the 1950s and 1960s
(Holme, 1953; Holme, 1966), and no settlement has been observed there subsequently.
These observations may indicate a northward movement of the southern limit of the range
of A. islandica, but it is also important to bear in mind that intermittent settlement with long
hiatuses is characteristic of such long-lived species and that several decades may
sometimes elapse between settlement events (Witbaard and Bergman, 2003).

Chapter 3 Distribution of Arctica islandica in the Irish Sea
3-9
The following sections (3.4.1 to 3.4.7) will analyse the details of the distribution of A.
islandica in the Irish Sea with reference to small scale hydrographic and topographic
provinces.
3.4.1 Belfast Lough
Records of A. islandica are common from the outer parts of Belfast Lough, and the area
sampled with the bespoke “Arctica” dredge (station C - larger red circle in Figure 3-4)
contains a particularly dense population where the catch rate was >50 individuals per km
towed. The spread of pre-existing records suggests that there may be other high density
concentrations in the same area. The substrate between the 20m and 60m isobaths is a
coarse sand that is suitable for settlement, and a cyclonic gyre at the mouth of Belfast
Lough, predicted by a three-dimensional model for the North Channel and with some
observational confirmation (Young et al., 2000), could act as a larval retention mechanism.
Figure 3-3 Mean bottom seawater temperatures (red plot) for the months August-
November measured at the Cypris station (see Figure 4-1) off Port Erin, Isle of Man. The
horizontal black lines show the means for the three periods shown.

Chapter 3 Distribution of Arctica islandica in the Irish Sea
3-10
3.4.2 The Isle of Man and the distribution of Arctica islandica around the western
Irish Sea gyre
The population distribution of A. islandica in the northwest Irish Sea appears to follow the
position of the tidal mixing front associated with the western Irish Sea gyre (Hill et al., 1994;
Horsburgh et al., 2000). Figure 3-5 shows typical potential energy anomaly contours (that
is, the amount of energy required to mix the stratified water column (Simpson, 1981))
associated with the outer margin of the gyre. The relatively dense population off the west
coast of the Isle of Man follows the margin of the gyre to the north. Although A. islandica
has not been recorded in the southern part of the North Channel, it is found along the
southward arm of the gyre from the Ards peninsula to Dublin Bay, and along the eastward
arm from Dublin Bay. Larval retention in the gyre has previously been suggested for
Figure 3-4 Recorded observations of A.
islandica from Belfast Lough and the east
coast of Northern Ireland. Symbols as
described for Figure 3-2. Text inside or next
to the red circles shows the station ID (from
Table 3-3) and the number of live A.
islandica collected per kilometre towed with
the bespoke dredge.

Chapter 3 Distribution of Arctica islandica in the Irish Sea
3-11
pelagic fish larvae (Dickey-Collas et al., 1997) and for the larvae of the commercially
important Norway lobster Nephrops norvegicus (Hill et al., 1996), and it seems likely that
the gyre is also a driving mechanism for the distribution of A. islandica. Larvae that are
entrained in the outer margin of the gyre are likely to settle in sandy sediments,
intermediate between the muds in the centre and the gravels that are typical of mixed
waters outside the gyre, and it is this type of sediments that is optimal for successful
settlement and growth of A. islandica (Witbaard and Bergman, 2003).
Figure 3-5 Distribution of A. islandica around the Isle of Man and the western Irish Sea
gyre. The contours are the 20 and 30 J m-3 potential energy anomaly contours showing the
boundary of the gyre as measured by Horsburgh et al. (2000) in July 1996. For key see
Figures 3-2 and 3-4.

Chapter 3 Distribution of Arctica islandica in the Irish Sea
3-12
3.4.3 Eastern Irish Sea
The eastern Irish Sea is a large,
shallow (<50 m) embayment
east of the Isle of Man. Existing
records of A. islandica are
isolated (Figure 3-6), and no
targeted dredging with the
bespoke “Arctica” dredge was
carried out in this area. The
benthos of the eastern Irish Sea
has, however, been extensively
sampled in recent decades
because of concerns about
pollution associated with the
nuclear reprocessing plant at
Sellafield, and it seems most
likely that A. islandica is indeed
uncommon there. The single
exception is Laxey Bay on the east coast of the Isle of Man, where substantial quantities of
recently dead shell have been recovered during scientific scallop surveys, and from where
live specimens have been collected for use in metabolism studies (Taylor and Brand, 1975).
In spite of the fact that there are no obvious physical or sedimentary barriers to the
occurrence of A. islandica populations east of the Isle of Man, the Laxey Bay community
remains the only substantial community that has so far been identified. Because of river
runoff from surrounding land masses, salinity in the eastern Irish Sea is low compared with
the rest of the Irish Sea, and can fall to around 31 in winter (Gowen et al., 2008), but this is
not a limiting factor for A. islandica, which tolerates considerably lower salinities in Baltic
Sea waters (Zettler et al., 2001). Apart from a muddy patch off Sellafield, the sediment type
is generally suitable for settlement. Summer temperatures are no higher than they are in
northern Cardigan Bay, where A. islandica is plentiful. It is possible that the development of
local populations is inhibited by the absence of any persistent circulation patterns in the
area. The production of larvae is contingent on the spatial distribution of the clams being
sufficiently dense for fertilization to take place, and the maintenance of the population is
Figure 3-6 Distribution of A. islandica in the eastern
Irish Sea. For key see Figure 3-2.
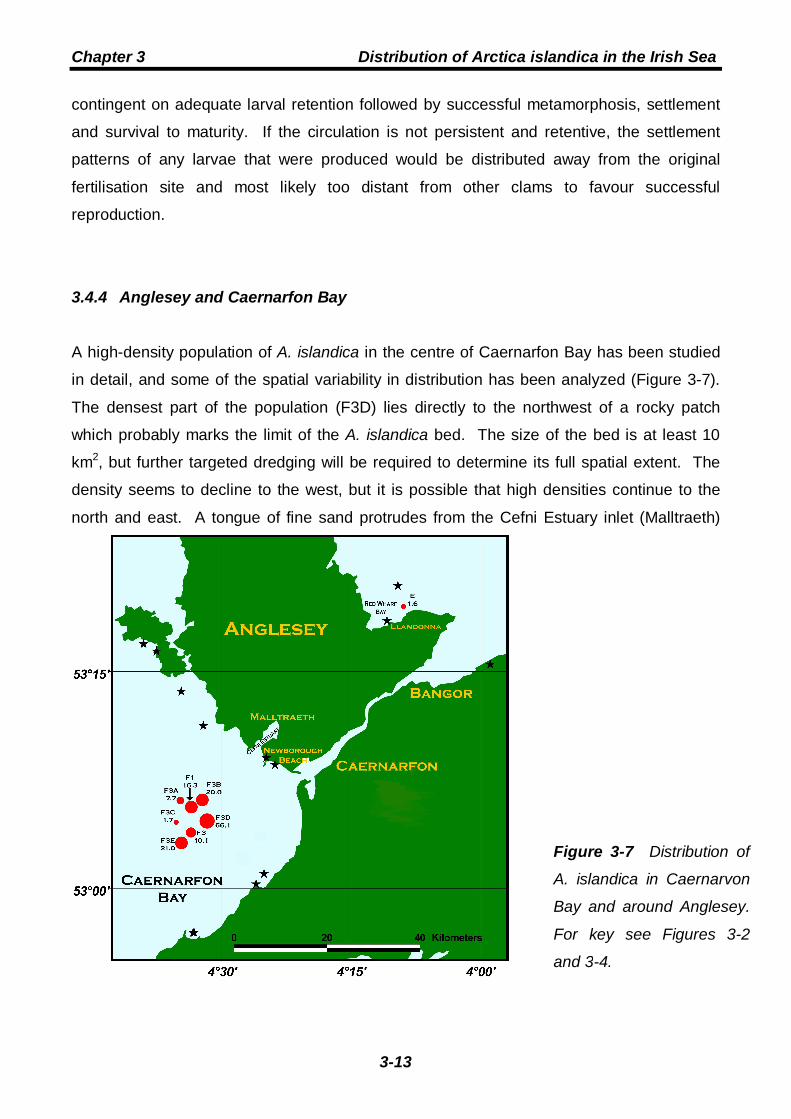
Chapter 3 Distribution of Arctica islandica in the Irish Sea
3-13
Figure 3-7 Distribution of
A. islandica in Caernarvon
Bay and around Anglesey.
For key see Figures 3-2
and 3-4.
contingent on adequate larval retention followed by successful metamorphosis, settlement
and survival to maturity. If the circulation is not persistent and retentive, the settlement
patterns of any larvae that were produced would be distributed away from the original
fertilisation site and most likely too distant from other clams to favour successful
reproduction.
3.4.4 Anglesey and Caernarfon Bay
A high-density population of A. islandica in the centre of Caernarfon Bay has been studied
in detail, and some of the spatial variability in distribution has been analyzed (Figure 3-7).
The densest part of the population (F3D) lies directly to the northwest of a rocky patch
which probably marks the limit of the A. islandica bed. The size of the bed is at least 10
km2, but further targeted dredging will be required to determine its full spatial extent. The
density seems to decline to the west, but it is possible that high densities continue to the
north and east. A tongue of fine sand protrudes from the Cefni Estuary inlet (Malltraeth)

Chapter 3 Distribution of Arctica islandica in the Irish Sea
3-14
inlet next to Newborough Beach as far as the position of station F3E (James and Wingfield,
1990), and this may indicate the stable centre of a circulation cell governed by the tidal
dynamics and topography of the bay. Significant numbers of A. islandica stranded alive on
Newborough Beach after a heavy storm in December 1968 (E.I.S.Rees, personal
communication) are unlikely to have originated directly from the area studied (a distance
approaching 20 km), but they may have come from its extension or from a separate patch
closer inshore. A single small population close to Llandonna (Red Wharf Bay) on the east
coast of Anglesey (station E) inhabits a patch of muddy sand surrounded by coarser sands.
It seems likely that a circulation pattern associated with the nearby headland is responsible
both for the sediment configuration and for the maintenance of the A. islandica population.
3.4.5 Cardigan Bay north
Suitable habitats for A. islandica in the northern part of Cardigan Bay (Figure 3-8) are
generally confined to the sands which stretch from the Dyfi and Mawddach estuaries as far
as the 20 m isobath and to the more complex sediments in Tremadoc Bay. Where targeted
dredging has been carried out, low densities have been found, but the distribution of
observations indicates that A. islandica may be found wherever the sediment is suitable.
Because of the status of the area as an SAC (Special Area of Conservation), it was not
possible to carry out any targeted dredging in Tremadoc Bay, but the distribution of
recorded observations between 1986 and 2007 indicates that it is common in the fine and
muddy sands in the eastern and southern parts of the bay. Sediments in the west of the
bay are gravelly and probably unsuitable, but A. islandica has been observed inshore of
these sediments and a shallow-water population at Llanbedrog has been observed exposed
above the water line at extreme spring tides.
Modelling studies indicate that with the exception of cells around the tip of the Llyn
Peninsula and to the west of Tremadoc Bay, tidal circulation in the northern part of
Cardigan Bay is generally weak and that a stronger and rather complex circulation
throughout Tremadoc Bay develops only at periods of peak wind stresses (Neill et al.,
2007).

Chapter 3 Distribution of Arctica islandica in the Irish Sea
3-15
The prevailing winds impacting on Cardigan Bay are south westerly and sometimes severe.
Since there is no net circulation away from Cardigan Bay, the effect of residual flow on
planktonic larvae will be retentive and shoreward. The effective area of settlement is
constrained by sediment type to precisely these shoreward areas, increasing the likelihood
that populations may develop which are sufficiently dense to facilitate fertilization.
3.4.6 Mid Cardigan Bay
The distribution of pre-existing records indicates that the frequent occurrence of A. islandica
continues south of the Dyfi estuary (Figure 3-9). Detailed sampling with the bespoke
dredge of an area to the southwest of Aberystwyth identified an A. islandica bed of about 3
km2, immediately west of a rare patch of the Mediterranean bivalve Callista chione (Figure
Figure 3-8 Distribution of A. islandica in the northern part of Cardigan Bay. For
key see Figures 3-2 and 3-4.

Chapter 3 Distribution of Arctica islandica in the Irish Sea
3-16
3-9). Shoreward, this patch is limited by a rocky substrate. No observations have been
recorded further than about 20 km from the shore, where the sediment is seen to grade
offshore into sandy gravels. South of Newquay, tidal and wind-driven flows become more
energetic as St George’s Channel narrows towards St David’s Head. The coarser
sediments extend to the shore, and records of A. islandica are unusual.
Figure 3-9 Distribution of A. islandica in mid Cardigan Bay. For key see
Figures 3-2 and 3-4.
3.4.7 The Celtic Sea front and St George’s Channel
The Celtic Sea front separates the mixed waters of the southern Irish Sea from the deeper,
seasonally stratified waters of the Celtic Sea to the south. Midway along the front a
cyclonic (anticlockwise) circulation, dynamically equivalent to the western Irish Sea gyre
described in section 3.4.2, is baroclinically steered around a pool of cold, saline (and
therefore dense) bottom water. The density gradient causes bottom water to be advected
north, then west and south at speeds in excess of 20 cm s-1 (Brown et al., 2003). Records

Chapter 3 Distribution of Arctica islandica in the Irish Sea
3-17
of A. islandica from the Celtic Sea frontal region appear to be associated with this
circulation (Figure 3-10), indicating that it functions as a larval retention mechanism.
Without such a mechanism, the establishment and maintenance of populations of A.
islandica in the region of the Celtic Sea front would be unlikely, in spite of the suitability of
the sediment on the stratified (southern) side. Even if a chance settlement of a large
number of larvae were to establish a single cohort, any developing larvae would likely be
dispersed too widely to maintain the existing population or to establish another. A more
likely direction of distribution is northwards. A narrow corridor of sandy sediment connects
the northern arm of the circulation cell to the northern part of Cardigan Bay (Figure 3-2),
Figure 3-10 Distribution of A. islandica around southwest Wales and in the
southern entrance to the Irish Sea. The contours show the Celtic front
circulation as the bottom density gradient in late summer 1998, adapted from
Brown et al (2003; Figure 13). Arrows show direction of flow. For key see
Figures 3-2 and 3-4.

Chapter 3 Distribution of Arctica islandica in the Irish Sea
3-18
and given the prevailing southerly and south westerly winds, it is possible that a chain of A.
islandica populations might have become established along this corridor.
3.5 Distribution of fossil shells
The distribution of living populations of A. islandica, both in the Irish Sea and in other shelf
sea waters around the North Atlantic region, indicates that the species has potential to be
used for the construction of networks of linked chronologies, making available a powerful
proxy for the temperate and boreal marine environments. However, the realistic time span
of chronologies based on live-caught specimens alone covers no more than one or two
centuries. The establishment of absolute dating control requires that a chronology be
anchored in time with the use of shells from clams whose date of death is known (most
likely because they have been live caught), but if the proxies are to be informative on multi-
centennial or millennial timescales, the acquisition of sufficient usable quantities of older
shell material is essential. This is the major challenge for the construction of long A.
islandica chronologies. The isolation of the common environmental signal for the Irish Sea
population used in this study requires that at least three or four coeval shells with well-
synchronized growth increment patterns (see references to the expressed population signal
(EPS) in chapters 4 and 5) are included in the chronology. The availability and condition of
fossil shells therefore constitute the primary constraints on the length of an absolutely-dated
chronology.
By default, the valves of dead clams remain in situ, vertically oriented and buried up to 6 cm
deep in the sediment. Although they disarticulate as the organic ligament connecting them
degrades, they may, if the sediment is sufficiently well compacted, remain in their live
configuration. In the case of A. islandica, which is not generally found in compacted
sediment, the valves are more likely to drift apart as shear stresses act on the sediment
matrix. Subsequent sediment accumulation may result in the valves becoming buried more
deeply; alternatively they may become exposed on the seabed and subject to mobilization
by bottom currents. Taking these and other considerations into account, it is possible to
identify the primary factors bearing on the availability of suitable fossil shells:
(a) A population must have been living on or around the site during the target period. This
is by no means trivial, since, except in very dense communities, A. islandica does not

Chapter 3 Distribution of Arctica islandica in the Irish Sea
3-19
typically reproduce continuously; rather, discrete settlement events may occur at multi-
decadal intervals (Witbaard et al., 1997a). A consequence of this is that A. islandica may
be absent from a site for significant periods, effectively truncating the absolutely-dated
chronology (see, for example the distribution of 14C ages of shells from the Isle of Man site
shown in Figure 4-7(A), where a hiatus, indicating a possible absence, is apparent during
the 14th century).
(b) The fossil shells must have remained in situ. While it is theoretically possible for shells
with chronological ages measured in hundreds or thousands of years to have travelled
great distances under the influence of bottom currents, it should be borne in mind that (i)
they would likely have undergone such a degree of physical degradation during the journey
as to render them unsuitable for increment-width measurement and (ii) even if they were in
a suitable condition to be measured, they could only be included in the chronology if their
increment-width patterns demonstrated synchronized growth with other fossil shells found in
the locality. The corollary of this is that any fossil shell whose increment widths can be
measured and which is sufficiently well synchronized to be included in the chronology
originated at a site which was ipso facto close enough. In practise, it is likely that any fossil
shells found on site originated at that site. Post mortem burial and topographically
prescribed shell lags represent very effective constraints on mobilization and transport of
the relatively heavy A. islandica shell.
(c) The fossil shells must be readily available on or close to the seabed and they must be in
good physical condition. While deep burial is an excellent method of preservation for shells
of great antiquity, it is also a significant obstacle to their recovery. The optimal fossil shell
for sclerochronological work is one that has been deeply buried throughout its post mortem
existence and has only recently been returned to the seabed or shallow burial by sediment
mobilization. If natural processes alone were operating, such shells would be extremely
difficult to identify and collect, since sediment transport is slow and difficult to model and
shells buried within the sediment are not detectable. It is likely, however, that these natural
processes have been anthropogenically short circuited. Dredging of the seabed by
commercial fisheries has accelerated sediment mobilization, bringing to the sediment
surface shells that had been buried for centuries or millennia. In waters off the Isle of Man
and the east coast of Northern Ireland, the shell of A. islandica is a common bycatch of
scallop dredging, and all the shells that have been radiocarbon dated for this project were
acquired from routine scientific scallop surveys in those two areas. A necessary condition
for the selection of a shell for submission for dating was that its physical condition showed

Chapter 3 Distribution of Arctica islandica in the Irish Sea
3-20
little or no indication of chemical diagenesis or bioerosion. The range of radiocarbon dates
includes 39 from the Isle of Man and one from Northern Ireland with calibrated ages greater
than 1,000 calendar years BP, the earliest of these having a calibrated age of ~8,670
calendar years BP (see Table 4-1 and Figure 4-4). The combination of antiquity and good
physical condition is a strong indication that these shells had been preserved in the
sediment until very recently. In Isle of Man waters, commercial scallop fishing began in
1937 and expanded rapidly after 1945 (Brand, 1999). Present-day populations of A.
islandica occur within the scallop grounds and just offshore from them; it seems likely that
these communities have existed in the area, possibly continuously, for several millennia and
that many thousands of shells from throughout that period have recently become available
for sclerochronological research.
The distribution of fossil A. islandica shells in the Irish Sea is shown (as blue circles) in
Figure 3-11. Although there are some pre-existing observations of fossil shells, these have
not generally been recorded as systematically or as extensively as live specimens and
hence most of the records shown in Figure 3-11 were obtained during the targeted surveys
with the RV Prince Madog, the routine scallop surveys in Isle of Man and Northern Ireland
waters, or opportunistically during other cruises of the Prince Madog and observations on
Irish and Welsh beaches in 2005 and 2006. As expected, fossil shells are most commonly
associated with living populations (red circles). There are a small number of instances
where this is not obviously the case. Several shells were found on beaches around
Rosslare (on the southeast Irish coast), indicating either that an unrecorded living
population exists or has existed nearby, or that the prevailing regional hydrography is
transporting shoreward shells from a wider offshore area. Some of the shells recovered
from the area to the south of the Isle of Man were of relatively recent appearance, perhaps
indicating the nearby presence of a sparse population associated with the western Irish Sea
gyre. The isolated records in the eastern Irish Sea tend to confirm the intermittent
distribution of living A. islandica there. The relative frequency of records of fossil shells
around Dublin reflects a curious set of observations from fisheries surveys in the early 20th
century.

Chapter 3 Distribution of Arctica islandica in the Irish Sea
3-21
3.6 Potential for the use of fossil shells in extended chronologies
The size, mass and external condition of fossil shells can be used to obtain a first indication
of their potential for use in a chronology. The size and mass of a shell (in relation to the
mean size and mass of shells in the local population) indicate its longevity, while its external
condition may reflect its chronological age (indicated by the degree of dissolution of the
organic periostracum and hinge ligament) or the internal state, and hence measurability, of
the growth increments (indicated by the extent of bioerosion). During the curation
procedure shell condition is assessed by assigning an index to each of five properties (state
Figure 3-11 Distribution of live A. islandica (red) and dead shells (blue) in the Irish
Sea. All records are shown, with no indication of relative density or of the collection
method.

Chapter 3 Distribution of Arctica islandica in the Irish Sea
3-22
of periostracum, state of hinge ligament, degree of erosion at the shell margin, degree of
bioerosion and state of the nacre layer), with high indices indicating live-collected or
recently dead shells and low indices showing erosion or dissolution. Figure 3-12 shows the
normalized condition indices for fossil shells from nine areas in the Irish Sea and for the
whole of the Irish Sea. The two extremes are the group from St. George’s Channel (low
indices) and that from Laxey Bay (high indices). The low bioerosion score of the shells from
St George’s Channel indicates that post mortem they have been lying on or close to the
sediment surface, leaving them vulnerable to boring and encrustations. This further
suggests that the low scores for periostracum and ligament condition of the same shells are
probably not an indication of antiquity; it is more likely that they result from accelerated
degradation processes. The high score for the shells from Laxey Bay indicates that many
of them are only recently dead. Most of the shells from Laxey Bay were obtained as
bycatch during scallop dredging, and Laxey Bay has not as yet been sampled with the
bespoke “Arctica” dredge. Scallop dredges generally do not dig into the sediment, and
therefore they rarely collect live A. islandica, but they will retain shells that are on the
seabed or partially exposed. Relatively few shells have been recovered from Laxey Bay
with degraded or absent periostracum and ligament suggesting either that the population
there is relatively recent, or alternatively that the older shells are swiftly buried or removed
to a lag outside the scallop ground. By contrast, the periostracum and ligament indices for
Caernarfon Bay are low, indicating that the shells recovered there have a considerably
wider temporal range.
-2.5-2.0-1.5-1.0-0.50.00.51.01.52.02.5
Periostracum Ligament Margin Bioerosion Nacre
Z-sc
ores
All (2457) St Georges Channel (85) NI coast (167) Caernarfon Bay (539) Red Wharf Bay (62)IOM west (1214) Eastern Irish Sea (30) Cardigan Bay (28) Belfast Lough (237) Laxey Bay (87)
Figure 3-12. Normalized mean condition indices of fossil A. islandica shells from nine
areas of the Irish Sea. Black columns show the values for all fossil shells. Numbers in
brackets show number of shells in the group.

Chapter 3 Distribution of Arctica islandica in the Irish Sea
3-23
The availability in each locality of fossil shells with potential for inclusion in a long
chronology can be indicated by the proportion of shells which (a) have little or no
periostracum and ligament (indices equal to 1 or 2) and which (b) are not bioeroded
(bioerosion index equal to 3 or 4). The first of these criteria is intended to select for
chronological age, while the second is intended to select for measurability. Table 3-2
shows the percentage of usable shells from each site. Suitable specimens can be obtained
from all sites, but IOM west, Caernarfon Bay and NI coast appear to have the greatest
potential to supply sufficient shells for extended chronologies.
Site Usable Total % usable
Laxey Bay 14 87 16.1
Belfast Lough 65 237 27.4 Red Wharf Bay 19 62 30.6
Eastern Irish Sea 12 30 40.0 IOM west 529 1215 43.5
Cardigan Bay 14 28 50.0 St George’s Channel 48 85 56.5
Caernarfon Bay 362 539 67.2 NI coast 114 167 68.3
3.7 Selection of chronology site and prospects for further research
The ultimate selection of the IOM west site for the chronology was governed not only by the
numbers of live and fossil shells available there, but also by the potential for instrumental
calibration afforded by the proximity of the Cypris and Port Erin breakwater monitoring
stations (Figure 4-1) and the results of the first phase of radiocarbon dating, which showed
a good spread of dates through the past millennium (Table 4-1). Five shells from the NI
coast site were also radiocarbon dated in the first phase. The results of four of these
indicated a date of death after 1950 (post-bomb pulse) and the 14C date of the other was
5338±26 14C years BP. This suggested that, while there may be a very wide range of
chronological ages available from the NI coast, targeting of specific periods within that
range might present a significant challenge. The Caernarfon Bay population was not
Table 3-2 Numbers of fossil shells from each of nine sites that are considered
to be usable in a long chronology. Usability determined by taphonomic
indications of antiquity and measurability (see text for criteria).

Chapter 3 Distribution of Arctica islandica in the Irish Sea
3-24
considered for the chronology because it was thought that the local hydrographic regime
there was not typical of the main body of Irish Sea waters and that the common
environmental signal there might be obscured by disturbance effects related to the
enhanced impact of severe storms on a shallow water community. Although synchronous
growth often appears to be absent in A. islandica from shallow water environments (Epplé
et al., 2006), the Caernarfon Bay site may be an appropriate focus for a close study of the
complex increment-width patterns that seem to be typical of such settings.
Sufficient material is available from most parts of the Irish Sea to study historical patterns of
recruitment in A. islandica populations during the period since the industrial revolution. By
increment counting and crossdating, it is possible to identify the exact years of settlement of
individual animals and by integrating this information for many animals, variations in
frequency and density of settlement in different localities can be reconstructed. The
scarcity of juvenile clams in all but the coldest parts of the Irish Sea strongly indicates that
the reproductive cycle in A. islandica is temperature stressed, and the recent history of
distribution and settlement can function as a proxy for temperature changes and possibly
also for changes in the hydrographic and sedimentary conditions that maintain populations.
Findings from this research can then inform further studies of equivalent trends in other
marginal areas for A. islandica, including the Celtic Sea, the North Sea and the most
southerly populations on the Atlantic coast of North America.
3.8 Notes on the lifecycle of Arctica islandica
Aspects of the lifecycle of A. islandica, including time spent in the larval and veliger stages,
age at sexual maturity and time of spawning are relevant to the use of the shell archive and
will be covered briefly here.
Age at sexual maturity. It is known that A. islandica produces growth lines in its shell
throughout its life, even during the juvenile phase before sexual maturity. If the growth
check in the mature animal is related to spawning, as some authors have suggested
(Thompson et al., 1980a, Schöne et al., 2005b), then there may be significant differences in
the mechanisms of growth line formation in juvenile and mature animals, so that the
decision whether or not to include juvenile increments in time series may impact on the
expression of the climate signal in the resultant chronology. It has not yet been possible to

Chapter 3 Distribution of Arctica islandica in the Irish Sea
3-25
study this distinction in detail, since only juvenile increments are sufficiently wide to permit
the analysis of microincrements (Schöne et al., 2005b). The age at sexual maturity seems
to be quite variable. Mature specimens as young as three years (Rowell et al.,1990, Ropes
et al., 1984b) have been described, while immature clams can be as old as 14 years
(Thompson et al., 1980b).
Time of spawning. Studies of the gametogenic cycle in A. islandica show that the time of
spawning is very variable and that it may vary, not only between different regions, but also
from year to year in the same region. For sites on the Atlantic seaboard of North America,
periods during which large proportions of populations of A. islandica have been reported as
spawning (here taken to mean that they are in the partially spent stage of the gametogenic
cycle) are: July-October (Loosanoff 1953), August-November 1977 and November 1978 to
January 1979 (Jones 1981a), September 1978 to January 1979 and August-September
1979 (Mann 1982) and July 1983 to October 1984 (a total of 16 months of apparently
continuous spawning activity; Rowell et al., 1990). In other regions spawning activity has
been reported in December 1997 to March 1998 and July-November 1998 for northwest
Iceland (Thorarinsdóttir 2000) and in May-September for the Baltic Sea (von Oertzen 1972).
Time spent in larval and veliger stages. Lutz et al., (1982) studied the larval and post-larval
development in clams from Rhode Island and New Jersey (depths between 27-50 m) and
found a minimum time from fertilization to settlement of 32 days at 13 ºC, with a longer
period of 55 days required at temperatures between 8.5 ºC and 10.0 ºC. These periods, in
combination with the regional hydrographic regime and the availability of suitable substrates
for settlement act to constrain the spread and expansion of populations.





















![Sonata Arctica - Best [Bandscore]](https://img.pdfslide.net/doc/110x75/55cf9dfd550346d033b02b92/sonata-arctica-best-bandscore.jpg)







