Embed Size (px)
Citation preview

Distribution of Relaxin-3 and RXFP3 WithinArousal, Stress, Affective, and CognitiveCircuits of Mouse Brain
Craig M. Smith,1 Pei-Juan Shen,1 Avantika Banerjee,1 Pascal Bonaventure,2 Sherie Ma,1
Ross A.D. Bathgate,1 Steven W. Sutton,2 and Andrew L. Gundlach1*1Florey Neuroscience Institutes, The University of Melbourne, Victoria 3010, Australia2Neuroscience Drug Discovery, Johnson & Johnson Pharmaceutical Research & Development, San Diego, California 92121, USA
ABSTRACTRelaxin-3 (RLN3) and its native receptor, relaxin family
peptide 3 receptor (RXFP3), constitute a newly identi-
fied neuropeptide system enriched in mammalian brain.
The distribution of RLN3/RXFP3 networks in rat brain
and recent experimental studies suggest a role for this
system in modulation of arousal, stress, metabolism,
and cognition. In order to facilitate exploration of the
biology of RLN3/RXFP3 in complementary murine mod-
els, this study mapped the neuroanatomical distribution
of the RLN3/RXFP3 system in mouse brain. Adult, male
wildtype and RLN3 knock-out (KO)/LacZ knock-in (KI)
mice were used to map the central distribution of RLN3
gene expression and RLN3-like immunoreactivity (-LI).
The distribution of RXFP3 mRNA and protein was deter-
mined using [35S]-oligonucleotide probes and a radiola-
beled RXFP3-selective agonist ([125I]-R3/I5), respectively.
High densities of neurons expressing RLN3 mRNA,
RLN3-associated b-galactosidase activity and RLN3-LI
were detected in the nucleus incertus (or nucleus O),
while smaller populations of positive neurons were
observed in the pontine raphe, the periaqueductal gray
and a region adjacent to the lateral substantia nigra.
RLN3-LI was observed in nerve fibers/terminals in nu-
cleus incertus and broadly throughout the pons, mid-
brain, hypothalamus, thalamus, septum, hippocampus,
and neocortex, but was absent in RLN3 KO/LacZ KI
mice. This RLN3 neural network overlapped the regional
distribution of RXFP3 mRNA and [125I]-R3/I5 binding
sites in wildtype and RLN3 KO/LacZ KI mice. These
findings provide further evidence for the conserved na-
ture of RLN3/RXFP3 systems in mammalian brain and
the ability of RLN3/RXFP3 signaling to modulate ‘‘be-
havioral state’’ and an array of circuits involved in
arousal, stress responses, affective state, and cognition.
J. Comp. Neurol. 518:4016–4045, 2010.
VC 2010 Wiley-Liss, Inc.
INDEXING TERMS: amygdala; arousal; nucleus incertus; relaxin-3; RXFP3 (GPCR135); septohippocampal system; stress
Relaxin-3 (RLN3) is a newly identified member of the
relaxin/insulin-like family of peptides (Bathgate et al.,
2002) and is highly conserved across a range of species
from fish to mammals (Wilkinson et al., 2005). Identified
by our laboratory via its sequence homology to human
RLN1 and -2 (Bathgate et al., 2002), RLN3 has an insulin-
like structure, with 51 amino acid residues arranged into
two separate alpha helix chains, joined by two disulfide
bonds (Rosengren et al., 2006). RLN3 expression is
largely restricted to the brain (Bathgate et al., 2002), and
in the mouse and rat, RLN3 mRNA is highly enriched in
neurons of the midline, pontine nucleus incertus (NI) or
nucleus O (Bathgate et al., 2002; Burazin et al., 2002; Liu
et al., 2003b). In more recent studies in the rat, popula-
tions of RLN3 mRNA-positive cells were also detected in
the pontine raphe nucleus, the anterior, lateral, and
Additional Supporting Information may be found in the online versionof this article.
Grant sponsor: National Health and Medical Research Council ofAustralia; Grant numbers: 327404 (to C.M.S.), 520299 (to S.M.), 277609,509246 (to A.L.G.); Grant sponsors: Collaborative Research Agreementwith Johnson & Johnson PR&D, LLC, San Diego, CA, Howard FloreyBiomedical Foundation USA (to A.L.G.), The University of Melbourne –Melbourne (Postgraduate) Research Scholarship (to A.B.), ANZ TrusteesMedical Research & Technology (Victoria, Australia), Perpetual Trustees (toA.L.G., S.M.).
*CORRESPONDENCE TO: Andrew L. Gundlach, Florey NeuroscienceInstitutes, The University of Melbourne, Victoria 3010, Australia. E-mail:[email protected]
VC 2010 Wiley-Liss, Inc.
Received December 22, 2009; Revised April 29, 2010; Accepted June 1,2010
DOI 10.1002/cne.22442
Published online June 17, 2010 in Wiley InterScience (www.interscience.wiley.com)
4016 The Journal of Comparative Neurology | Research in Systems Neuroscience 518:4016–4045 (2010)
RESEARCH ARTICLE

ventrolateral periaqueductal gray, and in an area dorsal
to the lateral substantia nigra (Tanaka et al., 2005; Ma
et al., 2007).
Interest in the connectivity and neurochemistry of the
NI was raised by the discovery that NI neurons expressed
high levels of corticotrophin-releasing factor receptor-1
Abbreviations
n. Nucleusaca Anterior commissure, anteriorAcb Accumbens n.AD Anterodorsal thalamic n.AHi Amygdalohippocampal areaalv Alveus of the hippocampusAOM Anterior olfactory n., medialApir Amygdalopiriform transition areaAq AqueductAStr Amygdalostriatal transition areaAuC Auditory cortexB Barrington’s n.BLA Basolateral amygdalaBMA Basomedial amygdalaBNST Bed n. stria terminalisCA1-3 CA1-3 fields of hippocampusCB Cerebellumcc Corpus callosumCe(M) Central amygdala (medial part)CG Central grayCg(1/2) Cingulate cortex (area 1/2)Cl ClaustrumCM Centromedial thalamic n.cp Cerebral peduncle, basalCPu Caudate putamenD3V Dorsal 3rd ventricleDCIC Dorsal cortex, inferior colliculusDEn Dorsal endopiriform n.df Dorsal fornixDG Dentate gyrus hippocampusDLG Dorsal lateral geniculate n.DLPAG Dorsal lateral periaqueductal grayDMH Dorsomedial hypothalamic n.DMPAG Dorsal medial periaqueductal grayDpG Deep layer of the superior colliculusDR Dorsal raphe n.dSN Region dorsal to substantia nigraDT Dorsal terminal n. of the accessory optic tractDTg Dorsal tegmental n.DTT Dorsal tenia tectaec External capsulef Fornixfmj Forceps major corpus callosumGrO Granule cell layer olfactory bulbGrDG Granular layer, dentate gyrusgVIIn Genu of facial nerveHb HabenulaHDB N. horizontal limb diagonal bandHip Hippocampusic Internal capsuleIC Inferior colliculusIGL Intergeniculate leafletInG Intermediate gray layer of the superior colliculusInWh Intermediate white layer of the superior colliculusIO Inferior oliveIPN Interpeduncular n.IPDL Interpeduncular n., dorsolateralIPDM Interpeduncular n., dorsomedialIPL Interpeduncular n., lateralLa Lateral amygdalalat LateralLC Locus coeruleusLDTg Laterodorsal tegmental n.LEnt Lateral entorhinal cortexLH Lateral hypothalamic areaLM Lateral mammillary n.LPAG Lateral periaqueductal grayLPG Lateral parabrachial n.LPO Lateral preoptic area hypothalamusLPT Lateral posterior thalamic n.LS Lateral septumLSD Lateral septal n., dorsalLT Lateral terminal n. of the accessory optic tractLV Lateral ventricleM1 Primary motor cortexM2 Secondary motor cortexMe5 Mesencephalic trigeminal n.
MeA Medial amygdalaMEnt Medial entorhinal cortexmfb Medial forebrain bundleMGV Medial geniculate n., ventralml Medial lemniscusm/l Midlinemlf Medial longitudinal fasciculusMnR Median rapheMO Medial orbital cortexMol Molecular layer of the dentate gyrusMS Medial septummt Mammillothalamic tractMVePC Medial vestibular n. parvicellular partNI Nucleus incertusOp Optic nerve layer, superior colliculusopt Optic tractOr Oriens layer, hippocampusPAG Periaqueductal graypc Posterior commissurePe Periventricular hypothalamic n.PH Posterior hypothalamic areaPIL Posterior intralaminar thalamic n.Pir Piriform cortexpm Principal mammillary tractPnC Pontine reticular n., caudalPnR Pontine raphe n.PoDG Polymorph layer dentate gyrusPP Peripeduncular n.PPT Posterior pretectal n.PPTg Pedunculopontine tegmental n.Pr Prepositus hypoglossal n.PrL Prelimbic cortexPVA Paraventricular thalamic n., anteriorPVN Paraventricular hypothalamic n.Py Pyramidal cell layer, hippocampusRC Raphe capRe Reuniens thalamic n.RLi Rostral linear rapheRLN3 Relaxin-3RMg Raphe magnus n.RS Retrosplenial cortexRSA Retrosplenial agranular cortexRSG Retrosplenial granular cortexRXFP3 Relaxin family peptide 3 receptorS SubiculumS1 Primary somatosensory cortexS2 Secondary somatosensory cortexSC Superior colliculusSCh Suprachiasmatic n.SFi Septofimbrial n.SG Supragenual n.SHi Septohippocampal n.SI Substantia innominataSN Substantia nigrasol Solitary tractSON Supraoptic n.SPF Subparafascicular thalamic n.st Stria terminalisSuG Superficial gray layer, superior colliculusSuM Supramammillary n.SuML Supramammillary n. lateral partSuMM Supramammillary n. medial partsumx Supramammillary decussationTS Triangular septal n.V1 Primary visual cortexV2 Secondary visual cortexVDB N. vertical limb diagonal bandVEn Ventral endopiriform cortexVIIn Facial nerveVLG Ventral lateral geniculate n.VLPAG Ventral lateral periaqueductal grayVMH Ventromedial hypothalamic n.VTA Ventral tegmental areavtgx Ventral tegmental decussation peduncleZI Zona incerta3V Third ventricle4V Fourth ventricle
Relaxin-3/RXFP3 distribution in mouse brain
The Journal of Comparative Neurology | Research in Systems Neuroscience 4017

(CRF1) mRNA and were activated following intracerebro-
ventricular (i.c.v.) injections of CRF (Potter et al., 1994;
Bittencourt and Sawchenko, 2000). Immunohistochemi-
cal studies reported that NI neurons in the rat express
the GABA-synthesizing enzyme, glutamate decarboxylase
(Olucha-Bordonau et al., 2003), and that the majority, if
not all, RLN3 neurons in the NI are GAD-positive (Ma
et al., 2007). Other peptides detected in the rat NI (or
equivalent) include cholecystokinin (Olucha-Bordonau
et al., 2003), galanin, dynorphin (Sutin and Jacobowitz,
1988), neurotensin (Jennes et al., 1982), and a ranaten-
sin-like peptide (probably neuromedin-B) (Chronwall
et al., 1985). Dense networks of RLN3-immunoreactive
nerve fibers have been described in rat brain (Tanaka
et al., 2005; Ma et al., 2007) and electron microscopy
studies indicate that RLN3 is translated in the rough
endoplasmic reticulum, packaged into dense-core
vesicles and trafficked to presynaptic sites in nerve termi-
nals (Tanaka et al., 2005), consistent with a role as a
transmitter/modulator.
Anterograde and retrograde tract-tracing studies in the
rat demonstrated that NI neurons have strong reciprocal
connections with the median raphe (or central superior)
and interpeduncular nuclei, forming a likely behavioral
control network (Goto et al., 2001; Olucha-Bordonau
et al., 2003) that uses GABA and/or a range of peptides
including RLN3 to influence various neural networks
involved in behavioral activation. A heavy innervation of
the septohippocampal system by the NI suggested an
involvement in the control of hippocampal theta rhythm—
a process implicated in memory and attentional states,
such as exploratory behavior and rapid eye movement
(REM) sleep (Buzsaki, 2005; Hasselmo, 2005; Vertes,
2005). In support of this view, electrical stimulation of
the NI in anesthetized rats induced hippocampal theta
rhythm, while electrolytic lesion or pharmacological
blockade of the NI disrupted hippocampal theta induced
by stimulation of the reticularis pontis oralis (Nunez et al.,
2006), a pontine region known to be an endogenous ini-
tiator of hippocampal theta rhythm that is strongly inter-
connected with the NI (Teruel-Marti et al., 2008).
Molecular and pharmacological studies have demon-
strated that the native RLN3 receptor is the relaxin family
peptide 3 receptor (RXFP3; Bathgate et al., 2006), which
was formally known as G-protein-coupled receptor 135
(GPCR135; Liu et al., 2003a,b; Sutton et al., 2006), and
somatostatin- and angiotensin-like peptide receptor
(SALPR; Matsumoto et al., 2000). Activation of RXFP3
stably expressed in Chinese hamster ovary CHO-K1 or
human embryonic kidney HEK-293 cells results in an inhi-
bition of cAMP accumulation via Gi/o-protein coupling
(Liu et al., 2003b; Ortinau et al., 2005) and activation of
extracellular signal-regulated kinase 1/2 in these cells
and the mouse septal neuronal cell line SN-56 (van der
Westhuizen et al., 2007), but the precise nature of endog-
enous RXFP3 signaling in neurons or particular neural net-
works is currently unclear.
The distribution of RXFP3 mRNA and binding sites in
rat brain has been mapped using in situ hybridization his-
tochemistry and radioligand autoradiography, respec-
tively, with the latter studies employing a radiolabeled
chimeric peptide ([125I]-RLN3 B-chain/INSL5 A-chain
(R3/I5)) that is highly selective for RXFP3 (Sutton et al.,
2004; Liu et al., 2005; Ma et al., 2007). The receptor dis-
tribution in forebrain largely overlapped the RLN3 inner-
vation, and together with experimental studies in rats,
suggests that RXFP3 signaling is involved in the regula-
tion of circuits involved in stress responses and feeding
behavior. Expression of CRF1 by RLN3 NI neurons is re-
sponsible for the elevation in c-fos/Fos expression in
these cells after i.c.v. injection of CRF (Bittencourt and
Sawchenko, 2000; Tanaka et al., 2005), and increased
RLN3 mRNA after water-immersion restraint stress
(Tanaka et al., 2005) or repeat forced swim stress (Ban-
erjee et al., 2005, 2010). Furthermore, RLN3 injected
i.c.v. or into hypothalamic nuclei of satiated rats pro-
duced an increase in feeding during the first hour post-
injection, and local injection into the paraventricular
hypothalamic nucleus (PVN) twice a day for 7 days or
i.c.v. infusion for 14 days resulted in increased food
intake and a sustained body weight gain (McGowan et al.,
2005, 2006, 2007; Hida et al., 2006).
In contrast to these comprehensive anatomical and ini-
tial functional studies in the rat, only limited reports on
the murine RLN3/RXFP3 system have appeared. And yet
the mouse represents an important experimental spe-
cies, due to the ability of genetically modified strains to
provide functional insights. Earlier studies identified
expression of RLN3 in a region of mouse brain homolo-
gous to the NI (Bathgate et al., 2002), and provided a
broad overview of RXFP3 mRNA and binding site distri-
butions (Liu et al., 2003b, 2005; Boels et al., 2004;
Sutton et al., 2006), but in the present study we aimed
to expand knowledge of the central RLN3/RXFP3 net-
works in the mouse. We mapped the distribution of
RLN3-positive neurons through the rostral-caudal extent
of the NI and described RLN3-positive neurons in
regions outside the NI, and produced a comprehensive
map of RLN3-positive nerve fibers. We mapped the dis-
tribution of RXFP3 throughout the forebrain axis and
conducted a semiquantitative assessment of the relative
regional abundance of RLN3-like immunoreactivity (-LI),
RXFP3 mRNA, and RXFP3 binding sites throughout the
entire brain.
Adult male C57BL/6J and mixed background
129S5:B6 wildtype (WT) and RLN3 knock-out (KO)/LacZ
Smith et al.
4018 The Journal of Comparative Neurology |Research in Systems Neuroscience

knock-in (KI) mice were used to map: 1) the distribution
of RLN3 gene expression by histochemistry for native
mRNA and LacZ reporter-gene activity; 2) RLN3-LI, using
an affinity-purified polyclonal antiserum (AS-R385-101)
against a conserved region of the RLN3 C-peptide (Ma
et al., 2007); 3) RXFP3 mRNA, using multiple [35S]-oligo-
nucleotide probes; and 5) RXFP3 binding sites, using
[125I]-R3/I5 (Sutton et al., 2004; Liu et al., 2005). The dis-
tribution of RXFP3 mRNA was examined at the cellular
level using nuclear emulsion, and these results were com-
pared to data in the open-access Allen Brain Institute
Gene Expression Atlas obtained using nonradioactive
RNA probes (Lein et al., 2007). Additionally, the RXFP3
protein distribution revealed by [125I]-R3/I5 binding was
analyzed in WT and RLN3 KO/LacZ KI mice. Preliminary
accounts of some of these data have appeared in
abstract form (Smith et al., 2006a,b) and in a recent con-
ference proceedings (Smith et al., 2009b).
MATERIALS AND METHODS
AnimalsAll procedures described were undertaken with the ap-
proval of the Howard Florey Institute Animal Welfare
Committee and in strict compliance with the ethical
guidelines issued by the National Health and Medical
Research Council of Australia. All efforts were made to
minimize the number of mice used and their discomfort.
Adult C57BL/6J male mice for the immunostaining and in
situ hybridization studies (18–25 g) were obtained from
the Australian Research Centre (Canning Vale, WA, Aus-
tralia). Mixed background (C57BL/6J � 129SV) RLN3
KO/LacZ KI mice, which express the LacZ reporter-gene
downstream, in-frame, and under the control of the en-
dogenous RLN3 promoter, were generated by Lexicon
Genetics (The Woodlands, TX). Adult (18–25 g) RLN3
KO/LacZ KI mice and WT littermates were generated via
heterozygous pairings and housed in the Howard Florey
Institute Animal Facilities. Mice were maintained on a
12:12-hour light dark cycle, with 3–4 mice per cage and
ad libitum access to standard chow and water.
Mouse genotypingWT (RLN3 (þ/þ)), heterozygous (RLN3 (þ/�)), and
KO (RLN3 (�/�)) mice were identified by polymerase
chain reaction (PCR) analysis of DNA extracted from tail
samples taken from each mouse using a REDExtract-N-
Amp Tissue PCR Kit (Sigma-Aldrich, Castle Hill, NSW, Aus-
tralia) following the manufacturer’s protocols. In the
RLN3 KO/LacZ KI mice used in this study, ablation of
functional RLN3 was achieved by deletion of exon 2 and
replacement with a LacZ/Neo cassette using targeting
vectors. For PCR identification of genotypes, a ‘‘forward’’
primer (which binds to an endogenous region upstream of
the LacZ/Neo cassette insertion site) was combined with
two downstream ‘‘reverse’’ primers, which bind to a
region within the endogenous WT genome that is deleted
in the KO allele (Reverse-WT primer), or to a region within
the LacZ/Neo cassette (Reverse-KO primer). The pres-
ence of a WT or KO allele resulted in the amplification of
a 621-bp or 281-bp product, respectively, which were
separated via gel electrophoresis. The sequences of pri-
mers used were as follows: Forward: 50-GGTTGGCAAGTAGTGTATGC-30; Reverse-WT: 50-TCTGATTTAGGGAGCCTAGC-30; Reverse-KO: 50-GCAGCGCATCGCCTTCTATC-30.The PCR reaction was conducted following the manufac-
turer’s protocols, using 50 pmol of forward primer and 25
pmol of each reverse primer, in a 20-lL reaction volume.
Amplification was conducted under the following condi-
tions: 94�C for 5 minutes; 35 cycles of 94�C for 40 sec-
onds, 58�C for 40 seconds, and 72�C for 2 minutes; fol-
lowed by a final 72�C for 10 minutes, and 4�C indefinitely.
Tissue preparationFor immuno- and X-GAL-histochemistry, mice were
killed by isoflurane inhalation overdose (IsoFLO; Abbott
Laboratories, Melbourne, VIC, Australia) and perfused
transcardially with 10 mL ice-cold phosphate buffer (PB)
solution (2.7 mM KCl, 11.2 mM Na2HPO4, 1.8 mM
KH2PO4, pH 7.4) followed by 50 mL ice-cold 4% parafor-
maldehyde in PB (pH 7.4) and then decapitated. Brains
were removed from the skull and immersed in fixative for
1 hour at 4�C before cryoprotection in 20% sucrose in PB
overnight at 4�C, coated in OCT embedding medium (Tis-
sue-Tek, Torrance, CA), and stored at �80�C. For in situ
hybridization and [125I]-R3/I5 binding assays, mice were
killed by isoflurane overdose and then decapitated.
Brains were quickly removed and embedded in OCT (Tis-
sue-Tek), frozen over dry ice, and stored at �80�C.
Antibody productionThe RLN3 antiserum used for mapping RLN3-LI was
raised against residues 85–101 of the C-chain of the
RLN3 propeptide that is identical in mouse and human,
with one conservative amino acid change from the rat
sequence (Bathgate et al., 2002; Burazin et al., 2002); it
has been characterized by our laboratory in studies of rat
and nonhuman primate brain (Ma et al., 2007, 2009b).
Thus, although this region of the RLN3 propeptide is pre-
dicted to be proteolytically cleaved during the processing
of mature RLN3, recent anatomical data suggest that the
distribution of RLN3-LI observed with this AS-R385-101C-chain antiserum is representative of the distribution of
mature RLN3 peptide (see Results) (Tanaka et al., 2005;
Ma et al., 2007). The specificity of the staining achieved
using affinity-purified AS-R385-101 has been demonstrated
Relaxin-3/RXFP3 distribution in mouse brain
The Journal of Comparative Neurology | Research in Systems Neuroscience 4019

in rats, where either omission of the primary antibody, use
of the preimmune serum, or preabsorption of the primary
antibody with 1 lg/mL pro-RLN3 C-peptide completely
abolished specific immunostaining of fibers/terminals (Ma
et al., 2007). In this study the specificity of this AS-R385-101antiserum was further validated by incubation with sections
through the medial septum of WT and RLN3 KO/LacZ KI
mice. These studies consistently resulted in strong immuno-
staining in the WT midline septal region and a total absence
of specific staining in equivalent sections from the RLN3 KO
mouse brain (Supporting Information Fig. S1).
ImmunohistochemistryCoronal sections (40 lm) through the rostrocaudal
brain axis of six brains were cut on a cryostat at �18�C(Cryocut 1800, Leica Microsystems, Heerbrugg, Switzer-
land) and collected in PB, pH 7.4. Free-floating sections
were preincubated in 70% methanol containing 0.6%
H2O2 for 30 minutes with gentle shaking, rinsed 3 � 10
minutes in PB, and then incubated in blocking buffer (10%
(v/v) normal horse serum and 0.3% (v/v) Triton X-100 in
PB) for 1 hour with shaking at room temperature. Sec-
tions were then incubated in PB containing polyclonal AS-
R385-101 RLN3 antiserum (dilution 1:2,500), 2% NHS,
0.3% Triton X-100, and 0.1% NaN3 (pH 7.4) with gentle
agitation for 72 hours at 4�C. Sections were washed 3 �10 minutes in PB and then incubated in PB containing bio-
tinylated antirabbit IgG (dilution 1:500, Vector Laborato-
ries, Burlingame, CA) with constant agitation for 1 hour at
room temperature, followed by 3 � 10 minute washes.
Sections were incubated in a 1:100 ABC/horseradish
peroxidase complex solution (Vectastain Elite, Vector
Laboratories) in PB for 1 hour at room temperature, fol-
lowed by 3 � 10 minute washes. Immunostaining was
visualized using a peroxidase-chromogen reaction by
placing the sections in 0.5 mg/mL 3, 30-diaminobenzidine
(DAB, Sigma-Aldrich) in PB containing 0.06% H2O2 for 1
minute before being washed for 3 � 10 minutes and
mounted onto gelatin-chrom alum-coated glass micro-
scope slides and dried overnight. Sections were then
dehydrated and cleared in ethanol and xylene and cover-
slipped using DePeX mounting media (Sigma-Aldrich).
X-GAL detection of LacZ reporter geneexpression in RLN3 KO/LacZ KI mice
Free-floating coronal sections (40 lm) from nine RLN3
KO/LacZ KI brains were incubated overnight at 37�C in
0.1% 4-chloro-5-bromo-3-indolyl-b-D-galactopyranoside(X-GAL, SciMAR, Templestowe, VIC, Australia), 2 mM
MgCl2, 5 mM EGTA, 0.01% (w/v) sodium deoxycholate,
0.02% (w/v) Nonidet P-40, 5 mM K3Fe(CN)6, and 5 mM
K4Fe(CN)6.6H2O in 0.1 M PB, pH 7.4. In brain regions
where the LacZ reporter gene was expressed under the
control of the endogenous RLN3 promoter, colorless solu-
ble X-GAL substrate was converted into a blue precipitate
that was visible in the neuronal soma and proximal proc-
esses. Sections were rinsed for 3 � 10 minutes in PB, pH
7.4, and mounted on 0.5% gelatin-chrom alum-coated
glass microscope slides, left to dry overnight, and then
counterstained in 0.01% neutral red solution for 2
minutes. After drying overnight, slides were dehydrated in
an ethanol series, cleared in xylene, and coverslipped
with DePeX mounting media (Sigma-Aldrich).
In situ hybridization histochemistryIn the current study, RLN3 mRNA was detected using
three DNA oligonucleotides complementary to nonoverlap-
ping regions of the mouse RLN3 sequence (50-GCACATCCGAATGAATCCGTCCATCCACTCCTCCGAGAC-30; 50-CAAGCAGAGCTGGCTCCTCCTGGCTCAAAGCCAATCTTC-30; 50-GTTGTAGCTCTGGGAGCGAGGCCTGAGCCTCAGACAGTA-30;NCBI Accession No. NM_173184). For detection of
RXFP3 mRNA, nine oligonucleotides complementary to
nucleotides of mouse RXFP3 cDNA were used (50-CAGTGAGTTGCCAGCGTTTGATGAGTTACATTGGCACCC-30; 50-GAAAACAGATCGTGGACATCTGTGTCTCGCTCTGACTGC-30;50-AGCAGGTGCCTGAGCTTTCTATGTAGACGCTCAAAAGGG-30; 50-GGCACAAACCACCCAGTAAACCGCGCTGATGAGG ATCCG-30; 50-GATGATGCTCAGCGGCAGCAGGAAGCCCAGCAGCACCTT-30; 50-CCAGGCGCTGCTGCTGCTCCTACTGCATCTGTTGTCCCA-30; 50-GCAGTAGAGGATCGG GTTGA
GGCAGCTGTTGGAGTGCGC-30; 50-CTTGGTGGTGGCGGTGAAAGGGCGCATGTTGGTGAGCGA-30; 50-AGCGCAGCCGAAGCCCAGATCAACCCACACAGCACCTTG-30; Accession
No. NM_178717). Although single oligonucleotide probes
are routinely used to detect abundant transcripts, exten-
sive experience within our laboratory (e.g., Ryan and
Gundlach, 1995), and other reports (e.g., Trembleau and
Bloom, 1995; Broide et al., 2004) demonstrate that multi-
ple oligonucleotide probes can be used to effectively
amplify the hybridization signal, with comparable or
improved specificity. Indeed, amplification is often essen-
tial for the successful detection of rare receptor tran-
scripts (e.g., Burazin et al., 2000).
Probes were produced commercially (Sigma-Aldrich)
and 30-end-labeled with [35S]-dATP (Amersham Life Sci-
ences, Amersham, UK) to a specific activity of �1–2 �109 dpm/lg, as described previously (Burazin et al.,
2002; Wisden and Morris, 2002). Using the GenBank
database, the sequence of each oligonucleotide was
checked for 100% homology to the target gene, less than
70% homology with other mammalian genes, and low sec-
ondary structure. Labeled probes were diluted in hybridiza-
tion buffer (1–5 pg/lL per probe) consisting of 50% (v/v)
formamide, 10% (v/v) dextran sulfate in 4 � SSC (1 �
Smith et al.
4020 The Journal of Comparative Neurology |Research in Systems Neuroscience

SSC: 0.15 M NaCl, 15 mM sodium citrate, pH 7.0). Dithio-
threitol (200 mM) was added to the solution to minimize di-
sulfide bridge formation and ‘‘nonspecific hybridization.’’
Brain sections from four mice were dehydrated in etha-
nol (70–100%) and delipidated in chloroform for 10
minutes to help decrease nonspecific ‘‘myelin binding’’ of
oligonucleotides during hybridization, and rinsed in 100%
ethanol (Burazin et al., 2002). Sections were then incu-
bated for 16 hours in hybridization buffer containing la-
beled probes at 42�C. Specificity of hybridization was
assessed in a 1:5 series of sections by the addition of a
100-fold excess of unlabeled oligonucleotides to the
hybridization buffer. In all experiments, specific hybridiza-
tion was successfully displaced by this procedure (data
not shown). Labeled brain sections were coated with nu-
clear emulsion (Ilford K5, diluted 1:1 with dH2O; Ilford
Imaging, Melbourne, VIC, Australia) and exposed for 48
hours (RLN3 mRNA) or 20 weeks (RXFP3 mRNA) prior to
development, counterstaining with 0.01% thionin, and
analysis using brightfield and darkfield microscopy. Some
sections were exposed to Kodak BioMax film (Integrated
Sciences, Sydney, NSW, Australia) for 5 weeks for detec-
tion of RXFP3 mRNA.
Radioligand binding autoradiography[125I]-R3/I5 binding to mouse brain sections was
assessed as described (Sutton et al., 2004, 2006; Ma
et al., 2007). Coronal sections (14 lm) from three WT
and three RLN3 KO mice on gelatin-chrom alum-coated
glass microscope slides were preincubated for 15
minutes at room temperature in incubation buffer (20
mM HEPES, pH 7.4, 120 mM NaCl2, 0.22 mM KH2PO4,
1.3 mM CaCl2, 0.8 mM MgSO4). Sections were dried and
incubated for 60 minutes with 7 pM [125I]-R3/I5 (specific
activity, 2,200 Ci/mmol) in incubation buffer containing
0.5% bovine serum albumin and a protease inhibitor cock-
tail (Sigma-Aldrich). Nonspecific binding of [125I]-R3/I5
was determined in the presence of 1 lM unlabeled RLN3.
Following incubation, slides were washed in ice-cold incu-
bation buffer for 3 � 10 minutes and rinsed in ice-cold
deionized water. Sections were allowed to air-dry over-
night and were apposed to Kodak Biomax film (Integrated
Sciences) for 2 weeks prior to development and fixation
in an automated processor.
Analysis of relative density of RLN3-LI,RXFP3 mRNA, and [125I]-R3/I5 binding sites
The relative density of RLN3-LI in different areas was
scored by visual inspection of sections from six brains
according to a 6-point scale: (�) no RLN3-LI detected;
(þ/�) infrequent but detectable RLN3-LI; (þ) low density
of scattered RLN3-LI in the region; (þþ) moderate;
(þþþ) high; and (þþþþ) very high density of staining in
a distinct nucleus/region. For example, RLN3-LI density
was very high in the medial septum, high in the cingulate
cortex, moderate in the periaqueductal gray, and low in
the claustrum (see Results).
The distribution of neurons expressing RXFP3 was
mapped using data from three independent in situ hybrid-
ization studies: 1) radiolabeled riboprobe studies, where
signal intensity was detected using x-ray film (preliminary
data from these studies has been previously reported;
Sutton et al., 2006); 2) radiolabeled oligonucleotides
studies, where the regional density of positive neurons
and intensity of silver grain accumulation over individual
neurons was detected using nuclear photographic emul-
sion; and 3) nonradioactive riboprobe studies, where data
are available in the Allen Brain Atlas (Lein et al., 2007)
(www.brainmap.org). The distribution of RXFP3 mRNA
was first mapped separately using data from each of
these three studies, which revealed overlapping distribu-
tion patterns and few notable discrepancies. Therefore,
for the majority of coronal brain levels data from these
three separate studies were available, which was aver-
aged to produce the data presented.
Hybridization density within an area was rated inde-
pendent of neuronal size, i.e., a region with a moderate
number of large RXFP3-positive cells scored a similar
density value to a region with smaller yet more numerous
RXFP3-positive cells and a region with a high number of
large cells each with relatively reduced signal intensity.
The relative density of [125I]-R3/I5 binding sites was
scored by visual analysis of sections from six brains col-
lected in the present study and images from an earlier
study (Sutton et al., 2006). RXFP3 mRNA and [125I]-R3/I5
binding site densities were scored using a 6-point scale:
(�) no RXFP3 mRNA or binding sites detected; (þ/�)
scarce, (þ) low, (þþ) moderate, (þþþ) high or (þþþþ)
very high density of RXFP3 mRNA or binding sites in a dis-
tinct nucleus/region. For example, the RXFP3 mRNA and
binding site density was very high in the medial nucleus
of the amygdala, high in the bed nucleus of the stria ter-
minalis, moderate in the nucleus incertus, and low in the
auditory cortex (see Results).
Photography and image productionBrightfield and darkfield digital images of RLN3 immu-
nostaining and RXFP3 mRNA-associated nuclear emul-
sion autoradiograms were captured using MCID-M2 soft-
ware (Imaging Research, St Catharine’s, ON, Canada) on
a Nikon Microphot SA microscope (FSE, Melbourne, VIC,
Australia) equipped with a Sony CCD camera. X-ray film
images were viewed using a Sony XC-77 camera mounted
above a light-box (Northern Lights; Berthold Australia,
Bundoora, VIC, Australia). All illustrative images were
Relaxin-3/RXFP3 distribution in mouse brain
The Journal of Comparative Neurology | Research in Systems Neuroscience 4021

archived as high-resolution images (at least 500 dpi) and
were cropped, adjusted for contrast, had obvious arti-
facts removed, and were arranged and labeled using
Adobe Photoshop 7.0 and Adobe Illustrator 10 for Win-
dows (Adobe Systems, San Jose, CA). Schematics were
created using Allen Brain Atlas software (Dong, 2008) to
illustrate the RLN3 neuron distribution in the NI and the
Mouse Brain Atlas software (Franklin and Paxinos, 1997)
to illustrate the distribution of RLN3-LI, RXFP3 mRNA and
[125I]-R3/I5 binding sites, using Adobe Photoshop 7 and
Illustrator 10.
RESULTS
The present study identified an enrichment of RLN3
mRNA and RLN3-LI in the NI of mouse brain (Fig. 1), simi-
lar to observations in the rat (Burazin et al., 2002; Tanaka
et al., 2005; Ma et al., 2007). High levels of RLN3 gene
expression were reflected by the detection of high levels
of mRNA by in situ hybridization of [35S]-oligonucleotides
and high levels of X-GAL staining, reflecting activity of the
LacZ reporter gene in RLN3 KO/LacZ KI mice (Fig. 2).
RLN3-LI was not detectable in RLN3 KO mice (Supporting
Information Fig. S1), but was present in neuronal cell
bodies within the NI in WT mice (Fig. 3). RLN3 was also
expressed by a number of small populations of neurons in
the periaqueductal gray, in a midbrain area dorsal to the
substantia nigra, and in the pontine raphe nucleus (Fig. 4).
RLN3-LI was observed in extensive networks of axons and
nerve terminals throughout the forebrain (Figs. 5–9) in a
pattern that overlapped the distribution of RXFP3 mRNA
(Figs. 5, 10, 11) and [125I]-R3/I5 binding sites (Figs. 5, 12);
again paralleling data from the rat (Sutton et al., 2004;
Tanaka et al., 2005; Ma et al., 2007). Further detailed
descriptions and semiquantitative assessments of these
findings are provided below (see Fig. 5; Table 1).
Distribution of RLN3 neurons in C57BL/6Jand RLN3 KO/LacZ KI mouse brain
RLN3-positive neurons were identified using three
methods. First, in situ hybridization was used to directly
detect RLN3 mRNA. The short 2-day exposure time to nu-
clear emulsion required to achieve a photographic image
indicates that RLN3 mRNA is highly abundant. Second,
RLN3 expression was detected indirectly via LacZ re-
porter gene activity, as a blue X-GAL associated precipi-
tate in the soma and proximal processes of NI neurons in
RLN3 KO/LacZ KI brain sections. Notably, X-GAL staining
offers advantages over in situ hybridization, such as
increased morphological detail and the ability to work
without any potential radioactive hazard. The observed
overlap in the distribution of RLN3 mRNA and reporter
gene activity suggests that X-GAL staining faithfully iden-
tifies neurons that normally express RLN3 in WT mice.
Lastly, immunohistochemistry using a polyclonal RLN3
antiserum detected RLN3-LI in cytoplasm and proximal
processes of NI neurons. The RLN3-LI present in NI soma
was of sufficient abundance to be detected without pre-
treatment of the mice with colchicine to enhance the cel-
lular accumulation of peptide (Ma et al., 2007). All three
techniques revealed overlapping distributions and pro-
vide an accurate description of the distribution of RLN3-
positive neurons. This distribution of RLN3-positive neu-
rons was summarized on Allen Brain Atlas schematic
images (Dong, 2008), as this atlas provides several plates
within the restricted rostral-caudal extent of the NI (Fig. 1).
Therefore, in these plates the borders of the NI illustrated
are those provided in the Allen Brain Atlas, but we have
noted a possible lack of consensus on whether the most
midline of cells within these borders are part of the NI or
part of the central gray. For example, Franklin and Paxi-
nos (1997) do not include cells along the midline in the
nucleus O (or NI) and several reports in the rat are con-
sistent with the bilateral nature of the NI (Goto et al.,
2001; Olucha-Bordonau et al., 2003; Tanaka et al., 2005;
Ma et al., 2007). Furthermore, several other genes
enriched in the mouse NI also display a bilateral appear-
ance (see, e.g., CRF binding protein and E430002G05Rik;
www.brainmap.org; Lein et al., 2007).
RLN3-positive neurons in the NIThe majority of RLN3-positive neurons were observed
within the NI, located medioventral to the locus coeruleus
and along the medial and ventral border of the dorsal teg-
mental nucleus (Fig. 1). At the rostral border of the mouse
NI defined in the Allen Brain Atlas (bregma �5.25 mm), a
high density of RLN3-positive neurons was present bilat-
erally in the ventral part of the nucleus, near the midline
and just dorsal to the medial longitudinal fasciculus, while
RLN3 neurons in more lateral and dorsal regions were
less tightly packed (Figs. 1A, 2). Neurons along the adja-
cent midline region were RLN3-negative. More caudally, a
high density of RLN3-positive cells was observed through-
out the extent of the NI (bregma �5.38 mm; Figs. 1B, 3),
with negative neurons present along the midline. Addi-
tionally, a low density of positive cells was observed at
the border of the NI and the pontine central gray (Dong,
2008).
The distribution of RLN3-positive neurons further cau-
dally (bregma �5.45 mm; Fig. 1C) was reminiscent of the
characteristic topography described in the rat, where the
NI consists of a tightly clustered and medial pars com-
pacta and a more lateral and diffuse pars dissipata
(Tanaka et al., 2005; Ma et al., 2007). At the caudal
boundary of the NI (bregma �5.55 mm), two diffuse pop-
ulations of RLN3-positive neurons were observed—a
Smith et al.
4022 The Journal of Comparative Neurology |Research in Systems Neuroscience
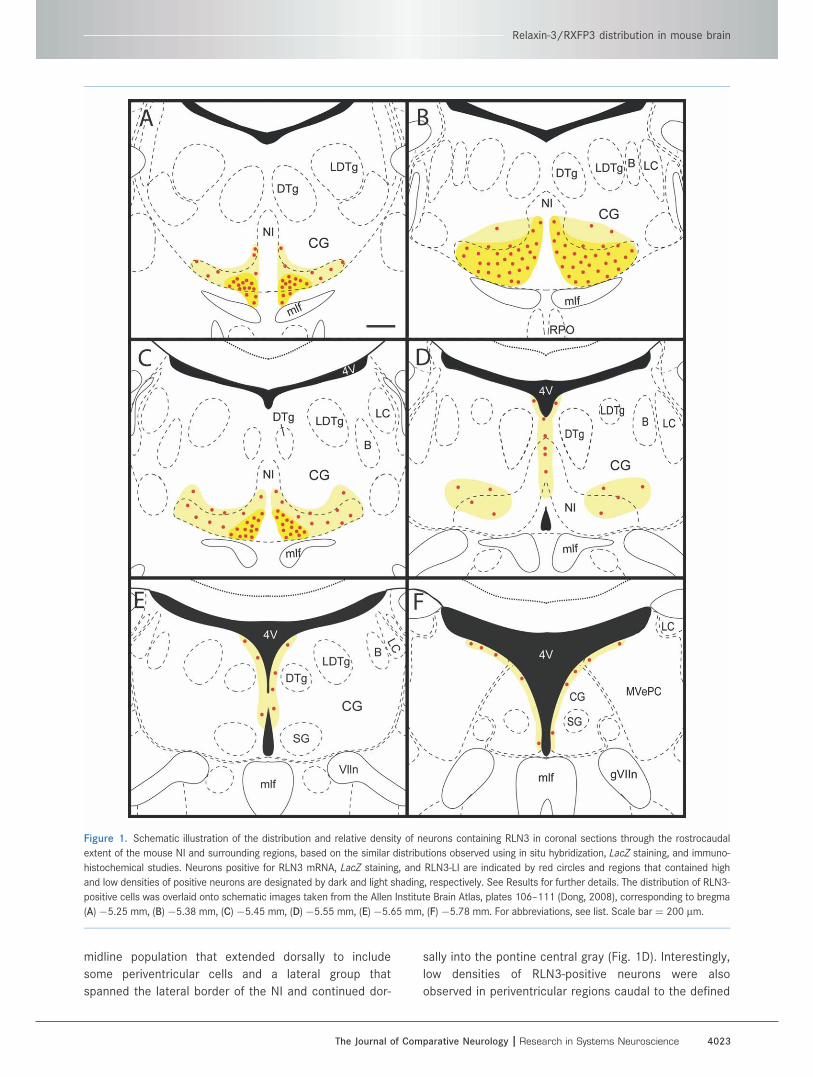
midline population that extended dorsally to include
some periventricular cells and a lateral group that
spanned the lateral border of the NI and continued dor-
sally into the pontine central gray (Fig. 1D). Interestingly,
low densities of RLN3-positive neurons were also
observed in periventricular regions caudal to the defined
Figure 1. Schematic illustration of the distribution and relative density of neurons containing RLN3 in coronal sections through the rostrocaudal
extent of the mouse NI and surrounding regions, based on the similar distributions observed using in situ hybridization, LacZ staining, and immuno-
histochemical studies. Neurons positive for RLN3 mRNA, LacZ staining, and RLN3-LI are indicated by red circles and regions that contained high
and low densities of positive neurons are designated by dark and light shading, respectively. See Results for further details. The distribution of RLN3-
positive cells was overlaid onto schematic images taken from the Allen Institute Brain Atlas, plates 106–111 (Dong, 2008), corresponding to bregma
(A)�5.25 mm, (B) �5.38 mm, (C) �5.45 mm, (D) �5.55 mm, (E)�5.65 mm, (F) �5.78 mm. For abbreviations, see list. Scale bar¼ 200 lm.
Relaxin-3/RXFP3 distribution in mouse brain
The Journal of Comparative Neurology | Research in Systems Neuroscience 4023
NI in the Allen Atlas (bregma �5.65 and �5.78 mm; Fig.
1E,F) (Dong, 2008).
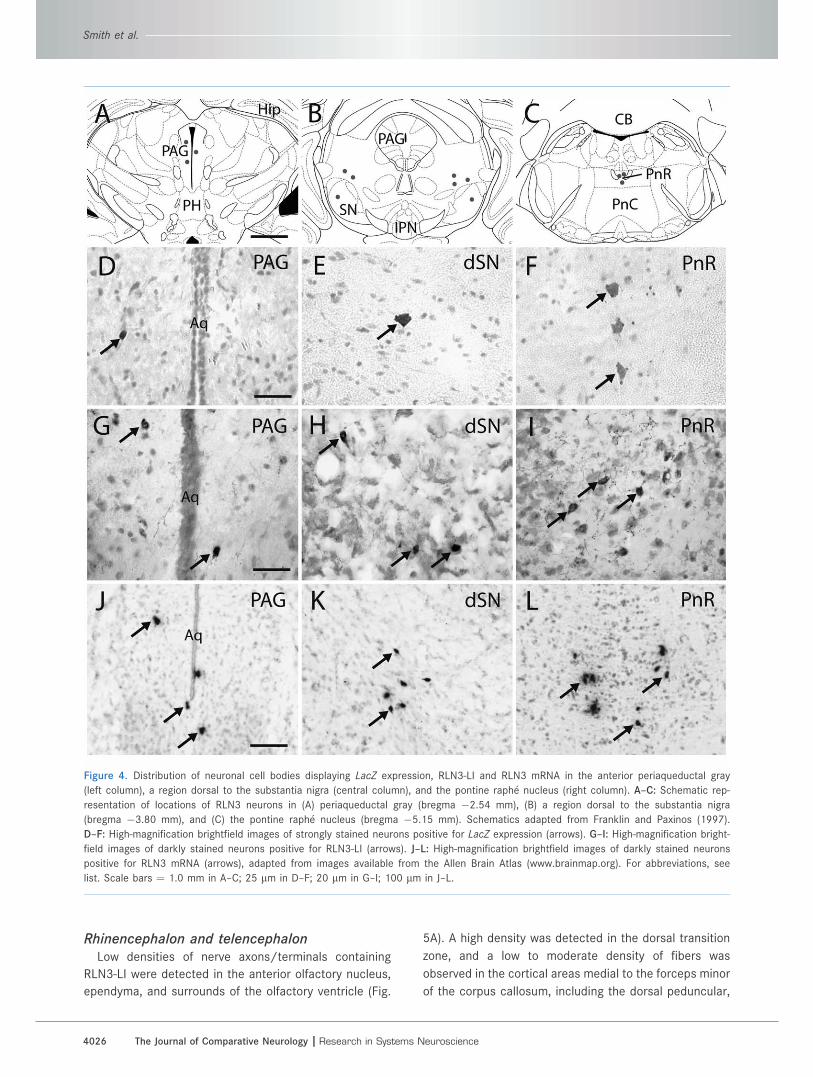
RLN3-positive neurons outside the NISeveral small populations of RLN3-positive neurons
were detected in the anterior periaqueductal gray, in the
pontine raphe nucleus, and in a region dorsal to the lat-
eral substantia nigra, and coinciding with or adjacent to
the lateral terminal and peripeduncular nuclei (Fig. 4A–
C). This distribution was observed with X-GAL staining of
RLN3 KO/LacZ KI brain sections (Fig. 4D–F), as RLN-LI
present in sections from C57BL/6J mice (Fig. 4G–I), and
was also observed in adult C57BL/6J mouse brain sec-
tions hybridized with a digoxigenin-labeled riboprobe
targeting RLN3 mRNA and detected using alkaline phos-
phatase (Fig. 4J–L; see Allen Brain Atlas; www.brainma-
p.org) (Gundlach et al., 2009). The authenticity of these
highly correlated findings is further supported by similar
observations in the rat (Tanaka et al., 2005; Ma et al.,
2007).
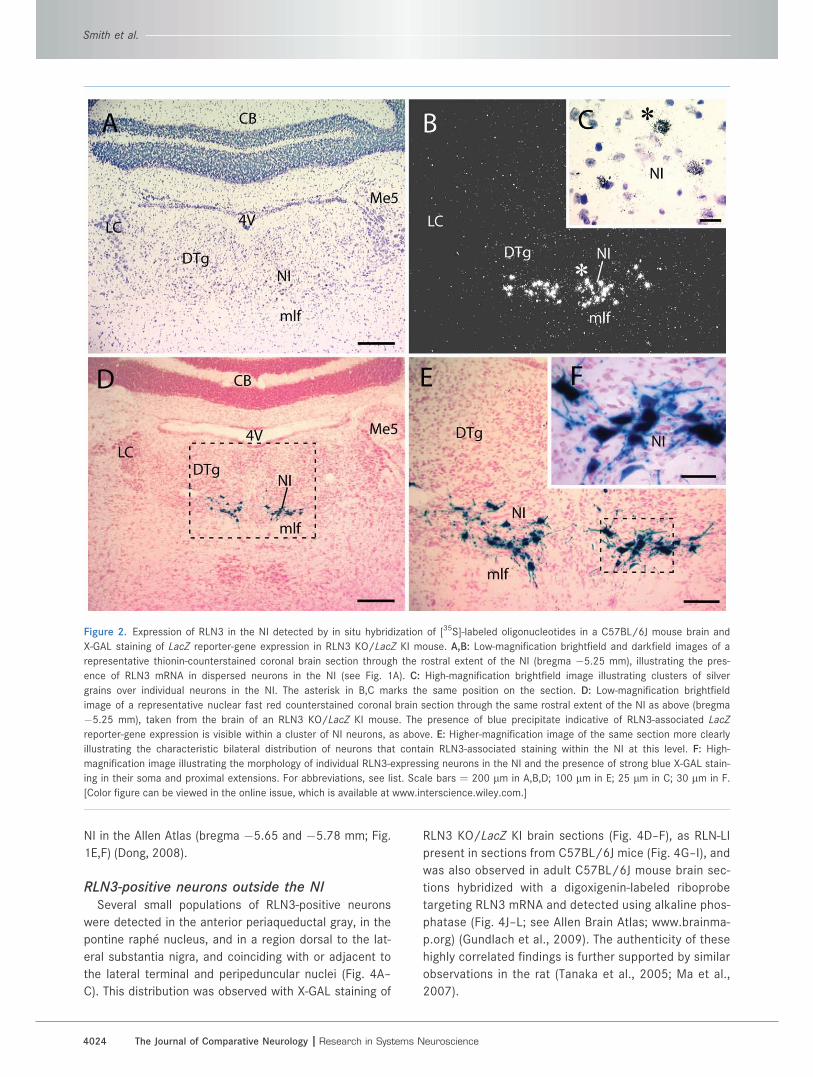
Figure 2. Expression of RLN3 in the NI detected by in situ hybridization of [35S]-labeled oligonucleotides in a C57BL/6J mouse brain and
X-GAL staining of LacZ reporter-gene expression in RLN3 KO/LacZ KI mouse. A,B: Low-magnification brightfield and darkfield images of a
representative thionin-counterstained coronal brain section through the rostral extent of the NI (bregma �5.25 mm), illustrating the pres-
ence of RLN3 mRNA in dispersed neurons in the NI (see Fig. 1A). C: High-magnification brightfield image illustrating clusters of silver
grains over individual neurons in the NI. The asterisk in B,C marks the same position on the section. D: Low-magnification brightfield
image of a representative nuclear fast red counterstained coronal brain section through the same rostral extent of the NI as above (bregma
�5.25 mm), taken from the brain of an RLN3 KO/LacZ KI mouse. The presence of blue precipitate indicative of RLN3-associated LacZ
reporter-gene expression is visible within a cluster of NI neurons, as above. E: Higher-magnification image of the same section more clearly
illustrating the characteristic bilateral distribution of neurons that contain RLN3-associated staining within the NI at this level. F: High-
magnification image illustrating the morphology of individual RLN3-expressing neurons in the NI and the presence of strong blue X-GAL stain-
ing in their soma and proximal extensions. For abbreviations, see list. Scale bars ¼ 200 lm in A,B,D; 100 lm in E; 25 lm in C; 30 lm in F.
[Color figure can be viewed in the online issue, which is available at www.interscience.wiley.com.]
Smith et al.
4024 The Journal of Comparative Neurology |Research in Systems Neuroscience

Specificity of RLN3 antiserum andverification of RLN3 deficiencyin KO mice
Brain sections from RLN3 KO/LacZ KI mice did not dis-
play any RLN3-LI in regions such as the medial septum
following incubation with the characterized polyclonal
RLN3 antiserum, despite high levels observed in equiva-
lent sections from WT mice (Supporting Information
Fig. S1). This finding strongly supports the specificity of
the antiserum for RLN3 (Saper and Sawchenko, 2003),
and is in line with our earlier reports in the rat and maca-
que brain (Ma et al., 2007, 2009b). The data also confirm
that RLN3 is indeed deficient in this strain of mice.
Distribution of RLN3-LI in neuronalprojections of mouse brain
Punctate RLN3-LI staining characteristic of nerve fibers
and terminal boutons was broadly distributed throughout
the mouse brain, in line with the trafficking, targeting, and
storage of RLN3 observed in the rat (Tanaka et al., 2005).
The distribution of RLN3-LI observed in six brains was col-
lated and plotted onto schematic brain sections (Franklin
and Paxinos, 1997; Fig. 5), and the relative density of
RLN3-LI in different brain regions was semiquantitatively
scored (Table 1). Descriptions of the topography of RLN3-
LI throughout the neuraxis follow, with reference to sum-
mary data and relevant micrographs (Figs. 6–9).
Figure 3. RLN3-LI in the NI of a C57BL/6J mouse brain labeled by a polyclonal C-peptide antiserum (AS-R385-101) and visualized by DAB
immunohistochemistry. A,B: Low- and high-magnification brightfield images of a representative coronal brain section through the ‘‘central’’
region of the NI (bregma �5.38 mm), illustrating the presence of RLN3-LI in dispersed neurons and their proximal processes in the NI
(see Fig. 1B). C,D: Darkfield images of the same section, illustrating the distribution of nerve fibers/terminals that contain RLN3-LI within
the NI region in addition to the nearby dorsal tegmental nucleus and area adjacent to the fourth ventricle. The region shown in B,D is out-
lined in A,C. For abbreviations, see list. Scale bars ¼ 100 lm in A,C; 50 lm in B,D. [Color figure can be viewed in the online issue, which
is available at www.interscience.wiley.com.]
Relaxin-3/RXFP3 distribution in mouse brain
The Journal of Comparative Neurology | Research in Systems Neuroscience 4025
Rhinencephalon and telencephalonLow densities of nerve axons/terminals containing
RLN3-LI were detected in the anterior olfactory nucleus,
ependyma, and surrounds of the olfactory ventricle (Fig.
5A). A high density was detected in the dorsal transition
zone, and a low to moderate density of fibers was
observed in the cortical areas medial to the forceps minor
of the corpus callosum, including the dorsal peduncular,
Figure 4. Distribution of neuronal cell bodies displaying LacZ expression, RLN3-LI and RLN3 mRNA in the anterior periaqueductal gray
(left column), a region dorsal to the substantia nigra (central column), and the pontine raphe nucleus (right column). A–C: Schematic rep-
resentation of locations of RLN3 neurons in (A) periaqueductal gray (bregma �2.54 mm), (B) a region dorsal to the substantia nigra
(bregma �3.80 mm), and (C) the pontine raphe nucleus (bregma �5.15 mm). Schematics adapted from Franklin and Paxinos (1997).
D–F: High-magnification brightfield images of strongly stained neurons positive for LacZ expression (arrows). G–I: High-magnification bright-
field images of darkly stained neurons positive for RLN3-LI (arrows). J–L: High-magnification brightfield images of darkly stained neurons
positive for RLN3 mRNA (arrows), adapted from images available from the Allen Brain Atlas (www.brainmap.org). For abbreviations, see
list. Scale bars ¼ 1.0 mm in A–C; 25 lm in D–F; 20 lm in G–I; 100 lm in J–L.
Smith et al.
4026 The Journal of Comparative Neurology |Research in Systems Neuroscience

Figure 5. Schematic illustration of the distribution and relative densities of RLN3-LI, RXFP3 mRNA, and RXFP3 binding sites in coronal
sections through the rostrocaudal extent of the C57BL/6J mouse brain. A–P: A series of coronal drawings, adapted from the stereotaxic
atlas of (Paxinos and Watson, 2007), illustrating the distribution and relative density of RLN3-LI (fine red lines), RXFP3 mRNA (blue dots),
and [125I]-R3/I5 binding sites (green areas, with light shading indicating a low density, and dark shading indicating a high density of bind-
ing sites). The position (in mm) of each plate relative to bregma is: A 2.68; B 1.98; C 1.18; D 0.62; E 0.02; F �0.94; G �1.34; H �1.94; I
�2.54; J �2.92; K �3.80; L �4.48; M �4.96; N �5.40; O �5.88; P �7.64. The distribution of RXFP3 mRNA was determined by combin-
ing data generated in the current study with data publicly available in the Allen Brain Atlas (www.brainmap.org). For abbreviations, see list.
Scale bar ¼ 1.0 mm.
Relaxin-3/RXFP3 distribution in mouse brain
The Journal of Comparative Neurology | Research in Systems Neuroscience 4027
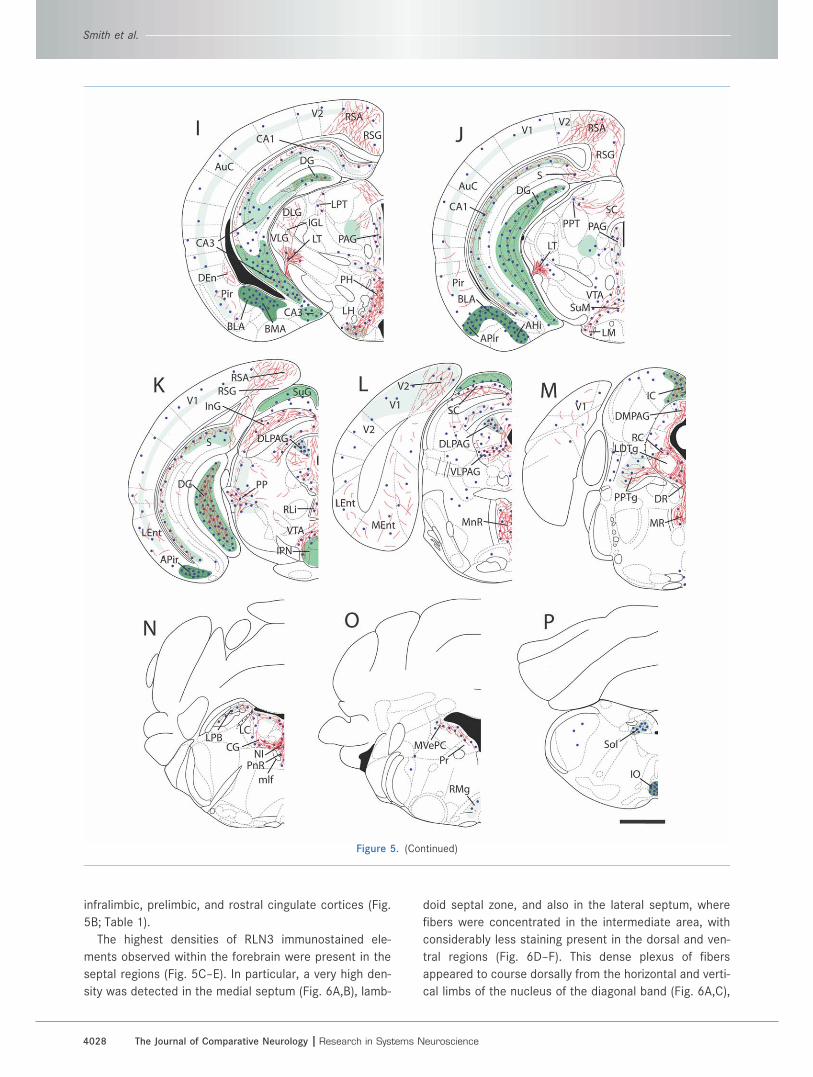
infralimbic, prelimbic, and rostral cingulate cortices (Fig.
5B; Table 1).
The highest densities of RLN3 immunostained ele-
ments observed within the forebrain were present in the
septal regions (Fig. 5C–E). In particular, a very high den-
sity was detected in the medial septum (Fig. 6A,B), lamb-
doid septal zone, and also in the lateral septum, where
fibers were concentrated in the intermediate area, with
considerably less staining present in the dorsal and ven-
tral regions (Fig. 6D–F). This dense plexus of fibers
appeared to course dorsally from the horizontal and verti-
cal limbs of the nucleus of the diagonal band (Fig. 6A,C),
Figure 5. (Continued)
Smith et al.
4028 The Journal of Comparative Neurology |Research in Systems Neuroscience
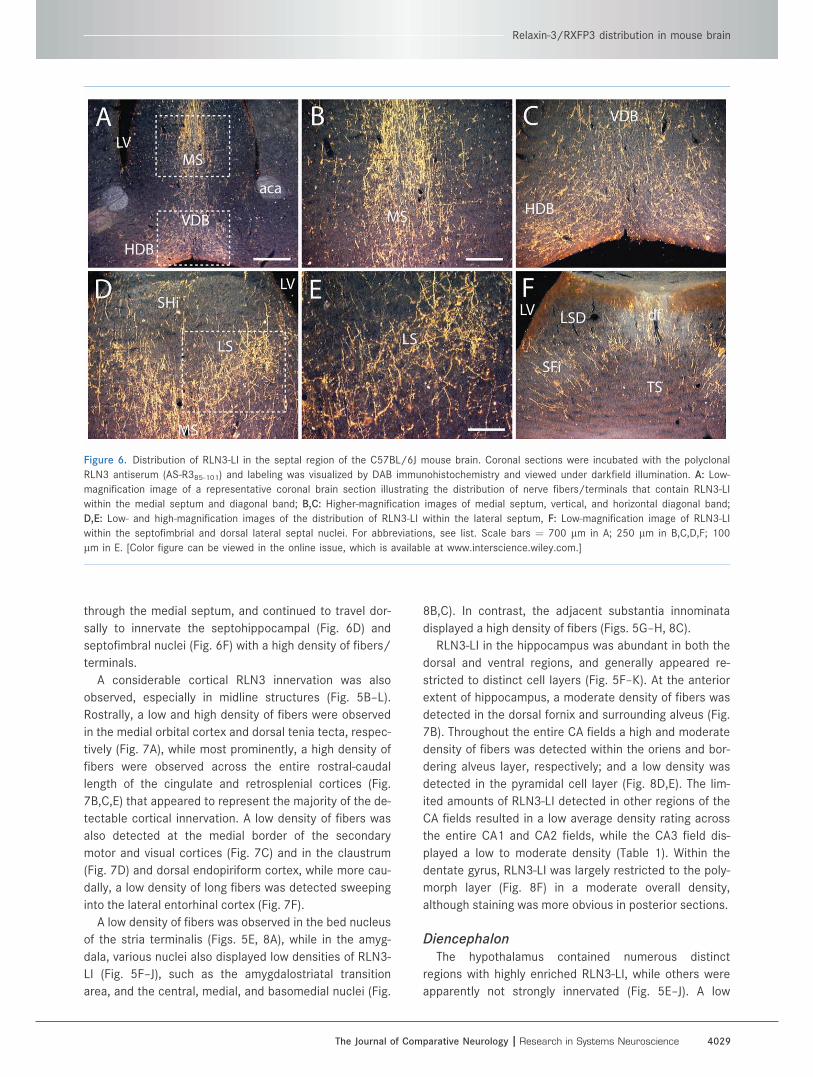
through the medial septum, and continued to travel dor-
sally to innervate the septohippocampal (Fig. 6D) and
septofimbral nuclei (Fig. 6F) with a high density of fibers/
terminals.
A considerable cortical RLN3 innervation was also
observed, especially in midline structures (Fig. 5B–L).
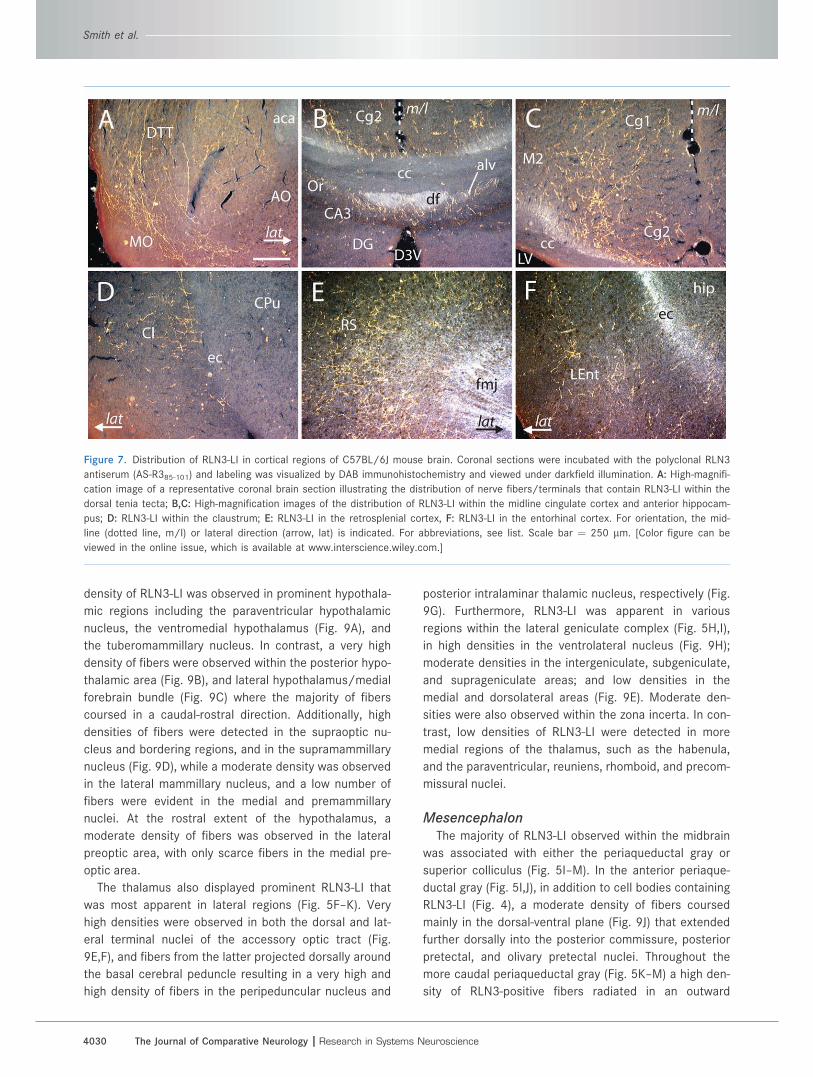
Rostrally, a low and high density of fibers were observed
in the medial orbital cortex and dorsal tenia tecta, respec-
tively (Fig. 7A), while most prominently, a high density of
fibers were observed across the entire rostral-caudal
length of the cingulate and retrosplenial cortices (Fig.
7B,C,E) that appeared to represent the majority of the de-
tectable cortical innervation. A low density of fibers was
also detected at the medial border of the secondary
motor and visual cortices (Fig. 7C) and in the claustrum
(Fig. 7D) and dorsal endopiriform cortex, while more cau-
dally, a low density of long fibers was detected sweeping
into the lateral entorhinal cortex (Fig. 7F).
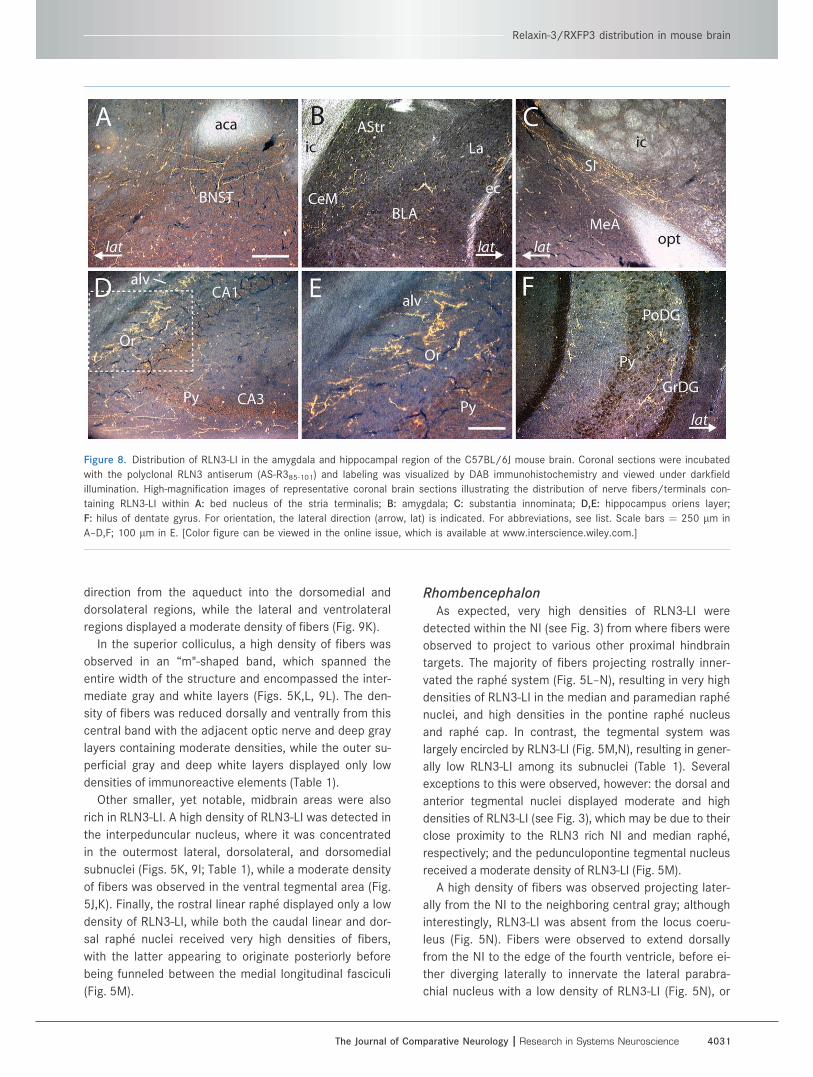
A low density of fibers was observed in the bed nucleus
of the stria terminalis (Figs. 5E, 8A), while in the amyg-
dala, various nuclei also displayed low densities of RLN3-
LI (Fig. 5F–J), such as the amygdalostriatal transition
area, and the central, medial, and basomedial nuclei (Fig.
8B,C). In contrast, the adjacent substantia innominata
displayed a high density of fibers (Figs. 5G–H, 8C).
RLN3-LI in the hippocampus was abundant in both the
dorsal and ventral regions, and generally appeared re-
stricted to distinct cell layers (Fig. 5F–K). At the anterior
extent of hippocampus, a moderate density of fibers was
detected in the dorsal fornix and surrounding alveus (Fig.
7B). Throughout the entire CA fields a high and moderate
density of fibers was detected within the oriens and bor-
dering alveus layer, respectively; and a low density was
detected in the pyramidal cell layer (Fig. 8D,E). The lim-
ited amounts of RLN3-LI detected in other regions of the
CA fields resulted in a low average density rating across
the entire CA1 and CA2 fields, while the CA3 field dis-
played a low to moderate density (Table 1). Within the
dentate gyrus, RLN3-LI was largely restricted to the poly-
morph layer (Fig. 8F) in a moderate overall density,
although staining was more obvious in posterior sections.
DiencephalonThe hypothalamus contained numerous distinct
regions with highly enriched RLN3-LI, while others were
apparently not strongly innervated (Fig. 5E–J). A low
Figure 6. Distribution of RLN3-LI in the septal region of the C57BL/6J mouse brain. Coronal sections were incubated with the polyclonal
RLN3 antiserum (AS-R385-101) and labeling was visualized by DAB immunohistochemistry and viewed under darkfield illumination. A: Low-
magnification image of a representative coronal brain section illustrating the distribution of nerve fibers/terminals that contain RLN3-LI
within the medial septum and diagonal band; B,C: Higher-magnification images of medial septum, vertical, and horizontal diagonal band;
D,E: Low- and high-magnification images of the distribution of RLN3-LI within the lateral septum, F: Low-magnification image of RLN3-LI
within the septofimbrial and dorsal lateral septal nuclei. For abbreviations, see list. Scale bars ¼ 700 lm in A; 250 lm in B,C,D,F; 100
lm in E. [Color figure can be viewed in the online issue, which is available at www.interscience.wiley.com.]
Relaxin-3/RXFP3 distribution in mouse brain
The Journal of Comparative Neurology | Research in Systems Neuroscience 4029
density of RLN3-LI was observed in prominent hypothala-
mic regions including the paraventricular hypothalamic
nucleus, the ventromedial hypothalamus (Fig. 9A), and
the tuberomammillary nucleus. In contrast, a very high
density of fibers were observed within the posterior hypo-
thalamic area (Fig. 9B), and lateral hypothalamus/medial
forebrain bundle (Fig. 9C) where the majority of fibers
coursed in a caudal-rostral direction. Additionally, high
densities of fibers were detected in the supraoptic nu-
cleus and bordering regions, and in the supramammillary
nucleus (Fig. 9D), while a moderate density was observed
in the lateral mammillary nucleus, and a low number of
fibers were evident in the medial and premammillary
nuclei. At the rostral extent of the hypothalamus, a
moderate density of fibers was observed in the lateral
preoptic area, with only scarce fibers in the medial pre-
optic area.
The thalamus also displayed prominent RLN3-LI that
was most apparent in lateral regions (Fig. 5F–K). Very
high densities were observed in both the dorsal and lat-
eral terminal nuclei of the accessory optic tract (Fig.
9E,F), and fibers from the latter projected dorsally around
the basal cerebral peduncle resulting in a very high and
high density of fibers in the peripeduncular nucleus and
posterior intralaminar thalamic nucleus, respectively (Fig.
9G). Furthermore, RLN3-LI was apparent in various
regions within the lateral geniculate complex (Fig. 5H,I),
in high densities in the ventrolateral nucleus (Fig. 9H);
moderate densities in the intergeniculate, subgeniculate,
and suprageniculate areas; and low densities in the
medial and dorsolateral areas (Fig. 9E). Moderate den-
sities were also observed within the zona incerta. In con-
trast, low densities of RLN3-LI were detected in more
medial regions of the thalamus, such as the habenula,
and the paraventricular, reuniens, rhomboid, and precom-
missural nuclei.
MesencephalonThe majority of RLN3-LI observed within the midbrain
was associated with either the periaqueductal gray or
superior colliculus (Fig. 5I–M). In the anterior periaque-
ductal gray (Fig. 5I,J), in addition to cell bodies containing
RLN3-LI (Fig. 4), a moderate density of fibers coursed
mainly in the dorsal-ventral plane (Fig. 9J) that extended
further dorsally into the posterior commissure, posterior
pretectal, and olivary pretectal nuclei. Throughout the
more caudal periaqueductal gray (Fig. 5K–M) a high den-
sity of RLN3-positive fibers radiated in an outward
Figure 7. Distribution of RLN3-LI in cortical regions of C57BL/6J mouse brain. Coronal sections were incubated with the polyclonal RLN3
antiserum (AS-R385-101) and labeling was visualized by DAB immunohistochemistry and viewed under darkfield illumination. A: High-magnifi-
cation image of a representative coronal brain section illustrating the distribution of nerve fibers/terminals that contain RLN3-LI within the
dorsal tenia tecta; B,C: High-magnification images of the distribution of RLN3-LI within the midline cingulate cortex and anterior hippocam-
pus; D: RLN3-LI within the claustrum; E: RLN3-LI in the retrosplenial cortex, F: RLN3-LI in the entorhinal cortex. For orientation, the mid-
line (dotted line, m/l) or lateral direction (arrow, lat) is indicated. For abbreviations, see list. Scale bar ¼ 250 lm. [Color figure can be
viewed in the online issue, which is available at www.interscience.wiley.com.]
Smith et al.
4030 The Journal of Comparative Neurology |Research in Systems Neuroscience
direction from the aqueduct into the dorsomedial and
dorsolateral regions, while the lateral and ventrolateral
regions displayed a moderate density of fibers (Fig. 9K).
In the superior colliculus, a high density of fibers was
observed in an ‘‘m"-shaped band, which spanned the
entire width of the structure and encompassed the inter-
mediate gray and white layers (Figs. 5K,L, 9L). The den-
sity of fibers was reduced dorsally and ventrally from this
central band with the adjacent optic nerve and deep gray
layers containing moderate densities, while the outer su-
perficial gray and deep white layers displayed only low
densities of immunoreactive elements (Table 1).
Other smaller, yet notable, midbrain areas were also
rich in RLN3-LI. A high density of RLN3-LI was detected in
the interpeduncular nucleus, where it was concentrated
in the outermost lateral, dorsolateral, and dorsomedial
subnuclei (Figs. 5K, 9I; Table 1), while a moderate density
of fibers was observed in the ventral tegmental area (Fig.
5J,K). Finally, the rostral linear raphe displayed only a low
density of RLN3-LI, while both the caudal linear and dor-
sal raphe nuclei received very high densities of fibers,
with the latter appearing to originate posteriorly before
being funneled between the medial longitudinal fasciculi
(Fig. 5M).
RhombencephalonAs expected, very high densities of RLN3-LI were
detected within the NI (see Fig. 3) from where fibers were
observed to project to various other proximal hindbrain
targets. The majority of fibers projecting rostrally inner-
vated the raphe system (Fig. 5L–N), resulting in very high
densities of RLN3-LI in the median and paramedian raphe
nuclei, and high densities in the pontine raphe nucleus
and raphe cap. In contrast, the tegmental system was
largely encircled by RLN3-LI (Fig. 5M,N), resulting in gener-
ally low RLN3-LI among its subnuclei (Table 1). Several
exceptions to this were observed, however: the dorsal and
anterior tegmental nuclei displayed moderate and high
densities of RLN3-LI (see Fig. 3), which may be due to their
close proximity to the RLN3 rich NI and median raphe,
respectively; and the pedunculopontine tegmental nucleus
received a moderate density of RLN3-LI (Fig. 5M).
A high density of fibers was observed projecting later-
ally from the NI to the neighboring central gray; although
interestingly, RLN3-LI was absent from the locus coeru-
leus (Fig. 5N). Fibers were observed to extend dorsally
from the NI to the edge of the fourth ventricle, before ei-
ther diverging laterally to innervate the lateral parabra-
chial nucleus with a low density of RLN3-LI (Fig. 5N), or
Figure 8. Distribution of RLN3-LI in the amygdala and hippocampal region of the C57BL/6J mouse brain. Coronal sections were incubated
with the polyclonal RLN3 antiserum (AS-R385-101) and labeling was visualized by DAB immunohistochemistry and viewed under darkfield
illumination. High-magnification images of representative coronal brain sections illustrating the distribution of nerve fibers/terminals con-
taining RLN3-LI within A: bed nucleus of the stria terminalis; B: amygdala; C: substantia innominata; D,E: hippocampus oriens layer;
F: hilus of dentate gyrus. For orientation, the lateral direction (arrow, lat) is indicated. For abbreviations, see list. Scale bars ¼ 250 lm in
A–D,F; 100 lm in E. [Color figure can be viewed in the online issue, which is available at www.interscience.wiley.com.]
Relaxin-3/RXFP3 distribution in mouse brain
The Journal of Comparative Neurology | Research in Systems Neuroscience 4031
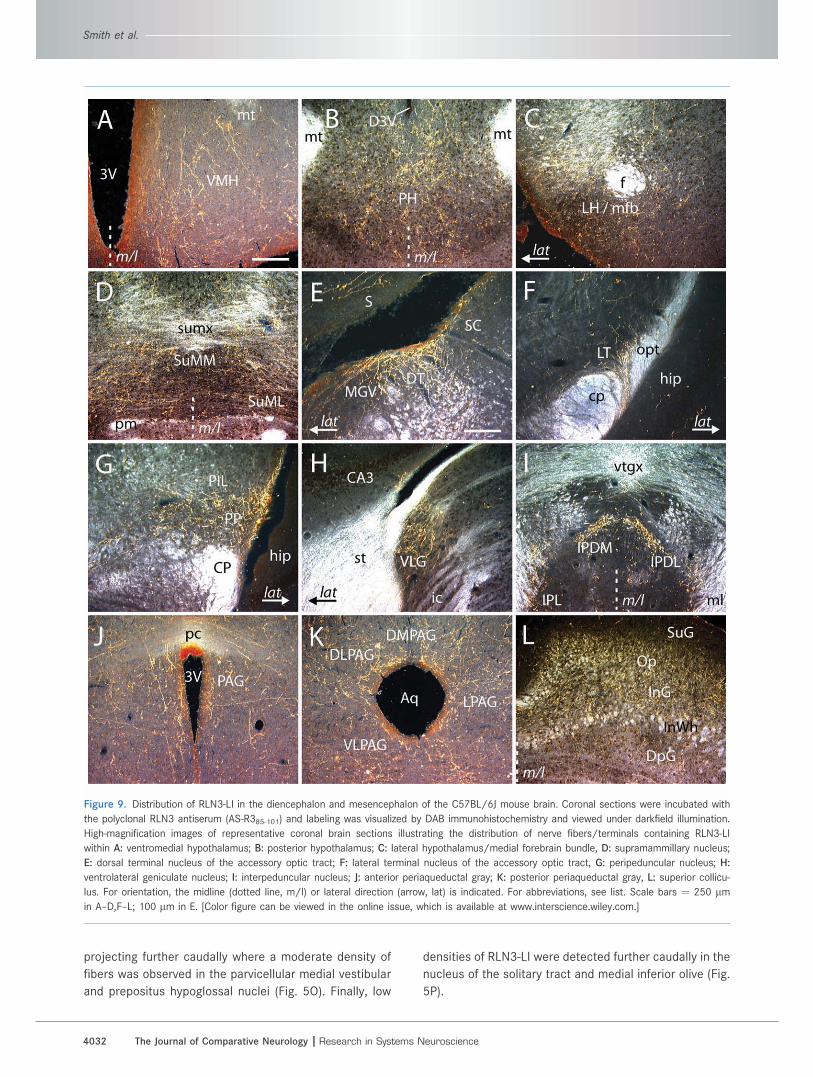
projecting further caudally where a moderate density of
fibers was observed in the parvicellular medial vestibular
and prepositus hypoglossal nuclei (Fig. 5O). Finally, low
densities of RLN3-LI were detected further caudally in the
nucleus of the solitary tract and medial inferior olive (Fig.
5P).
Figure 9. Distribution of RLN3-LI in the diencephalon and mesencephalon of the C57BL/6J mouse brain. Coronal sections were incubated with
the polyclonal RLN3 antiserum (AS-R385-101) and labeling was visualized by DAB immunohistochemistry and viewed under darkfield illumination.
High-magnification images of representative coronal brain sections illustrating the distribution of nerve fibers/terminals containing RLN3-LI
within A: ventromedial hypothalamus; B: posterior hypothalamus; C: lateral hypothalamus/medial forebrain bundle, D: supramammillary nucleus;
E: dorsal terminal nucleus of the accessory optic tract; F: lateral terminal nucleus of the accessory optic tract, G: peripeduncular nucleus; H:
ventrolateral geniculate nucleus; I: interpeduncular nucleus; J: anterior periaqueductal gray; K: posterior periaqueductal gray, L: superior collicu-
lus. For orientation, the midline (dotted line, m/l) or lateral direction (arrow, lat) is indicated. For abbreviations, see list. Scale bars ¼ 250 lmin A–D,F–L; 100 lm in E. [Color figure can be viewed in the online issue, which is available at www.interscience.wiley.com.]
Smith et al.
4032 The Journal of Comparative Neurology |Research in Systems Neuroscience
No RLN3-LI was observed within the cerebellum and
the distribution of RLN3-LI in the spinal cord was not
examined.
Distribution of neurons expressingRXFP3 mRNA
The distribution of RXFP3 mRNA was comprehensively
mapped using data collected in three independent in situ
hybridization studies, which all yielded overlapping distri-
butions that were averaged (see Materials and Methods
for details). The regional distribution of RXFP3 mRNA was
plotted onto schematic brain sections (Fig. 5), and the rel-
ative regional densities of RXFP3 mRNA were semiquanti-
tatively scored (Table 1). A description of this distribution
is provided below, with reference to illustrative autoradio-
grams (Figs. 10, 11).
Rhinencephalon and telencephalonNumerous regions within the telencephalon contained
high densities of RXFP3 mRNA-positive cells, including
the hippocampus and associated areas (Figs. 5F–K, 10),
the medial septum (Figs. 5C–E, 11A), the bed nucleus of
the stria terminalis (Fig. 11B), and the ventral part of the
lateral septum; while the horizontal limb of the diagonal
band and other septal regions displayed moderate den-
sities (Table 1). In hippocampus, high densities of RXFP3
mRNA were present throughout the oriens and pyramidal
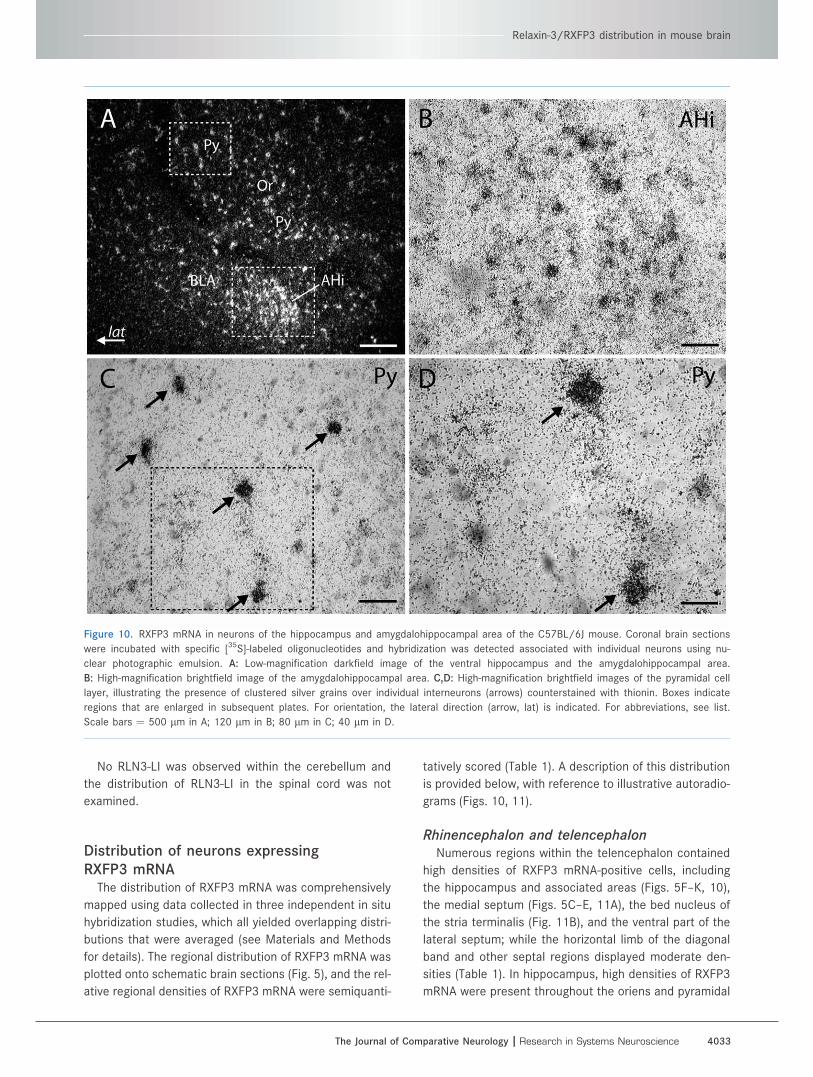
Figure 10. RXFP3 mRNA in neurons of the hippocampus and amygdalohippocampal area of the C57BL/6J mouse. Coronal brain sections
were incubated with specific [35S]-labeled oligonucleotides and hybridization was detected associated with individual neurons using nu-
clear photographic emulsion. A: Low-magnification darkfield image of the ventral hippocampus and the amygdalohippocampal area.
B: High-magnification brightfield image of the amygdalohippocampal area. C,D: High-magnification brightfield images of the pyramidal cell
layer, illustrating the presence of clustered silver grains over individual interneurons (arrows) counterstained with thionin. Boxes indicate
regions that are enlarged in subsequent plates. For orientation, the lateral direction (arrow, lat) is indicated. For abbreviations, see list.
Scale bars ¼ 500 lm in A; 120 lm in B; 80 lm in C; 40 lm in D.
Relaxin-3/RXFP3 distribution in mouse brain
The Journal of Comparative Neurology | Research in Systems Neuroscience 4033
cell layer, resulting in an overall moderate density of
RXFP3 mRNA expression throughout all the large CA
fields (Table 1). In the ventral hippocampus, cells that
expressed a very high level of RXFP3 mRNA displayed to-
pography similar to that of large inhibitory interneurons
(Fig. 10A,C,D). In the dentate gyrus, high densities of pos-
itive cells were detected in the polymorph layer and adja-
cent to or overlying the granule cell layer (Fig. 11C).
The amygdala also displayed strong RXFP3 mRNA
expression (Fig. 5G–K). High densities were detected in
the central nuclei, and along the medial border of the lat-
eral fork of the external capsule, corresponding to the lat-
eral nucleus. A large cluster of RXFP3 mRNA-positive cells
was observed just posterior and ventral to these neurons,
which resulted in an overall high density in the basomedial
and basolateral nuclei, while high and very high densities
were observed in the posteromedial cortical and medial
nuclei, respectively (Fig. 11D). The posterior amygdaloid
region was characterized by a ‘‘boomerang-shaped’’ group
of positive cells comprising the amygdalohippocampal and
amygdalopiriform transition areas, which displayed very
high and moderate densities, respectively (Figs. 5J, 10A,B).
A low density of RXFP3 mRNA expression was detected
throughout much of the mouse forebrain, including the
anterior olfactory nucleus and external plexiform layer of
the olfactory bulb, dorsal tenia tecta, and dorsal transi-
tion zone. Of particular note, throughout the entire rostro-
caudal extent of the cerebral cortex, a low density of
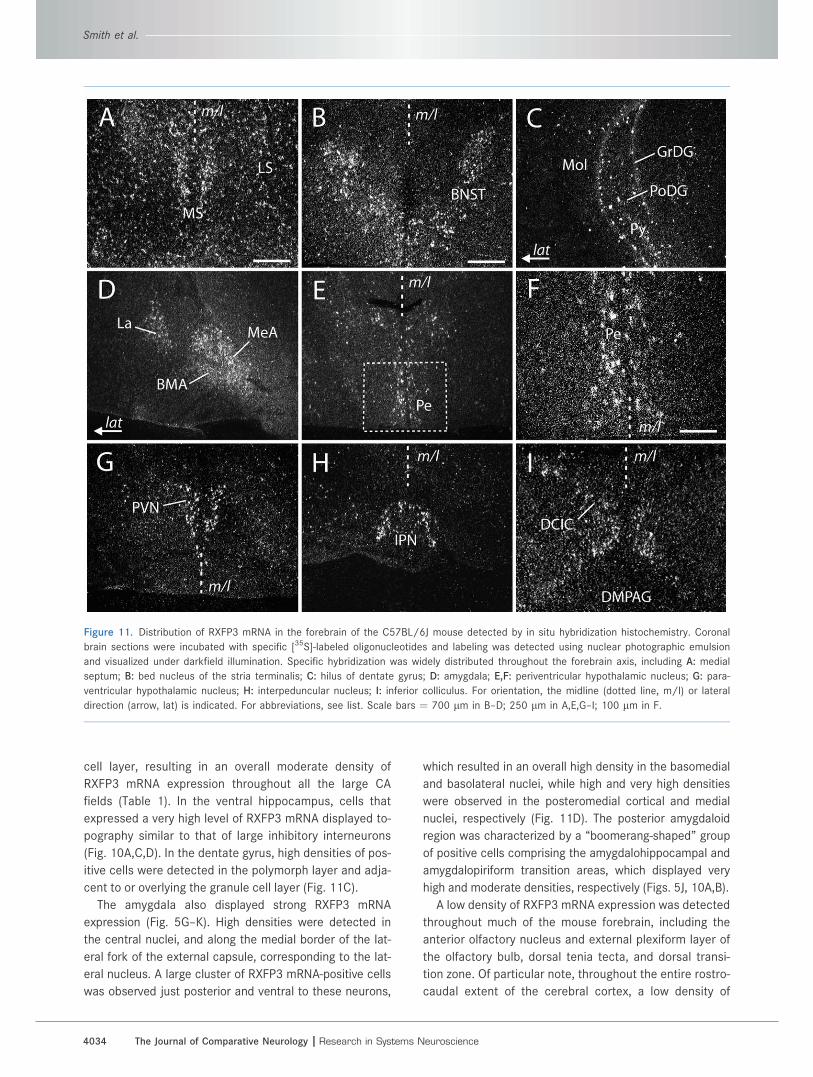
Figure 11. Distribution of RXFP3 mRNA in the forebrain of the C57BL/6J mouse detected by in situ hybridization histochemistry. Coronal
brain sections were incubated with specific [35S]-labeled oligonucleotides and labeling was detected using nuclear photographic emulsion
and visualized under darkfield illumination. Specific hybridization was widely distributed throughout the forebrain axis, including A: medial
septum; B: bed nucleus of the stria terminalis; C: hilus of dentate gyrus; D: amygdala; E,F: periventricular hypothalamic nucleus; G: para-
ventricular hypothalamic nucleus; H: interpeduncular nucleus; I: inferior colliculus. For orientation, the midline (dotted line, m/l) or lateral
direction (arrow, lat) is indicated. For abbreviations, see list. Scale bars ¼ 700 lm in B–D; 250 lm in A,E,G–I; 100 lm in F.
Smith et al.
4034 The Journal of Comparative Neurology |Research in Systems Neuroscience
RXFP3 mRNA was detected in the outer layers of almost
all regions, while the piriform cortex displayed a moderate
density (Fig. 5B–L; Table 1).
DiencephalonThe most prominent site of hypothalamic RXFP3 mRNA
expression was the paraventricular and supraoptic hypo-
thalamic nuclei, which displayed very high densities (Figs.
5F, 11G). High densities were also observed within the
periventricular hypothalamic nucleus (Fig. 11E,F), while
moderate densities were detected in the lateral hypo-
thalamic area / medial forebrain bundle, posterior hypo-
thalamus, and supramammillary nucleus (Fig. 5G–J).
Thalamic RXFP3 mRNA expression was generally low
(Fig. 5F–K), except for very high and high densities in the
habenula and subparafascicular nucleus, respectively,
and moderate densities in the precommissural nucleus
and ventrolateral geniculate nucleus. A cluster of RXFP3
mRNA-positive cells was also observed dorsal to the cere-
bral peduncle, resulting in very high and high densities in
the peripeduncular and posterior intralaminar nuclei,
respectively (Table 1).
MesencephalonRXFP3 mRNA expression was widespread throughout
the midbrain (Fig. 5I–M). The periaqueductal gray was
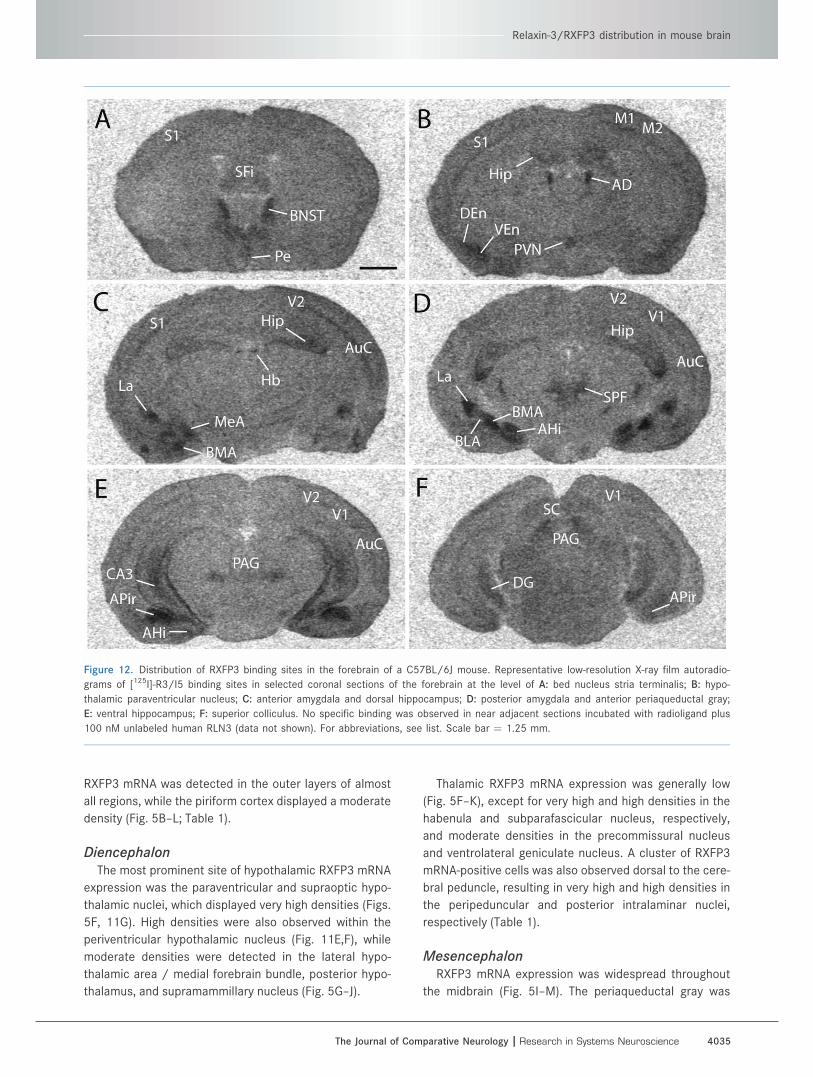
Figure 12. Distribution of RXFP3 binding sites in the forebrain of a C57BL/6J mouse. Representative low-resolution X-ray film autoradio-
grams of [125I]-R3/I5 binding sites in selected coronal sections of the forebrain at the level of A: bed nucleus stria terminalis; B: hypo-
thalamic paraventricular nucleus; C: anterior amygdala and dorsal hippocampus; D: posterior amygdala and anterior periaqueductal gray;
E: ventral hippocampus; F: superior colliculus. No specific binding was observed in near adjacent sections incubated with radioligand plus
100 nM unlabeled human RLN3 (data not shown). For abbreviations, see list. Scale bar ¼ 1.25 mm.
Relaxin-3/RXFP3 distribution in mouse brain
The Journal of Comparative Neurology | Research in Systems Neuroscience 4035

TABLE 1.
Regional Distribution and Relative Density of RLN3-LI, RXFP3 mRNA, and RXFP3 Binding Sites in Mouse Brain
Brain region RLN3-LI RXFP3 mRNA RXFP3 binding sites
RhinencephalonOlfactory bulb
anterior olfactory n. þ þ þependyma and olfactory ventricle þ � �external plexiform layer � þ �glomerular layer � � þinternal granule cell layer � � �
Telencephalonaccumbens n., shell þ/� þ �dorsal tenia tecta þþþ þ þdorsal transition zone þþþ � þsubstantia innominata þþþ þþ þventral pallidum � þ �ventral tenia tecta þ � þAmygdala
amygdalohippocampal area � þþþþ þþþþamygdalopiriform transition area � þþ þþþþamygdalostriatal transition area þ þ/� �anterior amygdaloid area � þ �anterior cortical amygdaloid n. � þ/� þbasolateral n. þ/� þþþ þþþbasomedial n. þ þþþ þþþþcentral n. þ þþþ þþcortex-amygdala transition area � þ/� þlateral n. þ/� þþþ þþþmedial n. þ þþþþ þþþþposterolat cortical amygdaloid n. � þ þposteromed cortical amygdaloid n. � þþþ þþ
Bed nucleus of the stria terminalis
intraamygdaloid division þ þþ þþþlateral division þ þþþ þþmedial division þ þþþ þþþventral division � þ þ/�
Cerebral cortex
auditory cortex � þ þcingulate cortex þþþ þ �claustrum þ þ/� �dorsal peduncular cortex þþ þ/� �ectorhinal cortex � þ þendopiriform n. dorsal þ þ þ/þþendopiriform n. ventral � þ þþþentorhinal cortex þ þ þfrontal association cortex � þ/� þ/�infralimbic cortex þ þ �insular cortex � þ þorbital cortex, medial þ/þþ þ þ/�perirhinal cortex � þ þpiriform cortex � þþ þ/þþposterior parietal associative area � þ þprelimbic cortex þ þ/� þ/�primary motor cortex þ/� þ þretrosplenial cortex þþþ þ �secondary motor cortex þ þ þsomatosensory cortex � þ þsubiculum þ þ þ/�temporal cortex � þ þvisual cortex þ þ þ
Hippocampal formation
alveus þþ � þ/�CA1 field þ þþ þþCA2 field þ þ þþCA3 field þ/þþ þþ þþDG, molecular layer þ/� � þDG, polymorph layer þþ þþþ þþþ
Smith et al.
4036 The Journal of Comparative Neurology |Research in Systems Neuroscience

TABLE 1 (Continued)
Brain region RLN3-LI RXFP3 mRNA RXFP3 binding sites
DG, granular layer þ/� þþ þþdorsal commissure, anterior þ � �dorsal fornix, anterior þþ þ/� �fimbra þ þ þ/�oriens layer þþþ þþþ þþþpyramidal cell layer þ þþþ þþ
Nucleus diagonal band
horizontal limb þþþ þþ þ/�vertical limb þþþ þþþ þ/�
Septum
lambdoid septal zone þþþþ þþ �lateral septal n., dorsal þþ þ �lateral septal n., intermediate þþþþ þþ þ/�lateral septal n., ventral þ þþþ þmedial septal n. þþþþ þþþ þ/�septofimbrial n. þþþ þþ þ/�septohippocampal n. þþþ þþ þ/�triangular septal n. þ þ/� þ
DiencephalonHypothalamus
anterior hypothalamic n. þ/� þ þ/�arcuate n. � þ �dorsomedial hypothalamic n. þ þ þlateral hypothalamic area þþþ þþ þlateroanterior hypothalamic n. þ/� þ �paraventricular hypothalamic n. þ þþþþ þþþperiventricular hypothalamic n. þ/� þþþ þþposterior hypothalamic area þþþþ þþ þpreoptic area, lateral þþ þþ þþpreoptic area, medial þ/� þ þ/�sub-paraventricular zone þ � �suprachiasmatic n. þ þ �supraoptic n. þþþ þþþ þþtuberomammillary n. þ þ þventromedial hypothalamic n. þ/þþ þ þ
Mammillary body
lateral mammillary n. þþ þ/� �medial mammillary þ � �premammillary n. þ þ þsupramammillary n. þþþ þþ þ/�
Thalamus
anterodorsal thalamic n. � þ þþþþcentromedial n. þ/� þ/� þ/�habenula þ þþþ þþþintermediodorsal thalamic n. þ � �Lateral posterior thalamic n. þ þ �n. access optic tract, dorsal terminal þþþþ þ �n. access optic tract, lateral terminal þþþþ þ �parafascicular thalamic n. � þ þþparaventricular thalamic n. þ þ/� þ/�peripeduncular n. þþþþ þþþþ þ/�periventricular fiber system � þþ �posterior intralaminar thalamic n. þþþ þþþ þ/�precommissural n. þ þþ �reuniens thalamic n. þ � þ/�rhomboid thalamic n. þ � þ/�subparafascicular thalamic n. � þþþ þþzona incerta þþ þ þ
Geniculate system
dorsolateral geniculate n. þ � �intergeniculate leaflet þþ þ/� �med geniculate n. þ � �subgeniculate n. þþ þ �suprageniculate n. þþ þ �ventrolateral geniculate n. þþþ þþ �
Relaxin-3/RXFP3 distribution in mouse brain
The Journal of Comparative Neurology | Research in Systems Neuroscience 4037
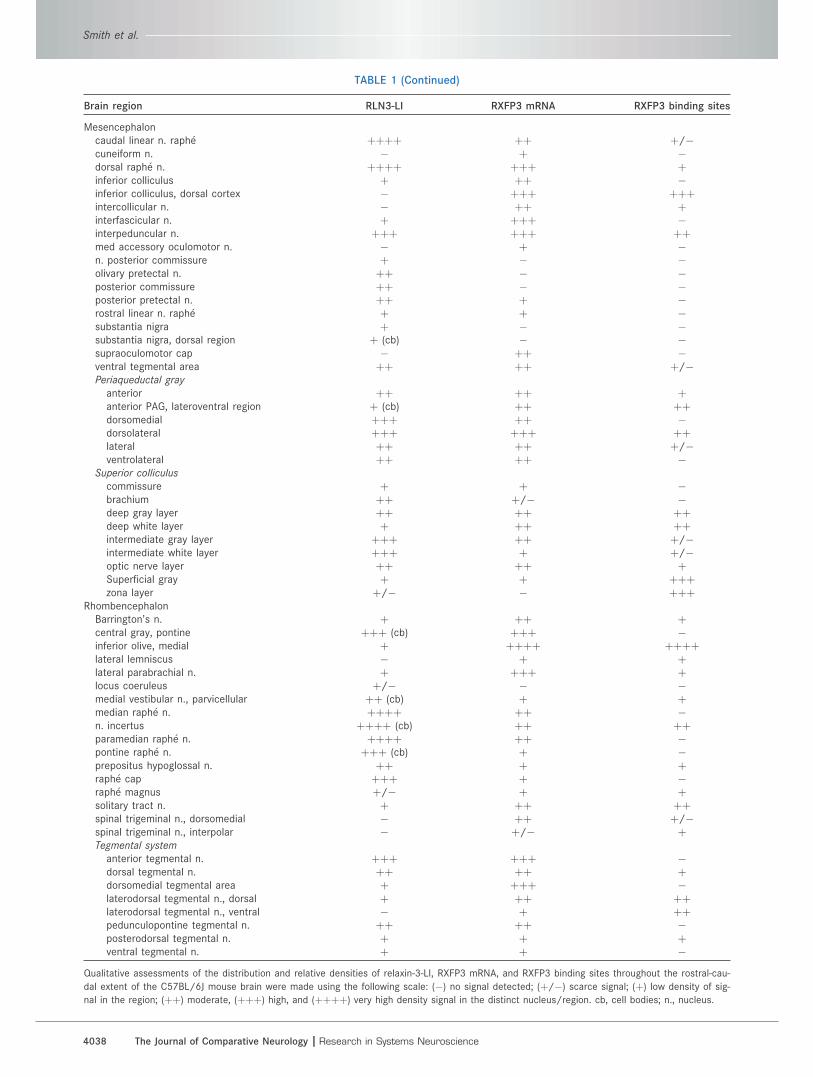
TABLE 1 (Continued)
Brain region RLN3-LI RXFP3 mRNA RXFP3 binding sites
Mesencephaloncaudal linear n. raphe þþþþ þþ þ/�cuneiform n. � þ �dorsal raphe n. þþþþ þþþ þinferior colliculus þ þþ �inferior colliculus, dorsal cortex � þþþ þþþintercollicular n. � þþ þinterfascicular n. þ þþþ �interpeduncular n. þþþ þþþ þþmed accessory oculomotor n. � þ �n. posterior commissure þ � �olivary pretectal n. þþ � �posterior commissure þþ � �posterior pretectal n. þþ þ �rostral linear n. raphe þ þ �substantia nigra þ � �substantia nigra, dorsal region þ (cb) � �supraoculomotor cap � þþ �ventral tegmental area þþ þþ þ/�Periaqueductal gray
anterior þþ þþ þanterior PAG, lateroventral region þ (cb) þþ þþdorsomedial þþþ þþ �dorsolateral þþþ þþþ þþlateral þþ þþ þ/�ventrolateral þþ þþ �
Superior colliculus
commissure þ þ �brachium þþ þ/� �deep gray layer þþ þþ þþdeep white layer þ þþ þþintermediate gray layer þþþ þþ þ/�intermediate white layer þþþ þ þ/�optic nerve layer þþ þþ þSuperficial gray þ þ þþþzona layer þ/� � þþþ
RhombencephalonBarrington’s n. þ þþ þcentral gray, pontine þþþ (cb) þþþ �inferior olive, medial þ þþþþ þþþþlateral lemniscus � þ þlateral parabrachial n. þ þþþ þlocus coeruleus þ/� � �medial vestibular n., parvicellular þþ (cb) þ þmedian raphe n. þþþþ þþ �n. incertus þþþþ (cb) þþ þþparamedian raphe n. þþþþ þþ �pontine raphe n. þþþ (cb) þ �prepositus hypoglossal n. þþ þ þraphe cap þþþ þ �raphe magnus þ/� þ þsolitary tract n. þ þþ þþspinal trigeminal n., dorsomedial � þþ þ/�spinal trigeminal n., interpolar � þ/� þTegmental system
anterior tegmental n. þþþ þþþ �dorsal tegmental n. þþ þþ þdorsomedial tegmental area þ þþþ �laterodorsal tegmental n., dorsal þ þþ þþlaterodorsal tegmental n., ventral � þ þþpedunculopontine tegmental n. þþ þþ �posterodorsal tegmental n. þ þ þventral tegmental n. þ þ �
Qualitative assessments of the distribution and relative densities of relaxin-3-LI, RXFP3 mRNA, and RXFP3 binding sites throughout the rostral-cau-
dal extent of the C57BL/6J mouse brain were made using the following scale: (�) no signal detected; (þ/�) scarce signal; (þ) low density of sig-
nal in the region; (þþ) moderate, (þþþ) high, and (þþþþ) very high density signal in the distinct nucleus/region. cb, cell bodies; n., nucleus.
Smith et al.
4038 The Journal of Comparative Neurology |Research in Systems Neuroscience

particularly enriched with high densities observed within
the dorsolateral region, while moderate densities were
detected in the anterior, dorsomedial, lateral, and ventro-
lateral regions, as well as the neighboring supraoculomo-
tor cap (Table 1). The superior colliculus was similarly
enriched, where two distinct bands of moderate density
RXFP3 mRNA expression were observed. The first was an
‘‘m-shaped’’ band that stretched the width of the superior
colliculus and included the optic nerve and intermediate
gray layers, while the second ‘‘straddled’’ the periaqueduc-
tal gray and included the deep gray and white layers (Fig.
5K,L). RXFP3 mRNA was also present in the inferior collicu-
lus, with high and moderate densities observed in the dor-
sal cortex (Fig. 11I) and adjoining posterior regions.
Other labeled regions include the interpeduncular nu-
cleus, with a high density of RXFP3 mRNA-positive cells
in the outer subnuclei (Fig. 11H), and the mesencephalic
raphe nuclei, where high and moderate densities were
detected in dorsal and caudal linear nuclei, respectively.
Finally, a high density was detected within the interfascic-
ular nucleus, and a moderate density was detected in the
intercollicular nucleus (Table 1).
RhombencephalonThe NI exhibited a moderate density of RXFP3 mRNA
(Fig. 5N), suggesting the existence of autoreceptors to
modulate the release of RLN3 from NI neurons. More lat-
erally, high and moderate densities were detected within
the central gray and lateral parabrachial nucleus, respec-
tively, while the locus coeruleus lacked RXFP3 mRNA
expression despite a moderate density present in the
neighboring area (possibly Barrington’s nucleus).
Various components of the tegmental system dis-
played RXFP3 expression, with high densities observed in
the anterior and dorsomedial regions, and moderate den-
sities detected in the pedunculopontine and dorsal
regions (Table 1). Interestingly, only low to moderate
RXFP3 mRNA expression was detected in the rhombence-
phalon raphe nuclei, including the raphe cap, median,
paramedian, and pontine raphe nuclei.
Caudal to the NI, low densities of RXFP3 mRNA-posi-
tive cells were present in the parvicellular medial vestibu-
lar and prepositus hypoglossal nuclei near the 4th ventri-
cle (Fig. 5O), and a moderate density was present in the
solitary tract and dorsomedial spinal trigeminal nucleus.
Finally, a very high density was present in the medial infe-
rior olive, the most caudal RXFP3-positive cells detected
in the mouse brain (Fig. 12N–P; Table 1).
Regional distribution of [125I]-R3/I5binding sites
In these experiments a low concentration (7–10 pM) of
[125I]-R3/I5 was used to determine the distribution of
RXFP3 binding sites in brain sections from WT and RLN3
KO/LacZ KI mice, and no discernable differences
between genotypes were detected. Relatively high levels
of nonspecific binding were observed in the presence of
100 nM unlabeled RLN3, but these were evenly distrib-
uted across brain sections (data not shown). Further-
more, due to the relatively low resolution inherent in x-ray
film, the regional sensitivity of this technique is below
that of those for the detection of RLN3 and RXFP3 mRNA.
Despite this, the present studies build upon earlier pre-
liminary experiments (Sutton et al., 2006); summarize the
distribution of [125I]-R3/I5 binding sites (Fig. 5), and pro-
vide semiquantitative scoring of the regional relative den-
sities (Table 1). Descriptions of this distribution through-
out the brain follow, with reference to relevant illustrative
autoradiograms.
[125I]-R3/I5 binding sites were most prominent within
the amygdala and hippocampus of the mouse brain (Fig.
5F–K). High to very high densities were observed in multi-
ple nuclei of the amygdala, including the lateral, medial,
basomedial, and basolateral subunits, in addition to the
amygdalohippocampal and amygdalopiriform transition
areas (Fig. 12C–E). In the hippocampus, binding sites were
enriched in the lateral and ventral regions, while more
medial regions displayed relatively sparse [125I]-R3/I5
binding sites (Fig. 12B–E). This resulted in an overall high
density of binding in the oriens layer and a moderate den-
sity of binding in the pyramidal cell layer and CA fields
(Table 1). In the dentate gyrus, high and moderate binding
was detected in the polymorph and granular layers,
respectively, while a low density was detected in the mo-
lecular layer (Table 1).
High to very high densities of [125I]-R3/I5 binding sites
were also observed in the bed nucleus of stria terminalis
(Fig. 12A), paraventricular hypothalamic nuclei (Fig.
12A,B), anterodorsal thalamic nucleus (Fig. 12B), habe-
nula (Fig. 12C), various subregions of the superior and in-
ferior colliculus (Fig. 12D; Table 1), and the inferior olive
(Fig. 5P).
Additionally, moderate densities of [125I]-R3/I5 binding
sites were detected in a range of other regions, including
the NI, supraoptic, lateral preoptic, and periventricular
hypothalamic nuclei (Fig. 12A), the interpeduncular nu-
cleus, subregions of the periaqueductal gray (Fig. 12E,F;
Table 1), the parafascicular and subparafascicular tha-
lamic nuclei (Fig. 12D), several tegmental subregions
(Table 1), and the nucleus of the solitary tract.
The distribution of [125I]-R3/I5 binding sites within the
cortex was of particular interest (Fig. 5B–K). Low, yet
readily detectable [125I]-R3/I5 binding was observed in a
mid-cortical ‘‘band’’ across the entire cortex at a level
equivalent to layer 4, which was absent from the more
medial regions of the cingulate and retrosplenial cortices
Relaxin-3/RXFP3 distribution in mouse brain
The Journal of Comparative Neurology | Research in Systems Neuroscience 4039

that lack this layer (Fig. 12A–E). Finally, a high density of
[125I]-R3/I5 binding sites was observed in the dorsal and
ventral endopiriform nuclei, which was only observable in
coronal sections collected close to bregma �0.94 mm
(Fig. 12B).
DISCUSSION
The present study employed a range of histological
techniques to produce the first comprehensive mapping
of the neuroanatomical distribution of RLN3 and its native
receptor, RXFP3, in the adult mouse brain. In situ hybrid-
ization of RLN3 mRNA, X-GAL histochemistry of a LacZ re-
porter gene in an RLN3 KO mouse and immunohisto-
chemical studies of RLN3-LI consistently revealed that
the major source of RLN3 expression in mouse brain is
large neurons that occupy the medial pontine NI, with
some scattered RLN3-positive neurons observed in the
adjacent pontine central gray and adjacent to the 4th
ventricle at more caudal levels; while separate small pop-
ulations of RLN3-positive neurons were detected in the
pontine raphe nucleus, anterior periaqueductal gray, and
a region dorsal to the substantia nigra and medial to the
peripeduncular nucleus. RLN3-LI was detected in nerve
axons, terminals, and boutons in a wide range of forebrain
areas, in a distribution characteristic of a transmitter/
neuromodulator. In general, a strong overlap was
observed between the distributions of the RLN3 nerve
fibers and RXFP3 mRNA and binding sites, in line with
RLN3 and RXFP3 representing a cognate ligand-receptor
pairing. Analysis of this distribution revealed several
RLN3/RXFP3-enriched brain regions and circuits associ-
ated with common or related functional modalities.
A majority of RLN3/RXFP3-rich brain regions can be
classified as functionally involved in ‘‘behavioral activa-
tion’’ (Goto et al., 2001; Swanson, 2003), and include the
septohippocampal, arousal, and sensory/visuospatial cir-
cuits. Notably, very high densities of RLN3/RXFP3 were
observed within the hippocampus and regions that regu-
late its function, including the medial septum/nucleus of
the diagonal band, posterior hypothalamus, and supra-
mammillary, interpeduncular, and median raphe nuclei,
which together constitute the septohippocampal pathway
(see e.g., Vertes, 1981; Vertes and Kocsis, 1997; Kirk,
1998; Pedemonte et al., 1998; Woodnorth et al., 2003;
Woodnorth and McNaughton, 2005; Jackson et al.,
2008). High levels of RLN3/RXFP3 were also observed in
arousal-related brain areas, such as the raphe nuclei, lat-
eral hypothalamus, and periaqueductal gray, while lower
levels were present in other arousal structures, such as
the ventral tegmental area and the pedunculopontine teg-
mental nucleus (see e.g., Mieda and Yanagisawa, 2002;
Saper et al., 2005; Lu et al., 2006; Monti and Jantos,
2008). Furthermore, RLN3 and/or RXFP3 were also
enriched in numerous structures involved in sensory/
visuospatial control, including the superior and inferior
colliculi, peripeduncular nucleus, lateral/dorsal terminals
of the nucleus of the optic tract, inferior olive, and cingu-
late and retrosplenial cortices (see e.g., Linke, 1999; Gib-
son et al., 2002; Vogt and Laureys, 2005; Dehmel et al.,
2008; Cappe et al., 2009; Halverson and Freeman, 2010).
Second, a number of RLN3/RXFP3-rich regions can be
classified as functionally involved in ‘‘stress responses,’’
and include hypothalamic and extrahypothalamic circuits
that play crucial roles in CRF mediated effects and regula-
tion of the hypothalamic–pituitary–adrenal (HPA) axis. A
large population of RXFP3-positive cells was observed
within the amygdala, while other positive stress- and anxi-
ety-related centers included the lateral septum, bed nu-
cleus of the stria terminalis, PVN, periaqueductal gray,
and dorsal raphe nucleus (see e.g., Potter et al., 1994;
Bittencourt and Sawchenko, 2000; Van Pett et al., 2000;
Justice et al., 2008; Lowry et al., 2008).
Comparative distribution of RLN3-LI, RXFP3mRNA, and [125I]-R3/I5 binding sites
In general, the distribution of RLN3-LI, RXFP3 mRNA,
and [125I]-R3/I5 binding sites appeared to overlap,
although some exceptions were noted. Such ‘‘mis-
matches,’’ which are commonly observed in ligand-recep-
tor systems (Herkenham, 1987; Justice et al., 2008), may
be due to several factors. For example, high densities of
RLN3-LI were observed in numerous raphe nuclei and the
cingulate and retrosplenial cortices, despite relatively low
densities of RXFP3 mRNA and binding sites, which may
indicate that RLN3-positive nerve fibers make few termi-
nal contacts in these regions, and instead project to more
distant structures. Conversely, regions including the
amygdala and paraventricular hypothalamic nucleus dis-
played relatively low densities of RLN3-LI and very high
densities of RXFP3 mRNA and binding sites, which apart
from technical difficulties of detection may indicate either
a high rate of local peptide release and relative depletion
of RLN3 from nerve terminals, and/or that RXFP3 present
in the amygdala and the paraventricular hypothalamic
nucleus are activated by RLN3 released from more dis-
tant sources, via dendritic (Justice et al., 2008) or volume
transmission (Zoli et al., 1999).
Furthermore, high levels of RXFP3 mRNA, but low lev-
els of [125I]-R3/I5 binding sites, were observed in regions
such as the septum and peripeduncular and posterior
intralaminar thalamic nuclei. This may reflect a lower sen-
sitivity and resolution of the binding assay or the reduced
[125I]-R3/I5 binding sites could result from high rates of
receptor internalization, turnover, and/or desensitization
Smith et al.
4040 The Journal of Comparative Neurology |Research in Systems Neuroscience

in ligand-rich regions, and high levels of endogenous
RLN3 binding to RXFP3 blocking [125I]-R3/I5 binding.
However, an explanation involving ligand regulation is not
supported by the observation that the distribution and rel-
ative densities of [125I]-R3/I5 binding were largely identi-
cal in RLN3 KO and WT mice (data not shown).
Interestingly, there were several instances where
bands of [125I]-R3/I5 binding sites were observed to be
differentially located to cells producing RXFP3 mRNA
within the same brain region, suggesting that within these
structures RXFP3 mRNA is produced in cell bodies and
the receptor protein is trafficked to the dendritic or nerve
terminal surface in a different anatomical location (layer/
region). For example, in the cerebral cortex RXFP3 mRNA
was expressed in neurons of the outer layers, whereas
low but robust [125I]-R3/I5 binding was restricted to a mid-
cortical ‘‘band’’ across the entire cortex at a level equiva-
lent to layer 4, which was absent from the more medial
regions of the cingulate and retrosplenial cortices. Also in
the superior colliculus, where the majority of RXFP3
mRNA-positive cells were within more internal layers,
[125I]-R3/I5 binding sites were more abundant peripherally
in the more dorsal zona and superficial gray layers.
Comparative analysis of RLN3/RXFP3distribution in different species
In addition to the highly conserved structure of RLN3
(Wilkinson et al., 2005), the enrichment of RLN3 in neu-
rons of the mouse NI further demonstrates strong homol-
ogy with the rat (Tanaka et al., 2005; Ma et al., 2007),
and with the nonhuman primate, as in a recent study high
levels of RLN3-LI were detected in a pontine region ho-
mologous to the rodent NI in the Macaca fascicularis (Ma
et al., 2009b). However, species differences will likely
exist in the distribution and neurochemistry of RLN3-posi-
tive and -negative neurons within the NI. For example,
based on its cytoarchitecture and efferent and afferent
connections, the rat NI has been defined as comprising a
densely packed medial pars compacta and a more diffuse
and lateral pars dissipata (Goto et al., 2001; Olucha-Bor-
donau et al., 2003). Due to the similar neuroanatomical
distribution of RLN3-positive neurons in the rat brain, it
has been postulated that RLN3 represents a useful
marker of the NI (Tanaka et al., 2005; Ma et al., 2007),
although there are clearly populations of RLN3-negative,
GABA neurons in the rat NI (Ma et al., 2007). An in-depth
anatomical and neurochemical analysis of the mouse NI
should reveal whether RLN3 is also a useful marker for
the NI in this species, or whether the mouse NI displays a
distinct morphology with a different proportion of RLN3-
positive and negative cells, particularly along the midline
(Franklin and Paxinos, 1997; Dong, 2008).
The presence of smaller numbers of RLN3 neurons in
the periaqueductal gray, pontine raphe, and in an area
dorsal to the substantia nigra and medial to the peripe-
duncular nucleus also demonstrates high homology with
the rat (Tanaka et al., 2005; Ma et al., 2007). Although
the contribution that each of these populations makes to
the forebrain pattern of RLN3 innervation is currently
unknown, it is likely that each population has differing
efferent and afferent connections, and may respond to
different local regulatory signals, as well as some stimula-
tory signals that activate the NI. Thus, further study of
these populations and their functional association with
each other and with neurons in the NI remains an impor-
tant step in elucidating the role of central RLN3/RXFP3
signaling.
In general, the distribution of RLN3 efferent projections
and RXFP3 expression/binding sites was very similar in
mice (current study; Sutton et al., 2006) and rats (Tanaka
et al., 2005; Ma et al., 2007), further supporting the exis-
tence of a conserved neuropeptide/receptor system.
However, some differences were apparent. For example,
the presence of RLN3-LI and RXFP3 in the substantia
innominata and the olivary and posterior pretectal nuclei
reported here in the mouse was not described in the rat,
whereas regions which may be more influenced by
RLN3/RXFP3 signaling in the rat than in the mouse
include the olfactory bulb, entorhinal cortex, arcuate
nucleus, and paraventricular thalamic nucleus. Analysis
of the distribution of RLN3 containing projections in the
M. fascicularis also revealed a potentially significant dif-
ference to rodents, as RLN3-LI was absent from midline
cortical regions of macaque brain (Ma et al., 2009b),
while high levels were observed in this region in mouse
and rat. The precise explanation for these differences is
currently unknown.
Putative functions of RLN3/RXFP3signaling in mouse brain
Although to date only a limited number of studies have
been conducted to investigate RLN3/RXFP3 function in
mice (Smith et al., 2009a; Sutton et al., 2009), based on
these and the neuroanatomical distribution of murine
RLN3/RXFP3, it is possible to speculate on the putative
functions of this system. As described above, high den-
sities of RLN3 and RXFP3 were observed in septohippo-
campal, arousal, and sensory/visuospatial control cir-
cuits, indicating that RLN3/RXFP3 signaling likely
modulates behavioral activation/control (Goto et al.,
2001; Swanson, 2003). Specifically, one of the major
functions of the septohippocampal pathway is the gener-
ation/modulation of hippocampal theta rhythm, which is
considered the ‘‘on’’ state of this important structure
Relaxin-3/RXFP3 distribution in mouse brain
The Journal of Comparative Neurology | Research in Systems Neuroscience 4041

involved in vigilance, exploratory behavior, and related
tasks such as navigation and working memory (Vertes
and Kocsis, 1997; Vertes, 2005; Villarreal et al., 2007).
Further to the demonstration that NI stimulation
increased hippocampal theta rhythm in anesthetized rats
(see introduction), we have recently shown that injection
of the RXFP3 agonist, R3/I5, into the medial septum
increased hippocampal theta rhythm, and that this effect
was blocked by pretreatment with an RXFP3 antagonist
(Ma et al., 2009a). These studies also demonstrated
effects of endogenous RXFP3 signaling in the medial sep-
tum on theta activity in conscious rats and on spatial
memory (Ma et al., 2009a).
Furthermore, central administration (i.c.v.) of R3/I5 in
rats produced a dose-related increase in locomotor activ-
ity in automated locomotor cells (Sutton et al., 2009),
while in a study of mixed-background RLN3 KO mice,
they displayed hypoactivity in multiple behavioral para-
digms compared to their WT littermates (Smith et al.,
2009a). These effects on locomotor behavior are in line
with the presence of RLN3/RXFP3 in brain regions associ-
ated with arousal such as the median raphe, lateral hypo-
thalamus, periaqueductal gray, ventral tegmental area,
tuberomammillary, and pedunculopontine tegmental nuclei;
however, interactions with arousal-related transmitters
such as orexin, which are expressed by subpopulations of
neurons within these regions are yet to be investigated.
In addition to arousal, the distribution of RLN3/RXFP3
in the mouse brain supports the idea that RLN3/RXFP3
signaling modulates stress responses through interac-
tions with CRF. Two of the most prominent regions that
displayed RXFP3 expression/binding sites were the
amygdala (Gray and Bingaman, 1996) and PVN (Herman
et al., 2002), while many other structures commonly
linked with stress responses also displayed robust
RLN3/RXFP3, such as the bed nucleus of the stria termi-
nalis (Lee et al., 2008), lateral septum (Henry et al.,
2006), periaqueductal gray (Bowers et al., 2003), and the
dorsal/median raphe nuclei (Graeff et al., 1996; Lowry
et al., 2008; Ohmura et al., 2008). A role of RLN3/RXFP3
in stress responses is further supported by the observed
activation of RLN3 neurons within the NI following neuro-
genic stressors and i.c.v. administration of CRF in the rat
(Tanaka et al., 2005; Banerjee et al., 2010), and the pres-
ence of a moderate density of CRF1 in the mouse NI (Van
Pett et al., 2000; Justice et al., 2008). However, as stress-
related regions such as the parvocellular PVN contain
populations of CRF-positive and -negative neurons (Her-
man et al., 2002), identifying the neurochemical pheno-
type of the cells that express RXFP3 within these regions
will be an important focus of future research.
Finally, the presence of high densities of RXFP3 in the
periventricular hypothalamic nucleus and magnocellular
PVN and supraoptic nucleus suggests a role for RLN3/
RXFP3 signaling in aspects of neuroendocrine and meta-
bolic control in mice. In particular, a possible modulation
of feeding behavior is predicted in light of the robust
increases in food intake and body weight elicited by cen-
tral infusion of relaxin-3 in rats (McGowan et al., 2005,
2006, 2007; Hida et al., 2006). In line with this, a cohort
of mixed background RLN3 KO mice displayed reduced
body weight, body fat, and plasma leptin and insulin lev-
els relative to WT controls when fed on a high fat diet
(Sutton et al., 2009). These findings have not been con-
firmed in backcrossed cohorts, however (Sutton S.W.,
unpubl. data), suggesting that further research is required
in this area. It is also unclear what proportion of the
increased feeding behavior observed in rats was driven
by actions of RLN3/RXFP3 signaling on arousal, as
reported for the peptide transmitters orexin (Willie et al.,
2001; Mieda and Yanagisawa, 2002) and melanin-con-
centrating hormone (Huang et al., 2007), and the mono-
amine histamine (Valdes et al., 2005).
CONCLUSIONS
These anatomical findings further support the con-
served native pairing of RLN3 and RXFP3 in mammals
and provide evidence for broad modulatory actions of
RLN3/RXFP3 networks. The main source of RLN3 in the
mouse is the pontine NI, and RLN3-positive projections
and RXFP3-positive cells are enriched in septohippocam-
pal, arousal, and sensory/visuospatial control circuits
associated with behavioral activation, and in hypothala-
mic and limbic circuits linked to stress and affective
responses. Our findings should assist the design and
interpretation of future anatomical studies to identify
RLN3 target neurons, pharmacological studies of the
effects of RXFP3 agonist and antagonist peptides on
mouse physiology and behavior, and studies to determine
the biological consequences of RLN3 deficiency in KO
mice (Smith et al., 2006a, 2009a,b). It is of importance to
determine the neurochemical identity of RXFP3-positive
cells in different brain regions, as this will provide further
insights into the function of endogenous RLN3/RXFP3
signaling. In this respect, we are currently attempting to
develop robust methods to histologically label RXFP3
using selective RXFP3 antibodies and/or reporter gene
mouse strains (e.g., Gould and Zingg, 2003; Liu et al.,
2003c), including a bacterial artificial chromosome trans-
genic mouse line in which the distribution of RXFP3 can
be conveniently tracked by assessing the distribution of a
fluorescent marker protein (Gong et al., 2003; Justice
et al., 2008). While the putative functional interactions
predicted by the observed neuroanatomical framework
are speculative, several are supported by existing findings
Smith et al.
4042 The Journal of Comparative Neurology |Research in Systems Neuroscience

in rats and represent proposed actions that are testable
experimentally in mice. Such studies should better estab-
lish the role of RLN3/RXFP3 networks in the mammalian
brain and identify their therapeutic potential in the treat-
ment of neurological and psychiatric diseases.
ACKNOWLEDGMENTS
This article is dedicated to Prof. Geoffrey Tregear on
the occasion of his retirement as Deputy Director of the
Howard Florey Institute. The authors thank Tania Ferraro
for assistance with the preparation of antisera used in
these studies. During these studies C.M.S. was the recipi-
ent of an NHMRC (Australia) Dora Lush Biomedical Post-
graduate Scholarship.
LITERATURE CITEDBanerjee A, Ma S, Ortinau S, Smith CM, Layfield S, Burazin
TCD, Shen P-J, Bathgate RAD, Gundlach AL. 2005. Relaxin-3 neurons in the nucleus incertus — projection patterns,response to swim stress and relaxin-3 neuronal signalling.Soc Neurosci Abstr 35:59.57.
Banerjee A, Shen P-J, Ma S, Bathgate RAD, Gundlach AL.2010. Swim stress excitation of nucleus incertus and rapidinduction of relaxin-3 expression via CRF1 activation. Neu-ropharmacology 58:145–155.
Bathgate RAD, Samuel CS, Burazin TCD, Layfield S, ClaaszAA, Reytomas IG, Dawson NF, Zhao C, Bond C, SummersRJ, Parry LJ, Wade JD, Tregear GW. 2002. Human relaxingene 3 (H3) and the equivalent mouse relaxin (M3) gene.Novel members of the relaxin peptide family. J Biol Chem277:1148–1157.
Bathgate RAD, Ivell R, Sanborn BM, Sherwood OD, SummersRJ. 2006. International Union of Pharmacology LVII: recom-mendations for the nomenclature of receptors for relaxinfamily peptides. Pharmacol Rev 58:7–31.
Bittencourt JC, Sawchenko PE. 2000. Do centrally adminis-tered neuropeptides access cognate receptors? An analy-sis in the central corticotropin-releasing factor system.J Neurosci 20:1142–1156.
Boels K, Hermans-Borgmeyer I, Schaller HC. 2004. Identifica-tion of a mouse orthologue of the G-protein-coupled recep-tor SALPR and its expression in adult mouse brain andduring development. Dev Brain Res 152:265–268.
Bowers LK, Swisher CB, Behbehani MM. 2003. Membraneand synaptic effects of corticotropin-releasing factor onperiaqueductal gray neurons of the rat. Brain Res 981:52–57.
Broide RS, Trembleau A, Ellison JA, Cooper J, Lo D, YoungWG, Morrison JH, Bloom FE. 2004. Standardized quantita-tive in situ hybridization using radioactive oligonucleotideprobes for detecting relative levels of mRNA transcriptsverified by real-time PCR. Brain Res 1000:211–222.
Burazin TCD, Larm JA, Ryan MC, Gundlach AL. 2000. Galanin-R1 and -R2 receptor mRNA expression during the develop-ment of rat brain suggests differential subtype involvementin synaptic transmission and plasticity. Eur J Neurosci 12:2901–2917.
Burazin TCD, Bathgate RAD, Macris M, Layfield S, GundlachAL, Tregear GW. 2002. Restricted, but abundant, expres-sion of the novel rat gene-3 (R3) relaxin in the dorsal teg-mental region of brain. J Neurochem 82:1553–1557.
Buzsaki G. 2005. Theta rhythm of navigation: link betweenpath integration and landmark navigation, episodic andsemantic memory. Hippocampus 15:827–840.
Cappe C, Rouiller EM, Barone P. 2009. Multisensory anatomi-cal pathways. Hear Res 258:28–36.
Chronwall BM, Skirboll LR, O’Donohue TL. 1985. Demonstra-tion of a pontine-hippocampal projection containing a rana-tensin-like peptide. Neurosci Lett 53:109–114.
Dehmel S, Cui YL, Shore SE. 2008. Cross-modal interactionsof auditory and somatic inputs in the brainstem and mid-brain and their imbalance in tinnitus and deafness. Am JAudiol 17:S193–209.
Dong HW. 2008. The Allen Reference Atlas: a digital colorbrain atlas of the C57BL/6J male mouse. Hoboken, NJ:John Wiley & Sons.
Franklin KBJ, Paxinos G. 1997. The mouse brain in stereotaxiccoordinates. Sydney: Academic Press.
Gibson AR, Horn KM, Pong M. 2002. Inhibitory control of oli-vary discharge. Ann NY Acad Sci 978:219–231.
Gong S, Zheng C, Doughty ML, Losos K, Didkovsky N, Scham-bra UB, Nowak NJ, Joyner A, Leblanc G, Hatten ME, Heintz N.2003. A gene expression atlas of the central nervous sys-tem based on bacterial artificial chromosomes. Nature425:917–925.
Goto M, Swanson LW, Canteras NS. 2001. Connections of thenucleus incertus. J Comp Neurol 438:86–122.
Gould BR, Zingg HH. 2003. Mapping oxytocin receptor geneexpression in the mouse brain and mammary gland using anoxytocin receptor-LacZ reporter mouse. Neuroscience 122:155–167.
Graeff FG, Guimaraes FS, De Andrade TG, Deakin JF. 1996.Role of 5-HT in stress, anxiety, and depression. PharmacolBiochem Behav 54:129–141.
Gray TS, Bingaman EW. 1996. The amygdala: corticotropin-releasing factor, steroids, and stress. Crit Rev Neurobiol10:155–168.
Gundlach AL, Ma S, Sang Q, Shen PJ, Piccenna L, SedaghatK, Smith CM, Bathgate RAD, Lawrence AJ, Tregear GW,Wade JD, Finkelstein DI, Bonaventure P, Liu C, LovenbergTW, Sutton SW. 2009. Relaxin family peptides and receptorsin mammalian brain. Ann NY Acad Sci 1160:226–235.
Halverson HE, Freeman JH. 2010. Ventral lateral geniculateinput to the medial pons is necessary for visual eyeblinkconditioning in rats. Learn Mem 17:80–85.
Hasselmo ME. 2005. What is the function of hippocampaltheta rhythm? Linking behavioral data to phasic propertiesof field potential and unit recording data. Hippocampus15:936–949.
Henry B, Vale W, Markou A. 2006. The effect of lateral sep-tum corticotropin-releasing factor receptor 2 activation onanxiety is modulated by stress. J Neurosci 26:9142–9152.
Herkenham M. 1987. Mismatches between neurotransmitterand receptor localizations in brain: observations and impli-cations. Neuroscience 23:1–38.
Herman JP, Cullinan WE, Ziegler DR, Tasker JG. 2002. Role ofthe paraventricular nucleus microenvironment in stressintegration. Eur J Neurosci 16:381–385.
Hida T, Takahashi E, Shikata K, Hirohashi T, Sawai T, Seiki T,Tanaka H, Kawai T, Ito O, Arai T, Yokoi A, Hirakawa T,Ogura H, Nagasu T, Miyamoto N, Kuromitsu J. 2006.Chronic intracerebroventricular administration of relaxin-3increases body weight in rats. J Recept Signal TransductRes 26:147–158.
Huang H, Acuna-Goycolea C, Li Y, Cheng HM, Obrietan K, vanden Pol AN. 2007. Cannabinoids excite hypothalamic mela-nin-concentrating hormone but inhibit hypocretin/orexinneurons: implications for cannabinoid actions on foodintake and cognitive arousal. J Neurosci 27:4870–4881.
Relaxin-3/RXFP3 distribution in mouse brain
The Journal of Comparative Neurology | Research in Systems Neuroscience 4043

Jackson J, Dickson CT, Bland BH. 2008. Median raphe stimula-tion disrupts hippocampal theta via rapid inhibition andstate-dependent phase reset of theta-related neural cir-cuitry. J Neurophysiol 99:3009–3026.
Jennes L, Stumpf WE, Kalivas PW. 1982. Neurotensin: topo-graphical distribution in rat brain by immunohistochemis-try. J Comp Neurol 210:211–224.
Justice NJ, Yuan ZF, Sawchenko PE, Vale W. 2008. Type 1corticotropin-releasing factor receptor expression reportedin BAC transgenic mice: implications for reconciling ligand-receptor mismatch in the central corticotropin-releasingfactor system. J Comp Neurol 511:479–496.
Kirk IJ. 1998. Frequency modulation of hippocampal theta bythe supramammillary nucleus, and other hypothalamo-hip-pocampal interactions: mechanisms and functional implica-tions. Neurosci Biobehav Rev 22:291–302.
Lee Y, Fitz S, Johnson PL, Shekhar A. 2008. Repeated stimula-tion of CRF receptors in the BNST of rats selectively inducessocial but not panic-like anxiety. Neuropsychopharmacology33:2586–2594.
Lein ES, Hawrylycz MJ, Ao N, Ayres M, Bensinger A, BernardA, Boe AF, Boguski MS, Brockway KS, Byrnes EJ, et al.2007. Genome-wide atlas of gene expression in the adultmouse brain. Nature 445:168–176.
Linke R. 1999. Differential projection patterns of superior andinferior collicular neurons onto posterior paralaminar nucleiof the thalamus surrounding the medial geniculate body inthe rat. Eur J Neurosci 11:187–203.
Liu C, Chen J, Sutton S, Roland B, Kuei C, Farmer N, SillardR, Lovenberg TW. 2003a. Identification of relaxin-3/INSL7as a ligand for GPCR142. J Biol Chem 278:50765–50770.
Liu C, Eriste E, Sutton S, Chen J, Roland B, Kuei C, Farmer N,Jornvall H, Sillard R, Lovenberg TW. 2003b. Identificationof relaxin-3/INSL7 as an endogenous ligand for the orphanG-protein-coupled receptor GPCR135. J Biol Chem 278:50754–50764.
Liu H, Kishi T, Roseberry AG, Cai X, Lee CE, Montez JM, Fried-man JM, Elmquist JK. 2003c. Transgenic mice expressinggreen fluorescent protein under the control of the melano-cortin-4 receptor promoter. J Neurosci 23:7143–7154.
Liu C, Chen J, Kuei C, Sutton S, Nepomuceno D, BonaventureP, Lovenberg TW. 2005. Relaxin-3/insulin-like peptide 5chimeric peptide, a selective ligand for G protein-coupledreceptor (GPCR)135 and GPCR142 over leucine-richrepeat-containing G protein-coupled receptor 7. Mol Phar-macol 67:231–240.
Lowry CA, Hale MW, Evans AK, Heerkens J, Staub DR, GasserPJ, Shekhar A. 2008. Serotonergic systems, anxiety, andaffective disorder: focus on the dorsomedial part of thedorsal raphe nucleus. Ann NY Acad Sci 1148:86–94.
Lu J, Jhou TC, Saper CB. 2006. Identification of wake-activedopaminergic neurons in the ventral periaqueductal graymatter. J Neurosci 26:193–202.
Ma S, Bonaventure P, Ferraro T, Shen PJ, Burazin TCD, Bath-gate RAD, Liu C, Tregear GW, Sutton SW, Gundlach AL.2007. Relaxin-3 in GABA projection neurons of nucleusincertus suggests widespread influence on forebrain cir-cuits via G-protein-coupled receptor-135 in the rat. Neuro-science 144:165–190.
Ma S, Olucha-Bordonau FE, Hossain MA, Lin F, Kuei C, Liu C,Wade JD, Sutton SW, Nunez A, Gundlach AL. 2009a. Mod-ulation of hippocampal theta oscillations and spatial mem-ory by relaxin-3 neurons of the nucleus incertus. LearnMem 16:730–742.
Ma S, Sang Q, Lanciego JL, Gundlach AL. 2009b. Localizationof relaxin-3 in brain of Macaca fascicularis: identificationof a nucleus incertus in primate. J Comp Neurol 517:856–872.
Matsumoto M, Kamohara M, Sugimoto T, Hidaka K, TakasakiJ, Saito T, Okada M, Yamaguchi T, Furuichi K. 2000. Thenovel G-protein coupled receptor SALPR shares sequencesimilarity with somatostatin and angiotensin receptors.Gene 248:183–189.
McGowan BM, Stanley SA, Smith KL, White NE, Connolly MM,Thompson EL, Gardiner JV, Murphy KG, Ghatei MA, BloomSR. 2005. Central relaxin-3 administration causes hyper-phagia in male Wistar rats. Endocrinology 146:3295–3300.
McGowan BM, Stanley SA, Smith KL, Minnion JS, Donovan J,Thompson EL, Patterson M, Connolly MM, Abbott CR,Small CJ, Gardiner JV, Ghatei MA, Bloom SR. 2006. Effectsof acute and chronic relaxin-3 on food intake and energyexpenditure in rats. Regul Pept 136:72–77.
McGowan BM, Stanley SA, White NE, Spangeus A, PattersonM, Thompson EL, Smith KL, Donovan J, Gardiner JV, GhateiMA, et al. 2007. Hypothalamic mapping of orexigenicaction and Fos-like immunoreactivity following relaxin-3administration in male Wistar rats. Am J Physiol EndocrinolMetab 292:E913–919.
Mieda M, Yanagisawa M. 2002. Sleep, feeding, and neuropep-tides: roles of orexins and orexin receptors. Curr OpinNeurobiol 12:339–345.
Monti JM, Jantos H. 2008. The roles of dopamine and sero-tonin, and of their receptors, in regulating sleep and wak-ing. Prog Brain Res 172:625–646.
Nunez A, Cervera-Ferri A, Olucha-Bordonau F, Ruiz-Torner A,Teruel V. 2006. Nucleus incertus contribution to hippocam-pal theta rhythm generation. Eur J Neurosci 23:2731–2738.
Ohmura Y, Yamaguchi T, Izumi T, Matsumoto M, Yoshioka M.2008. Corticotropin releasing factor in the median raphenucleus is involved in the retrieval of fear memory in rats.Eur J Pharmacol 584:357–360.
Olucha-Bordonau FE, Teruel V, Barcia-Gonzalez J, Ruiz-TornerA, Valverde-Navarro AA, Martinez-Soriano F. 2003.Cytoarchitecture and efferent projections of the nucleusincertus of the rat. J Comp Neurol 464:62–97.
Ortinau S, Lin F, Wade JD, Tregear GW, Bathgate RAD, Gund-lach AL. 2005. Insulin-relaxin family peptide signaling andreceptors in mouse brain membranes and neuronal cells.Ann N Y Acad Sci 1041:211–215.
Paxinos G, Watson C. 2007. The rat brain in stereotaxic coor-dinates. Sydney: Elsevier Academic Press.
Pedemonte M, Barrenechea C, Nunez A, Gambini JP, Garcia-Austt E. 1998. Membrane and circuit properties of lateralseptum neurons: relationships with hippocampal rhythms.Brain Res 800:145–153.
Potter E, Sutton S, Donaldson C, Chen R, Perrin M, Lewis K,Sawchenko PE, Vale W. 1994. Distribution of corticotropin-releasing factor receptor mRNA expression in the rat brainand pituitary. Proc Natl Acad Sci USA 91:8777–8781.
Rosengren KJ, Lin F, Bathgate RAD, Tregear GW, Daly NL,Wade JD, Craik DJ. 2006. Solution structure and novelinsights into the determinants of the receptor specificity ofhuman relaxin-3. J Biol Chem 281:5845–5851.
Ryan MC, Gundlach AL. 1995. Anatomical localisation of pre-proatrial natriuretic peptide mRNA in the rat brain by insitu hybridisation histochemistry: novel identification in ol-factory regions. J Comp Neurol 356:168–182.
Saper CB, Sawchenko PE. 2003. Magic peptides, magic anti-bodies: guidelines for appropriate controls for immunohis-tochemistry. J Comp Neurol 465:161–163.
Saper CB, Scammell TE, Lu J. 2005. Hypothalamic regulationof sleep and circadian rhythms. Nature 437:1257–1263.
Smith CM, Ma S, Shen P-J, Bathgate RAD, Tregear GW, Law-rence AJ, Sutton SW, Gundlach AL. 2006a. Central distri-bution of relaxin-3 and GPCR135 in C57BL/6 and relaxin-3KO mice: further evidence for a role in stress response,
Smith et al.
4044 The Journal of Comparative Neurology |Research in Systems Neuroscience

attentional states and behavioral control. Soc NeurosciAbstr 36:716.15.
Smith CM, Ma S, Shen P-J, Lin F, Wade JD, Bathgate RAD,Gundlach AL. 2006b. Distribution of relaxin-3 andGPCR135 in mouse brain: functional implications for sen-sory processing and stress. Proc Aust Neurosci Soc 17:POS110.
Smith CM, Lawrence AJ, Sutton SW, Gundlach AL. 2009a. Be-havioral phenotyping of mixed-background (129S5:B6)relaxin-3 knockout mice. Ann N Y Acad Sci 1160:236–241.
Smith CM, Shen PJ, Ma S, Sutton SW, Gundlach AL. 2009b.Verification of a relaxin-3 knockout/LacZ reporter mouseas a model of relaxin-3 deficiency. Ann NY Acad Sci 1160:259–260.
Sutin EL, Jacobowitz DM. 1988. Immunocytochemical localiza-tion of peptides and other neurochemicals in the rat later-odorsal tegmental nucleus and adjacent area. J CompNeurol 270:243–270.
Sutton SW, Bonaventure P, Kuei C, Roland B, Chen J, Nepo-muceno D, Lovenberg TW, Liu C. 2004. Distribution of G-protein-coupled receptor (GPCR)135 binding sites and re-ceptor mRNA in the rat brain suggests a role for relaxin-3in neuroendocrine and sensory processing. Neuroendocri-nology 80:298–307.
Sutton SW, Bonaventure P, Kuei C, Nepomuceno D, Wu J, ZhuJ, Lovenberg TW, Liu C. 2006. G-protein-coupled receptor(GPCR)-142 does not contribute to relaxin-3 binding inthe mouse brain: further support that relaxin-3 is the phys-iological ligand for GPCR135. Neuroendocrinology 82:139–150.
Sutton SW, Shelton J, Smith CM, Williams J, Yun S, Motley T,Kuei C, Bonaventure P, Gundlach AL, Liu C, et al. 2009.Metabolic and neuroendocrine responses to RXFP3 modu-lation in the central nervous system. Ann NY Acad Sci1160:242–249.
Swanson LW. 2003. The behavioral state system: intrinsiccontrol of sleep and wakefulness. In: Brain architecture:understanding the basic plan. New York: Oxford UniversityPress. p 139–155.
Tanaka M, Iijima N, Miyamoto Y, Fukusumi S, Itoh Y, OzawaH, Ibata Y. 2005. Neurons expressing relaxin 3/INSL 7 inthe nucleus incertus respond to stress. Eur J Neurosci 21:1659–1670.
Teruel-Marti V, Cervera-Ferri A, Nunez A, Valverde-Navarro AA,Olucha-Bordonau FE, Ruiz-Torner A. 2008. Anatomical evi-dence for a ponto-septal pathway via the nucleus incertusin the rat. Brain Res 1218:87–96.
Trembleau A, Bloom FE. 1995. Enhanced sensitivity for lightand electron microscopic in situ hybridization with multiple
simultaneous non-radioactive oligodeoxynucleotide probes.J Histochem Cytochem 43:829–841.
Valdes JL, Farias P, Ocampo-Garces A, Cortes N, Seron-FerreM, Torrealba F. 2005. Arousal and differential Fos expres-sion in histaminergic neurons of the ascending arousal sys-tem during a feeding-related motivated behaviour. Eur JNeurosci 21:1931–1942.
van der Westhuizen ET, Werry TD, Sexton PM, Summers RJ.2007. The relaxin family peptide receptor 3 activatesextracellular signal-regulated kinase 1/2 through a proteinkinase C-dependent mechanism. Mol Pharmacol 71:1618–1629.
Van Pett K, Viau V, Bittencourt JC, Chan RK, Li HY, Arias C,Prins GS, Perrin M, Vale W, Sawchenko PE. 2000. Distribu-tion of mRNAs encoding CRF receptors in brain and pitui-tary of rat and mouse. J Comp Neurol 428:191–212.
Vertes RP. 1981. An analysis of ascending brain stem sys-tems involved in hippocampal synchronization anddesynchronization. J Neurophysiol 46:1140–1159.
Vertes RP. 2005. Hippocampal theta rhythm: a tag for short-term memory. Hippocampus 15:923–935.
Vertes RP, Kocsis B. 1997. Brainstem-diencephalo-septohippo-campal systems controlling the theta rhythm of the hippo-campus. Neuroscience 81:893–926.
Villarreal DM, Gross AL, Derrick BE. 2007. Modulation of CA3afferent inputs by novelty and theta rhythm. J Neurosci 27:13457–13467.
Vogt BA, Laureys S. 2005. Posterior cingulate, precuneal andretrosplenial cortices: cytology and components of theneural network correlates of consciousness. Prog BrainRes 150:205–217.
Wilkinson TN, Speed TP, Tregear GW, Bathgate RAD. 2005.Evolution of the relaxin-like peptide family. BMC Evol Biol5:14.
Willie JT, Chemelli RM, Sinton CM, Yanagisawa M. 2001. Toeat or to sleep? Orexin in the regulation of feeding andwakefulness. Annu Rev Neurosci 24:429–458.
Wisden W, Morris BJ. 2002. In situ hybridization with oligonu-cleotide probes. Int Rev Neurobiol 47:3–59.
Woodnorth MA, McNaughton N. 2005. Different systemsin the posterior hypothalamic nucleus of rats controltheta frequency and trigger movement. Behav Brain Res163:107–114.
Woodnorth MA, Kyd RJ, Logan BJ, Long MA, McNaughton N.2003. Multiple hypothalamic sites control the frequency ofhippocampal theta rhythm. Hippocampus 13:361–374.
Zoli M, Jansson A, Sykova E, Agnati LF, Fuxe K. 1999. Volumetransmission in the CNS and its relevance for neuropsy-chopharmacology. Trends Pharmacol Sci 20:142–150.
Relaxin-3/RXFP3 distribution in mouse brain
The Journal of Comparative Neurology | Research in Systems Neuroscience 4045


![IEEE TRANSACTIONS ON AFFECTIVE COMPUTING VOL. XXX, …Several affective computing methods [8], [9] used semantic annotations in terms of arousal and valence to capture the under-lying](https://img.pdfslide.net/doc/110x75/5faf85500c275721d342a9ba/ieee-transactions-on-affective-computing-vol-xxx-several-affective-computing-methods.jpg)