Embed Size (px)
Citation preview



D O C T O R A L T H E S I S I N P H Y S I O L O G Y
Who is Who in the Adipose Organ:
A Look at the Heterogeneity of Adipocyte Biology
Jasper Martin Anton de Jong


Who is Who in the Adipose Organ: A look at the Heterogeneity of Adipocyte Biology
Jasper Martin Anton de Jong

© Jasper Martin Anton de Jong, Stockholm University 2017 Cover image by Jasper de Jong. Confocal microscopy image of 3T3-L1 adipocytes with stained lipid droplets (yellow) and nuclei (blue). ISBN print 978-91-7649-742-5 ISBN PDF 978-91-7649-743-2 Printed in Sweden by Universitetsservice US-AB, Stockholm 2017 Distributor: Department of Molecular Biosciences, the Wenner-Gren Institute, Stockholm University

Advisors: Prof. Jan Nedergaard Prof. Barbara Cannon

This thesis refers to the following published papers and manuscripts by their roman numbers (manuscripts are listed in order of appearance in the thesis):
I. De Jong J.M.A., Larsson O., Cannon B. and Nedergaard J. A stringent validation of mouse adipose tissue identity markers. Am J Physiol Endocrinol Metab. 2015;308:E1085-E1105
II. Jespersen N. Z., Larsen T. J., Peijs L., Daugaard S., Homøe P., Loft A., de Jong J., Mathur N., Cannon B., Nedergaard J., Klarlund Pedersen B., Møller K. and Scheele C. A classical brown adipose tissue mRNA signature partly overlaps with brite in the supraclavicular region of adult humans. Cell Metab. 2013;17:798–805.
III. De Jong J.M.A., Fischer A.W., Frontini A., von Essen G., Cinti S., Cannon B., Nedergaard J., Petrovic N. Classical brown adipose tissue in physiologically human-ized mice phenocopies human brown adipose tissue. Manuscript
IV. Shabalina I.G., Petrovic N., de Jong J.M.A., Kalinovich A.V., Cannon B. and Nedergaard. J. UCP1 in brite adipose tissue mitochondria is functionally thermogen-ic. Cell Rep. 2013;5:1196-1203.
V. Kalinovich A.V., de Jong J.M.A., Cannon B., Nedergaard J. UCP1 in adipose tissues: two steps to full browning. Biochimie. 2017 March;134:127-137.
VI. De Jong J.M.A., Wouters R.T.F., Boulet N., Cannon B., Nedergaard J., Pe-trovic N. The 3-adrenergic receptor is dispensable for cold-induced browning of adipose tissues. Am J Physiol Endocrinol Metab In Press. DOI: 10.1152/ajpendo.00437. 2016
VII. De Jong J.M.A., Cannon B., Nedergaard J. In primary brown adipose cultures, fetal and newborn bovine sera differently affect triglyceride storage and thermo-competence. Manuscript
VIII. De Jong J.M.A., Dethlefsen O., Cannon B., Nedergaard J. Utilization of fetal and newborn serum to uncover novel regulators of subcutaneous adipocyte differen-tiation. Manuscript

Contributions to papers that are not included in this thesis:
Abreu-Vieira G., Fischer A.W., Mattsson C., de Jong J.M.A., Shabalina I.G., Ry-dén M., Laurencikiene J., Arner P., Cannon B., Nedergaard J., Petrovic N. Cidea improves the metabolic profile through expansion of adipose tissue. Nat Comm. 2015;6:7433
Fischer A.W., Hoefig C.S., Abreu-Vieira G., de Jong J.M.A., Petrovic N., Mittag J., Cannon B., Nedergaard J. Leptin raises defended body temperature without activat-ing thermogenesis. Cell Rep. 2016;14(7):1621-31

Table of contents
Introduction and aim ---------------------------------------------------------------- 11
1. Heterogeneous adipose appearances ------------------------------------------- 131.1 Defining the term adipocyte ----------------------------------------------------------- 131.2 Brown adipose tissue -------------------------------------------------------------------- 141.3 White adipose tissue -------------------------------------------------------------------- 161.4 Brite/beige adipocytes ------------------------------------------------------------------ 171.5 Other adipose tissues -------------------------------------------------------------------- 17
1.5.1 Bone marrow adipose tissue ---------------------------------------------------------- 181.5.2 Dermal adipose tissue ------------------------------------------------------------------- 181.5.3 Intermuscular adipocytes -------------------------------------------------------------- 19
1.6 Dynamic appearances ------------------------------------------------------------------- 19
2. Functional heterogeneity -------------------------------------------------------- 212.1 Brown adipocyte primary function: Thermogenesis -------------------------------- 212.2 White adipocyte primary function: Energy storage and release ------------------- 222.3 Brite/beige adipocytes: Thermogenic brown-like cells among white cells ------ 242.4 Functions over short and long distances via secreted factors ---------------------- 26
2.4.1 Subcutaneous versus visceral white adipokines -------------------------------- 262.4.2 Brown and brite/beige adipokines -------------------------------------------------- 27
2.5 Adipocytes in supporting roles in other tissues -------------------------------------- 272.5.1 Marrow adipose tissue function ------------------------------------------------------ 272.5.2 Adipocytes in skin homeostasis ------------------------------------------------------ 282.5.3 Adipocytes of the mammary gland -------------------------------------------------- 28
3. Heterogenic adipose tissue dynamics ------------------------------------------ 303.1 Brown adipose remodeling under varying thermal conditions -------------------- 30
3.1.1 Two stages of classical brown adipose recruitment --------------------------- 303.1.2 Observations of BAT proliferation and differentiation ---------------------- 31
3.2 White adipose remodeling in varying thermal conditions ------------------------- 333.2.1 Varying degrees of cold-induced proliferation in white adipose depots ------------------------------------------------------------------------------------------------- 333.2.2 Brite/beige adipocyte formation: interconversion ----------------------------- 343.2.3 Brite/beige adipocyte formation: de novo formation ------------------------- 353.2.4 Brite/beige versus white predetermination --------------------------------------- 36
3.3 Do classical brown and brite/beige cells use different signaling mechanisms to respond to cold? --------------------------------------------------------------------------- 38

3.3.1 Requirement for different -adrenergic receptors in BAT ------------------ 383.3.2 Brite/beige adipocyte formation in 3-adrenergic receptor knockout mice -------------------------------------------------------------------------------------- 393.3.3 Formation of brite/beige adipocytes in different mouse strains ----------- 39
3.4 Diet-induced dynamics of brown and white adipose tissues ---------------------- 413.4.1 Hyperplasia and hypertrophy --------------------------------------------------------- 413.4.2 Diet-induced BAT recruitment ------------------------------------------------------- 443.4.3 Marrow adipose tissue dynamics ---------------------------------------------------- 45
3.5 Intrinsic (pre-)adipocyte differences: ex vivo responses to serum ---------------- 46
4. Developmental heterogeneity --------------------------------------------------- 484.1 Perinatal development of adipose tissues -------------------------------------------- 48
4.1.1 Brown adipose ontogeny --------------------------------------------------------------- 484.1.2 White adipose ontogeny ---------------------------------------------------------------- 494.1.3 Human adipose development --------------------------------------------------------- 50
4.2 Adipocytes derive from a variety of cellular origins ------------------------------- 504.2.1 Interscapular BAT originates in the dermomyotome ------------------------- 514.2.2 The brown fat-muscle relationship -------------------------------------------------- 524.2.3 Subsets of white adipocytes share the BAT/muscle origin ------------------ 524.2.4 Do developmental origins distinguish classical brown from brite/beige? ----------------------------------------------------------------------------------------- 544.2.5 Prx1 selectively labels pools of subcutaneous adipocytes ------------------ 544.2.6 Wilms’ Tumour gene 1 selectively labels subpopulations of visceral adipocytes ------------------------------------------------------------------------------- 55
4.3 Bone marrow adipocytes – a class apart ---------------------------------------------- 55
5. Molecular heterogeneity --------------------------------------------------------- 585.1 Identification of marker genes --------------------------------------------------------- 585.2 Determining the identity of human BAT with murine marker genes ------------- 595.3 Validation of suggested markers ------------------------------------------------------ 615.4 Implications of our marker validation for other studies ---------------------------- 62
5.4.1 What do our results mean for earlier observations? -------------------------- 625.4.2 How have our observations affected later studies? --------------------------- 64
Summary and conclusions --------------------------------------------------------- 66
Sammanfattning på Svenska ------------------------------------------------------- 69
Nederlandse samenvatting --------------------------------------------------------- 70
Acknowledgements ----------------------------------------------------------------- 72
References ---------------------------------------------------------------------------- 75


11
Introduction and aim
The mammalian body contains a plethora of functionally different cell types that together form an integrated organism. The cells that have specific tasks typically reside and operate within the environment of an organ or tissue. Most organs are clearly defined within one anatomical location, such as the heart or the pancreas. Other organs are more dispersed throughout the body, such as skin and blood. The adipose organ, the combined body of tissues containing adipocytes, can also be considered a “dispersed organ”. One of the key roles of adipose tissue is to store energy obtained from the diet for later times, when dietary energy is scarce. Imbalance of energy intake and combustion can lead to excessive deposition of so-called white adipose tis-sue (obesity), which can eventually result in the manifestation of complica-tions such as type 2 diabetes, cancer and cardiovascular diseases. Impaired development of adipose tissue, as is the case in lipodystrophy, also affects overall health (e.g. also through development of type 2 diabetes).
The obesity epidemic demands that we continue to increase our under-standing of how adipose tissue develops and functions, so we can one day find ways to deal with the burden of obesity on global health. One area that has received increasing attention in this respect is a type of adipose tissue that has the capacity to burn off energy to produce heat: brown adipose tis-sue.
It has become increasingly clear that the adipose organ is very dynamic and heterogeneous. Although new advanced techniques continue to enhance our understanding of the intrinsic complexity of adipose tissues, the dynamic nature and the varying characteristics of adipocytes throughout the body complicate the study of this organ. This variation among adipocytes and adipose tissue forms the main theme of this thesis. The heterogeneity among adipose cells and tissues is particularly apparent in their macroscopic and microscopic appearances (Chapter 1). Related to morphology, various adipose cell types fulfill different functions, yet we still put all of them under the umbrella of adipocytes (Chapter 2). The function of adipocytes heavily depends on the demands of the body imposed by the en-vironment (e.g. availability of dietary energy). In response to rapid changes in the environment, adipose tissue composition is uniquely dynamic (Chap-ter 3). Furthermore, the developmental timing and origins of the different – and also the seemingly similar – adipocytes display great variation (Chapter 4). To further understand the differences in adipose cells and tissues, much

12
effort is put into their molecular characterization (Chapter 5). Defining the molecular identities of adipocytes in various anatomical locations might be helpful in the translation of knowledge obtained from mouse models to hu-man adipose biology. Based on the morphology, functionality, responsiveness, development and molecular identity, the aim of this thesis is to provide a view of the complexity and heterogeneity within the adipose organ. The realization of this complexity and heterogeneity will influence how the field continues to study this intriguing organ.

13
1. Heterogeneous adipose appearances
1.1 Defining the term adipocyte For many cell types, their definition is rather clear. Once differentiated, many cells can be recognized from their morphological features. For exam-ple, myotubes are characteristically elongated, multinucleated cells. Neurons can be recognized by their distinctive dendrites and their uniquely long ax-ons. Furthermore, many cell types can be easily defined by their anatomical location; e.g. within defined organs, such as the pancreas or liver.
Adipocytes, or at least the cells we currently classify as such (see below), are different in the way that they are found at many anatomical sites throughout the body. Furthermore, adipocytes are morphologically uniquely dynamic (discussed in paragraph 1.6). Therefore, it is difficult to determine which cells should be considered adipocytes. For clarity, in this thesis, the term adipocyte refers to cells that display the morphological feature mostly associated with adipocyte identity: the presence of one or more lipid droplets that occupy a large fraction of the intracellular space. Additionally, although not entirely exclusive, the term adipocyte here refers to cells located in tis-sues where the majority of cells are lipid-filled (referred to as adipose tis-sues) under non-pathological conditions. In addition to cells that fulfill those two criteria, some other cells are included that may or may not completely fulfill the criterion of being located in an adipose tissue (e.g. bone marrow or dermis). However, those cells form only a minor part of this thesis (mostly because they are underrepresented in the current adipose-related scientific literature).
Although other cell types, such as hepatocytes, have been clearly demon-strated to be capable of storing large amounts of lipids, they are not consid-ered adipocytes as that only commit to lipid-storage under pathological con-ditions (e.g diet-induced obesity). As will become clear from this chapter, even among adipocytes, many dif-ferences exist related to morphology.
Adipocytes are typically divided into two groups: brown and white adipo-cytes, residing in brown and white adipose tissues (or depots), respectively. However, as will become clear in this and the following chapters, this brown-and-white classification is not sufficient to describe the whole of adi-pose tissues and adipose cells in an organism (also referred to as the adipose organ) (Cinti, 1999; Wells, 1940). To give a first view of the adipose organ

14
and to start distinguishing between different types of adipocytes and adipose tissues, this chapter will describe the morphological heterogeneity observed throughout the adipose organ.
1.2 Brown adipose tissue Brown adipose tissue (BAT), as the name indicates, has a typical brown-reddish color, clearly distinct from white adipose depots. However, just as the color of white adipose tissues can vary, the “brownness” of BAT is also dynamic depending on the physiological circumstances (Cinti, 1999) (Figure 1). The brown-reddish color is predominantly determined by the amount of mitochondria (iron-containing cytochrome C oxidase has a brownish color) and the degree of vascularization.
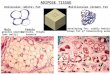
Under most conditions, brown adipocytes contain multiple lipid droplets and the cells are therefore referred to as “multilocular” (Figure 2). The cyto-plasm is typically fully packed with mitochondria that carry out the cell’s primary function: expending energy to generate heat (see paragraph 2.1). Brown adipocyte mitochondria have a round shape, quite different from the more typical rod-shaped mitochondria found in other cell types. In most mammals, brown adipose tissue can be found at several anatomi-cally distinct locations. The major BAT depot is typically located between the scapulae (hence termed interscapular BAT). Other clearly distinct depots are the cervical, subscapular, mediastinal (or periaortic) and perirenal BAT (Paper I) (Cinti, 1999; Symonds et al., 2015).
Figure 1. Overview of most adipose depots in the mouse. Adipose tissues are deposited throughout the body in distinct depots and are typically classified as either white (left) or brown (right). The mouse (9-week-old male NMRI, same as in Paper I) depicted on the left has been acclimated to thermoneutrality for 3 weeks; the mouse on the right was exposed to 4 °C for the same amount of time. Note the difference in color of most adipose depots.

15
In humans, brown adipose tissue has long been regarded as a tissue that is present and active in newborns and declines in size over time to physiologi-cally irrelevant amounts (reviewed in (Lean, 1989)). However, over the past decade numerous studies have shown that brown adipocytes can be identi-fied in adult humans as well (e.g. (Cypess et al., 2009; Nedergaard et al., 2007; Saito et al., 2009; van Marken Lichtenbelt et al., 2009; Virtanen et al., 2009; Zingaretti et al., 2009), Paper II). In infants, the major BAT depot is located in the interscapular area (Aherne and Hull, 1966; Heaton, 1972; Lidell et al., 2013). This depot is no longer observed in adult humans. The brown adipose tissues in adults are found at several anatomical sites, includ-ing the neck area and at perirenal and periadrenal sites (Figure 3) (Paper II) (Cypess et al., 2013; Lidell et al., 2013; Svensson et al., 2014). The “redis-covery” of BAT in adult humans sparked an enormous increase in the amount of research focused on BAT function as a potential therapeutic target to treat obesity and related diseases.
Figure 2. Confocal microscopy images of multilocular brown adipocytes (left) and unilocular white adipocytes (right). Samples were derived from C57BL/6 mouse interscapular brown (left) and inguinal white (right) adipose tissues. Lipid droplets are stained green, cell nuclei are blue and endothelial cells red. Samples were prepared by N. Petrovic; images were captured by A.W. Fischer.

16
1.3 White adipose tissue The majority of adipose cells in mice and humans are typically classified as white adipocytes – the cells that store energy in the form of lipids (see paragraph 2.2). When one looks at the white adipose tissues (WAT) within an animal, one can easily understand why these depots, and the adipocytes residing within them, are referred to as white (Figure 1). However, at the macroscopic level, influenced by varying physiological conditions, white adipose depots can display a range of colors; from white to yellowish to rather brown. The dynamic range of macroscopic appearance differs per depot (Figure 1) (Paper I) (Cinti, 1999). The color of the tissue is mostly influenced by the amount of stored lipids, the degree of vascularization and the mitochondrial density. Under most conditions, white adipose tissues contain much lipid, few mitochondria and are not very well vascularized – and thus have a white appearance. Based on that color, white fat depots can be observed in many places in the mammalian body. In general, white fat is divided into subcutaneous and visceral adipose depots (Figure 3). Subcuta-neous fat depots are located under the skin. In humans, subcutaneous adipose tissue can accumulate at most sites on the body, even in the face. The subcu-taneous adipose tissues in mice appear much more organized in defined de-pots, but can grow extensively. Visceral white fat depots are closely connected with organs (e.g. the gon-ads, heart, kidneys and gastro-intestinal tract) (Figure 1 and 3). In humans, accumulation of visceral fat is typically associated with the appearance of the “beer belly” (or apple shape) and has a clearly distinct appearance from subcutaneous fat accumulation (pear shape). Whether the localization in association with organs implies a direct functional connection with that or-gan is incompletely understood (Cinti, 1999). Distribution of fat mass in humans is generally sex-dependent. Overall, women typically display subcutaneous accumulation of fat in the lower part of the body, whereas men more often display accumulation of visceral fat (Vague, 1956). The size of adipocytes in the various white adipose depots can vary con-siderably. In humans, adipocyte size in subcutaneous and visceral depots displays a typical bimodal distribution including a small population of smaller adipocytes (diameter below 50 m) (Fang et al., 2015). Furthermore, the average size of subcutaneous adipocytes (the most-frequent cell diameter was around 125 m) has been observed to be larger than visceral (omental and mesenteric) adipocytes (in both depots, most cells had a diameter around 100 m) (Fang et al., 2015). Conversely, in one-year-old male C57BL/6 mice housed at room temperature, the average size of subcutaneous inguinal WAT was observed to be smaller than that of the visceral epididymal WAT depot (Sackmann-Sala et al., 2012). However, adipocyte size in the visceral mesenteric WAT depot was similar to that of inguinal WAT (Sackmann-Sala

17
et al., 2012). Thus, adipocyte size distribution in mice is rather heterogene-ous and is not related to anatomical location (subcutaneous or visceral). Another difference among white fat depots is the shape. As can be ob-served in mice, some WAT depots are rather rounded in all dimensions (e.g. epididymal WAT), whereas other depots form a thin layer (e.g. the anterior subcutaneous depot flanking the rib cage in mice) (Figure 1) (Paper I). Whether this variation in tissue shape reflects a difference in function, or whether it is a direct consequence of the available space in their particular anatomical areas is not well understood. White adipocytes have a seemingly simple architecture. Microscopically, they appear as nothing more than one large lipid droplet (they are “unilocu-lar”) with a peripheral, flattened nucleus together surrounded by the cell membrane (Figure 2). In the thin cytoplasmic rim, some mitochondria can be observed together with the Golgi complex and endoplasmic reticulum (Cinti, 1999). However, as described in paragraph 1.6, the cellular appearance can be rather dynamic.
1.4 Brite/beige adipocytes Under various physiological conditions (described in more detail in chap-
ter 3), adipocytes with morphological characteristics of brown adipocytes appear within white fat depots (Young et al., 1984). These adipocytes are multilocular and contain mitochondria that resemble those observed in “clas-sical” brown adipocytes and typically appear as islets surrounded by uni-locular (white) adipocytes (Figure 1 in Paper III) (Cinti, 1999; Young et al., 1984). Because these brown-like adipocytes appear among typical white adipocytes, they are considered a unique, third type of adipocyte. To distin-guish these cells from the “classical” brown and white adipocytes, they have been given termed “brite” (brown-in-white) (Petrovic et al., 2010) or “beige” (Ishibashi and Seale, 2010). As these terminologies refer to the same cell type, for clarity and consistency, they will be referred to in this thesis as “brite/beige” adipocytes. As will become clear in the following chapters, the discovery of brite/beige adipocytes has heavily influenced the field of adi-pose metabolism.
1.5 Other adipose tissues The vast majority of adipocyte research has focused on adipocytes that reside within the typical brown and white adipose depots (i.e. the macro-scopic masses of soft fatty tissue). However, increasingly more attention is given to adipocytes that reside in locations in the body that are (or were) not typically considered adipose tissues.

18
1.5.1 Bone marrow adipose tissue One such example is the presence of adipocytes within the bone marrow. Marrow adipose tissue (MAT) broadly consists of two types: constitutive MAT and regulated MAT (Scheller et al., 2015). Constitutive MAT is typi-cally located in the distal bones (e.g. hands and feet) and has a yellow ap-pearance. Regulated MAT is located in lumbar and thoracic bones, hips and in the proximal limb skeleton (reviewed in (Scheller et al., 2016)). Regulated MAT is much more dynamic. It normally has a more red color because it is a site of hematopoiesis, but can become much fatter under various physiologi-cal conditions (see paragraph 3.4.3). Adipocytes in bone marrow typically have a unilocular appearance, but size can change depending on physiologi-cal circumstances (Scheller et al., 2016).
1.5.2 Dermal adipose tissue Many vertebrate species, including mice and humans, contain a layer of unilocular adipocytes, located under the epidermis. Although sometimes
Figure 3. Distribution of the most-studied adipose depots in adult humans. (A) Brown adipose depots have been localized though observation by PET-scan with radioactively labeled glucose (18FDG) and is divided over several distinct depots. (B) White adipose tissue can be located at many locations in the body. The main depots are depicted in this figure). Distinction is made between sub-cutaneous (located at different sites in the body) and visceral depots (mostly named after the organ it is associated with).

19
confused, subcutaneous and dermal adipose tissues, in rodents, are physical-ly separated by a layer of muscle (Driskell et al., 2014). In humans, too, two anatomically and morphologically distinct layers of adipose tissue can be observed directly under the dermis. Dermal adipose tissue is often only a few cell layers thick and consists of clearly unilocular adipocytes (e.g. (Festa et al., 2011)).
1.5.3 Intermuscular adipocytes Adipocytes have been observed within muscle tissue in mice and humans (Joe et al., 2010). Intermuscular adipocytes have been described both as uni-locular (e.g. (Contreras-Shannon et al., 2007; Uezumi et al., 2010) and multi-locular (Almind et al., 2007). The abundance of multilocular intermuscular multilocular adipocytes is strain-dependent (i.e. they are more abundant in 129S6/SvEvTac than in C57BL/6 mice) (Almind et al., 2007). Although the presence of intermuscular adipocytes in mice is rare, even in obese mice, their abundance has been observed to increase under pathological conditions such as muscle injury (Uezumi et al., 2010).
1.6 Dynamic appearances Although brown and brite/beige adipocytes are generally described as multilocular and white adipocytes as unilocular, the “locularity” of adipo-cytes in the distinctive depots can be rather dynamic. The morphological characteristics that are often used to describe brown and white adipocytes (as also done in the previous paragraphs) are observed in lab rodents under nor-mal housing conditions (i.e. room temperature). With altered housing tem-perature, adipocytes in both brown and white adipose depots will display “atypical” morphological characteristics (e.g. (Loncar et al., 1988)). In Paper III, we showed in mice that prolonged exposure to thermoneu-trality results in a predominantly unilocular appearance of adipocytes in in-terscapular BAT in mice (Figure 1 in Paper III, see also (Sanchez-Gurmaches et al., 2016)). However, some islands of multilocular adipocytes remain. Under such conditions, BAT depots have a very similar microscopic appearance to subcutaneous adipose tissue of cold-exposed mice (Figure 1 in Paper III). Thus, under varying physiological conditions that require differ-ent adipose functionalities, adipose tissue appearances can cover a very wide range. Further discussion on the regulation of this dynamic nature will fol-low in chapter 3. Because of the observed dynamic range of adipocyte appearance, one cannot determine adipose tissue or cell identity based on morphology with-out knowing the physiological conditions and the anatomical origin. For

20
example, adipocytes within classical BAT depots can appear like unilocular white-like adipocytes under thermoneutral conditions. Conversely, adipo-cytes with the morphological characteristics of brown adipocytes are ob-served within typical white fat depots when sufficiently stimulated (e.g. un-der cold conditions). Thus, from the morphological appearance alone, one cannot determine adipocyte identity. Therefore, other characteristics, not visible under the microscope, are needed to aid in the classification and characterization in adipocyte studies.

21
2. Functional heterogeneity
As described in the previous chapter, large morphological variation exists between and within adipose tissues and cells. To further understand the dif-ferences among the various adipose cells and tissues, probably the most meaningful aspect to focus on is their physiological function. For the pur-pose of this chapter, function here is discussed as the use of energy (storage versus expenditure), substrate molecule preferences and the secretion of various factors into the circulation.
To further understand the variation among adipose types, this chapter will provide an overview of the various functions that are associated with differ-ent adipose cells. Based on the variation in metabolic functions, an attempt to further classify adipose cells will be made.
2.1 Brown adipocyte primary function: Thermogenesis Brown adipocytes have the unique ability to produce heat in a regulated manner. This heat-producing functionality is made possible by the presence of uncoupling protein 1 (UCP1), also known as Thermogenin. UCP1 is ex-pressed selectively in brown adipocytes (Cannon et al., 1982) (and in brite/beige adipocytes, see paragraph 2.3).
To understand the function of UCP1, we first have to look at the process of oxidative phosphorylation. Mitochondria typically generate ATP through the process of oxidative phosphorylation in which the electron transport chain generates a proton gradient across the inner mitochondrial membrane, which drives ATP synthase to phosphorylate ADP to generate ATP. In UCP1-containing mitochondria, this proton gradient generated by the elec-tron transport chain is uncoupled from ADP phosphorylation by the action of UCP1. UCP1 drives protons back across the inner mitochondrial membrane into the mitochondrial matrix. In doing so, energy is released as heat. Fur-thermore, levels of functional ATP synthase are kept very low through tran-scriptional regulation of the c-F0 subunit (Cannon and Vogel, 1977; Houstek et al., 1995; Kramarova et al., 2008). Given this primary function of brown adipocytes, heat production, it is thus only logical that brown adipocytes are packed with the characteristic mitochondria that can be physiologically uncoupled.

22
An important aspect, often overlooked, is that UCP1 has to be activated in order to carry out its uncoupling function. The presence of UCP1 in itself is not sufficient to drive uncoupling of oxidative phosphorylation and the re-sulting thermogenesis (Shabalina et al., 2010). The exact molecular mecha-nism through which UCP1 transfers protons back across the inner mitochon-drial membrane remains incompletely understood (Bertholet and Kirichok, 2017; Cannon and Nedergaard, 2004; Klingenberg, 2017; Nicholls, 2017). It is clear, however, that UCP1 function can be regulated in two ways. First, UCP1 activity is inhibited in the presence of purine nucleotides. Under basal conditions UCP1 is believed to be consistently bound - and thus inhibited - by purine nucleotides. Second, activation of UCP1 function is facilitated by fatty acids. Several models have been proposed explaining how fatty acids might be involved in the UCP1-dependent transport of protons back into the mitochondrial matrix (reviewed in (Bertholet and Kirichok, 2017; Cannon and Nedergaard, 2004; Klingenberg, 2017; Nicholls, 2017). All proposed mechanisms share a central role for fatty acids as activators/mediators of UCP1 function.
The availability of fatty acids as activators of UCP1, and indirectly as substrates for the electron transport chain, is thus of great importance for brown adipocyte function. Fatty acids are supplied either by lipolysis of the intracellular lipid stores or by uptake from the circulation, following fatty acid release from white adipocytes or through uptake of fatty acids released from triglyceride-rich lipoproteins containing lipids from the diet (Bartelt et al., 2011).
In addition to fatty acids, BAT also has a remarkably high capacity to glucose uptake upon cold exposure in an insulin-independent norepineph-rine-induced manner (Greco-Perotto et al., 1987; Shimizu et al., 1993). Glu-cose is suggested to serve in thermogenic brown adipocytes as a source of ATP and indirectly (converted to pyruvate) as a thermogenic substrate (Cannon and Nedergaard, 2004).
Combined, uptake of fatty acids and glucose by BAT is enhanced upon activation of thermogenic function, improving overall metabolic health and BAT has therefore taken a central position in the discussion of potential anti-obesity therapeutic targets.
2.2 White adipocyte primary function: Energy storage and release White adipocytes, as opposed to brown adipocytes, are primarily consid-ered to function as energy stores. Based on the morphology described in the previous chapter it can be readily deduced that white adipocytes are highly specialized in storing energy in the form of triglycerides. The stored energy can be made available to the rest of the body in situations where this is re-

23
quired (e.g. in the absence of food intake (starvation), during physical exer-cise or upon exposure to cold temperatures). White adipocytes contain the enzymatic toolbox to take up substrates and convert these into energy-rich triglycerides. Triglycerides are composed of three fatty acyl-CoA moieties and glycerol-3-phosphate. The fatty acids can be taken up from the circulation or produced de novo from carbohydrate substrates (in particular glucose). In addition to fatty acids, white adipocytes take up glucose in an insulin-dependent manner. In addition to serving as an ATP source, glucose contrib-utes to triglyceride synthesis through the glycolysis intermediate dihydroxy-acetone phosphate, which can be converted into glycerol-3-phosphate. In addition, the pyruvate resulting from glycolysis can be further metabolized into fatty acids (de novo lipogenesis). Through its glucose uptake capacity, white adipose tissue plays a key role in the regulation of systemic glucose levels. Triglycerides are stored in lipid droplets, which consist of a phospholipid monolayer with a core in which the triglycerides reside. Lipid droplets are formed at the endoplasmic reticulum and can fuse to form larger droplets (e.g. reviewed in (Wilfling et al., 2014)). Lipid droplets are heavily occupied by many proteins that regulate the formation and maintenance of the drop-lets. Regulation of these lipid droplet-associated proteins is a critical step in the release of the energy stored within the lipid droplets. By taking up lipids and storing them, adipose tissue functions as a lipid buffer (Frayn, 2002). With reduced adipose storing capacity, other organs are exposed to excess lipids. The result can be lipotoxicity, in which the excess of lipids may result in insulin resistance and/or apoptosis. The acute importance of the storage capacity of adipose tissue was elegantly exempli-fied in a mouse model in which insulin receptor could be deleted from ma-ture adipocytes using a tamoxifen-inducible Adiponectin-driven Cre-Lox model (Sakaguchi et al., 2017). Upon deletion of the insulin receptor (or both the insulin receptor and the insulin-like growth factor receptor), adipose depot mass decreased by more than 50 % as lipid stores were depleted. This diminished storage capacity of the adipose tissues had immediate effects on overall metabolism, as indicated by insulin resistance, increased liver tri-glyceride content and enhanced pancreatic -cell proliferation (Sakaguchi et al., 2017). All of these parameters were reversed as soon as the adipose tis-sue had recovered (newly differentiated “wild-type” adipocytes replaced those that had their insulin receptor deleted) (Sakaguchi et al., 2017). In addition to storing energy, when other tissues require fuel to support increased energy demands (e.g. muscle during physical exercise or BAT in the cold), adipose tissue can be induced to release fatty acids from its tri-glyceride stores. Most importantly, adrenergic stimulation ultimately results in the regulation of lipid droplet-associated proteins such as hormone sensi-

24
tive lipase (HSL), adipose triglyceride lipase (ATGL) and perilipins. Fatty acids can then be released into the circulation to be taken up by other tissues.
2.3 Brite/beige adipocytes: Thermogenic brown-like cells among white cells Brite/beige adipocytes can be identified under varying circumstances (described in more detail in paragraph 3.2). Under standard experimental housing conditions (i.e. adult mice living at room temperature) white adipo-cytes in most WAT depots appear rather homogeneous. As described in par-agraph 1.3, adipocyte appearance in white depots can change (e.g. with alter-ing ambient temperature) and become more heterogeneous in the cold. Im-portantly, the amounts of mitochondria and UCP1 increase with lower tem-perature, implying a shift towards thermogenic function of the newly appearing brite/beige adipocytes. In Paper IV, we showed that mitochondria isolated from the inguinal WAT of cold-acclimated mice are indeed func-tionally thermogenic and display functional characteristics similar to those of classical brown fat mitochondria (e.g. rapid substrate oxidation that is inhib-ited by the purine nucleotide GDP, as well as low ADP phosphorylation) and are qualitatively similar. However, the thermogenic respiratory capacity per mitochondrion is somewhat lower for brite/beige mitochondria (depending on the substrate) (Figure 2C-J in Paper IV). Interestingly, UCP1-ablated mice are able to survive several weeks in a cold environment (when preceded by acclimation at 18 °C) (Golozoubova et al., 2001). Although it was initially demonstrated that UCP1-knockout mice maintain their body temperature by shivering, further studies suggested addi-tional thermogenic processes (as determined by oxygen consumption) (Granneman et al., 2003; Grimpo et al., 2014; Kazak et al., 2015; Ukropec et al., 2006). Whereas the enhanced oxygen consumption in WAT of UCP1-knockout mice can be explained as increased lipolysis to provide the shiver-ing muscles with fuel, Kazak et al. proposed a mitochondrial futile cycle around creatine phosphorylation and dephosphorylation that might contrib-ute as a thermogenic mechanism (Kazak et al., 2015). This creatine-dependent thermogenic capacity appeared to occur predominantly in brite/beige adipose tissue as compared to classical brown. It has, however, still to be independently confirmed that such a mechanism exists. Thus, although classical brown and brite/beige mitochondria both display UCP1-dependent thermogenesis, some studies have implied additional brite/beige-selective, UCP1-independent, thermogenic mechanisms. The physiological importance of these processes seems limited since UCP1-ablated mice still fully depend on their shivering capacity (i.e. alternative thermogenic mechanisms are unable to compensate for the loss of UCP1)

25
(Golozoubova et al., 2001). Thus, uncoupled respiration is clearly the key to the major thermogenic capacity of brite/beige adipocytes and they are in that respect similar to classical brown adipocytes.
However, the significance of the contribution of brite/beige adipocytes to the total UCP1-dependent thermogenic capacity of wild-type animals is de-batable and differs per mouse strain (Paper IV). The total protein amounts in the combined WAT depots (in which the brite/beige adipocytes reside) might be larger than that of the combined classical brown depots; total UCP1 protein amounts are clearly much higher in the brown depots (Paper IV, Paper V and extrapolated from RNA data in Paper I). In Paper IV, we de-duced from data obtained from isolated mitochondrial that the thermogenic capacity of the inguinal depot (in 129Sv mice) is maximally 37 % of that of the interscapular BAT depot (this was only 10 % in C57BL/6 mice that are known to have lower “britening” capacity than 129Sv mice) (e.g. (Li et al., 2014; Vitali et al., 2012)).
Thus, whether total brite/beige thermogenic capacity is physiologically significant compared to that of classical BAT depots could be argued. How-ever, the white adipose depots of cold-exposed mice display increased levels of fatty acid release, and thus provide substrate for brown adipose tissue (and also acutely for muscle during the shivering response). However, upon pro-longed cold exposure, both brown and white adipose tissues also display enhanced levels of fatty acid synthesis (Lee et al., 2017; Mottillo et al., 2014; Trayhurn, 1981; Yu et al., 2002). The metabolic benefits of this increased fatty acid synthesis, is not directly evident as fatty acids could be taken up from the circulation (Cannon and Nedergaard, 2004). Regardless, it was reported that in ingWAT of mice treated with the 3-adrenergic receptor agonist CL-316,243 (considered to mimic cold exposure), generally two adipocyte populations could be observed that were different with regard to their expression of fatty acid oxidizing or synthesizing enzymes (Lee et al., 2017). Some (8 %) of the adipocytes expressed enzymes involved in both processes. Cells positive for medium chain acyl-coA dehydrogenase (MCAD, involved in fatty acid oxidation) mostly also were UCP1-expressing, whereas cells expressing fatty acid synthase (FASN) were most-ly negative for UCP1. Interestingly, this segregation of two populations was not observed in interscapular BAT (MCAD and FASN expression over-lapped).
Thus, although simultaneous adrenergic induction of fatty acid oxidation and fatty acid synthesis have been observed in BAT and inguinal WAT, there is a difference in the cellular distribution of these processes (Lee et al., 2017). Interestingly, the above-described results have revealed a level of heterogeneity within the inguinal depot. Further studies should point out whether this observation also occurs upon cold exposure and how this cellu-lar heterogeneity is established.

26
However, other than release of fatty acids into the circulation and contrib-uting to UCP1-dependent thermogenesis, brite/beige adipocytes (as well as white and brown) might serve other functions, as described in paragraph 2.4.2.
2.4 Functions over short and long distances via secreted factors Proper functioning of adipose tissues is obviously relevant for the rest of the body to function. This is indirectly the case through production of heat (brown fat) or storage and release of energy (white fat). Additionally, adi-pose tissues also communicate directly with other organs via the secretion of hormones called adipokines. Adipokines can act directly in the tissue micro-environment (autocrine or paracrine) or act on distant organs in an endocrine manner. This paragraph will cover a few examples to illustrate differences among adipose depot adipokine production.
2.4.1 Subcutaneous versus visceral white adipokines The most-studied adipokine was identified in 1994 as Leptin (Zhang et al., 1994) and its main ascribed function (in addition to many other biologi-cal effects) is to regulate food intake. In brief, with increased fat mass, circu-lating Leptin levels (typically some ng / mL) increase, resulting in decreased food intake via signaling in the hypothalamus. In contrast, in a fasted state, Leptin levels decrease, resulting in increased appetite. Mice, and humans, devoid of Leptin or its receptor (ob/ob and db/db mice, respectively) are hyperphagic and have highly increased fat mass. In humans, Leptin is ex-pressed at higher levels in subcutaneous than in visceral adipocytes (Hube et al., 1996; Van Harmelen et al., 1998; Vohl et al., 2004). Another well-studied adipokine, Adiponectin, has predominantly been recognized as a beneficial adipokine affecting cardiometabolic health and insulin sensitivity. Adiponectin is secreted at higher levels from visceral than subcutaneous adipose cells (Kovacova et al., 2012; Motoshima et al., 2002). In addition, bone marrow adipose tissue has been shown to secrete physio-logically relevant amounts of Adiponectin, contributing significantly to its circulating levels (Cawthorn et al., 2014). In general, visceral adipose tissue produces higher levels of pro-inflammatory cytokines (e.g. TNF , IL-6 and MCP-1) (Fontana et al., 2007; Pou et al., 2007). Indeed, visceral adipose tissue contains more macrophages and other immune cells than subcutaneous adipose tissue, particularly in a state of obesity (Cancello et al., 2006; Murano et al., 2008). One remaining question is how this apparent depot-specific infiltration of immune cells is established. Under artificial experimental conditions, treatment of mice with the lipolytic 3-adrenergic receptor agonist CL-316,243 results in the infiltra-

27
tion and activation of macrophages into visceral, but not subcutaneous adi-pose tissue in mice (Lee et al., 2016). CD44+ macrophages produce lipoxy-genase metabolites (e.g. 9-hydroxyoctadecadeionic acid (9-HODE) and 13-HODE) that serve as ligands for the adipogenic transcription factor PPAR , resulting in enhanced adipogenesis upon 3-adrenergic receptor activation in visceral adipose tissue (Lee et al., 2016). As visceral adipocytes are more sensitive to catecholamine-induced lipolysis than subcutaneous adipocytes (Arner, 1995, 2005), it might be so that higher rates of free fatty acid release attract macrophages predominantly to visceral adipose tissues. Whether fatty acid release, or similar processes, are responsible for macrophage infiltration in a state of obesity remains unknown.
2.4.2 Brown and brite/beige adipokines The secretome of brown adipocytes has thus far not been studied as ex-tensively as that of white fat, but some initial candidates have already been identified. Several identified factors produced by brown adipocytes have been related to autocrine or paracrine function. Vascular endothelial growth factor A (VEGF-A), for example, is increased in BAT in the cold and pro-motes angiogenesis, enabling cold-induced increases in blood flow through the tissue (Xue et al., 2009). Neuregulin 4 (NRG4) is another secreted factor that is induced with brown adipose function and possibly increases the neu-ronal innervation of BAT (Rosell et al., 2014). Probably the most studied brown adipokine is Fibroblast growth factor 21 (FGF21). Although predominantly produced by the liver, FGF21 production and release are induced adrenergically in brown (but also white) adipose tissue (Fisher et al., 2012; Hondares et al., 2011). FGF21 induces the expres-sion of thermogenic genes in BAT and WAT and has overall been shown to have many metabolically favorable effects in various tissues (reviewed in (Fisher and Maratos-Flier, 2016)). FGF21 is thus an example indicating an endocrine role for brown adipose tissue. However, Fgf21 is expressed at similar levels in many WAT depots (Paper I). Thus, FGF21 is not a selective brown (versus white) adipokine. However, the contribution to circulating levels cannot be determined only by mRNA levels.
2.5 Adipocytes in supporting roles in other tissues
2.5.1 Marrow adipose tissue function Adipocytes that reside in the bone marrow are surrounded by the bone environment with seemingly less room to expand than peripheral white adi-pose depots have. However, under obesogenic conditions, marrow adipose

28
tissue can expand rapidly within the limited space (described in more detail in paragraph 3.4.3). In that way, it appears to fulfill an energy storing func-tion similar to peripheral white adipocytes. However, also under calorie-restricted conditions marrow adipose tissue expands (see paragraph 3.4.3) (Cawthorn et al., 2014). Because of this counterintuitive behavior of marrow adipose tissue, it has been suggested that marrow adipocytes fulfill a more local role in protecting the bone marrow niche against lipotoxicity (Scheller et al., 2016). Addition-ally, bone marrow adipocytes have been proposed to play a mechanical role (Ma et al., 2014; Scheller et al., 2016).
2.5.2 Adipocytes in skin homeostasis Adipocytes found in the dermis may play a supporting role to the hair follicles. Dermal adipocytes are believed to be a source of Bone Morphogen-ic Proteins (BMPs), important regulatory ligands of hair follicle cycling (Shook et al., 2016). Furthermore, Leptin secreted by dermal adipocytes may influence hair growth by hair follicle stem cell activation and shaft length (Shook et al., 2016). The observation that dermal adipose tissue thickens in response to cold exposure links dermal adipocytes with hair development in what seems a coordinated response to cold stress (Kasza et al., 2014; Shook et al., 2016). Dermal adipocytes have also been implicated in wound healing. In partic-ular, Adiponectin stimulates re-epithelialization, and Leptin, in addition to re-epithelialization, promotes angiogenesis (Frank et al., 2000; Jin et al., 2015; Salathia et al., 2013). These two processes are critical steps in wound healing. Furthermore, fibroblast migration, another essential step in the wound-healing process, is induced by the presence of adipocytes (Schmidt and Horsley, 2013).
2.5.3 Adipocytes of the mammary gland In pregnant mice, subcutaneous adipose tissue undergoes drastic remodel-ing to support milk production. This includes the formation of lipid-filled epithelial-like cells (Morroni et al., 2004). It has been observed that a subset of these milk-producing secretory epithelial cells form through “transdiffer-entiation” of subcutaneous adipocytes during pregnancy (Morroni et al., 2004). These lipid-filled epithelial cells have been named “pink” adipocytes (Giordano et al., 2014). After the lactation period, the mammary tissue dis-plays strongly enhanced adipogenesis (Morroni et al., 2004). This wave of newly formed adipocytes has been implied to derive from epithelium-to-adipocyte transdifferentiation (Morroni et al., 2004). Thus, the observed transdifferentiation of adipocytes into secretory epi-thelial cells indicates an important role of adipocytes in the lactation process.

29
To conclude, a global functional characterization of brown and brite/beige versus white adipocytes has been established. However, within these three adipocyte subtypes, it appears that more functional differences exist than we now appreciate. Brown adipocytes in particular, are widely considered as one and the same type of cell, although located in various sites of the body. Whether brown adipocytes at different sites are – aside from thermogenesis - functionally different, remains to be determined. Brite/beige adipocytes too, although dispersed throughout the body, are typically generalized. Potential functional differences between brite/beige adipocytes at different anatomical locations have not yet been considered, although experimental evidence points at a rather heterogenic nature among these adipocytes (see following chapters). Among white adipocytes, subcutaneous and visceral adipocytes have been considered qualitatively different based on adipokine profiles and lipid stor-age capacity. However, the main function (energy storage) is ascribed to both white adipose types, but might be induced under different conditions. Other possible functions (e.g. hormone secretion and interaction with other tissues) are not as well described and those differences have yet to be further characterized. Identification and characterization of bone marrow, dermal and mammary gland adipocytes are examples of seemingly typical white adipocytes that appear to serve very specific functions within their local environment. These cell types are gaining attention, and further functional characterization is needed to help place them within the adipose spectrum. Thus, although functional differences are being characterized, the overall consensus is still limited to energy substrate handling in terms of storage (in white adipocytes) and expenditure (brown adipocytes). Bone marrow, der-mal and mammary adipocytes and their specific local functions are intri-guing exceptions to this.

30
3. Heterogenic adipose tissue dynamics
As mentioned earlier (e.g. paragraph 1.6), adipose tissues and cells are very dynamic. In response to the environment, adipocytes can grow or shrink, appear or disappear and change function. Observing how adipocytes adjust to physiological cues might help to further understand the heterogeneity among adipocytes. Because the analysis of adipose dynamics requires ad-vanced cell labeling and tracing systems, the vast majority of this chapter is focused on (transgenic) mouse models. The most studied environmental parameters that affect adipose behavior are temperature and diet and will thus form the main focus of this chapter.
3.1 Brown adipose remodeling under varying thermal conditions One of the questions regarding the differences between classical brown and brite/beige adipocytes concerns the origin of newly formed adipocytes in response to temperature. More specifically, are new cold-induced brown and brite/beige adipocytes formed from proliferating precursor cells or do exist-ing (pre-)adipocytes further functionally differentiate? As discussed below, classical brown and brite/beige appear to behave quite differently with re-gard to these developmental mechanisms. Furthermore, within classical BAT, these mechanisms seem to occur under different thermal conditions.
3.1.1 Two stages of classical brown adipose recruitment It can be noted, as shown in Paper III, that under thermoneutral (30 °C) and standard housing conditions (room temperature), brown adipocytes have different appearances. This could be expected based on thermogenic de-mands at 30 °C or 21 °C. In addition, as indicated for example by UCP1 quantification (either mRNA or protein), robust molecular changes occur (Paper III, Paper V). The temperature-induced increase in UCP1 could po-tentially be explained either by an increase in the total number of UCP1-expressing adipocytes in the tissue, an increase in the amount of UCP1 in the pre-existing brown adipocytes or a combination of both.
To analyze these quantitative tissue dynamics, histological analysis, as performed in Paper III, is typically not a very useful method. For example, tissue expansion (i.e. the total number of cells) is better quantified by meas-

31
urement of total DNA, RNA or protein amounts. In Paper V, we showed that between 30 °C and 21 °C, total (interscapular) BAT RNA and protein con-tent only modestly increased. This could be interpreted as the maintenance of the total cell number in the tissue. However, the amounts of UCP1 mRNA and protein measured per unit of RNA (UCP1 mRNA per reference gene) or protein (UCP1 per mg protein) increased rather drastically (Paper V). As a result, total UCP1 RNA and protein per whole tissue were also increased.
Together, these observations imply that between thermoneutrality and room temperature, the total number of brown (pre-)adipocytes stays at rather constant levels, but these adipocytes increase their individual UCP1 amounts (i.e. differentiation rather than proliferation).
Most published studies aimed at investigating cold-induced effects are typically performed at room temperature (control temperature, considered not cold) and 4 – 10 °C (cold). In classical BAT, between these two thermal conditions, we observed a rather different recruitment response than between thermoneutrality and room temperature. Described in Paper V, between 21 °C and 4 °C, we saw a robust increase in total tissue RNA and protein, im-plying an increase in total tissue cell number. When measuring UCP1 mRNA and protein per unit of RNA (per 18S ribosomal RNA) and mg pro-tein, respectively, only modest increases were observed at 4 °C compared to room temperature. However, the total tissue UCP1 mRNA and protein were again strongly increased between 21 °C and 4 °C. Thus, total tissue UCP1 again increased, but this time predominantly through formation of new brown adipocytes, rather than through differentiation of already existing non-proliferating (pre-) adipocytes.
Combined, between 30 °C and 4 °C, the total UCP1 RNA and protein amounts increased in a linear fashion (Figure 6 in Paper V). The mechanisms (proliferation or differentiation), by which this occurs, however, are different in the different thermal zones (Figure 4). How these two different responses may be regulated depending on the environmental temperature is unknown. Perhaps, variation in the degrees of nerve stimulation may be different in these different states. However, how this could mechanistically result in proliferation or differentiation is unclear.
Whereas we did not directly analyze rates of proliferation or differentia-tion in our study, some earlier studies have provided data indicating the oc-currence of these processes in brown adipose tissue under varying thermal conditions.
3.1.2 Observations of BAT proliferation and differentiation As mentioned, very few studies are performed with mice at thermoneu-trality. As a result, the comparison between thermoneutrality and room tem-perature is seldom made; comparing room temperature and colder is, for practical reasons, the norm. The studies described below indeed generally

32
follow that norm. Thus, studies on cellular dynamics currently are only in-formative about the effect of cold (4 – 10 °C) environments.
One of the first studies investigating the cytological responses of brown fat to varying temperatures was performed in rats (Cameron and Smith, 1964). Within the first 24 h after cold exposure (6 °C, after being kept at 26 °C), an increase in proliferating “reticuloendothelial” cells could be observed as indicated with 3H-thymidine incorporation into proliferating cells (Cameron and Smith, 1964). The data in this study indicated that prolifera-tion peaked after 4 days, after which proliferation decreased to control levels between 8 and 16 days in the cold. Further analysis indicated that these pro-liferating cells indeed gave rise to brown adipocytes (Cameron and Smith, 1964).
Similar studies on cold-induced proliferation in BAT quite consistently demonstrate a rapid hyperplastic response after cold exposure (e.g. (Bukowiecki et al., 1982; Cousin et al., 1996; Lee et al., 2015; Rehnmark and Nedergaard, 1989)) and that indeed UCP1-expressing brown adipocytes are formed from proliferating cells (Lee et al., 2015). Interestingly, most of the cold-induced proliferation was localized to the dorsal region of the inter-scapular brown adipose depot (Lee et al., 2015). Using cell surface markers, the authors showed that cold-induced proliferation was observed in endothe-lial cells (CD31+), macrophages (F4/80+), but also CD45-;PDGFR + pre-adipocytes. Tracing of PDGFR + cells indicated that PDGFR + cells indeed
Figure 4. Cellular dynamics of brown adipose tissue in varying thermal zones. Between thermoneutrality and room temperature, BAT recruitment seems to occur predominantly through differentiation of existing (pre-)adipocytes. UCP1 levels increase per unit of RNA and protein (and thus total tissue UCP1 increas-es). Between room temperature and colder temperatures, hyperplasia (prolifera-tion) appears to be the predominant mechanism of recruitment. In this thermal zone, UCP1 does not increase per unit of RNA or protein, but the total number of UCP1-containing cells increases, and thus total tissue UCP1 is increased).

33
gave rise to UCP1+ adipocytes in interscapular BAT of cold-exposed mice (Lee et al., 2015).
Combined, the current literature implies that proliferation (hyperplasia) in interscapular BAT is an important contributor to the thermogenic recruit-ment between room temperature and cold (4-6 °C). Of interest would be to use the same models to investigate the absence or presence of a proliferative response in BAT during the transition from thermoneutrality to room tem-perature.
Furthermore, to get a better understanding of BAT heterogeneity, it re-mains to be explored whether other classical BAT depots respond to cold in a similar way.
3.2 White adipose remodeling in varying thermal conditions Another question to be asked is whether the observed induction of brown-
like cells in WAT depots occurs through a proliferation- or differentiation-dependent mechanism and whether this is also temperature-dependent.
3.2.1 Varying degrees of cold-induced proliferation in white adipose depots The first study to look into cold-induced cellular dynamics in WAT de-pots found no evidence of cold-induced hyperplasia in rat periovarian and inguinal WAT over a time period of 9 days (Cousin et al., 1996). Similar results were obtained in ingWAT of mice as indicated by expression analysis of proliferation markers (Barbatelli et al., 2010; Lee et al., 2015) and BrdU1 incorporation after 3 days at 4 °C (Lee et al., 2015). Similar to inguinal WAT, retroperitoneal WAT did not display much proliferation (i.e. EdU-incorporation1) upon cold exposure. Interesting, however, was the observa-tion that proliferation in retroperitoneal WAT, interscapular WAT, and in particular gonadal WAT, were increased after 3 days at 4 °C (Lee et al., 2015). However, the EdU-incorporating cells, like in the retroperitoneal and interscapular WAT depots, were negative for CD31, F4/80 and PDGFR , making the nature of the proliferating cells unclear (Lee et al., 2015) (but also see (Lee et al., 2013), which does report cold-induced proliferation of PDGFR + cells in gonadal WAT). Whether any of these proliferating cells eventually formed (UCP1-expressing) adipocytes was not reported (Lee et al., 2015). However, treatment with the 3-adrenergic receptor agonist
1 BrdU (bromodeoxyuridine) and EdU (5’-ethynyl-2’-deoxyuridine) are synthetic thymidine analogs that are incorporated into the DNA during cell replication. BrdU and EdU can be made detectable using a fluorescent antibody (for BrdU) or a fluorescent substrate reaction (for EdU).

34
CL316,243 does lead to UCP1-expressing adipocyte formation from prolif-erating PDGFR + cells (Lee et al., 2013; Lee et al., 2012). The above observations imply varying proliferative responses in different white fat depots in response to cold. The observation that the largest prolif-erative response was observed in gonadal WAT – the depot with typically the lowest cold-induced browning – implies that in WAT depots prolifera-tion is not necessarily linked to browning. However, more dedicated experi-mental evidence exists to draw conclusions about proliferation and brite/beige adipocyte formation.
3.2.2 Brite/beige adipocyte formation: interconversion One of the questions about brite/beige adipocytes is how they are formed and from which cells. Currently, two models are mainly considered. First, interconversion (or transdifferentiation) suggests that brite/beige adipocytes are formed from already existing, differentiated adipocytes that have a uni-locular white-like appearance (Figure 5, points 3 and 4). Second, de novo formation implies brite/beige adipocyte formation through the differentiation or maturation of cells that were in an undifferentiated precursor state (Figure 5, point 2). As will become clear from the following, current evidence im-plies that both models (co-)exist. As described in the previous paragraph, little proliferation is observed in browning WAT as a result of cold exposure. Analysis of BrdU-incorporation into UCP1+ brite/beige cells in (ing)WAT generally indicates that brite/beige adipocytes do not directly derive from cells that had previously undergone proliferation (i.e. UCP1+ cells are negative for BrdU) (Lee et al., 2015). Additionally, fate-mapping of mature Adiponectin-expressing adipo-cytes implied that all multilocular UCP1-expressing cells in ingWAT were already in a differentiated state before cold exposure (Lee et al., 2015) (Fig-ure 5, points 3 and 4). Adding to the interconversion hypothesis are data indicating what happens to brite/beige cells when the cold stimulus is re-moved (Rosenwald et al., 2013). This study made use of a mouse model that allowed visualization of cells that had expressed UCP1 previously and cells that were expressing UCP1 at the time of analysis (Rosenwald et al., 2013). This model made it possible to follow brite/beige cells (induced at 4 °C) after the mice were put back at room temperature. Interestingly, cells that had expressed UCP1 in the cold acquired a unilocular white-like adipocyte phenotype and did not dedifferentiate or get eliminated through apoptosis (Rosenwald et al., 2013). Then, when mice were exposed to cold for a se-cond time, approximately 75 % of the second round of brite/beige cells had been labeled during the first round of cold exposure (i.e. those 75 % were “former brite cells”). This observation implies some sort of “memory” of their earlier brite/beige phenotype (Rosenwald et al., 2013). It might thus be that brite/beige cells are intrinsically different from their neighboring cells

35
that do not turn brite. Although this model provides clues about program-ming of brite cells after an initial cold stimulus, it does not provide evidence about how brite adipocytes were formed during the first round of cold expo-sure (i.e. through interconversion or de novo).
3.2.3 Brite/beige adipocyte formation: de novo formation Where the models above imply that brite/beige cells can form through interconversion of white-like adipocytes, studies with a technically different tracing model implied that brite/beige adipocytes can also originate from immature (i.e. Adiponectin-negative) cells (Wang et al., 2013). The observa-tion that brite/beige adipocytes mostly do not show signs of proliferation (e.g. BrdU incorporation) does not necessarily imply interconver-sion/transdifferentiation; newly formed brite/beige adipocytes might have resulted from differentiation of non-proliferating precursors (Figure 5, point 2) (Wang et al., 2013). Further evidence in favor of de novo formation of brite/beige adipocytes, comes from mouse models in which various cell types are labeled in different ways (Berry et al., 2016). Tracing of PPAR -expressing cells during 7 days of exposure to 6 °C, indicated that 30% of UCP1+ brite/beige adipocytes (in ingWAT) were derived from proliferating cells after the start of cold exposure (Berry et al., 2016). This observation indicates that during the first 7 days of cold exposure, a subset of brite/beige adipocytes is formed by differentiation of proliferating PPAR -expressing cells that have been described as perivascular mural cells (Berry et al., 2016; Tang et al., 2008). Additional mouse models in which mural cells can be traced (e.g. by tracing cells expressing Smooth Muscle Actin (SMA)), fur-ther added to the notion that brite/beige adipocytes have an origin in imma-ture perivascular cells (Berry et al., 2016). Importantly, whereas several studies described the interconversion of mature (Adiponectin-expressing, white-like) adipocytes into brite/beige adi-pocytes, Berry and colleagues reported different observations with similar mouse models (Berry et al., 2016). Labeling of Adiponectin-expressing cells or aP2-expressing cells implied that approximately 60 % of cold-induced brite/beige adipocytes were not derived from already-mature adipocytes. This observation contrasts greatly with that obtained in a very similar tamox-ifen-inducible cell-labeling model in which, after 7 days of cold exposure, 100 % of UCP1+ adipocytes were derived from Adiponectin-expressing mature cells (Lee et al., 2015). One of the discrepancies in these studies is that the Adipoq-CreERT2 mice that were used in these two studies were generated independently and the reporter alleles were also different. Also, the duration of tamoxifen treatment was different (5 consecutive days (Lee et al., 2015) and two consecutive days (Berry et al., 2016), prior to cold expo-sure). Whatever the underlying reason for the contrasting observations in

36
these two studies may be, it shows that this part of adipose biology and its tools require further refinement.
3.2.4 Brite/beige versus white predetermination Based on the current body of data, cold-induced brite/beige adipocytes are likely formed through a combination of interconversion of existing mature (white-like) adipocytes and through differentiation of immature precursor cells associated with the vasculature (Figure 5). The question that remains is whether interconverting adipocytes and differentiating precursors are prede-termined to become brite/beige adipocytes, or whether they are intrinsically identical to “real” white (pre-)adipocytes, but simply depend on receiving the right stimulus (i.e. norepinephrine).
Several studies have implied that various cell populations exist within WAT that have different browning capacities (Wang et al., 2014; Wu et al., 2012). When making adipogenic clonal cell lines from the stromal-vascular fraction of inguinal WAT, it was observed that some, but not all, clonal cell lines retained intrinsic browning capacity (as determined by the induction of Ucp1 mRNA in response to the -adrenergic receptor agonist isoproterenol) (Wu et al., 2012). Microarray-based gene expression analysis of the adipo-genic clonal cell lines (treated with forskolin1) revealed two separate clusters of cell lines. Of these two clusters of inguinal WAT-derived clonal cell lines, one had a gene expression pattern that was more similar to brown adipose clonal lines than to the other inguinal clonal lines (Wu et al., 2012). Fur-thermore, this analysis revealed a set of genes that could potentially allow distinction of brite/beige and white (and classical brown) (Wu et al., 2012) (further discussed in chapter 5). Importantly, this study implied the existence of separate populations between potentially brite/beige and non-brite/beige precursors. Additional results that argue for a predetermined brite/beige capacity came from a study that identified EBF2 (Early B cell factor 2) as a marker for embryonic classical brown pre-adipocytes (see paragraph 4.2.3) (Wang et al., 2014). EBF2, among PDGFR + cells in the stromal-vascular fraction of ingWAT, also marked a population of pre-adipocytes that, in vitro, displayed increased expression of functional brown genes (e.g. Ucp1) (Wang et al., 2014). This population of EBF2+;PDGFR + cells increased from 5% of the stromal-vascular fraction at thermoneutrality to 13% after 3 days of cold exposure. Although this increase could be due to changes in the abundance of other cell types in the tissues, it may imply a proliferative response of this brite/beige precursor pool. Furthermore, EBF2 is required for the browning of ingWAT by CL-316,243 treatment (Stine et al., 2016). Although EBF2
1 Forskolin is used to mimic adrenergic stimulation by increasing cellular cyclic AMP levels through activation of adenylate cyclase.

37
enhances the brite/beige phenotype in adipocytes, it remains to be shown whether EBF2- cell populations in vivo, in response to cold, are really una-ble to become functional brite/beige adipocytes. Another study implying that brite/beige adipocytes are intrinsically dif-ferent from neighboring white adipocytes analyzed translated RNAs from Ucp1+ and Adiponectin+ adipocytes (i.e. brite/beige and all adipocytes, respectively) from various adipose depots (Long et al., 2014). RNA-sequence analysis revealed that in inguinal WAT, expression of a set of smooth muscle-associated genes was enriched in Ucp1+ cells as compared to the whole pool of adipocytes (i.e. Adiponectin+ cells). In cell-tracing exper-iments using the smooth muscle marker Mysoin heavy chain 11 (Myh11) as a driver, the authors found that 10 % of all UCP1-expressing cells had a smooth muscle origin (i.e. Myh11+) (Long et al., 2014). This was later con-firmed in an independent study (Berry et al., 2016). Almost no UCP1-expressing adipocytes in BAT were labeled by the Myh11-reporter (Long et al., 2014). The smooth muscle-associated genes may be expressed selectively in a small subset of brite/beige adipocytes, but clearly not in all. This implies that among brite/beige adipocytes, heterogeneity exists with regard to the relation with smooth muscle cells. Whether these two populations differ in other (functional) ways is still unknown.
Figure 5. Brite/beige adipocytes can be derived from various cell types. 1-2) At the precursor stage, white and brite/beige fates might have already been estab-lished. Selection for CD137, TMEM26 (Wu et al., 2012) or EBF2 (Wang et al., 2014) would imply this. It is unclear whether the determined precursors are self-renewing stem cells or not. 3-4) Adipocytes have been shown to switch from multilocular UCP1+ to unilocular UCP1- states (Rosenwald et al, 2013). It is unclear whether all unilocular cells are identical or if they are preprogrammed to become brite/beige adipocytes (4).

38
3.3 Do classical brown and brite/beige cells use different signaling mechanisms to respond to cold? From the discussion above, it appears that brown and brite/beige for-mation depend on different underlying mechanisms (proliferation-dependent and –independent) and that this also depends on the thermal conditions (a shift from thermoneutrality to room temperature or to even colder condi-tions). Another observation, regarding the role of the 3-adrenergic receptor, initially suggested that brown and brite/beige responses to cold might be intrinsically different. However, as discussed below and in Paper VI, this is now debatable.
3.3.1 Requirement for different -adrenergic receptors in BAT In general, the thermogenic response to cold is considered to be primarily mediated through norepinephrine-induced signaling of the 3-adrenergic receptor ( 3-AR) (Cannon and Nedergaard, 2004; Zhao et al., 1998; Zhao et al., 1994). However, brown adipose function appears fully intact in mice lacking the 3-AR (Barbatelli et al., 2010; Jimenez et al., 2003; Mattsson et al., 2011; Revelli et al., 1997; Susulic et al., 1995). One observation, which seemingly has been ignored or missed, is the phenotype of BAT in 1/3-AR double knockout mice (Bachman et al., 2002). BAT of 1/2/3-AR triple knockout mice ( -less mice) has a characteristic unilocular white-like ap-pearance (Bachman et al., 2002). However, histological analysis of BAT of
1/3-AR double knockout mice appears normal and BAT weight is unaffected (Bachman et al., 2002). This implies that “the presence of 2-AR alone is sufficient for normal BAT morphology” (Bachman et al., 2002). Interesting-ly, the 2-AR has typically been disregarded because of an apparent lack of expression in brown adipocytes (Bengtsson et al., 2000; Cannon and Nedergaard, 2004) and because of the observation that a 2-AR antagonist does not affect norepinephrine-induced lipolysis or Ucp1 expression in cul-tured brown adipocytes (Chaudhry and Granneman, 1999), nor norepineph-rine-induced thermogenesis (Zhao et al., 1994). Other studies have suggested that cold-induced proliferation of precursors (see paragraph 3.1.2) is mediated by the 1-AR (Bronnikov et al., 1999; Lee et al., 2015), whereas the further maturation and functionality of already existing adipocytes is believed to be mediated by the 3-AR (Cannon and Nedergaard, 2004; Zhao et al., 1998; Zhao et al., 1994). Contrasting with this idea of the 1-AR driving proliferation and the 3-AR being responsible for norepinephrine-induced gene expression and thermogenesis, a study in-vestigating the effects of the absence of the 1-AR on BAT in mice implied that norepinephrine-induced BAT temperature is not as strongly induced in
1-AR knockout mice as in wild-type mice (Ueta et al., 2012). However, UCP1 protein levels do not appear to be different in BAT of 1-AR knockout

39
mice fed a chow or high-fat diet (Ueta et al., 2012). Those reported UCP1 levels were relative (i.e. UCP1 per unit protein), and thus are not informative for the total UCP1 amounts per depot. Together, it seems that the functional regulation of BAT by norepineph-rine is transferred by a complex system of combined actions of adrenergic receptors. Observations regarding the actions of adrenergic receptors studied in knockout models or in studies with receptor antagonists, could be further complicated by potential “compensatory” functioning from other (adrener-gic) receptors (as suggested e.g. by (Chernogubova et al., 2005; Mattsson et al., 2011)).
3.3.2 Brite/beige adipocyte formation in 3-adrenergic receptor knockout mice Although BAT of 3-AR knockout mice appears normal, two studies re-ported defects in the formation of brite/beige adipocytes (Barbatelli et al., 2010; Jimenez et al., 2003). Upon cold exposure, formation of multilocular cells and expression of cold-inducible genes were severely blunted in subcu-taneous and visceral white (but not brown) depots of 3-AR knockout mice (Barbatelli et al., 2010; Jimenez et al., 2003). The observation of the effects in the white adipose depots implies the absence of (sufficient) compensatory mechanisms in these depots. The observation that classical brown and brite/beige adipocytes were affected differently in the 3-AR knockout mice prompted us to further in-vestigate this difference. In Paper VI, we describe how we aimed to charac-terize the 3-AR-dependence for cold-induced gene expression throughout the adipose organ and further characterize fat depots as classical brown or brite/beige. Surprisingly, however, we did not observe any differences in BAT and WAT of 3-AR knockout compared to wild-type mice (Paper VI). Therefore, we could not perform our brown-versus-brite/beige characteriza-tion. This result did, however, open up the discussion about the role of the
3-AR in brite/beige adipocytes in different mouse models as described be-low.
3.3.3 Formation of brite/beige adipocytes in different mouse strains Although the following paragraph will be mostly speculative, some pre-liminary supportive data will be presented as well. We have observed that
3-AR knockout FVB/N mice display normal cold-induced thermogenic gene expression in all tissues analyzed (Paper VI). 3-AR knockout mice on the C57BL/6 or 129Sv background display impaired cold-induced thermo-genic gene expression, selectively in brite/beige depots (Barbatelli et al., 2010; Jimenez et al., 2003). For this reason, we asked whether the cellular

40
mechanisms that contribute to brite/beige adipocyte formation (described in paragraph 3.2.2) could be different between these mouse strains. In classical BAT, cold-induced proliferation of precursors is mediated by the 1-AR (Bronnikov et al., 1999; Lee et al., 2015), whereas the further maturation and functionality of already existing adipocytes is believed to be mediated by the
3-AR (Cannon and Nedergaard, 2004; Zhao et al., 1998; Zhao et al., 1994). It could thus be hypothesized that the same is true for brite/beige (pre-)adipocytes: the proliferative response of precursors might depend on the 1-AR, whereas interconversion of already existing adipocytes is 3-AR-dependent. If we now consider the possibility that the relative contributions of de novo formation and interconversion vary in different mouse strains, we could explain the different phenotypes of the 3-AR knockout models in the different mouse strains.
In interscapular BAT, Ucp1 expression at 24 °C and upon acute (48 h) cold exposure is very similar across different mouse strains (Figure 6A) and (Ferrannini et al., 2016). However, looking at Ucp1 gene expression in in-guinal WAT in response to this acute cold (Figure 6B), we saw that wild-type C57BL/6 and 129Sv mice have stronger fold-increase changes (13- and 10-fold, respectively) than wild-type FVB/N mice (only 2.6-fold) (Figure 6B). Although these data are very preliminary, they provide a first hint indi-cating that in C57BL/6 and 129Sv mice, in response to acute cold exposure, rapid interconversion ( 3-AR-dependent) could be the dominant britening mechanism, whereas the slower de novo formation from proliferating pre-cursors ( 1-AR-dependent) could be the dominant mechanism in FVB/N mice. However, although this might be true for wild-type mice, it does not explain through which mechanism FVB/N 3-AR knockout mice acquire and maintain full thermogenic gene expression in mature brite/beige adipocytes. Which signaling mechanisms are responsible in FVB/N mice – and whether this is really different in other mouse strains such as C57BL6 and 129Sv – has remained unclear (Paper VI). Furthermore, irrespective of the signaling mechanisms, the notion that the brite/beige phenotype varies much more among strains than the phenotype of classical brown adipocytes has been around for some time (Guerra et al., 1998; Xue et al., 2007). Whether these various strains really differ in the mechanisms underlying the formation of brite/beige cells should be further investigated. For proper analysis of de novo formation and interconversion, one would have to cross the existing tracing models to each of these backgrounds, which would be an enormous effort. A more realistic approach would be to perform EdU or BrdU labeling experiments in the three mouse strains in cold exposure ex-periments. Although the absence of EdU/BrdU incorporation into brite/beige adipocyte does not necessarily indicate the absence of de novo formation (Wang et al., 2013), this experiment has the potential to provide initial clues about the relative contributions of the two browning mechanisms in response to cold in different mouse strains.

41
The outcome of such experiments could affect how we should eventually approach the recruitment of brown or brite/beige adipocytes in humans. If genetically different mouse strains indeed display a dependence on different browning mechanisms, the same possibility should be considered for hu-mans.
3.4 Diet-induced dynamics of brown and white adipose tissues From the above paragraphs, it has become clear that various adipose de-
pots respond differently to varying temperatures. Another strong, often stud-ied parameter that affects adipose biology is dietary energy intake. The amount and the composition of the diet can dramatically influence the adi-pose organ. In the following paragraphs, variation among adipose depots in response to changes in dietary amount and composition will be discussed to further characterize differences between adipose tissues and cells.
3.4.1 Hyperplasia and hypertrophy In rodent experiments, the effects of excess caloric intake are typically assessed by feeding animals a control (chow) diet or a so-called high-fat diet for a few weeks. High-fat diet feeding readily results in an increase of body
Figure 6. Ucp1 expression normalized to TFIIB in ingWAT (A) and iBAT (B) in FVB, C57BL/6 and 129Sv wild-type female mice kept at 24 °C or exposed to 4 °C for 48 h. Presented values are mean ± S.E.M. Statistical significance was deter-mined by Student's t-test. * indicates temperature effect; P < 0.05. n = 3 (FVB and C57BL/6) or 4 (129Sv).

42
weight, which can be explained in large part by increased fat weight. In gen-eral, two mechanisms can be discerned as to how fat depots adapt to in-creased availability of energy. One way is through hyperplasia (an increase in the total number of adipocytes). The other mechanism, hypertrophy, com-prises the individual expansion of the existing adipocytes (Figure 7). Recent studies have pointed out that subcutaneous and visceral depots respond differently to high-fat diet feeding in male mice (Jeffery et al., 2015; Wang et al., 2013). Cell labeling experiments indicate that, in male mice, upon high-fat diet feeding, newly formed adipocytes appear in visceral fat, whereas almost no newly formed adipocytes are observed in subcutaneous WAT (Jeffery et al., 2015; Wang et al., 2013) (Figure 7). In female mice, a proliferative response is also observed in subcutaneous WAT (Jeffery et al., 2016) (Figure 7). The observed sex-dependent difference seems to be influ-enced, at least in part, by sex hormones. Ovariectomized females displayed cellular responses to high-fat feeding more similar to male mice. Alternative-ly, estrogen injection in male mice resulted in an enhanced proliferative re-sponse in inguinal fat in response to high-fat diet (Jeffery et al., 2016). Given the wide use of the estrogen receptor modulator tamoxifen in cell labeling models, effects of this compound and the role of estrogen in adipogenesis should be carefully taken into consideration and controlled for (Ye et al., 2015).
Within the first 4 days of high-fat feeding, adipocyte precursors (i.e. Lin-;CD34+:CD29+;Sca1+) in visceral WAT display increased BrdU incorpora-tion (i.e. proliferation), after which proliferation rates fall back to basal lev-els (Jeffery et al., 2015). However, not until after 4 weeks do mature adipo-cytes form from these proliferating adipocyte precursors (Jeffery et al., 2015). Thus, it appears that a very rapid proliferative response (already with-in 12 h) precedes a delayed differentiation (after more than 4 weeks) (Jeffery et al., 2015; Wang et al., 2013). These data clearly show how, at least in male mice, two different WAT depots display entirely different dynamics in response to a dietary stimulus. Interestingly, the hyperplastic and hypertrophic responses are not cell-intrinsic, but depend on the microenvironment (Jeffery et al., 2016). In male mice, when precursor cells from epididymal fat were transplanted into ingui-nal fat pads, the proliferative response to high-fat diet was absent. Converse-ly, precursors from inguinal WAT transplanted into epididymal WAT dis-played proliferation upon high-fat feeding (Jeffery et al., 2016). Which com-ponents of the tissue microenvironment play a role in the depot-specific reg-ulation of adipogenesis is presently unknown. Differences in the extracellular matrix composition have been observed and may provide clues in this context (Roca-Rivada et al., 2015). Furthermore, differences in sym-pathetic innervation between subcutaneous, retroperitoneal and mesenteric WAT have been reported (in rats and Syberian hamsters, respectively) (Adler et al., 2012; Nguyen et al., 2014). Of particular interest is the obser-

43
vation that relative neural projections into retroperitoneal WAT and inguinal WAT differ in male and female rats (Adler et al., 2012). The observation that in male rats, fewer neurons projecting into inguinal and retroperitoneal WAT expressed estrogen receptor compared to female rats (Adler et al., 2012). Thus, sympathetic innervations might be involved in the depot- and sex-specific regulation of adipogenesis. It will be of interest to know whether human visceral and subcutaneous depots display the same tissue dynamics in response to diet and whether this is different in men and women. One study concluded that, indeed, the weight of the human visceral major omental depot could be explained predominant-ly by adipocyte number, and to a lesser extent by adipocyte volume (ana-lyzed in women) (Arner et al., 2013). However, even between two subcuta-neous WAT depots (abdominal and femoral) differences in hyperplasia and hypertrophy can be observed (Tchoukalova et al., 2010). Upon overfeeding, femoral WAT responded with increased adipocyte number (hyperplasia),
Figure 7. White adipose tissue dynamics in response to high-fat diet feeding. Up-on high-fat diet feeding, male pgWAT responds through hyperplasia (CD24+ pre-cursors) and hypertrophy (top), whereas male ingWAT responds only through hypertrophy. In female mice, both depots display combined hyperplasia and hyper-trophy. Based on data presented in Jeffery et al., 2015 and 2016 and Wang et al.,

44
whereas abdominal subcutaneous adipocytes increased in size (hypertrophy), both in men and women (Tchoukalova et al., 2010). Whether the hyperplas-tic or hypertrophic nature of the WAT depots relate to their contributions to overall metabolic homeostasis remains to be determined.
3.4.2 Diet-induced BAT recruitment Brown adipose tissue has taken a central place in the field of diet-induced obesity and counteracting mechanisms. An important parameter measured in such studies is the so-called diet-induced thermogenesis (Rothwell and Stock, 1979): the recruitment of brown fat function in response to diet. Without going into the details of diet-induced thermogenesis, this paragraph describes the cytological dynamics of brown adipose tissue in response to various diets.
To really experimentally isolate the effects of diet, any influences of tem-perature should be omitted. Therefore, only studies conducted at thermoneu-trality are really informative about the diet-specific effects on BAT function and recruitment. However, although it is well established that room tempera-ture is a cold stress for rodents, the vast majority of metabolic studies are still not performed at thermoneutrality. Thus far, cellular dynamics in response to diet, such as those described in paragraph 3.4.1 for white fat, have only scarcely been reported for brown fat. Thus, without any direct evidence, one would have to obtain information about the recruitment mechanisms by deducing this from data describing tissue weights, tissue DNA/RNA/protein contents and UCP1 levels (total and per unit of RNA/protein). It is well established that in rodents, obe-sogenic diets result in increased brown adipose tissue weight (e.g (Rothwell and Stock, 1979)). As an example, short-term (30 days) and long-term (7-8 months) high-fat diet feeding of wild-type mice at thermoneutrality results in increased BAT weight (e.g. (Feldmann et al., 2009). This is mostly the result of increased lipid accumulation in the tissue. Additionally, long-term high-fat diet feeding at thermoneutrality also results in increased protein amounts in BAT (unpublished observations by Dr. N. Petrovic). However, this in-crease in protein content is not yet observed after 6 weeks of high-fat diet feeding at thermoneutrality (Feldmann et al., 2009). Also, in response to both short- and long-term high-fat diet feeding, UCP1 protein levels (per mg protein) increase, resulting in both cases in increased total amounts of UCP1 in interscapular BAT (Feldmann et al., 2009). Based solely on the protein content in interscapular BAT reported in the two studies discussed above, it may be speculated that the initial short-term (at least up to 6 weeks) response to high-fat diet is that of differentiation (increased UCP1 per mg protein, but no increase in total protein) (Feldmann et al., 2009). With extended exposure to high-fat diet, total tissue protein is increased (indicative of hyperplasia), but only a (statistically insignificant)

45
tendency to increased UCP1 per mg protein is observed (indicating little contribution of differentiation) (unpublished observations by Dr. N. Pe-trovic). The tissue dynamics suggested above are only speculative at this point. However, a study in rats (performed at 25 °C) indicated that “cafeteria” diet-feeding for 10 weeks resulted in a doubling of the number of brown adipo-cytes (Bukowiecki et al., 1982). This supports the notion that increased ca-loric intake indeed induced hyperplasia in BAT. However, studies directly aimed at analyzing proliferative responses and labeling of newly formed adipocytes, as has been performed in white fat (e.g. (Jeffery et al., 2015)), are needed to elucidate cellular dynamics in BAT in response to obesogenic diets.
3.4.3 Marrow adipose tissue dynamics As described in paragraph 1.5.1, marrow adipose tissue can be broadly
divided into two types: constitutive marrow adipose tissue and regulated marrow adipose tissue. The changes described below in marrow adiposity typically take place in areas of regulated marrow adipose tissue. Where the expansion of classical white adipose depots typically takes place in response to caloric excess, a reduction occurs in response to limited calor-ic availability. Marrow adipose tissue dynamics are not as intuitive as those observed in peripheral white adipose tissues. Indeed, in response to high-fat diet feeding, marrow adipose tissue increases in size (Doucette et al., 2015). However, counterintuitively, marrow adipose tissue also expands under ca-loric restriction (Cawthorn et al., 2014; Cawthorn et al., 2016; Devlin et al., 2010).
Because of its similar responses under contrasting physiological condi-tions (e.g. caloric excess versus restriction), it is difficult to determine what signals drive marrow adipose expansion and reduction. As an example, con-trasting effects of Leptin on marrow adiposity have been observed (Scheller et al., 2016). Several lines of evidence suggested that centrally administered Leptin inhibits bone marrow adiposity (Ambati et al., 2010; Hamrick et al., 2007). In contrast, a study in which the Leptin receptor was deleted selec-tively in limb bone marrow cells (using a Prx1-driven Cre) showed that this deletion resulted in decreased marrow adiposity (Yue et al., 2016). Further investigations should indicate how Leptin, centrally or directly on marrow cells, affects marrow adiposity under different physiological conditions. Another interesting observation regarding bone marrow and (peripheral) adipose tissue came from a study with patients that had undergone a bone marrow or peripheral blood stem cell transplantation (Ryden et al., 2015). DNA analysis of single peripheral adipocytes revealed that, in patients who had received a transplantation, donor-derived DNA was observed (Ryden et al., 2015). Over the entire remaining lifespan, donor-derived adipocytes

46
could contribute up to 10% of the subcutaneous adipocyte population. Inter-estingly, adipocytes were observed that either contained recipient DNA, donor-derived DNA or mixed DNA from both recipient and donor, implying cell fusion (Ryden et al., 2015). In mice, some experimental evidence exists for the contribution of bone marrow-derived peripheral adipocytes (Crossno et al., 2006; Majka et al., 2010), but negative findings also exist (Koh et al., 2007; Tomiyama et al., 2008). Thus, the contribution of bone marrow-derived cells to peripheral adipose tissues and its physiological significance remains to be further inves-tigated.
3.5 Intrinsic (pre-)adipocyte differences: ex vivo responses to serum Much of the adipose tissue dynamics discussed above come from in vivo experiments and thus mostly provide insights into tissue dynamics. Modern cell tracing models now allow visualizing individual cell populations within the context of the whole tissue. However, investigations to further study cell-autonomous behavior still depend heavily on ex vivo experiments. Without going into the details here of adipose cell culture systems, this paragraph will discuss some observations made in our lab in cell culture systems that indi-cate differences intrinsic to the (pre-)adipocytes from various adipose de-pots. The motivation to include this paragraph comes from personal observa-tions presented in Paper VIII. A large part of the experimental work focused on how precursor cells from various mouse adipose depots behave ex vivo. Here, ex vivo refers to the system in which the stromal-vascular fractions of adipose depots are allowed to grow and differentiate in a cell culture dish. This type of cell culture is also referred to as primary cell cultures. As described in the Materials and Methods sections of Papers VII and VIII, the cells are grown in a medium that contains 10% of serum derived from bovine animals. This is typically fetal bovine serum (FBS), but can also be from newborn calves (newborn calf serum, NCS). One of the most strik-ing observations made in cell culture was how cells derived from various adipose depots responded differently to the type of serum in which they were cultured. As described in Paper VII, brown pre-adipose cells could convert into adipocytes equally well in either type of serum. Among white adipose cell cultures, responses to serum type differed dramatically. Cells derived from white adipose depots displayed various responses to newborn and fetal serum (Paper VIII). This in itself shows the intrinsic differences among adi-pose cells located in various depots in mice. In addition to our own observations, different degrees of adipogenic ca-pacity have been reported in vitro for cells derived from different fat depots in mice (e.g. (Emont et al., 2015)) and in humans (reviewed in (Tchkonia et

47
al., 2013). However, the vast majority of cell culture studies are performed in immortalized 3T3-L1 cells. These cells were originally obtained from Swiss mouse late embryos and spontaneously immortalized in culture. Thus, to which depot these 3T3-L1 cells originally belonged is unknown. There-fore, in the light of the heterogenic nature of adipose precursors from various depots described above, interpretation and extrapolation of results obtained in 3T3-L1 cells should be done with care. The numerous studies discussed in this chapter collectively indicate a number of differences between and within adipose tissue types. First, within one adipose depot, cellular dynamics can differ depending on the type of environmental stimulus. As appears to be the case within interscapular BAT, different thermal zones induce different cellular responses (i.e. hyperplasia or hypertrophy) (Paper V). Furthermore, even within the interscapular BAT depot, regional differences exist in which cellular dynamics vary (e.g. the dorsal localization of cold-induced proliferation in interscapular BAT) (Lee et al., 2015). However, whether other BAT depots display different or simi-lar dynamic behavior has not yet been investigated. Second, the cold-induced formation of brite/beige adipocytes appears to occur in different ways. Both interconversion and de novo formation may occur in parallel within the inguinal depot (paragraph 3.2). The relative im-portance of each mechanism may well differ among various mouse strains and between adipose depots. Third, the requirement for the 3-AR for cold-induced expression of thermogenesis-related genes has been described to differ between classical brown and brite/beige adipocytes (Barbatelli et al., 2010; Jimenez et al., 2003). However, our own observations in another mouse strain indicate that brown and brite/beige adipocytes are not different in this respect (Paper VI). Thus, among mouse strains, there seem to be differences in signaling mech-anisms in brown and brite/beige cells in response to cold. Fourth, in response to high-fat diet feeding, cellular dynamics differ in gonadal and inguinal WAT depots in male mice, but not in female mice (Jeffery et al., 2015; Jeffery et al., 2016). In male mice, these differences depend on the microenvironment of the tissues, implying differences be-tween the depots rather than in the (pre-)adipocytes themselves. Finally, our own in vitro results with regard to the potential to differenti-ate in the presence of different serum types in the medium imply cell-intrinsic differences between cells from various adipose depots in mice (Pa-per VIII). Thus, the cellular dynamics in response to various physiological stimuli, both in vivo and in vitro, add to the overall view of heterogeneity among adipocytes, adipose depots in both sexes and in animals with different genet-ic backgrounds.

48
4. Developmental heterogeneity
As discussed in the previous chapter, various adipose depots and cells dis-play different dynamics in response to external stimuli. To add to our under-standing of how adipose cells and tissues are so inherently different, it helps to look at their differences in terms of developmental timing and cellular origin. This chapter describes the developmental timing of the formation of various adipose depots before and around birth. Furthermore, the cellular lineages from which different types of adipocytes derive will be discussed.
4.1 Perinatal development of adipose tissues Different stages of life have different demands for the functions of adipo-cytes (e.g. heat production or energy storage). Therefore, throughout life, functionally different adipocytes may appear (and disappear) at different stages. As described below, this is particularly apparent around the event of birth.
4.1.1 Brown adipose ontogeny Given the primary function of brown adipose tissue, heat production, its development should have taken place before an animal requires this function for the first time. This is typically considered to be the moment of birth. In older animals, acute exposure to cold is met with a shivering response, but the notion is that the shivering capacity of newborn animals is not yet suffi-ciently developed. Thus, brown adipose tissue must be fully developed at the time of birth. However, different mammalian species display variations in “maturity” at birth. Mice and rats, for example, are considered altricial ani-mals that are rather “helpless” and nest-dependent at the moment of birth. Altricial animals are born in large litters that huddle up and possibly also receive warmth from the mother. In altricial animals, maximum BAT ther-mogenic capacity is observed not directly at the moment of birth, but within the first week of life, probably at the time the animals move further away from the mother. This is paralleled by observation of Ucp1 expression, which starts to increase after birth depending on the environmental tempera-ture (Nedergaard et al., 1986; Obregon et al., 1989; Ricquier et al., 1986).

49
Precocial newborn animals (e.g. lamb and guinea-pig) that are born in a more mature, independent state, do show maximal thermogenic capacity at the moment of birth. Immature animals (e.g. hamsters, ground-squirrels and marsupials, are born underdeveloped. Such animals are unable, at this moment of birth, to induce BAT-dependent thermogenesis and respond behaviorally to thermic stress (Nedergaard et al., 1986). Thus, it is not the moment of physical birth (although it is this moment in precocial animals), but the moment of exposure to thermal stresses that de-mands fully recruited BAT. Although BAT development clearly differs among animals, the lab mouse and rat have been the ones most intensively studied in this context. The ini-tial indications about the developmental timing of BAT came from histolog-ical studies. Several studies in mice have shown that interscapular brown adipose tissue becomes apparent around embryonic day 14.5-15.5 (Atit et al., 2006; Hirning et al., 1989; Park et al., 2013; Schulz et al., 2013; Wang et al., 2014). The development starts with the formation of patches of pre-adipocytes that display a high rate of proliferation (as indicated by detection of the nuclear proliferation marker Ki67) (Park et al., 2013; Schulz et al., 2013). Lipid droplets become apparent around day 18 of embryonic devel-opment (Hirning et al., 1989; Park et al., 2013). The Ki67-indicated prolifer-ation decreases quickly upon birth (Schulz et al., 2013).
4.1.2 White adipose ontogeny Considering the primary function of white adipose tissue (energy stor-age), it can be expected to display a different developmental time line than brown adipose tissue. During the development of an embryo, all energy ob-tained can be expected to be utilized for growth, rather than being stored. However, as discussed here, this does not seem to be the case entirely. Although mouse embryonic development has been studied for a very long time, only with recent technical advancements has mouse adipose develop-ment become more visible. From a mouse model in which mature (i.e. Adi-ponectin-expressing) adipocytes can be followed over any chosen time frame, it has become clear that epididymal and inguinal adipocytes mostly develop during different developmental stages (Wang et al., 2013). Of the adipocytes present in inguinal adipose tissues of 28-day-old mice, basically all had already matured (i.e. expressed Adiponectin) before birth. This maturation started approximately around embryonic day 14 (Wang et al., 2013). Interestingly, only a few of the mature adipocytes in gonadal adipose tissues had matured before birth. More specifically, the vast majority of gon-adal adipocytes started maturing (i.e. expressing Adiponectin) later than 10 days after birth (Wang et al., 2013).

50
Together, the above observations visualized a striking difference between gonadal and inguinal adipocyte development. However, this model does not indicate whether pre-adipocytes were already present in the gonadal tissue and whether only the maturation stage was delayed in comparison to ingui-nal adipocytes. Whether the developmental timing relates to functional differences of the different WAT depots at these stages of life is unclear.
4.1.3 Human adipose development Similarly to mice (and many other species), human subcutaneous WAT is more developed at birth than visceral fat (Harrington et al., 2004). The first signs of adipose tissue development become evident at around 14 weeks of gestation (Poissonnet et al., 1983; Poissonnet et al., 1984). The adipose de-velopment was described as the initial formation of “mesenchymal lobules” that gradually develop into “primitive fat lobules” which already contain some lipid. These primitive fat lobules then further develop into “definitive fat lobules” representing adipose tissue as observed in infants and adults (Poissonnet et al., 1983; Poissonnet et al., 1984). The development of adipose tissues follows a cranial to caudal and prox-imal to distal gradient. The fat lobule development is preceded by the for-mation of vascular structures, implying the importance of angiogenesis for the development of adipose tissue (Poissonnet et al., 1983; Poissonnet et al., 1984). Much less information is available for the ontogeny of human BAT. One study claimed the detection of UCP1 in the “axial” BAT depot from 20 weeks of gestation (Velickovic et al., 2014). Earlier studies reported the presence of brown (i.e. multilocular) adipocytes in the interscapular region, around muscles and blood vessels in the neck region, under the clavicles and into the armpits. Brown adipocytes were also described in the thorax around the large vessels and in the abdomen around the aorta and mostly around the kidney and adrenals (Aherne and Hull, 1966). In some of these depots uni-locular cells were also observed. In samples of a 28-week-old fetus, the brown adipose tissue is described in terms of “immature adipose lobules” rich in blood vessels (Aherne and Hull, 1966) – a description similar to that for white adipose tissues (see above). From that moment, adipocytes in-crease in size, but probably not in number (Aherne and Hull, 1966).
4.2 Adipocytes derive from a variety of cellular origins For a long time, all adipocytes were considered similar in the way that they contain lipid droplets. Although the lipid droplet organization comes in a variety of flavors, all those lipid droplet-containing cells fell (or fall) in the

51
separate category adipocytes, distinguishing them from other cell types that do not typically (under non-pathological conditions) contain obvious lipid droplets. Studies of the embryonic cellular origins of adipose tissues changed the perspective of the inter-adipocyte relationships.
4.2.1 Interscapular BAT originates in the dermomyotome In 2006, Atit and colleagues investigated the fate of cells from the dorsal dermomyotome (Atit et al., 2006). The dermomyotome is the portion of so-mites that was initially thought to give rise to the dermis and skeletal muscle (Scaal and Christ, 2004). Within the dermomyotome, a group of cells ex-press the homeobox transcription factor Engrailed 1 (En1). To follow the fate of this group of En1-expressing cells, Atit et al. used a Cre-Lox mouse model (Atit et al., 2006). En1-positive cells express the Cre enzyme (from the En1-Cre allele) that induces the indelible activation of the LacZ gene, resulting in LacZ-signal in En1-expressing cells and all of its daughter cells (all generations, regardless of whether those cells still express En1 them-selves). In addition to dermis and skeletal muscle, cells of interscapular BAT were also labeled in this system. This indicated that iBAT was derived from the dermomyotome, sharing a cellular origin with muscle and dermis. Given that the authors concluded that the dermomyotome is tripotent, probably no white adipose cells were observed to be dermomyotome-derived. The relation be-tween brown adipocytes, muscle and dermis was also observed in an inde-pendent study (Wang et al., 2014). With regard to timing, it was observed that cells expressing En1 on day E9.5 labeled a subset of brown adipocytes observed on day E16.5, but not dermis or muscle. This indicates that (En1+) cells in the dermomyotome are primed on day E9.5 for a brown fat fate. This does not exclude the possibility that this “priming” occurs earlier, before En1-expression is established. In line with this observation, Lepper and Fan (2010) reported similar observations for Pax7-expressing dermomyotomal cells (Lepper and Fan, 2010). Togeth-er, these studies showed that BAT shares a dermomyotomal origin with dermis and muscle. Interestingly, Atit et al. noted, “the brown fat bundles in the most medial interscapular bundles and in the axillary depots were not labeled (not shown)”. This implies that the interscapular BAT depot contains cells of mixed origin and that the axillary BAT depot has a different cellular origin (En1-negative) than most of the interscapular BAT.

52
4.2.2 The brown fat-muscle relationship The observation that interscapular BAT is derived from cells that also give rise to skeletal muscle (and dermis) was later confirmed by a number of new studies. In 2007, our lab published a study in which extensive transcriptome anal-ysis was performed on cultured brown (combined interscapular, axillary and cervical) and white (epididymal) precursors (whole SVF) (Timmons et al., 2007). Among the genes enriched in brown versus white pre-adipocytes, a large set of genes belonged to the skeletal muscle Gene Ontology analysis class (Timmons et al., 2007). This implied that brown adipose cells share a cellular origin with myoblasts and that brown and (epididymal) white adi-pose cells have different developmental origins. Lineage tracing of cells known to give rise to muscle cells (i.e. cells that express the myogenic transcription factor Myf5) added another piece of evi-dence to the theory of a shared origin for myoblasts and brown adipocytes (Seale et al., 2008). Interscapular and perirenal BAT depots were both la-beled by a Myf5-induced YFP-reporter. However, whereas principally all interscapular BAT adipocytes were YFP-positive, perirenal BAT seemingly showed a mixture of YFP-positive and negative adipocytes (Seale et al., 2008). All analyzed WAT depots (gluteal WAT, inguinal WAT, and epidid-ymal WAT), were not derived from the Myf5+ lineage (Seale et al., 2008). Although Myf5 initially seemed to be a way to clearly distinguish be-tween brown and white adipocytes, this assumption did not hold over time. More thorough analysis of the Myf5 lineages throughout the adipose organ revealed that classical brown adipocytes exist that do not derive from the Myf5-lineage (Sanchez-Gurmaches and Guertin, 2014). In the cervical BAT depot, only approximately half of the adipocytes are derived from the Myf5-lineage. Furthermore, almost none of the brown adipocytes in perirenal BAT and periaortic (or mediastinal) BAT were labeled in the Myf5-lineage trac-ing model (Sanchez-Gurmaches and Guertin, 2014). Thus, some, but not all, brown adipocytes in mice originate from the same Myf5-expressing lineage.
4.2.3 Subsets of white adipocytes share the BAT/muscle origin Because of the initially apparent selective labeling of brown versus white adipocytes using a Myf5-driven Cre-Lox reporter system, this was later used several times in attempts to selectively manipulate the brown adipocyte line-age. In the first example, the PTEN gene was deleted from Myf5+ cells and their progeny with a knock-in floxed PTEN allele (Myf5-Cre;PTENfl/fl) (Sanchez-Gurmaches et al., 2012). Although the initial aim was to investi-gate the role of PTEN - an inhibitor of the PI3K signaling pathway that is involved in the adipogenic effects of insulin in vitro (Sanchez-Gurmaches et al., 2012) - in brown adipocyte development, the phenotype of the Myf5-

53
Cre;PTENfl/fl mouse resulted in new insights into adipocyte origins (Sanchez-Gurmaches et al., 2012). Deletion of PTEN in the Myf5 lineage resulted in a combination of lipomatosis (formation of lipomas) and partial lipodystrophy in which the adipose tissues were strikingly redistributed. The more anterior/dorsal adipose depots (interscapular, subscapular and cervical BAT, as well as interscapular and retroperitoneal WAT) increased in weight, whereas inguinal, mesenteric and epididymal WAT principally disappeared upon deletion of PTEN in the Myf5+ lineage (Sanchez-Gurmaches et al., 2012). Labeling of the Myf5-expressing lineage with a LacZ reporter showed that a subset of white adipocytes (in interscapular and retroperitone-al WAT) was indeed derived from the Myf5+ lineage. In contrast, almost no adipocytes in inguinal and gonadal WAT depots were labeled. More detailed investigation of the Myf5-lineage distribution throughout the adipose organ showed that Myf5+ and Myf5- lineages are heterogeneously distributed in BAT and WAT depots (Berry et al., 2016; Sanchez-Gurmaches and Guertin, 2014). More specifically, there is a tendency that dorsal/anterior depots are Myf5+ and the more posterior depots are Myf5- (Sanchez-Gurmaches and Guertin, 2014). Furthermore, progeny of cells expressing Pax3, another my-ogenic transcription factor that acts upstream or in concert with Myf5, are found in an even larger population of adipocytes, including perigonadal adi-pocytes in male, but not female mice (Sanchez-Gurmaches and Guertin, 2014). Interestingly, labeling of progeny of cells expressing MyoD1 – an-other myogenic transcription factor acting downstream of Myf5 and Pax3 - does not result in any labeled adipocytes (Sanchez-Gurmaches and Guertin, 2014). This implies that when activation of the muscle transcriptional pro-gram at the stage of MyoD1 expression occurs, the adipocyte-versus-muscle fate has been decided. Additional analysis of Myf5-expressing cells in developing interscapular BAT indicated that Myf5+ cells principally give rise to two populations of adipocytes. In mouse embryos on day E14.5, Myf5+-lineage derived cells that express the surface marker PDGFR , give rise in vitro to adipocytes that express brown fat functional genes at much higher levels than Myf5+-lineage cells that lack PDGFR expression (Wang et al., 2014). Expression analysis revealed Early B cell factor 2 (EBF2) as a key regulator of the brown phenotype in Myf5-derived PDGFR + brown pre-adipocytes (Wang et al., 2014). Thus, even within the Myf5+ population, two functionally different (pre-) adipose populations appear to exist, further emphasizing the heterogenic nature of the brown adipocyte population.

54
4.2.4 Do developmental origins distinguish classical brown from brite/beige? The developmental origin is being considered a means to distinguish be-tween various adipose cell types. When the myogenic lineage origin for BAT was established, an obvious question was whether brite/beige adipo-cytes also derive from this muscle lineage. Using the Myf5-tracing model and the 3-AR-agonist CL-316,243 to induce the formation of brite/beige adipocytes, Sanchez-Gurmaches et al., demonstrated that both Myf5+ and Myf5- multilocular (brite/beige) cells exist (Sanchez-Gurmaches and Guertin, 2014; Sanchez-Gurmaches et al., 2012). This Myf5+ or Myf5- origin corresponds to the origin of the surrounding white adipocytes; ingui-nal WAT (mostly Myf5-negative) contains Myf5-negative multilocular cells and retroperitoneal WAT (mostly Myf5-positive) contains Myf5-positive multilocular cells. Thus, the Myf5 and Pax3 lineages do not generally distin-guish between brown, brite/beige and white adipocytes. However, in vitro analysis of Myf5-lineage-derived and non-Myf5-lineage-derived adipose cells of the anterior subcutaneous WAT depot implied that expression of brown fat functional genes was higher in cells derived from the non-Myf5-lineage (Shan et al., 2013). Observations made in vitro in Pax3-lineage- and non-Pax3-lineage-derived cells from anterior subcutaneous WAT point in the same direction; muscle lineage-derived adipocytes in this depot display lower expression of brown fat functional genes (Liu et al., 2013a). Thus, although these in vitro observations imply some (negative) relation between an origin shared with muscle and adipocyte functionality within the anterior subcutaneous WAT depot, at this moment there has been no obser-vation of a clear distinctive origin characteristic for either brown or brite/beige adipocytes.
4.2.5 Prx1 selectively labels pools of subcutaneous adipocytes More selective labeling of a subset of adipose cells was established by using the gene Prx1 as a driver (Krueger et al., 2014; Logan et al., 2002; Sanchez-Gurmaches et al., 2015). Prx1-lineage tracing labels all adipocytes of the subcutaneous inguinal depot and about half of the adipocytes present in the anterior subcutaneous depot (Sanchez-Gurmaches et al., 2015). The Prx1-labeled ventral-dorsal gradient in anterior subcutaneous WAT opposes that of Pax3- and Myf5-labeled cells (Sanchez-Gurmaches et al., 2015). However, whether these two origins in this WAT depot are mutually exclu-sive was not addressed. Furthermore, in BAT located around the aorta (periaortic BAT or medias-tinal BAT), over 60% of adipocytes are derived from the Prx1-expressing lineage (Sanchez-Gurmaches et al., 2015).

55
Thus, labeling with Prx1 has added to our understanding of the lineage composition of the adipose organ, but we are still without a fully distinctive label or marker for various adipose subsets. However, the Prx1-Cre mouse model might serve as a tool to target ingWAT cells without affecting most, but not all, brown and visceral adipocytes. Marrow adipocytes, at least some, also appear to be targeted in the Prx1-Cre model (Krueger et al., 2014; Yue et al., 2016). Thus, as this model targets several populations of (pre-) adipo-cytes, metabolic observations would be difficult to be ascribed to one partic-ular adipose depot.
4.2.6 Wilms’ Tumour gene 1 selectively labels subpopulations of visceral adipocytes Whereas Prx1 rather selectively labels subcutaneous adipocytes, cells expressing the Wilms’ Tumour 1 gene (Wt1) were shown to give rise to visceral (i.e. epididymal, perirenal, retroperitoneal, epicardial, mesenteric and omental) adipocytes, but not subcutaneous white or interscapular brown adipose cells (Chau et al., 2014) (Figure 8). Labeling of Wt1-expressing cells of the lateral plate mesoderm on embryonic day 14.5, results in labeling of significant amounts of adipocytes in epididymal depots in mice over 1 year of age (Chau et al., 2014). However, Wt1 is not expressed in mature adipocytes, but rather in the stromal-vascular fraction and the mesothelium surrounding the adipose tissues.
4.3 Bone marrow adipocytes – a class apart With the increased interest in marrow adipose tissue, more studies have been dedicated to establish its developmental relation to other adipose de-pots. To this end, several cellular lineages were analyzed in mouse cell-tracing models (Berry et al., 2015; Chen et al., 2014; Liu et al., 2013b; Mizoguchi et al., 2014). The main shared observation is that all bone marrow adipocytes were labeled with an Osterix1-driven reporter. Osterix1 (Osx1) is a transcriptional regulator of osteoblast differentiation (Nakashima et al., 2002). No visceral, subcutaneous or brown adipocytes were derived from the Osx1-expressing lineage (Berry et al., 2015). Also, Myf5-driven labeling did not result in detection of any signal in bone marrow adipocytes. Thus, with regard to the analyzed cellular lineages, bone marrow adipocytes are an entirely isolated class of adipocytes and seem to share their lineage with osteoblasts as both cell types are derived from Osterix1-expressing cells.

56
Our knowledge of the cellular lineages that form the adipose organ has, to a major extent, developed during the past decade. The initial realization of the brown fat-muscle relationship spurred many studies looking into adipose cellular origins. This led to the finding that between and within adipose de-pots, adipocytes exist that can have different developmental origins. This makes it difficult, at this moment, to use this information to relate embryonic origin to adipocyte identity or function. However, it has clearly added to our understanding that the adipose organ is incredibly heterogeneous (Figure 8). With the advancements in mouse models to follow cellular fates through labelling, more insights have been acquired regarding the developmental timing of various adipose depots in mice. This information might become helpful to study defining events in adipose development and understand which processes are affected in pathological adipose development. The study of adipose development and origins has seen a promising start, but needs more work to further define the map of the origins of the adipose organ.

57
* Values were different in male and female mice and are here shown as male / female. ** Within the anterior subcutaneous depot, adipocytes are labeled in a dorsal-to-ventral gradient for both Pax3 and Myf5. A ventral-to-dorsal gradient is observed for Prx1-labeling. *** Microscopic evidence implies a high labeling percentage, but was not quantified (Chau et al., 2014).
Figure 8. Overview of the most commonly studied cellular lineages. Data indicate percentages of mature adipocytes derived from the indicated cellular lineages. Data are based on observations presented in Sanchez-Gurmaches et al. (2012, 2014, 2015), Chau et al. (2014), Atit et al. (2007) and Lepper and Fan (2010).
Cellular lineage
Tissue Pax3+ Myf5+ Prx1+ Wt1+ Other
cBAT 80 50 0 ?
iBAT 100 100 0 0 En1+, Pax7+
aBAT 100 100 0 ? En1-
mBAT 0 / 75* 0 65 ?
prBAT 70 0 0 ?
triWAT ? ? ? ?
isWAT 100 100 0 ?
asWAT 0-100** 0-100** 0-50 / 75** ?
ingWAT 0 0 100 0
cWAT ? ? ? 65
rpWAT 100 100 0 +***
prWAT ? ? ? +***
mWAT 0 0 0 30
pgWAT 50 / 0* 0 0 75

58
5. Molecular heterogeneity
Understanding the different natures of brown, brite/beige and white adipo-cytes will help to specifically target these cells therapeutically. Also, know-ing the relative abundance of the different adipocyte types will help guide therapeutic approaches. For these reasons, it is required to be able to reliably distinguish one adipocyte type from the others. To this end, much effort has been focused on finding genes that are expressed selectively in one adipose cell or tissue type as compared to the others. These selectively expressed genes are commonly referred to as marker genes.
To be of informative value, a proper marker gene would have to be ex-pressed at least one order of magnitude higher in one adipose cell or tissue as compared to another. Ideally, the presence or absence of a particular marker gene would reveal the identity of the analyzed cell or tissue. However, in reality as discussed below, the suggested marker genes that are used in our field often do not possess this selectivity. This chapter discusses the current status of marker genes and what they have taught us about brown, brite/beige and white identities in mice and humans.
5.1 Identification of marker genes The first genes that were suggested to distinguish between different adi-pose cell types came from a study that compared gene expression patterns in precursor cells derived from mouse interscapular brown fat and visceral (ep-ididymal) white fat (Timmons et al., 2007). The main finding of this study was that the gene expression “signature” of brown (but not white) preadipo-cytes showed great similarities with muscle cells. This led to the later finding that muscle cells and brown adipocytes are derived from the same precursor cell during development (see paragraph 4.2.2). Some years later, when brite/beige adipocytes started to become increas-ingly popular, two new studies used different adipose cell culture systems to identify new marker genes to distinguish between classical brown and brite/beige cells (Sharp et al., 2012; Wu et al., 2012). Wu et al. generated clonal cell lines from precursor cells obtained from mouse interscapular brown and subcutaneous (inguinal) white adipose tissue (described in para-graph 3.2.4). Multiple differentiated clonal cell lines were subjected to mi-croarray analysis to compare gene expression profiles. This resulted in a list

59
of marker genes that were differentially expressed between classical brown, brite/beige and white adipose clonal cell lines. Analysis of the expression of some of those brite/beige markers in human supraclavicular BAT and WAT (distinguished as such by UCP1 expression levels), led the authors to con-clude that human BAT gene expression more closely resembles murine brite/beige than murine classical brown (Wu et al., 2012). In a study with a similar aim, Sharp and colleagues used mouse primary adipose cultures from interscapular BAT and inguinal WAT (Sharp et al., 2012). To induce the formation of brite/beige cells, inguinal WAT (but not the brown) cultures were treated with the PPAR -agonist rosiglitazone. Rosiglitazone induces a brown-like phenotype in otherwise white adipocytes and enhances the brown phenotype in classical brown cells (Petrovic et al., 2008; Petrovic et al., 2010). Microarray analysis was performed on these cell cultures to compare gene expression patterns between brown, white and brite/beige cells. This resulted in the identification of a number of genes potentially distinguishing between classical brown and brite/beige cells. Similar to Wu et al., some of these new marker genes were analyzed in hu-man post-mortem brown adipose biopsies from various anatomical regions. Based on their observations, the authors concluded, “the molecular signature of human BAT resembles beige cells” (Sharp et al., 2012). The genes suggested to serve as markers in the above three studies have been widely used for adipose identification purposes, as discussed in the follow-ing paragraphs and in Paper I. In addition to the studies described above, a later study was aimed at find-ing adipocyte cell surface markers, but used quite a different approach (Ussar et al., 2014). With a combination of in silico searches and gene ex-pression studies, genes were selected that were expressed specifically in white adipocytes or brown adipocytes. This resulted in the identification of a new white adipocyte surface marker (Slc7a10/Asc1, solute carrier family 7 member 10) and two brown and brite/beige adipocyte surface markers (Pat2, Proton assistant amino acid transporter 2 and P2rx5, Purinergic receptor P2X, ligand-gated ion channel 5). Although these markers might not help determine the brown or brite/beige identity of human BAT, they might be-come useful tools to isolate cells for study purposes.
5.2 Determining the identity of human BAT with murine marker genes As described in the previous paragraph, two studies concluded that human BAT has the molecular appearance of murine brite/beige fat (Sharp et al., 2012; Wu et al., 2012). These reports were followed by a number of other studies analyzing marker gene expression in human BAT (Paper II) (Cypess

60
et al., 2013; Lidell et al., 2013). Our own work (Paper II) and that of others did not result in the same conclusions as the earlier work by Wu et al. and Sharp et al. As described in Paper II, using a similar set of marker genes, we found both suggested classical brown and brite/beige markers expressed in human supraclavicular BAT samples. Based on those observations, we con-cluded that classical brown and brite/beige mRNA signatures are overlap-ping in human BAT.
A study performed by Cypess and colleagues included a series of five ad-ipose depots from the neck area (Cypess et al., 2013). These five depots dis-played a wide range of UCP1 expression. Marker gene expression was ana-lyzed in depots with the highest UCP1 levels and subcutaneous neck WAT (low in UCP1) of 13 individuals. Of the analyzed markers, ZIC1 and LHX8 (proposed classical brown markers (Timmons et al., 2007)) were expressed most similarly to UCP1, implying a more classical brown identity in the UCP1-expressing depots. Other suggested brown markers (FBXO31, EBF3 and MPZL2 (Eva1 in mice) (Wu et al., 2012)) did not show enrichment in UCP1-high versus subcutaneous WAT depots. Furthermore, the brite/beige markers TBX1, TMEM26 and TNFRSF9 (Cd137 in mice) (Wu et al., 2012), also did not show enrichment in either of the depots. Thus, in this study, the marker genes provided very little distinctive value. Based on the two brown markers LHX8 and ZIC1, the authors concluded, “some human samples share many similarities with classical, also called constitutive, rodent BAT” (Cypess et al., 2013).
In a study performed by Lidell and colleagues, three fat depots were com-pared (Lidell et al., 2013). Supraclavicular BAT was obtained from healthy adult subjects, periadrenal BAT from patients undergoing surgery for re-moval of benign adrenal tumors (no other parameters reported) and inter-scapular BAT was obtained postmortem from infants, aged 0 – 74 weeks (Lidell et al., 2013). Although these samples came from very different popu-lations, the authors concluded that the imaging performed (MRI) and the gene expression analysis (including some of the proposed marker genes), “provide evidence for the presence of bona fide iBAT in human infants that consists of classical brown adipocytes as opposed to the beige/brite brown adipocytes found in other BAT depots”. From the expression patterns of the marker genes used, no firm conclusion could be drawn about the identity of the supraclavicular and periadrenal depots.
Taken together, the results obtained from marker gene expression analysis in human BAT samples have not resulted in a satisfactory conclusion. This could, for example, be due to issues concerning translation of mouse marker genes to human. However, the validation of the markers and their informa-tive value was limited, especially given what we now know about the heter-ogeneity between and within adipose depots.

61
5.3 Validation of suggested markers After publication of the proposed adipose marker genes (Sharp et al., 2012; Timmons et al., 2007; Wu et al., 2012) - and to a lesser extent (Ussar et al., 2014) – many laboratories included expression analysis of these mark-ers in their studies on brown, brite/beige and white adipose tissues and cells in both mouse and human (see previous paragraph). Because of their wide usage, and thus the acceptance of the informative value these markers are thought to provide, it is of great importance to thoroughly validate their in-formative value. For some of the earlier proposed markers (Timmons et al., 2007), this was done in a range of mouse adipose depots (Walden et al., 2012). This study already showed that the proposed classical brown marker Zic1 was not expressed in all BAT depots analyzed (Zic1 was absent in me-diastinal BAT). For the markers suggested in later studies (Sharp et al., 2012; Ussar et al., 2014; Wu et al., 2012), no validation had been independently performed. Importantly, the proposed markers had initially been identified in mouse primary and clonal cell cultures, followed by further expression analysis in a limited set of adipose tissues. The assumption was made that the brown, brite/beige and white markers identified in these limited set-ups could be extrapolated to all brown, brite/beige and white cells and tissues in the adi-pose organ, both in mouse and human. Given the observations of heteroge-neity in developmental cellular lineages (Chapter 4), extrapolation of the results obtained with marker genes should have been done with much more care. To validate the informative value of most suggested markers, we analyzed their expression in a much more extensive set of adipose tissues and cell culture conditions (Paper I). The data in Paper I indicated that the majority of suggested marker genes did not fulfill the basic criteria of distinguishing between interscapular BAT, inguinal WAT and epididymal WAT. The information obtained from mark-ers that did show selective expression in only one of these three tissues (Zic1, Cd137, Epsti1, Tbx1, Tmem26 and Tcf21) became much more difficult to interpret when analyzed in a set of 14 adipose depots (Figure 12 in Paper I). Importantly, these results clearly demonstrated large variation in molecu-lar signatures among adipose depots. Strikingly, the proposed brite/beige markers Cd137, Tbx1 and Tmem26 (Wu et al., 2012) displayed very similar expression patterns. Epsti1, which was originally proposed to be a classical brown marker (Sharp et al., 2012), followed the criteria of a brite/beige marker in our study and had an expression pattern similar to Cd137, Tbx1 and Tmem26. However, these four genes were all expressed in mesenteric WAT, the depot with the lowest Ucp1 levels (i.e. the “whitest” depot) (see also Table 1). Depending on one’s interpretation, this observation either invalidates these four markers as brite/beige-specific, or implies the presence

62
of potentially brite/beige cells in mesenteric WAT (but they are not suffi-ciently stimulated to become functionally brite/beige). Zic1, the most commonly used classical brown marker, was expressed in 3 out of 5 depots that we considered classical brown based on Ucp1 expres-sion levels at thermoneutrality (mediastinal BAT (Walden et al., 2012) and perirenal BAT lacked Zic1 expression (Figure 9A in Paper I)) (Table 1). Furthermore, Zic1 was highly expressed in interscapular WAT, a depot that does not express Ucp1 at thermoneutrality (i.e. is not considered classical brown). These observations make it difficult to keep considering Zic1 as a bona fide classical brown marker. Furthermore, positional analysis of Zic1 expression indicated that its expression is restricted entirely to the anterior-most fat depots in mice (Figure 14C in Paper I). The proposed white marker Tcf21 was indeed expressed in the 2 depots with the lowest Ucp1 expression (eWAT and mWAT), but also appeared in perirenal BAT and perirenal WAT. This makes it difficult to maintain Tcf21 as a clearly distinctive white marker (see also Table 1). To obtain further information about the behavior of some selected marker genes, we also analyzed their expression in BAT and inguinal WAT samples of C57BL/6 mice kept under normal housing conditions and mice fed a high-fat diet at thermoneutrality for 7-8 months (i.e. under “humanized” condi-tions) (Figure 4 in Paper III). The results obtained confirmed earlier BAT-versus-inguinal WAT differences at room temperature. However, important-ly, the markers became much less informative about tissue identity after humanization. Expression patterns of both classical brown and brite/beige markers were much more similar in humanized mice than in mice kept under standard housing conditions. This could explain why the markers had been identified as informative in the first place and why they are not consistently informative in human samples (paragraph 5.2). Taken together, our data show that the heterogeneous expression patterns of currently used marker genes make the information they provide regarding adipose identity very difficult (if not impossible) to interpret.
5.4 Implications of our marker validation for other studies
5.4.1 What do our results mean for earlier observations? Between the initial publications of the suggested markers and our publica-tion regarding their validity, several studies analyzed these marker genes. The main reason for marker analysis has been to determine the effects of pharmacological treatments and genetic manipulations on the functional nature of the derived adipocytes. For marker genes to be useful in this type of analysis, the assumption has to be that marker expression changes along

63
Tab
le 1
. Obs
erva
tions
and
inte
rpre
tatio
ns o
f mar
ker
gene
exp
ress
ion
M
arke
r ge
ne
Prop
osed
se
lect
ivity
O
bser
ved
in
Obt
aine
d in
form
atio
n O
ther
obs
erva
tions
E
bf3
Bro
wn
All
Mix
ed
E
psti1
B
row
n in
gWA
T, tr
iWA
T, m
WA
T B
rite/
beig
e*
E
va1
Bro
wn
All
Mix
ed
F
bxo3
1 B
row
n A
ll M
ixed
Lhx8
B
row
n iB
AT,
cB
AT,
isW
AT,
asW
AT
Mix
ed
Zi
c1
Bro
wn
iBA
T, a
BA
T, c
BA
T, is
WA
T M
ixed
Ex
pres
sed
in th
e an
terio
r-m
ost d
epot
s C
ar4
Brit
e/be
ige
All
Mix
ed
C
d137
B
rite/
beig
e in
gWA
T, tr
iWA
T, m
WA
T B
rite/
beig
e*
C
ited1
B
rite/
beig
e A
ll M
ixed
Fgf
21
Brit
e/be
ige
All
(exc
ept c
BA
T, c
WA
T an
d eW
AT)
M
ixed
Hox
c9
Brit
e/be
ige
prB
AT,
ingW
AT,
prW
AT,
rpW
AT,
eW
AT
Mix
ed
Ant
ero-
post
erio
r ex
pres
sion
gra
dien
t Tb
x1
Brit
e/be
ige
mB
AT,
ingW
AT,
triW
AT,
mW
AT
Mix
ed
Tm
em26
B
rite/
beig
e in
gWA
T, tr
iWA
T, m
WA
T an
d lo
wer
leve
ls in
all
othe
r tis
sues
exc
ept
iBA
T, a
BA
T an
d cB
AT
Mix
ed
Hox
c8
Whi
te
mB
AT,
prB
AT,
ingW
AT,
isW
AT,
prW
AT,
rpW
AT,
asW
AT,
eW
AT,
m
WA
T M
ixed
Po
sitio
nal e
xpre
ssio
n pa
ttern
Tc
f21
Whi
te
prB
AT,
prW
AT,
eW
AT,
mW
AT
Mix
ed
Posi
tiona
l exp
ress
ion
patte
rn
Tabl
e 1.
Ove
rvie
w o
f mar
ker g
enes
that
wer
e in
vest
igat
ed in
Pap
er I.
Pro
pose
d se
lect
ivity
is b
ased
on
the
orig
inal
stud
ies i
n w
hich
the
indi
cate
d ge
nes w
ere
iden
tifie
d. T
he “
obse
rved
in”,
“ob
tain
ed in
form
atio
n” a
nd “
othe
r obs
erva
tions
” co
lum
ns a
re b
ased
on
data
in P
aper
I.
* Th
e br
ite/b
eige
info
rmat
ion
that
Eps
ti1 a
nd C
d137
may
pro
vide
is c
ompl
icat
ed b
y th
eir e
xpre
ssio
n in
mes
ente
ric W
AT
(mW
AT)
, whi
ch
had
the
low
est l
evel
s of U
cp1
expr
essi
on a
nd c
ould
thus
be
cons
ider
ed a
whi
te a
dipo
se d
epot
(Pap
er I)
.

64
with activity. However, the original publications rather indicated that the markers are expressed either in brown, brite/beige or white cells, regardless of their functional status. This is illustrated by the fact that the markers that are mostly used have been identified in precursor cells (e.g. Zic1 and Lhx8 (Timmons et al., 2007)), or have even been used to select brite/beige cells at the precursor stage (e.g. Cd137 and Tmem26 (Wu et al., 2012)). Further-more, in our observations, expression of brite/beige markers during cold acclimation generally remained unchanged or was even downregulated (Pa-per I). Thus, the presence or absence of (changes in) expression of marker genes does not necessarily indicate a functional change of the adipocytes. The cur-rent methods to characterize brown, brite/beige and white adipose are insuf-ficient to be used to provide any information about the identity of human BAT. The best proof available so far is microscopic evidence and the detec-tion of UCP1. Although these methods indicate the presence of potentially thermogenic adipocytes, they do not distinguish between classical brown and brite/beige.
5.4.2 How have our observations affected later studies? At the moment of writing, Paper I has been cited 23 times (Thomson Reuters Web of Knowledge). Principally, the contexts in which the paper is cited can be divided into two categories. First, the description and illustration of the mouse adipose depots is referred to. Second, the expression and in-formative value of marker genes are interpreted from our data. One study in particular has built further on the data presented in our paper (Nakhuda et al., 2016). The authors performed microarray analysis of prima-ry cultures of inguinal stromal-vascular fractions derived from C57BL/6 and 129Sv mice in the presence or absence of rosiglitazone. A direct comparison of rosiglitazone-induced changes in the expression levels of several marker genes with the data presented in Paper I indicated around 60% consistency between the two studies (Nakhuda et al., 2016). Further analysis also indi-cated that observed gene expression levels could differ depending on which exons were analyzed (Nakhuda et al., 2016). Given the use of different mouse strains (C57BL/6 and 129Sv compared to NMRI in our study) and different analytical techniques (microarray versus quantitative PCR), the observed 60% consistency is rather high. However, the observed inconsist-encies further contribute to the notion that the validity of the proposed mark-er genes should have been scrutinized much more thoroughly before being used to draw conclusions about the nature of human (brown) adipose tissues. To conclude, it is clear that the field would benefit from having ways to specifically mark a particular type of adipocyte to distinguish it from other adipocyte types. Clearly UCP1 distinguishes brite/beige cells from surround-ing white adipocytes, but would not distinguish brite/beige from classical

65
brown adipocytes. Even more beneficial would be a cell surface marker to physically separate cells from each other (Ussar et al., 2014). The heterogeneity described throughout this thesis implies that it will be very difficult to define one type of brown adipocyte or one type of white adipocyte. Finding suitable markers to identify those (pre-)adipocytes specif-ically will then be even more difficult. Understanding the nature of human BAT (classical brown or brite/beige) will take more than just the expression of a handful of genes. Morphological and functional analyses are likely to be more informative.

66
Summary and conclusions
The adipose organ consists of a large number of adipose tissues that all have somewhat different colors and shapes. From this first macroscopic observa-tion, a division between brown and white adipose tissues can be readily made (at least in rodents). At the microscopic level, this classification does not change much, but under varying physiological conditions, one can ap-preciate the dynamics in the cellular morphology of adipocytes. Brown-like (i.e. multilocular) adipocytes can be observed within an otherwise seemingly white adipose tissue and white-like (i.e. unilocular) adipocytes can appear in brown adipose tissues. This dynamic nature makes it difficult to properly characterize and classify the different adipocytes. To further characterize and classify adipocytes and adipose depots, other aspects of adipose biology can be used. The aim of this thesis was to look, in addition to visual appearance, at functions, cell and tissue dynamics, devel-opment and molecular characteristics to further add to the classification. Based on function, it proves difficult to further distinguish between brown and white (and brite/beige) adipocytes. Among brown adipose depots, no differences in functionality have been described. Brite/beige adipocytes ap-pear thermogenically qualitatively similar to classical brown adipocytes, if sufficiently stimulated (e.g. in the cold). With regard to substrate uptake (glucose, fatty acids), brown adipose tissue has a much higher capacity for uptake of these substrates than subcutaneous WAT due to brown and brite/beige adipocyte density. Whether brite/beige adipocytes are qualitative-ly (and quantitatively) similar to classical brown adipocytes remains to be shown. Between white adipose tissues, a general distinction is made between subcutaneous and visceral WAT. This is in part due to adipokine secretion profiles and correlation studies looking at fat mass and overall metabolic health. Also, cellular responses to molecules such as catecholamines tend to differ between subcutaneous and visceral adipocytes. However, more de-tailed studies should clarify whether this subcutaneous-versus-visceral dis-tinction could be further complicated by functional differences among vari-ous subcutaneous adipocytes or various visceral adipocytes. In addition to the peripheral white adipocytes, morphologically similar adipocytes appear in bone marrow, dermis and the mammary gland. From the earliest studies, these adipocytes have been suggested to fulfill maintenance functions in their local environments.

67
Mouse studies are increasingly providing new insights into how adipose depots dynamically behave under various physiological situations (e.g. upon cold exposure or in response to high-fat diet feeding). This has led to the realization that the proliferative (hyperplastic) response upon cold exposure differs between various (white) adipose depots. However, these proliferative responses do not seem to relate to the visceral or subcutaneous location (par-agraph 3.2.1). The observation that brite/beige adipocytes can arise from either mature white-like adipocytes or through maturation of (proliferating) precursors, suggests that among brite/beige adipocytes different classes might exist, even within one depot. However, technical issues that are still being further developed may affect some of these observations. The observed dynamics of classical BAT imply that brown and brite/beige depots behave differently in the same thermal zones (paragraph 3.1 and 3.2). Hyperplastic responses in white adipose depots are also observed in re-sponse to high-fat diet feeding (paragraph 3.4.1). In male mice, this hyper-plastic response was observed in epididymal WAT, but not in inguinal WAT. In female mice, however, hyperplasia was observed in both WAT depots. These observations imply visceral-versus-subcutaneous differences in male mice, but not in female mice. Analysis of additional WAT depots would help to further define depot and gender differences. Much information about adipocyte identity has come from lineage tracing studies to find the embryonic origins of adipocytes. Adipose organ-wide analysis in mice has led to the view that much heterogeneity exists with re-gard to developmental origins and that this, at first sight, does not necessari-ly relate to adipocyte function. It will be a matter of time before more pieces of this puzzle are put together. Further complications in the classification of adipose depot identities have come from the use of marker genes. The experiments that put forward the suggested marker genes used rather limited experimental conditions (i.e. use of only few different adipose tissues and cells). Extrapolation of the suggest-ed markers to human adipose tissue has therefore led to contradictory results. Our study of the suggested markers in much more extensive mouse experi-ments indicated that interpretation of the informative value of the marker genes is much more complex than initially assumed. Overall, we currently generally distinguish brown, white and brite/beige adipocytes. Among white adipose tissues, increasing distinction is made between visceral WAT and subcutaneous WAT. Dermal, marrow and mam-mary adipocytes are being added to the field as additional adipocytes with “unusual” anatomical locations. With the current knowledge, this generaliza-tion starts to seem too simplistic, but as further classification is yet incom-plete, this simplistic distinction is all we can do for now. To add more detail to the classification of adipocytes, it is of great importance that studies are as

68
detailed as possible about which adipocytes are studied. Then, with time, our understanding of the complexity of the adipose organ can grow.

69
Sammanfattning på Svenska
Den ökande förekomsten av fetma och dess följdsjukdomar som typ-2 diabe-tes, hjärt-kärlsjukdomar och cancer, kräver en grundlig utredning av de bakomliggande mekanismerna. En av de primära vävnaderna som undersöks i detta sammanhang är fettvävnaden. Det blir alltmer klart att fettvävnaden är ett mycket dynamiskt och heterogent organ. Denna avhandling ger en överblick över olika aspekter av fettcellernas biologi som illustrerar dess heterogena natur och beskriver mitt vetenskapliga bidrag till detta område.
Fettceller delas vanligtvis upp i värmealstrande, så kallade termogena, en-ergi-förbrukande bruna fettceller och energi-lagrande vita fettceller. De bru-na och vita fettcellerna är anatomiskt lokaliserade i skilda fettdepåer. Utöver dessa finns så kallade ”brite” (eller ”beige”) fettceller som har termogena funktioner men som är spritt lokaliserade bland vita fettceller.
Relaterat till deras funktionella variation uppvisar fettcellerna ett brett morfologiskt spektrum. Ännu en egenskap som vittnar om fettcellernas och fettvävnadernas mångfasetterade natur är deras varierande cellulära svar på yttre fysiologiska stimuli, som förändrad diet eller ändringar i omgivningens temperatur. Fettcellernas mångsidighet speglas vidare i deras skilda embryo-nala ursprung, vilket delvis kan förklara varför de har så olika egenskaper.
I linje med komplexiteten bakom deras skilda embryonala ursprung, har uttrycket av molekylära markörer - som tidigare föreslagits särskilja mellan bruna, britea/beigea och vita fettceller- bidragit till att visa på en betydligt mer komplex bild av fettvävnadens uppbyggnad än vad man tidigare trott.
Mitt arbete inom området har bidragit till ökad insikt i fettcellernas heter-ogenitet. Mer specifikt har mina fynd om molekylära markörers ge-nuttrycksmönster betonat komplexiteten av dessa i fettceller, men även svårigheterna att översätta resultat erhållna från studier på möss till männis-kan. En annan del av mitt arbete har handlat om att bidra till ökad förståelse av likheter och skillnader gällande fettvävnadernas funktion och dynamik. I cellodlingsstudier har jag upptäckt nya egenskaper hos förstadier till fettcel-ler från olika fettvävnader som ytterligare visar på deras heterogenitet.
I dess helhet ger denna avhandling en överblick av fettvävnadens vari-erande egenskaper vilket illustrerar dess mångfasetterade natur. Erkännandet av denna heterogenitet är av stor betydelse för den fortsatta forskningen av fettvävnaden som organ och som mål för framtida terapeutiska intervention-er så att vi effektivt ska kunna bekämpa fetmarelaterade sjukdomar.

70
Nederlandse samenvatting
De toename van obesitas en gerelateerde aandoeningen zoals type 2 diabetes, hart- en vaatziekten en kanker vraagt om diepgaand onderzoek naar de on-derliggende oorzaken. Een van de voornaamste weefsels die worden bestu-deerd in dit verband is vetweefsel. Het wordt meer en meer duidelijk dat vetweefsel een dynamisch en veelzijdig weefsel is. Deze thesis geeft een overzicht van de veelzijdigheid van verscheidene aspecten van vetweefsel en het beschrijft mijn eigen bijdragen aan dit onderzoeksveld.
Vetcellen worden veelal opgesplitst in warmte producerende, energie-verbrandende bruine en energie-opslaande witte subgroepen. Bruine en witte vetcellen zijn gelokaliseerd in gescheiden weefsels. Daarnaast onderscheiden we ook “brite” (of “beige”) vetcellen die functioneel vergelijkbaar zijn met bruine vetcellen, maar zich tussen witte vetcellen begeven.
Gerelateerd aan functionele variatie, hebben vetcellen veel verschillende voorkomens. Een andere eigenschap die de veelzijdigheid van vetcellen en -weefsels weergeeft is de variatie in cellulaire reacties op fysiologische sti-muli zoals veranderingen in het dieet of in the omgevingstemperatuur. Daarnaast zijn de embryonale oorsprongen van verschillende vetcellen erg uiteenlopend. Dit kan mede verklaren waarom vetcellen zulke uiteenlopende eigenschappen vertonen.
In lijn met de complexiteit van de embryonale oorsprongen, hebben mo-leculaire markers (die in eerste instantie waren bedoeld om bruine, brite/beige en witte vetcellen van elkaar te kunnen onderscheiden) bijgedra-gen aan het begrip dat de opbouw van het vet-orgaan veel complexer is dan lang werd gedacht.
Mijn eigen werk heeft bijgedragen aan de toename van het begrip van de veelzijdigheid van vet subtypen. Met name mijn bevindingen aangaande genexpressie patronen hebben geleid tot toegenomen waardering van de complexiteit van deze expressie patronen en problemen om deze te vertalen van muizen naar mensen. Een ander deel van mijn werk betreft bijdragen aan de toename van ons begrip van de verschillen en overeenkomsten aangaande functies en dynamiek van warmte-producerende vetweefsels. Met celkweek studies heb ik nieuwe eigenschappen blootgelegd van pre-vetcellen van ver-schillende vetweefsels wat een extra kijk heeft gegeven in de verscheiden-heid van vetweefsels.
In zijn geheel geeft deze thesis een overzicht van de eigenschappen van vetweefsel die de veelzijdigheid van dit weefsel illustreren. Erkenning van

71
deze veelzijdigheid is van belang om dit orgaan op de juiste manier als ge-heel te bestuderen en om zo te kunnen begrijpen hoe vetweefsel als thera-peutisch doelwit kan worden gebruikt om effectief vetweefsel-gerelateerde ziekten te kunnen behandelen.

72
Acknowledgements
Here, I would like to take the opportunity to thank those who have played a part, big or small, during my time as a graduate student at Stockholm Uni-versity. Barbara and Jan: Thank you for letting me become part of the (brown) adipose community and for teaching me to be critical about any piece of data and to question everything. Because of your mentoring during the past 5-plus years, I have never questioned pursuing a career in science. The oppor-tunities you have given me to travel to conferences, to develop my own pro-jects and to make contacts with other labs have really helped to prepare tak-ing the next step in my career as a scientist. I truly hope I can pass on what I have learned from you. Natasa: Having the opportunity to work with you has added so much value to my PhD. Your enthusiasm, your drive to discover, your kindness and your willingness to help have made it so easy for me to complete this PhD. It has been a great experience to share ideas with you and to be part of your pro-jects. I hope you get to keep doing what you do best for as long as you like. Tore and Anders: Thank you both for your positive attitudes, the kind words of motivation and the jokes at always the right time. The Bengtsson Group: You guys are an amazing bunch of enthusiastic people. It has been a pleasure being around each and every one of you during the past years. Good luck to you all! The Jacobsson Group: Science-enthusiasts and very kind people. Thank you for the nice times, the helpful discussions and the memories! The Cannon/Nedergaard Group: Irina, it has been great to work in the same place as you. Your critical approach to experiments and data have helped me to also develop a critical view. Good luck continuing to further understand the wonders of mitochondria. Anastasia, your positive approach to everything that comes your way is very inspiring. Thank you for all the discussions and laughs! Yanling, thank you for your continuous efforts in your experimental work. It has had a motivating influence on me! Gustavo,

73
thanks for all the discussion about our work and your enthusiasm for science. Ida, thanks for all the laughs and the early morning coffees. I appreciated your efforts to keep maintain some level of order in the lab, although that’s never been easy. Ineke, dankje voor de nodige porties Nederlandse nuchter-heid en stroopwafels. Veel succes met de rest van je PhD en alles wat daarna komt! Gabriella, it has been very nice to share the office with you. Many thanks for all the discussion we had and for going through all the problems of thesis-practicalities before me so I did not have to. Nathalie, thanks for your positive and motivating attitude. Seeing you take on you project is in-spiring; I hope it gives you what you’re looking for. Erik, from c-course student to PhD-colleague. Thanks for all the discussions, your natural will to help out where needed and for taking over the plastic room and the frog lab! Petr, seeing you take on these huge projects has been very inspiring. I hope you get rewarded for your hard work. Good luck at your next position back home! Bosse, thank you for being so kind and helpful and sharing your pharma-experience with us! Alex, it is a delight to have you around in the lab. Your enthusiasm and drive for science are inspiring! Marie, Tania, Xisco, Frank, Alexia, Hamza, Martina, Laura, Kim, An-tonia, Ollie, Necati, Hoi, Ankita: It has been great to get to know all of you and it is a nice idea to know that there are so many people out there that share the experience of the F3-corridor. Animal house staff: Doing animal experiments was especially fun because you get to walk into the friendly environment that you have all created, even in the new location. Thank you for all the help and good luck in the coming chaotic but exciting times around the big move! Olga: Thank you for being the bioinformagician that everyone needs and helping me make sense of the RNA-seq data. Our discussion have been very motivating to me to learn the tools for big data analysis! Ola: Many thanks for your help on the marker gene project and advice on the RNA-seq experiment. Camilla: It was a great pleasure spending 2 weeks in your lab in Copenha-gen. Thank you for welcoming me there and for the continued scientific discussions! Charlie, Joshua, René My mentees. It has been an absolute pleasure to work with you guys. Seeing how all three of you have come out of this lab working on your futures is vey rewarding. I very much value that we still have contact and I look forward to seeing how you all build great futures for yourselves! Tobias, Touba, Alice, Josephine, Jelena, Marzia, Natalie: Thank you all for your efforts during the C-course project work. I hope you all enjoyed it as much as I did!

74
Decap: Hoe donker het ook wordt hier in Zweden, met Kerst is er altijd het licht van het Decap-diner! De studie-uren in de MB hebben me uiteindelijk naar deze thesis geleid. Die contactmomenten met jullie, in het echie of in de WhatsApp doen me altijd veel goed en ik hoop dat die niet verloren zullen gaan. Van deze generatie Decappers ben ik wellicht de laatste die promove-ert, maar de volgende generatie zit ons al op de hielen. Papoutsi/Svanberg familjen: Tack för att jag får vara del av familjen... Det är alltid jätte roligt att vara med er alla. Jag hoppas att vi kan fortsätta göra detta, även om jag kommer vara långt borta en längre tid. Tack och
! Pap, mam en David: Ondanks de grote afstand weet ik dat jullie altijd dicht achter me staan. Ik ben ontzettend dankbaar dat ik altijd de omgeving heb gekregen om mijn dromen te volgen, ook al ben ik daarom nu ver van huis. Ik hoop dat dit eindresultaat een voldoende goedmaker is voor m’n af-wezigheid. Anna: Thank you for sharing this adventure with me and helping me through the stressful times, especially towards the end. I am very happy to be able to share my ideas with you, even though I never write it down for you and all of it is so far from any clinical relevance. I’ll try to make my next projects a bit more interesting for you ;)

75
References
Adler, E.S., Hollis, J.H., Clarke, I.J., Grattan, D.R., and Oldfield, B.J. (2012). Neurochemical characterization and sexual dimorphism of projections from the brain to abdominal and subcutaneous white adipose tissue in the rat. The Journal of neuroscience : the official journal of the Society for Neuroscience 32, 15913-15921. Aherne, W., and Hull, D. (1966). Brown adipose tissue and heat production in the newborn infant. The Journal of pathology and bacteriology 91, 223-234. Almind, K., Manieri, M., Sivitz, W.I., Cinti, S., and Kahn, C.R. (2007). Ectopic brown adipose tissue in muscle provides a mechanism for differences in risk of metabolic syndrome in mice. Proceedings of the National Academy of Sciences of the United States of America 104, 2366-2371. Ambati, S., Li, Q., Rayalam, S., Hartzell, D.L., Della-Fera, M.A., Hamrick, M.W., and Baile, C.A. (2010). Central leptin versus ghrelin: effects on bone marrow adiposity and gene expression. Endocrine 37, 115-123. Arner, P. (1995). Differences in lipolysis between human subcutaneous and omental adipose tissues. Annals of medicine 27, 435-438. Arner, P. (2005). Human fat cell lipolysis: biochemistry, regulation and clinical role. Best practice & research. Clinical endocrinology & metabolism 19, 471-482. Arner, P., Andersson, D.P., Thorne, A., Wiren, M., Hoffstedt, J., Naslund, E., Thorell, A., and Ryden, M. (2013). Variations in the size of the major omentum are primarily determined by fat cell number. The Journal of clinical endocrinology and metabolism 98, E897-901. Atit, R., Sgaier, S.K., Mohamed, O.A., Taketo, M.M., Dufort, D., Joyner, A.L., Niswander, L., and Conlon, R.A. (2006). Beta-catenin activation is necessary and sufficient to specify the dorsal dermal fate in the mouse. Developmental biology 296, 164-176.

76
Bachman, E.S., Dhillon, H., Zhang, C.Y., Cinti, S., Bianco, A.C., Kobilka, B.K., and Lowell, B.B. (2002). betaAR signaling required for diet-induced thermogenesis and obesity resistance. Science (New York, N.Y.) 297, 843-845. Barbatelli, G., Murano, I., Madsen, L., Hao, Q., Jimenez, M., Kristiansen, K., Giacobino, J.P., De Matteis, R., and Cinti, S. (2010). The emergence of cold-induced brown adipocytes in mouse white fat depots is determined predominantly by white to brown adipocyte transdifferentiation. American journal of physiology. Endocrinology and metabolism 298, E1244-1253. Bartelt, A., Bruns, O.T., Reimer, R., Hohenberg, H., Ittrich, H., Peldschus, K., Kaul, M.G., Tromsdorf, U.I., Weller, H., Waurisch, C., et al. (2011). Brown adipose tissue activity controls triglyceride clearance. Nature medicine 17, 200-205. Bengtsson, T., Cannon, B., and Nedergaard, J. (2000). Differential adrenergic regulation of the gene expression of the beta-adrenoceptor subtypes beta1, beta2 and beta3 in brown adipocytes. The Biochemical journal 347 Pt 3, 643-651. Berry, D.C., Jiang, Y., and Graff, J.M. (2016). Mouse strains to study cold-inducible beige progenitors and beige adipocyte formation and function. Nature communications 7, 10184. Berry, R., Rodeheffer, M.S., Rosen, C.J., and Horowitz, M.C. (2015). Adipose Tissue Residing Progenitors (Adipocyte Lineage Progenitors and Adipose Derived Stem Cells (ADSC). Current molecular biology reports 1, 101-109. Bertholet, A.M., and Kirichok, Y. (2017). UCP1: A transporter for H+ and fatty acid anions. Biochimie 134, 28-34. Bronnikov, G., Bengtsson, T., Kramarova, L., Golozoubova, V., Cannon, B., and Nedergaard, J. (1999). beta1 to beta3 switch in control of cyclic adenosine monophosphate during brown adipocyte development explains distinct beta-adrenoceptor subtype mediation of proliferation and differentiation. Endocrinology 140, 4185-4197. Bukowiecki, L., Collet, A.J., Follea, N., Guay, G., and Jahjah, L. (1982). Brown adipose tissue hyperplasia: a fundamental mechanism of adaptation to cold and hyperphagia. The American journal of physiology 242, E353-359. Cameron, I.L., and Smith, R.E. (1964). Cytological responses of brown fat tissue in cold-exposed rats. The Journal of cell biology 23, 89-100.

77
Cancello, R., Tordjman, J., Poitou, C., Guilhem, G., Bouillot, J.L., Hugol, D., Coussieu, C., Basdevant, A., Bar Hen, A., Bedossa, P., et al. (2006). Increased infiltration of macrophages in omental adipose tissue is associated with marked hepatic lesions in morbid human obesity. Diabetes 55, 1554-1561. Cannon, B., Hedin, A., and Nedergaard, J. (1982). Exclusive occurrence of thermogenin antigen in brown adipose tissue. FEBS letters 150, 129-132. Cannon, B., and Nedergaard, J. (2004). Brown adipose tissue: function and physiological significance. Physiological reviews 84, 277-359. Cannon, B., and Vogel, G. (1977). The mitochondrial ATPase of brown adipose tissue. Purification and comparison with the mitochondrial ATPase from beef heart. FEBS letters 76, 284-289. Cawthorn, W.P., Scheller, E.L., Learman, B.S., Parlee, S.D., Simon, B.R., Mori, H., Ning, X., Bree, A.J., Schell, B., Broome, D.T., et al. (2014). Bone marrow adipose tissue is an endocrine organ that contributes to increased circulating adiponectin during caloric restriction. Cell metabolism 20, 368-375. Cawthorn, W.P., Scheller, E.L., Parlee, S.D., Pham, H.A., Learman, B.S., Redshaw, C.M., Sulston, R.J., Burr, A.A., Das, A.K., Simon, B.R., et al. (2016). Expansion of Bone Marrow Adipose Tissue During Caloric Restriction Is Associated With Increased Circulating Glucocorticoids and Not With Hypoleptinemia. Endocrinology 157, 508-521. Chau, Y.Y., Bandiera, R., Serrels, A., Martinez-Estrada, O.M., Qing, W., Lee, M., Slight, J., Thornburn, A., Berry, R., McHaffie, S., et al. (2014). Visceral and subcutaneous fat have different origins and evidence supports a mesothelial source. Nature cell biology 16, 367-375. Chaudhry, A., and Granneman, J.G. (1999). Differential regulation of functional responses by beta-adrenergic receptor subtypes in brown adipocytes. The American journal of physiology 277, R147-153. Chen, J., Shi, Y., Regan, J., Karuppaiah, K., Ornitz, D.M., and Long, F. (2014). Osx-Cre targets multiple cell types besides osteoblast lineage in postnatal mice. PloS one 9, e85161. Chernogubova, E., Hutchinson, D.S., Nedergaard, J., and Bengtsson, T. (2005). Alpha1- and beta1-adrenoceptor signaling fully compensates for beta3-adrenoceptor deficiency in brown adipocyte norepinephrine-stimulated glucose uptake. Endocrinology 146, 2271-2284. Cinti, S. (1999). The Adipose Rogan. (Milan, Italy: Kurtis).

78
Contreras-Shannon, V., Ochoa, O., Reyes-Reyna, S.M., Sun, D., Michalek, J.E., Kuziel, W.A., McManus, L.M., and Shireman, P.K. (2007). Fat accumulation with altered inflammation and regeneration in skeletal muscle of CCR2-/- mice following ischemic injury. American journal of physiology. Cell physiology 292, C953-967. Cousin, B., Bascands-Viguerie, N., Kassis, N., Nibbelink, M., Ambid, L., Casteilla, L., and Penicaud, L. (1996). Cellular changes during cold acclimatation in adipose tissues. Journal of cellular physiology 167, 285-289. Crossno, J.T., Jr., Majka, S.M., Grazia, T., Gill, R.G., and Klemm, D.J. (2006). Rosiglitazone promotes development of a novel adipocyte population from bone marrow-derived circulating progenitor cells. The Journal of clinical investigation 116, 3220-3228. Cypess, A.M., Lehman, S., Williams, G., Tal, I., Rodman, D., Goldfine, A.B., Kuo, F.C., Palmer, E.L., Tseng, Y.H., Doria, A., et al. (2009). Identification and importance of brown adipose tissue in adult humans. The New England journal of medicine 360, 1509-1517. Cypess, A.M., White, A.P., Vernochet, C., Schulz, T.J., Xue, R., Sass, C.A., Huang, T.L., Roberts-Toler, C., Weiner, L.S., Sze, C., et al. (2013). Anatomical localization, gene expression profiling and functional characterization of adult human neck brown fat. Nature medicine 19, 635-639. Devlin, M.J., Cloutier, A.M., Thomas, N.A., Panus, D.A., Lotinun, S., Pinz, I., Baron, R., Rosen, C.J., and Bouxsein, M.L. (2010). Caloric restriction leads to high marrow adiposity and low bone mass in growing mice. Journal of bone and mineral research : the official journal of the American Society for Bone and Mineral Research 25, 2078-2088. Doucette, C.R., Horowitz, M.C., Berry, R., MacDougald, O.A., Anunciado-Koza, R., Koza, R.A., and Rosen, C.J. (2015). A High Fat Diet Increases Bone Marrow Adipose Tissue (MAT) But Does Not Alter Trabecular or Cortical Bone Mass in C57BL/6J Mice. Journal of cellular physiology 230, 2032-2037. Driskell, R.R., Jahoda, C.A., Chuong, C.M., Watt, F.M., and Horsley, V. (2014). Defining dermal adipose tissue. Experimental dermatology 23, 629-631. Emont, M.P., Yu, H., Jun, H., Hong, X., Maganti, N., Stegemann, J.P., and Wu, J. (2015). Using a 3D Culture System to Differentiate Visceral Adipocytes In Vitro. Endocrinology 156, 4761-4768.

79
Fang, L., Guo, F., Zhou, L., Stahl, R., and Grams, J. (2015). The cell size and distribution of adipocytes from subcutaneous and visceral fat is associated with type 2 diabetes mellitus in humans. Adipocyte 4, 273-279. Feldmann, H.M., Golozoubova, V., Cannon, B., and Nedergaard, J. (2009). UCP1 ablation induces obesity and abolishes diet-induced thermogenesis in mice exempt from thermal stress by living at thermoneutrality. Cell metabolism 9, 203-209. Ferrannini, G., Namwanje, M., Fang, B., Damle, M., Li, D., Liu, Q., Lazar, M.A., and Qiang, L. (2016). Genetic backgrounds determine brown remodeling of white fat in rodents. Molecular metabolism 5, 948-958. Festa, E., Fretz, J., Berry, R., Schmidt, B., Rodeheffer, M., Horowitz, M., and Horsley, V. (2011). Adipocyte lineage cells contribute to the skin stem cell niche to drive hair cycling. Cell 146, 761-771. Fisher, F.M., Kleiner, S., Douris, N., Fox, E.C., Mepani, R.J., Verdeguer, F., Wu, J., Kharitonenkov, A., Flier, J.S., Maratos-Flier, E., et al. (2012). FGF21 regulates PGC-1alpha and browning of white adipose tissues in adaptive thermogenesis. Genes & development 26, 271-281. Fisher, F.M., and Maratos-Flier, E. (2016). Understanding the Physiology of FGF21. Annual review of physiology 78, 223-241. Fontana, L., Eagon, J.C., Trujillo, M.E., Scherer, P.E., and Klein, S. (2007). Visceral fat adipokine secretion is associated with systemic inflammation in obese humans. Diabetes 56, 1010-1013. Frank, S., Stallmeyer, B., Kampfer, H., Kolb, N., and Pfeilschifter, J. (2000). Leptin enhances wound re-epithelialization and constitutes a direct function of leptin in skin repair. The Journal of clinical investigation 106, 501-509. Frayn, K.N. (2002). Adipose tissue as a buffer for daily lipid flux. Diabetologia 45, 1201-1210. Giordano, A., Smorlesi, A., Frontini, A., Barbatelli, G., and Cinti, S. (2014). White, brown and pink adipocytes: the extraordinary plasticity of the adipose organ. European journal of endocrinology 170, R159-171. Golozoubova, V., Hohtola, E., Matthias, A., Jacobsson, A., Cannon, B., and Nedergaard, J. (2001). Only UCP1 can mediate adaptive nonshivering thermogenesis in the cold. FASEB journal : official publication of the Federation of American Societies for Experimental Biology 15, 2048-2050.

80
Granneman, J.G., Burnazi, M., Zhu, Z., and Schwamb, L.A. (2003). White adipose tissue contributes to UCP1-independent thermogenesis. American journal of physiology. Endocrinology and metabolism 285, E1230-1236. Greco-Perotto, R., Zaninetti, D., Assimacopoulos-Jeannet, F., Bobbioni, E., and Jeanrenaud, B. (1987). Stimulatory effect of cold adaptation on glucose utilization by brown adipose tissue. Relationship with changes in the glucose transporter system. The Journal of biological chemistry 262, 7732-7736. Grimpo, K., Volker, M.N., Heppe, E.N., Braun, S., Heverhagen, J.T., and Heldmaier, G. (2014). Brown adipose tissue dynamics in wild-type and UCP1-knockout mice: in vivo insights with magnetic resonance. Journal of lipid research 55, 398-409. Guerra, C., Koza, R.A., Yamashita, H., Walsh, K., and Kozak, L.P. (1998). Emergence of brown adipocytes in white fat in mice is under genetic control. Effects on body weight and adiposity. The Journal of clinical investigation 102, 412-420. Hamrick, M.W., Della Fera, M.A., Choi, Y.H., Hartzell, D., Pennington, C., and Baile, C.A. (2007). Injections of leptin into rat ventromedial hypothalamus increase adipocyte apoptosis in peripheral fat and in bone marrow. Cell and tissue research 327, 133-141. Harrington, T.A., Thomas, E.L., Frost, G., Modi, N., and Bell, J.D. (2004). Distribution of adipose tissue in the newborn. Pediatric research 55, 437-441. Heaton, J.M. (1972). The distribution of brown adipose tissue in the human. Journal of anatomy 112, 35-39. Hirning, U., Schmid, P., Schulz, W.A., Kozak, L.P., and Hameister, H. (1989). In developing brown adipose tissue c-myc protooncogene expression is restricted to early differentiation stages. Cell differentiation and development : the official journal of the International Society of Developmental Biologists 27, 243-248. Hondares, E., Iglesias, R., Giralt, A., Gonzalez, F.J., Giralt, M., Mampel, T., and Villarroya, F. (2011). Thermogenic activation induces FGF21 expression and release in brown adipose tissue. The Journal of biological chemistry 286, 12983-12990. Houstek, J., Andersson, U., Tvrdik, P., Nedergaard, J., and Cannon, B. (1995). The expression of subunit c correlates with and thus may limit the biosynthesis of the mitochondrial F0F1-ATPase in brown adipose tissue. The Journal of biological chemistry 270, 7689-7694.

81
Hube, F., Lietz, U., Igel, M., Jensen, P.B., Tornqvist, H., Joost, H.G., and Hauner, H. (1996). Difference in leptin mRNA levels between omental and subcutaneous abdominal adipose tissue from obese humans. Hormone and metabolic research 28, 690-693. Ishibashi, J., and Seale, P. (2010). Medicine. Beige can be slimming. Science (New York, N.Y.) 328, 1113-1114. Jeffery, E., Church, C.D., Holtrup, B., Colman, L., and Rodeheffer, M.S. (2015). Rapid depot-specific activation of adipocyte precursor cells at the onset of obesity. Nature cell biology 17, 376-385. Jeffery, E., Wing, A., Holtrup, B., Sebo, Z., Kaplan, J.L., Saavedra-Pena, R., Church, C.D., Colman, L., Berry, R., and Rodeheffer, M.S. (2016). The Adipose Tissue Microenvironment Regulates Depot-Specific Adipogenesis in Obesity. Cell metabolism 24, 142-150. Jimenez, M., Barbatelli, G., Allevi, R., Cinti, S., Seydoux, J., Giacobino, J.P., Muzzin, P., and Preitner, F. (2003). Beta 3-adrenoceptor knockout in C57BL/6J mice depresses the occurrence of brown adipocytes in white fat. European journal of biochemistry 270, 699-705. Jin, C.E., Xiao, L., Ge, Z.H., Zhan, X.B., and Zhou, H.X. (2015). Role of adiponectin in adipose tissue wound healing. Genetics and molecular research : GMR 14, 8883-8891. Joe, A.W., Yi, L., Natarajan, A., Le Grand, F., So, L., Wang, J., Rudnicki, M.A., and Rossi, F.M. (2010). Muscle injury activates resident fibro/adipogenic progenitors that facilitate myogenesis. Nature cell biology 12, 153-163. Kasza, I., Suh, Y., Wollny, D., Clark, R.J., Roopra, A., Colman, R.J., MacDougald, O.A., Shedd, T.A., Nelson, D.W., Yen, M.I., et al. (2014). Syndecan-1 is required to maintain intradermal fat and prevent cold stress. PLoS genetics 10, e1004514. Kazak, L., Chouchani, E.T., Jedrychowski, M.P., Erickson, B.K., Shinoda, K., Cohen, P., Vetrivelan, R., Lu, G.Z., Laznik-Bogoslavski, D., Hasenfuss, S.C., et al. (2015). A creatine-driven substrate cycle enhances energy expenditure and thermogenesis in beige fat. Cell 163, 643-655. Klingenberg, M. (2017). UCP1 - A sophisticated energy valve. Biochimie 134, 19-27. Koh, Y.J., Kang, S., Lee, H.J., Choi, T.S., Lee, H.S., Cho, C.H., and Koh, G.Y. (2007). Bone marrow-derived circulating progenitor cells fail to

82
transdifferentiate into adipocytes in adult adipose tissues in mice. The Journal of clinical investigation 117, 3684-3695. Kovacova, Z., Tencerova, M., Roussel, B., Wedellova, Z., Rossmeislova, L., Langin, D., Polak, J., and Stich, V. (2012). The impact of obesity on secretion of adiponectin multimeric isoforms differs in visceral and subcutaneous adipose tissue. International journal of obesity (2005) 36, 1360-1365. Kramarova, T.V., Shabalina, I.G., Andersson, U., Westerberg, R., Carlberg, I., Houstek, J., Nedergaard, J., and Cannon, B. (2008). Mitochondrial ATP synthase levels in brown adipose tissue are governed by the c-Fo subunit P1 isoform. FASEB journal : official publication of the Federation of American Societies for Experimental Biology 22, 55-63. Krueger, K.C., Costa, M.J., Du, H., and Feldman, B.J. (2014). Characterization of Cre recombinase activity for in vivo targeting of adipocyte precursor cells. Stem cell reports 3, 1147-1158. Lean, M.E. (1989). Brown adipose tissue in humans. The Proceedings of the Nutrition Society 48, 243-256. Lee, Y.H., Kim, S.N., Kwon, H.J., and Granneman, J.G. (2017). Metabolic heterogeneity of activated beige/brite adipocytes in inguinal adipose tissue. Scientific reports 7, 39794. Lee, Y.H., Kim, S.N., Kwon, H.J., Maddipati, K.R., and Granneman, J.G. (2016). Adipogenic role of alternatively activated macrophages in beta-adrenergic remodeling of white adipose tissue. American journal of physiology. Regulatory, integrative and comparative physiology 310, R55-65. Lee, Y.H., Petkova, A.P., and Granneman, J.G. (2013). Identification of an adipogenic niche for adipose tissue remodeling and restoration. Cell metabolism 18, 355-367. Lee, Y.H., Petkova, A.P., Konkar, A.A., and Granneman, J.G. (2015). Cellular origins of cold-induced brown adipocytes in adult mice. FASEB journal : official publication of the Federation of American Societies for Experimental Biology 29, 286-299. Lee, Y.H., Petkova, A.P., Mottillo, E.P., and Granneman, J.G. (2012). In vivo identification of bipotential adipocyte progenitors recruited by beta3-adrenoceptor activation and high-fat feeding. Cell metabolism 15, 480-491.

83
Lepper, C., and Fan, C.M. (2010). Inducible lineage tracing of Pax7-descendant cells reveals embryonic origin of adult satellite cells. Genesis (New York, N.Y. : 2000) 48, 424-436. Li, Y., Bolze, F., Fromme, T., and Klingenspor, M. (2014). Intrinsic differences in BRITE adipogenesis of primary adipocytes from two different mouse strains. Biochimica et biophysica acta 1841, 1345-1352. Lidell, M.E., Betz, M.J., Dahlqvist Leinhard, O., Heglind, M., Elander, L., Slawik, M., Mussack, T., Nilsson, D., Romu, T., Nuutila, P., et al. (2013). Evidence for two types of brown adipose tissue in humans. Nature medicine 19, 631-634. Liu, W., Shan, T., Yang, X., Liang, S., Zhang, P., Liu, Y., Liu, X., and Kuang, S. (2013a). A heterogeneous lineage origin underlies the phenotypic and molecular differences of white and beige adipocytes. Journal of cell science 126, 3527-3532. Liu, Y., Strecker, S., Wang, L., Kronenberg, M.S., Wang, W., Rowe, D.W., and Maye, P. (2013b). Osterix-cre labeled progenitor cells contribute to the formation and maintenance of the bone marrow stroma. PloS one 8, e71318. Logan, M., Martin, J.F., Nagy, A., Lobe, C., Olson, E.N., and Tabin, C.J. (2002). Expression of Cre Recombinase in the developing mouse limb bud driven by a Prxl enhancer. Genesis (New York, N.Y. : 2000) 33, 77-80. Loncar, D., Afzelius, B.A., and Cannon, B. (1988). Epididymal white adipose tissue after cold stress in rats. II. Mitochondrial changes. Journal of ultrastructure and molecular structure research 101, 199-209. Long, J.Z., Svensson, K.J., Tsai, L., Zeng, X., Roh, H.C., Kong, X., Rao, R.R., Lou, J., Lokurkar, I., Baur, W., et al. (2014). A smooth muscle-like origin for beige adipocytes. Cell metabolism 19, 810-820. Ma, H.T., Ren, R., Chen, Y., Griffith, J.F., Leung, P.C., and Zhang, P. (2014). A simulation study of marrow fat effect on bone biomechanics. Conference proceedings : ... Annual International Conference of the IEEE Engineering in Medicine and Biology Society. IEEE Engineering in Medicine and Biology Society. Annual Conference 2014, 4030-4033. Majka, S.M., Fox, K.E., Psilas, J.C., Helm, K.M., Childs, C.R., Acosta, A.S., Janssen, R.C., Friedman, J.E., Woessner, B.T., Shade, T.R., et al. (2010). De novo generation of white adipocytes from the myeloid lineage via mesenchymal intermediates is age, adipose depot, and gender specific. Proceedings of the National Academy of Sciences of the United States of America 107, 14781-14786.

84
Mattsson, C.L., Csikasz, R.I., Chernogubova, E., Yamamoto, D.L., Hogberg, H.T., Amri, E.Z., Hutchinson, D.S., and Bengtsson, T. (2011). beta(1)-Adrenergic receptors increase UCP1 in human MADS brown adipocytes and rescue cold-acclimated beta(3)-adrenergic receptor-knockout mice via nonshivering thermogenesis. American journal of physiology. Endocrinology and metabolism 301, E1108-1118. Mizoguchi, T., Pinho, S., Ahmed, J., Kunisaki, Y., Hanoun, M., Mendelson, A., Ono, N., Kronenberg, H.M., and Frenette, P.S. (2014). Osterix marks distinct waves of primitive and definitive stromal progenitors during bone marrow development. Developmental cell 29, 340-349. Morroni, M., Giordano, A., Zingaretti, M.C., Boiani, R., De Matteis, R., Kahn, B.B., Nisoli, E., Tonello, C., Pisoschi, C., Luchetti, M.M., et al. (2004). Reversible transdifferentiation of secretory epithelial cells into adipocytes in the mammary gland. Proceedings of the National Academy of Sciences of the United States of America 101, 16801-16806. Motoshima, H., Wu, X., Sinha, M.K., Hardy, V.E., Rosato, E.L., Barbot, D.J., Rosato, F.E., and Goldstein, B.J. (2002). Differential regulation of adiponectin secretion from cultured human omental and subcutaneous adipocytes: effects of insulin and rosiglitazone. The Journal of clinical endocrinology and metabolism 87, 5662-5667. Mottillo, E.P., Balasubramanian, P., Lee, Y.H., Weng, C., Kershaw, E.E., and Granneman, J.G. (2014). Coupling of lipolysis and de novo lipogenesis in brown, beige, and white adipose tissues during chronic beta3-adrenergic receptor activation. Journal of lipid research 55, 2276-2286. Murano, I., Barbatelli, G., Parisani, V., Latini, C., Muzzonigro, G., Castellucci, M., and Cinti, S. (2008). Dead adipocytes, detected as crown-like structures, are prevalent in visceral fat depots of genetically obese mice. Journal of lipid research 49, 1562-1568. Nakashima, K., Zhou, X., Kunkel, G., Zhang, Z., Deng, J.M., Behringer, R.R., and de Crombrugghe, B. (2002). The novel zinc finger-containing transcription factor osterix is required for osteoblast differentiation and bone formation. Cell 108, 17-29. Nakhuda, A., Josse, A.R., Gburcik, V., Crossland, H., Raymond, F., Metairon, S., Good, L., Atherton, P.J., Phillips, S.M., and Timmons, J.A. (2016). Biomarkers of browning of white adipose tissue and their regulation during exercise- and diet-induced weight loss. The American journal of clinical nutrition 104, 557-565.

85
Nedergaard, J., Bengtsson, T., and Cannon, B. (2007). Unexpected evidence for active brown adipose tissue in adult humans. American journal of physiology. Endocrinology and metabolism 293, E444-452. Nedergaard, J., Connolly, E., and Cannon, B. (1986). Brown adipose tissue in the mammalian neonate. In Brown Adipose Tissue. P. Trayhurn, and D.G. Nicholls, eds. (London Edward Arnold), pp. 152-213. Nguyen, N.L., Randall, J., Banfield, B.W., and Bartness, T.J. (2014). Central sympathetic innervations to visceral and subcutaneous white adipose tissue. American journal of physiology. Regulatory, integrative and comparative physiology 306, R375-386. Nicholls, D.G. (2017). The hunt for the molecular mechanism of brown fat thermogenesis. Biochimie 134, 9-18. Obregon, M.J., Jacobsson, A., Kirchgessner, T., Schotz, M.C., Cannon, B., and Nedergaard, J. (1989). Postnatal recruitment of brown adipose tissue is induced by the cold stress experienced by the pups. An analysis of mRNA levels for thermogenin and lipoprotein lipase. The Biochemical journal 259, 341-346. Park, J.H., Kang, H.J., Kang, S.I., Lee, J.E., Hur, J., Ge, K., Mueller, E., Li, H., Lee, B.C., and Lee, S.B. (2013). A multifunctional protein, EWS, is essential for early brown fat lineage determination. Developmental cell 26, 393-404. Petrovic, N., Shabalina, I.G., Timmons, J.A., Cannon, B., and Nedergaard, J. (2008). Thermogenically competent nonadrenergic recruitment in brown preadipocytes by a PPARgamma agonist. American journal of physiology. Endocrinology and metabolism 295, E287-296. Petrovic, N., Walden, T.B., Shabalina, I.G., Timmons, J.A., Cannon, B., and Nedergaard, J. (2010). Chronic peroxisome proliferator-activated receptor gamma (PPARgamma) activation of epididymally derived white adipocyte cultures reveals a population of thermogenically competent, UCP1-containing adipocytes molecularly distinct from classic brown adipocytes. The Journal of biological chemistry 285, 7153-7164. Poissonnet, C.M., Burdi, A.R., and Bookstein, F.L. (1983). Growth and development of human adipose tissue during early gestation. Early human development 8, 1-11. Poissonnet, C.M., Burdi, A.R., and Garn, S.M. (1984). The chronology of adipose tissue appearance and distribution in the human fetus. Early human development 10, 1-11.

86
Pou, K.M., Massaro, J.M., Hoffmann, U., Vasan, R.S., Maurovich-Horvat, P., Larson, M.G., Keaney, J.F., Jr., Meigs, J.B., Lipinska, I., Kathiresan, S., et al. (2007). Visceral and subcutaneous adipose tissue volumes are cross-sectionally related to markers of inflammation and oxidative stress: the Framingham Heart Study. Circulation 116, 1234-1241. Rehnmark, S., and Nedergaard, J. (1989). DNA synthesis in mouse brown adipose tissue is under beta-adrenergic control. Experimental cell research 180, 574-579. Revelli, J.P., Preitner, F., Samec, S., Muniesa, P., Kuehne, F., Boss, O., Vassalli, J.D., Dulloo, A., Seydoux, J., Giacobino, J.P., et al. (1997). Targeted gene disruption reveals a leptin-independent role for the mouse beta3-adrenoceptor in the regulation of body composition. The Journal of clinical investigation 100, 1098-1106. Ricquier, D., Bouillaud, F., Toumelin, P., Mory, G., Bazin, R., Arch, J., and Penicaud, L. (1986). Expression of uncoupling protein mRNA in thermogenic or weakly thermogenic brown adipose tissue. Evidence for a rapid beta-adrenoreceptor-mediated and transcriptionally regulated step during activation of thermogenesis. The Journal of biological chemistry 261, 13905-13910. Roca-Rivada, A., Bravo, S.B., Perez-Sotelo, D., Alonso, J., Castro, A.I., Baamonde, I., Baltar, J., Casanueva, F.F., and Pardo, M. (2015). CILAIR-Based Secretome Analysis of Obese Visceral and Subcutaneous Adipose Tissues Reveals Distinctive ECM Remodeling and Inflammation Mediators. Scientific reports 5, 12214. Rosell, M., Kaforou, M., Frontini, A., Okolo, A., Chan, Y.W., Nikolopoulou, E., Millership, S., Fenech, M.E., MacIntyre, D., Turner, J.O., et al. (2014). Brown and white adipose tissues: intrinsic differences in gene expression and response to cold exposure in mice. American journal of physiology. Endocrinology and metabolism 306, E945-964. Rosenwald, M., Perdikari, A., Rulicke, T., and Wolfrum, C. (2013). Bi-directional interconversion of brite and white adipocytes. Nature cell biology 15, 659-667. Rothwell, N.J., and Stock, M.J. (1979). A role for brown adipose tissue in diet-induced thermogenesis. Nature 281, 31-35. Ryden, M., Uzunel, M., Hard, J.L., Borgstrom, E., Mold, J.E., Arner, E., Mejhert, N., Andersson, D.P., Widlund, Y., Hassan, M., et al. (2015). Transplanted Bone Marrow-Derived Cells Contribute to Human Adipogenesis. Cell metabolism 22, 408-417.

87
Sackmann-Sala, L., Berryman, D.E., Munn, R.D., Lubbers, E.R., and Kopchick, J.J. (2012). Heterogeneity among white adipose tissue depots in male C57BL/6J mice. Obesity (Silver Spring, Md.) 20, 101-111. Saito, M., Okamatsu-Ogura, Y., Matsushita, M., Watanabe, K., Yoneshiro, T., Nio-Kobayashi, J., Iwanaga, T., Miyagawa, M., Kameya, T., Nakada, K., et al. (2009). High incidence of metabolically active brown adipose tissue in healthy adult humans: effects of cold exposure and adiposity. Diabetes 58, 1526-1531. Sakaguchi, M., Fujisaka, S., Cai, W., Winnay, J.N., Konishi, M., O'Neill, B.T., Li, M., Garcia-Martin, R., Takahashi, H., Hu, J., et al. (2017). Adipocyte Dynamics and Reversible Metabolic Syndrome in Mice with an Inducible Adipocyte-Specific Deletion of the Insulin Receptor. Cell metabolism 25, 448-462. Salathia, N.S., Shi, J., Zhang, J., and Glynne, R.J. (2013). An in vivo screen of secreted proteins identifies adiponectin as a regulator of murine cutaneous wound healing. The Journal of investigative dermatology 133, 812-821. Sanchez-Gurmaches, J., and Guertin, D.A. (2014). Adipocytes arise from multiple lineages that are heterogeneously and dynamically distributed. Nature communications 5, 4099. Sanchez-Gurmaches, J., Hsiao, W.Y., and Guertin, D.A. (2015). Highly selective in vivo labeling of subcutaneous white adipocyte precursors with Prx1-Cre. Stem cell reports 4, 541-550. Sanchez-Gurmaches, J., Hung, C.M., and Guertin, D.A. (2016). Emerging Complexities in Adipocyte Origins and Identity. Trends in cell biology 26, 313-326. Sanchez-Gurmaches, J., Hung, C.M., Sparks, C.A., Tang, Y., Li, H., and Guertin, D.A. (2012). PTEN loss in the Myf5 lineage redistributes body fat and reveals subsets of white adipocytes that arise from Myf5 precursors. Cell metabolism 16, 348-362. Scaal, M., and Christ, B. (2004). Formation and differentiation of the avian dermomyotome. Anatomy and embryology 208, 411-424. Scheller, E.L., Cawthorn, W.P., Burr, A.A., Horowitz, M.C., and MacDougald, O.A. (2016). Marrow Adipose Tissue: Trimming the Fat. Trends in endocrinology and metabolism: TEM 27, 392-403. Scheller, E.L., Doucette, C.R., Learman, B.S., Cawthorn, W.P., Khandaker, S., Schell, B., Wu, B., Ding, S.Y., Bredella, M.A., Fazeli, P.K., et al. (2015). Region-specific variation in the properties of skeletal adipocytes reveals

88
regulated and constitutive marrow adipose tissues. Nature communications 6, 7808. Schmidt, B.A., and Horsley, V. (2013). Intradermal adipocytes mediate fibroblast recruitment during skin wound healing. Development (Cambridge, England) 140, 1517-1527. Schulz, T.J., Huang, P., Huang, T.L., Xue, R., McDougall, L.E., Townsend, K.L., Cypess, A.M., Mishina, Y., Gussoni, E., and Tseng, Y.H. (2013). Brown-fat paucity due to impaired BMP signalling induces compensatory browning of white fat. Nature 495, 379-383. Seale, P., Bjork, B., Yang, W., Kajimura, S., Chin, S., Kuang, S., Scime, A., Devarakonda, S., Conroe, H.M., Erdjument-Bromage, H., et al. (2008). PRDM16 controls a brown fat/skeletal muscle switch. Nature 454, 961-967. Shabalina, I.G., Ost, M., Petrovic, N., Vrbacky, M., Nedergaard, J., and Cannon, B. (2010). Uncoupling protein-1 is not leaky. Biochimica et biophysica acta 1797, 773-784. Shan, T., Liang, X., Bi, P., Zhang, P., Liu, W., and Kuang, S. (2013). Distinct populations of adipogenic and myogenic Myf5-lineage progenitors in white adipose tissues. Journal of lipid research 54, 2214-2224. Sharp, L.Z., Shinoda, K., Ohno, H., Scheel, D.W., Tomoda, E., Ruiz, L., Hu, H., Wang, L., Pavlova, Z., Gilsanz, V., et al. (2012). Human BAT possesses molecular signatures that resemble beige/brite cells. PloS one 7, e49452. Shimizu, Y., Nikami, H., Tsukazaki, K., Machado, U.F., Yano, H., Seino, Y., and Saito, M. (1993). Increased expression of glucose transporter GLUT-4 in brown adipose tissue of fasted rats after cold exposure. The American journal of physiology 264, E890-895. Shook, B., Rivera Gonzalez, G., Ebmeier, S., Grisotti, G., Zwick, R., and Horsley, V. (2016). The Role of Adipocytes in Tissue Regeneration and Stem Cell Niches. Annual review of cell and developmental biology 32, 609-631. Stine, R.R., Shapira, S.N., Lim, H.W., Ishibashi, J., Harms, M., Won, K.J., and Seale, P. (2016). EBF2 promotes the recruitment of beige adipocytes in white adipose tissue. Molecular metabolism 5, 57-65. Susulic, V.S., Frederich, R.C., Lawitts, J., Tozzo, E., Kahn, B.B., Harper, M.E., Himms-Hagen, J., Flier, J.S., and Lowell, B.B. (1995). Targeted disruption of the beta 3-adrenergic receptor gene. The Journal of biological chemistry 270, 29483-29492.

89
Svensson, P.A., Lindberg, K., Hoffmann, J.M., Taube, M., Pereira, M.J., Mohsen-Kanson, T., Hafner, A.L., Rizell, M., Palming, J., Dani, C., et al. (2014). Characterization of brown adipose tissue in the human perirenal depot. Obesity (Silver Spring, Md.) 22, 1830-1837. Symonds, M.E., Pope, M., and Budge, H. (2015). The Ontogeny of Brown Adipose Tissue. Annual review of nutrition 35, 295-320. Tang, W., Zeve, D., Suh, J.M., Bosnakovski, D., Kyba, M., Hammer, R.E., Tallquist, M.D., and Graff, J.M. (2008). White fat progenitor cells reside in the adipose vasculature. Science (New York, N.Y.) 322, 583-586. Tchkonia, T., Thomou, T., Zhu, Y., Karagiannides, I., Pothoulakis, C., Jensen, M.D., and Kirkland, J.L. (2013). Mechanisms and metabolic implications of regional differences among fat depots. Cell metabolism 17, 644-656. Tchoukalova, Y.D., Votruba, S.B., Tchkonia, T., Giorgadze, N., Kirkland, J.L., and Jensen, M.D. (2010). Regional differences in cellular mechanisms of adipose tissue gain with overfeeding. Proceedings of the National Academy of Sciences of the United States of America 107, 18226-18231. Timmons, J.A., Wennmalm, K., Larsson, O., Walden, T.B., Lassmann, T., Petrovic, N., Hamilton, D.L., Gimeno, R.E., Wahlestedt, C., Baar, K., et al. (2007). Myogenic gene expression signature establishes that brown and white adipocytes originate from distinct cell lineages. Proceedings of the National Academy of Sciences of the United States of America 104, 4401-4406. Tomiyama, K., Murase, N., Stolz, D.B., Toyokawa, H., O'Donnell, D.R., Smith, D.M., Dudas, J.R., Rubin, J.P., and Marra, K.G. (2008). Characterization of transplanted green fluorescent protein+ bone marrow cells into adipose tissue. Stem cells (Dayton, Ohio) 26, 330-338. Trayhurn, P. (1981). Fatty acid synthesis in mouse brown adipose tissue. The influence of environmental temperature on the proportion of whole-body fatty acid synthesis in brown adipose tissue and the liver. Biochimica et biophysica acta 664, 549-560. Ueta, C.B., Fernandes, G.W., Capelo, L.P., Fonseca, T.L., Maculan, F.D., Gouveia, C.H., Brum, P.C., Christoffolete, M.A., Aoki, M.S., Lancellotti, C.L., et al. (2012). beta(1) Adrenergic receptor is key to cold- and diet-induced thermogenesis in mice. The Journal of endocrinology 214, 359-365. Uezumi, A., Fukada, S., Yamamoto, N., Takeda, S., and Tsuchida, K. (2010). Mesenchymal progenitors distinct from satellite cells contribute to

90
ectopic fat cell formation in skeletal muscle. Nature cell biology 12, 143-152. Ukropec, J., Anunciado, R.P., Ravussin, Y., Hulver, M.W., and Kozak, L.P. (2006). UCP1-independent thermogenesis in white adipose tissue of cold-acclimated Ucp1-/- mice. The Journal of biological chemistry 281, 31894-31908. Ussar, S., Lee, K.Y., Dankel, S.N., Boucher, J., Haering, M.F., Kleinridders, A., Thomou, T., Xue, R., Macotela, Y., Cypess, A.M., et al. (2014). ASC-1, PAT2, and P2RX5 are cell surface markers for white, beige, and brown adipocytes. Science translational medicine 6, 247ra103. Vague, J. (1956). The degree of masculine differentiation of obesities: a factor determining predisposition to diabetes, atherosclerosis, gout, and uric calculous disease. The American journal of clinical nutrition 4, 20-34. Van Harmelen, V., Reynisdottir, S., Eriksson, P., Thorne, A., Hoffstedt, J., Lonnqvist, F., and Arner, P. (1998). Leptin secretion from subcutaneous and visceral adipose tissue in women. Diabetes 47, 913-917. van Marken Lichtenbelt, W.D., Vanhommerig, J.W., Smulders, N.M., Drossaerts, J.M., Kemerink, G.J., Bouvy, N.D., Schrauwen, P., and Teule, G.J. (2009). Cold-activated brown adipose tissue in healthy men. The New England journal of medicine 360, 1500-1508. Velickovic, K., Cvoro, A., Srdic, B., Stokic, E., Markelic, M., Golic, I., Otasevic, V., Stancic, A., Jankovic, A., Vucetic, M., et al. (2014). Expression and subcellular localization of estrogen receptors alpha and beta in human fetal brown adipose tissue. The Journal of clinical endocrinology and metabolism 99, 151-159. Virtanen, K.A., Lidell, M.E., Orava, J., Heglind, M., Westergren, R., Niemi, T., Taittonen, M., Laine, J., Savisto, N.J., Enerback, S., et al. (2009). Functional brown adipose tissue in healthy adults. The New England journal of medicine 360, 1518-1525. Vitali, A., Murano, I., Zingaretti, M.C., Frontini, A., Ricquier, D., and Cinti, S. (2012). The adipose organ of obesity-prone C57BL/6J mice is composed of mixed white and brown adipocytes. Journal of lipid research 53, 619-629. Vohl, M.C., Sladek, R., Robitaille, J., Gurd, S., Marceau, P., Richard, D., Hudson, T.J., and Tchernof, A. (2004). A survey of genes differentially expressed in subcutaneous and visceral adipose tissue in men. Obesity research 12, 1217-1222.

91
Walden, T.B., Hansen, I.R., Timmons, J.A., Cannon, B., and Nedergaard, J. (2012). Recruited vs. nonrecruited molecular signatures of brown, "brite," and white adipose tissues. American journal of physiology. Endocrinology and metabolism 302, E19-31. Wang, Q.A., Tao, C., Gupta, R.K., and Scherer, P.E. (2013). Tracking adipogenesis during white adipose tissue development, expansion and regeneration. Nature medicine 19, 1338-1344. Wang, W., Kissig, M., Rajakumari, S., Huang, L., Lim, H.W., Won, K.J., and Seale, P. (2014). Ebf2 is a selective marker of brown and beige adipogenic precursor cells. Proceedings of the National Academy of Sciences of the United States of America 111, 14466-14471. Wells, H.G. (1940). Adipose tissue, a neglected subject. JAMA 114, 2177-2183. Wilfling, F., Haas, J.T., Walther, T.C., and Farese, R.V., Jr. (2014). Lipid droplet biogenesis. Current opinion in cell biology 29, 39-45. Wu, J., Bostrom, P., Sparks, L.M., Ye, L., Choi, J.H., Giang, A.H., Khandekar, M., Virtanen, K.A., Nuutila, P., Schaart, G., et al. (2012). Beige adipocytes are a distinct type of thermogenic fat cell in mouse and human. Cell 150, 366-376. Xue, B., Rim, J.S., Hogan, J.C., Coulter, A.A., Koza, R.A., and Kozak, L.P. (2007). Genetic variability affects the development of brown adipocytes in white fat but not in interscapular brown fat. Journal of lipid research 48, 41-51. Xue, Y., Petrovic, N., Cao, R., Larsson, O., Lim, S., Chen, S., Feldmann, H.M., Liang, Z., Zhu, Z., Nedergaard, J., et al. (2009). Hypoxia-independent angiogenesis in adipose tissues during cold acclimation. Cell metabolism 9, 99-109. Ye, R., Wang, Q.A., Tao, C., Vishvanath, L., Shao, M., McDonald, J.G., Gupta, R.K., and Scherer, P.E. (2015). Impact of tamoxifen on adipocyte lineage tracing: Inducer of adipogenesis and prolonged nuclear translocation of Cre recombinase. Molecular metabolism 4, 771-778. Young, P., Arch, J.R., and Ashwell, M. (1984). Brown adipose tissue in the parametrial fat pad of the mouse. FEBS letters 167, 10-14. Yu, X.X., Lewin, D.A., Forrest, W., and Adams, S.H. (2002). Cold elicits the simultaneous induction of fatty acid synthesis and beta-oxidation in murine brown adipose tissue: prediction from differential gene expression

92
and confirmation in vivo. FASEB journal : official publication of the Federation of American Societies for Experimental Biology 16, 155-168.
Yue, R., Zhou, B.O., Shimada, I.S., Zhao, Z., and Morrison, S.J. (2016). Leptin Receptor Promotes Adipogenesis and Reduces Osteogenesis by Regulating Mesenchymal Stromal Cells in Adult Bone Marrow. Cell stem cell 18, 782-796.
Zhang, Y., Proenca, R., Maffei, M., Barone, M., Leopold, L., and Friedman, J.M. (1994). Positional cloning of the mouse obese gene and its humanhomologue. Nature 372, 425-432.
Zhao, J., Cannon, B., and Nedergaard, J. (1998). Thermogenesis is beta3- but not beta1-adrenergically mediated in rat brown fat cells, even after cold acclimation. The American journal of physiology 275, R2002-2011.
Zhao, J., Unelius, L., Bengtsson, T., Cannon, B., and Nedergaard, J. (1994). Coexisting beta-adrenoceptor subtypes: significance for thermogenic process in brown fat cells. The American journal of physiology 267, C969-979.
Zingaretti, M.C., Crosta, F., Vitali, A., Guerrieri, M., Frontini, A., Cannon, B., Nedergaard, J., and Cinti, S. (2009). The presence of UCP1 demonstrates that metabolically active adipose tissue in the neck of adult humans truly represents brown adipose tissue. FASEB journal : official publication of the Federation of American Societies for Experimental Biology 23, 3113-3120.