Embed Size (px)
Citation preview

Department of Agricultural Sciences Faculty of Agriculture and Forestry
University of Helsinki Dissertationes Schola Doctoralis Scientiae Circumiectalis, Alimentariae, Biologiae
Publication 4/2017
DIVERSIFYING BOREAL - NEMORAL CROPPING SYSTEMS FOR SUSTAINABLE PROTEIN PRODUCTION
DOCTORAL THESIS Clara Isabel Lizarazo Torres
ACADEMIC DISSERTATION
To be presented, with the permission of the Faculty of Agriculture and Forestry, University of Helsinki, for public examination in lecture room Walter, Agnes Sjöberg street 2. Helsinki, on April
21st, at 12 o’clock noon.
Helsinki, Finland 2017

Supervisor: Professor Frederick Stoddard Department of Food and Environmental Sciences Department of Agricultural Sciences University of Helsinki, Finland. Reviewers: Dr. Glenn McDonald School of Agriculture, Food and Wine The University of Adelaide, Australia. Dr. Roxana Savin Department of Plant Production and Forestry Science University of Lleida, Spain. Opponent: Professor Frank Ewert Leibniz Centre for Agricultural Landscape Research (ZALF). Institute of Crop Science and Resource Conservation (INRES), University of Bonn, Germany. Cover picture: Faba bean and Narrow-leafed lupin flowering. Clara Lizarazo ISBN 978-951-51-3023-5 (Paperback) ISBN 978-951-51-3024-2 (Online PDF) ISSN 2342-5423 (Paperback) ISSN 2342-5431 (Online PDF)

“What shall I learn of beans or beans of me? I cherish them, I hoe them, early and late I have an eye to them…making the earth say beans instead of grass – this was my daily work.”
- Henry David Thoreau, Walden

2
CONTENTS LIST OF ORIGINAL PUBLICATIONS .......................................................................................................... 4
ABBREVIATIONS ........................................................................................................................................... 6
ABSTRACT ...................................................................................................................................................... 7
TIIVISTELMÄ .................................................................................................................................................. 9
RESUMEN ...................................................................................................................................................... 10
1 INTRODUCTION ................................................................................................................................... 12
1.1 Crop diversity in the Boreal – Nemoral region of Europe ............................................................... 12
1.1.1 The Boreal – Nemoral region of Europe ..................................................................................... 12
1.1.2 Crop production in the Boreal – Nemoral region ........................................................................ 12
1.1.3 Dependence on protein imports and fertilizers ............................................................................ 14
1.2 The role of legumes in cropping systems ........................................................................................ 16
1.2.1 Main characteristics of grain legumes ......................................................................................... 16
1.2.2 Constraints for legume crops in Boreal – Nemoral ecosystems .................................................. 18
1.2.3 Benefits of diversified crop rotations .......................................................................................... 19
1.3 Alternatives to solve the protein deficit: grain legumes as potential protein crops in Boreal – Nemoral cropping systems .......................................................................................................................... 21
1.3.1 Faba bean ..................................................................................................................................... 22
1.3.2 Narrow-leafed lupin ..................................................................................................................... 24
1.3.3 Lentil ............................................................................................................................................ 26
2 AIMS OF THIS STUDY ......................................................................................................................... 28
3 MATERIALS AND METHODS ............................................................................................................ 30
3.1 Experimental site ............................................................................................................................. 30
3.1. 1 Weather conditions ................................................................................................................... 30
3.2 Field characteristics ......................................................................................................................... 30
3.2.1 Latokartano field (I, III) ............................................................................................................... 30
3.2.2 Patoniitty field (II) ....................................................................................................................... 32
3.2.3 Bergman field (I, III) ................................................................................................................... 32
3.3 Experimental design ........................................................................................................................ 32
3.3.1 Grain legume germplasm screening (I, III) .................................................................................... 32
3.3.2 Screening for earliness (I) ............................................................................................................... 33
3.3.3 Temperature limits and flowering model for faba bean (III) .......................................................... 33
3.3.4 Screening for nutritive quality (I) ................................................................................................... 34
3.3.5 Crop rotation (II) ............................................................................................................................ 34
3.3.6 Sampling, measurements and indexes ............................................................................................ 34
3.3.7 Plant density ................................................................................................................................... 34
2

3
3.3.8 Statistical analyses .......................................................................................................................... 37
4 RESULTS AND DISCUSSION .............................................................................................................. 38
4.1 Importance and feasibility of protein crop cultivation .......................................................................... 38
4.1.1 Protein production in the Boreal – Nemoral region: potential for local protein yield .................... 38
4.1.2 Nutritive quality characteristics of legume crops grown in the Boreal – Nemoral region ................. 41
4.2 Crop diversity and nutrient availability: grain legumes in rotation ........................................................... 44
4.2.1 Designing sustainable crop rotations: The pre-crop effect ................................................................. 44
4.2.2 Soil health: nutrient availability, uptake and nutrient trade ............................................................ 45
4.3 Grain legume adaptation to short growing seasons ............................................................................... 47
4.3.1 Environmental control of flowering ............................................................................................... 47
4.3.2 Solar radiation ................................................................................................................................ 50
4.3.3 Water deficit ................................................................................................................................... 51
4.3.4 Cultivar temperature limits and ‘earliness per se’ .......................................................................... 51
4.4 Grain legume introduction to Boreal – Nemoral cropping systems and management challenges ........ 52
4.4.1 Faba bean ........................................................................................................................................ 52
4.4.2 NL lupin .......................................................................................................................................... 53
4.4.3 Lentil ............................................................................................................................................... 54
5 CONCLUSION AND FUTURE PROSPECTS ........................................................................................... 56
ACKNOWLEDGEMENTS ............................................................................................................................ 57
REFERENCES ................................................................................................................................................ 59

LIST OF ORIGINAL PUBLICATIONS
This dissertation is based on the following publications:
I. Lizarazo, C.I., Lampi, A-M., Sontag-Strohm, T., and Stoddard, F.L. 2015. Nutritive quality and protein production from grain legumes in a boreal climate. Journal of the Science of Food and Agriculture. 95 (10): 2053-2064.
II. Lizarazo, C.I., Yli-Halla, M., and Stoddard, F.L. 2015. Pre-crop effects on the nutrient composition and utilization efficiency of faba bean (Vicia faba L.) and narrow-leafed lupin (Lupinus angustifolius). Nutrient Cycling in Agroecosystems. 103: 311-327.
III. Lizarazo, C.I., Isotalo, J., Lindfors, A.V., and Stoddard, F.L. 2017. Progress towards flowering of faba bean (Vicia faba L.) is more than photothermal. Accepted in the Journal of Agronomy and Crop Science. 203 (5).
4

5
The author’s contribution to the original publications
I. Clara Lizarazo and Frederick Stoddard contributed to the research plan of the study. The experimental work, measurements and laboratory analyses were carried out by Clara Lizarazo and Jingwei Liu with guidance for fatty acid and starch analysis from Anna-Maija Lamppi, Tuula Sontag-Strohm and Vieno Piironen. Statistical analysis of the data was done by Clara Lizarazo under guidance of Frederick Stoddard. Clara Lizarazo was responsible for the writing of the manuscript and incorporation of the input of other authors.
II. All authors contributed to the research plan for this investigation. The experimental work, measurements and laboratory analyses excluding soil analysis were carried out by Clara Lizarazo. Guidance to soil sampling, analysis, soil science background, was given by Markku Yli-Halla. Statistical analysis of the data was done by Clara Lizarazo with guidance from Frederick Stoddard. Clara Lizarazo was responsible for the writing of the manuscript and incorporation of the input of other authors.
III. Clara Lizarazo and Frederick Stoddard contributed to the research plan. The
experimental work, measurements in field and growth chambers were carried out by Clara Lizarazo with guidance from Frederick Stoddard. Analyses of variance were done by Clara Lizarazo, while statistical models and permutation tests were done by Jarkko Isotalo, with input given by Frederick Stoddard and Clara Lizarazo. Estimations of solar radiation and PAR values were done by Anders Lindfors.

ABBREVIATIONS AA Amino acid
BN Boreal – Nemoral region
Cys Cysteine
DIAAS Digestible indispensable amino acid score
DM Dry matter
ER Efficiency ratio
His Histidine
IAA indispensable amino acid
Ile Isoleucine
K index Sielianinow hydrothermal index
LAI Leaf area index
Leu Leucine
Lys Lysine
Met Methionine
NL Narrow-leafed lupin
N2O Nitrous oxide
PAR Photosynthetic active radiation
PCA Principal Component Analysis
Phe Phenylalanine
PPD Photoperiod
BNF biological nitrogen fixation
Thr Threonine
Trp Tryptophane
Tyr Tyrosine
UI utilization index
UtE Utilization efficiency
Val Valine
VC Vicine-Convicine
6

7
ABSTRACT
Most cropping systems in the Boreal – Nemoral region of Europe are characterized by intensified cereal production, which has resulted in a heavy dependence on foreign vegetable protein imports for feed supplementation and high consumption of synthetic fertilizer. This in turn have caused numerous critical environmental impacts such as copious greenhouse gas emissions from fertilizer production and use, heavy reliance on a narrow range of crop protection methods, leading to risks of resistance to agrochemicals, nutrient runoff and losses in soil health locally, and in land-use change abroad. Hence, crop diversification is needed, and this work focuses on the potential to use grain legumes to help meet the demand for the local vegetable protein and to mitigate the environmental impacts resulting from the current narrow diversity on crop rotations and from the feed and fertilizer trade.
In this dissertation, three grain legume crops, namely faba bean (Vicia faba L.), narrow-leafed (NL) lupin (Lupinus angustifolius L.) and lentil (Lens culinaris Medik.) were grown in field trials in order to assess their potential adaptation to the Boreal – Nemoral region of Europe and to find their best place in the cereal-based crop rotations that are conventional in the region.
The research focused on 1) the protein yield potential of each crop and their nutritive quality particularities, 2) the pre-crop effect of cereals on grain legumes, and 3) the exploration of flowering time in faba bean as a key component of adaptation to high latitudes.
The results show, that faba bean was the crop with highest protein yield stability, and with higher protein concentration than is achieved at lower latitudes, whereas lentil and narrow-leafed lupin had comparable protein concentration as those achieved in other locations. Nutritive quality of all three crops was within the normal range, and amino-acid and DIAAS scores suggested that cultivar selection is important, since major variations in the content of lysine, cysteine and tryptophan influence the feed and food value.
The screening trials revealed that among the available lentil and NL lupin cultivars, earliness is sufficient with some reaching maturity in about 100 days, whereas significant improvement on the earliness of ‘Kontu’ faba bean is needed in order for the crop to be grown in the northern most part of the Boreal – Nemoral region.
The crop rotation trial showed that NL lupin produced equally high yields after turnip rape and oat, while faba bean gave higher yields after turnip rape and then after barley. Overall, the pre-crop effect on nutrient composition of NL lupin was less evident than on that of faba bean, the latter having 19, while the former 7 significant differences out of the 88 nutrient uptake variables measured. Among these 26 significant measures, barley was the best pre-crop in 9 variables, and oat in 5. The pre-crop effect was present on both the shoot and seed composition, and it was apparent that the pre-crop effect was able to influence soil nutrient availability and thus uptake. This study shows some insight about best pre-crop for grain legumes, but the effects need to be tested further to elucidate the mechanisms and to verify the reproducibility of such effect on crop sequences.
The upgraded flowering model showed that flowering control in faba bean in addition to photoperiod and temperature sum, depends on solar radiation (as measured by PAR or sunshine duration, the former providing a better model fit), and water deficit (as measured by the Sielianinow hydrothermal index ‘K’). Understanding the effect of these two new variables in flowering makes it possible to seek more types of variation in earliness be used to identify sources of variation that can serve as material for the selection and development of new cultivars for high latitudes or short seasons.
Overall, this study shows that faba bean and NL lupin have great potential for diversifying crop rotations in the Boreal – Nemoral region of Europe, whereas the susceptibility of lentil to the wet autumns typical of the region will make its management challenging. Each of the crops has different advantages, so they complement each other in terms of optimum soil type, nutrient uptake

and nutrient composition. It is recommended that their cultivation should be promoted not only to solve the vegetable protein deficit, but also to improve the sustainability of cropping systems in the region.
8

9
TIIVISTELMÄ
Viljelyjärjestelmät Boreaalis-Nemoraalisella vyöhykkeellä Euroopassa perustuvat intensiiviseen viljantuotantoon mikä on johtanut riippuvuuteen ulkomaisesta valkuaisrehusta ja runsaaseen lannoitteiden käyttöön. Tämä puolestaan on aiheuttanut runsaasti negatiivisia ympäristövaikutuksia. Esimerkiksi lannoitteiden valmistuksesta ja käytöstä johtuen kasvihuonekaasupäästöt ovat lisääntyneet, suppea kasvinsuojelumenetelmien valikoima on lisännyt rikkakasvien resistenssiriskiä, johtanut ravinteiden huuhtoutumiseen ja maaperän köyhtymiseen kotimaassa sekä aiheuttanut maankäytön muutoksia ulkomailla. Viljelykierron kasvilajiston monipuolistaminen on tarpeen, siksi tässä työssä tähdätään palkoviljojen käytön lisäämiseksi viljelykiertoon, vastaamaan alueellisen valkuaisrehun tuotannon tarpeisiin ja vähentämään nykyisestä yksipuolisen viljelykierron aiheuttamista ympäristövaikutuksista.
Tutkimuksessa on selvitetty härkäpavun (Vicia faba L.), sinilupiinin (Lupinus angustifolius L.) ja linssin (Lens culinaris Medik.) sopeutumista viljeltäväksi vilja-kasveihin pohjaavassa viljelykierrossa Boreaalis-Nemoraalisella vyöhykkeellä Euroopassa. Tutkimuksen painopisteitä ovat 1) proteiinintuotto ja ravitsemukselliset erityispiirteet, 2) edeltävän vuoden viljelykasvin vaikutus palkoviljan satoon, 3) härkäpavun kukinta-ajan selvitys, keskeisenä tekijänä sopeutuminen pohjoisille leveysasteille.
Tulostemme mukaan härkäpavun proteiinisadon stabiilius ja määrä olivat korkeammat kuin ne ovat olleet alhaisemmilla leveysasteilla, sen sijaan sinilupiinin ja linssin proteiinipitoisuudet vastasivat muilta alueilta saatuja tuloksia. Ravinnolliselta laadultaan kaikkien kolmen laatu vastasi yleistä tasoa. DIAAS-tulosten perusteella lajikevalinta osoittautui tärkeäksi, koska havaitut erot kolmen aminohapon, lysiinin, kysteiinin ja tryptofaanin pitoisuuksissa vaikuttavat myös rehun ja ruoan laatuun.
Tutkimustulokset paljastavat myös että linssi- ja sinilupiinilajikkeet ehtivät tuottaa satoa korkeilla leveysasteilla, joidenkin lajikkeiden sadon valmistuessa noin sadassa päivässä. Härkäpapu lajikkeista Kontu sen sijaan edellyttäisi huomattavaa aikaistamista ollakseen tuottoisa Boreaalis-Nemoraalisen vyöhykkeen pohjoisimmissa osissa. Viljelykiertokokeen perusteella sinilupiini oli yhtä satoisa rypsin ja kauran seuraajana, kun taas härkäpavun sato oli parempi rypsin seuraajana verrattaessa satoon ohran jälkeen. Esiviljelykasvilla ei myöskään ollut suurta vaikutusta sinilupiinin ravinnolliseen laatuun verrattuna härkäpapuun, jonka ravinteiden otossa ja käytön tehokkuudessa oli esiviljelykasvista riippuen 19 tilastollisesti merkittävää muutosta ja sinilupiinilla vain 7 muutosta 88:sta mitatusta muuttujasta. Ohra oli paras esikasvi 9 muuttujan suhteen ja kaura 5 muuttujan suhteen. Esiviljelykasvin vaikutus näkyi siementen laadun lisäksi verson ravinnekoostumuksessa. Oletettavasti esiviljelykasvi vaikutti maaperän ravinteiden saatavuuteen. Tutkimus antaa alustavaa tietoa siitä mitkä voisivat olla viljelykierrossa parhaita esiviljelykasveja kyseisille palkoviljoille, mutta lisätutkimuksia tarvitaan mekanismin selvittämiseksi ja osoittamaan kasvinvuorotuksen vaikutus satotuloksiin toistettavasti.
Päivitetty kukinnan malli osoitti että valojakson ja lämpösumman lisäksi fotosynteettisesti aktiivinen säteily (PAR) sekä säteilyn kesto ja vedenpuute (Sielianinowin hydroterminen indeksi K) vaikuttivat kukintaan. Edellä mainittujen kahden uuden muuttujan merkityksen ymmärtäminen voi auttaa löytämään aikaisuuteen vaikuttavia tekijöitä joita voidaan hyödyntää uusien, lyhyeen kasvukauteen sopivien lajikkeiden etsimisessä ja kehittämisessä.
Härkäpavulla ja sinilupiinilla on runsaasti potentiaalia viljelykierron monipuolistajina Boreaalis-Nemoraalisella vyöhykkeellä Euroopassa. Sen sijaan syksyn kosteus näillä alueilla tekee linssin viljelystä haasteellista. Kullakin lajilla on omat maaperä- ja ravinnevaatimuksensa ja ravintokoostumuksensa, joten ne voivat täydentää toisiaan. Palkoviljojen sisällyttäminen viljelyjärjestelmiin pitäisi edistää, ei pelkästään turvaamaan alueen kasviproteiini omavaraisuutta vaan parantamaan myös viljelyjärjestelmien kestävyyttä.

RESUMEN
La mayoría de sistemas de cultivo en la región boreal – nemoral de Europa se caracterizan por una intensa producción de cereales, la cual ha causado una fuerte dependencia en importaciones de proteína vegetal para suplementar el pienso animal y un alto consumo de fertilizantes sintéticos. Esto a su vez, ha provocado graves impactos ambientales, por ejemplo, considerables emisiones de gases de efecto invernadero originados por la producción y uso de fertilizantes, así como gran dependencia en una estrecha gama de métodos de protección de cultivos, lo cual ha conducido a un gran riesgo de resistencia a agroquímicos, lixiviación de nutrientes, pérdidas en la salud del suelo a nivel local y cambios en el uso de la tierra en el extranjero. Por lo tanto, la diversificación de cultivos es necesaria y este proyecto se enfoca en el uso potencial de leguminosas de grano para suplir la demanda local de proteína vegetal y también para mitigar los impactos medioambientales resultado de la estrecha diversidad en las rotaciones de cultivos y del comercio de fertilizantes y pienso para animales.
En esta tesis, tres leguminosas de grano, concretamente habas (Vicia faba L.), altramuces de hoja estrecha (NL de las siglas del inglés) (Lupinus angustifolius L.) y lentejas (Lens culinaris Medik.) fueron cultivadas en experimentos de campo abierto con el fin de evaluar su potencial de ser adaptadas a la región y encontrarles el mejor lugar en las rotaciones de cultivo, basadas en cereales, típicas de la región.
El proyecto se enfocó en 1) el rendimiento proteínico potencial de cada cultivo y en sus características nutricionales, 2) el efecto de cereales como cultivo precedente (pre-cultivo de aquí en adelante) a las leguminosas de grano, y 3) la investigación del tiempo de floración en las habas como componente importante para su adaptación a las altas latitudes nórdicas.
Los resultados indican que las habas son el cultivo con mayor estabilidad en rendimiento proteínico y con mayor concentración proteica que la obtenida a bajas latitudes; mientras que las lentejas y los altramuces presentaron concentraciones proteínicas comparables a aquellas obtenidas en otras latitudes. La calidad nutricional de las tres leguminosas de grano estuvo dentro de un rango normal y los aminoácidos y puntajes DIAAS sugirieron que la selección de variedades es importante, debido a que variaciones notables en su contenido de lisina, cisteína y triptófano pueden influenciar el valor nutritivo de los alimentos producidos a partir de estas.
Los ensayos de campo revelaron que, entre las variedades disponibles de lentejas y altramuces, la precocidad es suficiente con algunas variedades alcanzando su madurez en alrededor de 100 días, mientras que se necesita una mejora significativa en la precocidad de la variedad de haba ‘Kontu’ para efectuar su cultivo en la parte más al Norte de la región boreal – nemoral.
El experimento de rotación de cultivos evidenció que los altramuces produjeron un alto rendimiento después del cultivo de nabina y de avena, mientras que las habas produjeron un mayor rendimiento después del cultivo de nabina y luego del cultivo de cebada. En general, el efecto del pre-cultivo en la composición nutricional de los altramuces fue menos evidente que en la de las habas, pues ésta última presenta 19 diferencias en relación con 7 significativas de la primera dentro de 88 variables relacionadas con absorción de nutrientes. De esas 26 variables significativas, la cebada fue el mejor pre-cultivo en 9 variables, y la avena en 5 variables. El efecto del pre-cultivo estuvo presente tanto en la composición de semillas como en el de tallos, y fue evidente que el efecto del pre-cultivo era capaz de influenciar la disponibilidad de algunos nutrientes en el suelo y por lo tanto su absorción. Este experimento ha producido información útil sobre cuáles pueden ser los mejores pre-cultivos para las leguminosas de grano, pero los efectos necesitan ser evaluados con más detalle para dilucidar cuáles son los mecanismos y para verificar la reproducibilidad de tal efecto en las rotaciones de cultivo.
El modelo de floración que ha sido actualizado en esta tesis, ha demostrado que el control de floración en habas depende no solo del fotoperíodo y de la suma de temperaturas sino también de la radiación solar (medida como RFA o como duración de las horas de sol) y del déficit hídrico
10

11
(medido con el índice hidrotérmico de Sielianinow). El entendimiento de estas dos nuevas variables en la floración posibilita la búsqueda de otros tipos de variación en la precocidad, que pueden ser usados para identificar fuentes de variación y servir como material para la selección y desarrollo de nuevas variedades para latitudes altas o temporadas cortas.
Este proyecto ha demostrado que las habas y los altramuces tienen un gran potencial para diversificar las rotaciones de cultivo en la región boreal – nemoral de Europa, mientras que la susceptibilidad de las lentejas a los otoños húmedos típicos de la región, hacen que su manejo sea difícil. Cada leguminosa de grano estudiada tiene diferentes ventajas, así que se complementan unas a otras en términos de tipos óptimos de suelo, absorción de nutrientes y composición nutricional. Se recomienda que su cultivo sea promovido para solucionar el déficit de proteína vegetal actual y también para mejorar la sostenibilidad de los sistemas de cultivo de la región.

1 INTRODUCTION
1.1 Crop diversity in the Boreal – Nemoral region of Europe
1.1.1 The Boreal – Nemoral region of Europe The Boreal – Nemoral region of Europe - chosen as the area of influence of this research -
includes Norway, Sweden, Finland, Estonia, Latvia and Lithuania and it extends roughly from 55° to 70°N latitude. While the southernmost part of Sweden is considered part of the Continental environmental zone, most of the southern half of the Scandinavian Peninsula, the Baltic countries, and parts of the southern coast of Finland are Nemoral and the rest of the northern part of the region Boreal (Metzger et al. 2013).
In the Boreal – Nemoral region, daylength is one of the key features that changes considerably throughout the year. In winter the sun does not rise above the horizon above the Arctic Circle for some days or weeks, and this period is known as “polar night”, while at the southern edge the shortest days are about 7 hours at 60°N (in southern Finland) and about 10 hours at 55°N (Baldocchi et al. 2000; FMI (Finnish Meteorological Institute) 2016a; LHMS 2016; Time and date 2016). Conversely, in summer the sun does not set, above the Arctic Circle for some days or weeks, and the longest day is 19 hours at 60˚ and 17 at 55˚ (Baldocchi et al. 2000; Tveito et al. 2001; FMI (Finnish Meteorological Institute) 2016b).
Arable land is limited, and often exposed to harsh weather with relatively few frost free days and a small heat sum, so growing seasons are shorter than 120 days in the northernmost parts of the region (Baldocchi et al. 2000). Agricultural production in the Boreal – Nemoral region is challenging and will continue to be. Climate change is expected to prolong the growing season, with increases in temperature of 2- 6°C and in annual precipitation of 6-24% but mainly outside of the growing season (over 50% increase is expected during winter at some locations), which would have considerable effects on nutrient losses due to runoff and leaching (Øygarden et al. 2014). Thus the cropping systems of the Boreal – Nemoral region need to be adapted to both the current harsh conditions and future climate change. 1.1.2 Crop production in the Boreal – Nemoral region
In the Boreal – Nemoral region, the utilized agricultural area (UAA) represents 36% of the total area in Lithuania, 28% in Latvia, 20.8% in Estonia, while Sweden and Finland have only 7.5% and 7.4% respectively, and Norway barely 3% (EUROSTAT 2013).
Cropping systems in the region are simple, being heavily dominated by the small-grain cereals. According to production quantity, wheat is the most important arable crop in the Boreal – Nemoral region being first in all countries except in Finland; barley is the next most important crop in the region, being first in Finland, second in Estonia, third in Sweden and fourth in Latvia and Lithuania (FAOstat 2016). Other arable crops on the top 10 in the Boreal – Nemoral region are oat, rapeseed and rye. Grain legume production in the region is extremely limited and focused on field pea and faba bean, but areas and yields were rather low and stagnant until 2014, when the greening provisions of the EU’s revised common agricultural policy (CAP) started to take effect (Figure 1).
Currently, there is a considerable cultivation of forages (mixed grasses and legumes) in the region, which exceed the production of arable crops in all the countries (FAO 2016a). Although forage legumes in mixture with grasses fix a good amount of N2, their nitrogen fixation is highly susceptible to low temperature particularly in spring, so the mixtures are usually supplied with fertilizers (e.g. slurry) at the beginning of the growing season to improve early grass production, generating environmental costs (Duc et al. 2014; Elgersma and Søegaard 2016). The production of
12

13
grass-forage legume mixtures is mainly used for fodder or silage, but it is not enough to cover all the demand for ruminant feed, since depending on the animal species, growth stage and farm production type (meat or milk), other concentrate feeds are needed, whose ingredients may include by-products such as meal of rapeseed, soybean or other legumes such as faba bean.
The grass-forage mixtures cultivation still depends on fertilizer inputs and their range of end uses narrows down to silage and forage. In contrast, annual grain legumes do not need as much fertilizer inputs, and have a much wider range of possibilities for end uses, being a key source of food and feed in several forms; for example, they can be simply used as grain or to manufacture protein extracts, dehydrated feeds, as green forage, silage, hay. As well grain legumes can be used as green manure (Mihailovic et al. 2005; Mihailovic et al. 2011; Duc et al. 2014). Thus it would be very beneficial and useful to diversify and extend the legume production beyond forage legumes for grass mixtures, and also cultivate grain legumes.
The lack of crop diversity and small percentage of grain legumes on rotation, makes the cropping systems in the region vulnerable due to the loss of biodiversity that cereal mono-cropping represents, and the negative impact on soil fertility and water resources due to the heavy use of fertilizers and pesticides. Furthermore, in truly Boreal systems, crops are often exposed to challenging environmental conditions including frost-thaw cycles, prolonged snow cover, and summer drought spells, resulting in different yield levels, maturity dates and variations in protein concentration (Peltonen-Sainio et al. 2011) that in turn affect the end use and profitability of the crop.
Figure 1. Cultivated area (ha) and yield (t/ha) of faba bean (A, C) and field pea (B, D) in countries of the Boreal – Nemoral region. Source: (EUROSTAT 2016)

1.1.3 Dependence on protein imports and fertilizers
Cultivation of pulses has declined since 1961 throughout Europe (Bues et al. 2013; Voisin et al. 2014) and has always been neglected in the Boreal – Nemoral region (Stoddard 2010; Stoddard et al. 2010). In the 1970s the European Community had policies supporting the cultivation of protein crops (mainly pea, lupin and faba bean) so the cultivated area rose briefly, but in 1992, the CAP implemented a decoupling of subsidies, causing the end of price support for protein-rich crops and since then the land areas sown to legumes decreased sharply (Voisin et al. 2014).
Since 2009 their cultivation started to increase slightly, motivated by the search for protein self-sufficiency and the need for more sustainable cropping systems (Figure 1). After the greening of the CAP in 2013 (Bues et al. 2013), the effect of revisions to the CAP became more visible in the increase in areas in 2015 (Figure 1).
Nevertheless, the expansion of protein crop cultivation remains difficult due to several reasons, such as restrictions in available arable land, trends towards simplification and specialization of cropping systems, net economic value of outputs (marketed and non-marketed), food and feed market conditions, complexity of management (due to soil and weather), and non-recognition and underestimation of legumes ecosystem services (Zander et al. 2016). For example, in France, the obstacles that impede the use of grain legumes in cropping systems, are said to form a ‘technological lock-in’ that mainly promotes cereal cultivation, and this situation holds true in most of Europe (Magrini et al. 2016).
Consequently, for several decades the EU protein deficit has been and still is satisfied by a strong dependency on soybean imports from Brazil, USA and Argentina (Häusling 2011). These imports reached a peak of up to 21 million tonnes in 2002, but thanks to the above mentioned efforts of the EU Parliament and reforms to the CAP (LMC 2009), this amount has decreased notably, and by 2013 imports of soybeans were down to about 16.4 million tonnes (FAO 2016a). Beside soybean imports, there has also been a major import of rapeseed cake meal, with a peak of 51.4 million tonnes in 2013. In the Boreal – Nemoral zone increases in rapeseed cake imports started already in 2004 and coincided with a decrease in soybean imports, which since then have been consistently below 500 thousand tonnes. (FAO 2016a) (Figure 2 A).
Crop choice, both those which are grown and those which are imported, not only has the power to affect local food security, but can also have a strong impact on the environment as a consequence of the global trade in agricultural commodities, where developed countries often receive food supplies from developing ones, thus causing serious nutrient mining and deforestation due to land-use change in the latter (West et al. 2014). The clearest example is that of Brazilian soybean export causing 34% deforestation for the expansion of that country’s cropland and 383-773 kg CO2, 113-119 kg N2O (CO2 equivalent) and 11-41 kg CH4 (CO2 equivalent) (units given in kg CO2 equivalent per ton of soybean) emissions depending on the place of cultivation, transport and energy use (da Silva et al. 2010; West et al. 2014; Godar et al. 2015).
Currently, there is a large and undeniable imbalance between local vegetable protein production and demand, so a complex global trade of protein crops exists, causing severe impacts on land use allocation and resource use. In order to measure these impacts, some indicators have been created, such as the water and nitrogen foot print (NFP). NFP accounts for the total reactive Nitrogen (Nr) that is generated during the production, transport and consumption of a crop commodity (Oita et al. 2016). Oita et al. (2016) determined the NFP of 188 countries and, countries located in the Boreal – Nemoral region are clear major importers of agricultural commodities, being ranked as follows: Norway (15), Lithuania (32), Finland (34), Sweden (40), Estonia (50) and Latvia (53). Moreover, West et al. (2014) reported that the top 3 global crops namely maize, wheat and rice, occupied 33% of cropland but accounted for 68% of global N2O emissions. Apart from the gaseous loss of N, nutrient losses from global agricultural systems are high because inputs currently exceed nutrient removal. Using a simple nutrient mass balance based on nutrient inputs from
14

15
fertiliser and manure and removal in harvested products, West et al (2014) estimated that across the 17 major global crops, inputs of N were 60% higher than crop removal and inputs of P were 48% higher than crop removal causing pollution due to runoff and leaching of the excess nutrients.
Figure 2. Vegetable protein imports (Source: FAOstat 2016) and fertilizer consumption (EUROSTAT 2016) in the Boreal – Nemoral region (Estonia, Finland, Latvia, Lithuania, Norway, Sweden).
The problem of relying on soybean imports, has been aggravated due to the intensive cultivation of cereals, which means that fertilizer consumption in the region is remarkably high, and particularly that of N fertilizers (Figure 2 B). Much of the N fertilizer is lost to the environment, and it has been estimated that half of the N added to agricultural land in Europe is lost and ends up polluting air and water sources (Leip et al. 2014). Such N losses are not only a burden for the environment causing losses in biodiversity, and soil fertility, erosion, and greenhouse gas emissions, but also represent big economic losses that are estimated to be between 0.3 and 1.9% of the European gross domestic product (van Grinsven et al. 2015; Leip et al. 2014).
The steep increase in use of N fertilizers and trade of N-rich feed products (44% of N traded consists of soy-based products), has represented an eight-fold increase in the N trade just between

the period 1961-2010, and there is an evident imbalance worldwide where there are many net N importing countries but few net N exporting countries (Lassaletta et al. 2014). Consequently, there is a severe alteration to the global N cycle, generating several environmental issues for both N importing and exporting countries, related to N pollution and N mining respectively (Lassaletta et al. 2014).
In addition to N imports, the EU is also largely dependent on imports of P rock, and there is an evident excess of P in the environment; for example, in 2005 it was estimated that the net P import in EU-27 was 2392 Gg P and about half of it accumulated in agricultural soils (the accumulation was 4.9 kg P/ha/year) and the other half was lost in waste from different sectors (van Dijk et al. 2016). Such high accumulation of P in soils is often transferred through runoff and erosion to water bodies, and has contributed to eutrophication in the Baltic sea (Granstedt 2012). It is clear that P management is a key issue for the Boreal – Nemoral region, where the majority of countries particularly Finland, Lithuania and Sweden have an annual positive balance meaning net accumulation of P in the soil, while Latvia and Estonia have an annual negative balance meaning soil P depletion, and all countries release significant P loads to the Baltic sea (Granstedt 2012; van Dijk et al. 2016). 1.2 The role of legumes in cropping systems
1.2.1 Main characteristics of grain legumes
Grain legumes have several features that make them a key component for improving cropping
systems and human diets, the main being: 1) their ability to create symbiotic relationships that allow them to biologically fix nitrogen, 2) the high concentration of protein in their seed and of several essential amino acids, and 3) the nutrient richness of their seeds.
Reports of the ability of leguminous plants to fix N date back to 1830 and reports of the isolation of N fixing bacteria from root nodules date from 1888 (Nutman 1969; Burris and Evans 1993).
The process of biological N fixation (BNF) has been studied widely and can be characterized from genetic and biochemical perspectives, but in general can be described as follows: 1) legume roots produce exudates including sugars, amino acids, and flavonoids; 2) flavonoids interact with the soil Rhizobia bacteria through a chemotactic attraction, that in turn induces the transcription of the nodulation (nod) genes; 3) the nod genes are detected in the root epidermis by a receptor complex, which signal induces the curling of the root hair; 4) afterwards an ‘infection thread’ is formed through which the Rhizobia are able to enter the cell wall of the root hair; 5) once inside of the root, the Rhizobia induce cortical hypertrophy in the root cells to create nodule primordia, into which the bacteria are released; 6) when the bacteria infect the nodule primordia, the nodule tissue develops further and the bacteria create the N fixing region; 7) inside the nodules, the enzyme Nitrogenase is responsible for the conversion of atmospheric N2 to ammonia (NH3); 8) the fixed Nitrogen is delivered as either asparagine or ureides (depending on whether the nodule is determinate or indeterminate) through the xylem upwards to the shoot (Brewin 1991; Sinclair and Vadez 2012; Cooper and Scherer 2012; Gresshoff et al. 2015; Burris and Evans 1993).
The symbiotic association described above, can occur between legumes and different Alphaproteobacteria of the family known as Rhizobiaceae (mostly from the genera Rhizobium, Bradyrhizobium and Azorhizobium). A wide biodiversity of bacterial strains exists and they are reported to have different compatibility or affinity to infect a specific host (Lindström et al. 2010; Nutman 1969; Brewin 1991).
The N fixing symbiosis allows legumes to be a source of N for themselves and for the following crop, thus helping to reduce the N fertilizer use, so legumes have been an important tool for crop nutrition in rotations (Brewin 1991; Burris and Evans 1993; Voisin et al. 2014; Pampana et
16

17
al. 2016). The N balance of a crop sequence including legumes may vary depending on: legume species (since different legumes have different BNF capacity), frequency of the legume crop in the rotation, environmental stresses having an impact on BNF (see below), mineralization of crop residues, N leaching, N uptake by the crops (e.g. N removal in the harvested product, particularly by non-legume crops), and management practices (e.g. fertilizer choice and dosage). All these factors may alter the N dynamics in the crop rotation, and in the end the legume crop may contribute different amounts of N into the system depending on the effects of these different environment and management practices (Anglade 2015; Iannetta et al. 2016; Reckling et al. 2016a; Reckling et al. 2016b). Although BNF is responsible for most uncertainties in the N balance, several studies have shown that BNF by legumes such as alfalfa, clover and faba bean has a positive effect in the N balance of crop rotations (Anglade et al. 2015; Reckling et al. 2016a; Reckling 2016b).
Before the development of the Haber-Bosch process, at the beginning of the 20th century for the industrial production of ammonia, it was common to allocate 25-50% of farm land to legume cultivation, but as the N fertilizers became available, draught animals were replaced by machinery and meat consumption became widely affordable, thus legume cultivation was reduced dramatically (Crews and Peoples 2004).
Environmental stresses and host plant factors can alter the BNF rates, for example photosynthesis rates, carbon exchange rates and mineral nutrition (the most critical being Fe, P, K and S), soil pH, drought, salinity and heat (Divito and Sadras 2014; Dwivedi et al. 2015). In addition, there are key management practices that can influence the rates of BNF such as inoculation of seeds before sowing, a precise dose of starter N, and tillage practices (minimum tillage stimulating BNF) (Kessel and Hartley 2000).
The grain filling of legumes often requires not only all of the biologically fixed N, but also remobilization of N from vegetative tissues. Since there are differences in BNF levels among crops and also variations in BNF efficiency after flowering, there are differences in the need for N remobilization (Pampana et al. 2016).
The grain legumes are among the most protein-rich of crops, as their protein concentration ranges from 20% in pea and common bean to up to 40% in white lupin and soybean. From 50 to 90% of the protein is in the form of globulins (vicilin and legumin), which have been acknowledged to have medicinal and pharmacological as well as nutritional value, and are excellent for use in development of other food products (Roy et al. 2010; Nikolic et al. 2012). The rest of the protein fraction is in the form of albumins or glutelins, representing10-20% of the total protein concentration depending on species (Roy et al. 2010). Although legumes are generally deficient in the essential S-containing amino acids (methionine and cysteine) and tryptophan, they are rich in other essential amino acids, particularly lysine that is deficient in cereals.
After protein, the most important fractions of grain legume seeds are starch, fiber and oil (Table 1), the latter being particularly high in some species such as soybean which is an important source of edible oil (Gallardo et al. 2008). In addition, different legume species contain different ranges of bioactive compounds, including phenolic acids, protease inhibitors, lectins, isoflavones and flavones, phytosterols, saponins and pyrimidine glycosides. Compounds such as trypsin inhibitors, tannins and phytic acid, which are considered anti-nutritional and reduce the bioavailability of mineral nutrients, digestibility and palatability.
The seeds are also rich in mineral nutrients (mainly Ca, Cu, Fe, Mg, P and Zn), vitamins (e.g. folate, niacin, riboflavin, panthotenic acid, and tocopherol), and fatty acids (e.g. linoleic and linolenic acids) (Grela and Günter 1995; Campos-Vega et al. 2010; Nikolic et al. 2012; Zhou et al. 2013). The unique composition of grain legumes makes them cholesterol-free, gluten-free and they have low glycemic index among several other health benefits (FAO 2016b).

Table 1. General overview of main nutrient and anti-nutrient fractions present in the 3 grain legumes focus of the present study. Component Faba bean NL lupin Lentil Protein 26-38 % 29-37 % 15-31 % Starch 41-44 % <1% 42-44 % Non-starch polysaccharides <1% 49-50 % <1% Fiber 8-10 % 11-14 % 6-10 % Sugars 3.5-4.4 % 5-6 % 4-6 % Oil 1.3-2 % 5-8 % 0.3-3.5 % Phytic acid 11-20 g/kg 2.5-3.3 g/kg 3-12 g/kg Tannin 0-21 g/kg <0.02 mg/g 4-10 g/kg Vicine 5-7.6 mg/g NP NP Convicine 2-3.6 mg/g NP NP TIA* 1.7-3.5 mg/g ND 1,94-3,07(mg/g) Alkaloids NP 0.25-1 g/kg NP Reference (Makkar et al. 1997;
Crépon et al. 2010; Jiang et al. 2016; Pulkkinen et al. 2016).
(Lampart-Szczapa et al. 2003; Torres et al. 2005; Sujak et al. 2006; Beyer et al. 2015).
(Wang and Daun 2006; Erskine et al. 2011).
*Trypsin inhibitor activity NP= not present, ND= not detected 1.2.2 Constraints for legume crops in Boreal – Nemoral ecosystems
Increasing legume production in the Boreal – Nemoral region is feasible even though it is made difficult by environmental and production constraints. The main existing environmental constraints are the short growing season and night frosts (particularly during late spring and early autumn), and climate change that is expected to alter rainfall patterns and increase temperatures, which in turn will increase the frequency of heat and drought stress (Olesen and Bindi 2002; Iglesias et al. 2012; Peltonen-Sainio et al. 2013).
Among the environmental constraints, the most critical is the short growing season that in some parts of the region can be as little as 1000 GDD, which is considered insufficient for production of currently available grain legumes (Stoddard et al. 2009). Grain legumes need between 900 and 2000 GDD to reach maturity, depending on the species, cultivar, and on how other environmental fluctuations affect reproduction (Thomson et al. 1997). In addition, grain legumes are particularly susceptible to high temperatures near flowering time (Siddique et al. 2012). Thus, there is an urgent need that breeding programs develop not just early cultivars, but early cultivars tolerant to heat and drought stresses.
Besides breeding efforts, more studies on key phenological stages such as the onset of flowering are needed, to identify germplasm adapted to new target environments and climatic risks, and to adjust management practices in order to maximize productivity and reduce the exposure to environmental stresses (Chloupek and Hrstkova 2005; Vadez et al. 2012).
Among the production constraints is the lack of public policies and support systems to provide farmers the incentive to grow more grain legumes (Voisin et al. 2014). European farmers and hence those in the Boreal – Nemoral region have neglected legume cultivation, due to the view that legume crops are less competitive than cereals, in terms of yield levels, yield stability, market
18

19
price, and seeding costs (Von Richthofen and GL-Pro partners. 2006; Cernay et al. 2015). In addition, farmers are not positioned to take advantage of the wide potential end uses for grain legumes such as nutraceutical products and protein isolates in the food industry. Furthermore, there is insufficient local infrastructure and markets for these applications, so the current production chain is limited to feed uses.
Cernay et al. (2015) showed that although legumes have less yield stability than cereals, their environmental benefits and valuable potential uses for specialized markets can compensate for some of the yield penalty. The lower yields and yield stability of legumes is likely to be due to the long growing cycle that increases their exposure to environmental stresses, the lower speed for developing a closed canopy, and lower PAR interception during the life cycle when compared to cereals (Giunta et al. 2009; Cernay et al. 2015).
Moreover, the differences in yield stability among grain legumes vary depending on the region where they are grown: lupin was shown to have the highest variability, but faba bean and pea were shown to be the least variable for south-western and northern Europe, respectively (Cernay et al. 2015). The reported low yield stability of lupin in northern Europe may be debatable since there were only 22 observations for lupin while for most other species there were 53.
The need for reducing the dependency on legume imports should not be the only motive for increasing legume cultivation. Many ecological services are gained from a diverse, legume-supported crop rotation, and the need to protect the soil resources and to reduce the nitrous oxide emissions from agriculture are of utmost importance (Stoddard et al. 2009; Peltonen-Sainio et al. 2013). Legumes give many benefits to the productivity of agricultural systems (Peoples et al. 2009) but many of those cannot be monetarized, so financial incentives are needed in order to compensate for the losses caused by low yields and yield stability issues (Reckling et al. 2016a; Bues et al. 2013; Cernay et al. 2015; Zander et al. 2016) 1.2.3 Benefits of diversified crop rotations
Lack of diversity in crop rotations in the Boreal – Nemoral region is a general issue. As discussed in section 1.1.2, most crop rotations do not involve legumes often enough, and continuous cereal cropping of wheat, barley, grasses and pasture is the norm. Such oversimplified cropping systems have led to severe nitrogen losses, reported to be up to 30 kg/ha in Sweden, and less than 10 kg/ha in Estonia, due to the large fertilizer applications causing surpluses of N and P that are then lost due to water runoff (Vagstad et al. 2004). Consequently, modifications to management practices have been suggested, such as in tillage, green manure and catch crops (Myrbeck and Stenberg 2014; Valkama et al. 2015; Aronsson et al. 2016).
In contrast to the simple crop rotations practiced in the Boreal – Nemoral region, diversity in a rotation usually gives higher yields and a range of other benefits. The classic example is that of cereals after either a grain legume or an oilseed (such as turnip rape or linseed (Linum usitatissimum). The benefits of a diverse rotation and choice of a favorable preceding crop can be measured not only in terms of yield quantity and quality (which are easy to quantify), but also in terms of root growth and decrease in the pressure from pests and weeds (which are more difficult to quantify) (Reckling et al. 2015; Reckling et al. 2016b). The majority of studies that have measured the pre-crop effect, have focused on wheat, barley and canola, and consistently have shown a higher increase in yields when the pre-crop is a legume than when it is a non-legume. For example, Angus et al. (2015) reported that wheat yield increased by 0.5 t/ha after oats, while the increase was up to 1.5 t/ha after grain legumes. Other studies support that cereal yields consistently increase when the pre-crop is a broad-leafed crop, with the yield increase ranging from 20% after an oil crop to up to 60% after a legume crop (Kirkegaard et al. 2008; Angus et al. 2015).
Crop rotation experiments that seek to assess pre-crop effects can be arranged differently, varying either the frequency (in years) or the number of break crops. For example, it is possible to

have a single break crop1 (B-C-B-C), use two different break crops (B1-B2-C-B1-B2-C), or to test the persistence effect of the break crop on two consecutive crop years (B-C-C) (Angus et al. 2015). There is a wealth of studies about the benefits of break crops and pre-crops for cereals, wheat being the most studied case, while pre-crops for legumes are least studied.
Although legumes are a key crop for the diversification of cropping systems, evidence and characterization of a pre-crop effect for them is seldom reported, perhaps due to the minor role that they play in the world market, when compared to cereals. Nevertheless, it is possible to find some evidence of pre-crop effect for legumes when the rotations include two break crops, such as a 3 year study, where 10 break crop alternatives (including lupin, field pea, canola among others) for wheat were grown in a 10 x10 matrix over two years, resulting in 100 different crop sequence options (Malik et al. 2015). The second year of this crop rotation showed a significant effect of year 1 crops on year 2 crop in maximum dry matter, N mineralization, grain yield and weed levels; for example in NL lupin the lowest amount of weeds was observed when grown after barley, and highest grain yield were obtained after oaten hay 1050 kg/ha , field pea 1000 kg/ha and barley 930 kg/ha (Malik et al. 2015).
Benefits of legumes as pre-crops for cereals include increases in nutrient and water availability, soil mineral N budget, C sequestration, and energy efficiency, along with reductions in use of fossil fuels and weed levels (Gan et al. 2003; McConkey et al. 2003; Malhi and Lemke 2007; Nemecek et al. 2008; Peoples et al. 2009; Angus et al. 2015). Some of the benefits from crop rotations arise from the break of disease cycles, and from the influence that different crops have on available and total nutrient content in the soil. Different crops differ in their root residue composition, root channels and exudates, and they can cause variations in many soil chemical and physical parameters such as pH, soil organic carbon sequestration (SOC) and microbiota, thus altering micronutrient availability (Khoshgoftarmanesh et al. 2011). For example, phytoavailable Cu and P were reported to be higher in long-term leguminous cropping and cereal-legume rotations than in continuous wheat (Khoshgoftarmanesh et al. 2011), and P availability is also recognized to increase significantly after legumes in rotation (Pypers et al. 2007). Although it is known that differences in nutrient composition of crop parts and nutrient uptake among crops can affect the cycling of nutrients in crop sequences, few sequences have been examined in this regard.
Differences on synthetic fertilizer inputs and in the decay of different crop residues can lead to higher or lower nitrous oxide emissions depending on the order and diversity of crop sequences (Freibauer and Kaltschmitt 2003; Schwenke et al. 2015; Nemecek et al. 2008). For example, although oilseeds do not produce large crop residue pieces they are nitrogen-rich and depend solely on nitrogen input, potentially leading to high N2O emissions (McConkey et al. 2003; Freibauer and Kaltschmitt 2003; Schwenke et al. 2015). On the other hand, although legume crop residues are also rich in N, it has been shown that N2O emissions tend to be much higher in N fertilized crops than in grain legume crops and in legume pasture lands: for example, canola emitted a mean of 2.65 kg N2O-N/ha whereas faba bean, field pea and alfalfa emitted 0.41, 0.65 and 1.99 kg N2O-N/ha, respectively (Jensen et al. 2012). Thus legumes are a good option to reduce emissions from cropping systems, and thus should be included more often and strategically on rotations.
Finally, a complex crop rotation that includes legumes, increases the overall farm landscape heterogeneity and influences the diversity of pollinators, and so has considerable effects on crop pollination rates (Andersson et al. 2014). Increasing the proportion of legumes such as faba bean and lupins in farming systems is more beneficial than cereal crops for pollinator population density and species richness (e.g. bumblebees), since the papilionaceous flowers of legumes have evolved for bee-mediated pollination (Pywell et al. 2006; Andersson et al. 2014).
Efficient nutrient cycling, through the inclusion of legumes in rotation, could not only facilitate the rational use of fertilizers but also improve the nutritive composition of crops, so
1 B= break crop, C= main crop.
20

21
studies are still needed to evaluate the different patterns of nutrient uptake and how nutrient composition can be affected due to crop sequences. Identifying which pre-crops are best for grain legumes should be a priority, since it has the potential to help in their adaptation to the Boreal – Nemoral region, and in the designing efficient and productive cropping sequences, that may promote their cultivation.
1.3 Alternatives to solve the protein deficit: grain legumes as potential protein crops in Boreal
– Nemoral cropping systems
Currently, the protein deficit in the EU continues fluctuating between 70-80% (Martin 2014), representing about 20 million hectares of land cultivated elsewhere, from which many socio-environmental impacts ensue (Godar et al. 2015). Hence several alternatives have been proposed and some locally produced alternatives have been identified.
De Visser et al. (2014) showed that oil crops were more competitive than starch crops as alternatives to soybean, but warned that alternative protein crops will become feasible only when they have reached the yield level of soft wheat, which is not realistic for narrow-leafed lupin which would need the greatest increase, 334%. Parallel assessments by Martin (2014) indicated that more than one strategy is needed, mainly a) to ease the access to already existing sources of vegetable protein such as oilseeds or new protein sources for feed such as insect proteins, b) promote research to improve the competitiveness of EU protein crops, and c) increase feed efficiency to reduce protein consumption.
Other authors have proposed that the protein production from barley, wheat and oats should not be disregarded, and indicated that advanced breeding lines of cereals can exceed the protein yield of rapeseed and even out-compete modern pea cultivars (Peltonen-Sainio et al. 2012). Nevertheless, such cereal protein production may not be realistic under climate change conditions, because the increase in CO2 concentrations is likely to cause a reduction in the grain protein concentration of crops, and the percentage change is estimated to be much higher for cereals (e.g. wheat) than for legumes (e.g. field peas or soybeans) (Myers et al. 2014). Furthermore, this protein production can be achieved only with substantial inputs of nitrogen fertilizer.
There have been other strategies and approaches suggested to deal with the current protein deficit at the country level. In Sweden, for example, it has been estimated that there is an intake of about 35 g meat protein/day, that to replace 40% of it with grain legumes 60 000 ha would be needed (which is less than 1% of the country’s arable area), and common bean (Phaseolus vulgaris L.) and faba bean were identified as suitable crops for this purpose (Carlsson 2014).
In Finland, it has been estimated that current protein crop production can be increased, but it would be limited by the total arable land available, considering that protein crops are recommended to be grown only once in every 4-5 year interval (to avoid diseases and pests), so rapeseed and legumes could each have a 20% share of the total arable land in the country (Peltonen-Sainio and Niemi 2012). In addition, opportunities resulting from climate change will allow protein crop cultivation to reach 390 000ha by 2055, which could replace between 50% and 60% of the imported soybean meal (Peltonen-Sainio et al. 2013). Such massive changes in area of crops as result of climate change, will displace other mainstream crops such as barley and oat, but the implication of that is beyond the reach of this thesis. The suggested protein crop candidates with potential for increased cultivation were faba bean, field pea and rapeseed (B. rapa and B.napus, but mainly the latter) (Peltonen-Sainio and Niemi 2012; Peltonen-Sainio et al. 2013).
Although rapeseed cake is valuable and suitable to replace some of the imported soybean cake (OECD/FAO 2015), opportunities for expanding rapeseed areas are rather limited since it already is an important crop (Peltonen-Sainio et al. 2013).

More importantly, rapeseed does not provide key ecosystem services that grain legumes do, such as BNF (Peoples et al. 2009; Voisin et al. 2014) nor does it have the nutraceutical value of crops like NL lupin (Rajesh et al. 2015; Cabello-Hurtado et al. 2016).
Currently pea is the most cultivated grain legume crop in the region (EUROSTAT 2014), and it has long been a widely accepted crop in Europe, but its production is now challenged by the soil-borne fungus Aphanomyces euteiches (Peltonen-Sainio et al. 2013; Voisin et al. 2014), so other more resilient and productive grain legume species are needed to diversify the cropping systems and to provide protein.
At the start of this project, there was little information on the adaptation of other grain legumes to very long summers typical of the Boreal – Nemoral region. Preliminary field tests in 2009 including soybean, narbon bean (Vicia narbonensis L.), fenugreek (Trigonella foenum-graecum), white lupin (Lupinus albus L.), Andean lupin (Lupinus mutabilis), NL lupin faba bean , and lentil, highlighted the potential of the latter 3 species for deeper investigation for the Boreal – Nemoral region (Stoddard et al. 2010). At the moment the production of these 3 grain legume crops in Europe is not widespread (Figure 3), but they have potential to be adapted, so they are the focus of this study.
Figure 3. Average world production share during the period 1993-2013 for each of the candidate grain legumes. Data are shown for lupin spp. in general, since specific data for NL lupin are not available (FAO 2011; EUROSTAT 2016).
1.3.1 Faba bean
Faba bean is one of the most important grain legumes crops worldwide, being cultivated on 2.5 million hectares with an average production of 4.0 Tg in the period 1993-2013 (FAOstat 2016; FAO 2016b). In 2015, the top producing countries in Europe were UK with 170000 ha, followed by France with 86000 ha and Italy with 48000 ha, while in the Boreal zone the top producer was Lithuania with 61000 ha followed by Latvia and Sweden, both cultivating over 25000 ha (EUROSTAT 2016).
Faba bean is adapted to most European climates. It is grown as an autumn-sown crop where winters are mild, in the Mediterranean and Atlantic climatic zones, and as a spring sown crop where summers are sufficiently moist, outside the Mediterranean zone (Link et al. 2010).
Faba bean is a cool season crop, well adapted to the wet season of semi-arid regions with 400-600mm rainfall and a temperature range of between 18-27 °C with an optimum around 22-23 °C (Patrick and Stoddard 2010; Lim 2012c). Conditions outside this range cause considerable losses of yield stability (Jensen et al. 2010; Duc et al. 2011; Flores et al. 2013). Yield stability needs to be improved to renew European interest in faba bean cultivation (Flores et al. 2013).
22

23
Although there is wide variation in drought tolerance among faba bean genotypes, the crop is usually considered to be drought sensitive, since drought can cause losses of up to 50% in seed yield (Mwanamwenge et al. 1999; Khan et al. 2007). Some germplasm escapes terminal drought by early flowering and early set of pods and seeds, while the early podding stage is considered the most sensitive to drought stress (Siddique et al. 2001; Mwanamwenge et al. 1999). The crop is grown either irrigated or rain-fed, for example in South Australia the majority of the cultivation is in areas with about 400 mm of rainfall (Siddique et al. 2013; Mwanamwenge et al. 1999)
The crop grows best on heavy clay soils with a pH of 6.5-9, and is reputed to grow poorly on acid soils, which are reported to have detrimental effects on plant growth, root proliferation, nodulation and consequently on nitrogen fixation (Jensen et al. 2010; Ouertatani et al. 2011). Nevertheless, the study by Rose et al. (2010) which evaluated the growth and P acquisition of 50 faba bean genotypes on alkaline and acid soils, showed that the crop is able to grow on both soil types and can greatly modify the rhizosphere pH, by using the root exudates citrate and malate, which help it to decrease the soil pH in alkaline soils and increase it in acid soils. Its P acquisition in a pot experiment was 3 fold higher on acid soils, than on alkaline soils (Rose et al. 2010).
Faba bean leaves are paripinnate and consist mostly of 3 pairs of leaflets. Axillary racemes bear up to 15 flowers, most commonly 4-6 (Stoddard and Bond 1987; Stoddard 1993) typically with a black spot (Figure 4A). Two recessive mutations, zt1 and zt2, confer white flower color (Figure 4B) and low tannin in the seed coat (Crépon et al. 2010). High temperatures and water deficit often cause flower abortion and other flowers abscise due to lack of pollination, so only a small proportion of flowers become pods, and a mature plant will usually bear about 4-14 pods (Figure 4C), although up to 32 pods per plant have been reported depending on plant density and water stress (Mwanamwenge et al. 1999; López-Bellido et al. 2005; Patrick and Stoddard 2010; Flores et al. 2013). The plant height can reach 60-200 cm depending on cultivar and environmental conditions, although the stem is not wind resistant and lodging is a problem.
Faba bean has a vigorous tap root that reaches a depth of about 50-90 cm (Manschadi et al. 1998) but its root system is often said to be “shallow” (Figure 4D) (Muñoz-Romero et al. 2011) particularly in spring-sown cultivars (Jensen et al. 2010). Among the major cool season legume crops, faba bean is the one that biologically fixes the most N, with reported values of up to 326-648 kg ha/yr of N (Jensen et al. 2010; Bruning and Rozema 2013; Pampana et al. 2016).
The protein concentration of faba bean seeds is between 26 and 36% and tends to be slightly higher in zero-tannin and low-vicine-convicine (VC) genotypes than in their wild-type counterparts (Borisjuk et al. 1995; Duc et al. 1999; Lim 2012c). Starch is the main component, reaching up to 42% in seeds, and has a negative correlation with protein concentration (Crépon et al 2010). High quality protein isolates can be made from faba bean seeds as potential functional ingredients for food, which might be helpful in opening new markets and improving the crop value, but for achieving this the development of industrial processes is needed to improve the flavor and techno-functional properties of the faba bean flour (Vioque et al. 2012; Jiang et al. 2016).
The seeds are also rich in mineral nutrients (e.g., Ca, Mg, P), vitamins (e.g. B6, A, E, folate) and lysine (Crépon et al. 2010; Lim 2012c). They have been accounted to have several health properties such as cholesterol lowering, antibacterial activity, antioxidant activity due to the presence of flavonoids, and benefits in the treatment of Parkinson’s disease due to the high content of levo-dihydroxy phenylalanine (L-DOPA), the precursor of dopamine (DA) (Lim 2012c).
There are several anti-nutritional factors present in the seeds, such as vicine and convicine, raffinose oligosaccharides, tannins, protease inhibitors, trypsin inhibitors, and phytic acid, but their levels depend on the cultivar, so there are options suitable for food and feed.
Faba bean seeds have been proven to be suitable for pigs and poultry, but preferably using cultivars with low tannin and vicine-convicine content, since these have an additive effect that decreases the protein digestibility and energy value of the feed (Vilariño et al. 2009; Crépon et al. 2010). The most limiting components in faba bean are VC so cultivars with lower VC have been

bred using the recessive gene vc, but unfortunately they are not yet commercially available for the Boreal – Nemoral region (Pulkkinen et al. 2016). Hence the choice of faba bean cultivar for animal feed use depends on the animal species, since they may have different tolerances for VC levels, with effects not only on the apparent metabolizable energy but also on factors such as reduced egg size in poultry. It is recommended to use only up to 7% of faba bean in laying hen diets using cultivars with normal VC or 20% of cultivars with low VC (Crépon et al. 2010).
Figure 4. Faba bean morphology: axillary racemes of A) colored flowers, B) white flowers,
C) pods, and D) nodulated roots 1.3.2 Narrow-leafed lupin
NL lupin is one of the least popular grain legume crops worldwide, being mostly grown in Australia where it is cultivated on 517000 ha (FAOstat 2016). Currently the average world production is rather low at 0.77 Tg in 2013, when compared with record production that peaked in 1999 with over 2.1 Tg (FAOstat 2016). Europe accounts for only 12% of the world production, the top producing countries being Poland with 80000 ha, followed by Russia with 50000 ha and Germany with 21000 ha (FAOstat 2016). Most lupin cultivation takes place in Oceania (75.3%) while its cropping area is low in Europe (17.6%) (Lucas et al. 2015).
NL lupin is a cool-season grain legume that grows best at 21/16 °C in well drained, moderately acidic soils, but does not grow well on alkaline soils, due to its sensitivity to free Ca2+ and due to the rarity of the appropriate Bradyrhizobium in these soils (Brand et al. 1999; Lim 2012b; Lucas et al. 2015).
24

25
NL lupin has palmately compound leaves with 5-11 narrow leaflets, hence both the common and Latin names. The crop has an erect growing habit; the stem can reach up to 1.5 m in height and ends in a terminal inflorescence (Lim 2012b). Like faba bean, NL lupin produces an excess of flowers, many of which abort so only about 4-9 flowers in a raceme achieve pod and seed set (Figure 5 A-B). The main stem can have several orders of repeated branching or can have restricted branching depending on the cultivar. The latter are of two kinds: either with apical lateral branches being replaced by a solitary flower or a main stem with fewer leaves than in a normal branching type (Dracup and Thomson 2000).
NL lupin has a strong root system characterized by a long tap root (Figure 5C). A screening of wild genotypes of NL lupin in Western Australia estimated that mean root length is about 415 cm, but the range was from 99 cm to 1793 cm (Chen et al. 2012). NL lupin nitrogen fixation rates are reported to be between 26 and 244 kg N/ha (Williams et al. 2014).
NL lupin is known for the excellent nutritive content of its seeds, with a protein concentration ranging between 30-37% depending on the cultivar (Villarino et al. 2015). The chemical composition of its storage protein is made up by conglutin γ (water- and salt-soluble globulin) and conglutin δ (water-soluble albumin), which are both sulfur rich, and conglutins α and β which are the main globulins (Muranyi et al. 2016). The conglutin γ fraction is of great interest due to its relatively high content of methionine and cysteine, which are the most limiting amino acids in most grain legumes (Lim 2012b), and also due to its hypoglycemic activity, which make it useful to control insulin levels in diabetic patients (Foley et al. 2011; Lucas et al. 2015; Villarino et al. 2015; Carbonaro et al. 2015).
Figure 5. Narrow-leafed lupin morphology A) flower raceme B) Pod set C) Tap root (lateral roots are broken)
Besides their high protein concentration, NL lupin seeds are also characterized by their low starch content (which confers on them a low glycemic index) because they store energy in the form of β-galactan instead (Cheetham et al. 1993) and also other non-starch polysaccharides that are reported to have antioxidant and immunostimulatory properties (Rajesh et al. 2015). NL lupin seeds are rich in dietary fiber content, carotenoids, vitamins and minerals, which add to its unique nutritive composition and confer several health benefits (Pate et al. 1985; Sujak et al. 2006). Hence NL lupin flour or fractions have been added to wheat bread to improve the nutritional content and health functionality, and also has been used to prepare protein isolates for different food and non-

food industrial applications (Wong et al. 2013; Sussmann et al. 2013; Foley et al. 2011; Villarino et al. 2015).
Among the anti-nutritional factors present in NL lupin seeds, the most important are the quinolizidine alkaloids, at 0.25-1 g/kg which are of help against pathogens and herbivores, but that are undesirable for feed and food uses. Breeding programs have significantly reduced the content of alkaloids in NL lupin seeds, but it has not been eliminated completely, so there is a threshold of 0.05% and 0.02% for food and feed uses, respectively (Sujak et al. 2006; Vilariño and Ravetta 2008; Philippi et al. 2016). 1.3.3 Lentil
Lentil is one of the most popular grain legume crops worldwide, being cultivated on over 4.5
million ha (FAOstat 2016). The average world production for the period 1993-2013 was of 3.4 Tg (FAO 2016b). Europe accounts for barely 1.3% of the world production, the top producing countries being Spain with 31000 ha, followed by Russia with 26000 ha and France with 14000 ha (Erskine 2009; FAO 2016b).
Lentil is also a cool season grain legume, cultivated extensively in the semiarid regions of the world, where yields are relatively low (Lim 2012a). Its optimum temperature is 24 °C, but it can adapt to temperatures within the range of 6-27˚C and it grows well in all soil types, preferring well drained sandy loam soils rich in P and K with a neutral pH, but can tolerate pH within the range 4.5-9 (Yadav et al. 2009; Lim 2012a).
Lentil has herbaceous stems and a low bushy canopy that reaches 15-75 cm in height, depending on the environment. The leaves are pinnate with a tendril at the tip and are alternate with 4-12 pairs of leaflets, while the racemes bear 1-4 flowers and up to two pods with 1-3 seeds each (Figure 6) (Saxena 2009; Lim 2012a).
Figure 6. Lentil morphology. A) axillary raceme of flowers B) pinnate leaves with tendrils
at the tip, forming a bushy canopy C) nodulated roots. The root system is more slender than those of NL lupin and faba bean, but it is able to
produce nitrogen fixing nodules as early as 15 days after emergence (Saxena 2009). Nitrogen fixation levels in lentil have been estimated at between 5-191 kg/ha (Unkovich and Pate 2000; Cooper and Scherer2012).
26

27
The protein concentration of lentil seeds ranges between 24 and 30% depending on the cultivar (Stoddard et al. 1993; Wang and Daun 2006; Sharma 2009). Lentils are a rich source of mineral nutrients (e.g. K, P, Fe and Zn), vitamins (e.g. C, A, E, folate), and are relatively high in lysine and tryptophan (Grusak 2009). The main component of lentil seeds is 43%-47% starch (Grusak 2009).
Due to its lower yield and protein concentration, lentil is seldom used for feed purposes, but preferred for human consumption thanks also to its rich flavor and health benefits that include antioxidant activity, anti-carcinogen effects and ability to lower triacylglycerol and cholesterol levels (Lim 2012a). Lentils contain anti-nutritional factors such as trypsin inhibitors (1.9-3%), phytic acid (0.3-1.2%), tannins (0.4-1.01%) and oligosaccharides, the levels of which depend on cultivar, and can limit the bioavailability of nutrients and lower digestibility (Wang and Daun 2006).

2 AIMS OF THIS STUDY
The main aim of this study was to explore the feasibility of grain legume cultivation under the short growing season characteristic of the Boreal – Nemoral region, in order to diversify its conventional, cereal-based rotations.
First, the study focused on germplasm of faba bean, NL lupin and lentil and aimed to screen a wide range of cultivars of each, for evaluation of their yield, nutritive quality, and phenology and to assess their adaptation and potential cultivation in the Boreal – Nemoral region.
Second, the study included a crop rotation experiment, aimed at identifying best pre-crop sequences for grain legume crops, in order to design nutrient-efficient legume-supported crop rotations. The pre-crop effect was identified using various indexes of nutrient use efficiency.
Third, the study aimed to evaluate and describe additional environmental factors influencing flowering and thus earliness in faba bean. Information from field and growth chamber experiments was combined to build an empirical model to update the existing model of progress towards flowering. The research explored the following questions:
1. Can novel crops such as narrow-leafed lupin and lentil be grown under the challenging short growing season typical of the Boreal – Nemoral region? Is there earlier germplasm available of faba bean that could facilitate the expansion of its cultivation? Which cultivars are more stable across years (genotype x environment)? (I, III)
2. How can grain legumes be integrated into the conventional cereal-based rotations? Which cereal is the best pre-crop for legumes? How can a sequence influence the nutrient efficiency and composition of crops? How good is the achieved nutritive quality and protein yield of these crops? (I, II)
3. How can we model the progress towards flowering in grain legumes based on environmental factors? What are the key environmental variables influencing it? (I, III)
28

29
Figure 7. Diagram showing the connections between the 3 publications throughout the plant – cropping system

3 MATERIALS AND METHODS
A general description of the experiments conducted follows below. More details are given in the original publications (I-III). 3.1 Experimental site
All experiments were done at the Viikki Research Farm, University of Helsinki (60.224˚ N,
25.021˚ E). The farm has 42 ha of forest land and 153 ha of arable land. In an average year, 70 ha of arable land are allocated for pasture and grassland for silage production, 2.5 ha are left fallow and the rest is sown to grain crops, mostly cereals (barley, oat and wheat) and turnip rape.
The Viikki Research Farm has a history of cultivation that spans nearly 500 years. The fields are mostly clayey in texture and very near sea level with about <1 – 10 m of elevation, thus having a particularly high ground water table, so artificial drainage has been done for over 160 years (Mokma and Yli-Halla 2000), first with open ditches and for more than 50 years with subsurface drainage pipes.
3.1.1 Weather conditions
The farm is located on the shore of the Gulf of Finland, at the boundary between the Boreal
and Nemoral regions according to the environmental stratification of Europe (Metzger et al. 2013). Finland has a July isotherm of 17°C, an annual average air temperature of 5 °C and a very short growing season that in the north can be less than 120 frost free days and in the south can reach about 185 days (FMI 2016a). During the study period 2009-2012, the length of the growing season in Helsinki was 160, 165, 160 and 174 days respectively.
The region has a characteristically humid continental climate (Peel et al. 2006), but the weather was very variable during the four-year period of this study. By local standards, the growing season in 2010 was characterized by drought and extremely high temperatures during July and August, whereas that of 2012 was extremely wet and total precipitation was 480 mm. In contrast, the growing seasons in 2009 and 2011 were more moderate, the former being mild and wet, while the latter was warm at the start but had high precipitation towards the end (Figure 8).
3.2 Field characteristics
The fields included in this study are part of a catena of Cambisols (Latokartano and Bergman
field), Gleysols (Patoniitty) and some Podzols (other surrounding fields with coarse-textured soil at greater elevation above sea level) according to the FAO - UNESCO system (Mokma et al. 2000). More recently Stagnosols have been considered a more appropriate classification according to the WRB system, instead of Cambisols (Yli-Halla and Nyborg 2014). Among this catena, the most interesting soil group are the Gleysols, which include the acid sulphate soils that represent up to 130,000 ha of arable land in Finland, and often cause several on-site and off-site environmental issues (Šimek et al. 2011), such as acidification and metal loading of waters.
3.2.1 Latokartano field (I, III)
The Latokartano field area is located in the south-western edge of the fenced experimental
area of the University of Helsinki greenhouses and Viikintie. This area is made up of two main fields, Kasarmipelto and Koulupelto (A and B respectively in Figure 9), hereafter referred as Latokartano field (60.226˚ N, 25.018˚ E, 4 m.a.s.l.). The area has a silty clay loam soil texture with an approximate composition of 30–40% silt, 50–60% clay and 10–20% sand, and about 6–12% organic C concentration, conforming to the description of Luvic Stagnosols. This field is
30

31
approximately 4.5 ha and each year part of it is split into plots, for several small field experiments, while the rest is sown to barley so that it is as uniform as possible for the next year’s plot experiments.
Figure 8. Weather summary for the growing seasons 2009-2012 in Helsinki. Data from the Vaisala automatic weather transmitter WXT520 (Vaisala Oy, Finland) located 1-1.5 km from the experimental fields. Day of the year, 125 is the beginning of May and 275 is end of September
Figure 9. Viikki Research Farm fields included in this study. Latokartano field: A) Kasarmipelto B) Koulupelto. The map was built using 4 layers from the paikkatietoikkuna web tool: 1. Topographic map (National land survey open data CC 4.0 license, 2. Depth areas 3. Helsinki guide map and 4. CGI RS – map series (©CGI / MML 2016, publication permit: M. Hämäläinen 16.2.2016, © Liikennevirasto 2016).

3.2.2 Patoniitty field (II)
Patoniitty is located to the west of Latokartano field (60.224˚ N, 25.021˚ E and it is at sea
level), and it is representative of the acid sulphate soils common in the coasts of Finland. Its soil texture is silty clay loam, with approximately 33 % clay, 63 % silt and 4 % sand, and 3.8 % organic C concentration (Lizarazo et al. 2015), conforming to the criteria of Umbric Gleysol according to the WRB system and Sulfic Cryaquepts according to U.S. Soil Taxonomy expressing the acid sulphate characteristics. Due to its low elevation, this poldered field has to be drained extensively by pumping during the growing season, but in the off-season when the drainage pumps do not operate, the water table rises to the soil surface and in some years considerably above it (Figure 10)
This field is 8.6 ha and had been sown to grasses (1996-2001), rapeseed (2002) and cereals (2003 onwards).
Figure 10. Patoniitty field is at sea level and needs extensive drainage. The picture shows the water level risen above the soil surface and frozen, December 2015.
3.2.3 Bergman field (I, III)
The Bergman field is located to the east of the Latokartano field (60.222˚ N, 25.039˚ E, and 6
m.a.s.l.). The soil of this field has about 33–49% clay in the plough layer and 54–68% clay in the subsoil (30–40 cm). The pH in the plough layer was raised by liming to 5.0, while the pH in the subsoil was 3.8–4.8. The organic C concentration in the topsoil was about 6–10%. The area has a sandy clay soil texture in the plough layer, and can be classified as Endogleyic, Umbric Stagnosol (Dystric, Clayic) according to the WRB system. This field is 5 ha and its cultivation history did not include legumes: grasses (1996-1998), rapeseed (1999), wheat (2000-2002), grasses (2003-2005), camelina (Camelina sativa) (2006), wheat (2007-2009) and barley (2010-2011). 3.3 Experimental design
3.3.1 Grain legume germplasm screening (I, III)
A range of grain legume cultivars was screened during the period 2009-2012. The screening was focused on identifying early maturing, nutritive and high yielding cultivars and included 5 cultivars of narrow-leafed lupin, 14 of lentil and 21 of faba bean (complete list in Table 1 in I). All cultivars from each species were screened in the fields of Viikki Research Farm (Figure 9), and experiments were set up as a randomized complete block design per species with four replicates each.
32

33
3.3.2 Screening for earliness (I) Plots of each species were inspected daily and dates were recorded when 10, 50 and 90% of
the plants were flowering and when 10 and 90% of the pods in a plot were mature. For each day of the season, growing degree days (GDD) were calculated using a 5°C base
temperature: ((Daily minimum Temperature + Daily maximum Temperature) / 2) – 5. The accumulated GDD was calculated for each date corresponding to the three flowering and two maturity stages. 3.3.3 Temperature limits and flowering model for faba bean (III)
Considering the interesting patterns seen across years for faba bean phenology in response to varied field conditions (I), a set of experiments was done under controlled conditions to assess the temperature limits and response to photoperiod. Plants of four selected faba bean cultivars were grown in different growth chamber environments, made as a combination of two photoperiods (18 and 11 hours) with six daytime temperatures (18, 20, 22, 24, 26 and 28ºC) using similar settings as those used by Ellis et al. (1988). Flower appearance was checked daily on each of three plants per pot, and dates were recorded to calculate the rate of progress towards flowering (1/f), defined as the inverse of time in days from sowing to flowering. Calculations were done per individual plant, to then calculate the mean of three plants per replicate.
Based on field observations and growth chamber experiments, it was apparent that progress towards flowering in faba bean was governed not only by temperature and photoperiod, but possibly also by solar radiation, water deficit and cultivar. Progress towards flowering rates (1/f) were calculated using flowering dates of 20 cultivars from field screening trials during 2009-2012. The model was tested for early flowering (EF), middle flowering (MF) and full flowering (FF) (10, 50 and 90% plants with open flowers in a plot, respectively).
The former model of progress to flowering in faba bean, developed in the 1980s, focused on the effect of temperature (T) and photoperiod (P) where: 1/f = a + bT + cP (Evans 1959; Roberts and Summerfield 1987; Ellis et al. 1988; Ellis et al. 1988a; Ellis et al. 1988b; Ellis et al. 1990). The effect of adding two new variables was then tested: PAR (or sunshine duration, for cases when PAR is unavailable) and Sielianinow index (K, a hydrothermal index), which were fitted for each flowering stage forming a new ‘basic model’ (Eq.1). The model was expanded by adding a cultivar-variable specific for 'Kontu' and 'Witkiem Manita' (which behaved differently from the rest) and this formed an ‘extended model’ (Eq.2).
Hence the 'basic' model was
1/f ~ Photoperiod + Temperature + PAR + K index + Ɛ (Eq.1) and the 'extended' model was 1/f ~ Photoperiod + Temperature + PAR + K index + Kontu + Witkiem Manita + Ɛ (Eq.2)
The rates of progress towards flowering and the covariates were fitted using a linear model,
and the model fit was tested using the root square mean errors (RSME) and a permutation inference test, done following the Freedman-Lane method (Freedman and Lane 1983; Winkler et al. 2014) (III).

3.3.4 Screening for nutritive quality (I) To assess the feasibility of locally grown protein from grain legume crop production and the
stability of their nutritive characteristics in a Boreal – Nemoral climate, every year from 2009 to 2012 at maturity, a sample was collected from 1 m2 of each plot, then seeds were threshed, dried at 40°C and milled for analysis.
Nitrogen concentration was determined for each growing season, using the Dumas combustion method and a conversion factor of 6.25 for estimating protein concentration. Further analysis was done for cultivars that had early maturity dates, the main analysis including measured concentrations of starch, fiber, soluble sugars, oil, and the amino acid and fatty acid composition (list of methods in Table 2, and detailed description of methods in publication I).
3.3.5 Crop rotation (II)
The field trial used a randomized complete block design, and focused on the interaction of two grain legumes, NL lupin and faba bean, with several other spring crops that are conventionally grown in Boreal – Nemoral climate cropping systems, namely barley (Hordeum vulgare L., cv. ‘Streif’), oat (Avena sativa L., cv. ‘Ivory’), rye (Secale cereale L., cv. ‘Juuso’), wheat (Triticum aestivum L. emend Thell., cv. ‘Amaretto’), turnip rape (Brassica rapa L. ssp. oleifera (DC.) Metzg., cv. ‘Apollo’), and field pea (Pisum sativum L., cv. ‘Rocket’). The crops listed were cultivated in the first year of the rotation (2011), then in the following year these plots were split in half and each half was cultivated with faba bean or narrow-leafed lupin. In the third year of the rotation, each half was split again and sown with the same crops grown in the first year but excluding field pea (Figure 11). Only the results of the first two years are presented because the main interest was the effect of the pre-crop on the growth and yield of the following legume crop.
Plant and soil analyses were performed to assess the performance of each sequence. The key stage of the rotation was year 2012, where 50 mature plants were taken from 10 random places on each plot, then shoots and a seed fraction were sorted for drying, milling and digesting prior to chemical analyses by inductively coupled plasma-optical emission spectrometry (ICP-OES) that was used to determine the content of B, Ca, Cu, Fe, K, Mg, Mn, P, S, Si and Zn in each plant fraction. Based on the ICP results, a series of indexes (Table 3) were calculated for each mineral element, and these were used as indicators to identify the best cereal pre-crop for the two grain legumes (II).
3.3.6 Sampling, measurements and indexes
A brief list of the methods used in each publication is given in Table 2. To estimate the effect of weather conditions on moisture levels surrounding the legume crops
and its effect on crop phenology, the Sielianinow hydrothermal index was calculated at different time intervals (I, III; Table 3). For evaluating the pre-crop effect of cereals on the nutrient composition and utilization efficiency of grain legumes, 5 indexes were used and are summarized in Table 3 (I, II, III).
3.3.7 Plant density
Although the breeders of Haags Blaue (Saatzucht Steinach) recommend using a plant density of 125 plants/m2, in our farm trials this density required intensive management to control weeds. In order to identify the optimum plant density for cultivation in the Boreal – Nemoral region, a trial was established using a randomized complete block design with 4 replicates. Five different plant densities were tested: 100, 125, 150, 175 and 200 plants /m2, and phenology, yield, percentage of
34

35
canopy cover (measured as leaf area index), height of first pod and protein concentration were measured.
Figure 11. Crop rotation trial in Patoniitty (2011-2012). The first year of the rotation included the following crops: wheat (1), barley (2), rye (3), oat (4), turnip rape (5), faba bean (6), narrow-leafed lupin (7) and field pea (8). Field picture credit: AISA Eagle II, flight campaign 2011. Zou, X & Mõttus, M. Department of Geosciences and Geography, University of Helsinki.

Table 2. List of measurements and methods used during the research.
Measurement Description Publication Date and GDD to flowering Number of days until 10, 50 and 90% of the plants in a
plot started flowering and finished flowering. I
Date and GDD to maturity Number of days to until 10 and 90% of the plants in a plot reached maturity.
I
Progress towards flowering Number of days to first flower open. III Seed weight Weight of 100 or 500 mature dried seeds from 1 m2 hand
harvested. I, II
Seed N concentration Dumas combustion method in a Vario Max CN analyzer (Elementar, Hanau, Germany)
I, II
Starch content Megazyme total starch (K-TSTA 1107) kit I Total fiber content Megazyme total fiber (K-TDFR 1205) kit I Soluble sugars content Anthrone reagent method I Oil content Petroleum ether extraction using a Soxtec™ 2055 system
(AOAC 2003.06) I
Moisture and Ash content Overnight drying at 105˚ C and incineration in a muffle furnace at 600˚ C
I, II
Amino acid analysis UPLC Amino Acid Analysis Solution System (Waters, Milford, MA, USA) / Commission Directive 98/64/EC
I
Digestible indispensable amino acid score (DIAAS)
Calculated according to FAO 2011 as follows: 1) the Digestible IAA reference ratio= [(mg of digestible dietary indispensable amino acid in 1 g of the dietary protein)/(mg of the same dietary indispensable amino acid in 1 g of the reference protein)], where the reference protein refers to standard values from the scoring pattern established by FAO (2011) in its ‘Dietary protein quality evaluation in human nutrition’. 2) DIAAS % = 100 x lowest value of the Digestible IAA reference ratio for a given AA.
-
Fatty acid analysis Accelerated solvent extraction using acetone. Then methylated and analyzed by gas chromatography (GC)
I
Seed volume and seed coat thickness
Dimensions and seed coat thickness of 100 seeds of 3 cultivars were measured with a precision micrometer. This was only done for faba bean because of a) the higher concentration of tannins in their seed coat makes this information valuable for purposes of de-hulling seeds, and b) the wider variation of seed size and shape, than in the other two crops.
I
Leaf area index (LAI) SunScan SS1 ceptometer bar (Delta-T Devices, Cambridge, UK) canopy analysis system
II, III
Chlorophyll content Chlorophyll meter (SPAD-502, Minolta, Japan) II, III B, Ca, Cu, Fe, K, Mg, Mn, P, S, Si, and Zn content
Plant material was milled, digested with nitric acid and then mineral element content was measured using Inductively coupled plasma—optical emission spectrometry (ICP–OES) using an iCAP 6200 (Thermo Fisher Scientific Inc., Cambridge, UK)
II
36
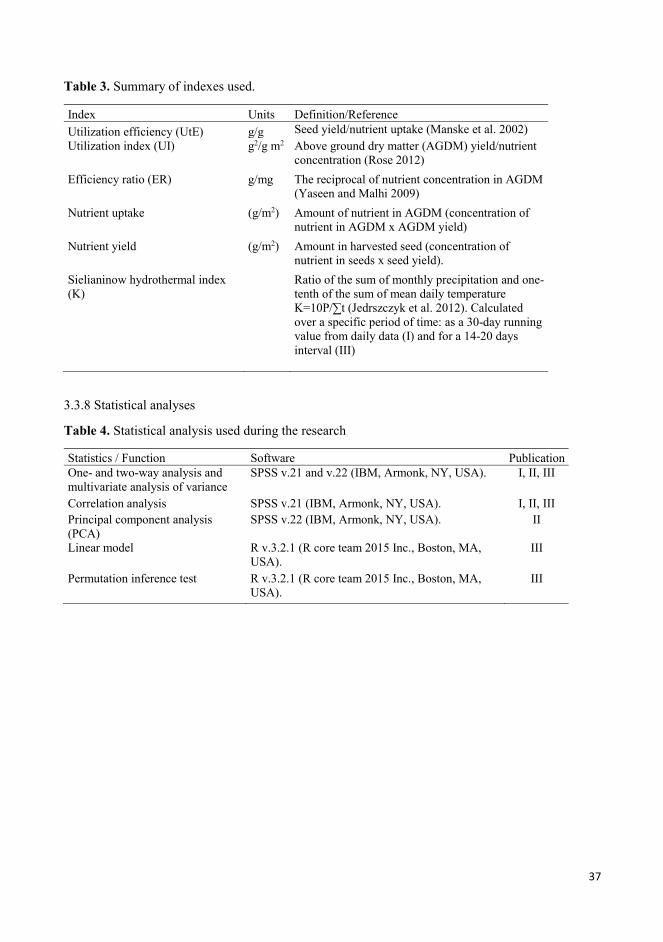
37
Table 3. Summary of indexes used.
Index Units Definition/Reference Utilization efficiency (UtE) g/g Seed yield/nutrient uptake (Manske et al. 2002) Utilization index (UI) g2/g m2 Above ground dry matter (AGDM) yield/nutrient
concentration (Rose 2012) Efficiency ratio (ER) g/mg The reciprocal of nutrient concentration in AGDM
(Yaseen and Malhi 2009) Nutrient uptake (g/m2) Amount of nutrient in AGDM (concentration of
nutrient in AGDM x AGDM yield) Nutrient yield (g/m2) Amount in harvested seed (concentration of
nutrient in seeds x seed yield). Sielianinow hydrothermal index (K)
Ratio of the sum of monthly precipitation and one-tenth of the sum of mean daily temperature K=10P/∑t (Jedrszczyk et al. 2012). Calculated over a specific period of time: as a 30-day running value from daily data (I) and for a 14-20 days interval (III)
3.3.8 Statistical analyses
Table 4. Statistical analysis used during the research
Statistics / Function Software Publication One- and two-way analysis and multivariate analysis of variance
SPSS v.21 and v.22 (IBM, Armonk, NY, USA). I, II, III
Correlation analysis SPSS v.21 (IBM, Armonk, NY, USA). I, II, III Principal component analysis (PCA)
SPSS v.22 (IBM, Armonk, NY, USA). II
Linear model R v.3.2.1 (R core team 2015 Inc., Boston, MA, USA).
III
Permutation inference test R v.3.2.1 (R core team 2015 Inc., Boston, MA, USA).
III

4 RESULTS AND DISCUSSION 4.1 Importance and feasibility of protein crop cultivation
4.1.1 Protein production in the Boreal – Nemoral region: potential for local protein yield
As discussed in section 1.1.3, the dependence on vegetable protein imports and N inorganic
fertilizers is a major issue in the Boreal – Nemoral region, as it is throughout Europe, thus local cultivation of crops with less N fertilizer requirements and that could be a good source of vegetable protein are needed.
The protein concentration achieved by NL lupin in these experiments was 33% on average, while that of faba bean was 32%. On the other hand, the protein concentration of lentil was much lower, being 29% on average (Table 4 in I). Overall the results show that the 3 suggested grain legumes, have a 5-18% higher protein concentration than that of turnip rape (Figure 12). There was a variation of 2% in protein concentration within cultivar across years for all 3 legumes (Table 5), as commonly found in grain legumes; for example, Stoddard et al (1993) reported that protein concentration had a variation of 4% in peas and of 3% in lentil depending on location and year.
The results also show the achievability of good protein yields at the local level (Table 5), particularly with faba bean cultivars that had a consistent protein yield level between 1130 and 1490 kg/ha, and NL lupin with less stable levels between 570 and 1250 kg/ha. Thus it is feasible to expand the protein production in the Boreal – Nemoral region not only with the conventional field pea and faba bean, but also with the locally novel NL lupin if suitable management practices are devised (I, II).
Lentil protein yield levels were significantly lower than those of faba bean or NL lupin (Table 5), so its potential use as protein source for feed is very limited, but that for human food is promising.
Since the protein yield stability across years was very low for NL lupin cultivars and rather low for the other grain legumes, improving protein self-sufficiency is not only an issue of availability and allocation of arable land, but also depends critically on both breeding and management solutions.
Considering the low protein yield and its dependence on N fertilizer, the current national focus on rapeseed as a protein crop in the region should be reduced, and the cultivation of faba bean and NL lupin should be promoted.
Initially, during the germplasm screening trials, in both faba bean and lentil there was a negative correlation between protein concentration and yield (Figure 6D and 7D in I), while NL lupin had a positive correlation (Figure 8D in I), although none was statistically significant. During the crop rotation trial it was found that there was no detectable correlation between the two variables in either crop (Figure 1 in II). The negative correlation between yield and protein concentration is widespread in cereals (Penning de Vries et al. 1974) but not in grain legumes. In pea and lentil in 27 environments in Australia, the correlation was usually non-significant and in 4 individual sites it was significant and negative (Stoddard et al. 1993) as also reported in chickpea (Gaur et al. 2016). In soybean, there was no significant correlation in one of 3 years, but a significant and positive correlation in the other two (Anthony et al. 2012). The results shown here on faba bean and lentil are consistent with the lack of correlation between protein concentration and yield found for these crops in the literature; whereas for narrow-leafed lupin there might be an exception, but more cultivars should be evaluated before reaching a conclusion on this.
38

39
Figure 12. Nutritive composition of grain legume crops tested in Finland (faba bean, lentil and NL lupin grown in Helsinki 2009-2012) and of other crops used as source of vegetable protein: soybean (FAO 1992; Stevenson et al. 2006) turnip rape (Musgrave et al. 2008; Henriksen et al. 2009) and field pea (Wang et al. 2008; Wang et al. 2010).

This fundamental difference between families of crops is associated with the BNF capacity of legumes that ensures that the plant is seldom N-limited, unlike a cereal. Part of the correlation in cereals is due to the greater sensitivity of starch synthesis than protein synthesis to water deficit and heat (Jenner et al. 1991). Thus, in a drought, the deposition of the major component of the grain (starch) is affected more than the minor component (protein), so protein concentration increases while yield is reduced.
Genotypic variability in protein concentration stability was found with cv. ‘SSNS-1’ being more stable than ‘Kontu’ (Table 5), and this trait is desirable by processors seeking consistent quality in raw materials.
Since the protein yield stability across years was very low for NL lupin cultivars and rather low for the other grain legumes, improving protein self-sufficiency is not only an issue of
It is evident that the Boreal – Nemoral region needs to incorporate more grain legumes in its cropping systems, not only to have a local source of protein for feed production, but also to comply with the local demand of grain legumes for human food. Moreover, the inclusion of grain legumes can help to improve the fitness of the cropping system by improvements in soil fertility, providing a break in disease cycles, and improving the quality of water systems by reducing nutrient leaching to the Baltic Sea.
Currently the strong impact of agriculture and food systems on the environment is mainly caused by the intensive cereal cropping systems that rely heavily on fertilizers (discussed further in section 4.1.3) and livestock farming systems that often rely on feed made with imported soybean meal. Both systems generate a significant amount of greenhouse gas emissions, need large amounts of fossil energy and have a considerably bigger environmental footprint, compared to that of legume-supported cropping systems (Fader et al. 2011; Reckling et al. 2016a; Reckling et al. 2016b). Grain legumes are particularly good components that minimize adverse environmental impacts not only of the cropping and farming systems, but also that of the human diets (e.g. nitrogen footprint, water footprint, land use footprint) (Fader et al. 2011; Cutforth et al. 2013; Leach et al. 2016; Gephart et al. 2016). Thus the local production and consumption of grain legumes could help to mitigate some of the negative environmental impacts of agriculture and food systems.
Achieving protein self-sufficiency in the Boreal – Nemoral region will require breeding efforts for earliness combined with protein stability (I, III), increase of subsidies and better planning of arable land allocation. This program needs to be implemented promptly to lessen the environmental impact that Boreal – Nemoral food and agricultural systems have through the imports of foreign vegetable protein and fertilizer use.
The correlation between seed yield and GDD to first flower was positive in NL lupin and faba bean, being significant in the former but not significant in the latter crop, while it was negative and non-significant in lentil (Figures 6A, 7A, 8A in I). Thus, earliness often implied a yield penalty in NL lupin (Figure 8 in I) and faba bean (Figure 6 in I and Table 3 in III). Nevertheless, the yields achieved by the earliest cultivars used in the studies were quite acceptable. For example, during the period 2009-2012 in the germplasm screening trials, the earliest NL lupin (cv. ‘Haags blaue’) had an average yield of 2.8 t/ha which is comparable to those achieved in Australia (1.5 t/ha) and Germany (2.7 t/ha) (Beyer et al. 2015). During the same period faba bean cv. ‘Kontu’ had an average yield of 4.4 t/ha which is certainly low when compared to the highest yielding cultivars available which can yield up to 6.6 t/ha, but is acceptable when compared to the average yields achieved in Estonia (3 t/ha), UK (3.9 t/ha), Denmark (4 t/ha) and Germany (4.5 t/ha) (Flores et al. 2013). Hence, although the yield penalty is not ideal, the performance of the early cultivars was acceptable, and earliness continues to be a top priority because of the need to reach maturity and harvest the crops before the typically harsh end of the short growing seasons in the Boreal – Nemoral region.
Earliness implies a short crop cycle and inevitably means a reduction in the time available to accumulate biomass, and thus in the feasibility to reach higher yields (Fischer et al. 2012, Tardieu et
40
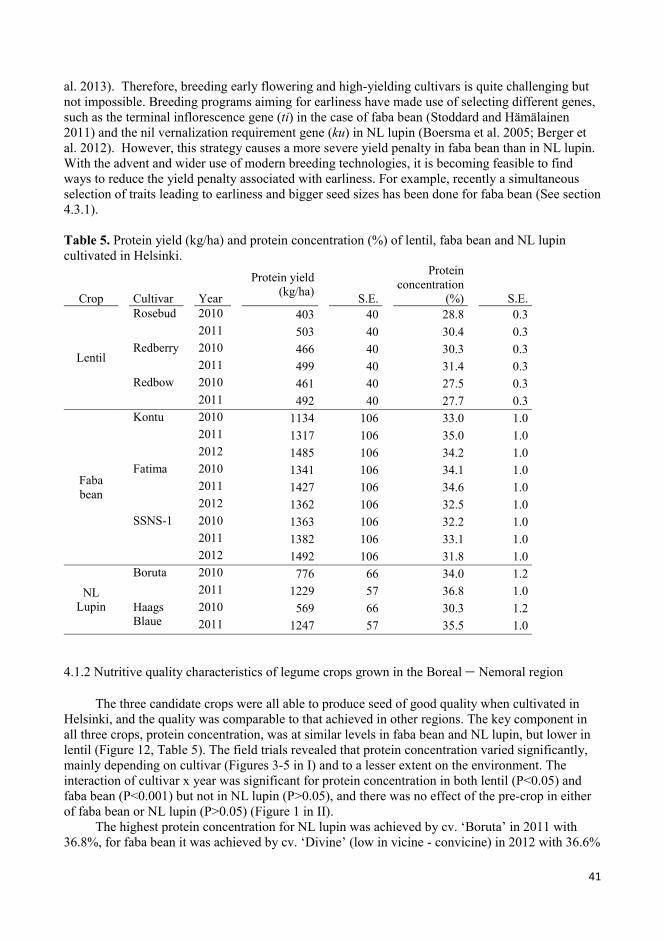
41
al. 2013). Therefore, breeding early flowering and high-yielding cultivars is quite challenging but not impossible. Breeding programs aiming for earliness have made use of selecting different genes, such as the terminal inflorescence gene (ti) in the case of faba bean (Stoddard and Hämälainen 2011) and the nil vernalization requirement gene (ku) in NL lupin (Boersma et al. 2005; Berger et al. 2012). However, this strategy causes a more severe yield penalty in faba bean than in NL lupin. With the advent and wider use of modern breeding technologies, it is becoming feasible to find ways to reduce the yield penalty associated with earliness. For example, recently a simultaneous selection of traits leading to earliness and bigger seed sizes has been done for faba bean (See section 4.3.1). Table 5. Protein yield (kg/ha) and protein concentration (%) of lentil, faba bean and NL lupin cultivated in Helsinki.
Crop Cultivar Year
Protein yield (kg/ha)
S.E.
Protein concentration
(%) S.E.
Lentil
Rosebud
2010
403 40 28.8 0.3
2011
503 40 30.4 0.3
Redberry
2010
466 40 30.3 0.3
2011
499 40 31.4 0.3
Redbow
2010
461 40 27.5 0.3 2011 492 40 27.7 0.3
Faba bean
Kontu 2010 1134 106 33.0 1.0
2011
1317 106 35.0 1.0
2012
1485 106 34.2 1.0 Fatima
2010
1341 106 34.1 1.0
2011
1427 106 34.6 1.0
2012
1362 106 32.5 1.0
SSNS-1
2010
1363 106 32.2 1.0
2011
1382 106 33.1 1.0
2012 1492 106 31.8 1.0
NL Lupin
Boruta 2010 776 66 34.0 1.2
2011
1229 57 36.8 1.0
Haags Blaue
2010
569 66 30.3 1.2
2011 1247 57 35.5 1.0
4.1.2 Nutritive quality characteristics of legume crops grown in the Boreal – Nemoral region
The three candidate crops were all able to produce seed of good quality when cultivated in Helsinki, and the quality was comparable to that achieved in other regions. The key component in all three crops, protein concentration, was at similar levels in faba bean and NL lupin, but lower in lentil (Figure 12, Table 5). The field trials revealed that protein concentration varied significantly, mainly depending on cultivar (Figures 3-5 in I) and to a lesser extent on the environment. The interaction of cultivar x year was significant for protein concentration in both lentil (P<0.05) and faba bean (P<0.001) but not in NL lupin (P>0.05), and there was no effect of the pre-crop in either of faba bean or NL lupin (P>0.05) (Figure 1 in II).
The highest protein concentration for NL lupin was achieved by cv. ‘Boruta’ in 2011 with 36.8%, for faba bean it was achieved by cv. ‘Divine’ (low in vicine - convicine) in 2012 with 36.6%

(I), which was much higher than the 29.5% reported by the French breeder (Agriobtentions 2016), while for lentil it was achieved by cv. ‘Black’ in 2010 with 32.6%. These values serve as an example of the maximum potential in grain protein concentration that each crop could reach, if management practices are improved and the environmental conditions are favorable.
However, these cultivars were not among the earliest in maturity. Among the earliest germplasm, the protein concentrations were acceptable, and high values were found in NL lupin cv. ‘Haags Blaue’ with 35.5%, faba bean cv. ‘Kontu’ with 35% and lentil cv. ‘Rosebud’ with 30%, all in 2011 (I). The coincidence of highest protein concentrations with the warmest year, is due to the well-known fact that at higher temperatures protein decreases less than starch deposition (Denyer and Hylton 1994; Patrick and Stoddard 2010; Zhen-zhen et al. 2015), as discussed earlier.
The grain protein concentrations in lentil and NL lupin were within normal ranges achieved at other latitudes, while that of faba bean was about 3% higher than at lower latitudes (32% cf 29%), and in all 3 crops it was possible to see how inter-annual variations in temperature and rainfall influenced the grain composition by using an agro-climatic index (I, II). The water deficit indicator ‘Sielianinow index’ (K) has been used in Poland (Jedrszczyk et al. 2012) to identify critical weather events such as drought and waterlogging across seasons (I, III), and can serve as an indicator to detect critical water deficits at specific developmental stages, and thus help to explain changes in development and grain composition (I, III).
In the present field experiments (particularly during 2010 and 2011), there were noticeable changes in grain composition mainly in protein and starch (I), associated with drought periods and high temperatures at critical developmental stages. Frequent dry periods (K index being lower than 1.3) experienced during the grain filling in the germplasm screening trials (Figure 2 in I), showed that severe drought stress periods (as that experienced in 2010) and even milder drought spells below optimal moisture, can affect starch deposition much more than protein deposition, contributing to the fluctuation on protein and starch levels across years (Bishop et al. 2016; Farooq et al. 2016). For example, in faba bean starch concentration was lower (~36%) in 2009 when the K index was often higher than 1.3 and exceeding 1.6 during the seed set and seed filling period, while the starch concentration was higher (~41.6%) in 2010 when the K index was often <1.3 during the same period. In contrast, protein concentration was less affected by drought having a variation of only ±1% depending on cultivar and being influenced by changes in the K index during the different growing seasons.
Moreover, heat and drought stresses are particularly damaging at sensitive phases such as during flower development, anthesis, gametophyte development and pollen production, consequently affecting yield levels. For example, faba bean yields were lower in 2010 and slightly higher in 2011, both years during which K index was often below optimal levels (Table 3 in I) and yields were much higher in the wetter year 2012 when K index was often >1.3 and even >2 (Table 4 in II).
Faba bean is able to form associations with mycorrhizas, which contribute to the uptake of metallic nutrients (II) and in a study by El Fiel et al. (2002) have been shown to increase the albumin and glutelin protein fractions when mycorrhizal infection was used in combination with a sulphur fertilizer. Among the grain legumes, faba bean seeds had the highest concentrations of Cu, Fe, K, P and Zn, while NL lupin had highest concentrations of B, Ca, Mg, Mn, S, Si in the crop rotation trial (II). The most notable difference in nutrient yield between these crops was in P and S yield. NL lupin had particularly high S yield, in accordance with its higher content of sulphur-containing amino acids, while faba bean had significantly higher P concentrations and yield, in agreement with its higher levels of phytic acid (I, II).
There were differences between cultivars in cysteine content: cv. ‘Kontu’ had the lowest and cv. ‘Aurora’ had the highest contents among faba bean germplasm, and cv. ‘Boruta’ the lowest and cv. ‘Sonet’ the highest among NL lupin germplasm (data not shown). Considering the consequences of AA composition for feed and food protein meal quality and digestibility, the AA composition per
42

43
cultivar was evaluated using the digestible indispensable amino acid score (DIAAS) recommended by FAO (2011). The DIAAS score has replaced the protein digestibility corrected amino acid score (PDCAAS) because the former makes use of ileal digestibility for each individual amino acid, while the latter uses a single value for crude protein digestibility, thus DIAAS is better in reflecting how much amino acid is available for absorption.
For the grain legume seeds from the germplasm screening trials, it was found that the sulphur containing amino acids have the lowest DIAAS score (Table 6), which was expected since legumes are known to have low concentrations of cysteine and methionine, but the DIAAS score of cv. ‘Kontu’ was exceptionally low. It is clear that not only the choice of legume crop, but also the choice of cultivar would make a considerable difference in the protein meal digestibility, mainly due to differences in lysine, cysteine and tryptophan content.
Table 6. DIAAS value for flour of faba bean cv. ‘Kontu’ and ‘Aurora’, and NL lupin cv. ‘Boruta’ and ‘Haags Blaue’. Flour was made from seed harvested on 2010. DIAAS was calculated using standardized ileal digestibility of indispensable amino acids (IAA), because although true ileal digestibility is preferable, values are not available.
Cultivar Digestible IAA reference ratio (for adult age group*) (mg/g protein) DIAAS %
Cys+Met His Ile Leu Lys Thr Trp Tyr Val
Kontu 0.24 1.11 0.94 0.84 0.95 0.93 0.65 N/A 0.74 24 SAA Aurora 0.53 1.15 0.91 0.88 0.95 0.98 0.83 N/A 0.71 53 SAA Boruta 0.57 1.52 1.09 0.94 0.81 1.14 1.14 N/A 0.76 57 SAA H.blaue 0.67 1.33 1.03 0.90 0.80 1.11 1.26 N/A 0.74 67 SAA
*calculated based on the reference protein requirement established by FAO (2011) for the third age group: ‘older child, adolescent, adult’.
In general, faba bean and NL lupin had AA profiles similar to those of crops grown elsewhere (Schumacher et al. 2011), and had enough content of essential AA to match the requirements for human and livestock nutrition (I). Several studies have tested their suitability as ingredients for enriching the nutritional value of food and feed products. For example, faba bean has recently been shown to improve the protein quality of pasta - although having a negative effects on its texture and sensorial qualities (Giménez et al. 2016) - and also successfully replaced soybean meal in diets for growing pigs, in a study where 3 experimental diets were made using 300 g/kg of different faba bean cultivars (White et al. 2015).
Among the interesting nutraceutical components present in NL lupin, it was estimated that the Helsinki-grown material had an average folate content of 3990 ng/g (DW basis) while Mikkeli-grown material had 3420 ng/g (DW basis)(Lizarazo et al. 2011). In contrast, whole-grain wheat flour had barely 500 ng/g folate (Kariluoto et al. 2010) while barley fractions were estimated to have between 563-1710 ng/g depending on the fraction (Edelmann et al. 2013). Thus outstanding levels of folate in NL lupin give it strong potential as an additive for enriching food products.
Stability in protein concentration and protein yields of crops is difficult to obtain as these traits are very responsive to the environment, for example protein yield of barley and protein concentration of oat in Finland have been shown to be significantly influenced by the genotype x environment interaction (Peltonen-Sainio and Peltonen 1993; Peltonen et al. 2011), and the present results indicated that this interaction was significant for both traits in faba bean and lentil.
Since faba bean had the highest biomass and seed yield, in turn it produced the highest protein yield (Figures 3-5 in I) compared to the other two legume crops. In addition, considering the differences in nitrogen metabolism among the crops and the abilities of some faba bean cultivars to better tolerate drought and to outcompete weeds, faba bean is potentially the more reliable source for high protein yield in the Boreal-Nemoral region. Nevertheless, due to the apparent (from small

number of cultivars tested) lower plasticity in both protein concentration and protein yield shown by NL lupin, and considering its unique nutritional attributes (e.g. folate content and higher DIAAS score), and adaptation to different soil types, NL lupin is a good alternative for protein yield.
4.2 Crop diversity and nutrient availability: grain legumes in rotation
4.2.1 Designing sustainable crop rotations: The pre-crop effect
In the present crop rotation experiment NL lupin produced equally high seed yields after turnip rape and oat, while faba bean gave higher yields after turnip rape and then after barley (Table 4 in II). Although the pre-crop effect was not significant in growth parameters (seed size and yield, harvest index, biomass and LAI), it was detectable and significant in the nutrient uptake (shoot composition), nutrient yield (seed composition) and in their nutrient utilization indexes, with barley giving the most favorable effect followed by oat (II).
Overall, there was a significant pre-crop effect in 19 of the 88 measurements on faba bean, while for NL lupin there were 7 (Figure 3 and 4 in II). Among these 26 measures, barley was the best pre-crop in 9 variables, and oat in 5. On the other hand, rye gave the lowest values in 8 variables, showing that any advantage was species-specific and not general across cereals (II). Both grain legume crops had a significantly positive pre-crop effect from barley and oat, the former being better for NL lupin and the latter for faba bean as evidenced by the significance on different efficiency indexes (Figure 3 and 4 in II). Specifically, for NL lupin the barley pre-crop improved Mn yield, Mn HI, and Mg, Mn and Zn concentrations in seed, while for faba bean it improved only the P yield and S concentration in seed. In addition, for faba bean the oat pre-crop improved CaUI, MnER, MnUI, MgER, MgUI and SiUI, while for NL lupin it improved only the Si uptake. The improvement of MgER and MgUI in faba bean by the oat pre-crop is of particular interest since it is known that the enzyme nitrogenase of Rhizobium leguminosarum (responsible for BNF) requires a minimum of 16 molecules of Mg-ATP to function, and so a more efficient MgER and MgUI have the potential to increase BNF (Kiss et al. 2004; Hood et al. 2015).
Barley had a particularly significant effect on the yield of P and in the seed S concentration of faba bean (Figure 3 in II), and on the Mn yield and seed concentration in NL lupin (Figure 4 in II), which could be used to improve the already rich grain composition of both legumes (I). Moreover, the fact that an oat pre-crop was able to significantly improve the MnER and MnUI is particularly relevant in this crop rotation, because the Mn field level in the farm was low (II), and because adequate levels of micronutrient in seed and in fertilizer have been reported to be important for achieving maximum seed yield production and reduction of split seed in NL lupin (Brennan and Longnecker 2001).
The suitability of cereals as a pre-crop for grain legumes was expected, since they do not have pathogens in common, and have different water and nutrient requirements, factors benefiting the following crop (Angus et al. 2015). What is more interesting and unusual, is the finding that a particular cereal seems to be a better pre-crop for a certain legume.
These results show that there are diverse dynamics between different crop species in rotation, and the potential of choosing specific pre-crops to design crop rotation sequences that could potentially improve the grain legume crop, and facilitate their integration into the traditional cereal-based cropping systems of the Boreal – Nemoral region (I, II).
Since the pre-crop effect was detected in both the shoot and seed nutrient composition and efficiency indexes, it may affect not only nutrient uptake but also, by extension, other vital processes such as cell signaling, chlorophyll production, photosynthesis, growth, and resistance to abiotic stresses, so it could improve the overall fitness of the whole plant and thus that of the cropping system (II). Nevertheless, the mechanisms responsible for the cereal pre-crop effect on legumes should be investigated further. For example, studies about exudate release and changes in
44

45
soil microbiology and chemistry during the pre-crop phase are needed. Furthermore, having detected evidence of a pre-crop effect from barley and oat, such evidence could be examined further by testing the crop sequences under different soil nutrient levels, which could make the significance of a pre-crop effect become more evident.
Overall, it is clear that diverse crop sequences are more productive than continued monocrops. Pre-crops not only are capable of influencing yield levels but more importantly, can improve the nutrient cycling efficiency (II), balance N budgets, manage water availability, soil fertility and break disease cycles. Rotations including legumes have remarkable extra advantages such as reductions in economic optimum nitrogen rates (O’Donovan et al. 2014; Luce et al. 2015) and nitrous oxide emissions (Freibauer and Kaltschmitt 2003; Schwenke et al. 2015) due to reductions in fertilizer input. Although sustainable crop rotations involving sequences of grain legumes, oilseeds, cereals and grasses are potentially available for the Boreal – Nemoral region, the main obstacles for their implementation (among those already mentioned in section 4.1.1) include the scarcity of early germplasm (I,III), and the belief among farmers that growing grain legumes is less profitable than that of growing cereals (Von Richthofen and GL-Pro partners. 2006; Malik et al. 2015; Zander et al. 2016). Nevertheless, in western Canada it has been shown that the inclusion in rotation of both dry pea and lentil to wheat-based cropping systems improved farm level returns due to their ability to reduce resource use by 17 to 25% and to reduce impact on ecosystem quality by 1 to 24% (MacWilliam et al. 2014).
Considering the superiority in protein concentration and BNF that faba bean and NL lupin have and their high ranking as pre-crops that increase cereal yields in many regions (Angus et al. 2015) as well as the set of environmental services they provide, it would be expected that they could lead to some farm economic benefits. However, grain prices are low, reductions of N2O release or NO-
3 leaching bring no financial benefit to the farmer, the reductions in N fertilizer use may be underestimated, yield stability is questioned, weed and disease control is challenging, and other environmental indicators are difficult to measure, so the determining balance of economic returns is not straightforward (Reckling et al. 2015; Reckling et al. 2016a). Several trade-offs between economic and environmental benefits need to be considered for the successful integration of legumes in cropping systems (Reckling et al. 2016a). 4.2.2 Soil health: nutrient availability, uptake and nutrient trade
In contrast to most studies, the crop rotation field experiment in this research was performed
under non-limiting mineral element supply. There was plenty of mineral N and P in the soil, while Mn concentration was low, as is observed elsewhere in the Viikki Research Farm (II). Faba bean was superior to NL lupin in the uptake and accumulation of metallic nutrients (Figure 2 in II), thus having great potential to improve nutrient cycling in crop rotations, and could be used for biofortification of metallic nutrients and even be involved in some types and stages of phytoremediation processes (II).
Although faba bean is not considered to be either a hyperaccumulator or a metallophyte (Austruy et al. 2013), it is considered a model plant in the field of eco-toxicity and genotoxicity studies (Rosa et al. 2003; Foltête et al. 2012; Silva et al. 2013) and has potential to be integrated in some stages of phytoremediation (Poonam et al. 2014). The growth rate of faba bean was reported not to be affected in cadmium-spiked soils during the first two months (Foltête et al. 2012), and has been used as a medium for conducting rhizoremediation of soils that were moderately contaminated with copper. Thus, it seems feasible that faba bean and other legumes as suggested by Safronova et al. (2011) would be useful as an alternative for revegetation and phytostabilization processes as part of phytoremediation of some polluted soils.

A set of nutrient efficiency indexes (UtE, UI and ER) was chosen to identify which crops have outstanding uptake of certain minerals and to determine their impact on soil nutrient status and their potential as pre-crop for legumes. Each of the chosen indexes measures the nutrient uptake in proportion to different plant parts: UI relates crop biomass yield to nutrient concentration in the shoots (giving stable results even at different levels of biomass production), UtE measures seed yield produced per gram of nutrient taken up, and ER measures the biomass yield per gram of nutrient taken up (Yaseen and Malhi 2009; Rose 2012; Bovill et al. 2013).
The two studied legume crops not only have marked differences in yield levels but also in their mineral element stoichiometry (Sadras 2005). Consequently, NL lupin generally produced more DM per unit of Cu, K and P, while faba bean produced more DM per unit of Mg, Mn and S. This resulted in some indexes being higher in NL lupin even if yields were not (Table 5 in II). Indeed, high utilization indexes are often equated to high yield, but exceptions may occur when comparing across species or even accessions (Rose and Wissuwa 2012).
Principal component analysis and analysis of variance results indicated that UI was the most informative among the 3 indexes, for further use. The high KMO values of UI showed that it was the best in separating clusters, and it was also the index that detected the most differences in pre-crop effects (Table 7 and Figure 3 and 6 in II).
In addition to detecting differences in uptake between crops, the nutrient efficiency indexes can be used to improve the understanding of internal and external nutrient requirements of crops in rotation, which could allow to improve the crop choice for rotations and precision of fertilizer dosages.
Barley and turnip rape are able to host endophytic rhizobia in their roots, and the former is also able to form mycorrhizal associations, both types of symbiosis improving the crop acquisition of nutrients (Lupwayi et al. 2004). Similarly, faba bean forms associations with mycorrhizal fungi (Duc et al. 1989; Kape et al. 1992; Kaschuk et al. 2009; Kaschuk et al. 2010) and releases a substantial amount of malate and citrate from its roots (Li et al. 2007; Chen et al. 2013). Although NL lupin is not able to form mycorrhizal associations (Avio et al. 1990; Oba et al. 2015), it can release a varied set of carboxylates, including tartrate, which eases the access of the crop to different nutrient pools (Pearse et al. 2006). Both mycorrhization and carboxylate release of the previous crop are key mechanisms able to influence the mycorrhiza formation (Sorensen, Larsen, and Jakobsen 2005) and nutrient uptake by the following crop (II), but more research on crop sequences and management practices, is needed in order to make full use of the pre-crop effect. Although several of the pre-crops included in this study are able to form symbioses with mycorrhiza, the colonization of roots by these symbionts is known to be reduced when the soil P supply is sufficient, hence the contribution of mycorrhiza to the pre-crop effect found here is likely to be minor. Studies by Jia et al. (2004) and Abd-Alla et al. (2014) have reported that mycorrhiza and rhizobia have a positive synergistic association in faba bean, thus for future studies it would be interesting to test specifically the pre-crop effect of crops able to form mycorrhiza on the BNF rates of the following legume crop.
Crops have significant differences in their mineral nutrient composition, so they have different nutrient supply requirements (I, II). For example, wheat has significantly higher N and P requirements for edible energy (in mg Kcal-1) and protein (mg/g) than many grain legumes but the latter have higher K requirements, so crop choice can have a notable influence on fertilizer use, nutrient uptake and global crop production (Jobbágy and Sala 2014). Furthermore, the author found a positive correlation between K fertilization and uptake, but no correlation for N or P, which indicates a severe disparity between fertilizer input and nutrient uptake (Jobbágy and Sala 2014), and that fertilizer use needs to be optimized to match crop needs and to protect soil health. The nutrient indexes described here could serve as a tool for detecting luxury nutrient provision and to plan a more precise and sustainable fertilizer use. The ‘Quantitative evaluation of the fertility of tropical soils’ (QUEPTS) approach has been used to estimate specific N-P-K fertilizer requirements
46

47
for crops based on their maximum and minimum nutrient uptake efficiencies and yield ranges for seed and AGDM (Pathak et al. 2003; Liu et al. 2006). Setiyono et al. (2010) derived a spherical model from QUEPTS taking into consideration differences in nutrient harvest index, to better describe the relationship between nutrient uptake and grain yield, and this method showed to be more effective for estimating precise fertilizer needs.
At the start of the present crop rotation experiment, both faba bean and NL lupin received the same N-P-K fertilizer dose (125 kg/ha of 16-7-13 of N-P-K). This study revealed significant differences in nutrient uptake (Figure 2 in II) and in nutrient harvest indexes (Table 5 in II), indicating that each crop has different nutrient efficiencies and requirements; for example, since the results shown here indicate that NL lupin has higher P and K efficiency, its fertilizer requirements for these minerals should be updated and are likely to be less than those required by faba bean.
The use of nutrient efficiency indexes to estimate fertilizer needs is promising as shown in maize by Setiyono et al. (2010), and the wide range of indexes used here for the assessment of a pre-crop effect could be used similarly. Since UI appeared to be the most informative index, it could be a promising candidate to update the QUEPTS model and improve the precision of fertilizer requirements, but this falls outside the scope of this thesis.
Taking into account the pre-crop effect on the nutrient efficiency indexes of grain legumes (II), which was observed in the current experiments and the existing complementarity between cereals and legumes for improving each other’s nutrient efficiency (Iannetta et al. 2016; Xue et al. 2016) and reducing the environmental impacts of the cropping system (Reckling et al. 2016a); it is apparent that cropping systems benefit from a diversification of crop species, among which grain legumes are a key crop that can help to improve the currently simplified cereal-based rotations. Both NL lupin and barley as pre-crops left the highest plant available P in the soil (Table 2 in II), while faba bean P uptake and yield were higher than those in NL lupin (Figure 2 in II). Faba bean residues are indeed rich in P and have been reported to be useful for the management of soil P pools in rotations with cereals (Alamgir et al. 2012). Thus NL lupin and faba bean have the potential to increase levels of plant-available P in the soil in different ways, either by mobilizing soil P during their growth or by improving the soil P during the decomposition of their residues respectively. That, combined with their ability to biologically fix N and their distinctive nutrient requirements due to grain composition, can potentially account for considerable N and P fertilizer savings in cropping systems (Jobbágy and Sala 2014; MacWilliam et al. 2014). Grain legume cultivation not only reduces the environmental impact of crop imports from distant regions to the Boreal – Nemoral region, but also may improve ecosystem services and land conservation. 4.3 Grain legume adaptation to short growing seasons 4.3.1 Environmental control of flowering
Field screening of the 3 grain legumes showed that cool and wet years were the most challenging for reaching maturity in a timely fashion and it was clear that among the available germplasm, no other available faba bean cultivar could compete with cv. ‘Kontu’ in terms of earliness (I, III). Glasshouse screening, however, has identified 3 landraces as possible sources of earliness and drought resistance in faba bean that can be tested further (Khazaei et al. 2012).
When comparing across species, the NL lupin cv. ‘Haags Blaue’ was the earliest in every year, and lentil cv. ‘Rosebud’ was equally early in 2010, and had comparable maturity dates to cv. ‘Kontu’ in 2011, but did not reach maturity in 2012 (Figure 13). This shows the great potential for adaptation of NL lupin to the short growing season of the Boreal – Nemoral region due to its consistent earliness across years (I). While the earliness of NL lupin increases its chances of exposure to heat and drought stress around flowering, the current study indicated that there is a range of tolerance to these stresses among the studied cultivars. Thus it would be advisable to

diversify not only the crop selection but also the cultivar selection in a given year, even more so under the challenging climate change scenarios (I, II, III).
The field screening trials of faba bean germplasm showed that cultivar, year and the interaction between them had a crucial effect and caused significant differences in flowering and maturity dates (I, II). Differences across years and crossovers in GDD patterns are likely due to the strongly contrasting weather extremes experienced mainly in 2010 (extreme drought by local standards) and 2012 (wettest year, with a record precipitation of 480mm in the growing season) (Table 2 in I).
Traditionally the progress towards flowering (1/f) in faba bean and in most other grain legumes has been described by the model equation developed during the 1980s (Roberts and Summerfield 1987; Ellis et al. 1988a; Ellis et al. 1988c; Ellis et al. 1988b; Ellis et al. 1990) which only considers two variables, Temperature and Photoperiod:
1/f = a + bT + cP (Eq. former model)
The initial field trials, suggested that cultivar Alexia had a particularly different phenological timing across years: in 2011 being faster than in 2010, this trend being opposite to that of most other cultivars which were all earlier in 2010 (Figure 2 in I), indicating that there might be other key factors controlling the progress towards flowering besides temperature and photoperiod. Hence further investigation on the environmental control of flowering in faba bean was initiated.
A series of growth chamber experiments was done to evaluate the effect of photoperiod and temperature on the rate of progress towards flowering; and field experiments across several years provided an opportunity to evaluate natural variation in solar radiation and water deficit as well as temperature and photoperiod. The listed environmental variables were fitted by a linear model in two steps: first the ‘basic model’ (Eq.2) including the four variables without cultivar, and then the ‘extended model’ (Eq.3) including the four variables plus some terms for differing cultivars (III):
1/f ~ Photoperiod + Temperature + PAR2 + K index + Ɛ (Eq.1)
1/f ~ Photoperiod + Temperature + PAR + K index + Kontu + Witkiem Manita + Ɛ (Eq.2)
When dates to flowering stages (EF= early flowering, MF= mid flowering, FF= full flowering) were fitted using the ‘basic’ model or the ‘extended’ model, the R2 values consistently indicated that the model variance was best explained for FF, then for MF and least well for EF, with all R2 being >0.9 (Table 4 and 5 in III). In addition, the K index, PAR and the conventionally used temperature and photoperiod were all significant terms in all models for each flowering stage (P<0.001) (Table 4 and 5 in III).
This trend is coherent with the fact that the number of days to early flowering is highly variable in annual crops, while the number of flowers peaks at mid flowering, giving more stability and reliability to the to the data recorded at MF and FF than at EF (Took et al. 2010). Flowering dates are indeed more consistent at mid and full flowering when 50% and 90% of the plants in a plot are in bloom, while the recorded dates of early flowering representing only 10% of the plants in a plot are more difficult to estimate, and is thus likely to be less consistent across years and plots (I, III). Faba bean cultivars are not genetically uniform as happens in most other crops, so variation in flowering date within a plot, is also a reflection of variation within a cultivar. Moreover, since faba bean is cross-pollinated by insects, there is often gene flow among plots that do not have an inter-
2 *PAR can be replaced by sunshine duration when needed (supplementary data in III).
48

49
plot barrier (Suso et al. 2006) as it was the case on our field trials, thus adding to the variation in flowering date within a plot when seeds for sowing were used from previous year’s plots.
Results from both flowering models, based on the field experiment data, indicated that flowering control in faba bean depends not only on photoperiod and temperature sum but also on solar radiation (as measured by PAR or sunshine duration), and water deficit (as measured by K index).
Figure 13. Phenological stages from sowing until maturity of the earlier cultivars from 3 grain legume species grown in Helsinki during 2010–2012. Phenological stages are expressed as growing degree days (GDD). EF= early flowering, FF= full flowering, EM= early maturity, FM= full maturity. Lentils did not reach maturity in 2012, hence the dotted bar.
The two cultivars that were outliers from the general model differed from the rest by earliness to flowering rather than lateness. They originated from different countries so probably represented different gene pools and possibly earliness genes. In spite of several years of experiments, there were not enough data points for either cultivar to test whether the difference was in the intercept or in the slope of one of the covariates, so further targeted experiments will be necessary to determine which component of progress to flowering differs in ‘Kontu’ and ‘Witkiem Manita’.
The fitting of both models allowed testing the effect of environmental variables on the progress towards flowering, and as expected, the coefficients for temperature and photoperiod were positive, which shows that higher temperatures and longer photoperiods promoted flowering.
Controlled environment experiments allowed the identification of the presence of a ceiling temperature, dependent on day-length and cultivar, that was sometimes reached in the field. Considering the importance of each environmental factor on the progress towards flowering of faba bean and potentially also in that of other grain legumes, the effect of the new variables will be discussed further.

4.3.2 Solar radiation
From all the available sunlight, plants use only that within the range of 400-700 nm wavelength (Taiz and Zeiger 2010). Due to the low solar elevation angles, at noon the values of incoming short-wave radiation (K↓) in the Boreal – Nemoral region usually peak below 1000 Wm-2 during the growing season (Baldocchi et al. 2000). Changes in diurnal radiation occur due to variations in Earth-sun geometry and the presence or absence of clouds modulating daily solar radiation levels (Baldocchi et al. 2000; Carbontree.fi 2016). Hence, over the years the field experiments were exposed to varying light levels due to the fluctuating weather conditions during the different years (I, III).
Earlier studies by Pearson et al. (1993) and then by Adams et al. (1997 and 1999) have identified that a ‘light integral’ (namely the sum of incident PAR) influenced the progress towards flowering in Chrysanthemum and other ornamentals. These studies and the present field observations triggered the decision to include solar radiation variables in the models of progress towards flowering (III), and both PAR and sunshine duration were tested. With PAR giving a better fit than sunshine duration (III). PAR had negative coefficients consistently for both models at all flowering stages, confirming that lower levels of PAR delayed flowering (Table 4 and 5 in III), and allowing the hypothesis that it is a question of sugar rather than of duration (photoperiod).
Although Yeang (2013) emphasized that the key for photoperiod-controlled 1/f was the timing of light transitions rather than its duration, the present results indicate that sunshine duration has an impact on 1/f, probably due to the above mentioned connection with PAR levels (supplementary data in III), and its influence on photosynthate production.
Changes in the pattern of incident solar radiation that occur daily and seasonally (Baldocchi et al. 2000), generate changes in ambient light in a pattern known as the circadian rhythm that controls the expression of the gene CONSTANS (CO) in plants. The CO protein is degraded in darkness, while the expression of the gene increases during the day when peaks in ambient light occur (Johansson and Staiger 2015; Yeang 2013). Extensive genetic studies in Arabidopsis have revealed that the gene CONSTANS (CO) and the stabilization of its protein CO is a requisite for the induction of the FLOWERING LOCUS T (FT), which is the ‘florigen’ that leads plants to flowering (Yeang 2013). In Arabidopsis, over 100 genes that control flowering have been identified, while fewer genes such as E, Hr, Lf and Sn have been identified and described for domesticated pea (Weller et al. 1997; Weller et al. 2009; Weller and Ortega 2015), wild pea and lentil (Weller et al. 2012), though not yet for faba bean (Patrick and Stoddard 2010). Other novel studies have shown two key factors that regulate flowering in Arabidopsis: a) the B-BOX 19 protein that interacts with CO to repress FT expression, a precise mechanism that defines flowering time (Wang et al. 2014) and b) the TREHALOSE- 6-PHOSPHATE SYNTHASE 1 (TPS1), a regulatory factor belonging to the recently found T6P pathway, where the sugar budget of a plant affects the occurrence of a single flowering time (FTi) (Wahl et al. 2013).
In addition to the long-known effects of red and far-red, other light components can also affect flowering: for example UV-A and blue light have been associated with an acceleration of flowering, while UV-B and green work in the opposite direction (Kakani et al. 2003; Kami et al. 2010). Other molecular studies have identified an interaction between the signaling pathways of temperature and light in Arabidopsis (Capovilla et al. 2015). Thus all this evidence supports the significant effect of solar radiation, expressed as PAR or sunshine duration in our model (III), and confirm that it is reasonable to include a specific parameter related to solar radiation or light quality into the flowering models in addition to photoperiod.
50

51
4.3.3 Water deficit
Since cloudy weather with low PAR is often, but not always, associated with rainfall, it was considered relevant to test the K index in the new models of progress towards flowering, based on our field data across several years (III). Climate change has already caused rainfall patterns to become more erratic, affecting both yield and flowering time in crops (Craufurd and Wheeler 2009). K-index values above 1.6 indicate excess water and those below 1.3 indicate deficit (Jedrszczyk et al. 2012), so a full range would be expected to have a curvilinear effect on sensitive aspects of plant growth.
During the years of field experiments included in the flowering model, the rainfall was not high enough to cause waterlogging or to cause the K-index to exceed a value of 1.6 so inhibitory levels of water excess could not be tested often, except for some periods during 2012. In contrast, dry intervals with K < 1.3 were frequent, allowing the testing of water deficit. Hence the fitted values of the model resulted in a linear relationship rather than in a curvilinear relationship (III).
In the ‘basic’ and ‘extended’ flowering models, the coefficients of K index were positive across all flowering stages, (Table 4 and 5 in III). A positive coefficient indicates that high values of K index result in slower time to flowering, while low values of K index resulted in faster time to flowering, which agrees with the early onset of flowering as a strategy of drought escape (Thomas et al. 2004; Anbazhagan et al. 2015). However, as discussed in section 4.1.1., the effect of drought stress may vary depending on the plant developmental stage and on cultivar, due to genotypic differences in temperature limits (III) and drought tolerance such as the reported drought tolerance of cv. ‘Melodie’ (Khan et al. 2007) and susceptibility of cv. ‘Kontu’ (Khazaei 2014).
Once cultivar terms were added in the ‘extended’ model for ‘Kontu’ and ‘Witkiem manita’, the influence of the hydrothermal index on progress to flowering became less marked, particularly at FF as seen in the estimate coefficients. Since this did not happen at EF, it can be suggested that for most cultivars, water deficits are more critical in earlier stages of flower development than at full flowering. Considering that cv. ‘Kontu’ is very sensitive to drought, when it has its own term in the ‘extended’ model equation, the effect of low values of K index becomes less marked for the rest of cultivars (Table 5 in III).
4.3.4 Cultivar temperature limits and ‘earliness per se’
The need for an ‘extended’ model that accurately predicts the flowering of the different range of cultivars screened in our field trials, is due to the wide diversity among the studied set of faba bean germplasm and remarkable differences in some cultivars, mainly in cvs. ‘Kontu’ and ‘Witkiem Manita’. Aspects differentiating these two cultivars from the rest included that the former cultivar had the smallest seed size and a thin seed coat (I), while the latter was the largest seeded, and had a flatter seed shape (I). Cv. ‘Kontu’ was by a significant margin the earliest to mature with a growing season of only 107 days (I,III) (Stoddard and Hämälainen 2011).
In the growth chamber experiments, temperatures of 20°C (short day) and 22°C (long day) were optimal for progress towards flowering, and there were differences between cultivars in the strength of effect of supra-optimal temperatures, with cv. ‘Fatima’ being particularly sensitive (I, III). In general, temperatures above 25°C were detrimental in the progress towards flowering (Figure 8 in III). With the forecasted increase in global temperatures of 2-4˚C (Vadez et al. 2012), the selection of cultivars than can tolerate and be stable along a wide range of temperatures (such as cv. ‘Alexia’ in the 18 h photoperiod) becomes more urgent.
Growth chamber test results showed that cv. ‘Kontu’ was the least sensitive to the shorter photoperiod, and it was the earliest to flower even at the highest temperatures (Figure 3 in III). Although being very drought sensitive, cv. ‘Kontu’ seems to have a much faster circadian clock or

specific ‘earliness per se’ genes, and the present results might also indicate that the faster circadian clock of Kontu may be associated with its high-latitude origin (I, III).
Unfortunately, similar information about temperature limits for cv. ‘Witkiem Manita’ is not available, since it was not included in the growth chamber experiments.
The term ‘earliness per se’ has mainly been used in earliness studies of wheat and barley, and sometimes is also referred to as intrinsic earliness (Slafer 1996). Detailed studies of earliness at the gene level are very scarce in legumes. QTL for earliness have been identified only a) in lentil on six linkage groups (Tullu et al. 2008), and b) in pigeonpea (Cajanus cajan L. Millsp), one major (qFL4.1) and one minor (qFL5.1) for days to flowering, and two major (qMT4.1 and qMT5.1) and one minor (qMT10.1) for days to maturity, all showing a positive additive effect except for locus qMT10.1 (Kumawat et al. 2012).
Other studies have identified a locus that regulates flowering by acting as a central integrator (CI) in several legume species, including PvTFL1y in common bean (Phaseolus vulgaris), MtFTa1 in Medicago truncatula, and GmFT2a, GmFT5a, and GmTFL1 in soybean (Glycine max) (Blümel et al. 2015).
In wheat, the flowering time has been shown to be dependent on vernalization (V), photoperiod (P) and ‘earliness per se’ (EPS) genes, of which the latter are poorly understood due the polygenic nature of the trait (Herndl et al. 2008; Griffiths et al. 2009). EPS drives the development rate of cultivars once the other flowering requirements (P and V) have been met, and EPS groups and germplasm origin groups are related in Argentinian wheat (Gomez et al. 2014).
4.4 Grain legume introduction to Boreal – Nemoral cropping systems and management challenges
Although cultivars from each of the grain legumes included in this research have proven to be early enough at the Boreal – Nemoral boundary, and have the potential to improve greatly the diversity and sustainability of cropping systems. Production and management challenges need to be met, to facilitate the expansion of cultivation further into the arable zone, and to assure sufficient earliness, guaranteeing both production levels and crop quality.
4.4.1 Faba bean
Due to it being the highest yielding crop with the highest nitrogen fixing capacity and fastest closing canopy, faba bean is already the most widely grown of the 3 and possibly the one with the greatest potential. This work has identified 3 key breeding objectives: 1) earliness, 2) resistance to water deficit and high temperature, and 3) resistance to lodging.
The most important need remains earliness. Cv. ‘Kontu’ has been the earliest maturing cultivar on the market for a long time, but while this work was in progress Boreal Plant Breeding has developed two new cultivars that are slightly earlier than ‘Kontu’, namely cvs. ‘Sampo’ and ‘Louhi’ (Laine 2015).
Although later cultivars usually are higher yielding, they are not considered a profitable option to be cultivated in the Boreal – Nemoral region due to the risk of failure of reaching maturity on time before the growing season reaches a harsh end, as is often the case at high latitudes. As discussed in section 4.1.1, the yield penalty certainly represents a challenge for breeders and farmers seeking for earlier cultivars, because by achieving earliness the number of days available for the crop to reach maximum yield is an unavoidably reduced. However, there has been progress in the development of higher yielding and early cultivars, such as cv. ‘Louhi’ that reaches maturity a day after cv. ‘Kontu’ but its bigger seed size allows it to yield 9% more than ‘Kontu’ (Laine et al. 2016).
52

53
Our field and growth chamber tests show that drought and heat spells pose a challenge for the crop, affecting both seed composition and progress towards flowering (I, III), and such challenges will become more frequent due to climate change (Kissoudis et al. 2016). Thus the second priority for breeding should be drought and heat tolerance.
In our experimental farm, there is a daily mean wind speed between 1.12 and 3.5 m/s and maximum wind speed reaches even >23 m/s in the growing season, thus causing faba bean plants of some cultivars to lodge (in some years even >50% plants in a plot suffered lodging). Thus, the third breeding aim should be to select germplasm with strong stems resistant to lodging so that the plant can mature evenly and harvest operations can be done easily.
Further breeding objectives should include decreased content of anti-nutrients in grain and increased biotic stress tolerance. For example, low or zero contents of vicine-convicine and tannins in faba bean are necessary to ensure markets, since their removal increases protein digestibility and feed palatability (Crépon et al. 2010).
Finally, farmers should be aware of the risk of key pest and diseases, and be prepared for field scouting. Black aphids (Aphis fabae), leaf weevils (Sitona lineatus), Lygus bugs and chocolate spot disease (Botrytis fabae and B. cinerea) were all found in these experiments (Figure 14). All can be controlled by integrated pest management (IPM) (Stoddard et al. 2010) and breeding for chocolate spot resistance is widely done (Torres et al. 2006; Sillero et al. 2010; Beyene et al. 2016).
Figure 14. Pest and diseases of faba bean: A) black aphids B) Damage to leaves by weevils C) Marks of chocolate spot disease.
4.4.2 NL lupin
This work has identified 3 key management issues for NL lupin, namely 1) yield levels, 2) soil acidity, and 3) development of seed processing methods for novel food and health uses.
In contrast to faba bean, in the case of NL lupin breeding is needed not so much for earliness but for reducing the penalty that earliness causes on yield levels. Results from all growing seasons showed that there is a significant positive correlation between GDD and yield in NL lupin (Figure 8A in I), the highest yielding cultivars being the latest to reach maturity. The yield penalty in 2009 was huge, considering that there was a difference of 1.6 t/ha between cv. ‘Haags Blaue’ (earliest – low yielding) and cv. ‘Sanabor’ (latest – highest yielding) (Stoddard et al. 2010).

Nevertheless, while higher yielding germplasm becomes available, strategies such as increasing plant density can be used to attain higher yields. The breeders of cv. ‘Haags Blaue’ (Saatzucht Steinach, Germany) recommend a plant density of 125 plants/m2, but this density requires intensive management to control weeds, and harvesting can be challenging due to the short stature of the plants. Hence a small trial was established in 2012 and some of its results were reported in a magazine article (Lizarazo et al. 2013). There were significant differences in TDM (p<0.05), leaf area index (p<0.05 on the 20th July) and height of first pod (p<0.01). Plants at densities of 175 and 200 per m2 set their first pods higher than those at other densities (28-30 cm vs. 25-26 cm). Thus, using higher plant densities could facilitate the action of the harvester cutter bar and potentially reduce harvesting losses. As expected, denser plots built a better canopy cover, and plots with 175 – 200 plants/m2 scored LAI of 5.3, while plots with 100-150 plants/m2 scored 4.8 or less. The intense competition in denser plots was also evident in the speed to reach maturity, with denser plots maturing significantly faster than less dense plots (p<0.05 for 90% maturity), although the difference was just a couple of days. The full canopy cover developed by denser plots represents a major advantage since it can shade weeds.
Yields were significantly higher in denser plots (p<0.05). The 200 plants/m2 yielded 2.8 t/ha, 175 plants/m2 yielded 2.7 t/ha, 125 and 150 plants/m2 yielded 2.5 t/ha and 100 plants/m2 yielded 2.2 t/ha. Thus the optimum plant density for this region may be 175 plants/m2 since the benefits of the highest density did not exceed the increased seeding cost. The effect of plant density needs to be tested in more years (Lizarazo et al. 2013) and it is likely that branching cultivars would not benefit from it.
NL lupin is considered to be better suited to acid soils than to alkaline soils, as a consequence of its calcifuge habit (Zohlen and Tyler 2004), and this is an advantage in many parts of Finland, Sweden and the Baltic countries where many fields are acidic (European Comission 2013).
Finally, it is important to invest in food processing and development programs that make use of NL lupin seeds as raw material, because if there is more demand other than for feed, the seed value will be higher and farmers will be more interested in cultivating it.
NL lupin has several health benefits such as low glycemic index, antioxidant activity, and lowering of cholesterol lowering and blood pressure (Lim 2012b), so there is potential for developing nutraceutical products. Although it is clear that NL lupin seeds are an excellent source of protein and fiber (Table 4 in I), some of their sensory qualities and storage stability still need to be improved (Lucas et al. 2015), and development of processing is needed.
4.4.3 Lentil
For lentil the key management issues are timely maturity, weed management, and risk of
diseases similar to those occurring in pea. Cultivation of lentil during wet years proved to be very challenging, and due to its indeterminate growth habit, the plant kept flowering, and it was not possible to achieve uniform maturity particularly in 2012 (Figure 13). The indeterminate growth habit of lentils, combined with the fact that excess moisture causes a non-uniform pod maturity along the canopy, means that chemical products that trigger desiccation such as “reglone” (Diquat dibromide) might need to be used in wet years (Saskatchewan Pulse Growers, 2000), and specially under future climate change scenarios when autumns are likely to be wetter than at present.
Like NL lupin, lentil closes its canopy rather late, thus giving room for weed development, and its canopy is rather short which might make harvesting difficult. Thus weed management and improvement of the canopy structure or support for it, are the second key issue that need to be considered. One solution is to sow lentil as an intercrop with oat, wheat or flax seed for example. Intercropping with oat and mustard (Brassica juncea L.) has the extra advantage of supporting the lentil vines, facilitating the harvesting of most pods and having extra benefits such as improved water use efficiency and crop productivity (Singh et al. 2009; Singh et al. 2011). However,
54

55
threshing of intercrops can be challenging depending on the selected end use, so other mechanical adjustments might be preferred for harvesting operations. Investing in a roller (to smooth the soil surface) or in other more precise tools such as a ‘swather’, ‘straight cut header’ or ‘vine lifter’ is recommended to improve harvesting efficiency and reduce seed losses (Saskatchewan Pulse-Growers 2000; Erskine et al. 2011).
Finally, there is also the risk that some pea pest and diseases could use lentil as a host. As seen in these field experiments, when pea plants were not available in the surroundings, some pea moths (Cydia nigricana) laid their eggs on lentil (and also NL lupin to a lesser extent). Thöming et al. (2014) reported that pea moth uses plant volatiles which vary depending on the development stage, to locate its host.

5 CONCLUSION AND FUTURE PROSPECTS
Three grain legume crops were studied for their potential to diversify the Boreal – Nemoral crop rotations that are currently narrow and based on cereals. Their high nutrient efficiency and their unique nutritive composition are highlighted in the present study, and it is shown, that the Boreal – Nemoral region needs to expand its cultivation of grain legumes, not only since they are a key ingredient for balanced feed diets and human diets, but also because they are essential components for the sustainable management of nutrient efficiency and emissions in cropping systems.
The field trials showed that all three grain legumes are able to reach maturity and produce seeds of good nutritive quality, a timely manner in most years. Earliness is needed to expand the borders of faba bean cultivation, and its outstanding biological nitrogen fixation, high protein concentration, and its competitive ability with weeds make it a very attractive crop for the diversification of crop rotations. Furthermore, its high metallic nutrient uptake, gives it potential use for phytoremediation and biofortification of foods.
Available cultivars of NL lupin were shown to be early enough for the Boreal – Nemoral region, to be an excellent source of protein and fiber, and their bioactive compounds confer health benefits, so it is an interesting crop. However, its yield levels need to be improved and industrial processing plants need to be set up locally to increase its demand. The earliness of lentil makes it suitable for cultivation in the region, but its low protein concentration and yields, along with its short stature, are likely to ensure that it will be a niche crop rather than a mainstream one. It could be grown as a specialty crop once seed supplies become easier to access and use of desiccants is adopted for a timely harvest.
The project showed that protein concentration and protein yield vary in ways that are not always predictable, and the causes of this variation need to be investigated further at both agronomic and genetic levels, in order that breeding and management strategies can be devised to stabilize protein delivery. Moreover, focus should be given to the value of all the other ecosystem services and nutraceutical uses that grain legumes provide, in order that a fair price for their seed is achieved. Indeed, the price of food grains generally exceeds that of feed grains so investing in food processing and development programs could provide a premium market for top-quality legume seeds, and help to improve and renew the interest of farmers in grain legume cultivation.
Barley and oat were shown to be the best pre-crops for faba bean and narrow-leafed lupin. The mechanisms responsible for the cereal pre-crop effect on legumes need to be investigated further. Studies about exudate release and changes in soil microbiology and chemistry during the pre-crop phase should be conducted. The UI index proved to be a useful indicator to identify significant pre-crop effects, and has potential to be used to assess fertilizer use and nutrient requirements. The UI and other indexes could be optimized to make fertilizer dosages match crop needs more closely and thus protect soil health. Excess of N and P fertilizer could also be controlled by strategically including faba bean or narrow-leafed lupin in rotations, since these crops have a key role in the efficient cycling of these elements, thanks to their BNF ability and root exudates.
European Boreal – Nemoral cropping systems are in need of diversification, in order to be more sustainable and to deal with the risks set by climate change. Diversification of cropping systems is needed not only by selecting legume crops but also by widening the cultivar selection in a given year.
56

57
ACKNOWLEDGEMENTS
This research was carried out in the Crop Science group of the Department of Agricultural Sciences, University of Helsinki. The Research Foundation, the Agriculture and Forestry Fund, the Future Fund and the Department of Agricultural Sciences from the Faculty of Agriculture and Forestry—University of Helsinki; the Häme Student Foundation (Hämääisten ylioppilassäätiö), the August Johannes and Aino Tiura Foundation, the Finnish Cultural Foundation (in particular the Pehr August and Saga Steniuksen Fund, and the A.E. Sandelin Fund), the Ministry of Agriculture and Forestry of Finland (MoniPalko project), the international research project “Legume Futures” funded by the European Union through the Framework 7 Programme (FP7) under Grant Agreement Number 245216 (FP7- KBBE-2009-3) and the FACCE ERA-Net Plus project, Climate Change Adaptability of Cropping and Farming systems for Europe, “Climate CAFE” are acknowledged for funding.
This thesis would not have been possible without the guidance of Professor Fred Stoddard. I have been fortunate to have you as supervisor since the beginning of my MSc studies. Thank you for all the valuable advice, for the countless revisions to the manuscripts and for sharing your knowledge of legume crops with me. Thank you also for giving me the opportunity to work for the Climate CAFE project, it has been a wonderful experience.
I would like to thank Professor Pirjo Mäkelä leader of the Crop Science group for her guidance and kind support. Many thanks to Glenn McDonald and Roxana Savin for the careful revision of this thesis, and to Arja Santanen, Mervi Seppänen, Matti Mõttus and Pedro Aphalo for scientific advice.
I would like to acknowledge my co-authors Jarkko Isotalo, Markku Yli-Halla, Anna-Maija Lampi, Anders Lindfors, Tuula Sontag-Strohm, Vieno Piironen and Jingwei Liu whose contribution made the articles possible. Many thanks to each one of you!
A thousand thanks to Markku Tykkyläinen, for all the assistance during the field and greenhouse experiments, it was heavy work and you were always ready to help and very kind. Thank you for all the talks in the field and the endurance while collecting samples.
Special thanks to all the greenhouse staff: Sanna Peltola, Jouko Närhi, Daniel Richterich, Sini Lindström, Matti Salovaara; and to the laboratory technicians Marjo Kilpinen, Ritva Lofman, Annika Korvenpää, Tuija Niininmäki, Marjut Wallner, each of them contributed significantly to the progress of the field and laboratory work.
Many thanks to all the colleagues from the Department of Agricultural Sciences for their kind help whenever needed and friendly work atmosphere. A very special thanks to my wonderful office-mates and friends Venla Jokela, Nashmin Ebrahimi, and Kiflemariam Yehuala Belachew. I am very grateful for your company, advice and support through the years, thank you for all the good times inside and outside of the office.
I am very grateful for all the technical assistance with field work and growth-chamber experiments received from current and former trainees, colleagues and friends. I am

indebted to, among many others: Marta Cwiertniewska, Heidi Sarkimaa, Tomas Rafsa, Tuula Puhakainen, Guillem Erra, Stefano Zanotto, Yao Lu, Jarno Taavitsainen, Hamid Khazaei, Priit Tammeorg, Antti Tuulos, Kenedy Etone Epie, Lin Chen and Mahmoud Seleiman. Also, many thanks to Mikko Hakojärvi for kindly providing the weather data every year.
I want to express my deepest gratitude to all the friends that have accompanied me through these years, and that have made Helsinki feel like home. I would not have started and survived the PhD without your encouragement and advice. This accomplishment would not have been possible without your constant presence in good and bad times; each one of you has been a positive influence and a constant inspiration. A very heartfelt thanks to: The brunch team: Windi, Anne, Beñat and Victor; Los parceros Leticia y Ali; The dessert lovers: Gina, Laine, Tuomas and Julius; Los guaraníes Ximena y Sascha; The Espoo team: Silvija, Sergey, Monika and Joonas; The UK team: Karolina, Pawel, Iza and Sesh; Os garotos: Carolina y Flavio; and to: Ale, Ciroline, Jenny, Aji, Efi, Zubi, Claudia, Marcela, Ola, Reza, and Waldo.
Finalmente, quiero dedicar esta tesis a mi mamá, por su amor, fuerza, determinación y dedicación, los cuales fueron el motor principal que me motivó a terminar este proyecto tan largo. También mil gracias a mis hermanos Carolina y Jorge, por ser mis cómplices, apoyarme y hacerme reír siempre. Gracias a mi papá y abuelos que, aunque no estén aquí, su amor incondicional, ejemplo y enseñanzas siempre me acompañarán.
Helsinki, April 2017.
Clara Lizarazo
58

59
REFERENCES
Abd-Alla, M.H., Abdel-Wahab E.E., Nivien A.N., David M.K., Fatthy M.M. 2014. Synergistic interaction of Rhizobium leguminosarum Bv. viciae and arbuscular mycorrhizal fungi as a plant growth promoting biofertilizers for Faba bean (Vicia faba L.) in alkaline soil. Microbiological Research 169 (1): 49–58.
Adams, S.R., Pearson, S., Hadley, P. 1997. The effects of temperature, photoperiod and light integral on the time to flowering of Pansy Cv. Universal Violet (Viola X Wittrockiana Gams.). Annals of Botany 80: 107–12.
Adams, S.R., Pearson, S., Hadley, P., Patefield, W.M. 1999. The effects of temperature and light integral on the phases of photoperiod sensitivity in Petunia X hybrida. Annals of Botany 83: 263–69.
Agriobtentions. 2016. http://www.agriobtentions.fr/images/fiches_pdf/Fiche_DIVINE.pdf (Last visited 25.4.2016)
Alamgir, M., McNeill, A., Tang, C., Marschner, P. 2012. Changes in soil P pools during legume residue decomposition. Soil Biology and Biochemistry 49: 70–77.
Anbazhagan, K., Bhatnagar-Mathur, P., Sharma, K.K., Baddam, R., Kavi Kishor, P.B., Vadez V. 2015. Changes in timing of water uptake and phenology favours yield gain in terminal water stressed Chickpea AtDREB1A transgenics. Functional Plant Biology. 42: 84–94.
Andersson, G.K.S., Ekroos, J., Stjernman, M., Rundlöf, M., Smith, H.G. 2014. Effects of farming intensity, crop rotation and landscape heterogeneity on field bean pollination. Agriculture, Ecosystems and Environment 184: 145–48.
Anglade, J., Billen, G., Garnier, J. 2015. Relationships for estimating N2 fixation in legumes : incidence for N balance of legume-based cropping systems in Europe. Ecosphere 6: 1–24.
Angus, J.F., Kirkegaard, J.A., Hunt, J.R., Ryan, M.H., Ohlander, L., Peoples M.B.. 2015. Break crops and rotations for wheat. Crop and Pasture Science 66 (6): 523–52.
Anthony, P., Malzer, G., Sparrow, S., Zhang, M.. 2012. Soybean yield and quality in relation to soil properties. Agronomy Journal 104 (5): 1443–58.
Aronsson, H., Hansen, E.M., Thomsen, I.K., Ogaard, A.F., Känkänen, H.., Ulén, B. 2016. The ability of cover crops to reduce nitrogen and phosphorus losses from arable land in Southern Scandinavia and Finland. Journal of Soil and Water Conservation 71: 41–55.
Austruy, A., Wanat, N., Moussard, C., Vernay, P., Joussein, E., Ledoigt, G., Hitmi, A. 2013. Physiological impacts of soil pollution and arsenic uptake in three plant species: Agrostis capillaris, Solanum nigrum and Vicia faba. Ecotoxicology and Environmental Safety 90: 28–34.
Avio, L., Sbrana, C., Giovannetti, M.. 1990. The response of different species of Lupinus to VAM endophytes. Symbiosis 9: 321–23.
Baldocchi, D., Kelliher, F.M., Black, T.A., Jarvis, P. 2000. Climate and vegetation controls on boreal zone energy exchange. Global Change Biol 6: 69–83.
Berger, J.D., Buirchell, B.J., Luckett, D.J., Palta, J.A., Ludwig, C., Liu, D.L. 2012. How has narrow-leafed lupin changed in its 1st 40 years as an industrial, broad-acre crop? A G×E-based characterization of yield-related traits in Australian cultivars. Field Crops Research 126: 152–64.
Beyene, A.T., Derera, J., Sibiya, J., Fikre, A. 2016. Gene action determining grain yield and chocolate spot (Botrytis fabae) resistance in faba bean. Euphytica 207 (2): 293–304.
Beyer, H., Schmalenberg, A.K., Jansen, G., Jürgens, H.U., Uptmoor, R., Broer, I., Huckauf, J., et al. 2015.

Evaluation of variability, heritability and environmental stability of seed quality and yield parameters of L. angustifolius. Field Crops Research 174: 40–47.
Bishop, J., Potts, S.G., Jones, H.E. 2016. Susceptibility of faba bean (Vicia Faba L.) to heat stress during floral development and anthesis. Journal of Agronomy and Crop Science 202 (6):508-517.
Blümel, M., Dally, M., Jung, C. 2015. Flowering time regulation in crops – what did we learn from Arabidopsis? Curr. Opin. Biotech. 32: 121–29.
Boersma, J.G., Pallotta, M., Li, C., Buirchell, B.J., Sivasithamparam, K., Yang, H. 2005. Construction of a genetic linkage map using Mflp and identification of molecular markers linked to domestication genes in narrow-leafed lupin (Lupinus Angustifolius L.). Cellular & Molecular Biology Letters 10: 331–44.
Borisjuk, L., Weber, H., Panitz, R., Manteuffel, R., Wobus, U. 1995. Embryogenesis of Vicia Faba L.: Histodifferentiation in relation to starch and storage protein synthesis. Journal of Plant Physiology 147: 203–18.
Brand, J.D., Tang, C., Rathjen, A.J. 1999. Adaptation of Lupinus angustifolius L. and Lupinus pilosus Murr. to calcareous soils. Australian Journal of Agricultural Research 50: 1027–33.
Brennan, R.F., Longnecker, N. 2001. Effects of the concentration of manganese in the seed in alleviating manganese deficiency of Lupinus Angustifolius L. Australian Journal of Experimental Agriculture 41: 1199–1205.
Brewin, N.J. 1991. Development of the legume root nodule. Annual Review of Cell Biology 7 (2): 191–226.
Bruning, B., Rozema, J. 2013. Symbiotic nitrogen fixation in legumes: perspectives for saline agriculture. Environmental and Experimental Botany 92: 134–43.
Bues, A., PreiBel, S., Reckling, M., Zander, P., Kuhlman, T., Topp, K., Watson, C., Lindström, K., Stoddard, F.L., Murphy, D. 2013. The environmental role of protein crops in the new common agricultural policy. Brussels.European Parliament (IP/B/AGRI/IC/2012-067) 113p.
Burris, R. H., Evans, H.J. 1993. Biological Nitrogen Fixation. Annual Review of Nutrition 13: 317–35.
Cabello-Hurtado, F., Keller, J., Ley, J., Sanchez-Lucas, R., Jorrín-Novo, J.V., Aïnouche, A. 2016. Proteomics for exploiting diversity of lupin seed storage proteins and their use as nutraceuticals for health and welfare. Journal of Proteomics 143: 57-68.
Campos-Vega, R., Loarca-Piña, G., Oomah, B.D. 2010. Minor components of pulses and their potential impact on human health. Food Research International 43: 461–82.
Capovilla, G., Schmid, M., Posé, D. 2015. Control of flowering by ambient temperature. Journal of Experimental Botany 66: 59–69.
Carbonaro, M., Maselli, P., Nucara, A. 2015. Structural aspects of legume proteins and nutraceutical properties. Food Research International 76: 19–30.
Carbontree.fi. 2016. Solar Radiation. http://www.carbontree.fi/articles/solar-radiation (Last visited 24.4.2016)
Carlsson, G. 2014. Increasing the share of domestic grain legumes in human diets - benefits for cropping and food system sustainability. Protein from plants in a Nordic bio-economy. Nordic Genetic Resource Center. Workshop, Copenhagen 20-21 November.
Cernay, C., Ben-Ari, T., Pelzer, E., Meynard, J.M., Makowski, D. 2015. Estimating variability in grain legume yields across Europe and the Americas. Nature Scientific Reports 5: 1–11.
Cheetham, N.W.H., Cheung, P.C.K., Evans, A.J. 1993. Structure of the principal non-starch polysaccharide from the cotyledons of Lupinus angustifolius (Cultivar Gungurru). Carbohydrate Polymers 22 (1): 37–47.
60

61
Chen, Q., Guo, C.L., Wang, P., Chen, X.Q.,Wu, K.H., Li, K.Z., Yu, Y.X., Chen, L.M. 2013. Up-regulation and interaction of the plasma membrane H(+)-ATPase and the 14-3-3 protein are involved in the regulation of citrate exudation from the broad bean (Vicia faba L.) under Al Stress. Plant Physiology and Biochemistry: PPB / Société Française de Physiologie Végétale 70: 504–11.
Chen, Y.L., Dunbabin, V.M., Diggle, A.J.,Siddique, K.H.M., Rengel, Z. 2012. Assessing variability in root traits of wild Lupinus angustifolius germplasm: basis for modelling root system structure. Plant and Soil 354 (1–2): 141–55.
Chloupek, O., Hrstkova, P. 2005. Adaptation of crops to environment. Theoretical and Applied Genetics. 111: 1316–21.
Cooper, J., Scherer, H. 2012. Nitrogen fixation. In Marschner’s Mineral Nutrition of Higher Plants, 3rd ed., 389–408. London: Academic Press.
Craufurd, P.Q., Wheeler, T.R. 2009. Climate change and the flowering time of annual crops. Journal of Experimental Botany. 60: 2529–39.
Crépon, K., Marget, P., Peyronnet, C., Carrouée, B., Arese, P., Duc, G. 2010. Nutritional value of faba bean (Vicia faba L.) seeds for feed and food. Field Crops Research 115 (3): 329–39.
Crews, T.E., Peoples, M.B. 2004. Legume versus fertilizer sources of nitrogen: ecological tradeoffs and human needs. Agriculture , Ecosystems and Environment 102: 279–97.
Cutforth, H.W., Angadi, S.V., McConkey, B.G., Entz, M.H.,Ulrich, D., Volkmar, K.M., Miller, P.R., Brandt, S. 2013. Comparing plant water relations for wheat with alternative pulse and oilseed crops grown in the semiarid Canadian prairie. Canadian Journal of Plant Science 93: 147–60.
da Silva, V.P., van der Werf, H.M.G., Spies, A., Soares, S.R. 2010. Variability in environmental impacts of Brazilian soybean according to crop production and transport scenarios. Journal of Environmental Management 91 (9): 1831–39.
De Visser, C., Leonardus, M., Schreuder, R., Stoddard, F.L. 2014. The EU’s dependency on soya bean import for the animal feed industry and potential for EU produced alternatives. Oilseeds & Fats Crops and Lipids Journal 21 (4): 1–8.
Denyer K., Hylton C.M., Smith A.M. 1994. The effect of high temperature on starch synthesis and the activity of starch synthase. Functional Plant Biol (21) 783–89.
Divito, G.A., Sadras, V.O. 2014. How do phosphorus, potassium and sulphur affect plant growth and biological nitrogen fixation in crop and aasture legumes? A meta-analysis. Field Crops Research 156: 161–71..
Dracup, M., Thomson, B. 2000. Narrow-leafed lupins with restricted branching. Annals of Botany 85 (1): 29–35.
Duc, G., Trouvelot, A., Gianinazzi-Pearson, V., Gianinazzi S. 1989. First report of non-mycorrhizal plant mutants (Myc-) obtained in pea (Pisum sativum L.) and faba bean (Vicia Faba L.). Plant Science 60: 215–22.
Duc, G., Marget, P., Esnault, R., Le Guen, J., Bastianelli, D. 1999. Genetic variability for feeding value of faba bean seeds (Vicia Faba): comparative chemical composition of isogenics involving zero-tannin and zero-vicine genes. Journal of Agricultural Science 133: 185–96.
Duc, G., Link, W., Marget, P., Redden, R.J., Stoddard, F.L. 2011. Genetic adjustment to changing climates: faba bean. In Crop Adaptation to Climate Change, edited by Lotze-Campen, H., Hall, E.A., Yadav, S.S., Redden, R.J., Hatfield, J.L. 269–86. John Wiley and Sons Ltd.
Duc, G., Agrama, H., Bao, S., Berger, J., Bourion,V., De Ron, A.M., Gowda, C.L.L., et al. 2014. Breeding annual grain legumes for sustainable agriculture: new methods to approach complex traits and target new cultivar ideotypes. Critical Reviews in Plant Sciences 34 (1–3): 381–411.

Dwivedi, S. L., Sahrawat, K.L., Upadhyaya, H.D., Mengoni, A., Galardini, M., Bazzicalupo, M., Biondi, E. G. et al. 2015. Advances in host plant and rhizobium genomics to enhance symbiotic nitrogen fixation in grain legumes. Advances in Agronomy 129: 1-110.
Edelmann, M., Kariluoto, S., Nyström, L., Piironen, V. 2013. Folate in barley grain and fractions. Journal of Cereal Science 58 (1): 37–44.
El Fiel, H.E.A., El Tinay, A.H., Elsheikh, E.A.E. 2002. Effect of nutritional status of faba bean ( Vicia faba L .) on Protein Solubility Profiles. Food Chemistry 76: 219–23.
Elgersma, A., Søegaard, K. 2016. Effects of species diversity on seasonal variation in herbage yield and nutritive value of seven binary grass-legume mixtures and pure grass under cutting. European Journal of Agronomy 78: 73–83.
Ellis, R.H., Roberts, E.H., Summerfield, R.J. 1988a. Photothermal time to flowering in faba bean (Vicia faba) and the analysis of potential vernalization responses. Annals of Botany 61: 73–82.
Ellis, R.H., Roberts, E.H., Summerfield, R.J. 1988b. Variation in the optimum temperature for rates of seedling emergence and progress towards flowering amongst six genotypes of faba bean (Vicia Faba). Annals of Botany 62: 119–26.
Ellis, R.H., Summerfield, R.J., Roberts, E.H. 1988. Effects of temperature, photoperiod and seed vernalization on flowering in faba bean (Vicia Faba). Annals of Botany 61: 17–27.
Ellis, R.H., Summerfield, R.J., Roberts, E.H.. 1990. Flowering in faba bean: genotypic differences in photoperiod sensitivity, similarities in temperature sensitivity, and implications for screening germplasm. Annals of Botany 65: 129–38.
Erskine, W. 2009. Global production, suppy and demand. In The lentil: botany, production and uses, 4–12.
Erskine, W., Sarker, A., Kumar, S. 2011. Crops that feed the world 3. Investing in lentil improvement toward a food secure world. Food Security 3: 127–39.
European Comission. 2013. Soil pH in Europe. European Soil Portal. http://esdac.jrc.ec.europa.eu/content/soil-ph-europe. (Last visited 15.5.2016)
EUROSTAT. 2013. Agricultural census. http://ec.europa.eu/eurostat/statistics-explained/index.php/Agricultural_census_2010. (Last visited 20.5.2016)
EUROSTAT. 2014. Harvested production and area of production of field peas and broad beans, EU-28. http://ec.europa.eu/eurostat/statistics-explained/index.php?title=File:Harvested_production_and_area_of_production_of_field_peas_and_broad_beans,_EU-28,_2014.png&oldid=259394 (Last visited 22.5.2016)
EUROSTAT. 2016. European statistics. http://ec.europa.eu/eurostat/data/database (Last visited 7.8.2016)
Evans, L.T. 1959. Environmental control of flowering in Vicia faba. L. Annals of Botany 23: 521–45.
Fader, M., Gerten, D., Thammer, M., Heinke, J., Lotze-Campen, H., Lucht, W., Cramer, W. 2011. Internal and external green-blue agricultural water footprints of nations, and related water and land savings through trade. Hydrology and Earth System Sciences 15 (5): 1641–60.
FAO. 1992. Technology of production of edible flours and protein products from soybeans. http://www.fao.org/docrep/t0532e/t0532e00.htm (Last visited 15.3.2016)
FAO. 2011. Dietary protein quality evaluation in human nutrition. FAO food and nutrition paper 92. Rome, Italy.
FAO. 2016a. FAOstat. http://www.fao.org/faostat/en/#data (Last visited 20.8.2016)
FAO. 2016b. Pulses: nutritious seeds for a sustainable future. FAO. Rome, Italy. 196 pp.
62

63
FAOstat. 2016. Production of crops. http://www.fao.org/faostat/en/#data/QC (Last visited 15.8.2016)
Farooq, M., Gogoi, N., Barthakur, S., Baroowa, B., Bharadwaj, N., Alghamdi, S.S., Siddique, K.H.M. 2016. Drought stress in grain legumes during reproduction and grain filling. Journal of Agronomy and Crop Science, 1–22.
Flores, F., Hybl, M., Knudsen, J.C., Marget, P., Muel, F., Nadal, S., Narits, L., et al. 2013. Adaptation of spring faba bean types across European climates. Field Crops Research 145.: 1–9.
FMI. 2016a. Finnish Meteorological Institute http://en.ilmatieteenlaitos.fi/climate (Last visited 10.4.2016)
FMI. 2016b. Finnish Meteorological Institute. Seasons in Finland. http://en.ilmatieteenlaitos.fi/seasons-in-finland (Last visited 10.4.2016)
Foley, R.C., Gao, L.L., Spriggs, A., Soo, L.Y.C., Goggin, D.E., Smith, P.M.C., Atkins, C.A. Singh, K.B. 2011. Identification and characterisation of seed storage protein transcripts from Lupinus angustifolius. BMC Plant Biology 11 (59).
Foltête, A.S., Masfaraud, J.F., Férard, J.F., Cotelle, S. 2012. Is there a relationship between early genotoxicity and life-history traits in Vicia faba exposed to cadmium-spiked soils? Mutation Research - Genetic Toxicology and Environmental Mutagenesis 747 (2): 159–63.
Freedman, D., Lane, D. 1983. A non-stochastic interpretation of reported significance levels. Journal of Business and Economic Statistics 1: 292–98.
Freibauer, A., Kaltschmitt, M. 2003. Controls and models for estimating direct nitrous oxide emissions from temperate and sub-boreal agricultural mineral aoils in Europe. Biogeochemistry 63: 93–115.
Gallardo, K., Thompson, R., Burstin, J. 2008. Reserve accumulation in legume seeds. Comptes Rendus - Biologies 331 (10): 755–62.
Gan, Y.T., Miller, P.R., MacConkey, B.G., Zentner, R.P., Stevenson, F.C., MacDonald, C.L. 2003. Influence of diverse cropping sequences on durum wheat yield and protein in the semi-arid Northern Great Plains. Agronomy Journal 95: 245–52.
Gaur, P.M., Singh, M.K., Samineni, S., Sajja, S.B., Jukanti, A.K., Kamatam, S., Varshney, R.K. 2016. Inheritance of protein content and its relationships with seed size, grain yield and other traits in chickpea. Euphytica 209 (1): 253–60.
Gephart, J. A., Davis, K.F., Emery, K.A., Leach, A.M., Galloway, J.N., Pace, M.L. 2016. The environmental cost of subsistence: optimizing diets to minimize footprints. Science of the Total Environment 553: 120–27.
Giménez, M.A., Drago, S.R., Bassett, M.N., Lobo, M.O., Sammán, N.C. 2016. Nutritional improvement of corn pasta-like product with broad bean (Vicia faba) and quinoa (Chenopodium quinoa). Food Chemistry 199: 150–56.
Giunta, F., Pruneddu, G., Motzo, R. 2009. Radiation interception and biomass and nitrogen accumulation in different cereal and grain legume species. Field Crops Research 110: 76–84.
Godar, J., Persson, U.M., Tizado, E.J., Meyfroidt, P. 2015. Towards more accurate and policy relevant footprint analyses: tracing fine-scale socio-environmental impacts of production to consumption. Ecological Economics 112: 25–35.
Gomez, D., Vanzetti, L., Helguera, M., Lombardo, L., Fraschina, J., Miralles, D.J. 2014. Effect of Vrn-1, Ppd-1 genes and earliness per se on heading time in Argentinean bread wheat cultivars. Field Crops Research 158: 73–81.
Granstedt, A. 2012. Farming for the future. BERAS Implementation Reports No. 2. Stockholm, Sweden.
Grela, E.R., Günter, K.D. 1995. Fatty acid composition and tocopherol content of some legume seeds.

Animal Feed Science and Technology 52: 325–31.
Gresshoff, P.M., Hayashi, S., Biswas, B., Mirzaei, S., Indrasumunar, A., Reid, D., Samuel, S. et al. 2015. The value of biodiversity in legume symbiotic nitrogen fixation and nodulation for biofuel and food production. Journal of Plant Physiology 172: 128–36.
Griffiths, S., Simmonds, J., Leverington, M., Wang, Y., Fish, L., Sayers, L., Alibert, L., Orford, S., Wingen, L., Herry, L., Faure, S., Laurie, D., Bilham, L., Snape, J. 2009. Meta-QTL analysis of the genetic control of ear emergence in elite European winter wheat germplasm. Theoretical and Applied Genetics 119: 383–95.
Grusak, M.A. 2009. Nutritional and health-beneficial quality. In: The Lentil: Botany, Production and Uses, 368–90.
Häusling, M. 2011. The EU protein deficit: what solution for a long-standing problem?. Committee on Agriculture and Rural Development. European Parliament. Brussels, Belgium.
Henriksen, B.I.F., Lundon, A.R., Prestløkken, E., Abrahamsen, U., Eltun, R. 2009. Nutrient supply for organic oilseed crops, and quality of potential organic protein feed for ruminants and poultry. Agronomy Reseach 7: 592–98.
Herndl, M., White, J., Hunt, L., Graeff, S., Claupein, W. 2008. Field based evaluation of vernalization requirement, photoperiod response and earliness per se in bread wheat (Triticum aestivum L.). Field Crops Research 105: 193–201.
LHMS. 2016. Climate in Lithuania. http://old.meteo.lt/english/climate_air.php (Last visited 17.4.2016)
Hood, G., Karunakaran, R., Downie, J.A. Poole, P.S. 2015. MgtE from Rhizobium leguminosarum is a Mg2+ channel essential for growth at low pH and N2 fixation on specific plants. Molecular Plant-Microbe Interactions 28 (12): 1281–87.
Iannetta, P., Young, M., Bachinger, J., Bergkvist, G., Doltra, J.,. Lopez-Bellido, R.J., Monti, M. et al. 2016. A comparative nitrogen balance and productivity analysis of legume and non-legume supported cropping systems: the potential role of biological nitrogen fixation. Frontiers in Plant Science 7 Article 1700.
Iglesias, A., Quiroga, S., Moneo, M. 2012. From climate change impacts to the development of adaptation strategies: challenges for agriculture in Europe. Climatic Change 112: 143–68.
Jedrszczyk, E., Skowera, B., Kopcinska, J., Ambroszczyk, A. 2012. The influence of weather conditions during vegetation period on yielding of twelve determinate tomato cultivars. Not. Bot. Hort. Agrobot. Cluj-Napoca 40: 203–9.
Jenner, C.F., Ugalde, T.D., Aspinall, D. 1991. The phisiology of starch and protein deposition in the endosperm of wheat. Journal of Plant Physiology 18: 211–26.
Jensen, E.S., Peoples, M.B., Hauggaard-Nielsen, H. 2010. Faba bean in cropping systems. Field Crops Research 115: 203–16.
Jensen, E.S, Peoples, M., Boddey, R., Gresshoff, P., Hauggaard Nielsen, H., Alves, B., Morrison, M. 2012. Legumes for mitigation of climate change and the provision of feedstock for biofuels and biorefineries. A review. Agronomy for Sustainable Development. Vol. 32.
Jia, Y., Gray, V.M., Straker, C.J. 2004. The influence of rhizobium and arbuscular mycorrhizal fungi on nitrogen and phosphorus accumulation by Vicia faba. Annals of Botany 94 (2): 251–58.
Jiang, Z.Q., Pulkkinen, M., Wang, Y., Lampi, A.M., Stoddard, F.L., Salovaara, H., Piironen, V., Sontag-Strohm, T. 2016. Faba bean flavour and technological property improvement by thermal pre-treatments. LWT - Food Science and Technology 68: 295–305.
Jobbágy, E.G., Sala, O.E. 2014. The imprint of crop choice on global nutrient needs. Environmental
64

65
Research Letters 9. 10pp.
Johansson, M., Staiger, D. 2015. Time to flower: interplay between photoperiod and the circadian clock. Journal of Experimental Botany 66: 719–730.
Kakani, V.G., Reddy, K.R., Zhao, D., Sailaja, K. 2003. Field crop responses to ultraviolet-B radiation: A review. Agricultural and Forest Meteorology 120: 191–218.
Kami, C., Lorrain, S., Hornitschek, P., Frankhauser, C. 2010. Light-regulated plant growth and development. In: Current Topics in Developmental Biology, edited by Timmermans, M. 29–66.
Kape, R., Wex, K., Parniske, M., Gorge, E., Wetzel, A., Werner, D. 1992. Legume root metabolites and VA-mycorrhiza development. Journal of Plant Physiology 141: 54–60.
Kariluoto, S., Edelmann, M., Piironen, V. 2010. Effects of environment and genotype on folate contents in wheat in the HEALTHGRAIN diversity screen. Journal of Agricultural and Food Chemistry 58: 9324–31.
Kaschuk, G., Kuyper, T., Leffelaar, P., Hungria, M., Giller, K. 2009. Are the rates of photosynthesis stimulated by the carbon sink strength of rhizobial and arbuscular mychorrhizal symbioses? Soil Biology and Biochemistry 41: 1233–44.
Kaschuk, G., Leffelaar, P., Giller, K., Alberton, O., Hungria, M., Kuyper, T. 2010. Responses of legumes to rhizobia and arbuscular mychorrhizal fungi: A meta-analysis of potential photosynthate limitation of symbioses. Soil Biology and Biochemistry 42: 125–27.
Kessel, C. V., Hartley, C. 2000. Agricultural management of grain legumes: Has it led to an increase in nitrogen fixation? Field Crops Research 65: 165–81.
Khan, H.U.R., Link, W., Hocking, T. J., Stoddard, F. L.. 2007. Evaluation of physiological traits for improving drought tolerance in faba bean (Vicia faba L.). Plant and Soil 292 (1–2): 205–17.
Khazaei, H., Stoddard, F.L., Lizarazo, C.I., Street, K. 2012. New sources of earliness for Finnish faba bean breeding. In: Maataloustieteen Päivät 2012, edited by Schulman, N. and Kauppinen, H. Helsinki.
Khazaei, H. 2014. Leaf traits associated with drought adaptation in faba bean (Vicia faba L.). University of Helsinki. 59pp. Helsinki, Finland.
Khoshgoftarmanesh, A.H., Schulin, R., Chaney, R.L., Daneshbakhsh, B. 2011. Micronutrient-efficient genotypes for crop yield and nutritional quality in sustainable agriculture. Agronomy for sustainable development 30 (1): 83-107.
Kirkegaard, J., Christen, O., Krupinsky, J., Layzell, D. 2008. Break crop benefits in temperate wheat production. Field Crops Research 107 (3): 185–95.
Kiss, S. A., Stefanovits-Bányai, E., Takács-Hájos, M. 2004. Magnesium-content of rhizobium nodules in different plants: the importance of magnesium in nitrogen-fixation of nodules. Journal of the American College of Nutrition 23 (6): 751S–753S.
Kissoudis, C., Van De Wiel, C., Visser, R.G.F.,Van Der Linden, G. 2016. Future-proof crops: challenges and strategies for climate resilience improvement. Current Opinion in Plant Biology 30: 47–56.
Kumawat, G., Raje, R.S., Bhutani, S., Pal, J.K., Mithra, A.S.V.C.R., Gaikwad, K., Sharma, T., Singh, N. 2012. Molecular mapping of QTLs for plant type and earliness traits in pigeon pea (Cajanus cajan L. Millsp.). BMC Genetics 13: 84–95.
Laine, A. 2015. Sampo ja Louhi ovat uudet härkäpavun lajikeet. Maaseudun Tiede. 1/2015, 6p.
Laine, A., Kujala, M., Niskanen, M. 2016. Virallisten lajikekokeiden tulokset 2008-2015. Helsinki. 239pp.
Lampart-Szczapa, E., Korczak, J., Nogala-Kalucka, M., Zawirska-Wojtasiak, R. 2003. Antioxidant properties of lupin seed products. Food Chemistry 83 (2): 279–85.

Lassaletta, L., Billen, G., Grizzetti, B., Garnier, J., Leach, A.M., Galloway, J.N. 2014. Food and feed trade as a driver in the global nitrogen cycle: 50-year trends. Biogeochemistry 118 (1–3): 225–41.
Leach, A.M., Emery, K.A., Gephart, J., Davis, K.F., Erisman, J.W., Leip, A., Pace, M.L., et al. 2016. Environmental impact food labels combining carbon, nitrogen, and water footprints. Food Policy 61: 213–23.
Leip, A., Weiss, F., Lesschen, J.P., Westhoek, H. 2014. The Nitrogen footprint of food products in the European Union. The Journal of Agricultural Science 152 (Supplement S1): 20–33.
Li, L., Li, S.M., Sun, J.H., Zhou, L.L., Bao, X.G., Zhang, H.G., Zhang, F.S. 2007. Diversity enhances agricultural productivity via rhizosphere phosphorus facilitation on phosphorus-deficient soils. Proceedings of the National Academy of Sciences of the United States of America 104 (27): 11192–96.
Lim, T.K. 2012a. Lens culinaris. In: Edible medicinal and non-medicinal plants. Vol 2, Fruits, 742–53. Springer Science+Business Media B.V. Dordrecth, Netherlands.
Lim, T.K. 2012b. Lupinus angustifolius. In: Edible medicinal and non-medicinal plants. Vol 2, Fruits, 770–78. Springer Science+Business Media B.V. Dordrecth, Netherlands.
Lim, T,K. 2012c. Vicia faba. In: Edible medicinal and non-medicinal plants. Vol 2, Fruits, 925–36. Springer Science+Business Media B.V. Dordrecth, Netherlands.
Lindström, K., Murwira, M., Willems, A., Altier, N. 2010. The biodiversity of beneficial microbe-host mutualism: the case of rhizobia. Research in Microbiology 161 (6): 453–63.
Link, W., Balko, C., Stoddard, F.L. 2010. Winter hardiness in faba bean: physiology and breeding. Field Crops Research 115 (3): 287–96.
Liu, M., Yu, Z., Liu, Y., Konijn, N.T. 2006. Fertilizer requirements for wheat and maize in China: the QUEFTS approach. Nutrient Cycling in Agroecosystems 74 (3): 245–58.
Lizarazo, C.I., Rekola, K., Edelmann, M., Kariluoto, S., Lampi, A.M., Piironen, V., Stoddard, F.L.. 2011. Nutritional quality of Finnish-grown lupin (L. angustifolius). In: Lupin Crops - an Opportunity for Today, a Promise for the Future. Proceedings of the 13th International Lupin Conference., edited by B. Naganowska, P. Kachlicki, and B. Wolko, 205–7. Poznán: International lupin Association. Poland.
Lizarazo, C.I., Jokela, V., Stoddard, F.L. 2013. Sinilupiini sopii Suomen kasvuoloihin ja viljelykiertoon. Maaseudun Tiede 8/2013. 8p.
LMC. 2009. Evaluation of measures applied under the common agricultural policy to the protein crop sector. Executive summary. Oxford, UK. 13pp.
López-bellido, F.J., López-bellido, L., López-bellido, R. . 2005. Competition, growth and yield of faba bean (Vicia faba L.). European Journal of Agronomy 23: 359–78.
Lucas, M.M., Stoddard, F.L., Annicchiarico, P., Frías, J., Martínez-Villaluenga, C., Sussmann, D., Duranti, M., Seger, A., Zander, P. M., Pueyo, J.J. 2015. The future of lupin as a protein crop in Europe. Frontiers in Plant Science 1–6.
Luce, M.S., Grant, C.A., Zebarth, B.J., Ziadi, N., Donovan, J.T.O., Blackshaw, R.E., Harker, K.N. et al. 2015. Legumes can reduce economic optimum nitrogen rates and increase yields in a wheat – canola cropping sequence in Western Canada. Field Crops Research 179: 12–25..
Lupwayi, N.Z., Clayton, G.W., Hanson, K.G., Rice, W.A., Biederbeck, V.O. 2004. Endophytic rhizobia in barley, wheat and canola roots. Canadian Journal of Soil Science 84: 37–45.
MacWilliam, S., Wismer, M., Kulshreshtha, S. 2014. Life cycle and economic assessment of Western Canadian pulse systems: the inclusion of pulses in crop rotations. Agricultural Systems 123: 43–53.
Magrini, M.B., Anton, M., Cholez, C., Corre-Hellou, G., Duc, G., Jeuffroy, M.H., Meynard, J.M., Pelzer, E.,
66

67
Voisin, A.S., Walrand, S. 2016. Why are grain-legumes rarely present in cropping systems despite their environmental and nutritional benefits? analyzing lock-in in the French agrifood system. Ecological Economics 126: 152–62.
Makkar, H.P.S., Becker, K., Abel, H., Pawelzik, E. 1997. Nutrient contents, rumen protein degradability and antinutritional factors in some colour- and white-flowering cultivars of Vicia faba beans. Journal of the Science of Food and Agriculture 75 (4): 511–20.
Malhi, S.S., Lemke, R. 2007. Tillage, crop residue and N fertilizer effects on crop yield, nutrient uptake, soil quality and nitrous oxide gas emissions in a second 4-yr rotation cycle. Soil & Tillage Research 96: 269–83.
Malik, R.S., Seymour, M., French, R.J., Kirkegaard, J.A., Lawes, R.A., Liebig, M.A. 2015. Dynamic crop sequencing in Western Australian cropping systems. Crop and Pasture Science 66: 594–609.
Manschadi, A.M., Sauerborn, J., Stu, H., Saxena, M.C., Go, W.. 1998. Simulation of faba bean (Vicia faba L.) root system development under Mediterranean conditions. European Journal of Agronomy 9: 259–72.
Manske, G.G.B., Van Ginkel, R.M., Rajaram, S., Vlek, P.L.G. 2002. Phosphorus use efficiency in tall, semi-dwarf and dwarf near-isogenic lines of spring wheat. Euphytica 125: 113–19.
Martin, N. 2014. What is the way forward for protein supply? The European perspective. Oilseeds & Fats Crops and Lipids Journal 21 (4): 1–8.
McConkey, B.G., Liang, B.C., Campbell, C.A., Curtin, D., Moulin, A., Brandt, S.A., Lafond, G.P. 2003. Crop rotation and tillage impact on carbon sequestration in Canadian prairie soils. Soil and Tillage Research 74: 81–90.
Metzger, M.J., Brus, D.J., Bunce, R.G.H., Carey, P.D., Gonçalves, J., Honrado, J.P., Jongman, R.H.G., Trabucco, A., Zomer, R. 2013. Environmental stratifications as the basis for national, European and global ecological monitoring. Ecological Indicators 33: 26–35.
Mihailovic, V., Mikic, A., Eric, P., Vasiljevic, S., Cupina, B., Katic, S. 2005. Protein pea in animal feeding. Biotechnology in Animal Husbandry 21 (5–6): 281–85.
Mihailovic, V., Mikic, A., Cupina, B., Krstic, D., Antanasovic, S., Eric, P., Pataki, I. 2011. Faba bean as a forage, biofuel and green manure crop. Grain Legumes 56 (56): 34.
Mokma D.L., Yli-Halla, M., Hartikainen H. 2000. Soils in a young landscape on the coast of Southern Finland. Agricultural and Food Science in Finland 9 (4): 291–302.
Muñoz-Romero, V., López-Bellido, L., López-Bellido, R.J. 2011. Field crops research faba bean root growth in a vertisol: tillage effects. Field Crop Research 120: 338–44.
Muranyi, I.S., Volke, D., Hoffmann, R., Eisner, P., Herfellner, T., Brunnbauer, M., Koehler, P., Schweiggert-Weisz, U. 2016. Protein distribution in lupin protein isolates from Lupinus angustifolius L. prepared by various isolation techniques. Food Chemistry 207: 6–15.
Musgrave, M., Allen, J., Blasiak, J., Tuominen, L., Kuang, A. 2008. In vitro seed maturation in Brassica rapa L.: relationship of silique atmosphere to storage reserve deposition. Environmental and Experimental Botany 62: 247–53.
Mwanamwenge, J., Loss, S.P., Siddique, K.H.M., Cocks, P. S.. 1999. Effect of water stress during floral inititation, flowering and podding on the growth and yield of faba bean (Vicia faba L.). European Journal of Agronomy 11: 1–11.
Myers, S.S, Zanobetti, A., Kloog, I., Huybers, P., Leakey, A.D.B., Bloom, A.J., Carlisle, E., et al. 2014. Increasing CO2 threatens human nutrition. Nature 510: 139–43.
Myrbeck, A., Stenberg, M. 2014. Changes in N leaching and crop production as a result of measures to

reduce N losses to water in a 6-yr crop rotation. Soil Use and Management 30: 219–30.
Nemecek, T., Von Richthofen, J.S., Dubois, G., Casta, P., Charles, R., Pahl, H. 2008. Environmental impacts of introducing grain legumes into European crop rotations. European Journal of Agronomy 28: 380–93.
Nikolic, Z., Ðorpevic, V., Torbica, A., Mikic, A. 2012. Legumes seed storage proteins characterization by SDS-PAGE and lab-on-a-chip electrophoresis. Journal of Food Composition and Analysis 28: 75–80.
Nutman, P.S. 1969. Genetics of symbiosis and nitrogen fixation in legumes. Proceedings of the Royal Society of London. Series B, Biological Sciences 172: 417–37.
O’Donovan, J.T., Grant, C.A., Blackshaw, R.E., Harker, K.N., Johnson, E.N., Gan, Y., Lafond, G.P., et al. 2014. Rotational effects of legumes and non-legumes on hybrid canola and malting barley. Crop Economics, Production and Management 106 (6): 1921–32.
Oba, H., Tawaray, K., Wagatsuma, T. 2015. Arbuscular mycorrizal colonization in Lupinus and related genera. Plant Nutrition 47: 685–94.
OECD/Food and Agriculture Organization of the United Nations. 2015. OECD-FAO Agricultural Outlook 2015-2024. Paris: OECD Publishing.
Oita, A., Malik, A., Kanemoto, K., Geschke, A., Nishijima, S., Lenzen, M. 2016. Substantial nitrogen pollution embedded in international trade. Nature Geoscience 9 (January): 6–10.
Olesen, J.E., Bindi, M. 2002. Consequences of climate change for European agricultural productivity, land use and policy. European Journal of Agronomy 16: 239–62.
Ouertatani, S., Regaya, K., Ryan, J., Gharbi, A. 2011. Soil liming and mineral fertilization for root nodulation and growth of faba beans in an acid soil in Tunisia. Journal of Plant Nutrition 34 (6): 850–60.
Øygarden, L., Deelstra, J., Lagzdins, A., Bechmann, M., Greipsland, I., Kyllmar,K., Povilaitis, A., Iital, A. 2014. Climate change and the potential effects on runoff and nitrogen losses in the Nordic – Baltic region. Agriculture, Ecosystems and Environment 198: 114–26.
Pampana, S., Masoni, A., Arduini, I. 2016. Grain legumes differ in nitrogen accumulation and remobilisation during seed filling. Acta Agriculturae Scandinavica 66: 127–32.
Pate, J.S., Williams, W., Farrington, P. 1985. Lupin (Lupinus spp.). In: Grain Legume Crops, 699–739.
Pathak, H., Aggarwal, P.K., Roetter, R., Kalra, N., Bandyopadhaya, S.K., Prasad, S., Van Keulen, H. 2003. Modelling the quantitative evaluation of soil nutrient supply, nutrient use efficiency, and fertilizer requirements of wheat in India. Nutrient Cycling in Agroecosystems 65 (2): 105–13.
Patrick, J.W., Stoddard, F.L. 2010. Physiology of flowering and grain filling in faba bean. Field Crops Research 115 (3): 234–42.
Pearse, S., Venekaas, E., Cathray, G., Bolland, M.A., Lambers, H. 2006. Carboxylate release of wheat, canola and 11 grain legume species as affected by phosphorus status. Plant and Soil 288: 127–39.
Pearson, S., Hadley, P., Wheldon, A.E. 1993. A Reanalysis of the Effects of Temperature and Irradiance on the Time to Flowering in Chrysanthemum (Dendranthema grandiflora). Journal of Horticultural Science 68: 89–97.
Peel, M.C., Finlayson, B.L., McMahon, T.A. 2006. Updated world map of the Köppen-Geiger climate classification. Meteorologische Zeitschrift 15 (3): 259–63.
Peltonen-Sainio, P., Peltonen, J. 1993. Stability of quality traits in spring cereals cultivated under the growing conditions of Southern Finland. Acta Agriculturae Scandinavica. Section B Soil and Plant Science 43: 45-52.
Peltonen-Sainio, P., Jauhiainen, L., and Sadras, V. O.. 2011. Phenotypic plasticity of yield and agronomic
68

69
traits in cereals and rapeseed at high latitudes. Field Crops Research 124: 261–69.
Peltonen-Sainio, P., Jauhiainen, L., Nissilä, E. 2012. Improving cereal protein yields for high latitude conditions. European Journal of Agronomy 39: 1–8.
Peltonen-Sainio, P., Niemi, J.K. 2012. Protein crop production at the Northern margin of farming: to boost, or not to boost. Agricultural and Food Science 21: 370–83.
Peltonen-Sainio, P., Hannukkala, A., Voutila, L., Niemi, J. 2013. Potential and realities of enhancing rapeseed- and grain legume-based protein production in a Northern Climate. Journal of Agricultural Science 151: 303–21.
Penning de Vries, F.W.T., Brunsting, A.H.M., van Laar, H.H. 1974. Products requirements and rfficiency of biosynthesis: a quantitative approach. Journal of Theoretical Biology 45: 339–77.
Peoples, M.B., Brockwell, J., Herridge, D.F., Rochester, I.J., Alves, B.J.R., Urquiaga, S., Boddey, R.M., et al. 2009. The contributions of nitrogen-fixing crop legumes to the productivity of agricultural systems. Symbiosis 48: 1–17.
Philippi, J., Schliephake, E., Jürgens, J.U., Jansen, G., Ordon, F. 2016. Correlation of the alkaloid content and composition of narrow-leafed lupins (Lupinus angustifolius L.) to aphid susceptibility. Journal of Pest Science 89 (2): 359–73.
Poonam, R.B., Sharma, R., Handa, N., Kaur, H., Kaur, R., Sirhindi, G., Thukral, A.K. 2014. Prospects of field crops for phytoremediation of contaminants. Emerging technologies and management of crop stress tolerance. In: Emerging technologies and management of crop stress tolerance. Second Edition. Vol. 2. Elsevier Inc. 449-470pp.
Pulkkinen, M., Zhou, X., Lampi, A.M., Piironen, V. 2016. Determination and stability of divicine and isouramil produced by enzymatic hydrolysis of vicine and convicine of faba bean. Food Chemistry 212: 10–19.
Pypers, P., Huybrighs, M., Diels, J., Abaidoo, R., Smolders, E., Merckx, R. 2007. Does the enhanced P acquisition by maize following legumes in a rotation result from improved soil P availability? Soil Biology and Biochemistry 39: 2555–66.
Pywell, R.F., Warman, E.A., Hulmes, L., Hulmes, S., Nuttall, P., Sparks, T.H., Critchley, C.N.R., Sherwood, A. 2006. Effectiveness of new agri-environment schemes in providing foraging resources for bumblebees in intensively farmed landscapes. Biological Conservation 129: 192–206.
Rajesh, S., Phillips, M., Rao, S., Reddy, N. 2015. Antioxidant activities and characterisation of polysaccharides isolated from the seeds of Lupinus angustifolius. Industrial Crops & Products 74: 950–56.
Reckling, M., Hecker, J.M., Bergkvist, G., Watson, C.A., Zander, P., Schläfke, N., Stoddard, F.L., et al. 2015. A cropping system assessment framework-evaluating effects of introducing legumes into crop rotations. European Journal of Agronomy 76: 186–97.
Reckling, M., Bergkvist, G., Watson, C.A., Stoddard, F.L., Zander, P.M., Walker, R.L., Pristeri, A., Toncea, I., Bachinger, J. 2016a. Trade-offs between economic and environmental impacts of introducing legumes into cropping systems. Frontiers in Plant Science 7 (May): 1–15.
Reckling, M., Hecker, J.M., Bergkvist, G., Watson, C.A., Zander, P., Schläfke, N., Stoddard, F.L., et al. 2016b. A cropping system assessment framework. Evaluating effects of introducing legumes into crop rotations. European Journal of Agronomy 76: 186–97.
Roberts, E., Summerfield, R. 1987. Measurement and prediction of flowering in annual crops. In: Manipulation of Flowering, Proceedings of previous Easter schools in Agricultural Science. Sutton Bonington, England. 17–50pp.
Rosa, E.V.C., Valgas, C., Souza-Sierra, M.M., Corrêa, A.X.R., Radetski, C.M. 2003. Biomass growth,

micronucleus induction, and antioxidant stress enzyme responses in Vicia faba exposed to cadmium in solution. Environmental Toxicology and Chemistry / SETAC 22 (3): 645–49.
Rose, TJ., Damon, P., Rengel, Z. 2010. Phosphorus-efficient faba bean (Vicia faba L.) genotypes enhance subsequent wheat crop growth in an acid and an alkaline soil. Crop and Pasture Science 1009–16.
Rose, T. J., Wissuwa, M. 2012. Rethinking internal phosphorus utilization efficiency: a new approach is needed to improve PUE in grain crops. Advances in Agronomy 116: 185–217.
Roy, F., Boye, J.I., Simpson, B.K. 2010. Bioactive proteins and peptides in pulse crops : pea, chickpea and lentil. Food Research International 43 (2): 432–42.
Safronova, V.L., Piluzza, G., Bullitta, S., Belimov, A.A. 2011. Use of legume-microbe symbioses for phytoremediation of heavy metal polluted soils: advantages and potential problems. In: Handbook for Phytoremediation, edited by I.A. Golubev, 443–69. NOVA Sci. Publ.
Sadras, O. 2005. The N: P stoichiometry of cereal, grain legume and oilseed crops. Field Crops Research 95: 13–29.
Saskatchewan Pulse-Growers, SPG. 2000. General production. In: Pulse Production Manual. Saskatchewan. Canda.
Saxena, M.C. 2009. Plant Morphology, Anatomy and Growth Habit. In: The Lentil: botany, production and uses, 34–46.
Schumacher, H., Paulsen, H.M., Gau, A.E., Link, W., Jürgens, H.U., Sass, O., Dieterich, R. 2011. Seed protein amino acid composition of important local grain legumes Lupinus angustifolius L ., Lupinus luteus L ., Pisum sativum L . and Vicia faba L . Plant Breeding 130: 156–64.
Schwenke, G.D., Herridge, D.F., Scheer, C., Rowlings, D.W., Haigh, B.M., Mcmullen, K.G. 2015. Soil N2O Emissions under N2 - fixing legumes and N-fertilised canola: a reappraisal of emissions factor calculations. Agriculture, Ecosystems and Environment 202: 232–42.
Setiyono, T.D., Walters, D.T., Cassman, K.G., Witt, C., Dobermann, A. 2010. Estimating maize nutrient uptake requirements. Field Crops Research 118 (2): 158–68.
Sharma, B. 2009. Genetics of economic rraits. In: The Lentil: botany, production and uses, 76–101.
Siddique, K.H.M., Regan, K.L., Tennant, D., Thomson, B.D. 2001. Water use and water use efficiency of cool season grain legumes in low rainfall Mediterranean-type environments. European Journal of Agronomy 15 (4): 267–80.
Siddique, K.H.M., Johansen, C., Turner, N.C., Jeuffroy, M.H., Hashem, A., Sakar, D., Gan, Y., Alghamdi, S.S. 2012. Innovations in agronomy for food legumes . A Review. Agronomy for Sustainable Development 32: 45–64.
Siddique, K.H.M., Erskine, W., Hobson, K., Knights, E.J., Leonforte, A., Khan, T.N., Paull, J.G., Redden, R., Materne, M. 2013. Cool-season grain legume improvement in Australia - use of genetic resources. Crop and Pasture Science 64 (4): 347–60.
Sillero, J.C., Villegas-Fernández, A.M., Thomas, J., Rojas-Molina, M.M, Emeran, A.A., Fernández-Aparicio, M., Rubiales, D. 2010. Faba bean breeding for disease resistance. Field Crops Research 115 (3): 297–307.
Silva, V.C., Almeida, S.M., Resgalla, C., Masfaraud, J.F., Cotelle, S., Radetski, C.M. 2013. Arsenate (As V) in water: quantitative sensitivity relationships among biomarker, ecotoxicity and genotoxicity endpoints. Ecotoxicology and Environmental Safety 92: 174–79.
Šimek, M., Virtanen, S., Krištůfek, V., Simojoki, A., Yli-Halla, M. 2011. Evidence of rich microbial communities in the subsoil of a boreal acid sulphate soil conducive to greenhouse gas emissions. Agriculture, Ecosystems & Environment 140 (1–2): 113–22.
70

71
Sinclair, T.R., Vadez, V. 2012. The future of grain legumes in cropping systems. Crop and Pasture Science 63 (6): 501–12.
Singh, T., Rana, K.S., Singh, S.Y., et al. 2009. Productivity and sustainability of mustard (Brassica juncea L.) and lentil (Lens culinaris L.) intercropping system as affected by moisture conservation practices and fertility levels under rainfed conditions. Archives of Agronomy and Soil Science 55 (2): 183–96.
Singh, U., Singh, S.R., Saad, A.A., et al. 2011. Yield advantage, reciprosity functions and energy budgeting of lentil (Lens culinaris) plus oat (Avena sativa) intercropping under varying row ratio and phosphorus management. Indian Journal of Agricultural Sciences 81 (3): 219–25.
Slafer, G.A. 1996. Differences in phasic development rate amongst wheat cultivars independent of responses to photoperiod and vernalization. A viewpoint of the intrinsic earliness hypothesis. Journal of Agricultural Science 126: 403–19.
Sorensen, J., Larsen, J., Jakobsen, I. 2005. Mycorrhiza formation and nutrient concentration in leeks. Plant and Soil 273 (1): 101–14.
Stevenson, D.G., Doorenbos, R.K., Jane, J.L., Inglett, G.E. 2006. Structures and functional properties of starch from seeds of three soybean ( Glycine max ( L .) Merr .) varieties. Starch 58: 509–19.
Stoddard, F.L., Bond, D.A. 1987. The pollination requirements of the faba bean. Bee World 68 (3): 144–52.
Stoddard, F.L., Marshall, D.R., Ali, S.M. 1993. Variability in grain protein concentration of peas and lentils grown in Australia. Australian Journal of Agricultural Research 44: 1415–19.
Stoddard, F.L. 1993. Limits to retention of fertilized flowers in faba beans (Vicia faba L.). Journal of Agronomy and Crop Science 171: 251–59.
Stoddard, F.L., Hovinen, S., Kontturi, M., Lindström, K., Nykänen, A. 2009. Legumes in Finnish Agriculture: History, present status and future prospects. Agricultural and Food Science 18: 191–205.
Stoddard, F.L. 2010. Improving food and feed security in the Nordic and Baltic region by using appropriate crop rotations. Agronomy Research 8 (II): 311–16.
Stoddard, F.L., Lizarazo, C.I., Mäkelä, P., Nykänen, A. 2010. New annual legume crops for Finnish conditions. In Maataloustieteen Päivät, edited by Hopponen, A. Helsinki. Finland.
Stoddard, F.L., Nicholas, A.H., Rubiales, D., Thomas, J. 2010. Integrated pest management in faba bean. Field Crops Research 115: 308–18.
Stoddard, F.L., Hämälainen, K. 2011. Towards the world’s earliest maturing faba bean. Grain Legumes 56: 9–10.
Sujak, A., Kotlarz, A., Strobel, W. 2006. Compositional and nutritional evaluation of several lupin seeds. Food Chemistry 98: 711–19.
Suso, M. J., Gilsanz, S., Duc, G., Marget, P., Moreno, M.T. 2006. Germplasm management of faba bean (Vicia faba L.): monitoring intercrossing between accessions with inter-plot barriers. Genetic Resources and Crop Evolution 53 (7): 1427–39.
Sussmann, D., Pickardt, C., Schweiggert, U., Eisner, P. 2013. Influence of different proccesing parameters on the isolation of lupin (Lupinus angustifolius L.) protein isolates: a preliminary study. Journal of Food Process Engineering 36: 18–28.
Taiz, L., Zeiger, E. 2010. Plant physiology and development. 6th ed. Sinauer. Massachusetts, USA.
Thomas., Robertson, M.J., Fukai, S., Peoples, M.B. 2004. The effect of timing and severity of water deficit on growth, development, yield accumulation and nitrogen fixation of mungbean. Field Crop Research 86: 67–80.
Thöming, G., Norli, H.R., Saucke, H., Knudsen, G.K. 2014. Pea plant volatiles guide host location behaviour

in the pea moth. Arthropod-Plant Interactions 8: 109–22.
Thomson, B.D., Siddique, K.H.M., Barr, M.D., Wilson, J.M. 1997. Grain legume species in low rainfall mediterranean-type environments I. Phenology and seed yield. Field Crops Research 54: 173–87.
Time and date. 2016. Time and date. https://www.timeanddate.com/ (Last visited 15.4.2016)
Took, F., Battey, N.H. 2010. Temperate flowering phenology. Journal of Experimental Botany 61: 2853–62.
Torres, A., Frias, J., Vidal-Valverde, C. 2005. Changes in chemical composition of lupin seeds (Lupinus angustifolius) after selective α-galactoside extraction. Journal of the Science of Food and Agriculture 85 (14): 2468–74.
Torres, A.M., Román, B., Avila, C.M., Satovic, Z., Rubiales, D., Sillero, J.C., Cubero, J.I., Moreno, M.T. 2006. Faba bean breeding for resistance against biotic stresses: towards application of marker technology. Euphytica 147 (1–2): 67–80.
Tullu, A., Tar’an, B., Warkentin, T., Vandenberg, A. 2008. Construction of an intraspecific linkage map and QTL analysis for earliness and plant height in lentil. Crop Science 48 (6): 2254–64.
Tveito, O. E., Forland, E. J., Alexandersson, H., Drebs, A., Jónsson, T., Tuomenvirta, H., Vaarby Laursen E.. 2001. Nordic climate maps. NORDKLIM Report 06/01. DNMI. Oslo, Norway. 29p.
Unkovich, M.J., Pate, J.S. 2000. An appraisal of recent field measurements of symbiotic N2 fixation by annual legumes. Field Crops Research 65 (2–3): 211–28.
Vadez, V., Berger, J.D., Warkentin, T., Asseng, S., Ratnakumar, P., Poorna Chandra Rao, K., Gaur, P.M. 2012. Adaptation of grain legumes to climate change : A review. Agrononmy for Sustainable Development 32: 31–44.
Vagstad, N., Stålnacke, P., Andersen, H., Deelstra, J., Jansons, V., Kyllmar, K., Loigu, E. 2004. Regional variations in diffuse nitrogen losses from agriculture in the Nordic and Baltic regions. Hydrology and Earth System Sciences 8 (4): 651–62.
Valkama, E., Lemola, R., Känkänen, H., Turtola, E. 2015. Meta-analysis of the effects of undersown catch crops on nitrogen leaching loss and grain yields in the Nordic countries. Agriculture, Ecosystems & Environment 203: 93–101.
van Dijk, Kimo C., Lesschen, J.P., Oenema, O. 2016. Phosphorus flows and balances of the European Union member states. Science of the Total Environment 542: 1078–93.
van Grinsven, H.J.M., Erisman, J.W., de Vries, W., Westhoek, H. 2015. Potential of extensification of European agriculture for a more sustainable food system, focusing on nitrogen. Environmental Research Letters 10 (2), 025002.
Vilariño, M., Ravetta, D.A. 2008. Tolerance to berbivory in lupin genotypes with different alkaloid concentration: interspecific differences between Lupinus albus L. and L. angustifolius L. Environmental and Experimental Botany 63 (1–3): 130–36.
Vilariño, M., Métayer, J. P., Crépon, K., Duc, G. 2009. Effects of varying vicine, convicine and tannin contents of faba bean seeds (Vicia faba L.) on nutritional values for broiler chicken. Animal Feed Science and Technology 150 (1–2): 114–21.
Villarino, C.B.J., Jayasena, V., Coorey, R., Chakrabarti-bell, S., Foley, R., Fanning, K., Johnson, S.K. 2015. The effects of lupin (Lupinus angustifolius) addition to wheat bread on its nutritional, phytochemical and bioactive composition and protein quality. Food Research International 76: 58–65.
Vioque, J., Alaiz, M., Girón-Calle, J. 2012. Nutritional and functional properties of Vicia faba protein isolates and related fractions. Food Chemistry 132: 67–72.
Voisin, A.S., Guéguen, J., Huyghe, C., Jeuffroy, M.H., Magrini, M.B., Meynard, J.M., Mougel, C., Pellerin,
72

73
S., Pelzer, E. 2014. Legumes for feed, food, biomaterials and bioenergy in Europe : A review. Agronomy for Sustainable Development 34 (2): 361–80.
Von Richthofen, J. S., GL-Pro partners. 2006.What do European farmers think about grain legumes? Grain Legumes, no. 45: 13–22.
Wahl, V., Ponnu, J., Sclereth, A., Arrivault, S., Langenecker, T., Franke, A., Feil, R., Lunn, J. E., Stitt, M., Schmid, M. 2013. Regulation of flowering by trehalose-6-phosphate signaling in Arabidopsis thaliana. Science 339: 704–7.
Wang, C.Q., Guthrie, C., Sarmast, M.K., Dehesh, K. 2014. BBX19 interacts with CONSTANS to repress FLOWERING LOCUS T transcription, defining a flowering time checkpoint in Arabidopsis. Plant Cell 26: 3589–3602.
Wang, N., Daun, J. K. 2006. Effects of variety and crude protein content on nutrients and anti-nutrients in lentils (Lens culinaris). Food Chemistry 95: 493–502.
Wang, N., Hatcher, D.W., Gawalko, E.J. 2008. Effect of variety and processing on nutrients and certain anti-nutrients in field peas (Pisum sativum). Food Chemistry 111: 132–38.
Wang, N., Hatcher, D.W., Warkentin, T. D., Toews, R. 2010. Effect of cultivar and environment on physicochemical and cooking characteristics of field pea ( Pisum sativum ). Food Chemistry 118 (1): 109–15.
Weller, J.L., Reid, J.B., Taylor, S.A., Murfet, I.C. 1997. The genetic control of flowering in pea. Trends in Plant Science 2: 412–18.
Weller, J.L., Hecht, V., Liew, L.C., Sussmilch, F.C., Wenden, B., Knowles, C.L., Vander Schoor, J.K. 2009. Update on the genetic control of flowering in garden pea. Journal of Experimental Botany 60: 2493–99.
Weller, J.L., Liew, L.C., Hecht, V.F.G., Rajandran, V., Laurie, R.E., Ridge, S., Wenden, B., Vander Schoor, J.K., Jaminon, O., Blassiau, C., Dalmais, M., Rameau, C., Bendahmane, A., Macknight, R.C., Lejeune-Hénaut, I. 2012. A conserved molecular basis for photoperiod adaptation in two temperate legumes. PNAS 109: 21158–63.
Weller, J.L., Ortega, R. 2015. Genetic control of flowering time in legumes. Frontiers in Plant Science 6 (April): 207.
West, P.C., Gerber, J.S., Engstrom, P.M., Mueller, N.D., Brauman, K.A., Carlson, K.M., Cassidy, E.S., et al. 2014. Leverage points for improving global food security and the environment. Science 345 (6194): 325–27.
White, G.A., Smith, L.A., Houdijk, J.G.M., Homer, D., Kyriazakis, I., Wiseman, J. 2015. Replacement of soya bean meal with peas and faba beans in growing / finishing pig diets: effect on performance, carcass composition and nutrient excretion. Animal Feed Science and Technology 209: 202–10.
Williams, C.M., King, J. R., Ross, S. M., Olson, M.A., Hoy, C.F., Lopetinsky, K. J. 2014. Effects of three pulse crops on subsequent barley, canola, and wheat. Agronomy Journal 106 (2): 343–50.
Winkler, A., Ridgway, G., Webster, M., Smith, S., Nichols, T. 2014. Permutation inference for the general linear model. NeuroImage 92: 381–97.
Wong, A., Pitts, K., Jayasena, V., Johnson, S. 2013. Isolation and foaming functionality of acid-soluble protein from lupin (Lupinus angustifolius) kernels. Journal of the Science of Food and Agriculture 93: 3755–62.
Xue, Y., Xia, H., Christie, P., Zhang, Z., Li, L., Tang, C. 2016. Crop acquisition of phosphorus, iron and zinc from soil in cereal/legume intercropping systems: A critical review. Annals of Botany 117 (3): 363–77.
Yadav, S.S., McNeil, D.L., Andrews, M., Chen, C., Brand, J., Singh, G., Shivakumar, B.G., Gangaiah, B. 2009. Soil nutrient management. In: The lentil: botany, production and uses, 194–212.

74
Yaseen, M., Malhi, S. 2009. Differential growth response of wheat genotypes to ammonium phosphate and rock phosphate phosphorus sources differential growth response of wheat genotypes to ammonium phosphate and rock phosphate. Journal of Plant Nutrition 32: 410–32.
Yeang, H.Y. 2013. Solar rhythm in the regulation of photoperiodic flowering of long-day and short-day plants. Journal of Experimental Botany 64: 2643–52.
Yli-Halla, M., Nyborg, Å. 2014. Update of the WRB soil classes in the 250K soil database of Finland: expression of soil moisture regime in mineral soils. In: Proceedings of the 20th World Congress of Soil Sciences. Jeju, South Korea.
Zander, P., Amjath-Babu, T.S., Preissel, S., Reckling, M., Bues, A. Schläfke, N., Kuhlman, T., et al. 2016. Grain legume decline and potential recovery in European agriculture: A review. Agronomy for Sustainable Development 36 (26).
Zhen-Zhen, C., Gang, P., Fu-Biao, W., Ke-Su, W., Zhao-Wei, L., Chun-Hai, S., Wei, G. 2015. Effect of high temperature on the expressions of genes encoding starch synthesis enzymes in developing rice endosperms. Journal of Integrative Agriculture 14 (4): 642–59.
Zhou, K., Slavin, M., Lutterodt, H., Whent, M., Eskin, N.A.M., Yu, L. 2013. Cereals and legumes. In: Biochemistry of Foods 3–41.
Zohlen, A., Tyler, G. 2004. Soluble inorganic tissue phosphorus and calcicole – calcifuge behaviour of plants. Annals of Botany 94: 427–32.





























