Embed Size (px)
Citation preview

1 2 Frequency and domain specificity of toxin-neutralizing paratopes in the human 3 antibody response to the anthrax vaccine AVA 4 5 6 7 Donald Reason1*, Justine Liberato1, Jinying Sun1, Wendy Keitel2, and Jianhui Zhou1 8
9
1Children's Hospital Oakland Research Institute, Oakland, CA, USA 10
2Baylor College of Medicine, Departments of Molecular Virology, Microbiology and 11
Medicine, Houston, TX, USA 12
13
running title: Frequency and specificity of PA-neutralizing paratopes 14
15
*Corresponding author: Donald C. Reason, Ph.D. 16
Children's Hospital Oakland 17
Research Institute 18
5700 Martin Luther King Jr. Way 19
Oakland, CA 94609 USA 20
Tel 510.450.7638 21
Fax: 510.450.7910 22
email: [email protected] 23
Copyright © 2009, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved.Infect. Immun. doi:10.1128/IAI.01254-08 IAI Accepts, published online ahead of print on 17 February 2009
on August 18, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

Abstract 1
Protective antigen (PA) is the cell surface recognition unit of the binary anthrax toxin 2
system, and the primary immunogenic component in both the current and proposed "next 3
generation" anthrax vaccines. Several studies utilizing animal models have indicated that 4
PA-specific antibodies, acquired either by active or passive immunization, are sufficient 5
to protect against infection with Bacillus anthracis. To investigate the human antibody 6
response to anthrax immunization, we have established a large panel of human PA-7
specific monoclonal antibodies derived from multiple individuals vaccinated with the 8
currently approved anthrax vaccine BioThrax™. We have determined that although 9
these antibodies bind PA in standard binding assays such as ELISA, Western blots, 10
capture assays, and dot blots, less than 25% are capable of neutralizing lethal toxin (LT) 11
in vitro. Non-neutralizing antibodies also fail to neutralize toxin when present in 12
combination with other non-neutralizing paratopes. Although neutralizing antibodies 13
recognize determinants throughout the PA monomer, they are significantly less common 14
among those paratopes that bind to the immuno-dominant amino terminal portion of the 15
molecule. These findings demonstrate that PA binding alone is not sufficient to 16
neutralize LT, and suggest that for an antibody to effectively block PA-mediated toxicity, 17
it must bind to PA such that one of the requisite toxin functions is disrupted. A vaccine 18
design strategy that directed a higher percentage of the antibody response towards 19
neutralizing epitopes may result in a more efficacious vaccine for the prevention of 20
anthrax infection. 21
on August 18, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

Introduction 1
The Bacillus anthracis binary toxin system contributes directly to anthrax 2
pathogenicity in the host (3, 14). The cell surface recognition element of this toxin 3
system is an 83kd protein known as protective antigen (PA83). Antibodies that bind PA 4
protect against infection (8, 12), and PA is the primary immunogenic component in the 5
anthrax vaccine currently licensed for use in the United States (BioThrax™ or AVA, 6
Emergent Biosystems). Ongoing attempts to develop a "next generation" anthrax vaccine 7
are relying on a recombinant form of PA as the sole immunogenic component. PA's role 8
as an important vaccine target has driven a significant amount of research into both the 9
biology of this protein toxin, and the immunobiology of its interaction with the immune 10
system of the vaccinated or infected host. 11
PA83 binds to the cell surface receptors tumor endothelial marker 8 (TEM8) and the 12
capillary morphogenesis gene 2 product (CMG2) (4, 20). Bound PA is cleaved by cell 13
surface associated furin proteases to release the 20kd amino-terminal portion of the 14
molecule (PA20), which has no further role in intoxication. Following proteolytic 15
cleavage, cell-bound PA63 self-assembles to form a heptameric pre-pore structure that can 16
bind several molecules of the catalytic toxin components lethal factor (LF) and/or edema 17
factor (EF). Receptor-mediated endocytosis results in the internalization of the complex, 18
which inserts into the membrane of the endocytic vacuole. LF and/or EF are then actively 19
translocated into the cytoplasm of the cell. The structure of PA, both as a monomer and 20
heptamer, has been determined (15, 19), and the regions of the molecule (domains) 21
involved in the various functions described above have been identified (1, 6, 15, 18, 19). 22
The immunobiology of the immune response to PA in vaccinated humans has only 23
on August 18, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

recently been explored at the molecular level. PA elicits a polyclonal antibody response 1
in vaccinated humans that utilizes a wide variety of immunoglobulin variable (V) region 2
genes. Preliminary studies have indicated that post vaccination antibodies undergo the 3
somatic hypermutation and class switch normally associated with affinity maturation 4
(22). We have previously demonstrated the human antibody response to PA to be 5
significantly biased towards epitopes associated with the amino-terminal domain of the 6
PA protein (PA20), and have postulated that these antibodies may be deficient in their 7
ability to neutralize toxin (16). 8
In this report, we determine the toxin neutralization potential of a large panel of 9
individual monoclonal antibodies isolated from seven individuals vaccinated with AVA 10
using a cell-based assay of LT-mediated cytotoxicity. We find that only 24% of the 11
component antibodies that comprise the overall response are capable of neutralizing PA-12
mediated cytotoxicity in vitro. We found no direct correlation between the relative PA 13
binding ability of the individual antibodies and their ability to neutralize anthrax toxin. 14
We also determine that toxin-neutralizing paratopes occur less frequently among those 15
antibodies that recognize the immunodominant epitopes associated with the amino-16
terminal domain of the PA monomer. These findings suggest that the efficacy of future 17
PA-based vaccines might be improved by modifying the immunogen such that a greater 18
proportion of the antibody response is directed toward those epitopes that lead to toxin 19
neutralization. 20
21
Materials and Methods 22
Subjects: The donors analyzed in this report were recruited from individuals taking part 23
on August 18, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

in a larger study of the response to AVA being conducted at Baylor College of Medicine. 1
Human subject protocols were reviewed and approved by the Institutional Review Boards 2
at both Children’s Hospital Oakland and Baylor College of Medicine. 3
Construction of Fab expression libraries: Fab expression libraries were constructed 4
from mononuclear cells (MNCs) enriched for PA-specific B cells in a manner similar to 5
that previously described for PA and polysaccharide-specific antibody expression 6
libraries (16, 17, 21, 22). PA83 was purchased from List Biological Laboratories, 7
Campbell, CA. PA-specific Fabs were identified using a sensitive 125I-labeled PA 8
capture assay and lysates of individual E. coli expression cultures. Positive isolates were 9
re-cloned, heavy (H) and light (L) chain gene sequence determined, and PA-specific 10
binding confirmed by ELISA. Initial sequence analysis utilized the NCBI IgBlast server 11
(http://www.ncbi.nlm.nih.gov/igblast/) to identify candidate germline gene (2). 12
Subsequent analysis, alignments and translations were performed using MacVector 13
(Accelrys Inc, Princeton, NJ). H and L chain V region gene nomenclature is as described 14
in the IMGT database (11). Complementarity determining regions (CDRs) are as defined 15
in (9). 16
Expression of PA-specific bivalent antibody in Chinese Hamster Ovary (CHO) cells: 17
In vitro expression of full chain IgG1 antibodies utilized the Flp-in CHO system from 18
Invitrogen (Carlsbad, CA). H and L chain V region gene segments were isolated from 19
PA-specific Fabs and inserted into the FRT vector as a bicistronic eukaryotic expression 20
cassette utilizing an Internal Ribosomal Entry Segment (IRES) sequence. Flp-In CHO 21
cells were plated at 3.5 x 105 cells per well (in 2 mL Flp-In medium) in Nunclon Delta 6-22
well plates and then incubated at 37˚C, 5% CO2 overnight. Once cells reached 80% 23
on August 18, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

confluence they were transfected with pOG44 and the FRT vector (9:1 ratio) using the 1
TransFast transfection reagent (Promega). Forty-eight hours after transfection, the cells 2
were trypsinized and placed in a fresh 6-well plate under drug selection with 600 µg/mL 3
hygromycin. Antibody was concentrated from the cell culture supernatant for use in 4
binding and toxin neutralization assays. 5
In vitro toxin neutralization assay: PA and LF were purchased from List Biological 6
Laboratories, Carlsbad, CA. RAW 264.7 cells were plated at 4 x 104 cells per well (in 65 7
µL assay medium) in Nunclon Delta 96-well plates and incubated at 37˚C, 5% CO2 for 8
approximately 4 to 5 hours to ensure proper settling and attachment. Antibodies and 9
PA/LF were pre-incubated in Greiner Bio-One 96-well plates at 37˚C, 5% CO2 for 1 10
hour, and then were transferred to the 96-well plate containing RAW 264.7 cells. The 11
plate was then placed back into the incubator at 37˚C, 5% CO2 for overnight incubation. 12
The following morning, 20 µL Cell-Titer Blue reagent (Promega) was added to each well 13
of the assay plate. Optical density (at 570/595nm) was determined for each well four 14
hours later using a microtiter plate reader. 15
Antigen concentration and binding assays: Antibody concentration was determined by 16
a capture ELISA in which goat anti-human Fc (The Binding Site, Birmingham, UK) 17
immobilized on a microtiter plate captures IgG, which is then detected by alkaline-18
phosphatase labeled goat anti-human L chain (Biosource International, Camarillo, CA). 19
This assay is standardized using a purchased IgG1 protein standard (Sigma). PA binding 20
in ELISA was determined for IgG1 antibodies on 96-well plates coated with 5 µg/ml 21
PA83 and developed with alkaline-phosphatase conjugated goat antibody specific for 22
human kappa or lambda light chains. 23
on August 18, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

Domain-specific blockade of neutralizing antibodies in the RAW cell assay: 1
Neutralizing antibodies were pre-incubated with modified forms of PA to verify the 2
specificity of the antibody-PA binding in the toxin neutralization assay. PA20 (residues 1 3
- 191) was expressed fused to GFP as previously described (16). A non-functional 4
mutant of PA (PArb-) was constructed using QuickChange™ Mutagenesis (Stratagene) 5
by mutating two residues in the Domain 4 region of wild type PA (N682A and D683A) to 6
remove its ability to bind to the cell surface receptor. This modification was necessary to 7
prevent PA added to the assay as an inhibitor from participating in LF-mediated toxicity. 8
Neutralizing antibodies were incubated with the modified PA constructs overnight at 9
concentrations sufficient for 50% inhibition of LT-mediated cytotoxicity. These were 10
then added to the neutralization assay as described above, and the degree to which the 11
pre-incubation blocked the antibodies ability to neutralize toxin was calculated. 12
Statistical analysis: Differences between groups were analyzed using the Mann-13
Whitney U test and the two-sided Fisher's exact test as implemented within the Prism™ 14
graphic software package. 15
on August 18, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

Results 1
Non-neutralizing paratopes predominate in the human antibody response to PA: 2
We have isolated and characterized a large panel of PA-specific monoclonal antibodies 3
from AVA-immunized donors (16, 22). Sequence analysis of heavy (H) chain joining (J) 4
region usage and variable (V) H chain complementarity determining region 3 (CDR3) 5
sequence and length allowed us to assign the over 120 somatically mutated and 6
differentiated antibodies isolated from these donors to 64 families that arose from unique 7
and independent B-cell rearrangement events (16). To determine the toxin neutralization 8
potential of the 64 individual members of the panel, the Ig V region coding genes of each 9
isolate were cloned into a bi-cistronic, eukaryotic expression vector and transfected into 10
CHO cells to facilitate the production of each antibody paratope as a bi-valent IgG1 11
molecule. CHO cells were maintained in culture, and expressed human monoclonal 12
antibody concentrated from the culture supernatant and quantitated prior to analysis. Six 13
of the 64 antibodies transferred into the CHO expression system failed to express 14
sufficient quantity of antibody for testing. To determine the lethal toxin neutralization 15
potential of the 58 expressed antibodies, we employed a well-established in vitro RAW 16
264.7 cell-based assay of LT-mediated cytotoxicity. Titrated quantities of antibody were 17
incubated with PA and LF to allow antibodies to bind PA. Antibody/toxin mixtures were 18
then added to previously seeded RAW 264.7 cells in 96-well plates, and cell viability 19
accessed using CellTiter-Blue™ (Promega) following overnight culture. Optical density 20
(O.D.) in each antibody-containing well was measured and calculated as a percentage of 21
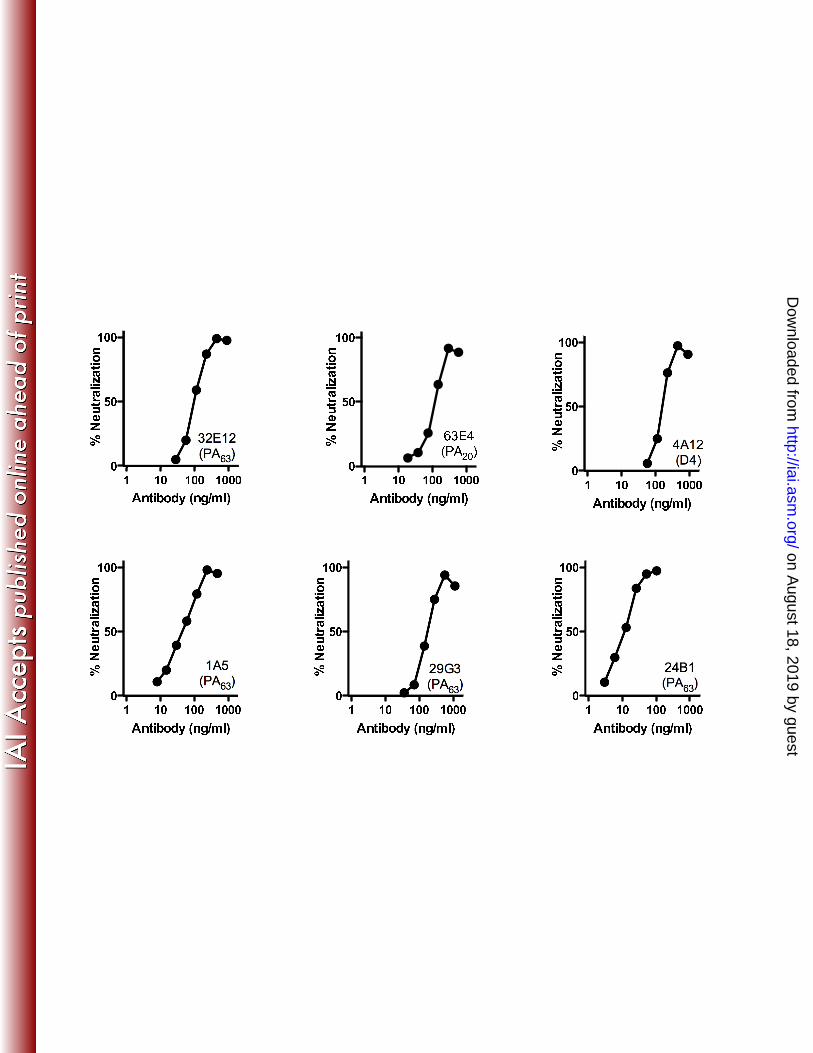
that in wells containing no toxin. Data were expressed as % viability versus antibody 22
concentration (Fig. 1). 23
on August 18, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

The majority of the individual paratopes failed to neutralize lethal toxin in culture 1
at any concentration tested. The maximum concentration of antibody tested varied for 2
each clone based on the level of expression we were able to achieve in the CHO cell 3
culture for that particular clone. All were tested at molar concentrations in excess of the 4
3.6 nM PA in the assay and in excess of that required to demonstrate neutralization by the 5
poorest neutralizing clone in the panel (see below). Several non-neutralizing clones, 6
when tested in combination of up to four antibodies, were also unable to neutralize toxin 7
(data not shown). To demonstrate the specificity of the antibody-mediated neutralization, 8
antibodies were pre-incubated with either a mutated form of PA (contains a 2-amino acid 9
substitution in the receptor binding region of the molecule and is non-functional) or the 10
amino-terminal PA20 region of PA fused to GFP prior to their addition to the RAW assay. 11
This pre-incubation abrogated the neutralizing potential of the antibodies in an epitope-12
specific manner (Fig. 2) for most antibodies, demonstrating that the toxin neutralization 13
we observe arose from the PA-specific antibody binding to the PA in the assay. 14
Monoclonal 4A12, which is specific for an epitope associated with the Domain 4 region 15
(D4) of the monomer (16) was not inhibited by pre-incubation with PArb- in this assay. 16
The two mutations introduced into PA to render it non-functional for this assay were in 17
the D4 region, and likely contribute to 4A12 binding. Monoclonal antibody 25G9 also 18
does not inhibit as might be expected in this assay. This antibody did not capture PA20 19
from solution in the capture assay, and specificity was assigned based on its ability to 20
bind PA20 in a solid phase ELISA. It is possible that the relevant PA20-associated epitope 21
is not retained or efficiently displayed by this construct in the soluble phase required for 22
this assay. Overall, approximately 24 percent of the monoclonal antibodies in the panel 23
on August 18, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

were capable of neutralizing LT in the in vitro assay. The toxin-neutralizing potential of 1
each monoclonal antibody in the panel is shown in Table 1 and summarized in Table 2. 2
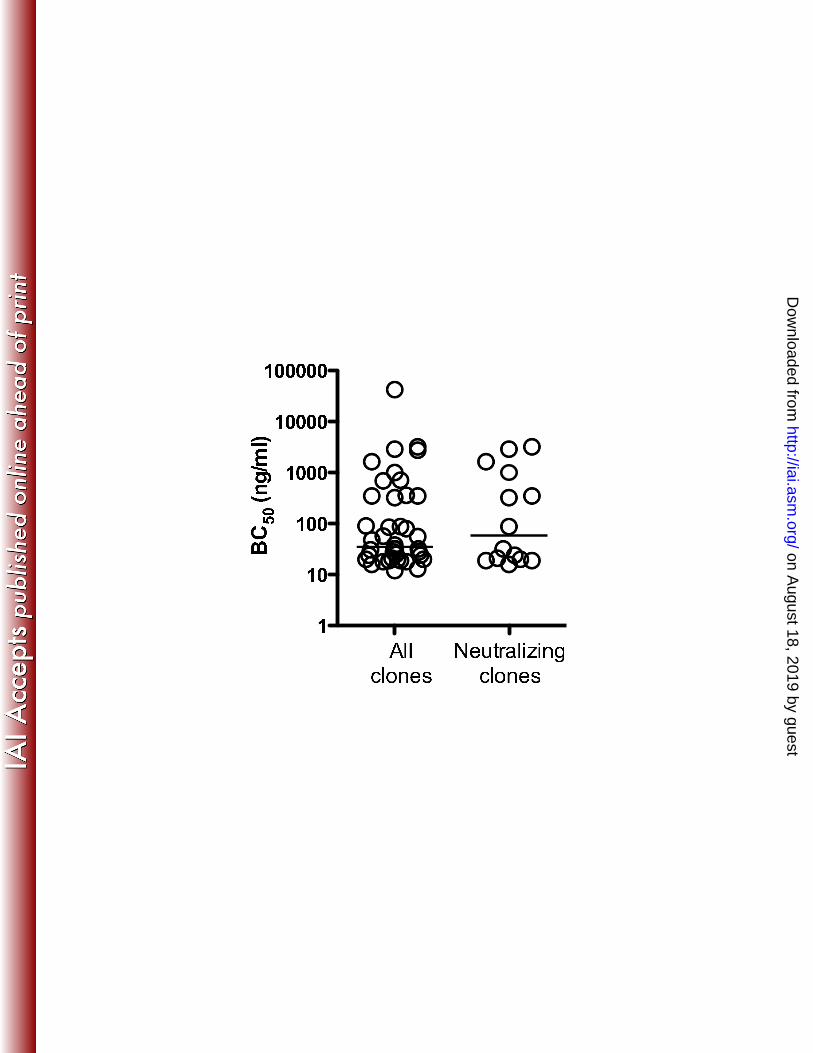
Neutralizing and non-neutralizing antibodies exhibit similar relative avidities for 3
PA: In addition to specificity, the affinity of the individual antibody combining sites for 4
the target antigen, and the region of the antigen bound (epitope) may determine the 5
ability of an individual toxin-specific antibody to block toxin function. To determine if 6
toxin neutralizing and non-neutralizing antibodies differed in their relative ability to bind 7
PA, monoclonal antibodies were titered in a PA-specific ELISA assay and the antibody 8
concentration resulting in 50% binding (BC50) ascertained. BC50 values for non-9
neutralizing antibodies varied from 12 to 42,500 ng/ml (median = 35 ng/ml), with the 10
majority of values falling between 16 and 3200 ng/ml. Neutralizing antibodies also varied 11
widely in their BC50 values (median = 60 ng/ml). When the relative avidities for the 12
neutralizing antibodies were compared to the overall antibody pool (Fig. 3), BC50 13
distributions for the two groups overlapped and no significant differences were observed 14
between antibodies capable of neutralizing toxin and those lacking efficacy in the 15
cytotoxicity assay (Mann-Whitney U test; p = 0.72). 16
Toxin-neutralizing antibodies are more frequent in the PA63-specific portion of the 17
antibody repertoire: We have previously demonstrated that the antibody response to 18
PA is highly biased towards epitopes present in the amino-terminal portion of the PA 19
monomer (PA20), with about 63% of the antibodies binding to this region of the molecule 20
(which represents only 25% of the PA monomer's mass) (16). PA20 is cleaved from the 21
remainder of the PA molecule rapidly following cell-surface binding and has no further 22
role in the intoxication process. It is possible that antibodies specific for epitopes located 23
on August 18, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

on this region of the molecule might be deficient in their ability to effect toxin 1
neutralization. When compared as a function of PA domain specificity (Table 1), toxin 2
neutralization was more frequently observed within the subset of antibodies specific for 3
epitopes associated with the PA63 region of the PA monomer. Seven of the 16 antibodies 4
(44%) specific for PA63-associated determinants (including those specific for domain 4) 5
were capable of neutralizing toxin. Seven of the 38 (18%) antibodies specific for PA20-6
associated determinants neutralized toxin in the in vitro RAW cell assay. PA63-specific 7
antibodies were therefore over twice as likely to neutralize lethal toxin as those specific 8
for PA20-related epitopes (p = 0.08; 2-sided Fishers exact test). 9
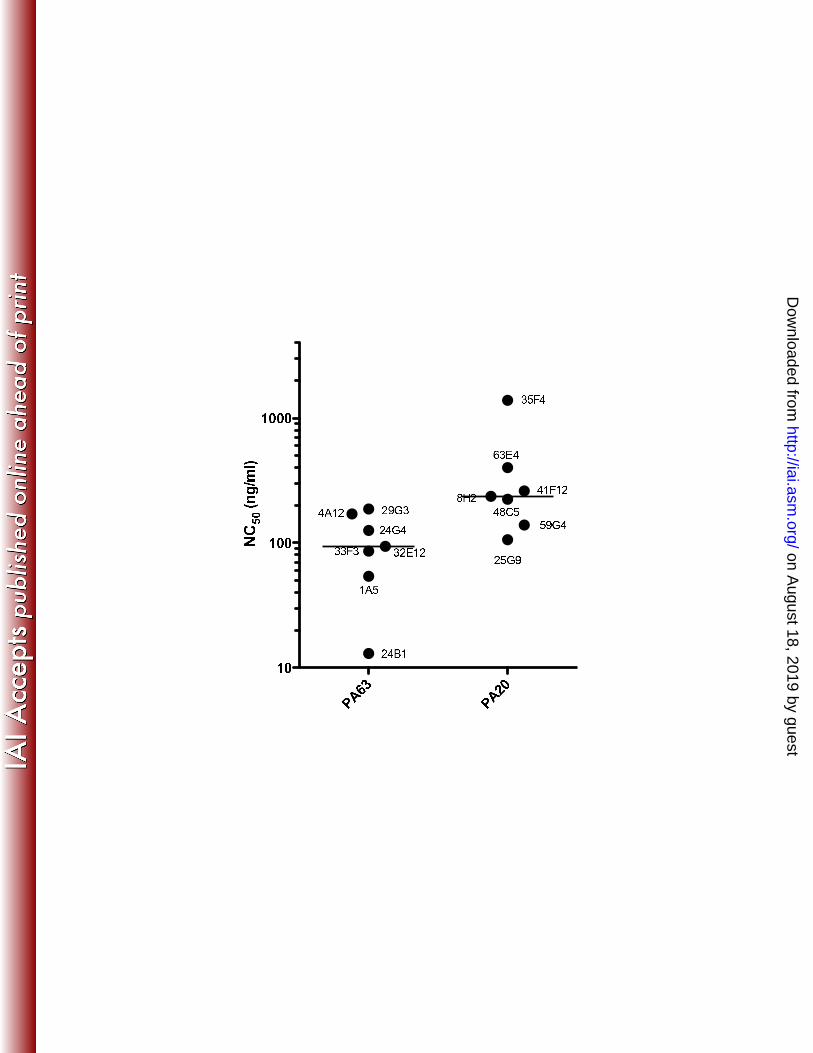
In addition to determining the relative frequency of toxin-neutralizing paratopes 10
within the two epitope-specific populations, we also sought to determine if the two 11
antibody populations varied in their relative efficacy in toxin neutralization. The 12
concentration of antibody resulting in 50% toxin neutralization (NC50) in the RAW cell 13
assay was determined for each antibody clone. NC50 values for the two groups were 14
compared to determine if this amount varied as a function of domain specificity. On 15
average, PA63-specific clones required about 3.8 fold less antibody to achieve 50% 16
neutralization when compared to clones specific for PA20-associated epitopes (Fig. 4). 17
Statistical comparison of the two groups indicated that PA63-specific antibodies were 18
significantly more efficient in neutralizing lethal toxin (p = 0.01; Mann-Whitney U test) 19
than PA20-specific antibodies. 20
on August 18, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

Discussion 1
The human antibody response to a large protein antigen such as PA is complex. 2
Exposure to such a molecule activates a large collection of B cells, each of which 3
produces an antibody specific for an individual antigenic epitope associated with the 4
immunogen. Most, if not all, of these antibodies undergo further somatic hypermutation 5
giving rise to a diverse assemblage of antibody paratopes, each characterized by the 3-6
dimentional structure on the target molecule it recognizes (its epitope), and the strength 7
with which it binds that structure (its affinity). For antibodies that bind determinants 8
physically associated with a pathogen (such as a bacterial cell wall), the functionality of 9
the antibody is often determined by the constant region of the antibody molecule. 10
Complement fixation and opsonization, for example, are facilitated by the constant 11
region, and are the primary means by which antibody mediates the destruction of many 12
pathogens. The mechanism by which antibodies neutralize soluble agents such as toxins 13
is not well understood. The findings we present here demonstrate that avid antibody 14
binding alone is insufficient to block PA’s role in intoxication, and suggests that the 15
antigenic epitope recognized is the primary determining factor in antibody function. 16
Given the complexity of the role PA plays in anthrax intoxication, the finding that 17
the majority of PA-specific antibodies in vaccinated individuals do not neutralize lethal 18
toxin is unexpected. For PA-mediated cell death to occur in the in vitro assay employed 19
in this study (and presumably in vivo as well), PA must bind to the cell surface receptor 20
and be cleaved by furin to yield cell-associated PA63. Cell bound PA63 must then form 21
homo-heptamers, the PA63 heptamers must bind LF, and the PA/LF complex must be 22
internalized and released into the cytosol. Our data demonstrates that this complicated 23
on August 18, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

chain of events can proceed unimpeded in the presence of antibody bound to each of the 1
participating PA monomers. The predominance of PA20-specific paratopes in the 2
response may offer a partial explanation of the phenomenon in serum. Antibodies 3
binding the PA20 region of the molecule would be detached from cell bound PA83 (along 4
with the PA20 fragment) following furin cleavage. Additionally, as free PA20 5
accumulates in the culture supernatant (or in the serum (13)), this proteolytic fragment 6
could compete for antibody binding with those PA20-associated epitopes still associated 7
with intact PA83. Nine of the sixteen PA63-specific antibodies we assayed were also non-8
neutralizing. It remains unclear how PA retains its functionality when complexed with 9
these antibodies. 10
While we believe the collection of antibodies we isolated to be representative of the 11
overall response, it is not complete, and further investigations of these same donors 12
would undoubtedly uncover additional PA-specific paratopes. Serum from donor 3, for 13
example, exhibited very low, but nevertheless above background, toxin neutralization 14
activity when tested at low dilutions. None of the antibodies we isolated from this 15
individual neutralized toxin. This could result from either insufficient sampling or the 16
fact that the respective B cells were not circulating at the time blood was collected. As a 17
group, the antibody panel represents a wide range of relative avidities, recognizes 18
epitopes distributed throughout the PA monomer, and has both neutralizing and non-19
neutralizing members. These factors suggest no obvious sampling bias, and that the 20
paratope distribution we observe reflects the paratope distribution present during the 21
ongoing immune response in a vaccinated individual. 22
Establishing the relationship between antibody paratope, antigenic epitope, and 23
on August 18, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

antibody function is crucial to the understanding of how toxin-based vaccines give rise to 1
efficacious antibody responses. If binding alone is insufficient, it is a reasonable 2
assumption that antibody binding must disrupt an essential toxin function in order for that 3
antibody to be effective. It has been postulated, for example, that the primary mechanism 4
of action by anti-PA antibodies would be the blockade of binding to the cell surface 5
receptor, and vaccine formulations based solely on the primary receptor binding domain 6
(domain 4) of PA have been proposed (5, 7). Our findings suggest that these assumptions 7
are premature. Only one of the neutralizing antibodies we isolated (4A12) reacts with the 8
domain 4 region of the molecule, and it is unlikely that any of the PA20-specific 9
antibodies interfere with receptor binding. Residues associated with heptamer formation, 10
LF/EF binding, or furin cleavage could also give rise to antigenic epitopes, and 11
antibodies recognizing these epitopes might also be effective in neutralizing toxin. We 12
have initiated studies to determine the mechanism by which each of the antibodies we 13
isolated neutralize PA-mediated toxicity in order to determine if they function through 14
any of these modalities. 15
Although a subset of PA20-speicifc antibodies are capable of neutralizing PA-16
mediated toxicity, their dominance in the response following vaccination may 17
nevertheless have negative implications for the protective efficacy of PA-based anthrax 18
vaccines. Only 18% of these antibodies neutralize the toxin, and they are less efficient, 19
requiring a higher concentration to achieve neutralization. And, in vivo, these antibodies 20
may be effectively blocked by free PA20. In comparison, the population of PA63-specific 21
paratopes contains a higher ratio of neutralizing specificities, and cell-free PA83 or PA63 22
encountered in serum would not diminish their effectiveness. 23
on August 18, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

Although other anthrax-derived antigens have been shown to elicit protective 1
immune responses and have been proposed for inclusion in new vaccine formulations, the 2
second-generation anthrax vaccines currently under development are based solely on PA 3
(10). The findings presented here, when considered along with our previous 4
demonstration of a profound domain bias in the PA-specific response towards PA20-5
specific epitopes in vaccinated humans (16), suggest that factors intrinsic to the 6
immunobiology of PA itself may diminish its effectiveness in inducing toxin-neutralizing 7
antibodies. The mechanisms underlying the domain bias of the antibody response to PA 8
remain unknown. We have postulated (16) that differential antigen processing of free 9
PA20 and cell bound PA63 may give rise to a preponderance of PA20-specific antibodies 10
following vaccination, and suggested that sequence alterations in the furin recognition 11
sequence that render PA protease resistant might produce a more immunogenic form of 12
the PA monomer. Such a design strategy aimed at shifting epitope dominance towards 13
neutralizing determinants might result in a more efficacious vaccine for the prevention of 14
anthrax infection. 15
Acknowledgments: The authors gratefully acknowledge Nanette Bond, PA-C for 16
assistance with sample collection, and Betty M. Ho for critically reading the manuscript. 17
This work was supported by Public Health Service Grants AI57932 and AI066508 from 18
the National Institute of Allergy and Infectious Diseases. This research was conducted in 19
a facility constructed with support from Research Facilities Improvement Program Grant 20
Number C06 RR-16226 from the National Center for Research Resources, NIH. 21
22
on August 18, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

Figure legends 1
Figure 1. Neutralization of anthrax lethal toxin by human PA-specific monoclonal 2
antibodies: Titered amounts of CHO-derived antibodies specific for PA were pre-3
incubated with fixed amounts of PA and LF prior to their addition to cultures of RAW 4
264.7 cells. Cell viability was determined approximately 16 hours later, and the degree 5
to which the added antibody blocked toxicity determined. The domain specificity (PA20, 6
PA63, or Domain 4) is indicated for each clone. 7
Figure 2. Domain-specific inhibition of antibody-mediated toxin neutralization by 8
pre-incubation with PA or its amino-terminal fragment: Toxin neutralizing 9
antibodies were pre-incubated with a receptor-deficient mutant of PA (PArb-) or the 10
amino-terminal 191 residues of PA expressed as a GFP fusion protein (PA20-GFP) prior 11
to their addition to the RAW267.4 cytotoxicity assay. Open bars represent the degree of 12
inhibition observed when antibodies were pre-incubated with PArb-. Filled bars 13
represent inhibition by PA20-GFP. Antibodies are grouped by their domain specificity. 14
Figure 3. Relative binding avidity of toxin neutralizing and non-neutralizing PA-15
specific monoclonal antibodies: CHO-derived human antibodies specific for PA were 16
titrated in an ELISA assay and the amount required to achieve 50% binding determined. 17
The median value for each group is indicated by a horizontal line. The two groups did 18
not differ significantly in their relative avidity for PA (p = 0.72; Mann-Whitney U test). 19
Figure 4. Relative toxin neutralizing ability of PA63- or PA20-specific antibodies: 20
CHO-derived human antibodies specific for PA were titrated in the RAW 264.7 21
cytotoxicity assay and the amount required to bring about 50% cell death determined. 22
The median value for each group is indicated by a horizontal line. PA63- and PA20-23
on August 18, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

specific antibodies differed significantly in the concentration required to effect 50% 1
neutralization of lethal toxin (p = 0.01; Mann-Whitney U test). 2
on August 18, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

Table footnotes 1
Table 1. Characteristics of the 58 human PA-specific monoclonal antibodies examined 2
in this study. aHeavy chain variable region gene utilized. bLight chain variable region 3
gene utilized. cHeavy chain joining region gene utilized. dAmino acid sequence of the 4
heavy chain third complementarity determining region. ePA domain specificity of the 5
monoclonal antibody (ND = not resolved). fAbility of the antibody to neutralize PA-6
mediated cytoxicity. 7
8
Table 2. Summary of data presented in Table 1 describing the PA-specific monoclonal 9
antibodies examined in this study. aNumber of sequence-unique monoclonal antibodies 10
isolated from each donor. bNumber of unique B-cell rearrangement events represented 11
by the antibodies isolated from each donor. c,d,e,fNumber of antibodies with specificity 12
for each PA domain (ND = not resolved). gNumber of antibodies that neutralize PA-13
mediated cytoxicity. 14
15
16
on August 18, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

Literature cited 1
2
1. Ahuja, N., P. Kumar, and R. Bhatnagar. 2001. Hydrophobic residues Phe552, 3
Phe554, Ile562, Leu566, and Ile574 are required for oligomerization of anthrax 4
protective antigen. Biochem Biophys Res Commun 287:542-549. 5
2. Altschul, S. F., T. L. Madden, A. A. Schaffer, J. Zhang, Z. Zhang, W. Miller, 6
and D. J. Lipman. 1997. Gapped BLAST and PSI-BLAST: a new generation of 7
protein database search programs. Nucleic Acids Res 25:3389-3402. 8
3. Bhatnagar, R., and S. Batra. 2001. Anthrax toxin. Crit Rev Microbiol 27:167-9
200. 10
4. Bradley, K. A., J. Mogridge, M. Mourez, R. J. Collier, and J. A. Young. 2001. 11
Identification of the cellular receptor for anthrax toxin. Nature 414:225-229. 12
5. Chichester, J. A., K. Musiychuk, P. de la Rosa, A. Horsey, N. Stevenson, N. 13
Ugulava, S. Rabindran, G. A. Palmer, V. Mett, and V. Yusibov. 2007. 14
Immunogenicity of a subunit vaccine against Bacillus anthracis. Vaccine 15
25:3111-3114. 16
6. Cunningham, K., D. B. Lacy, J. Mogridge, and R. J. Collier. 2002. Mapping 17
the lethal factor and edema factor binding sites on oligomeric anthrax protective 18
antigen. Proc Natl Acad Sci U S A 99:7049-7053. 19
7. Flick-Smith, H. C., N. J. Walker, P. Gibson, H. Bullifent, S. Hayward, J. 20
Miller, R. W. Titball, and E. D. Williamson. 2002. A recombinant carboxy-21
terminal domain of the protective antigen of Bacillus anthracis protects mice 22
against anthrax infection. Infect Immun 70:1653-1656. 23
on August 18, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

8. Ivins, B. E., S. L. Welkos, S. F. Little, M. H. Crumrine, and G. O. Nelson. 1
1992. Immunization against anthrax with Bacillus anthracis protective antigen 2
combined with adjuvants. Infect Immun 60:662-668. 3
9. Kabat, E. A., T. T. Wu, H. M. Perry, K. S. Gottesman, and C. Foeller. 1991. 4
Sequences of Proteins of Immunological Interest, 5 ed, vol. 2. U.S. Department of 5
Health and Human Services, Bethesday. 6
10. Keitel, W. A. 2006. Recombinant protective antigen 102 (rPA102): profile of a 7
second-generation anthrax vaccine. Expert Rev Vaccines 5:417-430. 8
11. Lefranc, M. P. 2004. IMGT-ONTOLOGY and IMGT databases, tools and Web 9
resources for immunogenetics and immunoinformatics. Mol Immunol 40:647-10
660. 11
12. McBride, B. W., A. Mogg, J. L. Telfer, M. S. Lever, J. Miller, P. C. Turnbull, 12
and L. Baillie. 1998. Protective efficacy of a recombinant protective antigen 13
against Bacillus anthracis challenge and assessment of immunological markers. 14
Vaccine 16:810-817. 15
13. Moayeri, M., J. F. Wiggins, and S. H. Leppla. 2007. Anthrax protective antigen 16
cleavage and clearance from the blood of mice and rats. Infect Immun 75:5175-17
5184. 18
14. Mock, M., and A. Fouet. 2001. Anthrax. Annu Rev Microbiol 55:647-671. 19
15. Petosa, C., R. J. Collier, K. R. Klimpel, S. H. Leppla, and R. C. Liddington. 20
1997. Crystal structure of the anthrax toxin protective antigen. Nature 385:833-21
838. 22
on August 18, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

16. Reason, D. C., A. Ullal, J. Liberato, J. Sun, W. Keitel, and J. Zhou. 2008. 1
Domain specificity of the human antibody response to Bacillus anthracis 2
protective antigen. Vaccine 26:4041-4047. 3
17. Reason, D. C., and J. Zhou. 2004. Correlation of antigenic epitope and antibody 4
gene usage in the human immune response to Streptococcus pneumoniae type 23F 5
capsular polysaccharide. Clin Immunol 111:132-136. 6
18. Rosovitz, M. J., P. Schuck, M. Varughese, A. P. Chopra, V. Mehra, Y. Singh, 7
L. M. McGinnis, and S. H. Leppla. 2003. Alanine-scanning mutations in 8
domain 4 of anthrax toxin protective antigen reveal residues important for binding 9
to the cellular receptor and to a neutralizing monoclonal antibody. J Biol Chem 10
278:30936-30944. 11
19. Santelli, E., L. A. Bankston, S. H. Leppla, and R. C. Liddington. 2004. Crystal 12
structure of a complex between anthrax toxin and its host cell receptor. Nature 13
430:905-908. 14
20. Scobie, H. M., G. J. Rainey, K. A. Bradley, and J. A. Young. 2003. Human 15
capillary morphogenesis protein 2 functions as an anthrax toxin receptor. Proc 16
Natl Acad Sci U S A 100:5170-5174. 17
21. Zhou, J., and D. C. Reason. 2007. The human antibody repertoire specific for 18
the capsular polysaccharide of Streptococcus pneumoniae serotype 14. in prep. 19
22. Zhou, J., A. Ullal, J. Liberato, J. Sun, W. Keitel, and D. C. Reason. 2008. 20
Paratope diversity in the human antibody response to Bacillus anthracis 21
protective antigen. Mol Immunol 45:338-347. 22
23
on August 18, 2019 by guest
http://iai.asm.org/
Dow
nloaded from
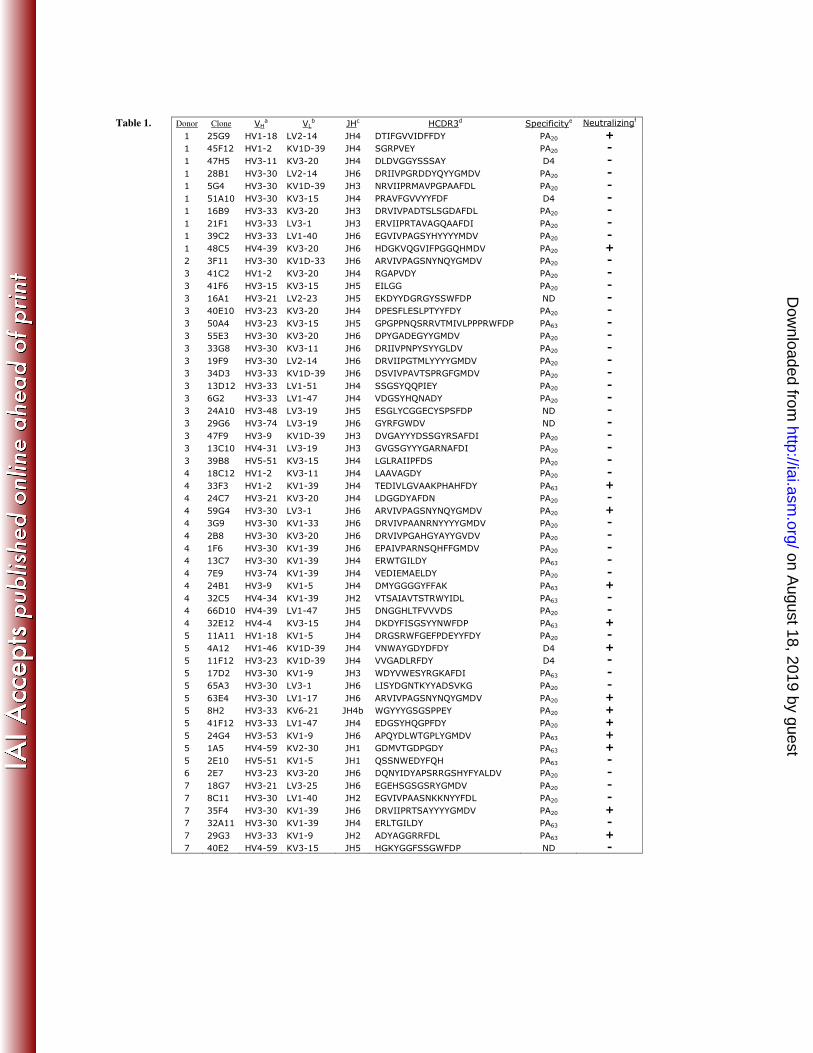
Table 1.
Donor Clone VHa VL
b JHc HCDR3d Specificitye Neutralizingf
1 25G9 HV1-18 LV2-14 JH4 DTIFGVVIDFFDY PA20 +
1 45F12 HV1-2 KV1D-39 JH4 SGRPVEY PA20 -
1 47H5 HV3-11 KV3-20 JH4 DLDVGGYSSSAY D4 -
1 28B1 HV3-30 LV2-14 JH6 DRIIVPGRDDYQYYGMDV PA20 -
1 5G4 HV3-30 KV1D-39 JH3 NRVIIPRMAVPGPAAFDL PA20 -
1 51A10 HV3-30 KV3-15 JH4 PRAVFGVVYYFDF D4 -
1 16B9 HV3-33 KV3-20 JH3 DRVIVPADTSLSGDAFDL PA20 -
1 21F1 HV3-33 LV3-1 JH3 ERVIIPRTAVAGQAAFDI PA20 -
1 39C2 HV3-33 LV1-40 JH6 EGVIVPAGSYHYYYYMDV PA20 -
1 48C5 HV4-39 KV3-20 JH6 HDGKVQGVIFPGGQHMDV PA20 +
2 3F11 HV3-30 KV1D-33 JH6 ARVIVPAGSNYNQYGMDV PA20 -
3 41C2 HV1-2 KV3-20 JH4 RGAPVDY PA20 -
3 41F6 HV3-15 KV3-15 JH5 EILGG PA20 -
3 16A1 HV3-21 LV2-23 JH5 EKDYYDGRGYSSWFDP ND -
3 40E10 HV3-23 KV3-20 JH4 DPESFLESLPTYYFDY PA20 -
3 50A4 HV3-23 KV3-15 JH5 GPGPPNQSRRVTMIVLPPPRWFDP PA63 -
3 55E3 HV3-30 KV3-20 JH6 DPYGADEGYYGMDV PA20 -
3 33G8 HV3-30 KV3-11 JH6 DRIIVPNPYSYYGLDV PA20 -
3 19F9 HV3-30 LV2-14 JH6 DRVIIPGTMLYYYYGMDV PA20 -
3 34D3 HV3-33 KV1D-39 JH6 DSVIVPAVTSPRGFGMDV PA20 -
3 13D12 HV3-33 LV1-51 JH4 SSGSYQQPIEY PA20 -
3 6G2 HV3-33 LV1-47 JH4 VDGSYHQNADY PA20 -
3 24A10 HV3-48 LV3-19 JH5 ESGLYCGGECYSPSFDP ND -
3 29G6 HV3-74 LV3-19 JH6 GYRFGWDV ND -
3 47F9 HV3-9 KV1D-39 JH3 DVGAYYYDSSGYRSAFDI PA20 -
3 13C10 HV4-31 LV3-19 JH3 GVGSGYYYGARNAFDI PA20 -
3 39B8 HV5-51 KV3-15 JH4 LGLRAIIPFDS PA20 -
4 18C12 HV1-2 KV3-11 JH4 LAAVAGDY PA20 -
4 33F3 HV1-2 KV1-39 JH4 TEDIVLGVAAKPHAHFDY PA63 +
4 24C7 HV3-21 KV3-20 JH4 LDGGDYAFDN PA20 -
4 59G4 HV3-30 LV3-1 JH6 ARVIVPAGSNYNQYGMDV PA20 +
4 3G9 HV3-30 KV1-33 JH6 DRVIVPAANRNYYYYGMDV PA20 -
4 2B8 HV3-30 KV3-20 JH6 DRVIVPGAHGYAYYGVDV PA20 -
4 1F6 HV3-30 KV1-39 JH6 EPAIVPARNSQHFFGMDV PA20 -
4 13C7 HV3-30 KV1-39 JH4 ERWTGILDY PA63 -
4 7E9 HV3-74 KV1-39 JH4 VEDIEMAELDY PA20 -
4 24B1 HV3-9 KV1-5 JH4 DMYGGGGYFFAK PA63 +
4 32C5 HV4-34 KV1-39 JH2 VTSAIAVTSTRWYIDL PA63 -
4 66D10 HV4-39 LV1-47 JH5 DNGGHLTFVVVDS PA20 -
4 32E12 HV4-4 KV3-15 JH4 DKDYFISGSYYNWFDP PA63 +
5 11A11 HV1-18 KV1-5 JH4 DRGSRWFGEFPDEYYFDY PA20 -
5 4A12 HV1-46 KV1D-39 JH4 VNWAYGDYDFDY D4 +
5 11F12 HV3-23 KV1D-39 JH4 VVGADLRFDY D4 -
5 17D2 HV3-30 KV1-9 JH3 WDYVWESYRGKAFDI PA63 -
5 65A3 HV3-30 LV3-1 JH6 LISYDGNTKYYADSVKG PA20 -
5 63E4 HV3-30 LV1-17 JH6 ARVIVPAGSNYNQYGMDV PA20 +
5 8H2 HV3-33 KV6-21 JH4b WGYYYGSGSPPEY PA20 +
5 41F12 HV3-33 LV1-47 JH4 EDGSYHQGPFDY PA20 +
5 24G4 HV3-53 KV1-9 JH6 APQYDLWTGPLYGMDV PA63 +
5 1A5 HV4-59 KV2-30 JH1 GDMVTGDPGDY PA63 +
5 2E10 HV5-51 KV1-5 JH1 QSSNWEDYFQH PA63 -
6 2E7 HV3-23 KV3-20 JH6 DQNYIDYAPSRRGSHYFYALDV PA20 -
7 18G7 HV3-21 LV3-25 JH6 EGEHSGSGSRYGMDV PA20 -
7 8C11 HV3-30 LV1-40 JH2 EGVIVPAASNKKNYYFDL PA20 -
7 35F4 HV3-30 KV1-39 JH6 DRVIIPRTSAYYYYGMDV PA20 +
7 32A11 HV3-30 KV1-39 JH4 ERLTGILDY PA63 -
7 29G3 HV3-33 KV1-9 JH2 ADYAGGRRFDL PA63 +
7 40E2 HV4-59 KV3-15 JH5 HGKYGGFSSGWFDP ND -
on August 18, 2019 by guest
http://iai.asm.org/
Dow
nloaded from
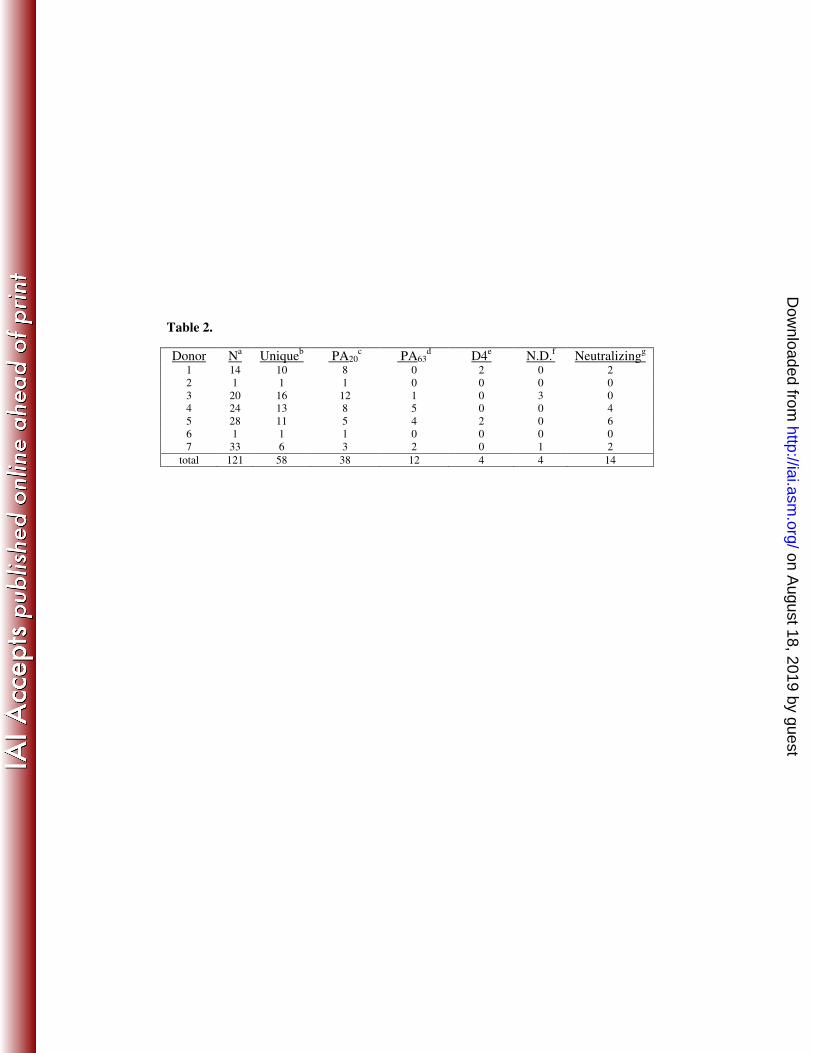
Table 2.
Donor Na Unique
b PA20
c PA63
d D4
e N.D.
f Neutralizing
g
1 14 10 8 0 2 0 2
2 1 1 1 0 0 0 0
3 20 16 12 1 0 3 0
4 24 13 8 5 0 0 4
5 28 11 5 4 2 0 6
6 1 1 1 0 0 0 0
7 33 6 3 2 0 1 2
total 121 58 38 12 4 4 14
on August 18, 2019 by guest
http://iai.asm.org/
Dow
nloaded from
1 1 0 1 0 0 1 0 0 005 01 0 0 1 A 5( P A 6 3 )A n t i b o d y ( n g / m l )%N eut rali zati on 1 1 0 1 0 0 1 0 0 005 01 0 0 2 9 G 3( P A 6 3 )A n t i b o d y ( n g / m l )%N eut rali zati on 1 1 0 1 0 0 1 0 0 005 01 0 0 4 A 1 2( D 4 )A n t i b o d y ( n g / m l )%N eut rali zati on
1 1 0 1 0 0 1 0 0 005 01 0 0 2 4 B 1( P A 6 3 )A n t i b o d y ( n g / m l )%N eut rali zati on1 1 0 1 0 0 1 0 0 005 01 0 0 3 2 E 1 2( P A 6 3 )A n t i b o d y ( n g / m l )%N eut rali zati on 1 1 0 1 0 0 1 0 0 005 01 0 0 6 3 E 4( P A 2 0 )A n t i b o d y ( n g / m l )%N eut rali zati on
on August 18, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

2 4 B 1 3 2 E 1 2 1 A 5 4 A 1 2 2 4 G 4 2 9 G 3 2 5 G 9 4 8 C 5 3 5 F 4 4 1 F 1 2 6 3 E 405 01 0 0P A 2 0 � s p e c i f i ca n t i b o d i e sP A 6 3 � s p e c i f i ca n t i b o d i e s%I nhibi ti on
on August 18, 2019 by guest
http://iai.asm.org/
Dow
nloaded from
11 01 0 01 0 0 01 0 0 0 01 0 0 0 0 0A l lc l o n e s N e u t r a l i z i n gc l o n e sBC 50( ng/ ml)
on August 18, 2019 by guest
http://iai.asm.org/
Dow
nloaded from
P A 6 3 P A 2 01 01 0 01 0 0 02 4 B 11 A 5 2 5 G 93 3 F 3 6 3 E 44 8 C 52 4 G 43 2 E 1 24 A 1 2 4 1 F 1 25 9 G 42 9 G 3 8 H 2 3 5 F 4NC 50( ng/ ml)
on August 18, 2019 by guest
http://iai.asm.org/
Dow
nloaded from





























