Embed Size (px)
Citation preview

Cell Cycle and Senescence
Dynamic and Nuclear Expression of PDGFRa and IGF-1R inAlveolar Rhabdomyosarcoma
M. Imran Aslam1,2,12, Simone Hettmer7,8,9,10,12, Jinu Abraham2, Dorian LaTocha1, Anuradha Soundararajan5,Elaine T. Huang2, Martin W. Goros6, Joel E. Michalek10, Shuyu Wang11, Atiya Mansoor3, Brian J. Druker1,4,Amy J. Wagers7,8,9,12, Jeffrey W. Tyner1, and Charles Keller2
AbstractSince the advent of tyrosine kinase inhibitors as targeted therapies in cancer, several receptor tyrosine kinases
(RTK) have been identified as operationally important for disease progression. Rhabdomyosarcoma (RMS) is amalignancy in need of new treatment options; therefore, better understanding of the heterogeneity of RTKs wouldadvance this goal. Here, alveolar RMS (aRMS) tumor cells derived from a transgenic mouse model expressing twosuch RTKs, platelet-derived growth factor (PDGFR)a and insulin-like growth factor (IGF)-1R, were investigatedby fluorescence-activated cell sorting (FACS). Sorted subpopulations that were positive or negative for PDGFRaand IGF-1R dynamically altered their cell surface RTK expression profiles as early as the first cell division.Interestingly, a difference in total PDGFRa expression and nuclear IGF-1R expression was conserved inpopulations. Nuclear IGF-1R expression was greater than cytoplasmic IGF-1R in cells with initially high cellsurface IGF-1R, and cells with high nuclear IGF-1R established tumorsmore efficiently in vivo. RNA interference–mediated silencing of IGF-1R in the subpopulation of cells initially harboring higher cell surface and total IGF-1Rresulted in significantly reduced anchorage-independent colony formation as compared with cells with initiallylower cell surface and total IGF-1R expression. Finally, in accordance with the findings observed in murine aRMS,human aRMS also had robust expression of nuclear IGF-1R.
Implications: RTK expression status and subcellular localization dynamics are important considerations forpersonalized medicine. Mol Cancer Res; 11(11); 1303–13. �2013 AACR.
IntroductionRhabdomyosarcoma is themost frequent soft tissue sarcoma
of childhood. Of the common subtypes for this aggressive
muscle cancer, patients with alveolar rhabdomyosarcoma havethe most dismal outcome despite maximal therapeutic inter-vention (1–3). We have developed and validated a geneticallyengineered, conditional mouse model of alveolar rhabdomyo-sarcoma (4, 5) and therein identified the receptor tyrosinekinase (RTK) targets platelet-derived growth factor receptor-a(PDGFRa) and insulin-like growth factor-1 receptor (IGF-1R) as potential therapeutic targets (6, 7). In agreement withour mouse model studies, careful work by other groups hasascribed biologic and clinical significance to the expression ofthese and other RTKs for alveolar rhabdomyosarcoma (8, 9).In the current era of biomarker-stratified and personalized
cancer therapy clinical trials, great importance is beingascribed to biomarker expression from the solitary biopsysample taken from each patient at a single time point (10).However, intra and intertumor heterogeneity for individualpatients is increasingly recognized (11). Several models devel-oped to interpret the heterogeneity of individual cells thatcompose a tumor have been proposed, with strong evidencethat solid tumors consist of subsets of cells distinct at thegenetic and phenotypic level (11–15). However, the conceptof isolating unique populations of cells by expression ofsurface membrane proteins lends itself toward controversybecause cells reverting to a state reflecting the heterogeneity ofthe original tumor have been described as well (16). Thus, theability to profile cell populations using cell surface markers,
Authors' Affiliations: 1Division of Hematology and Medical Oncology,Knight Cancer Institute; 2Department of Pediatrics, Pediatric Cancer BiologyProgram, Pap�e Family Pediatric Research Institute; 3Department of Pathol-ogy,OregonHealth& ScienceUniversity; 4HowardHughesMedical Institute,Portland, Oregon; 5Greehey Children's Cancer Research Institute; and6Department of Epidemiology & Biostatistics, University of Texas HealthScience Center, San Antonio, Texas; 7Department of Stem Cell and Regen-erative Biology, Harvard University; 8Harvard Stem Cell Institute; 9JoslinDiabetes Center, Cambridge; 10Division of Pediatric Hematology/Oncology,Department of Pediatric Oncology, Dana-Farber Cancer Institute, Children'sHospital; and 11Harvard University, Cambridge, MA; 12Howard HughesMedical Institute
Note: Supplementary data for this article are available at MolecularCancer Research Online (http://mcr.aacrjournals.org/).
M.I. Aslam and S. Hettmer contributed equally to this work.
Current address for M.I. Aslam: Howard Hughes Medical Institute MedicalResearch Fellows Program, 4000 Jones Bridge Rd., ChevyChase, MD 20815
Corresponding Authors: Charles Keller, Oregon Health & Science Uni-versity, 3181 SW Sam Jackson Park Rd., MC-L321, Portland, OR 97239.Phone: 503-494-1210; Fax: 503-418-5044; E-mail: [email protected];Jeffrey W. Tyner, E-mail: [email protected]; and Amy J. Wagers, E-mail:[email protected]
doi: 10.1158/1541-7786.MCR-12-0598
�2013 American Association for Cancer Research.
MolecularCancer
Research
www.aacrjournals.org 1303
on August 28, 2018. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst August 8, 2013; DOI: 10.1158/1541-7786.MCR-12-0598

particularly RTKs expressed by only a subpopulation oftumor cells, might be informative if RTK expression is stable.Otherwise, understanding mechanisms that dynamicallymodulate subcellular localization of RTKs is necessary(17–23). In this regard, the mechanism of intracellular(nuclear) trafficking of RTKs has been described with mostdetails for the EGF receptor (EGFR; refs. 18, 20, 21). OtherRTKs, such as FGFR, IGF-1R, and ErbB4 have also beenshown to localize to the cell nucleus in cancer (17, 19, 22).Because PDGFRa and IGF-1R are reported to act in
concert for resistance to IGF-1R small-molecule inhibitionin rhabdomyosarcoma (24), we sought to understandwheth-er tumor cell subpopulations can be subdivided into distinctRTK profiles and whether isolated cells behave as pheno-typically static or dynamic populations. Furthermore, weaimed to determine whether differential subcellular locali-zation of these RTKs had a biologically significant role intumor establishment.
Materials and MethodsCell cultureMouse tumor cell lines were established and cultured
previously described (7). Cells were grown in Dulbecco'smodified Eagle medium (DMEM) supplemented with 10%FBS and penicillin (100 U/mL)/streptomycin (100 mg/mL;Invitrogen) in 5% CO2 in air at 37�C.
Flow cytometryMurine alveolar rhabdomyosarcoma cells were grown to
approximately 50% confluency then trypsinized, washedwith PBS, and stained with primary APC-conjugated anti-body to PDGFRa (1:40, eBiosciences) in fluorescence-acti-vated cell sorting (FACS) buffer (2% FBS in PBS) on ice for30 minutes, washed in FACS buffer, then stained with IGF-1R (1:100, ab32823, Abcam) primary antibody for 30minutes on ice. Cells were again washed in FACS buffer,then stained with anti-chicken secondary antibody conjugat-ed to Cy3.5 (1:80, ab97146, Abcam) for 30 minutes on ice.Cells were washed in FACS buffer and resuspended in bufferand analyzed by a BD FACSAria III Cell Sorter (BD Bio-sciences). As a control, cells were stained with isotypeantibody conjugated to APC at the same concentration asPDGFRa antibody (#17-1401-81, eBiosciences) and subse-quentlywith secondary antibody conjugated toCy3.5 only, atthe aforementioned concentration. Stained cells were gatedaccording to control and sorted by quadrant for PDGFRaand IGF-1R expression. Once cells were sorted, reanalysis byFACS was done to assess purity of sorted populations beforecells were recounted and allowed to recover in culture beforefurther experiments were carried out. Analysis of .fcs files wasdone with FlowJo (Tree Star, Inc., PC version 7.6).
Immunoblotting and immunoprecipitationCell lysates were prepared using Cell Lysis Buffer (#9803,
Cell Signaling Technology) supplemented with the appro-priate protease and phosphatase inhibitors (Sigma-Aldrich),and standard immunoblotting procedures were subsequent-
ly conducted and proteins visualized with chemiluminescentsignal using Super Signal Western Pico or Dura Substrate(Pierce Biotechnology). Mouse tumors were washed in PBS,and then lysed in radioimmunoprecipitation assay buffersupplemented with protease and phosphatase inhibitors.Lysates were homogenized and centrifuged at maximumspeed for 10 minutes at 4�C and supernatant used forWestern blotting. Antibodies used for immunoblottingwere PDGFRa (1:1000, #3164, Cell Signaling Technolo-gies), IGF-1Rb (1:1000, sc-713, Santa Cruz Biotechnolo-gy), p-IGF-1Rb recognizing phospho-Tyr 1161 (1:1000, sc-101703, Santa Cruz Biotechnology), phospho and totalp42/44 MAP K from Cell Signaling Technology (used at1:1000), b-actin (1:10,000, Sigma), a-tubulin (1:10,000,Sigma), and Sp1 (1:1000, Millipore) and pan-Tyr antibody(1:10,000, 4G-10, Upstate Biotechnology). Immunopre-cipitation was conducted by standard protocol using ProteinA beads for rabbit immunoglobulin G (IgG; Invitrogen).
Cytosolic and nuclear fractionationThe nuclear fraction of cells was isolated by the following
protocol: cells were lifted from a 100-mm culture dish andwashed with PBS once. Cell pellet was resuspended in 600mL buffer 1 (25 mmol/L HEPES, pH 7.9, 5 mmol/L KCl,0.5 mmol/L MgCl2, 1 mmol/L dithiothreitol, 100 mmol/Lphenylmethylsulfonylfluoride in H2O), then 600 mL ofbuffer 2 was added (buffer 1þ 1%Nonidet-P40), and thencentrifuged at 2,600 rpm for 5 minutes. The supernatant(cytoplasmic fraction) was transferred to a new tube. Hun-dredmicroliter mL of buffer 3 (1:1mix of buffer 1 and 2) wasadded to the cell pellet and gently mixed, centrifuged at2,600 rpm, and supernatant added to cytoplasmic fraction.Cell pellet was then washed two to three times with buffer 1and supernatant discarded for purity of subsequent solublenuclear protein extraction. Five hundred microliter of buffer4 (25 mmol/L HEPES, pH 7.9, 360 mmol/L NaCl, 10%dextrose, 0.05% Nonidet-P40) was added to cell pellet andtube rotated for 1 hour at 4�C. Tube was centrifuged at13,200 rpm for 10minutes and supernatant (soluble nuclearfraction) was transferred to a new tube. At this point, boththe cytoplasmic and nuclear fractions were clarified again bycentrifuged at 13,200 rpm and transferring supernatant to anew tube. Protein was quantified using BCA Protein AssayReagent (Pierce) and immunoblotting was conducted asmentioned above.
RNA interference mediated silencing of IGF-1R andanchorage-independent colony formation assayFor RNA interference (RNAi) studies, siRNA targeted to
IGF-1R (#SI01074017) and nonspecific siRNA (#1027280)were purchased fromQiagen. Experiments with siRNA werecarried out at 100 nmol/L concentration out using Lipofec-tamine 2000 in Opti-MEMReduced SerumMedia (Invitro-gen). Cells were transfected with nonspecific siRNA orsiRNA-targeting IGF-1R and on day 2, trypsinized and ineach well of a 6-well plate, 6,000 cells were suspended in 1.5mL of 0.7% agarose (at 37�C) inDMEMwith 10% fetal calfserum. The cells in agarose were plated atop a 1.4% agarose
Aslam et al.
Mol Cancer Res; 11(11) November 2013 Molecular Cancer Research1304
on August 28, 2018. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst August 8, 2013; DOI: 10.1158/1541-7786.MCR-12-0598

layer and were allowed to grow for 2 weeks before visualizingthe colonies by light microscopy.
ImmunohistochemistryHuman alveolar rhabdomyosarcoma tissue microarrays
were obtained from the COG Biorepository (TMA #3000-30-P6897). Histology was conducted using standard pro-tocol. Briefly, formalin-fixed paraffin-embedded sections(FFPE) were dewaxed and dehydrated in xylene and gradedalcohol concentrations. Heat-induced epitope retrieval wasused with sodium citrate buffer (pH 6.0) for 20 minutes,slides blocked with 2% normal goat serum (NGS) and forendogenous biotin using an avidin/biotin blocking kit (Vec-tor Labs). Primary antibody for IGF-1R (Santa Cruz) wasused at 1:50 overnight in 2% NGS. VECTASTAIN EliteABC kit (Rabbit IgG) and ImmPACT DAB PeroxidaseSubstrate kit was used according to manufacturer's protocolto visualize staining (Vector Lab). Slides were counterstainedwith hematoxylin for 5 minutes, rinsed, dehydrated, andmounted with xylene-based mounting medium.
ImmunocytochemistryConfocal microscopy was conducted as follows: cells were
allowed to recover postsort in culture for 2 to 3 days thenreplated in 8 chamber slides (BD Biosciences) at low con-fluency and allowed to incubate for 2 to 3 days. Cells werethen fixed in 4% paraformaldehyde in PBS at room tem-perature for 20 minutes. Slides were carefully washed withPBS three times and incubated at room temperature in 5%NGS (Invitrogen) and 0.01%Triton-X in PBS for 1 hour toinhibit nonspecific binding of antibodies. Cells were washedagain, and primary antibody diluted in 5%NGS in PBS wasadded overnight at 4�C. Alexafluor 488-conjugated orAlexafluor 594-conjugated anti-rabbit IgG antibody (Invi-trogen) at 1:200 was added, and cells were incubated for 1hour at room temperature. Slides were mounted using theVECTASHIELDMountingMediumwith 40, 6-diamidino-2-phenylindole (Vector Laboratories) and visualized with aZeiss LSM 700 confocal microscope. PDGFRa antibodywas used at 1:100 (#3164, Cell Signaling Technologies) andIGF-1Rb at 1:50 (sc-713, Santa Cruz Biotechnology).Images were taken at �40 and �63 oil immersion forfurther magnification.
In vivo studiesAll animals were treated humanely and the experiments
were carried out in accordance with the Institutional AnimalCare and Use Committee approved protocols. SCID/hair-less/outbred (Crl:SHO-PrkdcscidHrhr) mice purchased fromCharles River at 8 weeks of age were injected with 1 � 105
U23674 cells into the gastrocnemius muscle that had beenpreinjured 24 hours prior with 0.85 mg/mouse cardiotoxin(fromNaja mossambica, Sigma). Tumor volumes (cm3) weremeasured three dimensionally with electronic calipers andcalculated from formula (p/6) � length � width �height,assuming tumors to be spheroid. Once tumor volumereached 2 cm3, mice were euthanized and the tumor washarvested at necropsy.
ResultsMurine alveolar rhabdomyosarcomas express PDGFRaand IGF-1R by FACS and can be isolated by cell sortingThe murine alveolar rhabdomyosarcoma primary cell
culture, U23674, was derived from a left upper extremityprimary tumor that developed in a 46-day-old mouse fromour genetically engineered, conditional mouse model ofalveolar rhabdomyosarcoma harboring the Pax3:Fkhr onco-gene and homozygous p53 deletion (4, 5). This lesion washistologically diagnosed as an alveolar rhabdomyosarcomatumor and used for these studies at low passage number (P <10). Initially, U23674 cells were stained for PDGFRa (APC)and IGF-1R (Cy3.5) and analyzed by FACS, indicating thatapproximately 35% of cells express both PDGFRa and IGF-1R, but approximately 15% of cells express either receptorexclusively (Fig. 1A). However, tumor cells displayed acontinuum of expression of PDGFRa and IGF-1R ratherthan neatly distinct populations expressing either RTKindividually or together. Similar results were seen in twoother murine alveolar rhabdomyosarcoma cell lines analyzedby flow cytometry for cell surface expression of PDGFRaand IGF-1R (Supplementary Fig. S1).Next, after U23674 cells were stained for PDGFRa and
IGF-1R, the cells were sorted by expression of each receptorinto four populations. Gates were determined using controlcells stained with isotype control antibody in addition toa Cy3.5 conjugated secondary antibody as a control.Cells positive or negative for PDGFRa or IGF-1R by theassigned gates were designated as PDGFRahi/IGF-1Rhi orPDGFRalo/IGF-1Rlo, respectively (Fig. 1B). For instance,cells sorted for high IGF-1R expression, but negative forPDGFRa were denoted as PDGFRaloIGF-1Rhi. Immedi-ately following sorting, cells were reanalyzed by flow cyto-metry to assess purity of the sort (Fig. 1B). Figure 1C showsdistinct morphology of cells grown in culture for 7 dayspostsort. IGF-1Rhi expression, but not PDGFRahi expres-sion, was associated with spindle morphology.
PDGFRa and IGF-1R cell surface expression is dynamicOnce U23674 cells were sorted by PDGFRa and IGF-
1R expression, cells were allowed to recover postsort inculture and reanalyzed by flow cytometry for surfaceexpression of both RTKs at serial time points. FACSanalysis revealed that cell surface expression of bothPDGFRa and IGF-1R varied dramatically as cells weregrown in culture postsort. Two such time points areshown in Fig. 2 at 2 days and 14 days postsort, comparedwith day 0 (day 7 was similar to day 14; SupplementaryFig. S2). These results illustrated an initial dramaticchange in expression of PDGFRa and IGF-1R postsortwithin a 48-hour time period, tending toward a presortprofile. With the observation that cell surface expressionprofile shifted considerably within a short period of time,we sought to understand whether altered receptor expres-sion was a function of many or few rounds of replication.Thus, after a purity check was conducted for sortedpopulations postsort, cells were recounted and plated ata known cell number. Exactly 48 hours after the initial
Nuclear Receptor Tyrosine Kinases in Rhabdomyosarcoma
www.aacrjournals.org Mol Cancer Res; 11(11) November 2013 1305
on August 28, 2018. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst August 8, 2013; DOI: 10.1158/1541-7786.MCR-12-0598

sort, cells were recounted and FACS analysis for PDGFRaand IGF-1R expression was done. The number of cellscomprising each of the four populations was calculatedfrom the FACS profile of the initial sort and 48-hourpostsort (Fig. 2). Doubling times were on the order of30 to 42 hours (Supplementary Table S1); therefore, dy-namic alterations of PDGFRa and/or IGF-1R cell surfaceexpression seem to start after 0 to 1 cell divisions.
Total expression of PDGFRa and IGF-1R byimmunoblot analysis in sorted cells differs from cellsurface expression profile by FACSTo determine whether total expression of either PDGFRa
or IGF-1R in the four sorted cell populations reflected thesurface expression profile of each RTK by FACS, we con-ducted immunoblot analysis on cells grown in culture for 7days postsort. Surprisingly, total cellular PDGFRa expression
Figure 1. Murine alveolar rhabdomyosarcoma isolated by PDGFRa and IGF-1R expression. A, U23674 (murine alveolar rhabdomyosarcoma) cell expressionprofile by FACS of PDGFRa (APC) and IGF-1R (Cy3.5) gated with respect to isotype and secondary only control. B, purity of isolated cells by FACS shown.Isolated populations according to cells positive and negative for each receptor designated as hi and lo, respectively. C, phase contrast microscopy of sortedpopulations showing differential morphology of cells grown in culture 7 days postsort. Scale bar, 200 mm.
Aslam et al.
Mol Cancer Res; 11(11) November 2013 Molecular Cancer Research1306
on August 28, 2018. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst August 8, 2013; DOI: 10.1158/1541-7786.MCR-12-0598

seemed to vary dramatically among the sorted populations(Fig. 3A). Higher PDGFRa expression at day 7 correlatedwith the populations, which were sorted for high PDGFRaexpression by FACS initially. Conversely, total IGF-1Rexpression by immunoblot analysis did not show a dramaticdifference in expression among sorted populations (Fig. 3A).In terms of activation, phosphorylation of PDGFRa waselevated in the PDGFRahiIGF-1Rlo and PDGFRahiIGF-1Rhi populations (Fig. 3A). Despite lack of differentialexpression for total IGF-1R, IGF-1R phosphorylation wasincreased in the PDGFRahiIGF-1Rhi populations, as well as(modestly) in the PDGFRaloIGF-1Rhi population. Becausewe observed that activated receptor levels varied across sortedpopulations, we looked at activation of common cell survivaland proliferation pathways downstream of both PDGFRa
and IGF-1R such as Akt and extracellular signal-regulatedkinase (Erk)-1/2, respectively. Although we did not see adifference in Akt phosphorylation levels between the sortedcells (data not shown), Erk1/2 phosphorylation levels wereremarkably elevated in the PDGFRahiIGF-1Rhi populationand conversely decreased in the PDGFRaloIGF-1Rlo popu-lation; p-Erk 1/2 levels seemed to correlate with the levels ofphosphorylated PDGFRa and IGF-1R.Thus far, our results indicated that PDGFRa and IGF-1R
in tumor cells are expressed in a dynamic manner withrespect to surface expression profile by FACS, as well as totalprotein expression by immunoblot analysis. However, thedramatic difference in total PDGFRa expression (Fig. 3A),for example, did not reflect the percentage of cells positive forPDGFRa by FACS analysis at the same time point (day
Figure 2. Murine alveolarrhabdomyosarcoma showdynamic expression of PDGFRaand IGF-1R by FACS in a temporalmanner. A, FACS profile ofPDGFRa and IGF-1R expression inU23674 cells at day of sort (Day 0),and 2 and 14 days postsort. Day 7was similar to Day 14.
Nuclear Receptor Tyrosine Kinases in Rhabdomyosarcoma
www.aacrjournals.org Mol Cancer Res; 11(11) November 2013 1307
on August 28, 2018. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst August 8, 2013; DOI: 10.1158/1541-7786.MCR-12-0598
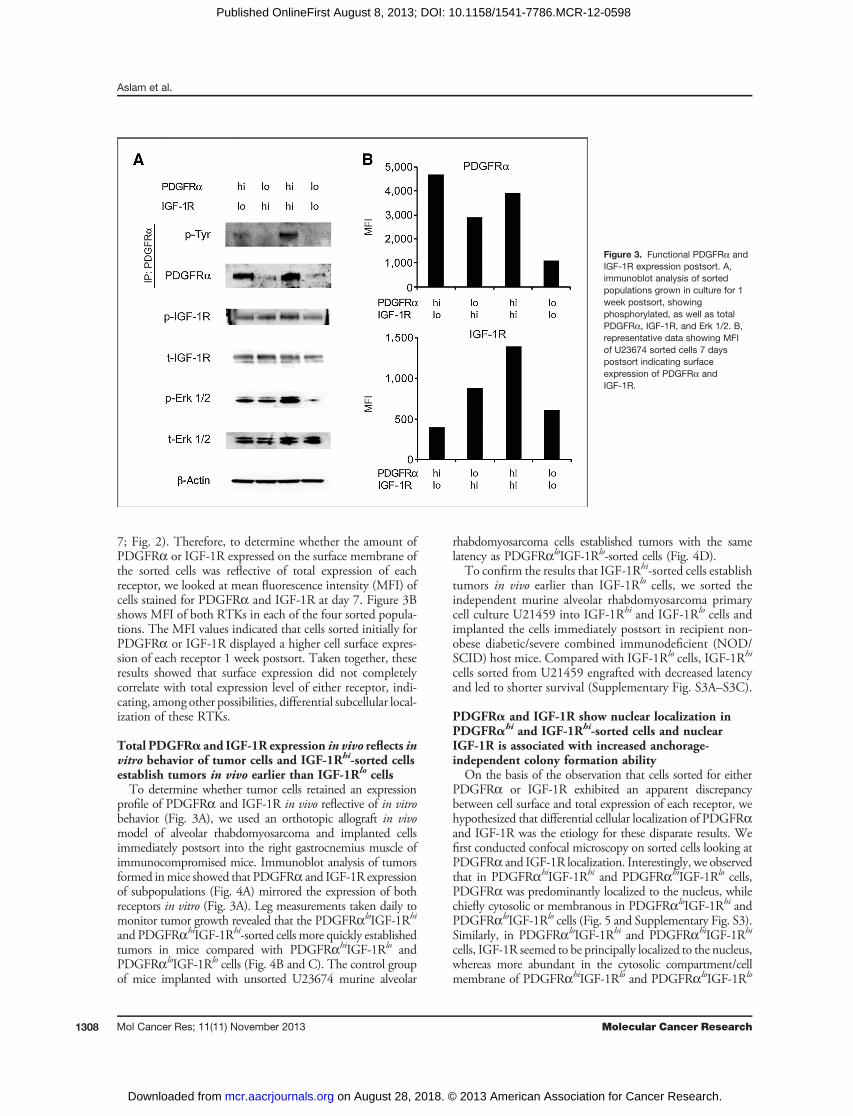
7; Fig. 2). Therefore, to determine whether the amount ofPDGFRa or IGF-1R expressed on the surface membrane ofthe sorted cells was reflective of total expression of eachreceptor, we looked at mean fluorescence intensity (MFI) ofcells stained for PDGFRa and IGF-1R at day 7. Figure 3Bshows MFI of both RTKs in each of the four sorted popula-tions. The MFI values indicated that cells sorted initially forPDGFRa or IGF-1R displayed a higher cell surface expres-sion of each receptor 1 week postsort. Taken together, theseresults showed that surface expression did not completelycorrelate with total expression level of either receptor, indi-cating, among other possibilities, differential subcellular local-ization of these RTKs.
Total PDGFRa and IGF-1R expression in vivo reflects invitro behavior of tumor cells and IGF-1Rhi-sorted cellsestablish tumors in vivo earlier than IGF-1Rlo cellsTo determine whether tumor cells retained an expression
profile of PDGFRa and IGF-1R in vivo reflective of in vitrobehavior (Fig. 3A), we used an orthotopic allograft in vivomodel of alveolar rhabdomyosarcoma and implanted cellsimmediately postsort into the right gastrocnemius muscle ofimmunocompromised mice. Immunoblot analysis of tumorsformed inmice showed that PDGFRa and IGF-1R expressionof subpopulations (Fig. 4A) mirrored the expression of bothreceptors in vitro (Fig. 3A). Leg measurements taken daily tomonitor tumor growth revealed that the PDGFRaloIGF-1Rhi
and PDGFRahiIGF-1Rhi-sorted cells more quickly establishedtumors in mice compared with PDGFRahiIGF-1Rlo andPDGFRaloIGF-1Rlo cells (Fig. 4B and C). The control groupof mice implanted with unsorted U23674 murine alveolar
rhabdomyosarcoma cells established tumors with the samelatency as PDGFRaloIGF-1Rlo-sorted cells (Fig. 4D).To confirm the results that IGF-1Rhi-sorted cells establish
tumors in vivo earlier than IGF-1Rlo cells, we sorted theindependent murine alveolar rhabdomyosarcoma primarycell culture U21459 into IGF-1Rhi and IGF-1Rlo cells andimplanted the cells immediately postsort in recipient non-obese diabetic/severe combined immunodeficient (NOD/SCID) host mice. Compared with IGF-1Rlo cells, IGF-1Rhi
cells sorted from U21459 engrafted with decreased latencyand led to shorter survival (Supplementary Fig. S3A–S3C).
PDGFRa and IGF-1R show nuclear localization inPDGFRahi and IGF-1Rhi-sorted cells and nuclearIGF-1R is associated with increased anchorage-independent colony formation abilityOn the basis of the observation that cells sorted for either
PDGFRa or IGF-1R exhibited an apparent discrepancybetween cell surface and total expression of each receptor, wehypothesized that differential cellular localization of PDGFRaand IGF-1R was the etiology for these disparate results. Wefirst conducted confocal microscopy on sorted cells looking atPDGFRa and IGF-1R localization. Interestingly, we observedthat in PDGFRahiIGF-1Rhi and PDGFRahiIGF-1Rlo cells,PDGFRa was predominantly localized to the nucleus, whilechiefly cytosolic or membranous in PDGFRaloIGF-1Rhi andPDGFRaloIGF-1Rlo cells (Fig. 5 and Supplementary Fig. S3).Similarly, in PDGFRaloIGF-1Rhi and PDGFRahiIGF-1Rhi
cells, IGF-1R seemed to be principally localized to the nucleus,whereas more abundant in the cytosolic compartment/cellmembrane of PDGFRahiIGF-1Rlo and PDGFRaloIGF-1Rlo
Figure 3. Functional PDGFRa andIGF-1R expression postsort. A,immunoblot analysis of sortedpopulations grown in culture for 1week postsort, showingphosphorylated, as well as totalPDGFRa, IGF-1R, and Erk 1/2. B,representative data showing MFIof U23674 sorted cells 7 dayspostsort indicating surfaceexpression of PDGFRa andIGF-1R.
Aslam et al.
Mol Cancer Res; 11(11) November 2013 Molecular Cancer Research1308
on August 28, 2018. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst August 8, 2013; DOI: 10.1158/1541-7786.MCR-12-0598

cells (Fig. 5 and Supplementary Fig. S4). In the biologicallyindependent U21459 cell culture, immunocytochemistry alsorevealed that IGF-1R was commonly localized to the nucleusin IGF-1Rhi cells and IGF-1Rlo cells, respectively (Supplemen-tary Fig. S3D). These results subsequently led us to questionwhether the ability of IGF-1Rhi cells to establish tumors earlierin vivo compared with IGF-1Rlo cells was due to higher nuclearexpression of IGF-1R in these cells. Focusing our efforts onIGF-1R because of the orthotopic allograft results, we nextconfirmed whether IGF-1R was indeed expressed at higherlevels in the nucleus in IGF-1Rhi cells, compared with IGF-1Rlo cells, by separating cytosolic and nuclear proteins of allfour sorted populations 7 days following isolation of cells byFACS. We subsequently immunoblotted for IGF-1R, usingSp1 and a-tubulin as confirmation of enrichment of nuclearand cytosolic cell fractions, respectively (Fig. 6A). These resultsconfirmed a higher nuclear expression of IGF-1R in IGF-1Rhi
cells compared with IGF-1Rlo cells.To confirm our hypothesis that nuclear expression of IGF-
1R contributed to the ability of tumor cells to establish moreaggressive tumors in vivo, we used RNAi to silence IGF-1R inPDGFRahiIGF-1Rhi and PDGFRaloIGF-1Rlo cells. We then
assessed the effect of IGF-1R knockdown on these cells' abilityto form colonies in an anchorage-independent colony forma-tion assay (Fig. 6B). Knockdown efficiencies of IGF-1R in bothsorted cells are shown (Fig. 6B). Colony formation ability ofPDGFRahiIGF-1Rhi was decreased approximately 45% com-pared with control (nonspecific siRNA) when IGF-1R wassilenced by RNAi, whereas PDGFRaloIGF-1Rlo cells showonly a approximately 27% reduction in colony formation abilitywith IGF-1R knockdown. Similarly, studies with the nuclearIGF-1R-expressingmurine alveolar rhabdomyosarcoma prima-ry cell culture U21459 showed that three independent siRNAfor IGF-1R suppressed colony formation in soft agar (Supple-mentary Fig. S3E). Taken together, these results indicate thatnuclear expression of IGF-1R confers an increased tumorigenicphenotype to murine alveolar rhabdomyosarcoma cells.
Normal human tissue expresses cytosolic IGF-1R,whereas human alveolar rhabdomyosarcoma has highfrequency of nuclear IGF-1R by immunohistochemistryTo determine whether nuclear IGF-1R occurs in
human alveolar rhabdomyosarcoma, we conducted immu-nohistochemistry for IGF-1R on a human alveolar
Figure 4. Cells sorted for IGF-1Restablish more aggressive tumorsin vivo. A, tumors established frommurine aRMS cells sorted forPDGFRa and IGF-1R show arelative dynamic and staticexpression of each receptor,respectively, reflective of cells invitro. B, PDGFRahiIGF-1Rlo-sortedcells show a significantly slowergrowth rate in an in vivo orthotopicallograft mouse model of alveolarrhabdomyosarcoma comparedwith PDGFRaloIGF-1Rhi-sortedcells. C, PDGFRahiIGF-1Rhi-sortedcells grow at a significantly fastergrowth rate in an in vivo orthotopicallograft mouse model of alveolarrhabdomyosarcoma comparedwith PDGFRaloIGF-1Rlo-sortedcells. D, tumor establishment orgrowth rate of unsorted U23674cells in vivo do not differ fromPDGFRaloIGF-1Rlo-sorted cells.
Nuclear Receptor Tyrosine Kinases in Rhabdomyosarcoma
www.aacrjournals.org Mol Cancer Res; 11(11) November 2013 1309
on August 28, 2018. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst August 8, 2013; DOI: 10.1158/1541-7786.MCR-12-0598

rhabdomyosarcoma tumor microarray with 31 sections of19 unique cases. We observed that 16 out of 19 (�84%)specimens had nuclear expression of IGF-1R (>10% nucleiof section positive). Tumor sections showed either cytosolic(Fig. 7A), or predominantly nuclear staining of IGF-1R(Fig. 7B). Normal human tissue (pancreatic and skeletalmuscle) was used as positive controls for IGF-1R, andshowed only cytosolic staining of IGF-1R; no nuclearIGF-1R was seen in these sections (data not shown). Detailsof sample scores are given in Supplementary Table S2.
DiscussionIn this report, we describe that the RTK PDGFRa and
IGF-1R exhibit a dynamic cell surface expression profile inmurine alveolar rhabdomyosarcoma cells. Although we wereable to isolate relatively pure populations of cells expressinghigh or low PDGFRa and/or IGF-1R, all sorted populationsrapidly and dynamically alter their cell surface expressionprofile before the second cell division. Kept isolated inculture, sorted populations eventually equilibrated their cellsurface expression profile toward a presort profile. In thecontext of previous studies illustrating the phenotypicallydynamic state of cancer cells, including stem and non-stemcancer cells (16, 25), our study is a key next step into
understanding the complexity and potential pitfalls ofassigning therapy based on the surface expression profilesor other static biomarkers. Previously, several studies haveused cell surface markers (e.g., CD133, CD44, CD24, etc.)as stable markers for a specific cell population (12, 15, 26).RTKs, which are known to undergo ligand-dependentsubcellular trafficking and recycling (18, 27, 28), mayhowever, be particularly undesirable as cell surface biomar-kers. Nevertheless, our results then pointed us to differencesin PDGFRa and IGF-1R subcellular localization that par-alleled membrane expression. IGF-1R nuclear localization,in particular, exhibited an important functional effect incontributing toward a more aggressive tumor phenotype invivo (summarized in Supplementary Fig. S5).Nuclear localization of RTKs or their substrates is an
interesting phenomenon, which has been described in selectearly reports (19–22, 29). IGF-1R had previously beenshown to be a poor prognostic indicator in clear cell renalcell carcinoma (30), and a new studywas able to further showthat among renal tumors expressing IGF-1R, intense orwidespread nuclear expression by immunohistochemistryportends a decrease in survival probability (17). Interesting-ly, nuclear localization of the IGF-1R holoreceptor wasdependent upon tyrosine phosphorylation of putative
Figure 5. Nuclear localization of PDGFRa and IGF-1R correlates with cells sorted by FACS for surface expression of the respective receptor. Representativeconfocal images of U23674 sorted populations showing localization of PDGFRa and IGF-1R (�40) with inset box enlarging image of a single cell to appreciatelocalization. Scale bar, 100 mm. Larger magnifications of insets are given in Supplementary Fig. S3.
Aslam et al.
Mol Cancer Res; 11(11) November 2013 Molecular Cancer Research1310
on August 28, 2018. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst August 8, 2013; DOI: 10.1158/1541-7786.MCR-12-0598

residues on the IGF-1Rb subunit of the receptor (31). In oursorted populations, the IGF-1Rhi cells, which displayed ahigher nuclear expression of IGF-1R, also had increasedphosphor-IGF-1Rb. A typical biologic response when an
RTK is stimulated by its respective ligand is receptorinternalization and subsequent degradation (31). However,phosphorylation of IGF-1Rb is one of the two necessaryposttranslation modifications of the receptor required for
Figure 6. IGF-1R is present insoluble nuclear fraction of sortedcells and is associated withanchorage-independent colonyformation ability. A, cytosolic andnuclear fractions of sortedU23674 cells show that cellssorted by IGF-1Rhi express higherlevels of nuclear IGF-1R. B, cellswith higher nuclear IGF-1R showasignificantly further decrease incolony formation ability comparedwith control (nonspecific siRNA)when IGF-1R is silenced bysiRNA. �, P < 0.05 compared withnonspecific siRNA by Studentt test.
Figure 7. IGF-1R localizationin human alveolarrhabdomyosarcoma (arms).A, human alveolarrhabdomyosarcoma sectionswith chiefly cytosolic IGF-1Rexpression. B, images of alveolarrhabdomyosarcoma sectionsshowing both cytosolic (left) andnuclear (right) IGF-1R expression.Scale bar, 100 mmol/L.
Nuclear Receptor Tyrosine Kinases in Rhabdomyosarcoma
www.aacrjournals.org Mol Cancer Res; 11(11) November 2013 1311
on August 28, 2018. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst August 8, 2013; DOI: 10.1158/1541-7786.MCR-12-0598

nuclear trafficking. SUMOylation of K1025, K1100, andK1120 residues in the kinase domain of IGF-1Rb is alsocritical to nuclear import of the holoreceptor (32). Furtherinvestigation has shown that nuclear IGF-1R originates fromsurface IGF-1R only (33). Of note, we used an antibody tosort cells based on surface IGF-1R expression (the a subunitof IGF-1R) and a different antibody to detect IGF-1Rb atthe nuclear fraction by immunoblot analysis, but given thecovalent assembly of a and b subunits in the maturereceptor, we do not expect this to have altered our results.Another intriguing aspect of our studies was that nuclearPDGFRa did not seem to have an adverse functional rolewith respect to tumor establishment in vivo.Although we were able to identify a difference in receptor
localization among the sorted populations ofmurine alveolarrhabdomyosarcoma, we did not observe a differential sen-sitivity of these cells to small-molecule inhibitors whichtarget PDGFRa or IGF-1R (data not shown). An interestingquestion these results raise is whether small-molecule inhi-bitors designed to inhibit kinase activity of RTKs are able toinhibit the function of these RTKs when localized to thenucleus. Understanding whether kinase activity contributesto the role these receptors play in the nucleus would be onestep toward answering this question. Presumably, small-molecule inhibitors would have an advantage over neutral-izing antibodies in this respect. Prior studies have alreadyshown that certain RTKs, such as EGFR, interact withDNA-binding proteins to directly regulate the transcriptionof putative gene targets (34). The exact role of IGF-1R in thenucleus of alveolar rhabdomyosarcoma will be an active areaof further investigation, particularly given the potentialtranslation from mouse to human and the expression ofnuclear IGF-1R in clinical FFPE samples. The mechanismby which nuclear IGF-1R contributes to the establishment
of new (metastatic) tumors will be a key subject of furtherstudy and perhaps an operative step toward a temporalbiomarker-driven and personalized medicine approach incancer therapy with regards to IGF-1R inhibition.
Disclosure of Potential Conflicts of InterestNo potential conflicts of interest were disclosed.
Authors' ContributionsConception and design:M.I. Aslam, S. Hettmer, A. Soundararajan, B.J. Druker, A.J.Wagers, J.W. Tyner, C. KellerDevelopment of methodology: M.I. Aslam, S. Hettmer, A. Soundararajan, J.W.Tyner, C. KellerAcquisition of data (provided animals, acquired and managed patients, providedfacilities, etc.):M.I. Aslam, Simone Hettmer, J. Abraham, D. LaTocha, E.T. Huang,S. Wang, A. Mansoor, B.J. Druker, J.W. Tyner, C. KellerAnalysis and interpretation of data (e.g., statistical analysis, biostatistics, compu-tational analysis): M.I. Aslam, Simone Hettmer, D. LaTocha, M.W. Goros, J.E.Michalek, A. Mansoor, J.W. Tyner, C. KellerWriting, review, and/or revision of the manuscript: M.I. Aslam, S. Hettmer,A. Soundararajan,M.W.Goros, J.E.Michalek, A.Mansoor, B.J. Druker, A.J.Wagers,J.W. Tyner, C. KellerAdministrative, technical, or material support (i.e., reporting or organizing data,constructing databases): M.I. Aslam, B.J. Druker, J.W. Tyner, C. KellerStudy supervision: M.I. Aslam, B.J. Druker, J.W. Tyner, C. Keller
Grant SupportThis work was supported by 5R01CA133229-05 and -06 to C. Keller. The studies
were also supported, in part, from a gift from the Ethan Jostad Foundation forChildhood Cancer. Pilot studies leading to this work were supported by an innovationaward from Alex's Lemonade Stand. J.W. Tyner is supported by grants from theWilliam Lawrence and Blanche Hughes Foundation, Leukemia & LymphomaSociety, and the National Cancer Institute. M.I. Aslam is a Howard Hughes MedicalInstitute Medical Research Fellow. B.J. Druker is an investigator of the HowardHughes Medical Institute.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be herebymarked advertisement in accordance with18 U.S.C. Section 1734 solely to indicate this fact.
Received October 17, 2012; revised June 25, 2013; accepted July 15, 2013;published OnlineFirst August 8, 2013.
References1. Arndt CA, Crist WM. Common musculoskeletal tumors of childhood
and adolescence. N Engl J Med 1999;341:342–52.2. Breneman JC, Lyden E, Pappo AS, Link MP, Anderson JR, Parham
DM, et al. Prognostic factors and clinical outcomes in childrenand adolescents with metastatic rhabdomyosarcoma–a report fromthe Intergroup Rhabdomyosarcoma Study IV. J Clin Oncol 2003;21:78–84.
3. Pappo AS, Shapiro DN, Crist WM. Rhabdomyosarcoma. Biology andtreatment. Pediatr Clin North Am 1997;44:953–72.
4. Keller C, Arenkiel BR, Coffin CM, El-Bardeesy N, DePinho RA, Capec-chi MR. Alveolar rhabdomyosarcomas in conditional Pax3:Fkhr mice:cooperativity of Ink4a/ARF and Trp53 loss of function. Genes Dev2004;18:2614–26.
5. Nishijo K, ChenQR, Zhang L,McCleish AT, RodriguezA, ChoMJ, et al.Credentialing a preclinical mouse model of alveolar rhabdomyosar-coma. Cancer Res 2009;69:2902–11.
6. Abraham J, Prajapati SI, Nishijo K, Schaffer BS, Taniguchi E, KilcoyneA, et al. Evasion mechanisms to IGF-1R inhibition in rhabdomyosar-coma. Mol Cancer Ther 2011;10:697–707.
7. Taniguchi E, Nishijo K,McCleishAT,Michalek JE,GraysonMH, InfanteAJ, et al. PDGFR-A is a therapeutic target in alveolar rhabdomyosar-coma. Oncogene 2008;27:6550–60.
8. Armistead PM, Salganick J, Roh JS, Steinert DM, Patel S, Munsell M,et al. Expression of receptor tyrosine kinases and apoptotic molecules
in rhabdomyosarcoma: correlationwith overall survival in 105 patients.Cancer 2007;110:2293–303.
9. Cen L, Arnoczky KJ, Hsieh FC, Lin HJ, Qualman SJ, Yu S, et al.Phosphorylation profiles of protein kinases in alveolar and embryonalrhabdomyosarcoma. Mod Pathol 2007;20:936–46.
10. Kim ES, Herbst RS, Wistuba II, Lee JJ, Blumenschein GR Jr, Tsao A,et al. The BATTLE trial: personalizing therapy for lung cancer. CancerDiscov 2011;1:44–53.
11. Gerlinger M, Rowan AJ, Horswell S, Larkin J, Endesfelder D,Gronroos E, et al. Intratumor heterogeneity and branched evolutionrevealed by multiregion sequencing. N Engl J Med 2012;366:883–92.
12. Bertolini G, Roz L, Perego P, Tortoreto M, Fontanella E, Gatti L, et al.Highly tumorigenic lung cancer CD133þ cells display stem-like fea-tures and are spared by cisplatin treatment. Proc Natl Acad Sci U S A2009;106:16281–6.
13. Patrawala L, Calhoun-Davis T, Schneider-Broussard R, Tang DG.Hierarchical organization of prostate cancer cells in xenograft tumors:theCD44þalpha2beta1þcell population is enriched in tumor-initiatingcells. Cancer Res 2007;67:6796–805.
14. Shmelkov SV, Butler JM, Hooper AT, Hormigo A, Kushner J, Milde T,et al. CD133 expression is not restricted to stem cells, and bothCD133þ and CD133- metastatic colon cancer cells initiate tumors.J Clin Invest 2008;118:2111–20.
Aslam et al.
Mol Cancer Res; 11(11) November 2013 Molecular Cancer Research1312
on August 28, 2018. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst August 8, 2013; DOI: 10.1158/1541-7786.MCR-12-0598

15. Singh SK, Hawkins C, Clarke ID, Squire JA, Bayani J, Hide T, et al.Identification of human brain tumour initiating cells. Nature 2004;432:396–401.
16. Gupta PB, Fillmore CM, Jiang G, Shapira SD, Tao K, Kuperwasser C,et al. Stochastic state transitions give rise to phenotypic equilibrium inpopulations of cancer cells. Cell 2011;146:633–44.
17. Aleksic T, Chitnis MM, Perestenko OV, Gao S, Thomas PH,Turner GD, et al. Type 1 insulin-like growth factor receptortranslocates to the nucleus of human tumor cells. Cancer Res2010;70:6412–9.
18. Ettenberg SA, Magnifico A, Cuello M, NauMM, Rubinstein YR, YardenY, et al. Cbl-b-dependent coordinated degradation of the epidermalgrowth factor receptor signaling complex. J Biol Chem 2001;276:27677–84.
19. Johnston CL, Cox HC, Gomm JJ, Coombes RC. Fibroblast growthfactor receptors (FGFRs) localize in different cellular compartments. Asplice variant of FGFR-3 localizes to the nucleus. J Biol Chem1995;270:30643–50.
20. Liao HJ, Carpenter G. Role of the Sec61 translocon in EGF receptortrafficking to the nucleus and gene expression. Mol Biol Cell 2007;18:1064–72.
21. Lin SY, Makino K, Xia W, Matin A, Wen Y, Kwong KY, et al. Nuclearlocalization of EGF receptor and its potential new role as a transcriptionfactor. Nat Cell Biol 2001;3:802–8.
22. Sardi SP, Murtie J, Koirala S, Patten BA, Corfas G. Presenilin-depen-dent ErbB4nuclear signaling regulates the timingof astrogenesis in thedeveloping brain. Cell 2006;127:185–97.
23. Duncan JS, Whittle MC, Nakamura K, Abell AN, Midland AA, Zawis-towski JS, et al. Dynamic reprogramming of the kinome in response totargeted MEK inhibition in triple-negative breast cancer. Cell2012;149:307–21.
24. Huang F, Hurlburt W, Greer A, Reeves KA, Hillerman S, Chang H, et al.Differential mechanisms of acquired resistance to insulin-like growthfactor-I receptor antibody therapy or to a small-molecule inhibitor,
BMS-754807, in a human rhabdomyosarcoma model. Cancer Res2010;70:7221–31.
25. Chaffer CL, Brueckmann I, Scheel C, Kaestli AJ, Wiggins PA, Rodri-gues LO, et al. Normal and neoplastic nonstem cells can spontane-ously convert to a stem-like state. Proc Natl Acad Sci U S A2011;108:7950–5.
26. Li C, Heidt DG, Dalerba P, Burant CF, Zhang L, Adsay V, et al.Identification of pancreatic cancer stem cells. Cancer Res 2007;67:1030–7.
27. Le Roy C, Wrana JL. Clathrin- and non-clathrin-mediated endocyticregulation of cell signalling. Nat Rev Mol Cell Biol 2005;6:112–26.
28. WangY, Pennock SD, Chen X, Kazlauskas A,Wang Z. Platelet-derivedgrowth factor receptor-mediated signal transduction from endo-somes. J Biol Chem 2004;279:8038–46.
29. Wu A, Chen J, Baserga R. Nuclear insulin receptor substrate-1 acti-vates promoters of cell cycle progression genes. Oncogene 2008;27:397–403.
30. Parker A, Cheville JC, Lohse C, Cerhan JR, Blute ML. Expression ofinsulin-like growth factor I receptor and survival in patients with clearcell renal cell carcinoma. J Urol 2003;170:420–4.
31. Mukherjee S, Tessema M, Wandinger-Ness A. Vesicular trafficking oftyrosine kinase receptors and associated proteins in the regulation ofsignaling and vascular function. Circ Res 2006;98:743–56.
32. Sehat B, Tofigh A, Lin Y, Trocme E, Liljedahl U, Lagergren J, et al.SUMOylation mediates the nuclear translocation and signaling of theIGF-1 receptor. Sci Signal 2010;3:ra10.
33. Deng H, Lin Y, Badin M, Vasilcanu D, Stromberg T, Jernberg-WiklundH, et al. Over-accumulation of nuclear IGF-1 receptor in tumor cellsrequires elevated expression of the receptor and the SUMO-conju-gating enzyme Ubc9. Biochem Biophys Res Commun 2011;404:667–71.
34. Lo HW. Nuclear mode of the EGFR signaling network: biology, prog-nostic value, and therapeutic implications. Discov Med 2010;10:44–51.
Nuclear Receptor Tyrosine Kinases in Rhabdomyosarcoma
www.aacrjournals.org Mol Cancer Res; 11(11) November 2013 1313
on August 28, 2018. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst August 8, 2013; DOI: 10.1158/1541-7786.MCR-12-0598

2013;11:1303-1313. Published OnlineFirst August 8, 2013.Mol Cancer Res M. Imran Aslam, Simone Hettmer, Jinu Abraham, et al. Alveolar Rhabdomyosarcoma
and IGF-1R inαDynamic and Nuclear Expression of PDGFR
Updated version
10.1158/1541-7786.MCR-12-0598doi:
Access the most recent version of this article at:
Cited articles
http://mcr.aacrjournals.org/content/11/11/1303.full#ref-list-1
This article cites 34 articles, 17 of which you can access for free at:
Citing articles
http://mcr.aacrjournals.org/content/11/11/1303.full#related-urls
This article has been cited by 1 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications Department at
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://mcr.aacrjournals.org/content/11/11/1303To request permission to re-use all or part of this article, use this link
on August 28, 2018. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst August 8, 2013; DOI: 10.1158/1541-7786.MCR-12-0598