Embed Size (px)
Citation preview

seminars in CELL & DEVELOPMENTAL BIOLOGY, Vol 9, 1998: pp 445]450 Article No. sr980202
Dynamics of meiotic prophase I duringspermatogenesis: from pairing to divisionJohn Cobb and Mary Ann Handel
This review focuses on recent developments in ourunderstanding of meiotic chromosome behavior inmammalian spermatogenesis, with a special emphasis onprophase I events in the best characterized organism, thelaboratory mouse. The dynamics of chromosome movementprior to pairing and synapsis of chromosomes are complexand implicate function for both centromeres and telomeres ingetting homologous chromosomes together in intimatesynapsis. Likely candidates for mediating pairing andrecombination include a host of proteins implicated in DNArepair and recombination, which have been shown to localizeto the synaptonemal complex during meiotic prophase I.Precocious induction of meiotic metaphase in culturedpachytene spermatocytes has led to new information aboutrequirements for MPF and topoisomerase II activity duringthe transition from meiotic prophase to metaphase. Together,the studies reviewed here increase our understanding of howchromosomes get together with their homologous partnersand how these partners subsequently come apart.
Key words: chromosome pairing r DNA repair proteins rmeiosis r synaptonemal complex rtopoisomerase II
Q1998 Academic Press
Introduction
MEIOSIS ACCOUNTS FOR ONE-THIRD of the time re-quired to make a sperm cell in the mouse. Over 90%of the time for meiosis is spent in prophase I andmost of that in the pachytene stage. The tasks facingthe germ cell during prophase I are tremendous.Homologous chromosomes must find each other,align and then intimately synapse. At least one re-combination event must occur within each homologpair; the penalty for failure is malsegregation and loss
From the Department of Biochemistry and Cellular and Molecu-lar Biology, University of Tennessee, Knoxville, TN 37996-0840,USA
Q1998 Academic Press1084-9521r98r040445q06 $30.00r0
of offspring due to aneuploidy. Progress of recombi-nation must be monitored to ensure proper timing ofthe end of meiotic prophase. Finally, in transition outof meiotic prophase to metaphase I, chromosomesmust be desynapsed and disentangled and attachedto an assembled spindle apparatus.
What are the mechanisms governing this intricatebehavior of chromosomes and how are they coordi-nated? In spite of the fact that meiotic prophase isdefined by pairing and recombination of homolo-gous chromosomes, the underlying mechanismslargely are unknown. Recent studies on chromosomemovement in early meiotic prophase, on the localiza-
Ž .tion to the synaptonemal complex SC of proteinsimplicated in DNA repair and recombination and onthe dynamics in vitro of homolog desynapsis andprogress to metaphase, all combine to give us insightsand direction to analysis of meiotic prophase mecha-nisms.
Chromosomal movements associated withpairing
Although in some species homologous chromosomesoccupy common nuclear territory prior to meiosis, inmost species the homologs must find each other atthe onset of meiotic prophase. The study of thedynamics of the pairing of meiotic chromosomes hasbeen difficult because of the absence of methods tostudy this process in vitro. Scherthan et al1 haveovercome the lack of such methods by exploiting acombination of techniques to address the questions:are there any specific chromosomal movements orpositions associated with the onset of pairing andwhen do these movements occur? The difficulty inanswering these questions lies in being able to de-termine simultaneously the precise meiotic stage, theprogress of chromosome pairing and synapsis, andthe position of specific chromosomes within nuclei.By studying fixed, sectioned testes of prepuberal miceof various ages, the authors were able to determine
445

J. Cobb and M. A. Handel
precisely the different stages of meiotic prophase asthey appeared sequentially during development. Thenovelty of this study resides in combining the precise
Ž .staging with fluorescence in situ hybridization FISHto follow chromosome behavior, and with immuno-staining of proteins of the SC to follow pairing andsynapsis. The SC is a proteinaceous structure foundexclusively in meiotic cells lying between the axes ofpaired homologous chromosomes; its structure andfunction recently have been reviewed in depth byMoens.2
Previously, it was unknown if chromosomes arealigned or paired prior to the tight synapsis initiatedat zygonema. Scherthan et al1 used FISH to definepairing as the point at which the separate signals
Žfrom homologous chromosomes merge as one FISHsignals are represented diagrammatically as the rect-
.angles in Figure 1 . Alignment was followed withŽ .Chromosome 8 Chr-8 -specific probes by noting the
distance between the signals within a given nucleus.Leptotene cells showed no significant pairing withthe FISH assay and the first stage with a significantnumber of fused signals was zygonema. Thus, it canbe concluded that chromosomes are not aligned be-
fore zygonema. What movements bring the chromo-somes into a paired position at zygonema so that theycan synapse?
Chromosome movements were followed by usingtelomere or centromere FISH probes. These probeswere used in combination either with Chr-8 FISHprobes to show pairing, or with indirect immunoflu-orescence of SC proteins to show synapsis. This al-lowed, for the first time in mammalian meiotic cells,the visualization of chromosome positions as pairingbecomes established. Centromere movement to thenuclear envelope correlated with the beginning ofmeiotic prophase. Late in preleptonema, thetelomeres move to the nuclear periphery and alongthe nuclear envelope until they all cluster at one endof the nucleus, defining the ‘bouquet’ stage at thebeginning of zygonema. The authors showed thatChr-8 FISH signals become paired at the same time
Ž .as this telomere clustering Figure 1B and that for-mation of the SC, indicating synapsis, also begins atthe bouquet stage. Thus, a specific chromosomalconformation, the bouquet, has been linked to thebeginning of pairing and synapsis.
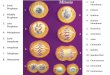
Figure 1 illustrates how bouquet formation might
Figure 1. Diagrammatic representation of chromosome movements associated with chromosomeŽ .pairing and unpairing during meiosis I prophase of spermatogenesis. The rectangles ST
represent homologous regions on a pair of chromosomes. These rectangles are analogous toChr-8 FISH signals.1. For clarity only one chromosome pair is shown. Pairing and synapsis beginin B; synapsis is illustrated diagrammatically in C and completed in D. Unpairing, chromosomecondensation and individualization correlate with the activation of MPF at the end of thepachytene stage and require the action of topo II. At metaphase I, unpairing is complete exceptfor connections at the chiasmata, which disappear at anaphase I. Proteins presumed to mediatepairing and recombination associate with chromosomes at various stages of pairing: v, proteinsŽ . Ž .such as RAD51 that associate with unpaired chromosomal axes and S, proteins such as MLH1that associate only with paired axes.
446

Meiotic dynamics in spermatogenesis
aid in the homology search thought to be necessaryfor pairing.1 By anchoring all chromosomes near thesame point, homologous regions of chromosomescould associate more readily with each other as theyshould be the same distance from the cluster point.Once pairing is initiated, the telomeres move awayfrom the cluster point and the homologs are zippedfurther together until synapsis is complete. Unfortu-nately, there currently is no means to directly test thishypothesis. Scherthan et al1 have shown only a corre-lation between these movements and the onset ofpairing. In fact the bouquet formation could have afunction other than chromosome alignment, perhapsin crossover control.3
Proteins implicated in meiotic prophasedynamics
Only recently have some of the likely players in thepairing and recombination processes been identifiedin mammalian spermatogenesis. The evidence to dateprimarily is inferential, with likely candidates beingidentified by virtue of ‘being in the right place at theright time’. This is evidenced by immunolocalizationof specific candidate proteins in cytological prepara-tions adapted for visualization of the SC. Promising‘candidates’ are those showing homology to proteinsrequired for chromosome pairing andror recombi-
Ž .nation in other organisms e.g. yeast, Drosophila andalso those where a mouse null mutation generated byhomologous recombination leads to meiotic arrest ofgerm-cell development and hence infertility.
Logical candidates for essential roles in chromo-some pairing or homologous recombination are pro-teins of the SC. Two of these have been studied in
Ž .depth, the rat and mouse SCP1 also called SYN1Ž . 4,5and SCP3 also referred to as COR1 . Antibodies to
SCP1rSYN1 localize to the space between synapsedchromosome cores in tripartite SC, whereas anti-bodies to SCP3rCOR1 identify chromosome coresprior to and after full synapsis.5 Both proteins havenon-specific DNA-binding properties, but their possi-ble roles in pairing, synapsis and recombination re-main unknown. SCP1rSYN1 and SCP3rCOR1 arethe only well-characterized proteins that have beenidentified as components of isolated SCs. Other pro-teins mentioned below show association with or local-ization to the SC in cytological preparations, butwhether they are structural andror functional com-ponents is not yet known.
One of the most intriguing proteins localizing to
Ž .the SC is HSP70-2 see also Eddy, this volume . Thisis a unique variant of the HSP70 heat-shock proteinswhose synthesis is restricted to meiotic prophasespermatocytes.6] 8 Targeted disruption of the Hsp70-2gene results in male infertility, with arrest of germcells prior to the meiotic divisions.9 Recently, it hasbeen found that HSP70-2 is required for associationof the cell-cycle regulator CDC2 with its partnercyclin B1 and subsequent kinase activity.10,11 Theseinteresting results suggest that regulation of theG2rM transition in spermatocytes may be tied tocompletion or monitoring of events occurring at theSC.
Recently, a number of proteins implicated in DNArepair andror recombination have been reported tobe associated with the SC. One of these is RAD51, amurine homolog of the yeast Rad51 protein, relatedto the bacterial RecA protein, which binds to single-stranded DNA and promotes strand invasion to forma heteroduplex molecule with proper sequencealignment for recombination.12 Yeast S. cerevisiaeRad51 protein is required for recombination afterformation of the single-stranded intermediate and isimplicated as a member of the recombination com-plex.13 In mouse spermatocytes, RAD51 protein hasbeen reported to be associated with chromosomalcores in foci prior to synapsis, but to disappear aftersynapsis, in early to mid-pachynema,14 ,15 implyingfunction in homology recognition or in recombina-
Ž .tion in mammals as in yeast Figure 1B . The RAD51protein is assumed to be involved in the processing ofDNA double-strand breaks, but this gets at the heartof our ignorance about molecular events of mam-malian meiosis. We do not know when the putativedouble-strand breaks form, or when recombinationintermediates appear, or when these events occurrelative to the homology search. Thus, it is difficult toset these results on RAD51 expression in mouse sper-matocytes into a biochemical context or to define itsprecise role in mammalian chromosome pairingandror recombination.
There are reports of localization at the SC of anumber of proteins implicated in both DNA repairprocesses and tumor susceptibility; these includeMLH1, ATM, ATR and BRCA1. The murine MLH1protein is a homolog of bacterial MutL proteins,highly conserved throughout evolution and involvedin mismatch repair of DNA. Interestingly, these genesare also implicated in susceptibility to various can-cers.16 Infertility and meiotic impairment in the malegerm line is a consequence of production of nullmutants for the Mlh1 gene. In both oocytes and
447

J. Cobb and M. A. Handel
spermatocytes of the mouse, MLH1 was found associ-ated with the SC, in discrete foci in pachytene sper-matocytes and at chiasmata in diplotene oocytes17
Ž .Figure 1 . Of interest are differences in the numberand distribution of MLH1 foci in oocytes and sper-matocytes, which reflect sex-specific differences inrate and distribution of chiasma sites. This observa-tion is consistent with recent data suggesting thatyeast Mlh1 protein promotes crossing-over duringmeiosis.18 Another protein implicated in DNA da-mage recognition and repair is the mouse homolog
Žof ATM, product of the human ATM mutated in.ataxia telangiectasia gene. The ATM protein has a
region of homology to the phosphatidylinositol 3-kinase family, whose members have been implicatedin cell-cycle checkpoints, meiotic recombination,DNA-damage response and cell signalling.19 Micehomozygous for an Atm null mutation are infertile,with germ-cell arrest in males at the zygotene or earlypachytene stage of meiotic prophase.20 In addition toATM, another mouse homologous protein is ATR,
Ž . 21the product of the Atr AT and rad-related gene.It has been reported that both ATM and ATR can bedetected with antibodies at the SC,22 but the natureand function of this association is unknown as yet.Two additional tumor suppressor proteins, impli-cated in susceptibility to breast cancer, BRCA1 andBRCA2, also may be involved in the dynamics ofmeiotic prophase, but here the evidence for a directrole is more tenuous. Both proteins are highly ex-pressed in mouse testes,23,24 interact with RAD51 andtherefore may be involved in the repair of DNAdouble-strand breaks25,26 and are implicated in tran-scription.27,28 The BRCA1 protein can be detected inasynapsed regions of paired chromosomes.25 Geneticanalysis of meiotic function of these two proteins willbe a challenge, as the null mutations result in embry-onic lethality.25,26
These studies on immunolocalization of candidateproteins implicated in DNA repair and recombina-tion do not yet provide definitive or conclusive evi-dence about their meiotic function, or, indeed, thenature of their association with the SC. For example,proteins with activity as transcription factors may belocalized close to the SC in spermatocytes, but play arole in the active transcription of meiotic prophaserather than in recombination. Nonetheless, observa-tions thus far are intriguing. It will be reassuringwhen they are repeated in different laboratories andwith a variety of methods, including immunolocaliza-tion to sectioned testes and identification inbiochemically isolated SCs. Taken together, these
studies implicate a host of interesting proteins andprovide a foundation for exploring hypotheses abouttheir biochemical functions. They give us a glimpseof what is probably only the tip of the iceberg in thecomplexity of meiotic prophase events.
Desynapsis and unpairing
Just as chromosomes pair during early meioticprophase, they must desynapse and unpair to exit
Ž .prophase Figure 1 . It is assumed that this occursafter all or most of recombination is completed; theprimary basis for this assumption is the presence ofchiasmata, the visual manifestation of prior crossoverevents, between paired homologs. The nature of thesignal that recombination is complete, and that allother prophase events have been executed, is un-known. This process is analogous to the progressionto metaphase in mitosis except that it involves theunpairing of homologous chromosomes. Theprocesses of desynapsis and completion of chromatin
Žcondensation to metaphase I the meiosis I G2rM.transition are very rapid in spermatocytes; therefore
these cells are rare in the testis and difficult to studyin situ. The study of this unpairing has been facili-tated by the development of a method to inducemetaphase precociously by treating culturedpachytene spermatocytes with the phosphatase in-
Ž . 29hibitor okadaic acid OA . This treatment causesspermatocyte chromosomes to desynapse and con-dense to chiasmate bivalents within 6 h and thusallows us to follow the unpairing process.
We do not know what signals completion of thepachytene stage, but OA can mimic this signal androrabrogate an inhibitory signal. The progression fromG2 to metaphase in mitosis is controlled by the uni-versal cell cycle regulator MPF, a complex of the
Ž .kinase CDC2 p34cdc2 and cyclin B1, which can beactivated indirectly by okadaic acid treatment.30 Thehistone H1 kinase activity of MPF is activated whenpachytene spermatocytes are induced to metaphaseby OA.29 Does MPF cause the paired chromosomesto desynapse? OA induces the desynapsis of pairedchromosomes with SC disassembly similar to thatseen in the natural transition to metaphase I.31 WhenMPF activation by OA is blocked by the kinase inhibi-tors staurosporine or roscovitine, the SC remainsintact and the transition to metaphase is blocked.While these observations do not prove definitivelythat MPF is the signal that initiates desynapsis, theydo provide a strong correlation between MPF activa-
448

Meiotic dynamics in spermatogenesis
tion, desynapsis, chromosome condensation andtransition to the first meiotic metaphase.
Precocious induction of the G2rM transition hasrevealed a requirement during condensation ofmetaphase I chromosomes for the DNA decatenation
Ž . 31activity of topoisomerase II topo II . When cul-tured in the presence of OA and topo II inhibitors,pachytene spermatocytes do not condense metaphasechromosomes as they do when treated with OA alone;nonetheless the SC still is disassembled in these con-ditions. Thus, topo II appears to be unnecessary fordesynapsis but necessary for condensing chromo-somes into individualized units, which can be sepa-rated at anaphase. MPF is still activated after OAtreatment in the presence of topo II inhibitors.Therefore, MPF activation is not sufficient to con-dense MI chromosomes in the absence of topo IIactivity.
Interestingly, different topo II inhibitors cause dif-ferences in chromatin condensation in OA-treatedpachytene spermatocytes.31 Treatment with tenipo-side, which inhibits topo II after the generation ofDNA double strand breaks by the enzyme, results inlittle or no chromosome condensation even thoughthe SC is disassembled. Possibly the double strandbreaks caused by teniposide activate a DNA damagecheckpoint that arrests G2rM chromosome conden-sation. In interesting contrast, treating cells withICRF-193, which inhibits topo II in a closed clampposition prior to generation of DNA breaks, results inchromatin condensation into elongated, pairedstrands, similar to that seen in diplotene. Indirectimmunofluorescence shows that in many cells treatedwith ICRF-193 and OA, the localization of the 170-kDa isoform of topo II shifts from centromeric hete-rochromatin to the axes of the elongated chromo-
Ž .somes unpublished observations . This suggests thattopo II may be trapped by ICRF-193 in chromatin atthe site of its enzymatic action. These results suggesta dynamic role for topo II in condensing pairedhomologs, during which the enzyme’s catalytic activ-ity is required and its localization changes.
Conclusion
The studies reviewed here raise more questions thansolutions about mechanisms governing meioticprophase chromosomal dynamics. For example, whatproteins mediate the successful initiation of pairingat the bouquet stage preceding synapsis? What is thebiochemical nature of the association with the SC of
proteins mediating recombination? What is the tem-poral ordering of recombination events and how arethey linked to pairing and synapsis? Could some ofthe DNA repair proteins localizing to the SC beinvolved in checkpoint surveillance, monitoringfidelity andror completion of recombination? Mightthese proteins be involved in activating MPF andsignalling the end of meiotic prophase, perhaps byinteracting with HSP70-2? How are these events cou-pled with the tremendous growth that occurs duringmeiotic prophase in spermatogenesis? Takentogether, the information reviewed here on the dy-namics of meiotic prophase suggest an intriguingsynthesis of mechanisms governing chromosomemovements, recombination and condensation for themeiotic division first. Precision and accuracy of theseevents is essential to make good gametes and avoidaneuploidy.
Acknowledgements
We are grateful to Drs. John Eppig, Monica Justice, JohnKoontz, Bruce McKee, Peter Moens and Laura Richardsonfor critical comments on the manuscript. Work from theauthors’ laboratory cited here was supported by grants HD31376 and 33816 from the NIH to MAH.
References
1. Scherthan H, Weich S, Schwegler H, Heyting C, Harle M,Ž .Cremer T 1996 Centromere and telomere movements dur-
ing early meiotic prophase of mouse and man are associatedwith the onset of chromosome pairing. J Cell Biol134:1109]1125
Ž .2. Moens PB 1994 Molecular perspectives of chromosomepairing at meiosis. BioEssays 16:101]106
3. Bass HW, Marshall WF, Sedat JW, Agard DA, Cande WZŽ .1997 Telomeres cluster de novo before the initiation ofsynapsis: A three dimensional spatial analysis of telomerepositions before and during meiotic prophase. J Cell Biol137:5]18
4. Heyting C, Dettmers RJ, Dietrich AJJ, Redeker EJW, VinkŽ .ACG 1988 Two major components of synaptonemal com-
plexes are specific for meiotic prophase nuclei. Chromosoma96:325]332
5. Dobson MJ, Pearlman RE, Karaiskakis A, Spyropoulos B,Ž .Moens PB 1994 Synaptonemal complex proteins: occur-
rence, epitope mapping and chromosome disjunction. J CellSci 107:2749]2760
6. Allen RL, O’Brien DA, Jones CC, Rockett DL, Eddy EMŽ .1988 Expression of heat shock proteins by isolated mousespermatogenic cells. Mol Cell Biol 8:3260]3266
Ž .7. Zakeri ZF, Wolgemuth DJ, Hunt CR 1988 Identification andsequence analysis of a new member of the mouse HSP70gene family and characterization of its unique cellular anddevelopmental pattern of expression in the male germ line.Mol Cell Biol 8:2925]2932
8. Rosario MO, Perkins SL, O’Brien DA, Allen RL, Eddy EM
449

J. Cobb and M. A. Handel
Ž .1992 Identification of the gene for the developmentallyŽ .expressed 70 kDa heat-shock protein P70 of mouse sper-
matogenic cells. Dev Biol 150:1]119. Dix DJ, Allen JW, Collins BW, Mori C, Nakamura N, Poor-
Ž .man-Allen P, Goulding EH, Eddy EM 1996 Targeted genedisruption of Hsp70-2 results in failed meiosis, germ cellapoptosis, and male infertility. Proc Natl Acad Sci USA93:3264]3268
Ž .10. Zhu D, Dix DJ, Eddy EM 1997 HSP70-2 is required forCDC2 kinase activity in meiosis I of mouse spermatocytes.Development 124:3007]3014
11. Allen JW, Dix DJ, Collins BW, Merrick BA, He C, Selkirk JK,Ž .Poorman-Allen P, Dresser ME, Eddy EM 1996 HSP70-2 is
part of the synaptonemal complex in mouse and hamsterspermatocytes. Chromosoma 104:414]421
Ž .12. Camerini-Otero RD, Hsieh P 1995 Homologous recombina-tion proteins in prokaryotes and eukaryotes. Annu Rev Genet29:509]552
Ž .13. Bishop DK 1994 RecA homologs Dmc1 and Rad51 interactto form multiple nuclear complexes prior to meiotic chromo-some synapsis. Cell 79:1081]1092
Ž .14. Plug AW, Xu JH, Reddy G, Golub EI, Ashley T 1996 Presy-naptic association of Rad51 protein with selected sites inmeiotic chromatin. Proc Natl Acad Sci USA 93:5920]5924
15. Moens PB, Chen DJ, Shen Z, Kolas N, Tarsounis M, HengŽ .HHQ, Spyropoulos B 1997 Rad51 immunocytology in rat
and mouse spermatocytes and oocytes. Chromosoma106:207]215
Ž .16. Fishel R, Wilson T 1997 MutS homologs in mammaliancells. Curr Opin Genet Develop 7:105]113
17. Baker SM, Plug AW, Prolla TA, Bronner CE, Harris AC, YaoX, Christie DM, Monell C, Arnheim N, Bradley A, Ashley T,
Ž .Liskay RM 1996 Involvement of mouse Mlh1 in DNA mis-match repair and meiotic crossing over. Nat Genet13:336]342
Ž .18. Hunter N, Borts RH 1997 Mlh1 is unique among mismatchrepair proteins in its ability to promote crossing-over duringmeiosis. Gene Develop 11:1573]1582
Ž .19. Brown L, McCarthy N 1997 DNA repair } A sense-ablresponse? Nature 387:450]451
20. Xu Y, Ashley T, Brainerd EE, Bronson RT, Meyn MS, Balti-Ž .more D 1996 Targeted disruption of ATM leads to growth
retardation, chromosomal fragmentation during meiosis, im-mune defects, and thymic lymphoma. Gene Develop10:2411]2422
Ž .21. Elledge SJ 1996 Cell cycle checkpoints: preventing an iden-tity crisis. Science 274:1664]1672
22. Keegan KS, Holtzman DA, Plug AW, Christenson ER, Brain-erd EE, Flaggs G, Bentley NJ, Taylor EM, Meyn MS, Moss SB,
Ž .Carr AM, Ashley T, Hoekstra MF 1996 The Atr and Atmprotein kinases associate with different sites along meioticallypairing chromosomes. Gene Develop 10:2423]2437
Ž .23. Zabludoff SD, Wright WW, Harshman K, Wold BJ 1996BRCA1 mRNA is expressed highly during meiosis andspermiogenesis but not during mitosis of male germ cells.Oncogene 13:649]653
24. Connor F, Smith A, Wooster R, Stratton M, Dixon A, Camp-Ž .bell E, Tait TM, Freeman T, Ashworth A 1997 Cloning,
chromosomal mapping and expression pattern of the mouseBrca2 gene. Hum Mol Genet 6:291]300
25. Scully R, Chen JJ, Plug A, Xiao YH, Weaver D, Feunteun J,Ž .Ashley T, Livingston DM 1997 Association of BRCA1 with
Rad51 in mitotic and meiotic cells. Cell 88:265]27526. Sharan SK, Morimatsu M, Albrecht U, Lim D-S, Regel E,
Ž .Dinh C, Sands A, Eichele G, Bradley A 1997 Embryoniclethality and radiation hypersensitivity mediated by Rad51 inmice lacking Brca2. Nature 386:804]810
27. Milner J, Ponder B, Hughes-Davies L, Seltmann M, KouzaridesŽ .T 1997 Transcriptional activation functions in BRCA2. Na-
ture 386:772]77328. Scully R, Anderson SF, Chao DM, Wei W, Ye L, Young RA,
Ž .Livingston DM, Parvin JD 1997 BRCA1 is a component ofthe RNA polymerase II holoenzyme. Proc Natl Acad Sci USA94:5605]5610
Ž .29. Wiltshire T, Park C, Caldwell KA, Handel MA 1995 Inducedpremature G2rM transition in pachytene spermatocytes in-cludes events unique to meiosis. Dev Biol 169:557]567
Ž .30. Goris J, Hermann J, Hendrix P, Ozon R, Merlevede H 1989Okadaic acid, a specific protein phosphatase inhibitor, in-duces maturation and MPF formation in Xenopus laevis oo-cytes. FEBS Lett 245:91]94
Ž .31. Cobb J, Reddy RK, Park C, Handel MA 1997 Analysis ofexpression and function of topoisomerase I and II duringmeiosis in male mice. Mol Reprod Dev 46:489]498
450