Embed Size (px)
Citation preview

Early development of Rhizobium-induced root nodules of Parasponia rigida. 11. Nodule morphogenesis and symbiotic development
SUSAN A. LANCELLE' A N D JOHN G. TOKREY' Cnbor Forrrlclnrion, Hnrvnrcl Ur~iversity, Perer:skorn, MA, U.S.A. 01366
Received June 8, 1984
LANCELLE, S. A., and J. G. TORREY. 1985. Early development of Rhizobilrrrl-induced root nodules of Pora.sl>orlicz rigicln. 11. Nodule morphogenesis and symbiotic development. Can. J. Bot. 63: 25-35.
The Rhizobiurn-induced root nodules of Pnrcrsporlin rigidn (Ulmaceae) outwardly resemble those formed on actinorhizal plants, being coralloid in shape and consisting of multiple, branched lobcs. The details of nodule morphogenesis also resemble more closely those which occur in an actinorhizal association than a typical Rhizobium-legume association and include prenodule formation, initiation of modified lateral roots which are termed nodule lobe primordia, and rhizobial colonization of tissues derived from the nodule lobe primordia to form the primary nodule lobes. Mature nodule lobe structure is actinorhizallike. Each lobe has an apical meristem and a central vascular cylinder which is surrounded by an uninfected inner cortex and then a zone of infected tissue. Peripheral to the infected tissue is an uninfected outer cortex. Infection threads and intercellular rhizobia progress continuously toward the apical mcristem but do not infect the meristcm itself. The establishment of the symbiosis in the host cells involves continuous thread formation after the initial infection until the host cells are nearly filled with rhizobia enclosed in threads. The rhizobia remain in threads throughout the symbiotic relationship and are not released from the threads as occurs in bacteroid formation in legumes.
LANCELLE, S. A , , et J . G. TORREY. 1985. Early development of Rhizobiur~l-induced root nodules of P~rn.sporlitr rigitln. 11. Nodule morphogenesis and symbiotic development. Can. J. Bot. 63: 25-35.
De forme corallo'ide et constituCs dc multiples lobes ramifiCs, les nodules radiculaires du Portrsporlin rigitln (Ulmaceae), induits par le Rhizobi~rrn ressemblent extkrieurement aux nodosites dcs plantes actinorhizkes. Lcs dCtails de la morphogCnkse des nodules ressemblent aussi a ceux que I'on voit dans une association actinorhiziennc plutBt qu ' i ccux d'une association Rhizobiurn-ICgumineuse typique; sont inclus: la formation de prCnodules, I'initiation dcs racines laterales modifites appelCes primordiums de lobes nodulaires et la colonisation rhizobienne des tissus dCrivCs des primordiums de lobes nodulaires pour former les lobes nodulaires primaires. La structure du lobe nodulaire mature cst d'aspect actinorhizien. Chaque lobe a un mCristkme apical et un cylindre vasculaire central entour6 d'un cortex interne non infect6 et d'une zone de tissu infectC. A la p6riphCrie du tissu infect6 se trouve le cortex externe non infect&. Des cordons d'infection et des rhizobiums intercellulaires progressent de f a ~ o n continue vers le mtristkme apical qui, cependant, Cchappe a I'infection. L'Ctablisscment de la symbiose dans les cellules de I'h8te nCccssite la formation continue de cordons aprks I'infection initiale et jusqu'i cc que les cellules soient presque remplies de rhizobiums inclus dans des cordons. Lcs rhizobiums demcurent dans Ics cordons pendant la durCe du rapport symbiotique et, contrairement a ce qui se passe au cours de la formation dc bactCro'ides chez Ics ICgumineuses, ils ne sont pas IibtrCs des cordons.
[Traduit par le journal]
Introduction Rhizobium-induced root nodules of the nonleguminous trop-
ical tree Parasponia (Ulmaceae) are of considerable interest in view of the unique combination of symbionts. Structural studies of mature nodules of P . rligosa (Trinick and Galbraith 1976), P . andersonii (Trinick 1979), and P . parvij7ora (Becking 1979) indicate that the root nodules resemble more closely the actinorhizal type induced by the actinomycete Frankia on various nonlegumes (Bond 1974) than those typi- cally formed on legumes in association with Rhizobium (Dart 1977). Mature Parasponia root nodules are coralloid in shape and consist of multiple, branched lobes. Like actinorhizal nodules, each nodule lobe has an apical meristem and a single central vascular bundle surrounded by the nodule cortex which includes the zones of infected cells (Trinick and Galbraith 1976; Trinick 1979). In contrast, legume root nodules, although varying in shape depending on host species, have in general a central infected zone surrounded by unin- fected cortical cells and numerous, usually peripheral, vascular traces (Dart 1977).
Although the studies by Trinick and Galbraith (1976), Trinick (1979), and Becking (1979) described the structure of
'Present address: Department of Botany, University of Massachu- setts, Amherst, MA, U.S.A. 01003.
'Author to whom reprint requests should be addressed.
mature nodules of Parasponia, there exists IIO published ac- count of early nodule development in this genus. Recent progress in aseptic germination of Parasponia seed and inocu- lation of the sterile seedlings with appropriate Rhizobilim strains (Becking 1983; Lancelle and Torrey 1984) has aided in the study of the earliest stages of nodule initiation. The inva- sion of seedling roots of Parasponia rigida by Rhizobium and the infection process have been described (Lancelle and Torrey 1984). In the present study, using correlated light and electron micrographs, we describe the early stages of nodule morpho- genesis and the development of the-symbiotic association between the rhizobial cells and the host cells of the root nodule. Although the root invasion and infection process resemble some aspects of Rhizobium-legume nodule initiation, the subsequent events more closely approximate the develop- mental sequence in actinorhizal nodule formation as described by Becking (1975), Angulo et al. (1975), Callaham and Torrey (1977), and Torrey and Callaham (1979). Thus Parasponia-Rhizobium nodules represent an intermediate condition between the two major types of symbiotic nodule systems, those of the legumes and those of actinorhizal plants.
Materials and methods Seed germination, seedling establishment, inoculation, and prepa-
ration of very young nodules for microscopy were carried out as described previously (Lancelle and Torrey 1984). Older nodules were
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y H
AR
VA
RD
UN
IVE
RSI
TY
HE
RB
AR
IA o
n 08
/30/
11Fo
r pe
rson
al u
se o
nly.

36. CAN. J . BOT. I IOL. 63, 1985
removed from roots of plants grown aeroponically (Zobel et 01. 1976) in one-quarter strength nitrogen-free Hoagland's solution (Hoagland and Arnon 1950) supplemented with 0.175 mM NH4N03, pH 6.5. Individual lobes were removed from the nodules and sliced open before fixation in 6% glutaraldehyde in 75 mM cacodylate buffer, pH 7.3, for 2 h at room temperature. Postfixation, dehydration, infil- tration, embedment, and staining for light and electron microscopy were carr~ed out as described previously (Lancelle and Torrey 1984).
Results Appearance of mature nodules
The root nodules of Parasponia rigida are coralloid in ap- pearance, as has been described for other species of Parasponia (Trinick and Galbraith 1976; Trinick 1979; Becking 1979). Figure I shows a relatively small nodule with multiple, branched lobes. Nodule roots, which are seen in some actino- rhizal plants (Bond 1974), were not observed.
Lateral root initiation Because the earlier structural studies (Trinick and Galbraith
1976; Trinick 1979; Becking 1979) had indicated that the nodule lobes were actually modified lateral roots as is the case in actinorhizal nodules (Becking 1975; Angulo et al. 1975; Callaham and Torrey 1977; Torrey and Callaham 1979), we examined normal lateral root initiation in P. rigida. Figure 2 shows a young lateral root which has just emerged through the parent root cortex. The young root has arisen in the pericycle and in this section is still surrounded by parental endodermis. No evidence of cortical participation in lateral root initiation was observed.
Prenodule development The infection process and early nodule initiation events in
P. rigida were described previously (Lancelle and Torrey 1984). In that account, the proliferation of subdivided cortical cells that occurred in response to rhizobial penetration of the epidermis via intercellular spaces was described as calluslike because of the randomlv oriented host cell divisions. In de- scribing nodule morphogenesis it is appropriate to term the calluslike mass of cells a "prenodule" to be consistent with the actinorhizal literature, where a prenodule is defined as an initial, unorganized proliferation of root cortical cells formed in response to the invasion of the endophyte (Becking 1975; CaIlaham and Torrey 1977).
The extent of development of the prenodule prior to lateral root initiation is evident in Fig. 3, where just a few pericyclic divisions have taken place. Serial sections of this entire speci- men showed no other pericyclic activity. As revealed by electron microscopy in the earlier study (Lancelle and Torrey 1984), rhizobia are present in some of the intercellular spaces of prenodule areas such as that shown in Fig. 3. Continued cell divisions in the prenodule result in its enlargement while cells of the pericycle and their derivatives independently divide at approximately the same level to give rise to a IateraI root (Fig. 4). This modified lateral root is termed a nodule lobe primordium, again to be consistent with the actinorhizal litera- ture (Callaham and Torrey 1977) and to distinguish it from a true lateral root. The nodule lobe primordium is very rootlike in appearance (see Fig. 1) and, like the true lateral root, does not appear to merge with subdividing inner cortical cells in these early stages (Figs. 4 and 5).
Usually only one prenodule and one nodule lobe primordium are formed in each nodule at the early stage of nodule formation (Fig. 4), but occasionally several prenodules and two or three
nodule lobe primordia form in close proximity (Fig. 5). Serial sections revealed that the young nodule shown in Fig. 5 had two nodule lobe primordia and five separate prenodule areas, two of which are visible in this section. Areas of intercellular invasion of rhizobia can be seen in the prenodules in Fig. 5, and although this particular specimen was not examined at the electron microscope level, evidence presented previously (Lancelle and Torrey 1984) showed that in other specimens at this stage of development, some of the prenodule cells were already infected by rhizobia and infection thread formation had begun to occur. Primordium-derived tissue remains uninvaded at this stage.
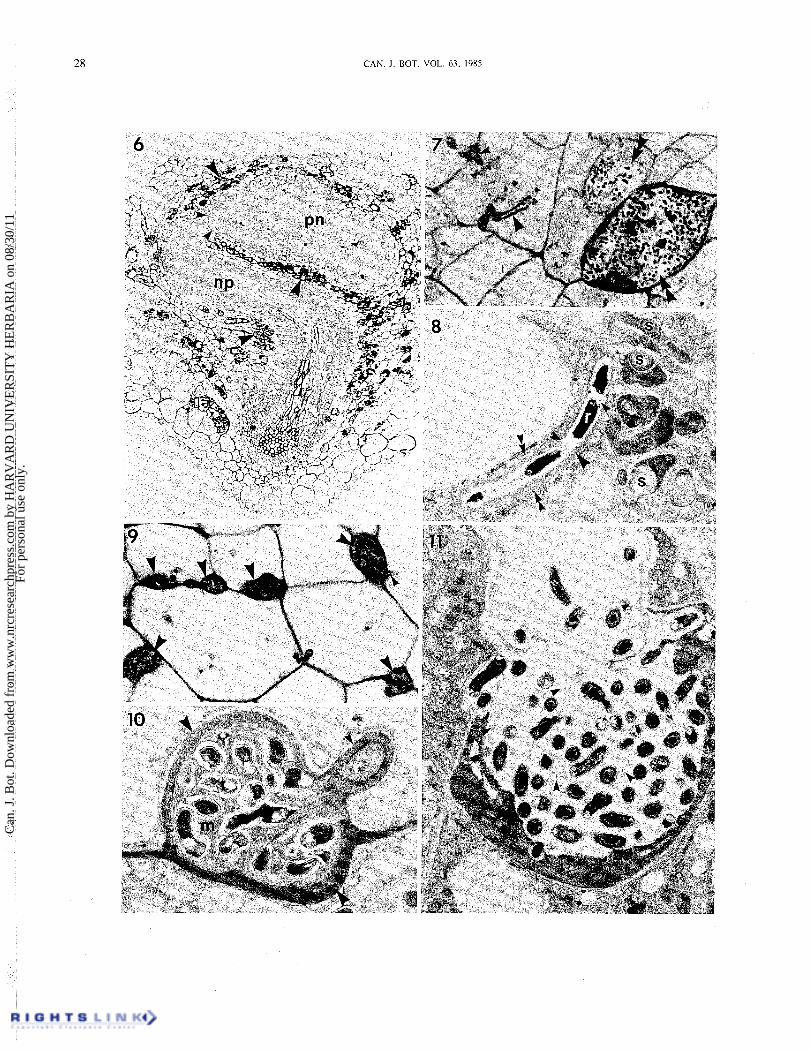
The prenodule continues to develop into a large mass of tissue as the nodule lobe primordium grows toward it and in some cases around it (Fig. 6). In Fig. 6, tissue derived from the primordium appears to merge with prenodule tissue, but this area has not yet been invaded by rhizobia. The prenodule and the primordium are both nearly surrounded by darkly staining cells containing phenolic compounds, except near the growing tip of the primordium.
Host-symbiont interactions in the well-developed pre- nodule are varied. Infection thread formation may be wide- spread, with a large number of the host cells infected in this way (Figs. 7 and 8). Figure 7 shows a dividing cell with infec- tion threads. The threads advance from cell to cell, or host cells are infected by rhizobia which progressively invade intercel- lular spaces (Figs. 7, 9, 10). Infection thread structure resem- bles that seen in the earliest stages (Lancelle and Torrey 1984), with rhizobia usually in single file (Fig. 8). The rhizobia are often surrounded by an electron-translucent zone and a small amount of thread matrix material. The rhizobia embedded in matrix are in turn surrounded by a thread wall and then a cytoplasmic membrane which delimits the thread from host cell cytoplasm. The host cytoplasm is dense with large numbers of organelles which include plastids with prominent starch grains (Fig. 8). In the prenodule stage, host cells containing infection threads may be highly vacuolate with a relatively small amount of peripheral cytoplasm (Fig. 9).
Rhizobia in intercellular spaces may proliferate to form enlarged local populations (Figs. 9, 10). When confined to intercellular spaces and surrounded by host wall material, the rhizobia are embedded in a homogeneous matrix (Fig. 10). Such large accumulations of rhizobia are in contrast to the orderly, single-file strands of rhizobia invading intercellular spaces in earlier stages of nodule initiation (~ancel le and Torrey 1984).
Commonly in large prenodules such as that shown in Fig. 6, another expression of enlarged populations of rhizobia occurs in which host cells appear to have been invaded and destroyed (Figs. 7 and 11). The rhizobia invade the cells in wide, un- walled masses (Fig. 1 l) , enlarged perhaps from a smaller inter- cellular population. A very sparse matrix material is present around the bacteria, which at this stage still look healthy. In Fig. 1 I , the host cell cytoplasm is very dense and appears to have degenerated in comparison with surrounding uninfected cells.
Development of the primary nodule lobe As nodule development continues, infection threads advance
toward the apex of the nodule lobe primordium, finally pene- trating cells derived from its apical region (Figs. 12, 13). In this manner of infection, the Parasponia-Rhizobium inter- action cIosely resembles that occurring typically in actinorhizal
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y H
AR
VA
RD
UN
IVE
RSI
TY
HE
RB
AR
IA o
n 08
/30/
11Fo
r pe
rson
al u
se o
nly.
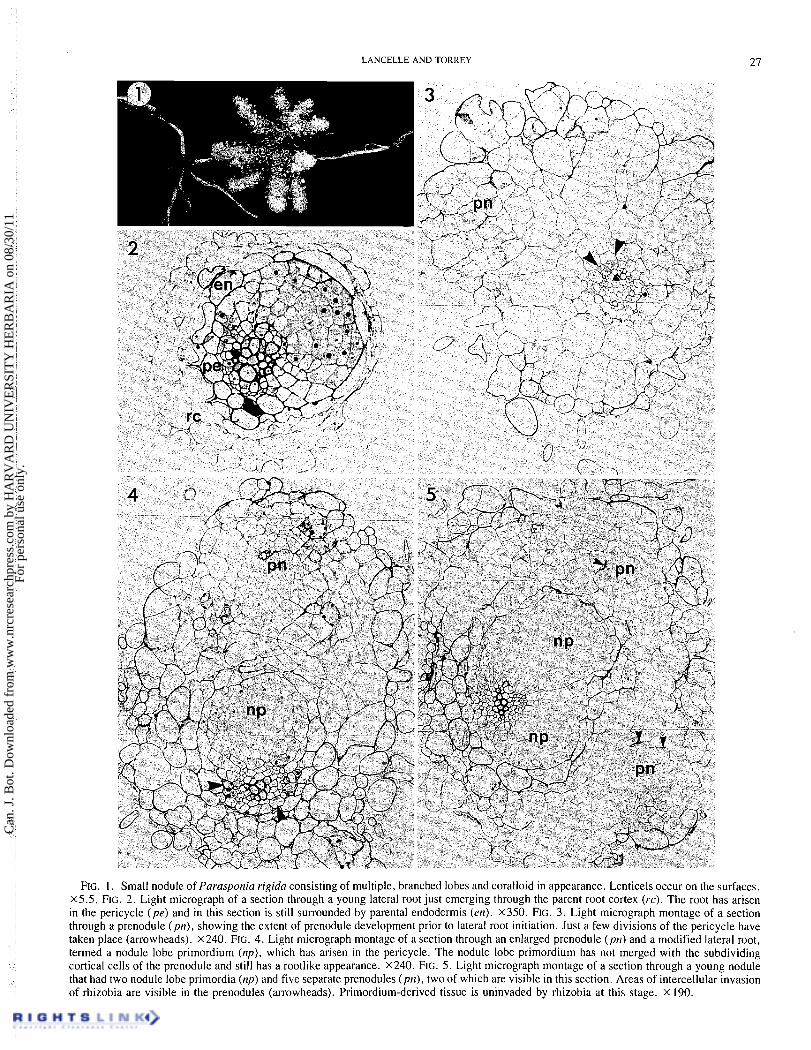
LANCELLE AND TORREY 27
FIG. 1. Small nodule of Parasponia rigida consisting of multiple, branched lobes and coralloid in appearance. Lenticels occur on the surfaces. X5.5. FIG. 2. Light micrograph of a section through a young lateral root just emerging through the parent root cortex (rc). The root has arisen in the pericycle (pe) and in this section is still surrounded by parental endodermis (en). X350. FIG. 3. Light micrograph montage of a section through a prenodule (pn), showing the extent of prenodule development prior to lateral root initiation. Just a few divisions of the pericycle have taken place (arrowheads). X240. FIG. 4. Light micrograph montage of a section through an enlarged prenodule (ptl) and a modified lateral root, termed a nodule lobe primordium (np), which has arisen in the pericycle. The nodule lobe primordium has not merged with the subdividing cortical cells of the prenodule and still has a rootlike appearance. x240. FIG. 5. Light micrograph montage of a section through a young nodule that had two nodule lobe primordia (np) and five separate prenodules (ptl), two of which are visible in this section. Areas of intercellular invasion of rhizobia are visible in the prenodules (arrowheads). Primordium-derived tissue is uninvaded by rhizobia at this stage. x 190.
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y H
AR
VA
RD
UN
IVE
RSI
TY
HE
RB
AR
IA o
n 08
/30/
11Fo
r pe
rson
al u
se o
nly.
CAN. 1. BOT. VOL. 63. I985
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y H
AR
VA
RD
UN
IVE
RSI
TY
HE
RB
AR
IA o
n 08
/30/
11Fo
r pe
rson
al u
se o
nly.

LANCELLE AND TORREY 29
nodules where the filamentous endophyte invades successive cortical cells of the nodule lobe primordium in a longitudinal progression toward the root apex (Callaham and Torrey 1977). Cells of the apical meristem itself were not observed to have been infected. It is at this point that the nodule lobe primordium can be considered to have been colonized by the symbiont and organization of the primary nodule lobe begins.
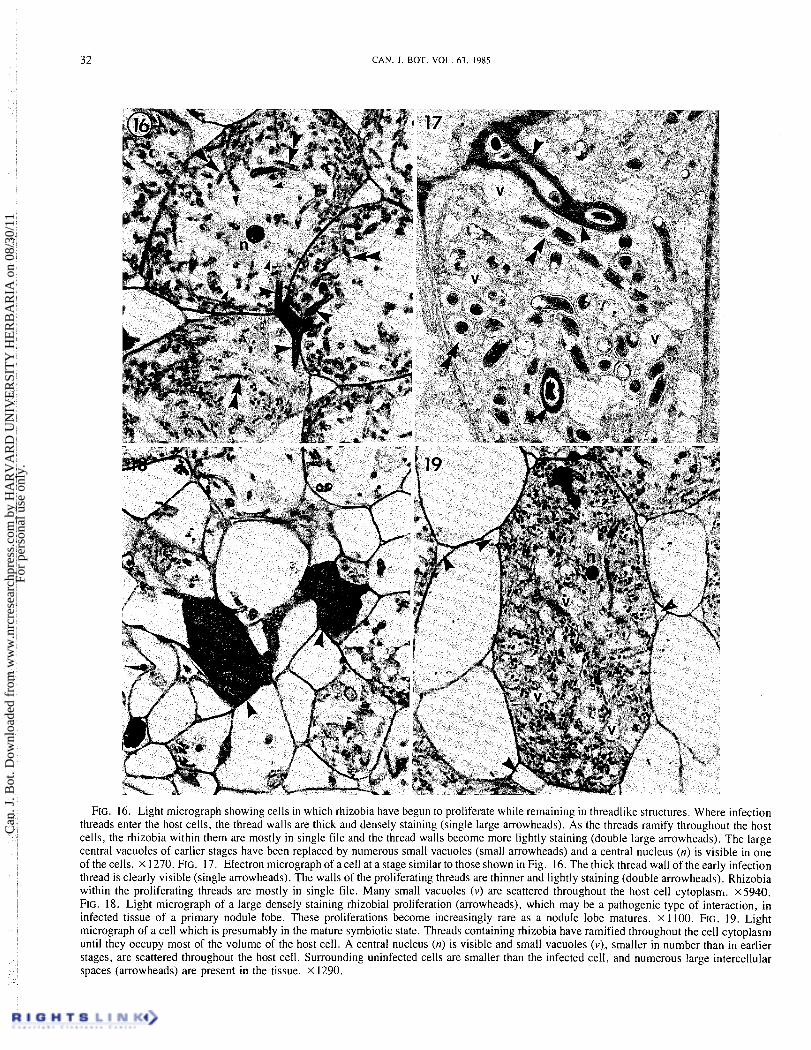
Rhizobia continue to proliferate within cells of the pre- nodule until most of the host cell volume is filled with bacteria (Fig. 12). Cell divisions cease in the prenodule'and it becomes
evident as large, very densely stained bacterial populations (Fig. 18). It is not known if these bacteria are capable of nitrogen fixation, but they are evidently a dead end in nodule development as colonization of host cells by infection thread formation and continued proliferation within thread structures is the prevailing host-symbiont interaction in the nodule lobe. Earlier in the prenodule stage this type of proliferation is relatively common, but it becomes increasingly rare as the lobe matures.
incorporated into the nodule lobe, although the prenodule actu- Establishment of the mature symbiotic state ally originated independently of the nodule lobe primordium. The threads containing dividing bacteria continue to ramify A layer of uninfected phenolic-containing cells surrounds the throughout the host cell cytoplasm until they occupy most of infected area except near the nodule apex. the volume of the cell (Figs. 19 and 20). The host cell cyto-
Closer examination of the host cells in the infected area of plasm is mainly around the cell periphery and contains large the primary nodule lobe reveals advancing threads near the numbers of mitochondria (Fig. 20). Plastids, when present, apex of the lobe have moved from cell to cell across cortical appear dense with little or no starch (Fig. 20). The cell nucleus cell walls (Figs. 14 and 15). Host cells can also be infected remains in a central position (Figs. 19 and 20) and may become from populations of intercellular bacteria (Fig. 15) as described multilobed (Figs. 20 and 21), but otherwise it appears healthy. for the prenodule stage (Figs. 9 , 10) with infection thread for- Small vacuoles appear to decrease in number and are scattered mation and no observed release of rhizobia from the thread. throughout the cell (Figs. 19 and 20). The infected cells have Newly infected cells are usually highly vacuolate with a large central vacuole and often many smaller ones around the periph- ery (Figs. 14 and 15). As was seen in the prenodule stage (Fig. 8), newly infected host cells may contain plastids with prominent starch grains (Figs. 14 and 15).
Where the infection threads enter the cells, they have thick, densely staining thread walls (Figs. 8 , 14, 15, 16, 17). As the rhizobia continue to proliferate within the host cells, they remain in threadlike structures and never have been ob- served being released from the threads as occurs in legumes (Bergersen 1974; Newcomb 1981). 'The threads are always delimited from host cell cytoplasm by a cytoplasmic mem- brane and usually have a relatively thin, lightly staining wall (Fig. 17). This distinction in thread types was also observed in other species of Parasponicz (Trinick and Galbraith 1976; Trinick 1979; Becking 1979). As the threads ramify throughout the host cells, the bacteria within them are most often in single file (Figs. 16 and 17). Small amounts of thread matrix material may be present, although this is not apparent in Fig. 17. The large vacuoles of the host cell are replaced progressively by smaller ones as bacterial proliferation continues (Figs. 16 and 17). Central nuclei are often visible (Fig. 16).
At this stage, remnants of the large pathogenic-type prolif- erations seen earlier in the prenodule (Figs. 7 and I I) are still
enlarged in comparison with nearby uninfected cells, which are highly vacuolate (Fig. 19) and may be comparable with the interstitial cells observed in soybean nodules (Bergersen and Goodchild 1973). Similar cells have been observed in some actinorhizal plants (Newcomb et al. 1978; Schwintzer et al. 1982). Large intercellular spaces are evident in the tissue (Figs. 19 and 20).
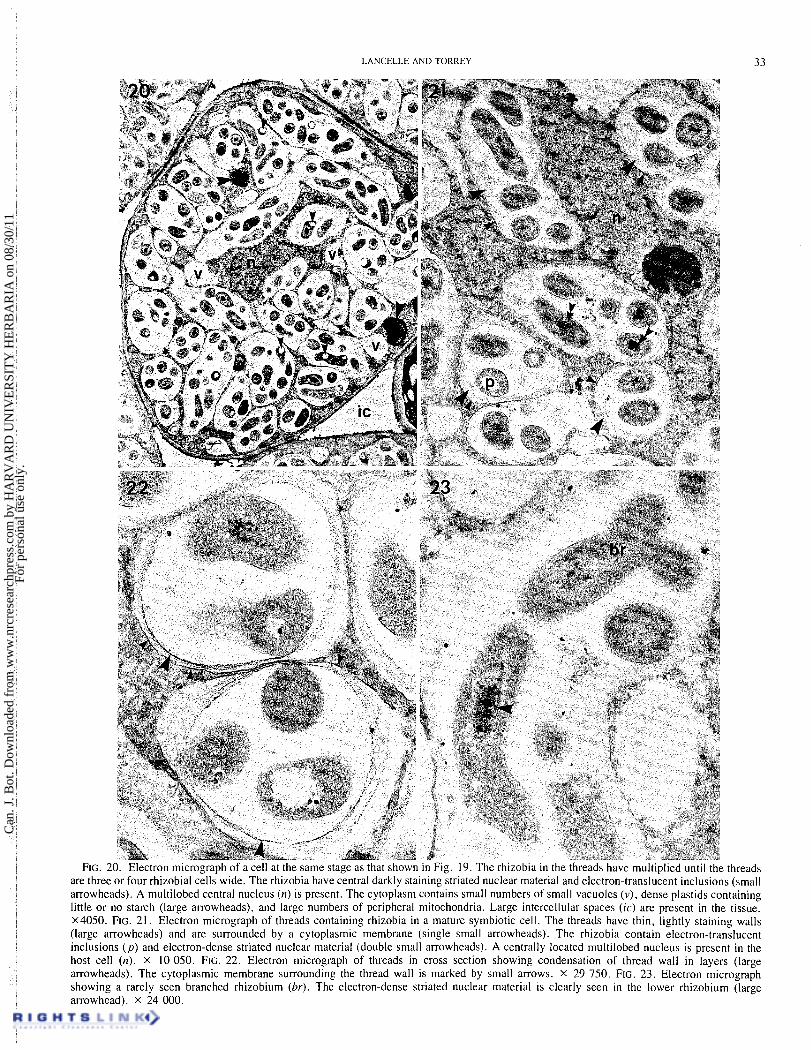
The threads widen because of rhizobial cell divisions and in the mature symbiotic state are three or four rhizobial cells wide (Figs. 20 and 21). The threads usually retain a lightly staining wall layer and are always surrounded by a cytoplasmic mem- brane (Figs. 2 0 , 2 1 , 2 2 ) . No "inner membrane" as described by Trinick (1979) for P. andersonii was observed in threads of P. rigidcz. However, the wall material sometimes condensed in layers, often leaving a densely staining layer surrounding the rhizobia (Fig. 22). This layer does not appear to have mem- brane structure and at present is ascribed to a fixation artifact.
Rhizobia were not observed released from the threads. The rhizobia d o not enlarge at this stage and remain mostly elon- gate and rod shaped (Figs. 20 and 21); very rarely a Y-shaped cell typical of bacteroids in some legumes (Newcomb 1981) was observed (Fig. 23). The rhizobia contain central darkly staining striated nuclear material and prominent electron- translucent inclusions considered to be poly-P-hydroxybutyrate
FIG. 6. Light micrograph of a section through a well-developed prenodule (pn) and nodule lobe primordium (np) which has curved around the prenodule. Tissue derived from the nodule lobe primoridum has begun to merge with the prenodule (small arrowheads), but this area is not yet invaded by rhizobia. Both the nodule lobe primordium and the prenodulc arc ncarly surrounded by a layer of cells containing darkly staining phenolic compounds (large arrowheads). X 120. FIGS. 7- 11. Scries of micrographs illustrating different types of host-symbiont interactions in a well-developed prenodule such as that shown in Fig. 6. Fig. 7. Light micrograph showing an infection thread (single large arrowhead) in a cell undergoing mitosis. The same cell is being infected from a population of intercellular rhizobia (small arrowhead). Some cells contain enlarged populations of rhizobia (double large arrowheads) that appear to have a deleterious effect on the host cells. X 1390. Fig. 8. Electron micrograph showing infection thread structure in early infected cells of a prcnodule. The rhizobia (r) are usually in single file and may be surrounded by a small amount of thread matrix material (single small arrowheads). 'The rhizobia embedded in the matrix are surrounded by a thread wall (single large arrowhead) and then cytoplasmic membrane (double small arrowheads) delimiting the thread from the host cytoplasm, which is dense with organelles and may contain plastids with prominent starch grains (s). x7200. Fig. 9. Light micrograph showing enlarged local populations of intercellular rhizobia (large arrowheads). Host cells may become infected from these populations via typical infection thread formation (small arrowhead). The early infected host cells are highly vacuolatc. X 1240. Fig. 10. Electron micrograph of an enlarged intercellular rhizobial population such as that shown in Fig. 9. The rhizobia are embedded in a homogeneous matrix ( m ) and arc surrounded by a thick host cell wall (large arrowheads). An infection thread (small arrow) has entered the cell from the intercellular population. X9540. Fig. I I. Electron micrograph of a large intracellular proliferation of rhizobia such as that shown in Fig. 7. The rhizobia have entered the cell in a wide, unwalled mass. The matrix material surrounding the rhizobia is very sparse (small arrowheads). The cytoplasm of the invaded cell is very dense, appearing degenerate in comparison with nearby uninfected cells. X7200.
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y H
AR
VA
RD
UN
IVE
RSI
TY
HE
RB
AR
IA o
n 08
/30/
11Fo
r pe
rson
al u
se o
nly.

CAN. J . BOT. VOL. 63, 1985
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y H
AR
VA
RD
UN
IVE
RSI
TY
HE
RB
AR
IA o
n 08
/30/
11Fo
r pe
rson
al u
se o
nly.

LANCELLE AND TORREY 3 1
(Craig et al. 1973) (Figs. 20, 2 1, 22, 23). (Becking 1975; Angulo et al. 1975; Callaham and Torrey 1977;
Structure o f the mature noclule lobe A section through a mature nodule lobe (Fig. 24) reveals an
organization that is actinorhizallike and agrees with the general description given for mature nodules of other Parczsponia spe- cies (Trinick and Galbraith 1976; Trinick 1979). The apex of the lobe is organized into a broad meristematic zone. Deriva- tives of the nodule lobe meristem may become infected either by advancing infection threads moving cell to cell or by popu- lations of intercellular bacteria, as described for earlier stages (Figs. 14 and 15).
As is seen in Fig. 24, the threads continue to ramify through- out the infected host cells, resulting in broad bands of cells which are nearly filled with threads. Each lobe has one central vascular cylinder which is surrounded by nodule cortex. An uninfected inner cortex surrounds the vascular cylinder and an uninfected outer cortex surrounds the infected tissue. In cross section, the infected tissue of P. rigida occurs in zones around the vascular cylinder (Tjepkema and Cartica 1982). In Fig. 24, an uninfected area is evident on the lower side of the vascular cylinder.
Discussion The structure of mature lobes of Parasponia rigida resem-
bles that of actinorhizal plants. The process of nodule mor- phogenesis also bears much more resemblance to that found in actinorhizal plants (Becking 1975; Angulo et al. 1975; Callaham and Torrey 1977; Torrey and Callaham 1979) than that described for legumes (Newcomb et al. 1979); however, there are also details that appear to be unique to Parasponia.
The prenodules which form in P. ridiga in response to rhi- zobial invasion and infection constitute a much more well- developed stage than that reported for actinorhizal plants (Becking 1975; Angulo et al. 1975; Callaham and Torrey 1977; Torrey and Callaham 1979), where the prenodule is a very transient stage. In P. rigida randomly oriented host root corti- cal cell divisions occur over an extended period of time as the rhizobia ramify throughout the prenodule, resulting in a large unorganized mass of tissue (Fig. 6).
Nodule morphogenesis in both actinorhizal plants and Parasponia involves bacterial colonization of tissues derived from a lateral root primordium, here termed the nodule lobe primordium, so that the nodule lobe is actually a modified lateral root. In Comptonia peregrina (Callaham and Torrey 1977) and Myrica gale (Torrey and Callaham 1979), both acti- norhizal plants, the innermost root cortical cells start sub- dividing almost simultaneously with nodule lobe primordium initiation and contribute to the tissues of the developing nodule lobe primordium. These tissues are very rapidly invaded by the endophyte which has been proliferating in the prenodule
Torrey and callaham 1979). In contrast, in P. rigida the nodule lobe primordium, like the
lateral root, arises only from the pericycle and its early devel- opment does not include involvement of inner root cortical cells. The well-developed prenodule, derived from the root cortex, remains independent from the nodule lobe primordium for a relatively long time before cells derived from the nodule lobe primordium itself are infected.
Once infection thread formation occurs in cells derived from the nodule lobe primordium, organization of the primary nodule lobe begins. Infection threads and intercellular invasion of rhizobia progress continuously toward the apical meristem, infecting newly derived cells but not the meristem itself. The infected host cells become progressively filled with rhizobia enclosed in threads. Scattered among the infected cells are smaller uninfected cells.
Dispersal of rhizobia in the host tissues appears to occur only by continuous infection of cells and not by host cell division after bacterial infection as occurs in some legume nodules (Dart 1977). In the organized lobe, host cell division was only ob- served in the lobe meristem. Cell divisions in the prenodule cease and it becomes incomorated into the nodule iobe. 'The infected tissue occurs in zones separated from each other by broad bands of uninfected parenchyma cells. This arrangement is in contrast to actinorhizal plants and was also noted for other species of Parasponia (Trinick and Galbraith 1976; Trinick 1979; Becking 1979).
Well-developed prenodules of P . rigidcz are surrounded by a layer of uninfected phenolic-containing cells and this continues in the mature lobe, where the infected zones of tissue are also surrounded by such cells. The function of these cells is unknown, but uninfected phenolic-containing cells are also present in many actinorhizal nodules and Newcomb et al. (1978) suggested for Comptonia roles for such cells that may include prevention of secondary infections by other bacteria or fungi-and prevention of herbivory.
'The prenodule is characterized by a variety of host- symbiont interactions, most commonly infection of host cells by intercellular bacteria and concomitant infection thread for- mation. Another type of interaction which is relatively com- mon in the prenodule stage is the large proliferation of rhizobia which may invade a cell and appear to destroy it (Fig. 1 1). This type of interaction does not appear to be the same i s the wide infection thread formation observed in mung bean (Newcomb and Mclntyre 1981), which was accompanied by typical bacte- roid release and had no discernible ill effects on host cell cytoplasm. The proliferation of rhizobia and apparent take-over of host cells observed in P. rigida may be an indication of an imbalance in the association such that the rhizobia are behaving more as pathogens than symbionts. Other such interactions
FIG. 12. Light micrograph montage of a section through a young nodule, showing that tissue derived from the nodule lobe primordium has been invaded by infection threads (boxed-in area) and organization of the primary nodule lobe has begun. Rhizobia in threads have multiplied in host cells of the prenodule area (pn) until some of them have become nearly filled with threads. The young nodule has a central vascular cylinder (vc) and the infected tissue is nearly surrounded by a layer of phenolic-containing cells (arrowheads). X 140. FIG. 13. Light micrograph at higher magnification of boxed-in area of Fig. 12, showing that infection threads (arrowheads) have advanced toward the meristematic apex (a) of the nodule lobe. The central vascular cylinder (vc) of the nodule lobe is visible in the upper part of the micrograph. X 325. FIG. 14. Light micrograph of early infected cells near the apex of a young nodule. Infection threads have moved from cell to cell across cortical cell walls (large arrowheads). Plastids containing starch (small arrowheads) are often visible in early infected cells, which also contain large central vacuoles. x 1280. FIG. 15. Light micrograph of a section through early infected cells in a young nodule. Infection threads have advanced from cell to cell by passing through cortical cell walls (large arrowhead). Host celIs can also be infected from populations of intercellular bacteria with de novo infection thread formation (double small arrowheads). Early infected host cells are highly vacuolate and may contain plastids with starch grains (single small arrowheads). X 1030.
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y H
AR
VA
RD
UN
IVE
RSI
TY
HE
RB
AR
IA o
n 08
/30/
11Fo
r pe
rson
al u
se o
nly.
CAN. J . DOT. VOL. 63, 1985
FIG. 16. Light micrograph showing cells in which rhizobia have begun to proliferate while remaining in threadlike structures. Where infection threads enter the host cells, the thread walls are thick and densely staining (single large arrowheads). As the threads ramify throughout the host cells, the rhizobia within them are mostly in single file and the thread walls become more lightly staining (double large arrowheads). The large central vacuoles of earlier stages have been replaced by numerous small vacuoles (small arrowheads) and a central nucleus (n) is visible in one of the cells. x 1270. FIG. 17. Electron micrograph of acell at a stage similar to those shown in Fig. 16. The thick thread wall of the early infection thread is clearly visible (single arrowheads). The walls of the proliferating threads are thinner and lightly staining (double arrowheads). Rhizobia within the proliferating threads are mostly in single file. Many small vacuoles (v) are scattered throughout the host cell cytoplasnl. x.5940. FIG. 18. Light micrograph of a large densely staining rhizobial proliferation (arrowheads), which may be a pathogenic type of interaction, in infected tissue of a primary nodule lobe. These proliferations become increasingly rare as a nodule lobe matures. X 1100. FIG. 19. Light micrograph of a cell which is presumably in the mature symbiotic state. Threads containing rhizobia have ramified throughout the cell cytoplasm until they occupy most of the volume of the host cell. A central nucleus (n) is visible and small vacuoles (v), smaller in number than in earlier stages, are scattered throughout the host cell. Surrounding uninfected cells are smaller than the infected cell, and numerous large intercellular spaces (arrowheads) are present in the tissue. X 1290.
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y H
AR
VA
RD
UN
IVE
RSI
TY
HE
RB
AR
IA o
n 08
/30/
11Fo
r pe
rson
al u
se o
nly.
LANCELLE AND TORREY 33
thre ads are three or four rhizobial cells wide. The rhizobia have central darkly staining striated nuclear material and electron-translucent inclusions (small arrowheads). A multilobed central nucleus (n) is present. The cytoplasm contains small numbers of small vacuoles (v) , dense plastids containing little or no starch (large arrowheads), and large numbers of peripheral mitochondria. Large intercellular spaces (ic) are present in the tissue. x4050. FIG. 21. Electron micrograph of threads containing rhizobia in a mature symbiotic cell. The threads have thin, lightly staining walls (large arrowheads) and are surrounded by a cytoplasmic membrane (single small arrowheads). The rhizobia contain electron-translucent inclusions (p) and electron-dense striated nuclear material (double small arrowheads). A centrally located multilobed nucleus is present in the host cell (n). x 10 050. FIG. 22. Electron micrograph of threads in cross section showing condensation of thread wall in layers (large arrowheads). The cytoplasmic membrane surrounding the thread wall is marked by small arrows. x 29 750. FIG. 23. Electron micrograph showing a rarely seen branched rhizobium (br). The electron-dense striated nuclear material 1s clearly seen in the lower rhizobium (large arrowhead). x 24 000.
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y H
AR
VA
RD
UN
IVE
RSI
TY
HE
RB
AR
IA o
n 08
/30/
11Fo
r pe
rson
al u
se o
nly.

34 CAN. J . BOT. VOL. 63. 1985
FIG. 24. Light micrograph montage of a longitudinal section through part of a mature nodule lobe. The apex of the lobe is organized into a meristematic zone (m). Some of the derivatives of the nodule meristem form a zone of early infected cells (ei) which may become infected by advancing infection threads moving cell to cell or by populations of intercellular bacteria. Beyond the zone of early infection is a zone of host cells containing ramifying threads (r t ) . The threads containing rhizobia continue to ramify throughout the host cells until most of the cell volume is occupied by threads and what is presumably the mature symbiotic state is achieved (ms). In this zone, enlarged infected host cells are interspersed with smaller uninfected cells. The zones of infected cells are surrounded by a layer of cells containing phenolic compounds (arrowheads). Peripheral to the infected zone is an uninfected outer cortex (oc). Surrounding the central vascular cylinder (vc) is an uninfected inner cortex (ic). In cross section, the infected cells occur in zones around the vascular cylinder; in this longitudinal section an uninfected area is visible in the lower part of the micrograph (uc). X 125.
were o b s e ~ e d in earlier stages of nodule initiation (Lancelle and Torrey 1984). It is interesting to note that as the nodule lobe matures, this type of interaction becomes increasingly rare and appears to be a dead end in nodule development. The establishment of true syn~biosis occurs with infection thread formation and subsequent ramification throughout host cells.
The composition of the thread wall at different stages in symbiotic growth is unknown and warrants further investi- gation. The observed differences in staining properties indicate a change in the nature of the wall material. Gresshoff et al. (1984) propose the term "fixation threads" for threads with
more lightly stained walls. However, while it is possible that the observed change in wall type coincides with the onset of nitrogenase activity, a possibility first pointed out by Trinick (1979), there is no direct evidence supporting this view nor the contention that nitrogen fixation occurs in one thread type only.
The lack of release of rhizobia from the threads is reminis- cent of the situation in actinorhizal plants, where the endophyte remains encapsulated by a material that appears to be pectic (Lalonde and Knowles 1975). Lack of rhizobial release from threads was also noted for P. andersonii (Trinick 1979). Trinick and Galbraith (1976) reported that some of the host
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y H
AR
VA
RD
UN
IVE
RSI
TY
HE
RB
AR
IA o
n 08
/30/
11Fo
r pe
rson
al u
se o
nly.

LANCELLE A N D TORREY 35
cells of P. rugosa contained released bacteroids, but electron microscopy was not utilized in this study and this detail should be reinvestigated.
The structure of the host cells changes as the symbiotic relationship becomes established. Early infected cells may be highly vacuolate, but the peripheral cytoplasm is dense with organelles, including plastids with starch grains (Figs. 8 , 14, 15). In mature symbiotic cells, the plastids are dense and con- tain little or no starch (Fig. 20). There is no large buildup of starch in infected cells during symbiotic development as is observed in many legumes (Newcomb 198 1). In actinorhizal plants (Newcomb et al. 1978; Schwintzer et 01. 1982; VandenBosch and Torrey 1983) starch may be present in plastids of early infected cells, but it decreases as the symbiotic relationship develops until in mature symbiotic cells little or no starch is present; starch may accumulate in nearby uninfected cells, but this was not observed for P. rigida.
As the threads ramify throughout the cells, large numbers of mitochondria occur in the peripheral cytoplasm, but the cell nuclei remain in a central position. The large central vacuoles are replaced by many small ones, which eventually decrease in number and are scattered throughout the cell. This sequence is in contrast both to some legumes, where a large central vacuole often is present in the mature symbiotic state (Newcomb 1976; Goodchild 1977), and actinorhizal plants, where many small vacuoles are located in the peripheral cytoplasm (Newcomb et al. 1978; Schwintzer et al. 1982; VandenBosch and Torrey 1983).
Acknowledgements 'This research was supported in part by the Maria Moors
Cabot Foundation for Botanical Research of Harvard Uni- versity. The authors thank P. K. Hepler for use of his labora- tory and the electron microscope facility in the Botany Department at the University of Massachusetts, D . A. Callaham for technical advice and assistance, R. Lundquist for care of plants in the greenhouse and F. O'Brien for secretarial assistance.
ANGULO, A. F., C. VAN DIJK, and A. QUISPEL. 1975. Symbiotic interactions in nonleguminous root nodulcs. 111 Symbiotic nitrogcn fixation in plants. Edited by P. S. Nutman. Cambridgc Univcrsity Press, Cambridge. pp. 474-484.
BECKING, J. H. 1975. Root nodules in nonlcgumcs. 111 Dcvclopmcnt and function of roots. Edited by J. G. Torrcy and D. T. Clarkson. Academic Prcss, London. pp. 508-566.
1979. Root nodule symbiosis bctwccn Rhizobi~rtn and Parasponia (Ulmaceae). Plant Soil, 51: 289-296.
1983. The Parasponia parvitlora - Rhizobi~rtn symbiosis. Host specificity, growth and nitrogcn fixation undcr various con- ditions. Plant Soil, 75: 309-342.
BERGERSEN, F. J ,1974. Formation and function of bactcroids. 111 Thc biology of nitrogen fixation. Edited by A. Quispel. North Holland Publ. Co., Amsterdam. pp. 473-498.
BERGERSEN, F. J., and A. J. GOODCHILD. 1973. Acration pathways in soybean root nodules. Aust. J. Biol. Sci. 26: 729-740.
BOND, G. 1974. Root nodule symbioscs with actinomycetc-likc organisms. It1 The biology of nitrogcn fixation. Edited by A. Quispel. North Holland Publ. Co., Amstcrdam. pp. 342-380.
CALLAHAM, D. A,. and J . G. TORREY. 1977. Prenodulc formation and primary nodulc dcvclopmcnt in roots of Cottlprot~icr (Myricaccac). Can. J . Bot. 55: 2306-2318.
CRAIG, A. S . , R. M. GREENWOOD, and K. I. WILLIAMSON. 1973. Ultrastructural inclusions of rhizobial bactcroids of lotus nodules and thcir taxonomic significancc. Arch. Mikrobiol. 89: 23-32.
DART. P. J . 1977. lnfcction and dcvclopmcnt of leguminous root nodulcs. 111 A trcatisc on dinitrogcn fixation. Scction 111. Biology. Edited by R. W. F . Hardy and W. S. Silvcr. John Wilcy and Sons, Ncw York, London, Sydncy, 'Toronto. pp. 367-472.
GOODCHILD, D. J. 1977. Thc ultrastructure of root nodulcs in rclation to nitrogcn fixation. Int. Rcv. Cytol. Suppl. 6: 235-288.
GRESSHOFF, M.. S. NEWTON. S. S. MOHAPATRA, K. F. SCOTT. S. HOWITT, G. D. PRICE, G. L. BENDER. J. SHINE, and B. G. ROLFE. 1984. Symbiotic nitrogcn fixation involving Rhizobi~rtn and thc non-lcgumc Pnrnsporlirr. In Advances in nitrogcn fixation research. Editerl by C. Vecgcr and W. E. Newton. Nijhoff/Junk, The Haguc; Pudoc. Wagcningcn. pp. 483-489.
HOAGLAND, D. R., and D. I. ARNON. 1950. The watcr-culturc mcthod for growing plants without soil. Rcviscd cd. Calif. Agric. Exp. Stn. Circ. 347.
LALONDE, M., and R. KNOWLES. 1975. Ultrastructurc, composition, and biogcncsis of thc encapsulation matcrial surrounding thc cndo- phytc in A111rr.s crispa var. rno1li.s root nodulcs. Can. J. Bot. 53: 1951-1971.
LANCELLE, S. A., and J. G. TORREY. 1984. Early dcvclopmcnt of Rhizobi~rrn-induced root nodulcs of Parcrspor~irr rigitln. 1.-lnfcction and carly nodulc initiation. Protoplasma, 123: 26-37.
NEWCOMB, W. 1976. A corrclatcd light and electron microscopic study of symbiotic growth and diffcrcntiation in Pislrrn scrrivrrtn root nodulcs. Can. J . Bot. 54: 2163-2186.
1981. Nodulc morphogencsis and diffcrcntiation. Int. Rcv. Cytol. Suppl. 13: 247-298.
NEWCOMB. W., and L. MCINTYRE. 1981. Dcvclopmcnt of root nodulcs of mung bean (Vigncr rcrditrtcr): a reinvestigation of cndo- cytosis. Can. J. Bot. 59: 2478-2499.
NEWCOMB, W., R. L. PETERSON, D. A. CALLA~IAM, and J. G. TORREY. 1978. Structurc and host -actinomycctc interactions in dcvcloping root nodules of Comptor~irr pt,regritlcr. Can. J. Bot. 56: 502-53 1 .
NEWCOMB, W., D. SIPPEL, and R. L. PETERSON. 1979. Thc carly morphogcncsis of Glycitlt~ tncr,r and Pislrrn .sotivrrtn root nodulcs-. Can. J. Bot. 57: 2603-2616.
SCHWINTZER, C. R., A. M. BERRY, and L. D. DISNEY. 1982. Scasonal pattcrns of root nodulc growth, cndophytc morphology, nitrogcnasc activity, and shoot dcvclopmcnt in Myricrr gtrle. Can. J. Bot. 60: 746-757.
TJEPKEMA, J. D., and R. J. CARTICA. 1982. Diffusion limitation of oxygcn uptakc and nitrogcnasc activity in thc root nodulcs of Parasponia rigida Mcrr. and Pcrry. Plant Physiol. 69: 728-733.
TORREY, J. G., and D. CALLAHAM. 1979. Early nodule dcvclopmcnt in Myriccl gale. Bot. Gaz. (Chicago) Suppl. 140: S 10-S 14.
TRINICK, M. J . 1979. Structurc of nitrogen-fixing nodulcs formed by Rhizobi~rtt~ on roots of Parcrsponin (~tld~r.sotlii Planch. Can. J. Microbiol. 25: 565-578.
TRINICK. M. J., and J . GALBRAITH. 1976. Structurc of root nodulcs formed by Rhizobi~rm on thc non-lcgumc Trpttrcl t~at~nabitla var. Scabra. Arch. Microbiol. 108: 159- 166.
VANDENBOSCH, K. A,, and J . G. TORREY. 1983. Host-endophyte intcractions in cffcctivc and incffcctivc nodules induced by the endophytc of Myrica gtrle. Can. J. Bot. 61: 2898-2909.
ZOBEL, R. W., P. DELTREDICI, and J. G. TORREY. 1976. A method for growing plants acroponically. Plant Physiol. 57: 344-346.
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y H
AR
VA
RD
UN
IVE
RSI
TY
HE
RB
AR
IA o
n 08
/30/
11Fo
r pe
rson
al u
se o
nly.





























