Embed Size (px)
Citation preview

Effect of spontaneous seizures onGABAA receptor a4subunit expression in an animalmodel of temporal lobe
epilepsy*Heidi L Grabenstatter, †MeaghanCogswell, *Yasmin CruzDel Angel, *Jessica Carlsen, *Marco
I. Gonzalez, *YogendraH. Raol, †Shelley J. Russek, and *‡AmyR. Brooks-Kayal
Epilepsia, **(*):1–8, 2014doi: 10.1111/epi.12771
Heidi Grabenstatter,postdoctoral fellow atthe University ofColorado AnschutzMedical Campus.
SUMMARY
Objective: Temporal lobe epilepsy (TLE) is frequently medically intractable and often
progressive. Compromised inhibitory neurotransmission due to altered c-aminobu-
tyric acid (GABA)A receptor a4 subunit (GABAARa4) expression has been emphasized
as a potential contributor to the initial development of epilepsy following a brain insult
(primary epileptogenesis), but the regulation of GABAARa4 during chronic epilepsy,
specifically, how expression is altered following spontaneous seizures, is less well
understood.
Methods: Continuous video–electroencephalography (EEG) recordings from rats with
pilocarpine-induced TLE were used to capture epileptic animals within 3 h of a sponta-
neous seizure (SS), or >24 h after the last SS, to determine whether recent occurrence
of a seizure was associated with altered levels of GABAARa4 expression. We further
evaluated whether this GABAARa4 plasticity is regulated by signaling mechanisms
active in primary epileptogenesis, specifically, increases in brain-derived neurotrophic
factor (BDNF) and early growth response factor 3 (Egr3).
Results: Elevated levels of GABAARa4 messenger RNA (mRNA) and protein were
observed following spontaneous seizures, and were associated with higher levels of
BDNF and Egr3mRNA.
Significance: These data suggest that spontaneous, recurrent seizures that define
chronic epilepsy may influence changes in GABAARa4 expression, and that signaling
pathways known to regulate GABAARa4 expression after status epilepticus may also
be activated after spontaneous seizures in chronically epileptic animals.
KEYWORDS: Spontaneous seizure, GABAA receptor, Pilocarpine.
Fast synaptic inhibition is mediated primarily by postsyn-aptic c-aminobutyric acid (GABA)A receptors (GABAARs).The most abundant receptor subtype, a1b2c2, is localized at
the synapse, and mediates phasic inhibition.1,2 Alterationsin subunit expression and composition that affect the locali-zation, function, and pharmacology of GABAARs havebeen demonstrated during primary epileptogenesis and fol-lowing the onset of spontaneous seizures in animal modelsof epilepsy,3–9 as well as in tissue resected from patientswith intractable temporal lobe epilepsy (TLE).10,11 Statusepilepticus (SE) results in changes in the expression andmembrane localization of several GABAAR subunits (e.g.,a1, a4, c2, and d) in hippocampal dentate granule cell neu-rons; specifically, a1 and d subunit expression decreasesand a4 and c2 subunit expression increases, resulting in achange in receptor subunit composition and localization inthese neurons7,12,13 that is predicted to impair inhibitory
Accepted July 31, 2014.*Department of Pediatrics, Section of Neurology, Translational Epilepsy
Research Program, University of Colorado, AMC, Aurora, Colorado,U.S.A.; †Department of Pharmacology and Experimental Therapeutics,Programs in Biomolecular Pharmacology and Neuroscience, BostonUniversity School of Medicine, Boston, Massachusetts, U.S.A.;and ‡Children’s Hospital Colorado, Aurora, Colorado,U.S.A.
Address correspondence to Amy R. Brooks-Kayal, Pediatrics, Divisionof Neurology, University of Colorado, AMC, Pharmacy Building (V20),Mail Stop 8605, 12850 E. Montview Blvd., Aurora, CO 80045, U.S.A.E-mail: [email protected]
Wiley Periodicals, Inc.© 2014 International League Against Epilepsy
1
FULL-LENGTHORIGINALRESEARCH

function.14 Preventing these changes using viral gene trans-fer can inhibit development of epilepsy in the pilocarpinemodel of TLE.15
The signaling mechanisms that mediate GABAAR a4subunit (GABAARa4) plasticity after status epilepticus (SE;in primary epileptogenesis) have been defined. Increases inGABAARa4s are transcriptionally regulated by brain-derived neurotrophic factor (BDNF) activation of thetropomyosin-receptor-kinase B (TrkB) receptor, its down-stream signaling cascades (protein kinase C [PKC] andmitogen-activated protein kinase [MAPK]), as well asupregulation of its target (early growth response factor 3[Egr3]).16 Moreover, levels of both BDNF and Egr3increase,12,16,17 and binding of Egr3 to the Egr response ele-ment (ERE) in the endogenous GABAARa4 gene (Gabra4)increases17 following SE. Whether similar mechanismsmediate changes in GABAARa4 during chronic epilepsy,and the role, if any, spontaneous seizures may play in thisregulation remain unknown.
The current studies investigate whether GABAARa4 pro-tein expression in the dentate gyrus of chronically epilepticrats is altered following recent spontaneous seizure (SS),and if this is regulated by activation of signal transductionpathways similar to those that are activated during primaryepileptogenesis. We now report for the first time increasesin levels of BDNF and GABAARa4 within 3 h after SS thatparallel those seen following SE, and that increases inGABAARa4 following SS are transient, but associated withoverall higher mean seizure frequencies.
Materials and MethodsPilocarpine injections
SE was induced in adult Sprague-Dawley rats (Charles-River Labs, Kingston, PA, U.S.A.) as described previ-ously12 using 385 mg/kg intraperitoneal (i.p.) pilocarpineafter pretreatment with scopolamine (1 mg/kg, i.p.), andfollowed by diazepam (6 mg/kg, i.p.) 1 h after onset ofSE, with additional 3 mg/kg every 2 h as needed to ablatepersistent motor seizures. Age-matched control ratsreceived subconvulsive doses of pilocarpine (38.5 mg/kg,i.p.), did not develop SE or spontaneous seizures, and werehandled and housed identically to the epileptic animals.All animal experiments were approved by the institutionalanimal care and use committee at the University of Colo-rado Anschutz Medical Campus, and approved and con-ducted in accordance with the United States Public HealthService’s Policy on Humane Care and Use of LaboratoryAnimals.
Surgical implantation of electrodesChronically epileptic rats (4 weeks post-SE) and controls
were implanted with two stainless steel screws placed 4 mmcaudal to bregma and 2.5 mm lateral from midline bilat-erally, with reference and ground electrodes placed bilater-
ally behind lambda, as described previously.18Animals wereallowed to recover from surgery for 1 week before proceed-ing with any further experimentation.
EEG acquisition and analysisRats subjected to pilocarpine-induced SE and control
rats were video–electroencephalography (EEG) recorded24 h per day using a Pinnacle digital video-EEG systemwith flexible cables to monitor seizure frequency asdescribed previously.18 EEG signals were sampled at1 kHz, amplified by 1009, and band-pass filtered between0.3 Hz and 600 Hz. Prior to inclusion in the study, all pilo-carpine-treated SE animals were confirmed to have at leasttwo EEG-documented SS and thus meet criteria for epi-lepsy. EEG records of these chronically epileptic animalswere then reviewed daily, and animals were sacrificed ifthey had (1) experienced a seizure within the past 3 h(recent SS or <3 h group) or (2) had been seizure free for>24 h (no recent SS or >24 h group). After sacrifice, andtissue collection, electrographic recordings spanning the7 days prior to sacrifice were examined visually by atrained technician blinded to all experimental parametersto determine the number of electrographic seizures thathad occurred for each animal. Electroencephalographic sei-zures were differentiated from background noise by theappearance of large-amplitude (>3 times baseline), high-frequency (>5 Hz) activity, with progression of the spikefrequency lasting >10 s. Seizure quantification was con-ducted according to seizure number and type (convulsiveor nonconvulsive). Motor seizures were scored by standardbehavioral classes,19 with Racine class 3 or greater seizurescategorized as convulsive seizures and class 2 or belowconsidered nonconvulsive.
Real-time (RT)-PCR andWestern blotsEpileptic animals (56.21 � 13.99 days after pilocarpine-
induced SE) confirmed to have had at least two SS were sac-rificed <3 h from the time of an SS or >24 h after the lastSS. Tissue from age-matched controls was analyzed toassess changes in protein and messenger RNA (mRNA)potentially induced by spontaneous seizure activity. RNAwas extracted from microdissected hippocampal dentategyrus tissue using RNeasy Mini RNA extraction kit (QiagenVenlo, The Netherlands). Primers and probe for BDNF(Rn02531967),Gabra4 (Rn00589846_m1), and cyclophilin(Rn00690933_m1) were purchased from Applied Biosys-tems (Foster City, CA, U.S.A.). Primers and probes for Egr3were designed using primer express software. Primer andprobe sequences for Egr3 were the following: Egr3 forward50-GAGATCCCCAGCGCGC-30, Egr3 reverse 50-CATCTGAGTGTAATGGGCTACCG-30, and Egr3 TaqMan 50-CAACCTCTTCTCCGGCAGCAGTGAC-30. Samples wererepeated in duplicate, with each reaction split into two wellsin a total volume of 20 ll containing 16 ng of RNA usingan ABI Prism 7900HT machine (ABI Biosystems, Foster
Epilepsia, **(*):1–8, 2014doi: 10.1111/epi.12771
2
H. L. Grabenstatter et al.

City, CA, U.S.A.). Polymerase chain reaction (PCR)cycling parameters were 50°C for 30 min, 95°C for 10 min,50 cycles of 95°C for 15 s, and 60°C for 1 min. All valueswere normalized to cyclophilin expression to control forloading variability and expressed as fold change withrespect to the mean control values (defined as 1).
Western blot was performed on protein (25 lg forGABAARa4; 30 lg for Egr3) extracted from microdissecteddentate gyrus (DG) as previously published.15 Membraneswere incubated with rabbit polyclonal antibodies raisedagainst GABAARa4 (anti-GABARa4, Millipore, Billerica,Massachusetts, U.S.A. AB5457; Research Research Identi-fier (RRID): AB_177479; 1:2,000) in 1% milk/Tris-Buf-fered Saline and Tween 20 (TBST) or Egr3 (anti-Egr-3 (H-180) N –terminus, sc22801; Research Research Identifier(RRID): AB_2097199; 1:400; Santa Cruz, Dallas, Texas,U.S.A.) in 2% milk/Tris-buffered Saline and Tween 20(TBST) overnight in 4°C. Membranes were then washedand incubated with anti-rabbit secondary antibody(1:10,000 in 1% milk/TBST for GABAAR; GE Healthcare,Little Chalfont, Buckinghamshire, United Kingdom) or(Veryblot, Abcam, Cambridge, United Kingdom AB131366;1:500 in 2% milk/TBST for Egr3) for 1 h at room tempera-ture. Bands were detected using chemiluminescent solution(Pierce, Rockford, IL, U.S.A.), and membranes werestripped and reprobed with rabbit polyclonal antibodyraised against b-actin (1:40,000, Sigma, St. Louis, MO,U.S.A.) in 1% milk/TBS-T overnight. GABAARa4 andEgr3 values were normalized to b-actin expression in thesame samples and expressed as percent change relative tomean control values in the same run (defined as 1). Densi-tometry was performed with National Institutes of Health(NIH, Bethesda, MD, U.S.A.) Image J version 1.42q(RRID:nif-0000-30467). Statistical significance was calcu-lated with Prism software (Irvine, CA, U.S.A.) (RRID:rid_000081) using a one-way analysis of variance (ANO-VA) with a Tukey’s test for multiple comparisons or anunpaired Student’s t-test as indicated.
Chromatin immunoprecipitationRats were perfused with cold 19 phosphate-buffered
saline (PBS; with 1:1,000 phosphatase inhibitors) fol-lowed by 4% formaldehyde. Whole brains wereimmersed overnight in 4% formaldehyde and sectioned at600 lm. Microdissected DG was sonicated to produce300–500 bp fragments of crosslinked protein–DNA com-plexes and precipitated using an Egr3 specific antibody(10 lg; Santa Cruz Biotechnology) or normal rabbitimmunoglobulin G (IgG; 2 lg; Santa Cruz Biotechnol-ogy) to account for nonspecific DNA pull down.Immunoprecipitated protein–DNA complexes werereverse-crosslinked and purified for polymerase chainreaction (PCR) analysis using the protein A–conjugatedmagnetic bead ChIP kit (Millipore). Quantitative real-time RT-PCR was performed using primers that flank the
Egr3 binding site in the Gabra4 promoter and a TaqManprobe: forward, 50- GAACAAACTTGCCTAGCTTCGCGT-30; reverse, 50- TCCTCCAGCCTAGCCGC-30; TaqManprobe, 50- AAGTTCACCGGCGAGCAGCGCTTTCA-30.Data were normalized to input signal [Egr3/Input] andexpressed as fold change with respect to control (definedas 1). Input represents the DNA signal from sample prep-aration prior to immunoprecipitation.
ResultsRegulation of GABAARa4 protein by spontaneousseizure activity
Previous experiments have demonstrated that increasedtranscription of Gabra4 following prolonged pilocarpine-induced seizures (SE) is mediated by binding of Egr3 to theGabra4 ERE site in the core promoter region 24 h followingSE.17 For the first time, we evaluated whether increasedGABAARa4 expression may also occur during secondaryepileptogenesis in response to acute SS. Animals sacrificedwithin 3 h of a recent SS had significantly higher GABARa4protein levels than control rats (Fig. 1A and 1C), yet epilep-tic rats that had not had a seizure for >24 h haveGABAARa4 protein levels similar to control rats (Fig. 1Band 1C). Most importantly, epileptic rats having an SSwithin the last 3 h also had significantly higher GABAARa4protein levels when compared to epileptic rats without a sei-zure for >24 h (Fig. 1C), suggesting that recent SS activityinduces increased GABAARa4 protein expression in chroni-cally epileptic animals. Continuous video-EEG collectedfrom the same animals for 7 days prior to sacrifice was ana-lyzed to determine whether there was a difference in overallseizure frequency between the two groups. Video-EEG datademonstrated that rats sacrificed <3 h after SS, which hadhigher levels of GABAARa4 protein expression, also hadsignificantly higher daily seizure frequency than the rats sac-rificed >24 h after the last seizure (Fig. 1D). These resultssuggest that spontaneous seizures may induce a cascade ofevents that increase GABAARa4 on a relatively short time-scale, and that animals with frequent SS (i.e., more severeepilepsy) may have higher overall GABAARa4 levels.
Spontaneous seizures increase GABAARa4, BDNF, andEgr3 mRNA levels
Evaluation of the mechanisms leading to increasedGABAARa4 protein levels demonstrated that rats with arecent SS had significantly higher levels of GABAARa4mRNAs in the DG than controls (Fig. 2A), whereas epilep-tic rats with no seizures for >24 h had GABAARa4 mRNAlevels similar to controls. Levels of BDNF mRNAs in theDG were increased fivefold within 3 h of SS in chronicallyepileptic rats relative to control rats (Fig. 2B). BDNFmRNA levels in epileptic rats with no SS within 24 h ofsacrifice were similar to controls, suggesting changes inBDNF expression may be driven by recent seizure activity
Epilepsia, **(*):1–8, 2014doi: 10.1111/epi.12771
3
Spontaneous Seizures and GABAARa4 Expression

rather than being a residual effect of SE, or inherent to theepileptic state. Because BDNF has been shown previouslyto regulate GABAARa4 expression via its control overEgr3 levels, and the binding of Egr3 to the Gabra4 corepromoter after SE,16 we examined whether Egr3 mRNAlevels also change following recent SS. Levels of Egr3mRNAs increased by greater than eightfold following arecent (<3 h) spontaneous seizure and were significantlyhigher than in epileptic rats with no seizures for >24 h(Fig. 2C). However, the 14.37% increase in mean Egr3protein expression following recent spontaneous seizures(i.e., in the <3 h SS group) in the DG was not significantlydifferent from that of controls. Chromatin immunoprecipi-tation was used to examine whether increased binding ofEgr3 protein to the promoter of the Gabra4 subunit geneoccurred within 3 h of a spontaneous seizure and mightaccount for the increased GABAARa4 mRNA levels.Although Egr3 protein binding to the ERE promoter of theGabra4 subunit gene was observed in the DG followingspontaneous seizures, the levels were not significantly dif-ferent from those seen in controls (Fig. 2D).
Changes in seizure semiology related to GABAARdifferences
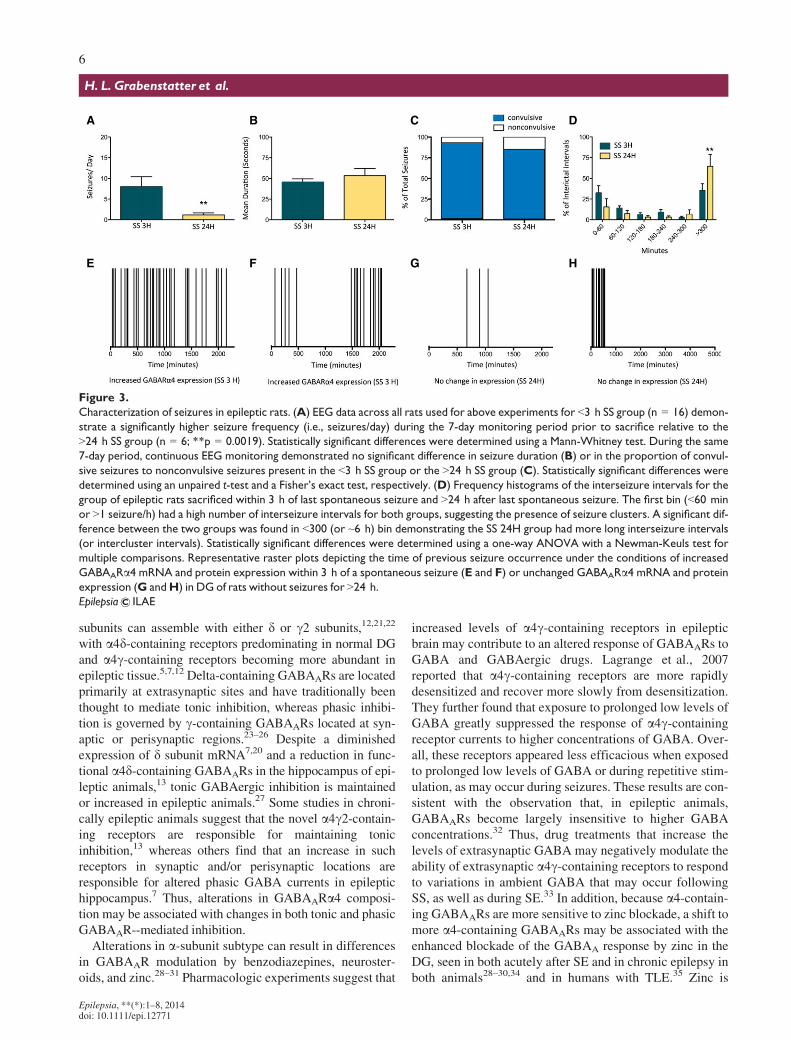
Continuous video-EEG monitoring was used to charac-terize seizure frequency, seizure type, and temporal distri-bution for 7 days before sacrifice (Fig. 3). Rats sacrificedwithin 3 h of a recent seizure had significantly higher
seizure frequencies overall than rats sacrificed >24 h afterthe last spontaneous seizure (Fig. 3A), suggesting they hadmore severe epilepsy in general. However, despite the dif-ference in seizure frequency between the <3 h SS and>24 h SS groups, each group had similar seizure durationsand severity (i.e., convulsive vs. nonconvulsive [Fig. 3Band 3C]). Of interest, rats sacrificed >24 h after SS, hadsignificantly more prolonged interictal intervals (defined as>300 min or ~6 h) than animals sacrificed <3 h after SS(3D). Increases in GABAARa4 mRNA and protein levelsoccurred following high frequency seizure clusters with orwithout prolonged intercluster intervals (Fig. 3A, 3E, and3F), suggesting that spontaneous seizure clusters caninduce GABAARa4 plasticity even when they occur infre-quently. High frequency seizure clusters were more com-mon in <3 h SS animals, but were present in some of the>24 h SS group; however, the group consistently hadGABAARa4 mRNA and protein levels similar to controlrats when sacrificed following long periods of seizure free-dom (i.e., long interictal intervals; Fig. 3G–H). This sug-gests that GABAARa4 levels are plastic, increasingtransiently following a spontaneous seizure and then fallingagain after a long period of seizure freedom.
DiscussionThe results of these studies are the first demonstration
that spontaneous seizures may acutely regulate GABAARa4
A B
C D
Figure 1.
Spontaneous seizures increase GABAARa4 protein expression in DG. Western blots of DG protein homogenates of control rats or epi-
leptic rats sacrificed within (A) 3 h of a spontaneous seizure (n = 6 controls, n = 6 SS) or (B) >24 h after last spontaneous seizure
(n = 3 controls, n = 4 No SS) reacted with anti-a4 and anti-Actin antibodies. (C) Densitometry analysis demonstrates that rats with
recent seizure activity show ~23% increase in GABAARa4 expression relative to controls (p < 0.01) and ~20% increase relative to epilep-
tic rats without recent spontaneous seizures (p < 0.05) using a one-way ANOVAwith Tukey’s test for multiple comparisons. Normalized
data are presented as mean � standard error of the mean (SEM) and expressed as percent change. (D) The same rats that express
increased GABAARa4 protein levels in DG following a recent SS (i.e., <3 h SS; SS 3H, n = 6) demonstrate a significantly higher seizure fre-
quency (i.e., seizures/day) during the 7-day monitoring period prior to sacrifice relative to the >24 h SS group (SS 24H, n = 4;
*p = 0.0139). Significant differences in seizure frequency were detected using a Mann-Whitney test. For all graphs, error bars,
mean � SEM.
Epilepsia ILAE
Epilepsia, **(*):1–8, 2014doi: 10.1111/epi.12771
4
H. L. Grabenstatter et al.

RNA and protein expression, suggesting that spontaneousseizures (regardless of whether convulsive or nonconvul-sive) result in acute transcriptional upregulation of theGabra4 gene in the DG of epileptic rats.
Spontaneous seizures, GABAARa4 increases, andchronic epilepsy severity
Increased levels of GABAARa4 RNAs and protein wereassociated with higher overall seizure frequencies in epilep-tic rats sacrificed within 3 h of an SS. These data couldsupport two different, but not mutually exclusive, interpre-tations. One interpretation is that in chronic epilepsy thespontaneous, recurrent seizures drive GABAARa4 increasesvia acute activation of cellular signaling pathways after eachepileptic event (i.e., reactive plasticity of GABAAR subun-its). Long periods of seizure freedom allow the system to“reset” and GABAAR subunit levels to return to control lev-els until an SS again induces GABAARa4 increases. In
addition, it is possible that the increased seizure frequencyobserved in these animals may, in part, be a consequence oflong-term changes in GABAA-R subunit expression. Themost parsimonious explanation of our data is that these sce-narios exist together, with spontaneous seizures themselvesstimulating the change in GABAARa4 subunit expression,and that these alterations, in turn, may contribute to higheroverall seizure frequencies that then perpetuate and/or exac-erbate the GABAAR alterations.
Changes in GABAAR function in chronically epilepticrats with spontaneous seizures
Increases in GABAARa4 expression have been demon-strated in epileptic animals in several TLE models.3,5,7,20
The functional consequences of these increases inGABAARa4 levels depend in large part on what othersubunits it pairs with to compose a receptor. Immunoprecipi-tation and immunogold labeling studies suggest that a4
A B
C D
Figure 2.
Spontaneous seizures increase GABAARa4, BDNF, and Egr3 mRNA expression in DG. (A) Quantification of RT-PCR analysis of
GABAARa4 mRNA expression in DG <3 and >24 h after SS (controls, n = 4; SS 3H, n = 5; SS 24H, n = 4) demonstrates a significant
(p < 0.05) increase in GABAARa4 mRNA levels following recent spontaneous seizure activity (within 3 h) relative to control rats. Epilep-
tic rats that had not had a seizure for >24 h have GABAARa4 mRNA levels similar to controls. (B) DG from epileptic rats with recent SS
(within 3 h, n = 8) showed significant (p < 0.05) increases in BDNF mRNA expression compared to controls (n = 7), and compared to
rats without SS for >24 h (n = 6). (C) DG from epileptic rats with recent SS (within 3 h) showed significant (p < 0.05) increases in Egr3
mRNA expression compared to rats that had not had an SS for >24 h and controls (controls, n = 4; SS 3H, n = 8; SS 24H, n = 4). Expres-
sion of all mRNAs was normalized to cyclophilin and expressed as fold change compared to controls (defined as 1). (D) Quantification of
protein–DNA binding in DG <3 h and >24 h after SS (controls, n = 3; SS 24H, n = 4) demonstrates no difference in Egr3 protein binding
to the ERE promoter of Gabra4 following recent spontaneous seizure activity (within 3 h) relative to control rats. Data were normalized
to input signal (Egr3/Input) and expressed as fold change with respect to control (defined as 1). Input represents the DNA signal from
sample preparation prior to immunoprecipitation. Statistically significant differences in mRNA levels and in protein–DNA binding were
determined using a one-way ANOVAwith Tukey’s test for multiple comparisons. For all graphs, error bars, mean � SEM.
Epilepsia ILAE
Epilepsia, **(*):1–8, 2014doi: 10.1111/epi.12771
5
Spontaneous Seizures and GABAARa4 Expression
subunits can assemble with either d or c2 subunits,12,21,22
with a4d-containing receptors predominating in normal DGand a4c-containing receptors becoming more abundant inepileptic tissue.5,7,12 Delta-containing GABAARs are locatedprimarily at extrasynaptic sites and have traditionally beenthought to mediate tonic inhibition, whereas phasic inhibi-tion is governed by c-containing GABAARs located at syn-aptic or perisynaptic regions.23–26 Despite a diminishedexpression of d subunit mRNA7,20 and a reduction in func-tional a4d-containing GABAARs in the hippocampus of epi-leptic animals,13 tonic GABAergic inhibition is maintainedor increased in epileptic animals.27 Some studies in chroni-cally epileptic animals suggest that the novel a4c2-contain-ing receptors are responsible for maintaining tonicinhibition,13 whereas others find that an increase in suchreceptors in synaptic and/or perisynaptic locations areresponsible for altered phasic GABA currents in epileptichippocampus.7 Thus, alterations in GABAARa4 composi-tion may be associated with changes in both tonic and phasicGABAAR--mediated inhibition.
Alterations in a-subunit subtype can result in differencesin GABAAR modulation by benzodiazepines, neuroster-oids, and zinc.28–31 Pharmacologic experiments suggest that
increased levels of a4c-containing receptors in epilepticbrain may contribute to an altered response of GABAARs toGABA and GABAergic drugs. Lagrange et al., 2007reported that a4c-containing receptors are more rapidlydesensitized and recover more slowly from desensitization.They further found that exposure to prolonged low levels ofGABA greatly suppressed the response of a4c-containingreceptor currents to higher concentrations of GABA. Over-all, these receptors appeared less efficacious when exposedto prolonged low levels of GABA or during repetitive stim-ulation, as may occur during seizures. These results are con-sistent with the observation that, in epileptic animals,GABAARs become largely insensitive to higher GABAconcentrations.32 Thus, drug treatments that increase thelevels of extrasynaptic GABA may negatively modulate theability of extrasynaptic a4c-containing receptors to respondto variations in ambient GABA that may occur followingSS, as well as during SE.33 In addition, because a4-contain-ing GABAARs are more sensitive to zinc blockade, a shift tomore a4-containing GABAARs may be associated with theenhanced blockade of the GABAA response by zinc in theDG, seen in both acutely after SE and in chronic epilepsy inboth animals28–30,34 and in humans with TLE.35 Zinc is
A B C D
E F G H
Figure 3.
Characterization of seizures in epileptic rats. (A) EEG data across all rats used for above experiments for <3 h SS group (n = 16) demon-
strate a significantly higher seizure frequency (i.e., seizures/day) during the 7-day monitoring period prior to sacrifice relative to the
>24 h SS group (n = 6; **p = 0.0019). Statistically significant differences were determined using a Mann-Whitney test. During the same
7-day period, continuous EEG monitoring demonstrated no significant difference in seizure duration (B) or in the proportion of convul-
sive seizures to nonconvulsive seizures present in the <3 h SS group or the >24 h SS group (C). Statistically significant differences were
determined using an unpaired t-test and a Fisher’s exact test, respectively. (D) Frequency histograms of the interseizure intervals for the
group of epileptic rats sacrificed within 3 h of last spontaneous seizure and >24 h after last spontaneous seizure. The first bin (<60 min
or >1 seizure/h) had a high number of interseizure intervals for both groups, suggesting the presence of seizure clusters. A significant dif-
ference between the two groups was found in <300 (or ~6 h) bin demonstrating the SS 24H group had more long interseizure intervals
(or intercluster intervals). Statistically significant differences were determined using a one-way ANOVA with a Newman-Keuls test for
multiple comparisons. Representative raster plots depicting the time of previous seizure occurrence under the conditions of increased
GABAARa4 mRNA and protein expression within 3 h of a spontaneous seizure (E and F) or unchanged GABAARa4 mRNA and protein
expression (G andH) in DG of rats without seizures for >24 h.
Epilepsia ILAE
Epilepsia, **(*):1–8, 2014doi: 10.1111/epi.12771
6
H. L. Grabenstatter et al.

distributed by aberrant mossy fiber axons of DG cells thatinnervate the inner molecular layer of the DG in latestages of epileptogenesis (i.e., after the onset of SS). Thea4-containing GABAARs present on dendrites of epilepticDG neurons are exquisitely sensitive to blockade by zinc,and thus may contribute to loss of inhibitory function andincreased excitability.29,35 The transcriptional upregulationof GABAARa4 gene expression seen in association withspontaneous seizures may thus result in changes in bothtonic and phasic GABAAR-mediated inhibition that couldimpact seizure susceptibility in chronically epileptic rats.
Caveats related to assessing Egr3 regulation ofGabra4changes
The increases in BDNF levels following recent spontane-ous seizure activity (i.e., within 3 h) suggest that SS mayrecurrently “reactivate” signaling pathways that could con-tribute to the perpetuation and progression of epilepsy.However, although there was a marked and significantincrease in levels of Egr3 mRNA, as well as BDNF, follow-ing recent SS, we did not detect an associated increase inEgr3 binding to the Gabra4 promoter. Without this evi-dence, we cannot definitively conclude that BDNF-inducedincreases in Egr3 are driving the induction of GABAARa4transcription following SS. Our evidence does allow us toconclude, however, that Egr3 is present at the promoterregion and is part of the transcriptional complex that mostlikely underlies increased rates of transcription in our stud-ies. Little is known about the mechanism of Egr3-directedgene regulation in neurons, and research in this area hasbeen hampered by the lack of antibodies with which toprobe the expression of its many variants, an active area ofinvestigation in our laboratories.
ConclusionsThese studies provide the first evidence of acute molecu-
lar regulation of GABAAR subunit expression in associationwith spontaneous seizures during chronic epilepsy. The cur-rent study specifically examines changes in GABAARa4,and as it is known that expression of multiple GABAAR su-bunits may be alerted in chronic epilepsy, the role of sponta-neous seizures in regulating other subunits and thesubsequent effects on receptor composition and function,merit additional study. Better understanding of the molecu-lar changes induced by SS and their functional conse-quences could contribute to the development of disease-modifying therapies that do not just symptomatically treatseizures but potentially inhibit their progression and long-term adverse consequences.
AcknowledgmentsThe authors would like to thank the University of Colorado Neurophysi-
ology Core for assistance related to EEGmonitoring. Funding was provided
by the National Institutes of Health (NIH), National Institute of Neurologi-cal Disorders and Stroke (NINDS) R01NS051710 (to ABK and SJR) andthe Epilepsy Foundation (to HG).
DisclosureNone of the authors has any conflict of interest to disclose. We confirm
that we have read the Journal’s position on issues involved in ethical publi-cation and affirm that this report is consistent with those guidelines.
References1. Sieghart W. Structure, pharmacology, and function of GABAA
receptor subtypes. Adv Pharmacol 2006;54:231–263.2. Rudolph U, Mohler H. GABA-based therapeutic approaches:
GABAA receptor subtype functions. Curr Opin Pharmacol2006;6:18–23.
3. Brooks-Kayal AR, Shumate MD, Jin H, et al. Selective changes insingle cell GABA(A) receptor subunit expression and function intemporal lobe epilepsy.Nat Med 1998;4:1166–1172.
4. Lauren HB, Pitkanen A, Nissinen J, et al. Selective changes in gamma-aminobutyric acid type A receptor subunits in the hippocampus inspontaneously seizing rats with chronic temporal lobe epilepsy.Neurosci Lett 2003;349:58–62.
5. Peng Z, Huang CS, Stell BM, et al. Altered expression of the deltasubunit of the GABAA receptor in a mouse model of temporal lobeepilepsy. J Neurosci 2004;24:8629–8639.
6. Sperk G, Furtinger S, Schwarzer C, et al. GABA and its receptors inepilepsy. Adv Exp Med Biol 2004;548:92–103.
7. Zhang N, Wei W, Mody I, et al. Altered localization of GABA(A)receptor subunits on dentate granule cell dendrites influences tonic andphasic inhibition in a mouse model of epilepsy. J Neurosci2007;27:7520–7531.
8. Kamphuis W, De Rijk TC, Lopes da Silva FH. Expression of GABAAreceptor subunit mRNAs in hippocampal pyramidal and granularneurons in the kindling model of epileptogenesis: an in situhybridization study. Brain Res Mol Brain Res 1995;31:33–47.
9. Schwarzer C, Tsunashima K, Wanzenbock C, et al. GABA(A)receptor subunits in the rat hippocampus II: altered distribution inkainic acid-induced temporal lobe epilepsy. Neuroscience1997;80:1001–1017.
10. Brooks-Kayal AR, Shumate MD, Jin H, et al. Human neuronalgamma-aminobutyric acid(A) receptors: coordinated subunit mRNAexpression and functional correlates in individual dentate granule cells.J Neurosci 1999;19:8312–8318.
11. Sperk G, Drexel M, Pirker S. Neuronal plasticity in animal models andthe epileptic human hippocampus. Epilepsia 2009;50(Suppl. 12):29–31.
12. Lund IV, Hu Y, Raol YH, et al. BDNF selectively regulates GABAAreceptor transcription by activation of the JAK/STAT pathway. SciSignal 2008;1:ra9.
13. Rajasekaran K, Joshi S, Sun C, et al. Receptors with low affinity forneurosteroids and GABA contribute to tonic inhibition of granule cellsin epileptic animals.Neurobiol Dis 2010;40:490–501.
14. Lagrange AH, Botzolakis EJ, Macdonald RL. Enhanced macroscopicdesensitization shapes the response of alpha4 subtype-containingGABAA receptors to synaptic and extrasynaptic GABA. J Physiol2007;578:655–676.
15. Raol YH, Lund IV, Bandyopadhyay S, et al. Enhancing GABA(A)receptor alpha 1 subunit levels in hippocampal dentate gyrus inhibitsepilepsy development in an animal model of temporal lobe epilepsy.J Neurosci 2006;26:11342–11346.
16. Roberts DS, Hu Y, Lund IV, et al. Brain-derived neurotrophic factor(BDNF)-induced synthesis of early growth response factor 3 (Egr3)controls the levels of type A GABA receptor alpha 4 subunits inhippocampal neurons. J Biol Chem 2006;281:29431–29435.
17. Roberts DS, Raol YH, Bandyopadhyay S, et al. Egr3 stimulation ofGABRA4 promoter activity as a mechanism for seizure-induced up-regulation of GABA(A) receptor alpha4 subunit expression. Proc NatlAcad Sci USA 2005;102:11894–11899.
Epilepsia, **(*):1–8, 2014doi: 10.1111/epi.12771
7
Spontaneous Seizures and GABAARa4 Expression

18. Grabenstatter HL, Cruz Del Angel Y, Carlsen J, et al. The effect ofSTAT3 inhibition on status epilepticus and subsequent spontaneousseizures in the pilocarpine model of acquired epilepsy. Neurobiol Dis2013;62:73–85.
19. Racine RJ. Modification of seizure activity by electrical stimulation.II. Motor seizure. Electroencephalogr Clin Neurophysiol 1972;32:281–294.
20. Nishimura T, Schwarzer C, Gasser E, et al. Altered expression ofGABA(A) and GABA(B) receptor subunit mRNAs in thehippocampus after kindling and electrically induced status epilepticus.Neuroscience 2005;134:691–704.
21. Sur C, Farrar SJ, Kerby J, et al. Preferential coassembly of alpha4 anddelta subunits of the gamma-aminobutyric acidA receptor in ratthalamus.Mol Pharmacol 1999;56:110–115.
22. Jia F, Pignataro L, Schofield CM, et al. An extrasynaptic GABAAreceptor mediates tonic inhibition in thalamic VB neurons.J Neurophysiol 2005;94:4491–4501.
23. Nusser Z, Mody I. Selective modulation of tonic and phasicinhibitions in dentate gyrus granule cells. J Neurophysiol 2002;87:2624–2628.
24. Wisden W, Cope D, Klausberger T, et al. Ectopic expression of theGABA(A) receptor alpha6 subunit in hippocampal pyramidal neuronsproduces extrasynaptic receptors and an increased tonic inhibition.Neuropharmacology 2002;43:530–549.
25. Mody I. Distinguishing between GABA(A) receptors responsible fortonic and phasic conductances.Neurochem Res 2001;26:907–913.
26. Chandra D, Jia F, Liang J, et al. GABAA receptor alpha 4 subunitsmediate extrasynaptic inhibition in thalamus and dentate gyrus andthe action of gaboxadol. Proc Natl Acad Sci USA 2006;103:15230–15235.
27. Zhan RZ, Nadler JV. Enhanced tonic GABA current in normotopic andhilar ectopic dentate granule cells after pilocarpine-induced statusepilepticus. J Neurophysiol 2009;102:670–681.
28. Gibbs JW 3rd, Shumate MD, Coulter DA. Differential epilepsy-associated alterations in postsynaptic GABA(A) receptor function indentate granule and CA1 neurons. J Neurophysiol 1997;77:1924–1938.
29. Cohen AS, Lin DD, Quirk GL, et al. Dentate granule cell GABA(A) receptors in epileptic hippocampus: enhanced synapticefficacy and altered pharmacology. Eur J Neurosci 2003;17:1607–1616.
30. Buhl EH, Otis TS, Mody I. Zinc-induced collapse of augmentedinhibition by GABA in a temporal lobe epilepsy model. Science1996;271:369–373.
31. Sun C, Mtchedlishvili Z, Erisir A, et al. Diminished neurosteroidsensitivity of synaptic inhibition and altered location of the alpha4subunit of GABA(A) receptors in an animal model of epilepsy.J Neurosci 2007;27:12641–12650.
32. Overstreet LS, Jones MV, Westbrook GL. Slow desensitizationregulates the availability of synaptic GABA(A) receptors. J Neurosci2000;20:7914–7921.
33. Naylor DE, Wasterlain CG. GABA synapses and the rapid loss ofinhibition to dentate gyrus granule cells after brief perforant-pathstimulation. Epilepsia 2005;46(Suppl. 5):142–147.
34. Leroy C, Poisbeau P, Keller AF, et al. Pharmacological plasticity ofGABA(A) receptors at dentate gyrus synapses in a rat model oftemporal lobe epilepsy. J Physiol 2004;557:473–487.
35. Shumate MD, Lin DD, Gibbs JW 3rd, et al. GABA(A) receptorfunction in epileptic human dentate granule cells: comparison toepileptic and control rat. Epilepsy Res 1998;32:114–128.
Epilepsia, **(*):1–8, 2014doi: 10.1111/epi.12771
8
H. L. Grabenstatter et al.





















![[A]petyxa;_by_Simos_Vassilis [Α4]](https://img.pdfslide.net/doc/110x75/54a11a0fac7959c87d8b4646/apetyxabysimosvassilis-4.jpg)







