Embed Size (px)
Citation preview

Immunity 24, 787–799, June 2006 ª2006 Elsevier Inc. DOI 10.1016/j.immuni.2006.03.024
Efficient T Cell Activationvia a Toll-Interleukin 1Receptor-Independent Pathway
Edith Janssen,1 Koichi Tabeta,3 Michael J. Barnes,3
Sophie Rutschmann,3 Sara McBride,1 Keith S. Bahjat,4
Stephen P. Schoenberger,2
Argyrios N. Theofilopoulos,3 Bruce Beutler,3
and Kasper Hoebe3,*1Department of Developmental Immunology2Laboratory of Cellular ImmunologyLa Jolla Institute for Allergy and ImmunologySan Diego, California 921213Department of ImmunologyThe Scripps Research InstituteLa Jolla, California 920374Cerus CorporationConcord, California 94520
Summary
Here, we describe a previously unrecognized pathway
for activation of antigen-specific adaptive immune re-sponses that was independent of Toll-Interleukin 1 Re-
ceptor signaling and directed toward detection of anti-gens expressed by apoptotic cells. This pathway is
represented within Flt-3 Ligand-derived dendritic cells(DCs) that represent immature lymphoid DCs, but not
within GM-CSF-treated bone marrow-derived dendriticcells. Exposure of these DCs to apoptotic cells resulted
in production of type I interferon and favored the devel-opment of cytotoxic T cell responses. The N-Ethyl-N-Ni-
trosourea-induced germline mutation 3d (Unc3b13d/3d)abolished both MHC class I and II responses elicited
by this pathway, whereas a null allele of Cd36 selectivelyabolished class II responses. We propose that this mode
ofadaptive immune activation evolved to permit the sen-sitive detection of intracellular microbial infections,
particularly viral infections, which frequently induceapoptotic cell death, but may also be important in trans-
plantation, autoimmunity, and vaccine development.
Introduction
It is commonly thought that apoptosis is a bland processthat does not stimulate innate or adaptive immune re-sponses (Gallucci et al., 1999; Blander and Medzhitov,2004; Li et al., 2001; Steinman et al., 2000). Yet pro-grammed cell death is induced by a variety of viral infec-tions, and teleological reasoning suggests that the hostwould benefit from exercise of a mechanism serving theimmune detection of foreign proteins expressed in thecontext of a dying cell, an idea that has previously beenposed as the danger hypothesis (Matzinger, 1994). In-deed, evidence suggests that apoptosis contributes tothe onset of an adaptive immune response either directly(Albert et al., 1998; Finberg et al., 1979; Ronchetti et al.,1999) or in the context of viral and/or bacterial infections(Gorla et al., 1994; Yrlid and Wick, 2000; Winau et al.,2006). However, the underlying mechanisms for these ob-
*Correspondence: [email protected]
servations remain elusive. Toll-like receptors (TLRs) arewell recognized for their ability to sense molecules ofmicrobial origin (Akira and Takeda, 2004) and to mediatemicrobial adjuvanticity (Iwasaki and Medzhitov, 2004;Hoebe et al., 2003c; Schnare et al., 2001). It has been sug-gested that the promiscuity of TLR receptors permitsactivation not only by molecules derived from microbesbut alsobyendogenousmolecules thatmay possess sim-ilar chemical properties (Seong and Matzinger, 2004). Inthis report, however, we describe a powerful Toll/Interleu-kin-1 Receptor domain (TIR)-independent pathway foradaptive immune activation. This pathway senses anti-gens expressed by cells undergoing programmed deathand depends upon specific proteins that contribute to in-nate immune responses. It is represented within a subsetof Flt3L-induced bone marrow-derived dendritic cells(BMDC) that, upon maturation, transform into CD8+
lymphoid-related DCs.
Results
CTL Responses Induced by Cell Death
To assess the requirements for the onset of an adaptiveimmune response induced by cell death, we first estab-lished an immunization model by using syngeneic cellstransgenic for an actin promoter-driven, membrane-associated form of ovalbumin (act-mOVA) (Ehst et al.,2003). C57BL/6 mice were immunized with 10 millionact-mOVA cells that were either treated with g irradiation(1500 rad) to induce apoptosis or left untreated prior toinjection (see Figure S1 in the Supplemental Data avail-able with this article online). After 8 days, splenocyteswere isolated from each mouse and restimulated withOVA257-264 peptide. The frequency of antigen-specificCD8+ T cells was determined by measuring intracellularinterferon-g (IFN-g) production as well as specific bind-ing of OVA-H2kb tetramers, as previously described(Hoebe et al., 2003c). In parallel, cells were restimulatedfor 6 days to address their cytolytic activity and theircapacity for secondary expansion, a hallmark of T cellmemory. Although immunization with apoptotic cellsresulted in a strong IFN-g+ response in CD8+ T cells, im-munization with nonirradiated act-mOVA cells did not(Figure 1A). To exclude direct priming by apoptotic cells,we immunized mice with either irradiated act-mOVAsplenocytes or act-mOVA splenocytes on a Kbm1 back-ground to avoid direct priming via MHC class I. Bothimmunizations of mice with g-irradiated (1500 rad) act-mOVA and act-mOVA-Kbm1 splenocytes induced strongCD8+ T cell responses as measured by the number ofCD8+IFN-g+ cells after restimulation with the OVA257-264
peptide (Figure 1B). To test whether the responsecould be induced by different apoptotic stimuli, weimmunized mice with act-mOVA splenocytes that wereeither UV radiated (254 nm wavelength; 240 mJ/cm2)or treated with Fas-activating antibody (5 mg/ml). Eachprotocol resulted in the development of strong cytotoxicT lymphocyte (CTL) responses with a high frequencyof antigen-specific CD8+IFNg+ cells and efficient killingof target cells (Figures 1C and 1D). In addition,
Immunity788
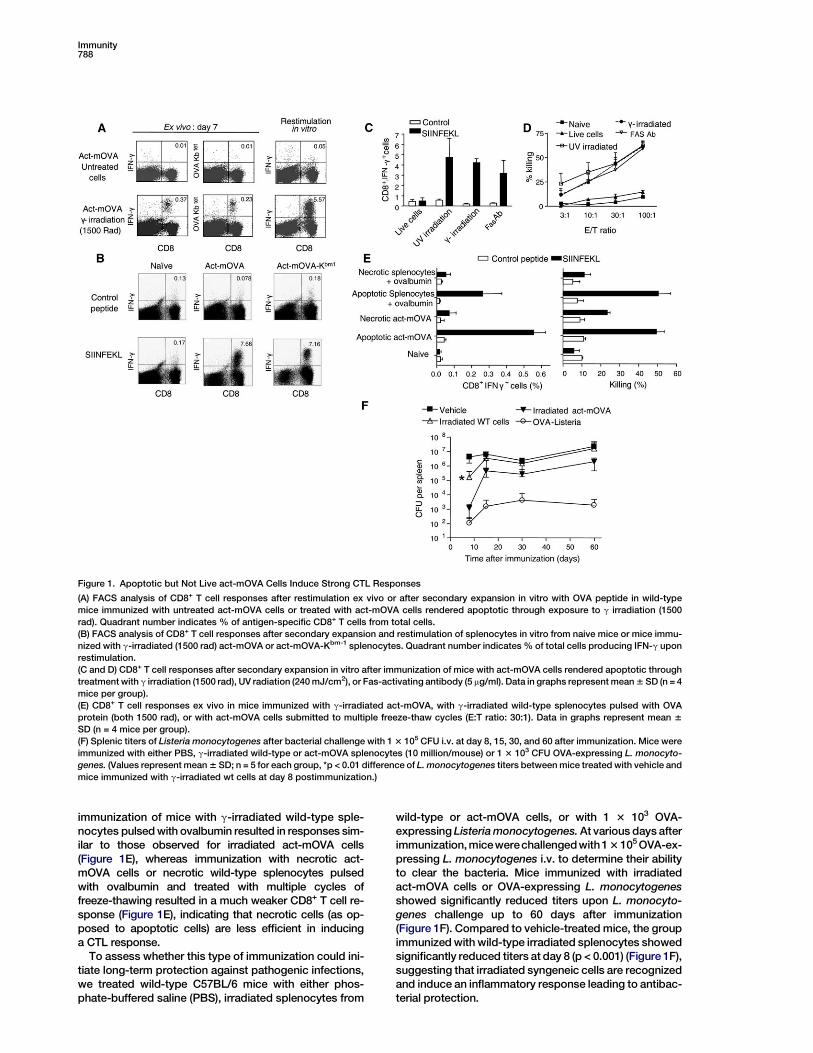
Figure 1. Apoptotic but Not Live act-mOVA Cells Induce Strong CTL Responses
(A) FACS analysis of CD8+ T cell responses after restimulation ex vivo or after secondary expansion in vitro with OVA peptide in wild-type
mice immunized with untreated act-mOVA cells or treated with act-mOVA cells rendered apoptotic through exposure to g irradiation (1500
rad). Quadrant number indicates % of antigen-specific CD8+ T cells from total cells.
(B) FACS analysis of CD8+ T cell responses after secondary expansion and restimulation of splenocytes in vitro from naive mice or mice immu-
nized with g-irradiated (1500 rad) act-mOVA or act-mOVA-Kbm-1 splenocytes. Quadrant number indicates % of total cells producing IFN-g upon
restimulation.
(C and D) CD8+ T cell responses after secondary expansion in vitro after immunization of mice with act-mOVA cells rendered apoptotic through
treatment with g irradiation (1500 rad), UV radiation (240 mJ/cm2), or Fas-activating antibody (5 mg/ml). Data in graphs represent mean 6 SD (n = 4
mice per group).
(E) CD8+ T cell responses ex vivo in mice immunized with g-irradiated act-mOVA, with g-irradiated wild-type splenocytes pulsed with OVA
protein (both 1500 rad), or with act-mOVA cells submitted to multiple freeze-thaw cycles (E:T ratio: 30:1). Data in graphs represent mean 6
SD (n = 4 mice per group).
(F) Splenic titers of Listeria monocytogenes after bacterial challenge with 1 3 105 CFU i.v. at day 8, 15, 30, and 60 after immunization. Mice were
immunized with either PBS, g-irradiated wild-type or act-mOVA splenocytes (10 million/mouse) or 1 3 103 CFU OVA-expressing L. monocyto-
genes. (Values represent mean 6 SD; n = 5 for each group, *p < 0.01 difference of L. monocytogenes titers between mice treated with vehicle and
mice immunized with g-irradiated wt cells at day 8 postimmunization.)
immunization of mice with g-irradiated wild-type sple-nocytes pulsed with ovalbumin resulted in responses sim-ilar to those observed for irradiated act-mOVA cells(Figure 1E), whereas immunization with necrotic act-mOVA cells or necrotic wild-type splenocytes pulsedwith ovalbumin and treated with multiple cycles offreeze-thawing resulted in a much weaker CD8+ T cell re-sponse (Figure 1E), indicating that necrotic cells (as op-posed to apoptotic cells) are less efficient in inducinga CTL response.
To assess whether this type of immunization could ini-tiate long-term protection against pathogenic infections,we treated wild-type C57BL/6 mice with either phos-phate-buffered saline (PBS), irradiated splenocytes from
wild-type or act-mOVA cells, or with 1 3 103 OVA-expressing Listeria monocytogenes. At various days afterimmunization, micewerechallengedwith1 3 105 OVA-ex-pressing L. monocytogenes i.v. to determine their abilityto clear the bacteria. Mice immunized with irradiatedact-mOVA cells or OVA-expressing L. monocytogenesshowed significantly reduced titers upon L. monocyto-genes challenge up to 60 days after immunization(Figure 1F). Compared to vehicle-treated mice, the groupimmunized with wild-type irradiated splenocytes showedsignificantly reduced titers at day 8 (p < 0.001) (Figure 1F),suggesting that irradiated syngeneic cells are recognizedand induce an inflammatory response leading to antibac-terial protection.
TLR-Independent Adjuvanticity for CTL Responses789
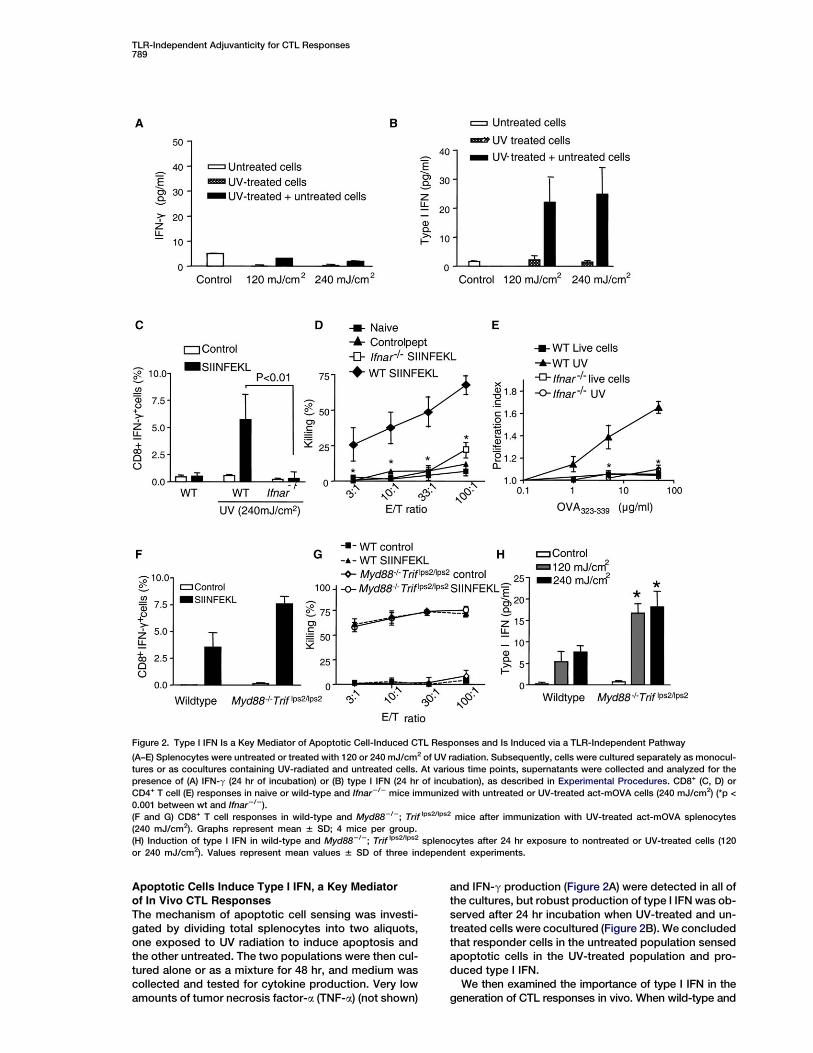
Figure 2. Type I IFN Is a Key Mediator of Apoptotic Cell-Induced CTL Responses and Is Induced via a TLR-Independent Pathway
(A–E) Splenocytes were untreated or treated with 120 or 240 mJ/cm2 of UV radiation. Subsequently, cells were cultured separately as monocul-
tures or as cocultures containing UV-radiated and untreated cells. At various time points, supernatants were collected and analyzed for the
presence of (A) IFN-g (24 hr of incubation) or (B) type I IFN (24 hr of incubation), as described in Experimental Procedures. CD8+ (C, D) or
CD4+ T cell (E) responses in naive or wild-type and Ifnar2/2 mice immunized with untreated or UV-treated act-mOVA cells (240 mJ/cm2) (*p <
0.001 between wt and Ifnar2/2).
(F and G) CD8+ T cell responses in wild-type and Myd882/2; Trif lps2/lps2 mice after immunization with UV-treated act-mOVA splenocytes
(240 mJ/cm2). Graphs represent mean 6 SD; 4 mice per group.
(H) Induction of type I IFN in wild-type and Myd882/2; Trif lps2/lps2 splenocytes after 24 hr exposure to nontreated or UV-treated cells (120
or 240 mJ/cm2). Values represent mean values 6 SD of three independent experiments.
Apoptotic Cells Induce Type I IFN, a Key Mediatorof In Vivo CTL Responses
The mechanism of apoptotic cell sensing was investi-gated by dividing total splenocytes into two aliquots,one exposed to UV radiation to induce apoptosis andthe other untreated. The two populations were then cul-tured alone or as a mixture for 48 hr, and medium wascollected and tested for cytokine production. Very lowamounts of tumor necrosis factor-a (TNF-a) (not shown)
and IFN-g production (Figure 2A) were detected in all ofthe cultures, but robust production of type I IFN was ob-served after 24 hr incubation when UV-treated and un-treated cells were cocultured (Figure 2B). We concludedthat responder cells in the untreated population sensedapoptotic cells in the UV-treated population and pro-duced type I IFN.
We then examined the importance of type I IFN in thegeneration of CTL responses in vivo. When wild-type and

Immunity790
IFN type I receptor knockout mice (Ifnar2/2) were immu-nized with apoptotic act-mOVA cells, the latter showeda marked reduction in the percentage of CD8+IFN-g+ cells(Figure 2C) as well as the ability to kill OVA-expressing tar-get cells (Figure 2D), and CD4 T cells failed to proliferatewhen exposed to MHC class II-specific OVA peptides(Figure 2E). These results indicate that type I IFN is a keymediator of immune responses induced by apoptotic cells.
Cell Death-Induced CTL Responses Do Not Depend
upon TLR SignalingBecause it was suggested that TLRs are essential for rec-ognition of molecular constituents released from injuredtissues or apoptotic cells of the host (Seong and Mat-zinger, 2004), we studied responses to apoptotic cells inmice with homozygous mutations in both Myd88 andTrif genes (Myd882/2 Trif lps2/lps2) (Hoebe et al., 2003a).TLR-mediated macrophage responses to all known TLRligands are abolished in these mice except for dsRNA,which is known to signal via a TLR-independent pathwayandelicits type I IFNproduction, but notTNF-a production(Figures S2A–S2H; Hoebe et al., 2003c).
In MyD882/2 Trif lps2/lps2 mice, the number ofCD8+IFN-g+ T cells generated in response to immuniza-tion with UV-treated act-mOVA cells appeared to be in-creased compared to the number observed in wild-typemice (Figure 2F), and both wild-type and MyD882/2
Trif lps2/lps2 mice supported the generation of equivalentcytotoxic activity against OVA-expressing target cells(Figure 2G). Moreover, type I IFN production was slightlyincreased in splenocyte cultures from mice lacking TLRsignaling compared to wild-type mice when exposed toUV-treated (240 mJ/cm2) splenocytes (Figure 2H). Theseresults effectively challenge the assumption that TLRsare involved in cell death-induced immune responses,and instead suggest the involvement of a previously un-described TLR-independent pathway leading to type IIFN production.
Flt3L-Derived B2202, but Not GM-CSF-Derived DCs,Respond to Apoptotic Cells
Dendritic cells (DCs) and macrophages have previouslybeen implicated as the major cell types involved in up-take of apoptotic cells, and DCs are potent inducers ofboth CD4+ and CD8+ T cell responses. We hypothesizedthat either conventional myeloid (MDCs) or plasmacytoidDCs (PDCs) might be responsible for recognition of apop-totic cells and subsequent type I IFN production. Wetherefore generated GM-CSF-derived DCs and B220+
and B2202 Flt3 Ligand (Flt3L)-derived DCs from bonemarrow of C57BL/6 mice and examined these cells forexpression of myeloid- and lymphoid-specific markers(Figure S3A) and for responses to CpG and LPS(Figure S3B). We then exposed these cells to apoptoticsyngeneic splenocytes and monitored type I IFN produc-tion over 48 hr. GM-CSF-derived DCs did not producesubstantial amounts of type I IFN, whereas B2202 and,to a lesser extent, B220+ Flt3L-derived DCs were efficienttype I IFN producers when exposed to apoptotic cells(Figure 3A). The type I IFN production by DCs becameapparent after 24 hr, consistent with our observationswhen we used total splenocytes as responders, andwas independent of the apoptotic stimulus used. To spe-cifically address the possibility that PDCs were able to
produce type I IFN in response to apoptotic cells, weseparated PDCs from non-PDCs by using a specific an-tibody recognizing mouse plasmacytoid dendritic cellantigen-1 (mPDCA-1). Whereas Flt3L-derived mPDCA12
DCs showed a similar type I IFN production as observedfor B2202 DC, mPDCA1+ cells were unable to respond toapoptotic cells (Figure S3C), suggesting that the type IIFN response primarily depends upon Flt3L-derivedmPDCA12B2202 DCs.
In addition to the poor ability to induce CTL responses,necrotic cells were also unable to induce type I IFN in anyof the DC subtypes (Figure 3B). Apoptotic cells obtainedafter UV irradiation (240 mJ/cm2), g-irradiation (1500 rad),or exposure to Fas-activating antibody (5 mg/ml) inducedsimilar amounts of type I IFN production (Figure 3C).
Flt3L-Derived B2202 DCs Acquire Apoptotic
Cell-Derived Antigen via ‘‘Nibbling’’ andAre Highly Efficient in Crosspriming
UV-treated splenocytes were labeled with a fluorescentmarker (CFSE) and exposed to Flt3L-derived B220+
and B2202 DCs as well as GM-CSF-derived myeloidDCs (mDCs) for 24 hr. Although GM-CSF-derivedmDCs were able to engulf apoptotic cells (Figure 3D,Figure S3D), Flt3L-derived B2202 (Figure 3D) andB220+ (not shown) DCs were not. However, both B2202
and B220+ Flt3L-derived DCs acquired apoptotic mate-rial via a process that resembled ‘‘nibbling,’’ which wasobservable after 24 hr of coincubation (Figure 3D andFigure S3E). The uptake of apoptotic material could bequantitated by FACS (Figure 3E).
The ability of each DC subtype to crossprime OT-ICD8+ T cells was examined after 24 hr pre-exposure toapoptotic cells that carried the act-mOVA transgeneon a C57BL/6 Kbm-1 background (thus excluding directactivation of OT-I cells via act-mOVA cells). B2202
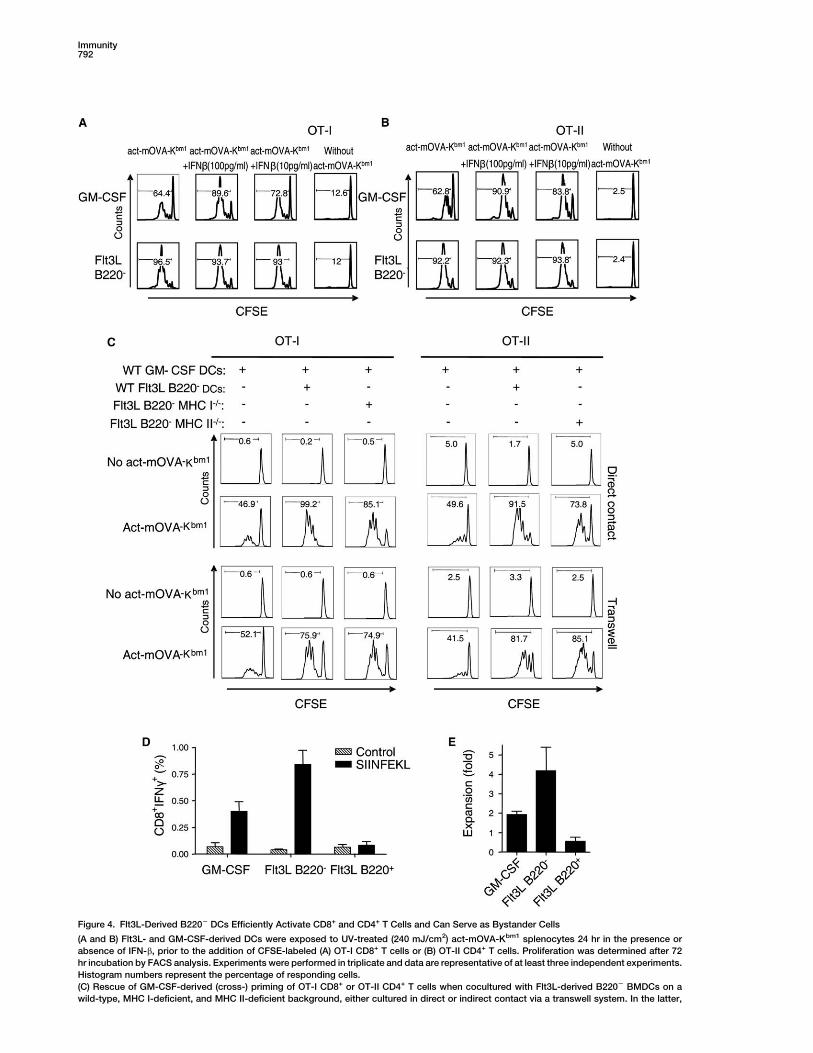
Flt3L-derived DCs were highly efficient in crossprimingOT-I cells, whereas both GM-CSF- and B220+ Flt3L-de-rived DCs induced little or no crosspriming (Figure 3F).The same results were observed for priming of OT-IICD4 T cells (results not shown). Coincubation of GM-CSF-derived mDCs with irradiated act-mOVA cells inthe presence of type I IFN (Figures 4A and 4B) orB2202 Flt3L-derived DCs that were physically separatedthrough transwell systems (Figure 4C) restored the abil-ity of these mDCs to (cross-) prime OT-I and OT-II cells.In the latter case, crosspriming of OT-I cells by GM-CSF-derived DCs was only observed when B2202 Flt3L-de-rived DCs were in direct contact with apoptotic cells(Figure S4), suggesting that the apoptotic cell-derivedIFN-inducing factor is unable to pass the membraneand therefore does not represent a soluble factor re-leased from apoptotic cells.
To confirm crosspriming ability of these various DCsubtypes in vivo, we generated GM-CSF and Flt3L-de-rived DCs and incubated them overnight with a 1:1 ratioof g-irradiated act-mOVA Kbm-1 splenocytes. We thenimmunized mice with 10 million (MACS isolated) DC sub-sets i.v., and after 8 days the antigen-specific CD8+ T cellresponse was analyzed ex vivo. Whereas Flt3L-derivedB220+ PDCs and GM-CSF-derived DCs induced CD8+
T cell responses that were modest at most, Flt3L-derivedB2202 DCs elicited a strong CD8-specific T cell response

TLR-Independent Adjuvanticity for CTL Responses791
Figure 3. Flt3L- and GM-CSF-Derived BMDCs Respond Differently to Apoptotic Cells
(A) Type I IFN production by GM-CSF and Flt3L-derived BMDCs during a 48 hr incubation period with UV-treated wild-type splenocytes. Values
are mean 6 SD of three independent experiments.
(B) Type I IFN production in different BMDC subsets exposed to g-irradiated cells or cells subjected to multiple freeze-thawing cycles. Values
represent mean 6 SD of three independent experiments.
(C) Induction of type I IFN by BMDCs exposed to cells treated with UV radiation (120 mJ/cm2), g irradiation (1500 rad), or exposed to Fas-acti-
vating antibody (5 mg/ml). Values are mean 6 SD and depict representative experiment of three independent experiments.
(D) Uptake of UV-treated splenocytes (CFSE-labeled) by B2202 Flt3L- and GM-CSF-derived BMDCs at 6 or 24 hr of coculture as analyzed by laser
scanning confocal microscopy. Red stain represents mitochondrial activity as assessed by mitotracker red treatment.
(E) FACS analysis of GM-CSF and Flt3L-derived B2202 or B220+ DCs after 24 hr incubation with or without CFSE-labeled apoptotic (UV-treated)
splenocytes.
(F) Proliferation of CFSE-labeled OT-I cells by GM-CSF and Flt3L-derived BMDCs, pre-exposed to apoptotic act-mOVA-Kbm1 cells for 24 hr.
Proliferation was determined by FACS analysis after an additional 72 hr incubation. Data are representative of four independent experiments.
and secondary expansion (Figures 4D and 4E), thusconfirming in vivo crosspriming ability of this DC subset.
Type I IFN has emerged as an important mediator forCTL responses and costimulatory molecule expression(Hoebe et al., 2003c; Le Bon and Tough, 2002; Montoyaet al., 2002). We therefore tested costimulatory moleculeexpression on GM-CSF and B2202 Flt3L-treated bonemarrow-derived DCs after exposure to apoptotic cells.Although CD40, CD80, and CD86 expression were in-creased on B2202 Flt3L-derived DCs, expression wasunchanged or decreased on GM-CSF-derived DCs(Figure S5A). Furthermore, Flt3L-derived DCs fromIfnar2/2 mice showed limited ability to activate OT-I cellsin vitro (Figure S5B), suggesting that type I IFN acts onDCs in an autocrine and/or paracrine manner indepen-dent of T cells. Finally, addition of type I IFN directlyinduced costimulatory molecule expression on Flt3L-de-rived B2202 DCs (Figure S5C), confirming the importantrole of type I IFN in priming DCs ultimately leading to effi-cient T cell activation.
Maturation of Flt3L-Derived B2202CD82 DCs into
B2202CD8+ Lymphoid-Related DCsCD8+ but not CD82B2202 DCs have previously beenshown to be the principal DC subset involved in CTL
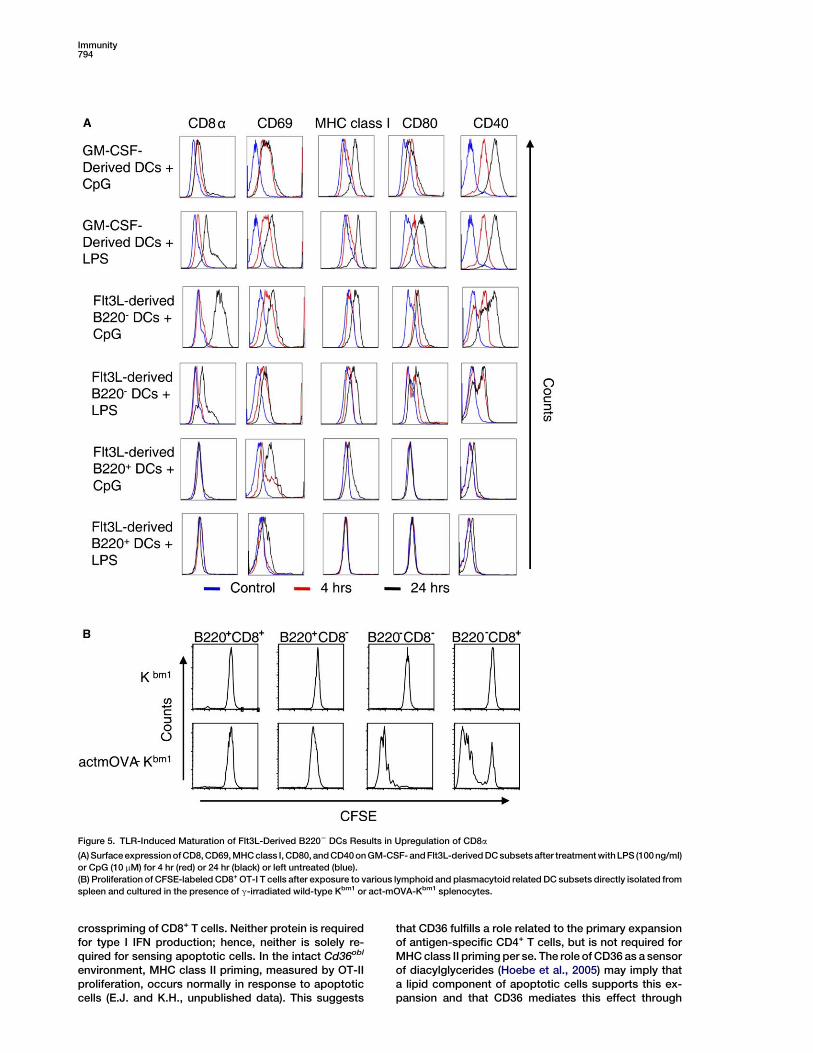
responses in the context of viral infections, i.e. HSVtype 1, influenza, and vaccinia virus infections, as wellas bacterial infections (Belz et al., 2004, 2005; Smithet al., 2003). They have been shown to elicit CTL re-sponses to antigens administered in the context ofcell death (den Haan et al., 2000). We therefore followedmaturation of GM-CSF and Flt3L-derived DC subsetsupon exposure to TLR ligands. Whereas all subtypesshowed upregulation of CD40, CD80, MHC class I, andthe activation marker CD69, CD8a expression washighly induced on Flt3L-derived B2202 DCs and tosome degree on GM-CSF-derived DCs (Figure 5A).Based on these results, we concluded that Flt3L-derivedCD82B2202 DCs represent immature precursors of con-ventional CD8+ lymphoid-related DCs.
We next confirmed the crosspriming ability of lym-phoid-related DC subsets ex vivo by isolating phenotyp-ically identical dendritic cells directly from the spleen.CD11b2CD11c+ DCs were isolated from spleen by meansof MACS and sorted based on their MHC II, CD8, and/orB220 expression (Figure S5D). While CD82B220+ andCD8+B220+ cells were unable to stimulate crossprimingafterexposure toapoptoticact-mOVAcells, the immatureCD82B2202 and mature CD8+B2202 DCs were bothhighly efficient in activating CD8+ OT-I cells (Figure 5B),
Immunity792
Figure 4. Flt3L-Derived B2202 DCs Efficiently Activate CD8+ and CD4+ T Cells and Can Serve as Bystander Cells
(A and B) Flt3L- and GM-CSF-derived DCs were exposed to UV-treated (240 mJ/cm2) act-mOVA-Kbm1 splenocytes 24 hr in the presence or
absence of IFN-b, prior to the addition of CFSE-labeled (A) OT-I CD8+ T cells or (B) OT-II CD4+ T cells. Proliferation was determined after 72
hr incubation by FACS analysis. Experiments were performed in triplicate and data are representative of at least three independent experiments.
Histogram numbers represent the percentage of responding cells.
(C) Rescue of GM-CSF-derived (cross-) priming of OT-I CD8+ or OT-II CD4+ T cells when cocultured with Flt3L-derived B2202 BMDCs on a
wild-type, MHC I-deficient, and MHC II-deficient background, either cultured in direct or indirect contact via a transwell system. In the latter,

TLR-Independent Adjuvanticity for CTL Responses793
confirming previous results obtained in vitro with bonemarrow-derived Flt3L-treated DCs. Together, these find-ings suggest that both immature and mature lymphoidDCs act either directly or as a source of type I IFN toinduce CTL responses stimulated by apoptotic cells.
Involvement of Unc93b1 and Cd36 in CD4and CD8 Responses
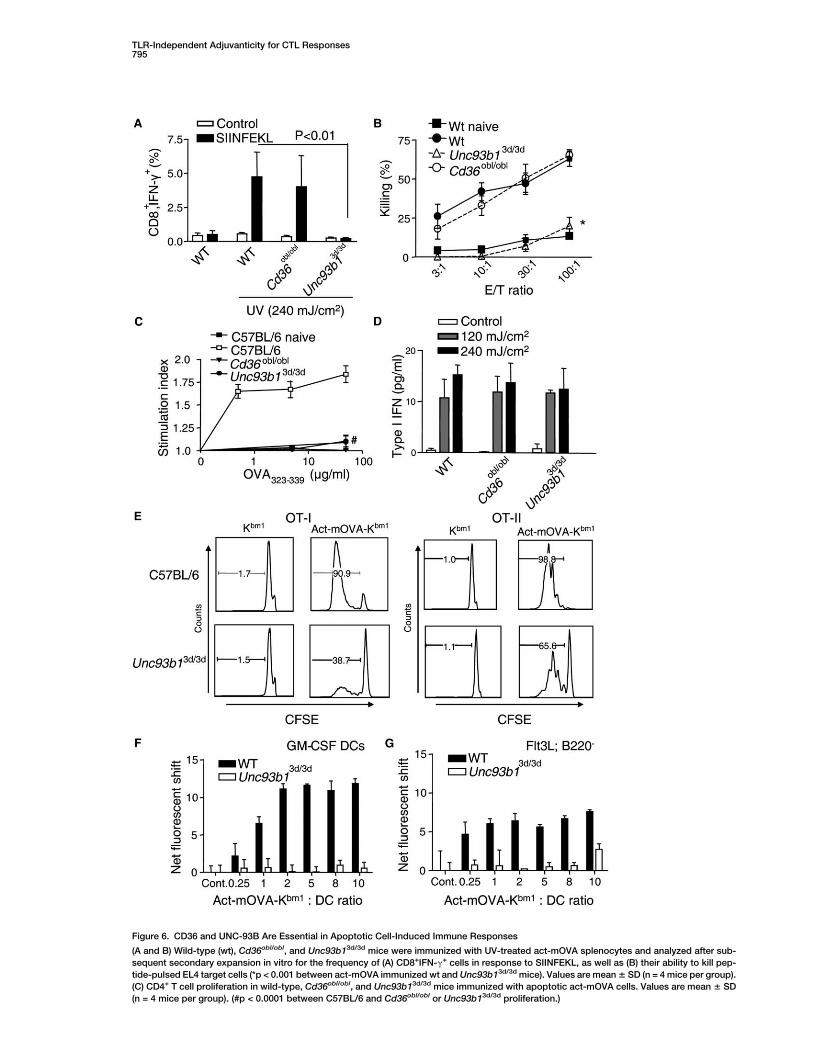
Among numerous ENU-induced mutations known to af-fect innate immune responses, two exhibited impressiveinhibitory effects on adaptive responses induced by ap-optotic cells. Oblivious, a nonsense allele of Cd36 knownto impair sensing of microbial diacylglycerides (Hoebeet al., 2005), abrogated CD4 responses, but not CD8responses, in homozygous mutant mice (Figures 6A–6C). This mutation had no effect on type I IFN production(Figure 6D). The 3d allele of Unc93b1, which encodesUNC-93B, a 12-spanning ER membrane protein requiredfor responses to TLR3, 7, and 9 ligands (Tabeta et al.,2006), abrogated both CD8+ and CD4+ T cell responsesin homozygotes (Figures 6B and 6C). Again, type I IFNproduction was unaffected (Figure 6D). Neither mutationaffected the uptake of apoptotic cells by GM-CSF- orFlt3L-derived DCs (Figure S6A and data not shown) ordecreased expression of MHC class I or class II antigenson the cell surface (Figure S6B and data not shown).However, crosspriming of OT-I CD8+ cells was abro-gated, and reduced priming of OT-II CD4+ cells was ob-served in Unc93b13d/3d mice (Figure 6E). Additional stud-ies with antibodies specific for the OVA-MHC I complexshowed that Unc93b13d/3d mice were defective in MHCclass I antigen presentation when exposed to either ap-optotic act-mOVA cells (Figures 6G and 6F) or solubleovalbumin (Tabeta et al., 2006). Finally, Unc93b13d/3d
DCs directly loaded with low concentrations of OVA pep-tides were able to (cross-) prime OT-I or OT-II cells(Figure S6C). Together, these findings suggest that thedefect observed in Unc93b13d/3d mice is related to de-fective antigen processing rather than recognition or up-take of apoptotic cells.
Discussion
TLRs mediate many and perhaps most phenomena as-sociated with microbial infections, including the well-known immunoadjuvant effect of infection. However,mice that are incapable of TIR signal transductionhave grossly normal lymphoid tissues, produce IgG,and are capable of allograft rejection (unpublished ob-servations), indicating that TLRs are not obligatory foradaptive immune responses per se. We show hereinthat apoptotic cells trigger an innate and subsequentadaptive immune response that is independent of TIRsignaling. Of key importance in this pathway is the rec-ognition of apoptotic cells by (precursor) lymphoidDCs and their concomitant production of type I IFN.
Type I IFN is an important mediator of adjuvanticity viaTIR-dependent pathways (Hoebe et al., 2003c; Le Bonet al., 2003), and, as shown here, is also a central medi-ator of adjuvanticity elicited by apoptotic cells, but viaa TIR-independent pathway. Diverse subsets of DCswith specialized functions have been described (Liu,2005; Shortman and Liu, 2002). Among them, pDCs arethought to be specialized type I IFN-producing cells dur-ing viral infections, yet clearly myeloid or conventionalDCs can serve a similar function utilizing alternative sig-naling pathways (Asselin-Paturel and Trinchieri, 2005;Kato et al., 2005). The current data suggest that produc-tion of type I IFN induced by cell death depends ondirect recognition of dying cells by (precursor) lymphoidDCs and not by pDCs, whereas the removal of apop-totic cells primarily depends upon GM-CSF-derived ormDCs. In addition, immature lymphoid DCs are capableof ‘‘sampling’’ apoptotic cells through ‘‘nibbling’’ and arehighly efficient in crosspriming CD8+ T cells.
Several triggers of apoptosis are capable of initiatingevents that lead to a strong adaptive immune response.CTL responses were elicited with act-mOVA cells ren-dered apoptotic via UV or g irradiation and also aftertreatment with Fas-activating antibodies. The lattereffect implicates an apoptotic pathway that is intimatelyinvolved in immune responses. Fas receptor is ex-pressed on many cell types in different organs and in-duces apoptosis via a caspase-dependent mechanism(Barnhart et al., 2003). The engagement of Fas with itscognate ligand (FasL) is used by virtually every effectorimmune cell that is activated to mediate cytotoxicity to-wards infected cells or tumor cells (Dennert, 2002; Smythet al., 2005). In addition, many viruses or bacteria can di-rectly induce apoptosis that can lead to inflammatoryresponses. The TLR-independent pathway thereforemay detect intracellular infections and also aids the de-velopment of CTL responses when cell death occursvia NK cells or T cell-mediated cytotoxicity. Recent stud-ies report that both dsRNA and mammalian DNA inducetype I IFN via TLR-independent pathways (Ishii et al.,2006; Kato et al., 2005; Okabe et al., 2005; Yoneyamaet al., 2004). However, these pathways are present inconventional myeloid DCs, which we show here are de-ficient in type I IFN production upon exposure to apopto-tic cells. Furthermore, we found no effect on apoptoticcell-mediated type I IFN production in the presence ofDNase I and/or RNase (results not shown). These obser-vations suggest that the production of type I IFN is inde-pendent of DNA and RNA, yet the possibility that thesenucleotide structures enter the cytosol directly withoutexposure to the extracellular environment cannot beexcluded.
Although not all of the molecular components of thedeath-induced immunoadjuvant pathway are known,we have shown that CD36 and UNC-93B both contributeto CD4+ T cell priming, while UNC-93B is required for
both top and bottom wells contain apoptotic act-mOVA cells, with top wells containing Flt3L-derived B2202 BMDCs and bottom wells containing
GM-CSF-derived DCs and OT-I CD8 or OT-II CD4 T cells. Control experiments include apoptotic act-mOVA cells in the bottom or top well only
(Figure S3). Experiments were performed in triplicate and data are representative for at least three independent experiments. Histogram numbers
represent the percentage of responding cells.
(D and E) Immunization of C57BL/6 mice with 10 million GM-CSF- or Flt3L-derived B220+ and B2202 DC subsets exposed for 24 hr to g-irradiated
act-mOVAKBm-1 splenocytes. After 8 days, the antigen-specific CD8+ T cell response was determined after ex vivo restimulation with SIINFEKL
and measurement of the % CD8+IFN-g+ T cells (D) and their ability to proliferate after exposure to OVA-expressing EL-4 cells in vitro (E). Values
are mean values 6 SD (n = 4 mice per group).
Immunity794
Figure 5. TLR-Induced Maturation of Flt3L-Derived B2202 DCs Results in Upregulation of CD8a
(A) Surface expression of CD8, CD69, MHC class I, CD80, and CD40 on GM-CSF- and Flt3L-derived DC subsets after treatment withLPS (100 ng/ml)
or CpG (10 mM) for 4 hr (red) or 24 hr (black) or left untreated (blue).
(B) Proliferation of CFSE-labeled CD8+ OT-I T cells after exposure to various lymphoid and plasmacytoid related DC subsets directly isolated from
spleen and cultured in the presence of g-irradiated wild-type Kbm1 or act-mOVA-Kbm1 splenocytes.
crosspriming of CD8+ T cells. Neither protein is requiredfor type I IFN production; hence, neither is solely re-quired for sensing apoptotic cells. In the intact Cd36obl
environment, MHC class II priming, measured by OT-IIproliferation, occurs normally in response to apoptoticcells (E.J. and K.H., unpublished data). This suggests
that CD36 fulfills a role related to the primary expansionof antigen-specific CD4+ T cells, but is not required forMHC class II priming per se. The role of CD36 as a sensorof diacylglycerides (Hoebe et al., 2005) may imply thata lipid component of apoptotic cells supports this ex-pansion and that CD36 mediates this effect through
TLR-Independent Adjuvanticity for CTL Responses795
Figure 6. CD36 and UNC-93B Are Essential in Apoptotic Cell-Induced Immune Responses
(A and B) Wild-type (wt), Cd36obl/obl, and Unc93b13d/3d mice were immunized with UV-treated act-mOVA splenocytes and analyzed after sub-
sequent secondary expansion in vitro for the frequency of (A) CD8+IFN-g+ cells in response to SIINFEKL, as well as (B) their ability to kill pep-
tide-pulsed EL4 target cells (*p < 0.001 between act-mOVA immunized wt and Unc93b13d/3d mice). Values are mean 6 SD (n = 4 mice per group).
(C) CD4+ T cell proliferation in wild-type, Cd36obl/obl, and Unc93b13d/3d mice immunized with apoptotic act-mOVA cells. Values are mean 6 SD
(n = 4 mice per group). (#p < 0.0001 between C57BL/6 and Cd36obl/obl or Unc93b13d/3d proliferation.)

Immunity796
a TLR-independent process. As for UNC-93B, its diversefunctions in immunity include support of signaling viaendosomal TLRs (3, 7, 8, and 9) as well as MHC antigenpresentation leading to (cross-) priming of CD8+ andCD4+ T cells (Tabeta et al., 2006). The exact mechanismwhereby it exerts these effects has yet to be deciphered.
Autoamplification loops, based upon the binding ofpre-existing immune complexes either to B cells (viaTLR-dependent but also -independent pathways)(Boule et al., 2004; Lau et al., 2005; Martin and Elkon,2005;Vollmer et al., 2005; Leadbetter et al., 2002; Barratet al., 2005) or to PDCs (via FcgRIIa) (Bave et al., 2003;Lovgren et al., 2004) have been proposed as a funda-mental mechanism in systemic lupus erythrematosus(SLE), the prototypic systemic autoimmune disease. Inthe former case, the adaptive immune response was de-pendent upon TLR activation, while the latter was ac-companied by type I IFN production. The death-drivenimmunoadjuvant pathway described here may also am-plify the autoimmune response in SLE, a view consistentwith defects in removal of apoptotic bodies (Herrmannet al., 1998), the existence of a type I IFN ‘‘signature’’in blood cells and afflicted tissues of patients, and therequirement for type I IFN in some spontaneous modelsof this disease (Theofilopoulos et al., 2005). As such, itwould represent a proximal autoimmune pathway thatcould function independent of pre-existing autoanti-bodies. Mutations that alter the normal pattern of celldeath, impair the removal of apoptotic cells, or enhanceimmune perception of apoptotic cells might be primarycontributors to autoimmunity. Moreover, interruptionof the apoptotic cell-induced immune response mighthave a substantial mitigating effect in autoimmunedisease.
Experimental Procedures
Mice and Materials
All experiments were performed according to the US National Insti-
tutes of Health guidelines. C57BL/6 wild-type, CD36obl/obl,
unc93b13d/3d mutant, OT-I, OT-II, and MyD882/2; Trif lps2/lps2 dou-
ble-deficient mice were bred on a C57BL/6 background and housed
in the Scripps Research Institute Vivarium. All knockout mice used
in the experiments were transferred on a C57BL/6 background after
repeated backcrossing to C57BL/6 mice (at least 6 times). IFNAR
knockout mice were kindly provided by Dr. Jonathan Sprent (The
Scripps Research Institute, La Jolla, CA), and act-mOVA transgenic
mice were a kind gift from Dr. Mark Jenkins (University of Minnesota
Medical School, Minneapolis, MN) and were bred onto a Kbm1 back-
ground.
Escherichia coli RE595 LPS was obtained from Alexis (San Diego,
CA), and dsRNA (poly (I:C)) was obtained from Amersham Pharma-
cia Biotech (Piscataway, NJ). Resiquimod was a kind gift from No-
vartis (Basel, Switzerland). PAM3CSK4 and Malp2 were obtained
from EMC microcollections GmbH (Tubingen, Germany). Phosphor-
othioate-stabilized CpG oligodeoxynucleotide (CpG ODN) 50-TCC-
ATG-ACG-TTC-CTG-ATG-CT-30 was obtained from Integrated
DNA Technologies (Coralville, IA). Zymosan A was obtained from
Sigma (St. Louis, MO). Fluorescent-labeled antibodies directed
against IFN-g, CD11b, CD11c, MHC-II, B220, CD4, and CD8 were
obtained from eBioscience (San Diego, CA), and microbeads for
negative selection of CD4 and CD8 T cell and positive selection of
B220, mPDCA-1, and CD11c were obtained from Miltenyi Biotec
(Auburn, CA). Human Flt3 Ligand was purchased from Preprotech,
Inc. (London, UK), and GM-CSF was from Becton Dickinson (Frank-
lin Lakes, NJ). CFSE and mitotracker Red CM-H2XRos were ob-
tained from Molecular Probes (Eugene, OR). Ovalbumin-specific
peptide for MHC class I (OVA257-264: SIINFEKL) or MHC class II
(OVA323-339; ISQAVHAAHAEINEAGR) were purchased from A&A
Labs LLC (San Diego, CA).
Immunization and Listeria monocytogenes Challenge
The experiments involving immunizations and challenges with Liste-
ria monocytogenes were performed as detailed elsewhere (Brock-
stedt et al., 2005). In brief, five 6- to 8-week-old C57BL/6 male
mice per group were subcutaneously injected either with 200 ml
PBS, with 200 ml irradiated splenocytes from wild-type or act-
mOVA cells (1 3 107 in 200 ml), or with 200 ml containing 1 3 103
CFU of OVA expressing Listeria monocytogenes. Subsequently,
mice were challenged with 1 3 105 OVA expressing L. monocyto-
genes i.v. at day 8, 15, or 30 postimmunization. After 3 days, spleens
were isolated and the L. monocytogenes titer was determined after
homogenization of total spleens in 5 ml dissociation buffer contain-
ing dH2O (GIBCO) + 0.2% NP-40 (Sigma-Aldrich). Series of 10-fold
dilutions in PBS were prepared and 100 ml of each dilution was
plated onto bacterial culture plates containing Brain Heart Infusion
agar + 200 mg/mL streptomycin (Difco). After additional 24 hr incuba-
tion at 37ºC, the numbers of colonies were counted to determine the
splenic titer.
Induction of Cell Death
Splenocytes were isolated via collagenase and a cell strainer (70 mM)
and subsequently washed in Iscove’s modified Dulbecco’s medium
(IMDM) supplemented with 10% FCS, 2% penicillin/streptomycin
(Invitrogen, San Diego, CA) and 50 mM b-mercaptoethanol (Sigma)
followed by lysis of erythrocytes via red blood cell lysing buffer
(Sigma). Cells were subsequently brought to a concentration of
10 3 106 cells/ml in medium and were either left untreated or
rendered apoptotic through exposure to either g irradiation (1500
Rad), UV irradiation (240 mJ/cm), or Fas-activating antibody (5 mg/
ml). For treatment with Fas-activating antibody, splenocytes were in-
cubated at 37ºC and 5% CO2 for 2 hr, followed by two thorough wash-
ings with medium and one wash with PBS. Alternatively, cells were
rendered necrotic through 5 cycles of freeze-thawing at 280ºC in
PBS. The induction of apoptosis/necrosis was analyzed by flow cy-
tometry via annexin V and propidium iodide as apoptotic/necrotic
markers. Cells used for immunization were washed once with PBS
and resuspended in PBS at a concentration of 5 3 107 cells/ml.
Each mouse was injected s.c. with 200 ml cell suspension.
Cytokine Measurements
To measure cytokine responses in vitro, either total splenocytes (5 3
105/well) or 5 3 104 DCs/well in a 96-well plate were cocultured with
5 3 105 cells/well of UV- or g-irradiated cells for 24 hr. Control cul-
tures included 106 cells/well of apoptotic or nontreated cells. After
24 hr, supernatants were collected and cytokines measured by bio-
assays or ELISAs. TNF-a production was measured via the L929 bio-
assay as previously described (Hoebe et al., 2003b). The concentra-
tion of type I IFN was determined by means of a cell line containing
an ISRE-responsive element luciferase reporter construct as de-
scribed elsewhere (Jiang et al., 2005). Production of IFN-g was mea-
sured by a commercial ELISA (R&D Biosystems, Minneapolis, MN).
(D) Induction of type I IFN in wild-type, Cd36obl/obl, and Unc93b13d/3d splenocytes after 24 hr exposure to untreated or UV-treated cells (120 or 240
mJ/cm2). Values represent mean values 6 SD of three independent experiments.
(E) Activation of CFSE-labeled OT-I and OT-II T cells by B2202 Flt3L-derived DCs from wild-type and Unc93b13d/3d mutant mice pre-exposed to
apoptotic act-mOVA-Kbm-1 cells for 24 hr. Proliferation was determined by FACS analysis after an additional 72 hr incubation.
(F and G) GM-CSF and Flt3L-treated B2202 BMDCs from Unc93b13d/3d mice show abrogated MHC/SIINFEKL-complex formation. DCs were
incubated for 24 hr with g-irradiated act-mOVA-Kbm-1 splenocytes. Formation of MHC class I-SIINFEKL-complex was determined via a specific
antibody, as described in Experimental Procedures. Values are mean 6 SD from triplicate values and graph is representative of three indepen-
dent experiments.

TLR-Independent Adjuvanticity for CTL Responses797
Measurement of Cell Death-Induced CD4+ and CD8+ T Cell
Responses
Induction of cytolytic T lymphocyte responses via subcutaneous in-
jection of apoptotic cells has been extensively described (Finberg
et al., 1979; Huang et al., 1996; Janssen et al., 2003, 2005). In brief,
mice were immunized with act-m-OVA splenocytes that were either
left untreated or rendered apoptotic through exposure to either g ir-
radiation (1500 Rad), UV irradiation (240 mJ/cm), or Fas-activating
antibody (5 mg/ml). Mice were immunized subcutaneously with
10 3 106 cells in PBS. Alternatively, mice were immunized with
equivalent volume/numbers of necrotic cells obtained through mul-
tiple cycles (5 times) of freeze-thawing (280ºC) in PBS. After 7 days,
spleens were isolated and single-cell suspensions were made in
IMDM supplemented with 10% FCS, 2% penicillin/streptomycin,
and 50 mM b-mercaptoethanol. The frequency of OVA-specific
CD8+ T cells was determined after OVA257-264-Kb tetramer staining
or by intracellular IFN-g staining upon 5 hr incubation with OVA257-264
(5 mg/ml) in the presence of Brefeldin (Janssen et al., 2003, 2005).
To study the cytolytic capacity of the CD8+ T cells, splenocytes
were cultured with irradiated MEC.B7.Sig-OVA cells (1:10) for 6
days. Restimulated splenocytes were evaluated by JAM test as
previously described (Janssen et al., 2003), by means of 3[H]thymi-
dine-labeled EL-4 cells loaded with OVA257-264 or control peptide.
Specific killing was calculated as follows: (spontaneous c.p.m. 2
experimental c.p.m.) 3 100/spontaneous c.p.m. The proliferative
capacity of OVA-specific CD4+ T cells was determined by standard
lymphocyte stimulation assay. In brief, splenocytes were cultured in
96-well plates (2 3 106 cells/well) with medium or increasing doses
of OVA323-339 for 72 hr, after which the cells were pulsed for 8 hr
with 3[H]thymidine (0.1 mCi/well). Proliferation was calculated by
dividing the incorporated c.p.m. of the OVA323-339-stimulated cells
by the c.p.m. of medium-stimulated cells.
Generation of DC Subsets and T Cell Purification
BM cells were isolated and cultured with Flt3L (200 ng/ml) or with
GM-CSF (20 ng/ml) as previously described (Brasel et al., 2000;
Lutz et al., 1999). After 8 days, DCs were sorted in subpopulations
based on their expression of B220 and CD11c or expression of
mouse plasmacytoid dendritic cell antigen-1 (mPDCA-1) by MACS
microbeads. DCs were thoroughly analyzed for surface marker ex-
pression and cytokine expression upon TLR ligation. Purity of all
the different populations was >97%. OT-I (CD8+) and OT-II (CD4+)
T cells were isolated by negative selection by T cell isolation kits
(Miltenyi) and labeled with CFSE as described previously (Tabeta
et al., 2006).
Determination of Priming and Crosspriming by Different DC
Subsets
To determine the ability of DC subsets to prime purified CFSE-la-
beled OT-I or OT-II T cells, we cultured different DC subsets in
IMDM (supplemented with 10% FCS, 2% penicillin/streptomycin,
50 mM b-mercaptoethanol) at a concentration of 1 3 106 cells/well
in the presence or absence of 3 3 106 UV-treated act-mOVA cells
(240 mJ/cm2) on a Kbm1 background (to avoid direct presentation
of act-mOVA cells to T cells). In addition, different DC subsets
were cultured either in direct or indirect contact with apoptotic
act-mOVA cells via 0.02 mm Anapore membrane Tissue culture
inserts (Nalgene Nunc International, Rochester, NY). Cells were in-
cubated at 37ºC, 5% CO2, and after 24 hr incubation, either 2 3
105 CFSE-labeled OT-I or OT-II T cells were added to each well. After
an additional 72 hr of incubation, CFSE intensity of OT-I or OT-II cells
was determined by flow-cytometry analysis with specific fluores-
cent-labeled antibodies for CD4 or CD8 in combination with Va2.
In addition, OT-I or OT-II T cells were cultured alone or in the pres-
ence of act-mOVA/Kbm1 cells as negative controls, or with CD4- or
CD8-specific OVA peptides as a positive control.
Isolation of DC Subsets from Spleen
Spleens were treated with 2 mg/ml of collagenase D (Roche Diag-
nostics, Germany) in 10 mM HEPES-NaOH, 150 mM NaCl, 5 mM
KCl, 1 mM MgCl2, 1.8 mM CaCl2 to obtain a single-cell suspension.
Subsequently, splenocytes were washed with PBS containing 0.5%
FBS and 2% EDTA, and CD11b2/CD11c+ cells were isolated with
microbeads (Milteny Biotec, Auburn, CA). After 15 min incubation,
cells were washed and separated by means of a LS MACS column.
Subsequently, DCs were labeled with fluorochrome-conjugated an-
tibodies specific for MHC-II, CD8, and B220 and directly separated
by FACS sort. CD11b2/CD11c+ DCs were sorted based on MHC
class II+ and CD8 and B220 surface expression, and CD8+/B220+,
CD82/B220+, CD8+/B2202, and CD82/B2202 DC subsets were
collected and exposed to g-irradiated act-mOVA-Kbm-1 cells. OT-I
and OT-II proliferation was assessed as described above.
Immunostaining of SIINFEKL/Kb Complex
Bone marrow-derived GM-CSF and B2202 Flt3L-treated DCs from
wild-type and 3d mice were cocultured in the presence of g-irradi-
ated act-mOVA splenocytes (at various ratios). After 24 hr, cells
were stained with Alexa Fluor-750 coupled mAb 25-D1.16 specifi-
cally recognizing the SIINFEKL/Kb complex (Porgador et al., 1997).
Statistical Analysis
All experiments were repeated at least three times, and comparison
between groups was performed by one-way analysis of variance
(ANOVA).
Supplemental Data
Supplemental Data include six figures and two movies and can be
found with this article online at http://www.immunity.com/cgi/
content/full/24/6/787/DC1/.
Acknowledgments
The work was supported by funding from the National Institutes of
Health (#AI 054523). E.J.’s work was supported by a Leukemia and
Lymphoma Society Grant (3248-05).
Received: October 5, 2005
Revised: March 15, 2006
Accepted: March 30, 2006
Published: June 20, 2006
References
Akira, S., and Takeda, K. (2004). Toll-like receptor signalling. Nat.
Rev. Immunol. 4, 499–511.
Albert, M.L., Sauter, B., and Bhardwaj, N. (1998). Dendritic cells ac-
quire antigen from apoptotic cells and induce class I-restricted
CTLs. Nature 392, 86–89.
Asselin-Paturel, C., and Trinchieri, G. (2005). Production of type I in-
terferons: plasmacytoid dendritic cells and beyond. J. Exp. Med.
202, 461–465.
Barnhart, B.C., Alappat, E.C., and Peter, M.E. (2003). The CD95 type
I/type II model. Semin. Immunol. 15, 185–193.
Barrat, F.J., Meeker, T., Gregorio, J., Chan, J.H., Uematsu, S., Akira,
S., Chang, B., Duramad, O., and Coffman, R.L. (2005). Nucleic acids
of mammalian origin can act as endogenous ligands for Toll-like re-
ceptors and may promote systemic lupus erythematosus. J. Exp.
Med. 202, 1131–1139.
Bave, U., Magnusson, M., Eloranta, M.L., Perers, A., Alm, G.V., and
Ronnblom, L. (2003). Fc gamma RIIa is expressed on natural IFN-
alpha-producing cells (plasmacytoid dendritic cells) and is required
for the IFN-alpha production induced by apoptotic cells combined
with lupus IgG. J. Immunol. 171, 3296–3302.
Belz, G.T., Smith, C.M., Eichner, D., Shortman, K., Karupiah, G., Car-
bone, F.R., and Heath, W.R. (2004). Cutting edge: conventional CD8
alpha+ dendritic cells are generally involved in priming CTL immu-
nity to viruses. J. Immunol. 172, 1996–2000.
Belz, G.T., Shortman, K., Bevan, M.J., and Heath, W.R. (2005).
CD8alpha+ dendritic cells selectively present MHC class I-restricted
noncytolytic viral and intracellular bacterial antigens in vivo. J. Im-
munol. 175, 196–200.
Blander, J.M., and Medzhitov, R. (2004). Regulation of phagosome
maturation by signals from toll-like receptors. Science 304, 1014–
1018.
Boule, M.W., Broughton, C., Mackay, F., Akira, S., Marshak-Roth-
stein, A., and Rifkin, I.R. (2004). Toll-like receptor 9-dependent

Immunity798
and -independent dendritic cell activation by chromatin-immuno-
globulin G complexes. J. Exp. Med. 199, 1631–1640.
Brasel, K., De Smedt, T., Smith, J.L., and Maliszewski, C.R. (2000).
Generation of murine dendritic cells from flt3-ligand-supplemented
bone marrow cultures. Blood 96, 3029–3039.
Brockstedt, D.G., Bahjat, K.S., Giedlin, M.A., Liu, W., Leong, M.,
Luckett, W., Gao, Y., Schnupf, P., Kapadia, D., Castro, G., et al.
(2005). Killed but metabolically active microbes: a new vaccine
paradigm for eliciting effector T-cell responses and protective
immunity. Nat. Med. 11, 853–860.
den Haan, J.M., Lehar, S.M., and Bevan, M.J. (2000). CD8(+) but not
CD8(-) dendritic cells cross-prime cytotoxic T cells in vivo. J. Exp.
Med. 192, 1685–1696.
Dennert, G. (2002). Elimination of virus-specific cytotoxic T cells in
the liver. Crit. Rev. Immunol. 22, 1–11.
Ehst, B.D., Ingulli, E., and Jenkins, M.K. (2003). Development of
a novel transgenic mouse for the study of interactions between
CD4 and CD8 T cells during graft rejection. Am. J. Transplant. 3,
1355–1362.
Finberg, R., Greene, M.I., Benacerraf, B., and Burakoff, S.J. (1979).
The cytolytic T lymphocyte response to trinitrophenyl-modified syn-
geneic cells. I. Evidence for antigen-specific helper T cells. J. Immu-
nol. 123, 1205–1209.
Gallucci, S., Lolkema, M., and Matzinger, P. (1999). Natural adju-
vants: endogenous activators of dendritic cells. Nat. Med. 5, 1249–
1255.
Gorla, R., Imberti, L., Prati, E., Brugnoni, D., Caligaris, S., Cattaneo,
R., Albertini, A., and Primi, D. (1994). Differential priming to pro-
grammed cell death of superantigen-reactive lymphocytes of HIV
patients. AIDS Res. Hum. Retroviruses 10, 1097–1103.
Herrmann, M., Voll, R.E., Zoller, O.M., Hagenhofer, M., Ponner, B.B.,
and Kalden, J.R. (1998). Impaired phagocytosis of apoptotic cell
material by monocyte-derived macrophages from patients with
systemic lupus erythematosus. Arthritis Rheum. 41, 1241–1250.
Hoebe, K., Du, X., Georgel, P., Janssen, E., Tabeta, K., Kim, S.O.,
Goode, J., Lin, P., Mann, N., Mudd, S., et al. (2003a). Identification
of Lps2 as a key transducer of MyD88-independent TIR signalling.
Nature 424, 743–748.
Hoebe, K., Du, X., Goode, J., Mann, N., and Beutler, B. (2003b). Lps2:
a new locus required for responses to lipopolysaccharide, revealed
by germline mutagenesis and phenotypic screening. J. Endotoxin
Res. 9, 250–255.
Hoebe, K., Janssen, E.M., Kim, S.O., Alexopoulou, L., Flavell, R.A.,
Han, J., and Beutler, B. (2003c). Upregulation of costimulatory
molecules induced by lipopolysaccharide and double-stranded
RNA occurs by Trif-dependent and Trif-independent pathways.
Nat. Immunol. 4, 1223–1229.
Hoebe, K., Georgel, P., Rutschmann, S., Du, X., Mudd, S., Crozat, K.,
Sovath, S., Shamel, L., Hartung, T., Zahringer, U., and Beutler, B.
(2005). CD36 is a sensor of diacylglycerides. Nature 433, 523–527.
Huang, A.Y., Bruce, A.T., Pardoll, D.M., and Levitsky, H.I. (1996).
In vivo cross-priming of MHC class I-restricted antigens requires
the TAP transporter. Immunity 4, 349–355.
Ishii, K.J., Coban, C., Kato, H., Takahashi, K., Torii, Y., Takeshita, F.,
Ludwig, H., Sutter, G., Suzuki, K., Hemmi, H., et al. (2006). A Toll-like
receptor-independent antiviral response induced by double-
stranded B-form DNA. Nat. Immunol. 7, 40–48.
Iwasaki, A., and Medzhitov, R. (2004). Toll-like receptor control of the
adaptive immune responses. Nat. Immunol. 5, 987–995.
Janssen, E.M., Lemmens, E.E., Wolfe, T., Christen, U., von Herrath,
M.G., and Schoenberger, S.P. (2003). CD4+ T cells are required for
secondary expansion and memory in CD8+ T lymphocytes. Nature
421, 852–856.
Janssen, E., Droin, N., Lemmens, E., Pinkoski, M., Bensinger, S.,
Ehst, B., Griffith, T., Green, D., and Schoenberger, S. (2005). CD4
T cell help controls CD8 T cell memory via trail-mediated activa-
tion-induced cell-death. Nature 434, 88–93.
Jiang, Z., Georgel, P., Du, X., Shamel, L., Sovath, S., Mudd, S.,
Huber, M., Kalis, C., Keck, S., Galanos, C., et al. (2005). CD14 is
required for MyD88-independent LPS signaling. Nat. Immunol. 6,
565–570.
Kato, H., Sato, S., Yoneyama, M., Yamamoto, M., Uematsu, S., Mat-
sui, K., Tsujimura, T., Takeda, K., Fujita, T., Takeuchi, O., and Akira,
S. (2005). Cell type-specific involvement of RIG-I in antiviral
response. Immunity 23, 19–28.
Lau, C.M., Broughton, C., Tabor, A.S., Akira, S., Flavell, R.A., Ma-
mula, M.J., Christensen, S.R., Shlomchik, M.J., Viglianti, G.A., Rifkin,
I.R., and Marshak-Rothstein, A. (2005). RNA-associated autoanti-
gens activate B cells by combined B cell antigen receptor/Toll-like
receptor 7 engagement. J. Exp. Med. 202, 1171–1177.
Le Bon, A., and Tough, D.F. (2002). Links between innate and adap-
tive immunity via type I interferon. Curr. Opin. Immunol. 14, 432–436.
Le Bon, A., Etchart, N., Rossmann, C., Ashton, M., Hou, S., Gewert,
D., Borrow, P., and Tough, D.F. (2003). Cross-priming of CD8+
T cells stimulated by virus-induced type I interferon. Nat. Immunol.
4, 1009–1015.
Leadbetter, E.A., Rifkin, I.R., Hohlbaum, A.M., Beaudette, B.C.,
Shlomchik, M.J., and Marshak-Rothstein, A. (2002). Chromatin-IgG
complexes activate B cells by dual engagement of IgM and Toll-
like receptors. Nature 416, 603–607.
Li, M., Carpio, D.F., Zheng, Y., Bruzzo, P., Singh, V., Ouaaz, F., Medz-
hitov, R.M., and Beg, A.A. (2001). An essential role of the NF-kappa
B/Toll-like receptor pathway in induction of inflammatory and
tissue-repair gene expression by necrotic cells. J. Immunol. 166,
7128–7135.
Liu, Y.J. (2005). IPC: professional type 1 interferon-producing cells
and plasmacytoid dendritic cell precursors. Annu. Rev. Immunol.
23, 275–306.
Lovgren, T., Eloranta, M.L., Bave, U., Alm, G.V., and Ronnblom, L.
(2004). Induction of interferon-alpha production in plasmacytoid
dendritic cells by immune complexes containing nucleic acid re-
leased by necrotic or late apoptotic cells and lupus IgG. Arthritis
Rheum. 50, 1861–1872.
Lutz, M.B., Kukutsch, N., Ogilvie, A.L., Rossner, S., Koch, F., Ro-
mani, N., and Schuler, G. (1999). An advanced culture method for
generating large quantities of highly pure dendritic cells from mouse
bone marrow. J. Immunol. Methods 223, 77–92.
Martin, D.A., and Elkon, K.B. (2005). Autoantibodies make a U-turn:
the toll hypothesis for autoantibody specificity. J. Exp. Med. 202,
1465–1469.
Matzinger, P. (1994). Tolerance, danger, and the extended family.
Annu. Rev. Immunol. 12, 991–1045.
Montoya, M., Schiavoni, G., Mattei, F., Gresser, I., Belardelli, F., Bor-
row, P., and Tough, D.F. (2002). Type I interferons produced by
dendritic cells promote their phenotypic and functional activation.
Blood 99, 3263–3271.
Okabe, Y., Kawane, K., Akira, S., Taniguchi, T., and Nagata, S. (2005).
Toll-like receptor-independent gene induction program activated by
mammalian DNA escaped from apoptotic DNA degradation. J. Exp.
Med. 202, 1333–1339.
Porgador, A., Yewdell, J.W., Deng, Y., Bennink, J.R., and Germain,
R.N. (1997). Localization, quantitation, and in situ detection of spe-
cific peptide-MHC class I complexes using a monoclonal antibody.
Immunity 6, 715–726.
Ronchetti, A., Rovere, P., Iezzi, G., Galati, G., Heltai, S., Protti, M.P.,
Garancini, M.P., Manfredi, A.A., Rugarli, C., and Bellone, M. (1999).
Immunogenicity of apoptotic cells in vivo: role of antigen load, anti-
gen-presenting cells, and cytokines. J. Immunol. 163, 130–136.
Schnare, M., Barton, G.M., Holt, A.C., Takeda, K., Akira, S., and
Medzhitov, R. (2001). Toll-like receptors control activation of adap-
tive immune responses. Nat. Immunol. 2, 947–950.
Seong, S.Y., and Matzinger, P. (2004). Hydrophobicity: an ancient
damage-associated molecular pattern that initiates innate immune
responses. Nat. Rev. Immunol. 4, 469–478.
Shortman, K., and Liu, Y.J. (2002). Mouse and human dendritic cell
subtypes. Nat. Rev. Immunol. 2, 151–161.
Smith, C.M., Belz, G.T., Wilson, N.S., Villadangos, J.A., Shortman, K.,
Carbone, F.R., and Heath, W.R. (2003). Cutting edge: conventional
CD8 alpha+ dendritic cells are preferentially involved in CTL priming

TLR-Independent Adjuvanticity for CTL Responses799
after footpad infection with herpes simplex virus-1. J. Immunol. 170,
4437–4440.
Smyth, M.J., Cretney, E., Kelly, J.M., Westwood, J.A., Street, S.E.,
Yagita, H., Takeda, K., Dommelen, S.L., Degli-Esposti, M.A., and
Hayakawa, Y. (2005). Activation of NK cell cytotoxicity. Mol. Immu-
nol. 42, 501–510.
Steinman, R.M., Turley, S., Mellman, I., and Inaba, K. (2000). The
induction of tolerance by dendritic cells that have captured apopto-
tic cells. J. Exp. Med. 191, 411–416.
Tabeta, K., Hoebe, K., Janssen, E.M., Du, X., Georgel, P., Crozat, K.,
Mudd, S., Mann, N., Sovath, S., Goode, J., et al. (2006). The Unc93b1
mutation 3d disrupts exogenous antigen presentation and signaling
via Toll-like receptors 3, 7 and 9. Nat. Immunol. 7, 156–164.
Theofilopoulos, A.N., Baccala, R., Beutler, B., and Kono, D.H. (2005).
Type I interferons in immunity and autoimmunity. Annu. Rev. Immu-
nol. 23, 307–335.
Vollmer, J., Tluk, S., Schmitz, C., Hamm, S., Jurk, M., Forsbach, A.,
Akira, S., Kelly, K.M., Reeves, W.H., Bauer, S., and Krieg, A.M.
(2005). Immune stimulation mediated by autoantigen binding sites
within small nuclear RNAs involves Toll-like receptors 7 and 8.
J. Exp. Med. 202, 1575–1585.
Winau, F., Weber, S., Sad, S., de Diego, J., Hoops, S.L., Breiden, B.,
Sandhoff, K., Brinkmann, V., Kaufmann, S.H., and Schaible, U.E.
(2006). Apoptotic vesicles crossprime CD8 T cells and protect
against tuberculosis. Immunity 24, 105–117.
Yoneyama, M., Kikuchi, M., Natsukawa, T., Shinobu, N., Imaizumi,
T., Miyagishi, M., Taira, K., Akira, S., and Fujita, T. (2004). The RNA
helicase RIG-I has an essential function in double-stranded RNA-
induced innate antiviral responses. Nat. Immunol. 5, 730–737.
Yrlid, U., and Wick, M.J. (2000). Salmonella-induced apoptosis of in-
fected macrophages results in presentation of a bacteria-encoded
antigen after uptake by bystander dendritic cells. J. Exp. Med.
191, 613–624.





























