Embed Size (px)
Citation preview

[CANCER RESEARCH 33, 2034-2041, September 1973]
SUMMARY
It was found that one Reuber hepatoma and seven Morris hepatomas with different growth rates and variousdegrees of differentiation contained pyruvate carboxylase(PC) activity ranging from 3 to 113% of control values. Thelevels of PC activity in the regenerating liver, spontaneous mouse hepatoma, and host livers of tumor-bearing ratswere estimated in the range of normal rat liver. In the groupof tumors studied, two hepatomas of intermediate growthrate and four highly and well-differentiated, slow-growingtumors, like rat fetal liver, had negligible or little PC activity. However, one hepatoma, 9618A, had a normal levelof PC activity. Antibodies prepared in rabbits against thepurified mitochondrial enzyme showed identical titrationcurves against the nuclear and cytosol PC of liver. A singlecontinuous precipitation line was obtained on Ouchterlonydouble-diffusion precipitation analysis when purified mitochondrial enzyme, crude mitochondrial preparations of rattissues and hepatomas, were compared. It was also foundthat the immunological, enzymatic, and electrophoreticproperties of the tumor PC were the same as those of normal adult and newborn rat liver. The amount of enzymeprotein was quantitatively reduced in all but one of theliver tumors studied.
INTRODUCTION
PC4 [pyruvate:carbon dioxide ligase (ADP), EC6.4. 1. 1j catalyzes the carboxylation of pyruvate to formoxalacetate which is an important intermediate in the reactions involved in the processes of gluconeogenesis, lipogenesis, energy production, and other tricarboxylic acidcycle activities (17, 28). Its role in these functions is supported by the finding that when rat liver mitochondria areincubated with an excess of pyruvate, NaH “CO3,activators, and cofactors necessary for carboxylation, theprincipal products containing incorporated ‘4Care citrate,malate, fumarate, and aspartate (9, 25), which are precursors of glucose, fatty acids, proteins, and nucleic acids.
1 This investigation was supported in part by American Cancer Society
Grant BC-72-2l3.2 Recipient of General Research Support Grant 5 sol RR05431.
3 Recipient of USPHS Grant Ca 10729.
4 The abbreviations used are: PC, pyruvate carboxylase; STE. 0.3 M
sucrose-S m@ N-Tris-sulfonic acid- I m@t EDTA; TCA, trichloroaceticacid.
Received March 16, 1973; accepted May 23, 1973.
It is probable that the function of this enzyme is importantfor the normal growth and metabolism of liver.
It was shown (23, 29, 33, 34) that the activities of enzymes important in gluconeogenesis are negatively relatedto the growth rate of liver tumors on the basis of studiesof glucose 6-phosphatase, fructose-l ,6-diphosphatase,and phosphoenolpyruvate carboxykinase. However, thescant data on PC were based on assays of total enzyme activity, and no data were available on mitochondrial PC inthe highly differentiated, slow-growing hepatomas.
The elegant demonstration of isoenzyme alterations asanomalies ofgene expression in the experimental hepatomasand the characteristic resemblance of the rapidly growing,poorly differentiated hepatoma to the rat fetal liver provide important knowledge for our understanding of themolecular basis of cancer (7, 35—37).Preliminary observations that rat fetal liver and slow-growing minimal-deviation hepatoma 9633 have negligible PC activity promptedthis comparative study of PC in the liver-liver tumor modelsystem. Comparisons were made of: rapidly growingtissues versus quiescent and slow-growing tissues, hostor normal liver versus hepatoma, well-differentiated hepatoma versus poorly differentiated hepatoma, transplantable rat hepatoma versus spontaneous mouse hepatomaand fetal versus adult liver.
Some of the specific questions we wish to ask are: Areisoenzymes of PC involved in liver-liver tumor systems?Is there a correlation between PC activity and the growthrate of livers and liver tumors, including the highly differentiated hepatomas? Is the observed reduction in PCactivity in tumor mitochondria due to a difference in thecontent of enzyme protein or to a difference in quality ofthe enzyme protein? This paper is concerned only with theproblems related to these 3 questions.
MATERIALS AND METHODS
Animals and Hepatomas. Tumor-bearing and normalmale rats were housed in individual cages under standardconditions (12-hr-cycle lighting schedule, 72°F). Purinalaboratory chow and water were available ad libilum. Thetumors were transplanted bilaterally s.c. or i.m. The growthrate, histological classification, and generation of the 8hepatoma lines used in this study are summarized in Table1. Host livers and livers from 1 strain of normal male rats(Buffalo) and 2 stocks of male rats (Sprague-Dawley and
2034 CANCER RESEARCH VOL. 33
Enzymatic and Immunological Studies on Pyruvate Carboxylasein Livers and Liver Tumors'
Lillie 0. Chang2 and Harold P. Morris3Department of Pediatrics, University of Virginia School of Medicine, Charlottesville, Virginia, 22901 [L. 0. C.], and Department of Biochemistry,Howard University School of Medicine, Washington, D. C. 21'XXH[H. P. M.]
on March 26, 2018. © 1973 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from
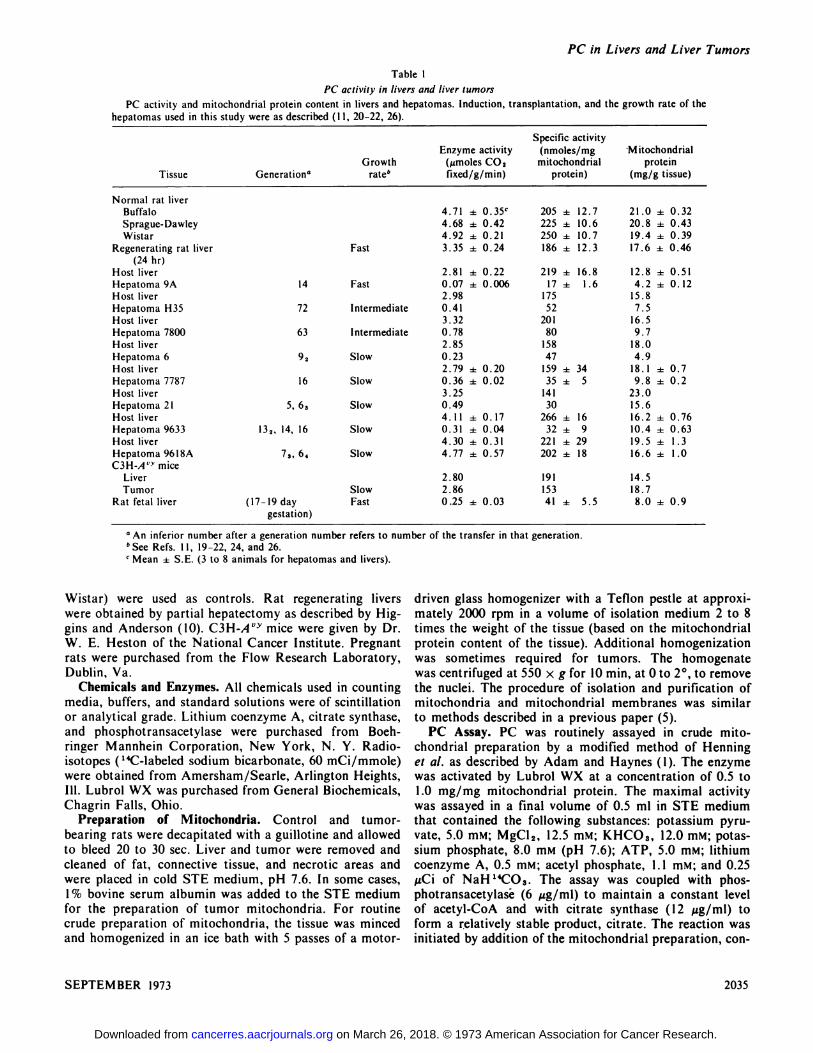
SpecificactivityEnzymeactivity(nmoles/mgMitochondrialGrowth(@imoles
CO,mitochondrialproteinTissueGeneration―ratebfixed/g/min)protein)(mg/gtissue)Normal
ratliverBuffalo4.71± 0.35c205 ± 12.721.0 ±0.32Sprague-Dawley4.68±0.42225 ± 10.620.8 ±0.43Wistar4.92± 0.21250 ± 10.719.4 ±0.39Regenerating
rat liverFast3.35 ± 0.24186 ± 12.317.6 ±0.46(24hr)Host
liver2.81 ± 0.22219 ± 16.812.8 ±0.51Hepatoma9A14Fast0.07±0.00617 ± 1.64.2 ±0.12Host
liver2.9817515.8HepatomaH3572Intermediate0.41527.5Hostliver3.3220116.5Hepatoma780063Intermediate0.78809.7Hostliver2.8515818.0Hepatoma692Slow0.23474.9Hostliver2.79
±0.20159 ± 3418.1 ±0.7Hepatoma778716Slow0.36 ± 0.0235 ± 59.8 ±0.2Host
liver3.2514123.0Hepatoma215,63Slow0.493015.6Hostliver4.11
± 0.17266 ± 1616.2 ±0.76Hepatoma9633132, 14,16Slow0.31 ± 0.0432 ± 910.4 ±0.63Hostliver4.30
±0.31221 ± 2919.5 ±1.3Hepatoma96l8A73,64Slow4.77±0.57202 ± 1816.6 ±1.0C3H-A@
miceLiver2.8019114.5TumorSlow2.8615318.7Rat
fetal liver(17-19 daygestation)Fast0.25
± 0.0341 ± 5.58.0 ± 0.9
PC in Livers and Liver Tumors
Table I
PC activity in livers and liver tumors
PC activity and mitochondrial protein content in livers and hepatomas. Induction, transplantation, and the growth rate of thehepatomas used in this study were as described (I I, 20—22,26).
a An inferior number after a generation number refers to number of the transfer in that generation.
b See Refs. I 1, 19—22, 24, and 26.
C Mean ± SE. (3 to 8 animals for hepatomas and livers).
driven glass homogenizer with a Teflon pestle at approximately 2000 rpm in a volume of isolation medium 2 to 8times the weight of the tissue (based on the mitochondrialprotein content of the tissue). Additional homogenizationwas sometimes required for tumors. The homogenatewas centrifuged at 550 x g for 10 mm, at 0 to 2°,to removethe nuclei. The procedure of isolation and purification ofmitochondria and mitochondrial membranes was similarto methods described in a previous paper (5).
PC Assay. PC was routinely assayed in crude mitochondrial preparation by a modified method of Henninget a!. as described by Adam and Haynes (1). The enzymewas activated by Lubrol WX at a concentration of 0.5 to1.0 mg/mg mitochondrial protein. The maximal activitywas assayed in a final volume of 0.5 ml in STE mediumthat contained the following substances: potassium pyruvate, 5.0 mM; MgCl2, 12.5 mM; KHCO3, 12.0 mM; potassium phosphate, 8.0 mM (pH 7.6); AlP, 5.0 mM; lithiumcoenzyme A, 0.5 mM; acetyl phosphate, 1. 1 mM; and 0.25zCi of NaH 14(@@ The assay was coupled with phosphotransacetylasé (6 @cg/ml)to maintain a constant levelof acetyl-CoA and with citrate synthase (1 2 zg/ml) toform a relatively stable product, citrate. The reaction wasinitiated by addition of the mitochondrial preparation, con
Wistar) were used as controls. Rat regenerating liverswere obtained by partial hepatectomy as described by Higgins and Anderson (10). C3H-A@' mice were given by Dr.W. E. Heston of the National Cancer Institute. Pregnantrats were purchased from the Flow Research Laboratory,Dublin, Va.
Chemicals and Enzymes. All chemicals used in countingmedia, buffers, and standard solutions were of scintillationor analytical grade. Lithium coenzyme A, citrate synthase,and phosphotransacetylase were purchased from Boehringer Mannhein Corporation, New York, N. Y. Radioisotopes ( ‘IC-labeled sodium bicarbonate, 60 mCi/mmole)were obtained from Amersham/Searle, Arlington Heights,Ill. Lubrol WX was purchased from General Biochemicals,Chagrin Falls, Ohio.
Preparation of Mitochondria. Control and tumorbearing rats were decapitated with a guillotine and allowedto bleed 20 to 30 sec. Liver and tumor were removed andcleaned of fat, connective tissue, and necrotic areas andwere placed in cold STE medium, pH 7.6. In some cases,1% bovine serum albumin was added to the STE mediumfor the preparation of tumor mitochondria. For routinecrude preparation of mitochondria, the tissue was mincedand homogenized in an ice bath with 5 passes of a motor
SEPTEMBER 1973 2035
on March 26, 2018. © 1973 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from

Li/lie 0. Chang and Harold P. Morris
taming approximately 0.2 mg mitochondrial protein, tothe reaction mixture and incubation at 37°with slow shaking for 5 mm. The reaction was stopped by addition of I mlof 5% TCA. Following centrifugation at 550 x g for 5 mm,0.5 ml of the supernatant fluid was cleared of acid-labile“CO2by 100% CO2, which was bubbled through it for2 mm. The “4Cwas counted in a liquid scintillation spectrometer. Enzyme activity is defined as 1ttmoles CO2 fixedper g of tissue per mm or nmoles CO2 fixed per mg mitochondrial protein per mm at 37°. Incubations in the absence of pyruvate or enzyme were carried out as blanks. Ablank incorporation of about 80 to 200 cpm was found;this value was subtracted from all assay measurements.The mitochondrial suspension without Lubrol treatmentwas also included and gave an incorporation of “CO2thatwas usually very low, indicating that the mitochondria wereintact.
Measurement of Radioactivity. 14(@radioactivity wasmeasured by scintillation counting in a Beckman liquidscintillation spectrometer. The scintillant contained PPO,naphthalene, Cellosolve, and toluene (1 : 20: 100: 150)(w/w, v/v). Sufficient counts were recorded to give anaccuracy of 1.0 to 1.5%; the efficiency ofcounting was about96% for “C.
Identification of Reaction Products. Separation of radioactive carboxylic acids on Dowex I-formate resin was performed according to the method of Busch et a!. (4) afterremoval of TCA with ether and neutralization of the extracts. After the nonacid components had been washed offthe column with water, the acids were eluted with 4 Nformic acid. The fractions containing the peak activity werepooled, air dried, and taken up in a small quantity of water.Aliquots of the sample from each step were counted, andthe recovery of the activity was calculated. The labeledcompounds were identified by means of silica gel thin-layerchromatography (3 1) and high-voltage paper electrophoresis (13).
Thin-Layer Chromatography. The labeled acids andstandard solutions were applied on an Eastman Type K301 R silica gel sheet and were developed in a well-saturatedtank with methanol : 5 N ammonia solution (80 : 20) for2 hr. The acids were located with a Bromcresol green aerosol (yellow spots on blue background). They were scrapedoff the plate with the silica gel and transferred directly tocounting vials containing 15 ml of counting solution (5 gPPO, 0.3 g POPOP, and 4 g Cab-O-Sil in 1 liter toluene).
High-Voltage Paper Electrophoresis (HorizontalPlates). The labeled acids and standard solutions were applied to a strip of Whatman No. 3MM paper (6 x 28.5inches). Electrophoresiswas carried out in 2 N aceticacid solution (pH 2.2) at 7 kV, 29 to 33 ma, for 2 hr. Thepaper was dried and treated with steam to evaporate tracesof acetic acid. Citric acid, malate, and other organic acidswere detected by spraying with 0.05% methyl red in 0.01 Mphosphate buffer, pH 7.7. The acid appeared as red spotson a yellow background. The distribution of radioactivityon the electrophoretogram was located by a Packard scanncr. The areas in the electrophoretogram correspondingto the radioactive peaks were cut and placed in countingvials containing the scintillant.
Purification of PC. The procedure for the preparationof rat liver mitochondrial PC described by McClure et a!.(18) was followed, with slight modifications. The initial10 to 40% homogenate (depending on the mitochondrialprotein content of the tissue) was centrifuged at 50 x g for10 mm, and the pellet was resuspended in STE mediumand centrifuged again at 50 x g for 10 mm. The combinedsupernatant fractions were centrifuged at 9750 x g for 10mm. The resulting pellets were washed twice with the sameisolating medium. The remainder of the procedure of PCextraction and purification was the same as that describedby McClure et al. (18). The elution pattern of PC from thesucrose gradient contained 2 protein peaks correspondingto the refractive indices I .3600 and 1.3650. The PC activitypeak coincided with the 1.3650 protein peak. The purityof this peak was examined immunologically and electrophoretically. The average specific activity of PC peakswas 25 units/mg protein. Peaks were pooled and used forpreparation of antisera.
Immunization. Rabbit antiserum against pure PC wasprepared according to the method described by Williamsand Chase (38) and Work and Work (39). Approximately2 mg PC protein in 2 ml solution were well suspended inan equal volume of complete Freund's adjuvant. Theemulsion was injected into 8 sites of a 2 to 3-lb rabbit (3i.m. injections in each side of the rabbit and 2 s.c. injectionsin back of the neck). Six rabbits were used in this experiment. Blood samples taken before immunization served ascontrol sera.
Three additional injections of enzyme were made duringa I-month period. The potency of the antiserum was measured weekly by Ouchterlony double-diffusion-precipitationanalysis.
Double Immunodiffusion in Agar Gel. The circular-wellmethod of double immunodiffusion was used which involves the use of agar plates with wells for both antigensand antibodies. One % lonagar No. 25 was made in barbitone:acetate buffer, pH 8.6 (final concentration of 0.05ionic strength). For the double immunodiffusion, initialtrials to fix a balanced proportion between antigen and antiserum were performed in 2 series, e.g., with a fixed amountof the antigen diffusing against a series of antiserum dilutions and vice versa. The application of the reactants wasstandardized as much as possible in terms of proteinconcentration. After 48 hr of diffusion, the slides werewashed for 48 hr in 0.15 M NaCl and for 24 hr in multiple changes of deionized distilled water. Following thesewashings, the slides were dried and then stained I mm with0.5% Amido schwarz dye in methanol : acetic acid : water(5 : 1: 5) and were washed with the solvent until well decolorized.
Single Radial Immunodiffusion in Agar Gel. The procedure of Mancini et a!. (16) was used for the quantitativeimmunological determination of PC protein. One part of3% (w/v) agar in barbitone : acetate buffer containing 0.01%Merthiolate was mixed with 2 parts of anti-PC antiserumat 48°. The agar: antiserum mixture was poured ontoprewarmed slides to give a layer 2.0 mm thick. After solidification of the gel in a humid atmosphere, wells 2.5 mm indiameter were punched out. These wells were filled with
CANCER RESEARCH VOL. 332036
on March 26, 2018. © 1973 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from

PC in Livers and Liver Tumors
antigen solutions by means of a calibrated microsyringe,with care being taken to deliver accurately measured volumes containing known amounts of protein. At the end of5 days of incubation at 22°,the agar slides were washed for2 to 3 days in 0.15 M NaC1 and then in distilled water.They were dried and stained with Amido schwarz. A standard curve was plotted relating the area of the precipitateto the amount of purified PC preparation added. The relative concentration of PC protein in the crude mitochondrialsamples developed under standard conditions was estimated by the standard curve.
Protein Determination. Protein concentrations were determined by the method of Lowry et al. (15) with bovineserum albumin as a standard. A spectrophotometricmethod (14) was used for the determination of proteincontent in the fractions from the sucrose gradient that wasused in the purification of PC.
RESULTS
Enzyme Activation. The enzyme can be activated eitherby sonic disruption or by the non-ionic detergent LubrolWX. The effectiveness of these 2 methods in dismembration ofmitochondrial preparations was compared by following PC activity and the changes in turbidity at A520. ThePC activity rose during the 1st mm of sonic disruption andremained at a similar level for up to 8 mm, although therate of clearance of the mitochondrial suspension wasfound to be proportional to the time of sonic disruption.
The results of the Lubrol effect on the dismembrationand PC activity in the crude mitochondrial preparationsof liver showed that PC activity increased with increasingLubrol concentration. Four mm after Lubrol treatment,8 to 10-fold increases in activity were observed in mostcases. Concentrations of 0.5 to 1.0 mg Lubrol per mg mitochondrial protein were chosen for routine assay, but concentrations as high as 1.8 mg Lubrol per mg proteincaused no inhibition of the enzyme. The effect of Lubrolon tumor PC activity was similar.
Maximal PC Activity in Liver and Tumor Mitochondria.In order to ensure that the observed differences betweenliver and tumor were not the results of preparative artifacts, the following experiments were performed. (a) Tumormitochondria prepared with the isolation media [used bySordahl and Schwartz (30) and Schnaitman and Greenawait (27)] and 0.25 M sucrose, with and without bovineserum albumin, appeared to be metabolically functional.In the presence of the substrate succinate, the addition ofADP to the mitochondrial preparation caused an increasein respiratory activity. The respiratory control values obtamed for hepatoma H35 were 2.7 to 5.6. (b) STE mediumwas routinely used for the crude preparation of liver mitochondria. The addition of 1% bovine serum albumin toSTE medium for the preparation of tumor mitochondriadid not change the measured amount of PC activity in themitochondria; therefore, STE medium was used for bothliver and tumor mitochondrial preparations. (c) The response of tumor mitochondria to Lubrol treatment in terms
of turbidity change and activation of PC was another mdication that the mitochondria were intact. (d) The PC activity in tumor mitochondria was studied in relation to theconcentrations of the substrates, activator, and cofactors(either reduced to one-fifth or increased to 2-fold the concentrations described in “PCassay―). The PC activity ofhepatoma 9633, at barely detectable levels, was not noticeably altered by the changes. On the other hand, hepatoma9618A and host liver responded to the variations in theconcentrations ofthese reactants in a similar fashion (Chart1).
Intracellular Location of PC. The PC activity in thesubcellular fractions of hepatoma 9633 and host liver wasexamined. About 70% of the enzyme activity in liver islocalized in the crude mitochondrial preparation. About10% was observed in the postmitochondrial fraction and20% was observed in the nuclear fraction.
Qualitative characteristics of the PC present in thesesubcellular fractions of liver were examined immunologically. A single continuous precipitation line was obtainedby Ouchterlony double-diffusion analysis when the mitochondrial, nuclear, and cytosol PC were compared. Furthermore, the results of immunochemical precipitationtests showed that antibodies prepared in rabbits againstthe purified mitochondrial enzyme gave identical titrationcurves against the mitochondrial, nuclear, and cytosolPC of liver (Chart 2). Therefore, there is a high probabilitythat all of the PC activity observed in all the subcellularcompartments is derived from mitochondria and represents preparative artifacts and mitochondrial leakage.
Furthermore, the low enzyme activity in tumor 9633 doesnot appear to be caused by the presence of a dissociatingendogenous inhibitor since the mixing experiment of hostliver and tumor gave the mean activity of the 2 tissues.
Identification of Reaction Products. TCA extracts of thelabeled reaction production products in a 5-mm incubationwere prepared and analyzed by silica gel thin-layer chromatography and high-voltage paper electrophoresis. Asingle distinct peak was observed on the radioactive scanning tracing of the electrophoretogram. This peak, which
005101.520 0 2 4 6 810
Chart I . Effect of concentrations of substrates, activators, and cofactors on PC activity. •, normal rat liver; 0, hepatoma 9618A; and @,
hepatoma 9633. For enzyme assay, see “Materialsand Methods.―
CoA Pyruvate KHCO3
AlP
0 S 10 10 20 2S..M
KCI
0 20 40 60 80.,M
Acetyl P0@
SEPTEMBER 1973 2037
4
.2‘@ mgCl2
0 10 2030
on March 26, 2018. © 1973 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from
Lillie 0. Chang and Harold P. Morris
the 2 minimal-deviation, very slow-growing hepatomas9633 and 9618A. Hepatoma 96l8A had a normal level ofPC activity and a relatively high glycogen content (Ref 24,and our unpublished observations) compared with otherhepatomas, whereas hepatoma 9633, with only about 8% ofhost liver PC activity, contained a negligible amount ofglycogen.
The mitochondrial protein content in the hepatomasvaried from 30 to 80% of their host liver values. These dataare also listed in Table 1.
Table 2 summarizes the results of the Ouchterlonydouble-diffusion precipitation analysis of the anti-PC antiserum of rat liver mitochondria against PC from variousrat tissues and hepatomas. A single continuous precipitation line was observed on Ouchterlony double-diffusionprecipitation pattern when purified mitochondrial PCenzyme, crude mitochondrial preparations of rat livers,and hepatomas were compared. There was a good correla
25 tion between the immunoprecipitation data and the amount
of PC activity measured enzymatically in these tissues. Itcan be said, from these data, that the mitochondrial PC ofrat liver and rat hepatomas appear to have a commonantigen.
Validity of the Immunochemical Evidence for PC ProteinDeficiency in Hepatomas. The homogeneity of the PCpreparations used for preparation of antibodies was exammed by 4 and 7.2% polyacrylamide gel electrophoresis in0.04 M Tris:glycine buffer at pH 8.8 (6, 8). The originalliver homogenate separated 16 protein fractions on 7.2%gel. The number of protein bands gradually decreased inthe following purification steps. The purified enzyme obtamed from the sucrose gradient purification step separatedon 7.2% gel as a single band with a mobility similar to thebovine serum albumin dimer. The enzyme preparation hadan average specific activity of 25 units/mg protein. Asingle immunoprecipitation line produced by the anti-PCantiserum is additional evidence of the purity of the enzyme preparation.
Attempts were made to purify PC from hepatoma H35.It was found that the purified PC from both liver and hepatoma had the same electrophoretic mobility in 4.0 and 7.2%polyacrylamide gel in Tris:glycine buffer. Both liver andtumor PC activity were inhibited by avidin, and the preincubation of the avidin and biotin alleviated the inhibitionaction of avidin. Both liver and tumor PC had the sameimmunoprecipitation reaction with the antiserum madeagainst liver mitochondrial PC. However, the yield andspecific activity of PC were low in the tumor, about 13%of the normal rat liver value.
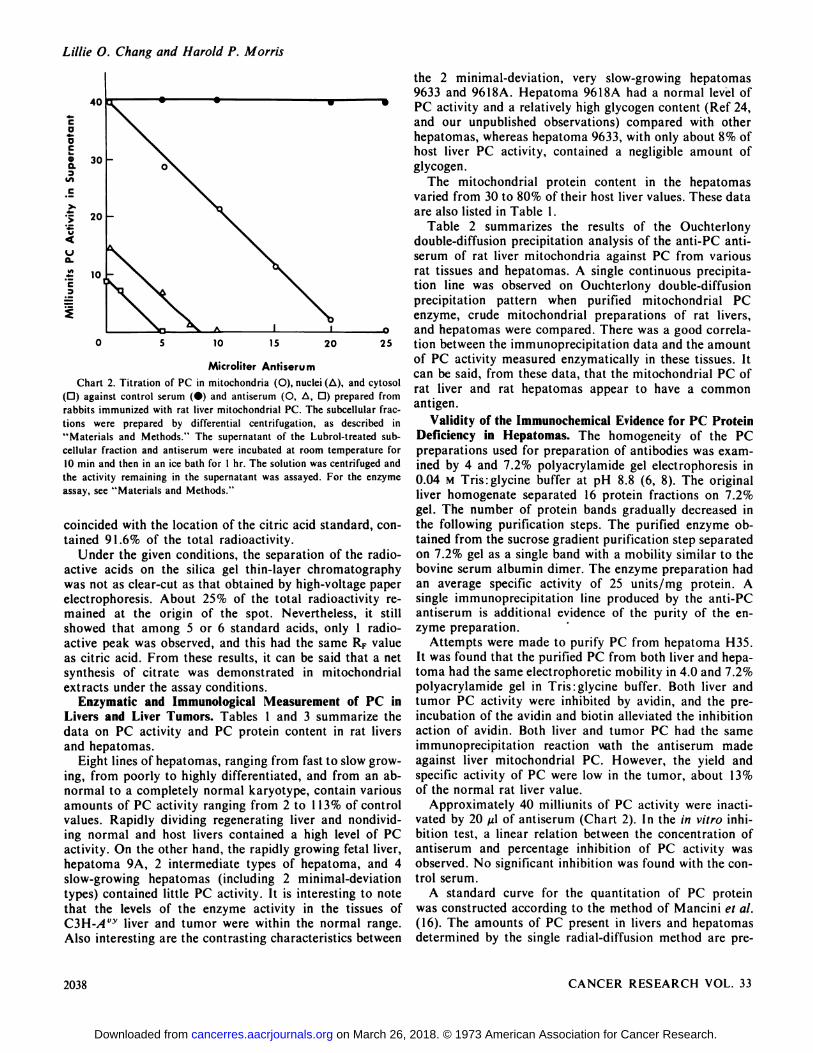
Approximately 40 milliunits of PC activity were mactivated by 20 zl of antiserum (Chart 2). In the in vitro inhibition test, a linear relation between the concentration ofantiserum and percentage inhibition of PC activity wasobserved. No significant inhibition was found with the control serum.
A standardcurve for the quantitation of PC proteinwas constructed according to the method of Mancini et a!.(16). The amounts of PC present in livers and hepatomasdetermined by the single radial-diffusion method are pre
CaaC
a.3
C
>
V
Ua.88
C3
40
30
20
10
0 5 10 15 20
Microliter AntiserumChart 2. Titration of PC in mitochondria (0), nuclei (ti), and cytosol
(0) against control serum (•)and antiserum (0, @,0) prepared fromrabbits immunized with rat liver mitochondrial PC. The subcellular fractions were prepared by differential centrifugation, as described in“Materialsand Methods.― The supernatant of the Lubrol-treated subcellular fraction and antiserum were incubated at room temperature for10 mm and then in an ice bath for I hr. The solution was centrifuged andthe activity remaining in the supernatant was assayed. For the enzymeassay, see “Materialsand Methods.―
coincided with the location of the citric acid standard, contamed 9 1.6% of the total radioactivity.
Under the given conditions, the separation of the radioactive acids on the silica gel thin-layer chromatographywas not as clear-cut as that obtained by high-voltage paperelectrophoresis. About 25% of the total radioactivity remained at the origin of the spot. Nevertheless, it stillshowed that among 5 or 6 standard acids, only 1 radioactive peak was observed, and this had the same RF valueas citric acid. From theseresults,it can be saidthat a netsynthesis of citrate was demonstrated in mitochondrialextracts under the assay conditions.
Enzymatic and Immunological Measurement of PC inLivers and Liver Tumors. Tables 1 and 3 summarize thedata on PC activity and PC protein content in rat liversand hepatomas.
Eight lines of hepatomas, ranging from fast to slow growing, from poorly to highly differentiated, and from an abnormal to a completely normal karyotype, contain variousamounts of PC activity ranging from 2 to I 13% of controlvalues. Rapidly dividing regenerating liver and nondividing normal and host livers contained a high level of PCactivity. On the other hand, the rapidly growing fetal liver,hepatoma 9A, 2 intermediate types of hepatoma, and 4slow-growing hepatomas (including 2 minimal-deviationtypes) contained little PC activity. It is interesting to notethat the levels of the enzyme activity in the tissues ofC3H-A@―liver and tumor were within the normal range.Also interesting are the contrasting characteristics between
2038 CANCER RESEARCH VOL. 33
on March 26, 2018. © 1973 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from
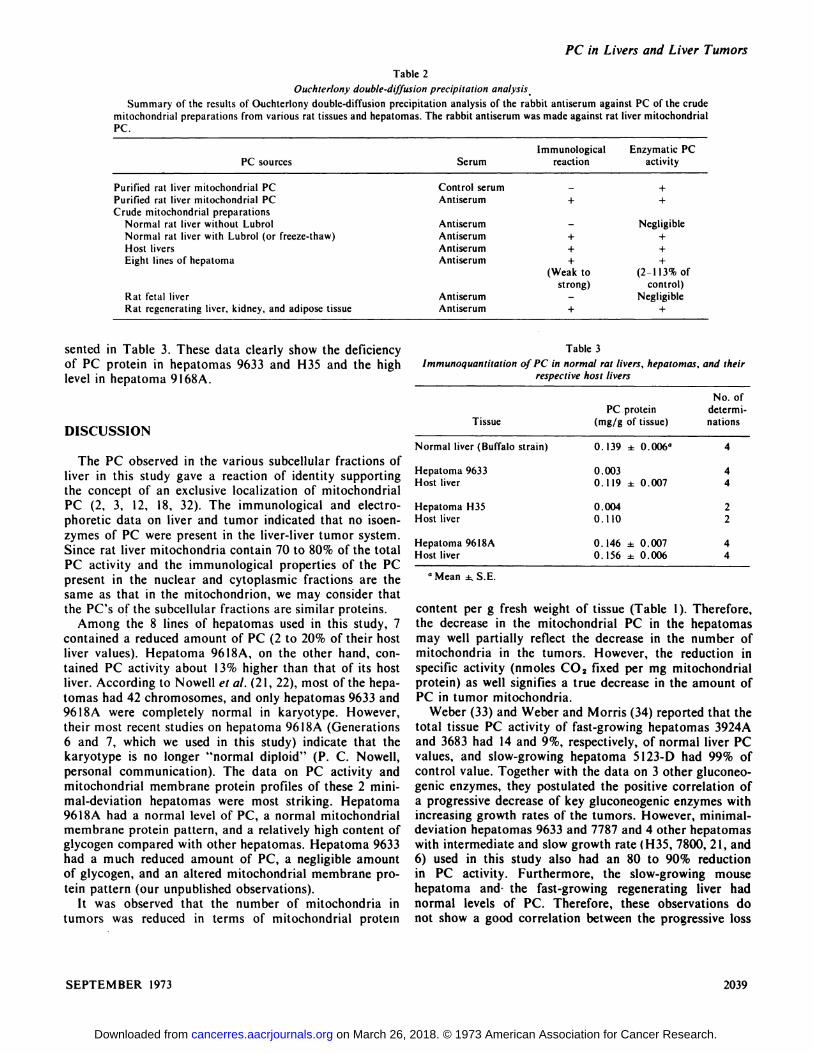
PC sourcesSerumImmunological reactionEnzymaticPC
activityPurified
rat liver mitochondrial PCControlserum—+Purifiedrat liver mitochondrialPCAntiserum++Crude
mitochondrialpreparationsNormalrat liver withoutLubrolAntiserum—NegligibleNormalrat liver with Lubrol (orfreeze-thaw)Antiserum++Host
liversAntiserum++Eightlines of hepatomaAntiserum+
(Weaktostrong)+
(2-lI3%ofcontrol)Rat
fetalliverAntiserum—NegligibleRatregenerating liver, kidney, and adipose tissueAntiserum++
Immunoquantitation crf PC in normal rat livers, hepatomrespective host liversas,
andtheirPC
proteinTissue (mg/g of tissue)No.
ofdetermi
nationsNormal
liver (Buffalo strain) 0. 139 ±0.006―4Hepatoma9633
0.003Hostliver 0.119 ±0.00744Hepatoma
H35 0.004Hostliver 0.11022Hepatoma96l8A
0.146 ±0.007Hostliver 0.156 ±0.0064 4
PC in Livers and Liver Tumors
Table 2
Ouchterlony double-diffusion precipitation analysis
Summary of the results of Ouchterlony double-diffusion precipitation analysis of the rabbit antiserum against PC of the crudemitochondrial preparations from various rat tissues and hepatomas. The rabbit antiserum was made against rat liver mitochondrialPC.
sented in Table 3. These data clearly show the deficiencyof PC protein in hepatomas 9633 and H35 and the highlevel in hepatoma 9l68A.
DISCUSSION
The PC observed in the various subcellular fractions ofliver in this study gave a reaction of identity supportingthe concept of an exclusive localization of mitochondrialPC (2, 3, 12, 18, 32). The immunological and electrophoretic data on liver and tumor indicated that no isoenzymes of PC were present in the liver-liver tumor system.Since rat liver mitochondria contain 70 to 80% of the totalPC activity and the immunological properties of the PCpresent in the nuclear and cytoplasmic fractions are thesame as that in the mitochondrion, we may consider thatthe PC's of the subcellular fractions are similar proteins.
Among the 8 lines of hepatomas used in this study, 7contained a reduced amount of PC (2 to 20% of their hostliver values). Hepatoma 9618A, on the other hand, contamed PC activity about 13% higher than that of its hostliver. According to Nowell et a!. (21, 22), most ofthe hepatomas had 42 chromosomes, and only hepatomas 9633 and96l8A were completely normal in karyotype. However,their most recent studies on hepatoma 9618A (Generations6 and 7, which we used in this study) indicate that thekaryotype is no longer “normaldiploid― (P. C. Nowell,personal communication). The data on PC activity andmitochondrial membrane protein profiles of these 2 minimal-deviation hepatomas were most striking. Hepatoma9618A had a normal level of PC, a normal mitochondrialmembrane protein pattern, and a relatively high content ofglycogen compared with other hepatomas. Hepatoma 9633had a much reduced amount of PC, a negligible amountof glycogen, and an altered mitochondrial membrane protein pattern (our unpublished observations).
It was observed that the number of mitochondria intumors was reduced in terms of mitochondrial protein
Table 3
“Mean @S.E.
content per g fresh weight of tissue (Table I). Therefore,the decrease in the mitochondrial PC in the hepatomasmay well partially reflect the decrease in the number ofmitochondria in the tumors. However, the reduction inspecific activity (nmoles CO2 fixed per mg mitochondrialprotein) as well signifies a true decrease in the amount ofPC in tumor mitochondria.
Weber (33) and Weber and Morris (34) reported that thetotal tissue PC activity of fast-growing hepatomas 3924Aand 3683 had 14 and 9%, respectively, of normal liver PCvalues, and slow-growing hepatoma 5l23-D had 99% ofcontrol value. Together with the data on 3 other gluconeogenic enzymes, they postulated the positive correlation ofa progressive decrease of key gluconeogenic enzymes withincreasing growth rates of the tumors. However, minimaldeviation hepatomas 9633 and 7787 and 4 other hepatomaswith intermediate and slow growth rate (H35, 7800, 21, and6) used in this study also had an 80 to 90% reductionin PC activity. Furthermore, the slow-growing mousehepatoma and- the fast-growing regenerating liver hadnormal levels of PC. Therefore, these observations donot show a good correlation between the progressive loss
SEPTEMBER 1973 2039
on March 26, 2018. © 1973 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from

Li/lie 0. Chang and Harold P. Morris
of PC and growth rate and degree of differentiation ofthe tumor cells. Except for hepatoma 96l8A, the PC inthe other 7 hepatomas was reduced to 2 to 20% of the control values.
Furthermore, immunological and enzymatic studiesindicate thatthe PC proteins of rat liver and liver tumorsappear to be the same. As far as we are aware, this is thefirst report showing that the changes in the amount of PCactivity in the hepatomas result from variations in theamount of PC protein present.
ACKNOWLEDGM ENTS
We are grateful to Dr. R. C. Haynes, Jr., and Dr. J. Lamer for valuable discussions during the course of this work and helpful assistance inthe preparation of this manuscript; to Dr. G. Goldstein for helpful discussions on immunological data; to E. C. Pancake for her technical assistance in the biochemistry work; to C. M. Jackson, J. W. Lewis, andLouise Lawson for their technical assistance in maintaining the hepatomas;and to Cole Squire for her assistance in the immunization work. Wethank Dr. W. E. Heston for his generous gift of C3H.AvY mice and Dr.R. K. Yamazaki for his interest and helpful suggestionsin this investigation.
REFERENCES
I. Adam, P. A. J., and Haynes, R. C., Jr. Control ofHepatic Mitochondrial CO, Fixation by Glucagon, Epinephrine, and Cortisol. J. Biol.Chem., 244: 6444-6450, 1969.
2. Ballard, F. J. Regulation of Gluconeogenesis. Discussion. In: H. D.Soling and B. WilIms (eds.), Ninth Conference of the Gesellschaft furBiologische Chemie, pp. 41-43, 1971.
3. Bottger, J., Wieland, 0., Brdiczka, D., and Pette, D. IntracellularLocalization of Pyruvate Carboxylase and Phosphoenolpyruvate Carboxykinase in Rat Liver. European J. Biochem., 8: 113-I 19, 1969.
4. Busch, H., Hurlbert, R. B., and Potter, V. R. Anion Exchange Chromatography of Acids of the Citric Acid Cycle. J. Biol. Chem., 196:717-727,1952.
5. Chang, L. 0., Schnaitman, C. A., and Morris, H. P. Comparison ofthe Mitochondrial Membrane Proteins in Rat Liver and Hepatomas.CancerRes.,31: 108-113, 1971.
6. Chang, L. 0., Srb, A. M., and Steward, F. C. Electrophoretic Separations of the Soluble Proteins of Neurospora. Nature, 193: 756-759,
1962.7. Criss, W. E. A Review of Isoenzymes in Cancer. Cancer Res., 31:
1523-1542, 1971.8. Davis, B. J. Disc Electrophoresis. II. Method and Application to
Human Serum Proteins. Ann. N. Y. Acad. Sci., 121: 404-427, 1964.9. Haynes, R. C., Jr. The Fixation ofCarbon Dioxide by Rat Liver Mito
chondria and Its Relation to Gluconeogenesis. J. Biol. Chem., 240:4103-4106,1965.
10. Higgins, G. M., and Anderson, R. M. Experimental Pathology of theLiver: Restoration of the Liver of the White Rat following PartialSurgical Removal. Arch. Pathol., 12: 186-202, 1931.
II. Hruban, Z., Morris, H. P., Mochizuki, Y., Meranze, D. R., andSlesers, A. Light Microscopic Observations of Morris Hepatomas.Cancer Res., 31: 752-762, 1971.
12. Keech, D. B., and Utter, M. F. Pyruvate Carboxylase. II. Properties.J. Biol. Chem., 238: 2609-2614, 1963.
13. Lamer, J., and Sanger, F. The Amino Acid Sequence ofthe Phosphorylation Site of Muscle Uridine Diphosphoglucose a-I ,4-Glucan a-4-Glucosyl Transferase. J. Mol. Biol., 11. 491-500, 1965.
14. Layne, E. Spectrophotometric and Turbidimetric Methods for Mea
suring Proteins. Methods Enzymol. 3: 447 454, 1957.15. Lowry, 0. H., Rosebrough, N. J., Farr, A. L., and Randall, R. J. Pro
tein Measurement with the Folin Phenol Reagent. J. BioI. Chem., 193:
265-275,1951.
16. Mancini, G., Vaerman, J. P., Carbonara, A. 0., and Heremans, J. F.A Single Radial Diffusion Method for the Immunological Quantitation of Proteins. In: H. Peters (ed.) Protides of the Biological Fluids.Proceedings of the I Ith Colloqium, Bruges, Belgium, 1963, pp. 370-373. New York: American Elsevier Publishing Co., Inc., 1964.
17. McClure, W. R., and Lardy, H. A. Factors Affecting the Regulationin Vivo.J. Biol.Chem.,246:3591-3596, 1971.
18. McClure, W. R., Lardy, H. A., and Kneifel, H. P. Rat Liver PyruvateCarboxylase. I. Preparation, Properties and Cation Specificity. J.Biol.Chem., 246:3569-3578,1971.
19. Morris, H. P. Studies on the Development, Biochemistry and Biologyof Experimental Hepatomas. Advan. Cancer Res., 9:227-302, 1965.
20. Morris, H. P., and Wagner, B. P. Induction and Transplantation ofRat Hepatomas with Different Growth Rate (Including Minimal Dcviation Hepatomas). In: H. Busch (ed), Methods in Cancer Research,Vol. 4, pp. 125-152. New York: Academic Press, lnc., 1968.
21. Nowell, P. C., and Morris, H. P. Chromosomes of “MinimalDeviation― Hepatomas: A Further Report on Diploid Tumors. Cancer Res.,
29: 969-970, 1969.22. Nowell, P. C., Morris, H. P., and Potter, V. R. Chromosomes of
“MinimalDeviation―Hepatomas. Cancer Res., 27: 1565-1579, 1967.23. Pitot, H. C. The Comparative Enzymology and Cell Origin of Rat
Hepatomas. II. Glutamic Dehydrogenase, Choline Oxidase, and Glucose-6-Phosphatase. Cancer Res., 20: 1262- 1268, 1960.
24. Potter, V. R., Wantanabe, M., Pitot, H. C., and Morris, H. P. Systematic Oscillations in Metabolic Activity in Rat Liver and Hepatomas.Survey of Normal Diploid and Other Hepatoma Lines. Cancer Res.,
29: 55-78, 1969.25. Prinz, W., Schoner, W., Haag, U., and Seubert, W. On the Mecha
nism of Gluconeogenesis and its Regulation. IV. An Isotope Assay
for the Determination of Microquantities of Acetyl CoA. Biochem.z.,346:206-211,1966.
26. Reuber, M. D. Histopathology ofTransplantable Hepatic CarcinomasInduced by Chemical Carcinogens in Rats. Gann Monograph, 1: 43-54, 1966.
27. Schnaitman, C. A., and Greenawalt, J. W. Enzymatic Properties ofthe Inner and Outer Membranes of Rat Liver Mitochondria. J. CellBiol.,38: 158-175,1968.
28. Scrutton, M. C., and Utter, M. F. The Regulation of Glycolysis andGluconeogenesis. Ann. Rev. Biochem., 37: 249-302, 1968.
29. Shonk, C. E., Morris, H. P., and Boxer, G. E. Patterns of GlycolyticEnzymes in Rat Liver and Hepatoma. Cancer Res., 25: 671-676,1965.
30. Sordahl, L. A., and Schwartz, A. Tumor Mitochondria. In: H. Busch(ed), Methods in Cancer Research, Vol. 6, pp. 159-184. New York:Academic Press, Inc., 1971.
31. Stahl, E. (ed.) Thin-layer Chromatography. In: A Laboratory Handbook. pp. 357-358. New York: Academic Press, Inc., 1965.
32. Walter, P., and Anabitarte, M. On the Use of Glutamate Dehydrogenase as a Mitochondrial Marker Enzyme for the Determination ofthe Intracellular Distribution of Rat Liver Pyruvate Carboxylase.Federation European Biochem. Soc. Letters, 12: 289-292, 1971.
33. Weber, G. The Molecular Correlation Concept: Studies on the Metabolic Pattern of Hepatomas. Gann Monograph, 1: 151-178, 1966.
34. Weber, G., and Morris, H. P. Comparative Biochemistry of Hepatomas. III. Carbohydrate Enzymes in Liver Tumors of DifferentGrowth Rates. Cancer Res., 23: 987-994, 1963.
35. Weinhouse, S. Glycolysis, Respiration, and Anomalous Gene Expression in Experimental Hepatomas: G. H. A. Clowes Memorial Lecture.Cancer Res., 32: 2007-2016, 1972.
36. Weinhouse, S., and Ono, T. (eds.) Isoenzymes and Enzyme Regula
2040 CANCER RESEARCH VOL. 33
on March 26, 2018. © 1973 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from

PC in Livers and Liver Tumors
tion in Cancer. Gann Monograph on Cancer Research No. 13.Tokyo:University of Tokyo Press, 1972.
37. Weinhouse, S., Shatton, J. B., Criss, W. E., Farina, F. A., and Morris, H. P. Isozymes in Relation to Differentiation in TransplantableRat Hepatomas. Gann Monograph, 13: I - 18, 1972.
38. Williams, C. A., and Chase, N. W. Methods in Immunology and Im
munochemistry, Vols. I, 2, and 3. New York: Academic Press, Inc.,1967,1968,1971.
39. Work, T. S., and Work, E. (eds.) Laboratory Techniques in Biochemistry and Molecular Biology, Vols. I and 2. Amsterdam: NorthHolland Publishing Co., 1969.
SEPTEMBER 1973 2041
on March 26, 2018. © 1973 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from

1973;33:2034-2041. Cancer Res Lillie O. Chang and Harold P. Morris in Livers and Liver TumorsEnzymatic and Immunological Studies on Pyruvate Carboxylase
Updated version
http://cancerres.aacrjournals.org/content/33/9/2034
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/33/9/2034To request permission to re-use all or part of this article, use this link
on March 26, 2018. © 1973 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from