Embed Size (px)
Citation preview

Epidermal Nerve Fibers Modulate KeratinocyteGrowth via Neuropeptide Signaling in an InnervatedSkin ModelDennis Roggenkamp1, Sarah Kopnick1, Franz Stab1, Horst Wenck1, Martin Schmelz2 and Gitta Neufang1
Atopic eczema is a chronic inflammatory skin disease characterized by cutaneous nerve fiber sprouting andepidermal hyperplasia, pointing to an involvement of the peripheral nervous system in cutaneous homeostasis.However, the interaction of sensory neurons and skin cells is poorly understood. Using an innervated skinmodel, we investigated the influence of sensory neurons on epidermal morphogenesis. Neurons induced theproliferation of keratinocytes, resulting in an increase in the epidermal thickness. Inhibition of calcitonin gene–related peptide (CGRP), but not substance P (SP) signaling, reversed this effect. Human CGRP enhancedkeratinocyte proliferation and epidermal thickness in skin models, demonstrating a key role of CGRP inmodulating epidermal morphogenesis, whereas SP had only a moderate effect. Innervated skin modelscomposed of atopic skin cells showed increased neurite outgrowth, accompanied by elevated CGRP release.As atopic keratinocytes were sensitized to CGRP owing to higher expression levels of the CGRP receptorcomponents, receptor activity–modifying protein 1 (RAMP1) and receptor component protein (RCP), atopicinnervated skin models displayed a thicker epidermis than did healthy controls. We conclude that neuralCGRP controls local keratinocyte growth. Our results show that the crosstalk of the cutaneous peripheralnervous system and skin cells significantly influences epidermal morphogenesis and homeostasis in healthy andatopic skin.
Journal of Investigative Dermatology (2013) 133, 1620–1628; doi:10.1038/jid.2012.464; published online 3 January 2013
INTRODUCTIONThe cutaneous peripheral nervous system has a pivotal role inskin homeostasis and disease. It can be divided in twofunctional units: afferent sensory innervation is responsiblefor the detection of mechanical, thermal, itchy, and painfulstimuli, whereas efferent autonomic innervation controlsglandular activity or vasoregulation (Schulze et al., 1997;Vetrugno et al., 2003; Lumpkin and Caterina, 2007). Sensoryepidermal innervation is provided by long axons originatingfrom cell bodies in the dorsal root ganglia (DRGs), close tothe spinal cord. DRG neurons are pseudounipolar with abifurcated axon, connecting the sensory endings in the skinwith the synapses in the dorsal horn of the spinal cord(Tandrup, 1995).
Cutaneous nerve fibers terminate either freely in theepidermis and dermis or around blood vessels, hair follicles,and sweat glands (Reilly et al., 1997; Hendrix et al., 2008).They are partly uncovered from their Schwann-cell sheaths,forming varicosities, which enable the nonsynaptic release ofneurotransmitters and neuropeptides along the axon (Hilarp,1959). The close contact of peptidergic nerve fibers and skincells, such as dermal fibroblasts, keratinocytes, mast cells, orLangerhans cells, has been suggested as the basis of functionalinteractions (Hosoi et al., 1993; Botchkarev et al., 1997; Peterset al., 2006b). Upon activation, sensory neurons releaseneuropeptides such as calcitonin gene–related peptide (CGRP)and substance P (SP), which subsequently bind their specificreceptors on skin cells and thereby modulate cutaneousinflammation (Black, 2002; Steinhoff et al., 2003; Peterset al., 2006a). Together with calcitonin and amylin, CGRPbelongs to the calcitonin gene peptide superfamily. It consistsof 37 amino acids and exists in two different isoforms: CGRPaand CGRPb. CGRPa is predominantly expressed in sensoryneurons and is one of the most potent endogenousvasodilators (Wimalawansa, 1997; Arulmani et al., 2004).CGRPb was detected in enteric neurons (Mulderry et al.,1988) and was recently shown to be released by keratinocytes(Hou et al., 2011). CGRP binds to a G protein–coupledreceptor composed of the seven transmembrane domainprotein calcitonin receptor–like receptor (CLR) and the receptoractivity–modifying protein 1 (RAMP1) (Walker et al., 2010).
See related commentary on pg 1465ORIGINAL ARTICLE
1Research and Development, Beiersdorf AG, Hamburg, Germany and2Department of Anesthesiology Mannheim, Heidelberg University, Mannheim,Germany
Correspondence: Gitta Neufang, Research and Development, Beiersdorf AG,Hamburg 20253, Germany. E-mail: [email protected]
Received 27 June 2012; revised 25 September 2012; accepted 16 October2012; published online 3 January 2013
Abbreviations: CALCB, calcitonin-related polypeptide-b; CGRP, calcitoningene–related peptide; CLR, calcitonin receptor–like receptor; DRG, dorsal rootganglion; Flg, filaggrin; NK1R, neurokinin 1 receptor; RAMP1, receptoractivity–modifying protein 1; RCP, receptor component protein;SP, substance P
1620 Journal of Investigative Dermatology (2013), Volume 133 & 2013 The Society for Investigative Dermatology
The intracellular receptor component protein (RCP) isessential for CGRP receptor functionality (Prado et al.,2002). The undecapeptide SP and the neurokinins A and Bare members of the tachykinin peptide family (Severini et al.,2002). SP binds to the seven transmembrane domainG protein–coupled neurokinin 1 receptor (NK1R) (Regoliet al., 1987).
Both neuropeptides, CGRP and SP, are involved in neuro-genic inflammation and are said to exacerbate chronicinflammatory skin diseases such as atopic eczema (Peterset al., 2006b). Patients with atopic eczema display anincreased cutaneous nerve fiber density, which might havefunctional consequences for skin homeostasis (Tobin et al.,1992; Ostlere et al., 1995). Some studies suggest aninvolvement of the peripheral nervous system in theepidermal turnover. Denervation of rat skin induces thinningof the epidermis (Li et al., 1997; Hsieh and Lin, 1999).However, the mechanisms of this communication betweenneurons and keratinocytes remain unknown.
Several in vitro models of neurons and keratinocytes havebeen developed that allow investigating functional interac-tions such as neuropeptide release (Pereira et al., 2010) andstructural interactions upon reinnervation of skin explants(Lebonvallet et al., 2012). Although providing a morephysiological environment for the neuronal outgrowth, skinexplants are prone to higher intersubject variability ascompared with in vitro systems. As we have previouslyshown, atopic skin cells mediate increased neuriteoutgrowth of CGRP-positive nerve fibers in a spatiallycompartmented coculture model of porcine DRG neuronsand human skin cells (Roggenkamp et al., 2012). In thismodel, sensory nerve endings interact with monocultivateddermal fibroblasts or keratinocytes. Here, we introduce aninnervated organotypic skin model based on the coculture ofhuman dermal fibroblasts, keratinocytes, and porcine DRGneurons, resulting in an air-exposed skin equivalent withinnervated epidermis. We used the innervated skin model toinvestigate the trophic effects of epidermal innervation andclarify its mechanisms. Using keratinocytes and dermalfibroblasts from patients with atopic eczema, we aimed tomimic atopic skin and investigate potential hyperinnervationof the diseased skin.
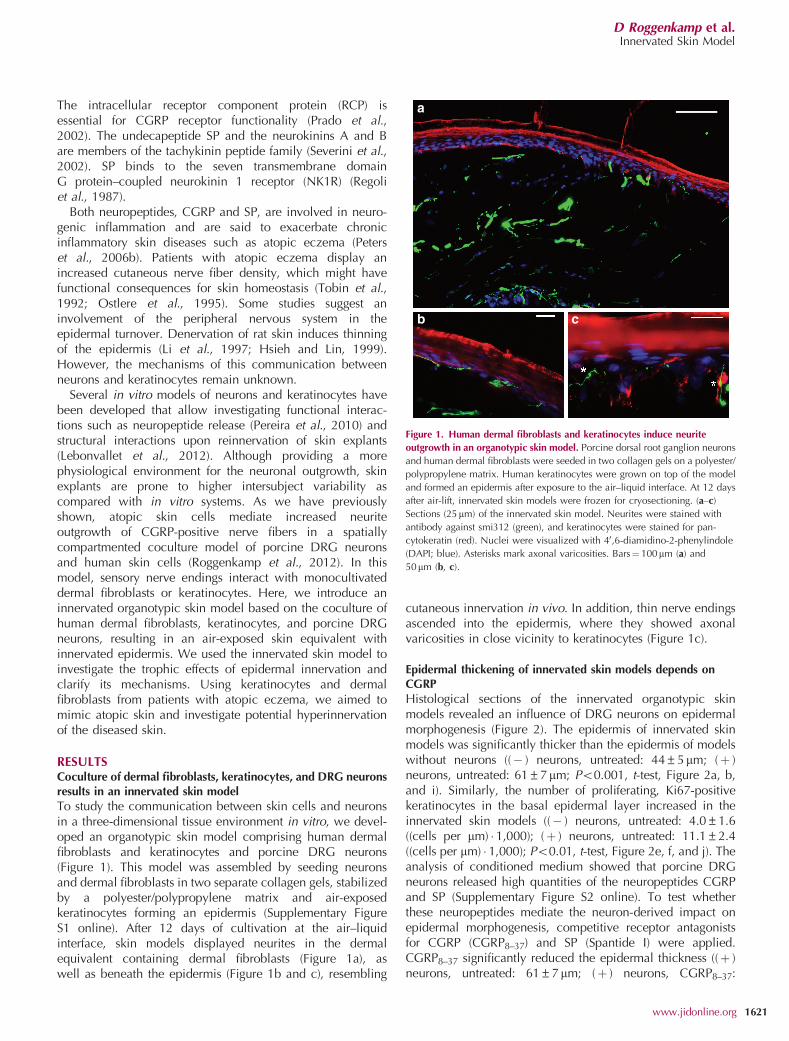
RESULTSCoculture of dermal fibroblasts, keratinocytes, and DRG neuronsresults in an innervated skin modelTo study the communication between skin cells and neuronsin a three-dimensional tissue environment in vitro, we devel-oped an organotypic skin model comprising human dermalfibroblasts and keratinocytes and porcine DRG neurons(Figure 1). This model was assembled by seeding neuronsand dermal fibroblasts in two separate collagen gels, stabilizedby a polyester/polypropylene matrix and air-exposedkeratinocytes forming an epidermis (Supplementary FigureS1 online). After 12 days of cultivation at the air–liquidinterface, skin models displayed neurites in the dermalequivalent containing dermal fibroblasts (Figure 1a), aswell as beneath the epidermis (Figure 1b and c), resembling
cutaneous innervation in vivo. In addition, thin nerve endingsascended into the epidermis, where they showed axonalvaricosities in close vicinity to keratinocytes (Figure 1c).
Epidermal thickening of innervated skin models depends onCGRP
Histological sections of the innervated organotypic skinmodels revealed an influence of DRG neurons on epidermalmorphogenesis (Figure 2). The epidermis of innervated skinmodels was significantly thicker than the epidermis of modelswithout neurons ((� ) neurons, untreated: 44±5mm; (þ )neurons, untreated: 61±7mm; Po0.001, t-test, Figure 2a, b,and i). Similarly, the number of proliferating, Ki67-positivekeratinocytes in the basal epidermal layer increased in theinnervated skin models ((� ) neurons, untreated: 4.0±1.6((cells per mm) � 1,000); (þ ) neurons, untreated: 11.1±2.4((cells per mm) � 1,000); Po0.01, t-test, Figure 2e, f, and j). Theanalysis of conditioned medium showed that porcine DRGneurons released high quantities of the neuropeptides CGRPand SP (Supplementary Figure S2 online). To test whetherthese neuropeptides mediate the neuron-derived impact onepidermal morphogenesis, competitive receptor antagonistsfor CGRP (CGRP8–37) and SP (Spantide I) were applied.CGRP8–37 significantly reduced the epidermal thickness ((þ )neurons, untreated: 61±7mm; (þ ) neurons, CGRP8–37:
a
b c
Figure 1. Human dermal fibroblasts and keratinocytes induce neurite
outgrowth in an organotypic skin model. Porcine dorsal root ganglion neurons
and human dermal fibroblasts were seeded in two collagen gels on a polyester/
polypropylene matrix. Human keratinocytes were grown on top of the model
and formed an epidermis after exposure to the air–liquid interface. At 12 days
after air-lift, innervated skin models were frozen for cryosectioning. (a–c)
Sections (25mm) of the innervated skin model. Neurites were stained with
antibody against smi312 (green), and keratinocytes were stained for pan-
cytokeratin (red). Nuclei were visualized with 40,6-diamidino-2-phenylindole
(DAPI; blue). Asterisks mark axonal varicosities. Bars¼ 100mm (a) and
50mm (b, c).
D Roggenkamp et al.Innervated Skin Model
www.jidonline.org 1621
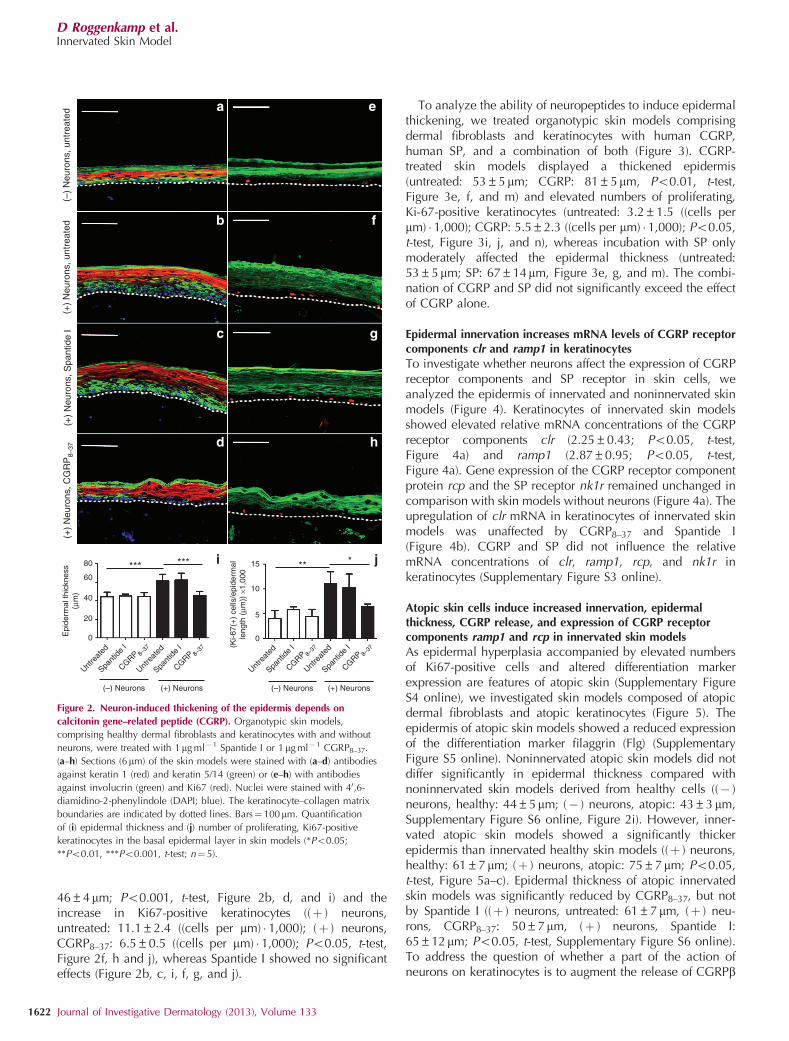
46±4mm; Po0.001, t-test, Figure 2b, d, and i) and theincrease in Ki67-positive keratinocytes ((þ ) neurons,untreated: 11.1±2.4 ((cells per mm) � 1,000); (þ ) neurons,CGRP8–37: 6.5±0.5 ((cells per mm) � 1,000); Po0.05, t-test,Figure 2f, h and j), whereas Spantide I showed no significanteffects (Figure 2b, c, i, f, g, and j).
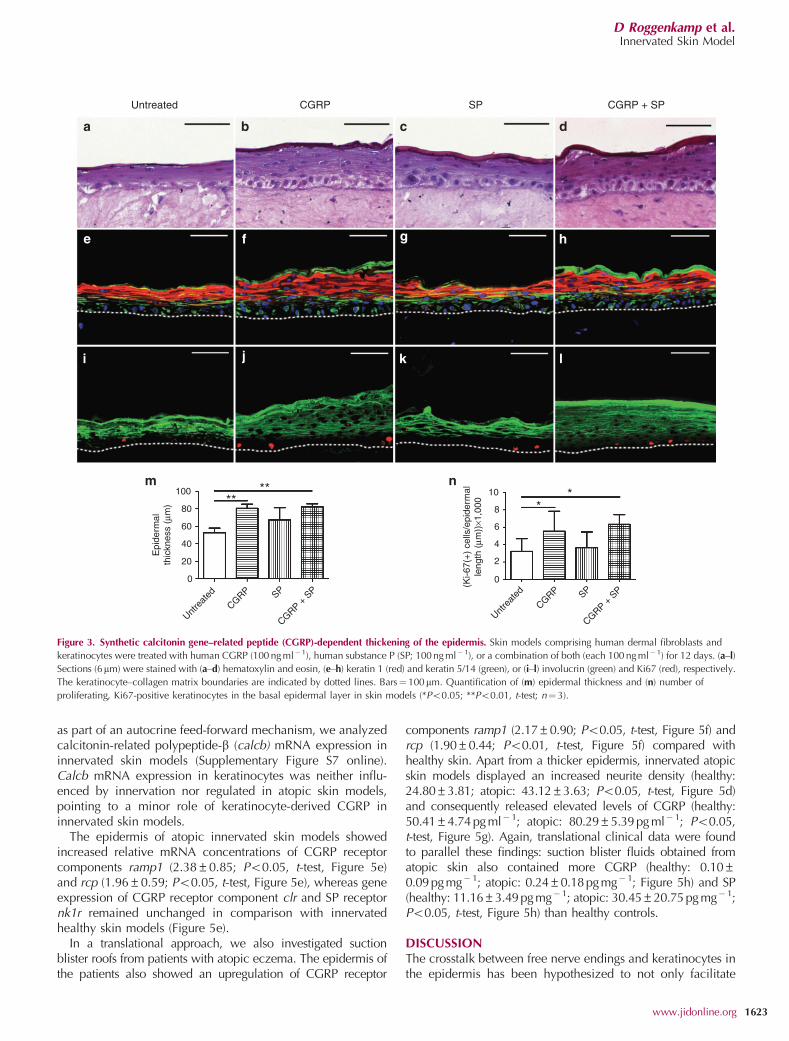
To analyze the ability of neuropeptides to induce epidermalthickening, we treated organotypic skin models comprisingdermal fibroblasts and keratinocytes with human CGRP,human SP, and a combination of both (Figure 3). CGRP-treated skin models displayed a thickened epidermis(untreated: 53±5mm; CGRP: 81±5mm, Po0.01, t-test,Figure 3e, f, and m) and elevated numbers of proliferating,Ki-67-positive keratinocytes (untreated: 3.2±1.5 ((cells permm) � 1,000); CGRP: 5.5±2.3 ((cells per mm) � 1,000); Po0.05,t-test, Figure 3i, j, and n), whereas incubation with SP onlymoderately affected the epidermal thickness (untreated:53±5mm; SP: 67±14mm, Figure 3e, g, and m). The combi-nation of CGRP and SP did not significantly exceed the effectof CGRP alone.
Epidermal innervation increases mRNA levels of CGRP receptorcomponents clr and ramp1 in keratinocytes
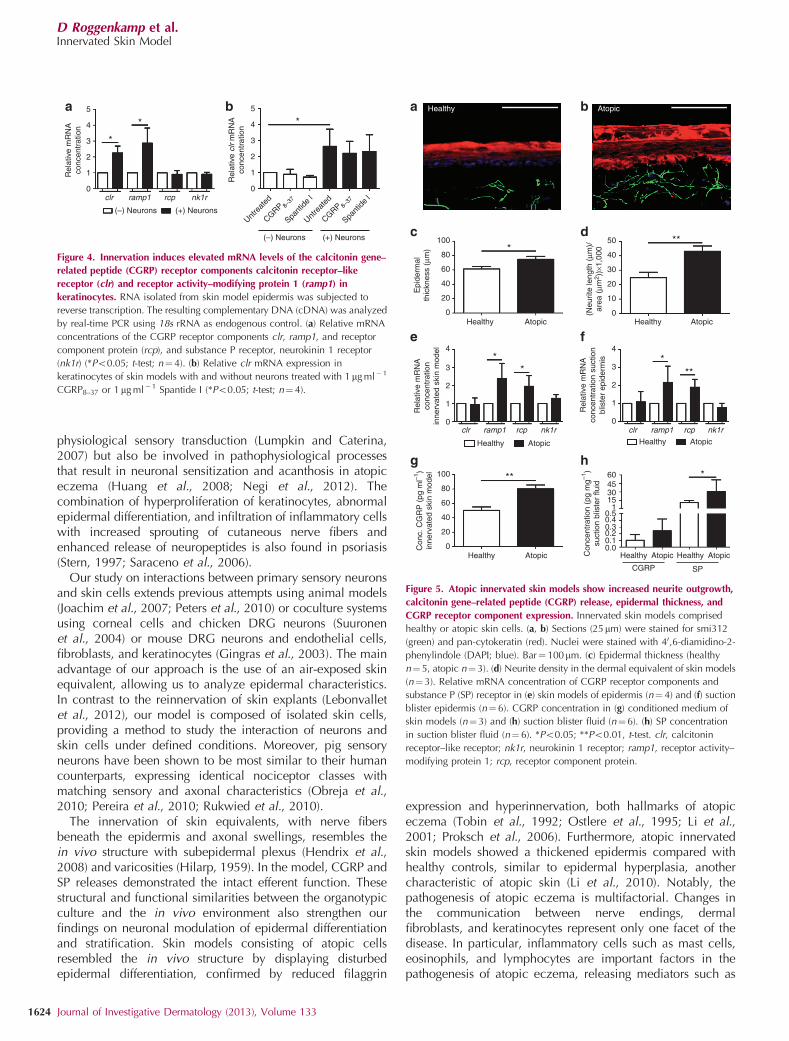
To investigate whether neurons affect the expression of CGRPreceptor components and SP receptor in skin cells, weanalyzed the epidermis of innervated and noninnervated skinmodels (Figure 4). Keratinocytes of innervated skin modelsshowed elevated relative mRNA concentrations of the CGRPreceptor components clr (2.25±0.43; Po0.05, t-test,Figure 4a) and ramp1 (2.87±0.95; Po0.05, t-test,Figure 4a). Gene expression of the CGRP receptor componentprotein rcp and the SP receptor nk1r remained unchanged incomparison with skin models without neurons (Figure 4a). Theupregulation of clr mRNA in keratinocytes of innervated skinmodels was unaffected by CGRP8–37 and Spantide I(Figure 4b). CGRP and SP did not influence the relativemRNA concentrations of clr, ramp1, rcp, and nk1r inkeratinocytes (Supplementary Figure S3 online).
Atopic skin cells induce increased innervation, epidermalthickness, CGRP release, and expression of CGRP receptorcomponents ramp1 and rcp in innervated skin models
As epidermal hyperplasia accompanied by elevated numbersof Ki67-positive cells and altered differentiation markerexpression are features of atopic skin (Supplementary FigureS4 online), we investigated skin models composed of atopicdermal fibroblasts and atopic keratinocytes (Figure 5). Theepidermis of atopic skin models showed a reduced expressionof the differentiation marker filaggrin (Flg) (SupplementaryFigure S5 online). Noninnervated atopic skin models did notdiffer significantly in epidermal thickness compared withnoninnervated skin models derived from healthy cells ((� )neurons, healthy: 44±5mm; (� ) neurons, atopic: 43±3mm,Supplementary Figure S6 online, Figure 2i). However, inner-vated atopic skin models showed a significantly thickerepidermis than innervated healthy skin models ((þ ) neurons,healthy: 61±7mm; (þ ) neurons, atopic: 75±7mm; Po0.05,t-test, Figure 5a–c). Epidermal thickness of atopic innervatedskin models was significantly reduced by CGRP8–37, but notby Spantide I ((þ ) neurons, untreated: 61±7mm, (þ ) neu-rons, CGRP8–37: 50±7mm, (þ ) neurons, Spantide I:65±12mm; Po0.05, t-test, Supplementary Figure S6 online).To address the question of whether a part of the action ofneurons on keratinocytes is to augment the release of CGRPb
(–)
Neu
rons
, unt
reat
ed(+
) N
euro
ns, u
ntre
ated
(+)
Neu
rons
, Spa
ntid
e I
(+)
Neu
rons
, CG
RP
8–37
(–) Neurons (+) Neurons (–) Neurons (+) Neurons
Untre
ated
Spant
ide I
CGRP 8–37
Untre
ated
Spant
ide I
CGRP 8–37
Untre
ated
Spant
ide I
CGRP 8–37
Untre
ated
Spant
ide I
CGRP 8–37
Epi
derm
al th
ickn
ess
(µm
)
60
40
0
20
80
(Ki-6
7(+
) ce
lls/e
pide
rmal
leng
th (
µm))
×1,
000
15
10
5
0
*** *** ***
a e
b f
c g
d
i j
h
Figure 2. Neuron-induced thickening of the epidermis depends on
calcitonin gene–related peptide (CGRP). Organotypic skin models,
comprising healthy dermal fibroblasts and keratinocytes with and without
neurons, were treated with 1mg ml�1 Spantide I or 1mg ml� 1 CGRP8–37.
(a–h) Sections (6mm) of the skin models were stained with (a–d) antibodies
against keratin 1 (red) and keratin 5/14 (green) or (e–h) with antibodies
against involucrin (green) and Ki67 (red). Nuclei were stained with 40,6-
diamidino-2-phenylindole (DAPI; blue). The keratinocyte–collagen matrix
boundaries are indicated by dotted lines. Bars¼100mm. Quantification
of (i) epidermal thickness and (j) number of proliferating, Ki67-positive
keratinocytes in the basal epidermal layer in skin models (*Po0.05;
**Po0.01, ***Po0.001, t-test; n¼ 5).
D Roggenkamp et al.Innervated Skin Model
1622 Journal of Investigative Dermatology (2013), Volume 133
as part of an autocrine feed-forward mechanism, we analyzedcalcitonin-related polypeptide-b (calcb) mRNA expression ininnervated skin models (Supplementary Figure S7 online).Calcb mRNA expression in keratinocytes was neither influ-enced by innervation nor regulated in atopic skin models,pointing to a minor role of keratinocyte-derived CGRP ininnervated skin models.
The epidermis of atopic innervated skin models showedincreased relative mRNA concentrations of CGRP receptorcomponents ramp1 (2.38±0.85; Po0.05, t-test, Figure 5e)and rcp (1.96±0.59; Po0.05, t-test, Figure 5e), whereas geneexpression of CGRP receptor component clr and SP receptornk1r remained unchanged in comparison with innervatedhealthy skin models (Figure 5e).
In a translational approach, we also investigated suctionblister roofs from patients with atopic eczema. The epidermis ofthe patients also showed an upregulation of CGRP receptor
components ramp1 (2.17±0.90; Po0.05, t-test, Figure 5f) andrcp (1.90±0.44; Po0.01, t-test, Figure 5f) compared withhealthy skin. Apart from a thicker epidermis, innervated atopicskin models displayed an increased neurite density (healthy:24.80±3.81; atopic: 43.12±3.63; Po0.05, t-test, Figure 5d)and consequently released elevated levels of CGRP (healthy:50.41±4.74 pg ml�1; atopic: 80.29±5.39 pg ml� 1; Po0.05,t-test, Figure 5g). Again, translational clinical data were foundto parallel these findings: suction blister fluids obtained fromatopic skin also contained more CGRP (healthy: 0.10±0.09 pg mg�1; atopic: 0.24±0.18 pg mg� 1; Figure 5h) and SP(healthy: 11.16±3.49 pg mg� 1; atopic: 30.45±20.75 pg mg�1;Po0.05, t-test, Figure 5h) than healthy controls.
DISCUSSIONThe crosstalk between free nerve endings and keratinocytes inthe epidermis has been hypothesized to not only facilitate
Untreated CGRP SP CGRP + SP
** ****
100
80
60
40
20
0
10
8
6
4
2
0
Untre
ated
CGRP
CGRP + S
PSP
Untre
ated
CGRP
CGRP + S
PSP
Epi
derm
alth
ickn
ess
(µm
)
(Ki-6
7(+
) ce
lls/e
pide
rmal
leng
th (
µm))
×1,0
00
Figure 3. Synthetic calcitonin gene–related peptide (CGRP)-dependent thickening of the epidermis. Skin models comprising human dermal fibroblasts and
keratinocytes were treated with human CGRP (100 ng ml�1), human substance P (SP; 100 ng ml� 1), or a combination of both (each 100 ng ml�1) for 12 days. (a–l)
Sections (6mm) were stained with (a–d) hematoxylin and eosin, (e–h) keratin 1 (red) and keratin 5/14 (green), or (i–l) involucrin (green) and Ki67 (red), respectively.
The keratinocyte–collagen matrix boundaries are indicated by dotted lines. Bars¼100mm. Quantification of (m) epidermal thickness and (n) number of
proliferating, Ki67-positive keratinocytes in the basal epidermal layer in skin models (*Po0.05; **Po0.01, t-test; n¼3).
D Roggenkamp et al.Innervated Skin Model
www.jidonline.org 1623
physiological sensory transduction (Lumpkin and Caterina,2007) but also be involved in pathophysiological processesthat result in neuronal sensitization and acanthosis in atopiceczema (Huang et al., 2008; Negi et al., 2012). Thecombination of hyperproliferation of keratinocytes, abnormalepidermal differentiation, and infiltration of inflammatory cellswith increased sprouting of cutaneous nerve fibers andenhanced release of neuropeptides is also found in psoriasis(Stern, 1997; Saraceno et al., 2006).
Our study on interactions between primary sensory neuronsand skin cells extends previous attempts using animal models(Joachim et al., 2007; Peters et al., 2010) or coculture systemsusing corneal cells and chicken DRG neurons (Suuronenet al., 2004) or mouse DRG neurons and endothelial cells,fibroblasts, and keratinocytes (Gingras et al., 2003). The mainadvantage of our approach is the use of an air-exposed skinequivalent, allowing us to analyze epidermal characteristics.In contrast to the reinnervation of skin explants (Lebonvalletet al., 2012), our model is composed of isolated skin cells,providing a method to study the interaction of neurons andskin cells under defined conditions. Moreover, pig sensoryneurons have been shown to be most similar to their humancounterparts, expressing identical nociceptor classes withmatching sensory and axonal characteristics (Obreja et al.,2010; Pereira et al., 2010; Rukwied et al., 2010).
The innervation of skin equivalents, with nerve fibersbeneath the epidermis and axonal swellings, resembles thein vivo structure with subepidermal plexus (Hendrix et al.,2008) and varicosities (Hilarp, 1959). In the model, CGRP andSP releases demonstrated the intact efferent function. Thesestructural and functional similarities between the organotypicculture and the in vivo environment also strengthen ourfindings on neuronal modulation of epidermal differentiationand stratification. Skin models consisting of atopic cellsresembled the in vivo structure by displaying disturbedepidermal differentiation, confirmed by reduced filaggrin
expression and hyperinnervation, both hallmarks of atopiceczema (Tobin et al., 1992; Ostlere et al., 1995; Li et al.,2001; Proksch et al., 2006). Furthermore, atopic innervatedskin models showed a thickened epidermis compared withhealthy controls, similar to epidermal hyperplasia, anothercharacteristic of atopic skin (Li et al., 2010). Notably, thepathogenesis of atopic eczema is multifactorial. Changes inthe communication between nerve endings, dermalfibroblasts, and keratinocytes represent only one facet of thedisease. In particular, inflammatory cells such as mast cells,eosinophils, and lymphocytes are important factors in thepathogenesis of atopic eczema, releasing mediators such as
5a b4
3
2
1
0
*
*
Rel
ativ
e m
RN
Aco
ncen
trat
ion
Rel
ativ
e cl
r m
RN
Aco
ncen
trat
ion
clr ramp1 rcp nk1r
(–) Neurons (+) Neurons
5
4
3
2
1
0
*
Untre
ated
CGRP 8–37
Spant
ide l
Untre
ated
Spant
ide l
CGRP 8–37
(–) Neurons (+) Neurons
Figure 4. Innervation induces elevated mRNA levels of the calcitonin gene–
related peptide (CGRP) receptor components calcitonin receptor–like
receptor (clr) and receptor activity–modifying protein 1 (ramp1) in
keratinocytes. RNA isolated from skin model epidermis was subjected to
reverse transcription. The resulting complementary DNA (cDNA) was analyzed
by real-time PCR using 18s rRNA as endogenous control. (a) Relative mRNA
concentrations of the CGRP receptor components clr, ramp1, and receptor
component protein (rcp), and substance P receptor, neurokinin 1 receptor
(nk1r) (*Po0.05; t-test; n¼4). (b) Relative clr mRNA expression in
keratinocytes of skin models with and without neurons treated with 1mg ml�1
CGRP8–37 or 1mg ml�1 Spantide I (*Po0.05; t-test; n¼ 4).
Healthy Atopic
100
80
60
40
20
0
Epi
derm
alth
ickn
ess
(µm
) *
a b
c d
e f
g h
50
40
30
20
10
0(Neu
rite
leng
th (
µm)/
area
(µm
2 ))×
1,00
0
**
4
3
2
1
0
**
Rel
ativ
e m
RN
Aco
ncen
trat
ion
inne
rvat
ed s
kin
mod
el
Rel
ativ
e m
RN
Aco
ncen
trat
ion
suct
ion
blis
ter
epid
erm
is
clr ramp1 rcp nk1r clr ramp1 rcp nk1r
4
3
2
1
0
***
100
80
60
40
20
0
** *604530151
0.50.40.30.20.10.0
Con
cent
ratio
n (p
g m
g–1)
suct
ion
blis
ter
fluid
Con
c. C
GR
P (
pg m
l–1)
inne
rvat
ed s
kin
mod
el
CGRP SP
AtopicHealthy
Atopic Healthy AtopicHealthyHealthy Atopic
Healthy Atopic
Healthy Atopic Healthy Atopic
Figure 5. Atopic innervated skin models show increased neurite outgrowth,
calcitonin gene–related peptide (CGRP) release, epidermal thickness, and
CGRP receptor component expression. Innervated skin models comprised
healthy or atopic skin cells. (a, b) Sections (25mm) were stained for smi312
(green) and pan-cytokeratin (red). Nuclei were stained with 40,6-diamidino-2-
phenylindole (DAPI; blue). Bar¼ 100mm. (c) Epidermal thickness (healthy
n¼ 5, atopic n¼3). (d) Neurite density in the dermal equivalent of skin models
(n¼ 3). Relative mRNA concentration of CGRP receptor components and
substance P (SP) receptor in (e) skin models of epidermis (n¼ 4) and (f) suction
blister epidermis (n¼6). CGRP concentration in (g) conditioned medium of
skin models (n¼ 3) and (h) suction blister fluid (n¼6). (h) SP concentration
in suction blister fluid (n¼ 6). *Po0.05; **Po0.01, t-test. clr, calcitonin
receptor–like receptor; nk1r, neurokinin 1 receptor; ramp1, receptor activity–
modifying protein 1; rcp, receptor component protein.
D Roggenkamp et al.Innervated Skin Model
1624 Journal of Investigative Dermatology (2013), Volume 133

histamine or IL-31 (Stander and Luger, 2010). IL-31 wasshown to have a role in itch induction (Dillon et al., 2004;Grimstad et al., 2009), but also to elicit a differentiation defectin organotypic skin models, associated with a repression offilaggrin (Cornelissen et al., 2012), which was also observed inatopic skin models. Our innervated skin model offers theopportunity to integrate immune cells, and thereby enablesthe investigation of the cutaneous neuroimmune crosstalk infuture studies.
In our model, the integration of neurons induced a thicken-ing of the epidermis, which was even more pronounced inmodels consisting of atopic skin cells. As paracrine interactionby soluble mediators contributes to epidermal integrity andstructural development (Maas-Szabowski et al., 2000), theimpact of innervation on the epidermis is most probablybased on neuron-derived secreted factors. Moreover, a directinteraction of neurites and skin cells is conceivable, as cell–cell contacts between keratinocytes and neurons wereobserved in our model and also reported by others (Chateauand Misery, 2004; Suuronen et al., 2004; Chateau et al., 2007).
Apart from epidermal hyperplasia, atopic innervated skinmodels showed increased neurite outgrowth, presumablyaffecting epidermal morphogenesis. In vivo, especially CGRP-and SP-positive nerve fibers are much more abundant inlesional atopic skin (Jarvikallio et al., 2003), indicating animportant role of these neuropeptides in the pathogenesis ofthe skin disease. CGRP and SP enhanced proliferation ofkeratinocytes in organotypic skin models, pointing to acorrelation between epidermal hyperinnervation andhyperplasia. So far, the influence of neuropeptides onkeratinocyte proliferation was only studied in monoculturesand mouse models. CGRP and SP increase the proliferation ofHaCaT keratinocytes and murine Pam212 keratinocytesin vitro (Tanaka et al., 1988; Yu et al., 2009). In mice,neurogenic inflammation induces epidermal thickening in anSP-dependent manner (Pavlovic et al., 2008). Furthermore,intracutaneous injection of CGRP in mouse skin increases thenumber of BrdU-labeled basal keratinocytes and epidermalthickness (Seike et al., 2002).
As a result of the higher innervation density, elevated CGRPlevels were detected in atopic skin models. Recently, weshowed that atopic keratinocytes induce increased outgrowthof CGRP-positive nerve fibers in a two-dimensional coculturemodel (Roggenkamp et al., 2012). Here, atopic skin cellsexerted their effect on neurite outgrowth in a three-dimensional tissue-like environment and induced elevatedlevels of CGRP in atopic innervated skin models displayingan impact on epidermal morphogenesis.
In accordance with the atopic innervated skin model, wefound elevated levels of CGRP and SP in suction blister fluidsderived from atopic eczema patients, demonstrating thein vivo relevant simulation of neuron–skin cell interactionsin these models. Higher concentrations of CGRP and SP werealso detected in the plasma of patients with atopic eczema(Salomon and Baran, 2008) or in the inflamed skin of NC/Ngamice (Katsuno et al., 2003). An elevated level of CGRP wasshown to stimulate neuronal expression of proteins involved inperipheral sensitization (Cady et al., 2011). Increased
concentrations of CGRP in the skin might thereforecontribute to the induction of itching in atopic dermatitispatients. Because of their effects on epidermal morphogenesis,we assumed that elevated CGRP levels induced epidermalhyperplasia in atopic innervated skin models. Consistently,the application of a competitive CGRP receptor antagonistreduced epidermal thickness to control the levels ofnoninnervated skin models, whereas an SP receptorantagonist had only a moderate effect. Our results suggestthat primarily CGRP, and to a lesser extent DRG-neuron-derived SP, influenced epidermal thickness and proliferationof keratinocytes in innervated skin models. Recently, it wasdemonstrated that keratinocytes release CGRPb (Hou et al.,2011), which might contribute to epidermal thickening.However, in our model, neurons did not increase calcbmRNA expression in innervated keratinocytes. Thus, ourresults suggest a direct effect of neuron-derived CGRP onkeratinocyte proliferation.
The strong effect of CGRP on keratinocytes was mediatedby the upregulation of CGRP receptor components clr, ramp1,and rcp mRNA in healthy and atopic innervated skin models.However, the expression of SP receptor was unaffected byinnervation. As the cellular concentrations of RAMP1 and RCPcorrelate with CGRP responsiveness (McLatchie et al., 1998;Zhang et al., 2007), sensitized CGRP signaling and increasedavailability might synergistically lead to hyperproliferation ofatopic keratinocytes. This assumption is also corroborated bythe translational results from suction blister roof and fluid fromatopic dermatitis patients, confirming increased CGRP contentand upregulation of CGRP receptor components.
The upregulation of CGRP receptor components was notinhibited by CGRP/SP receptor antagonists, and both neuro-peptides failed to regulate the expression of their receptors inkeratinocytes. Thus, the impact of neurons on the expressionof CGRP receptor components in keratinocytes might be dueto other neuron-derived mediators. Apart from CGRP- and SP-positive nerve fibers, the skin also contains, e.g., somatostatin-and neuropeptide Y–expressing sensory fibers (Gibbins et al.,1987; Tobin et al., 1992). As keratinocytes express a multitudeof receptors, specific for neuron-derived mediators such aspituitary adenylate cyclase–activating polypeptide (PACAP),vasoactive intestinal peptide (VIP), acetylcholine, or adreno-medullin (Grando et al., 1995; Albertin et al., 2003; Pozo,2003), a further characterization of their effects onkeratinocyte biology is warranted.
This study introduced the innervated skin model as anexcellent tool to study the interaction of sensory neurons andhuman skin cells in a three-dimensional tissue-like in vitrosystem. Neuron-derived CGRP induced a thickening of theepidermis, which was even more pronounced in atopic skinmodels because of increased neurite outgrowth, resulting inelevated CGRP levels. Moreover, atopic keratinocytes weresensitized to CGRP signaling as they expressed increasedconcentrations of CGRP receptor components RAMP1 andRCP. These in vitro findings were confirmed in clinicalspecimens from eczematous skin of atopic dermatitis patients.Our models extend the options to study the crosstalk betweenprimary sensory neurons and skin cells in healthy and atopic
D Roggenkamp et al.Innervated Skin Model
www.jidonline.org 1625

skin. It will be of major interest to verify the clinical relevanceof our findings in future studies.
MATERIALS AND METHODSCell culture
Dermal fibroblasts and keratinocytes were isolated from punch
biopsies obtained from similar sites on adult inner upper arm skin
as described before (Roggenkamp et al., 2012). Punch biopsies from
atopic dermatitis patients were obtained from perilesional sites of
eczema. Atopic dermatitis patients displayed an intensity of local
SCORAD (SCORing Atopic Dermatitis) of 6.36±0.99 on the test area
arm. All volunteers provided written, informed consent. Patients and
volunteers of the control group did not differ in their age (control:
37.45±17.08; atopic: 35.56±10.32 years). In a second clinical
study, the epidermis of atopic dermatitis patients and healthy
controls was obtained using the suction blister technique (Kiistala,
1968). Suction blister roofs were obtained from the same area of
interest and subjected to RNA isolation. Suction blister fluids were
collected for quantification of neuropeptides CGRP and SP. Local
SCORAD of atopic dermatitis patients and ages of patients and
controls were equivalent. The clinical studies were conducted
according to the Declaration of Helsinki Principles and approved
by the ethical committees of the Medical Association, Kiel and
Freiburg, Germany.
DRG neurons were isolated from the spines of 3–5-day-old pigs
(Sus scrofa domestica) as described before (Roggenkamp et al., 2012).
Organotypic skin model
For the dermal equivalents of the organotypic skin models, a gel
containing rat tail collagen type I (Sigma-Aldrich, Munich, Germany)
was prepared (3 mg ml� 1 in 0.1% acetic acid). Dermal fibroblasts
were growth-arrested with 2mg ml� 1 mitomycin C (Sigma-Aldrich)
for 4 hours and then seeded in the collagen gel (100,000 cells per ml).
A volume of 600ml of this suspension was transferred to a cell culture
insert (pore size 3mm, 12-well plate; Falcon, Grafelfing-Lochham,
Germany). Dermal equivalents were cultivated in rFAD medium,
DMEM, and Ham’s F12 medium (3:1) supplemented with 1%
penicillin/streptomycin, 10% fetal bovine serum, 50mg ml� 1 ascorbic
acid (Sigma-Aldrich), and 0.4mg ml� 1 hydrocortisone (Sigma-Aldrich)
at 37 1C and 7% CO2. After 24 hours, keratinocytes were suspended
in rFAD and seeded on the gel (250,000 per model). After 48 hours,
keratinocytes were air-exposed and the skin models were supplied
with rFAD medium supplemented with 100 ng ml� 1 human CGRP
(Sigma-Aldrich) or 100 ng ml� 1 human SP (Sigma-Aldrich). The
medium was changed every day. At 12 days after air exposure, the
skin models were frozen in Tissue-Tek (Leica, Wetzlar, Germany) for
further analysis.
Innervated organotypic skin model
The scaffold of the innervated skin model was a polyester/polypro-
pylene matrix (FibraCel Disk, Bibby Sterilin, Staffordshire, UK) glued
with silicone grease in a cell culture insert (pore size 3mm, 12-well
plate; Falcon). As described above, a collagen gel was prepared.
Isolated DRG neurons were seeded in the gel (180,000 cells per
model) and 30ml was transferred on the matrix. Next, 350ml of a
collagen gel comprising dermal fibroblasts (100,000 cells per ml) was
applied on top of the matrix. In all, 250,000 keratinocytes were seeded
on the collagen gels 24 hours later. After 48 hours, keratinocytes were
air-exposed, and the innervated skin models were cultivated in a 1:1
mixture of supplemented Ham’s F12 medium (except nerve growth
factor, compare cultivation of DRG neurons) and rFAD medium at
37 1C and 5% CO2. The medium was additionally supplemented with
1mg ml� 1 human CGRP8–37 (Sigma-Aldrich) or 1mg ml� 1 human
Spantide I (Sigma-Aldrich) and changed every day. After a cultivation
period of 12 days, innervated skin models were frozen in Tissue-Tek
(Leica). Alternatively, the epidermis was removed from innervated
skin models and subjected to RNA isolation.
Immunohistochemistry
Cryosections (6 or 25mm) were fixed with acetone for 15 minutes,
and nonspecific binding was blocked with 1% BSA (Sigma-Aldrich) in
phosphate-buffered saline (PAA, Linz, Austria) for 1 hour at room
temperature. Sections were either stained with hematoxylin and eosin
solution (Sigma-Aldrich) or incubated with primary antibodies. The
6-mm sections were incubated with primary antibodies directed either
against keratin 1 (PRB-149B, Covance, Munich, Germany) and
keratin 5/14 (X1255M, Exalpha Biologicals, Shirley, IN) or involucrin
(I9018, Sigma-Aldrich) and Ki-67 (ab16667, Abcam, Cambridge, UK).
Furthermore, a mouse mAb antibody against filaggrin (15C10; Leica,
Newcastle, UK) was used. The 25-mm sections were incubated with
primary antibodies against pan-cytokeratin (ab9377; Abcam) and
pan-axonal neurofilament marker smi312 (Covance). The sections
were stained with secondary antibodies (Invitrogen, Darmstadt,
Germany), mixed with 40,6-diamidino-2-phenylindole (Sigma-
Aldrich) for 2 hours at room temperature. Immunofluorescence was
visualized with Axiovert S100 (Zeiss, Gottingen, Germany) applying
the AxioVision 4.8 software (Zeiss). Epidermal thickness was quanti-
fied as the median distance between the basal cell layer and the
stratum corneum (ImageJ, NIH, Bethesda, MD). Proliferating Ki-67-
positive keratinocytes in the basal epidermal layer were counted and
normalized to section length measured by using the ImageJ software
(NIH). For each condition, 10 sections were analyzed.
Real-time quantitative reverse transcription–PCR
RNA was isolated from keratinocytes, skin model epidermis, or
human epidermis by using the RNeasy Kit (Qiagen, Hilden,
Germany), and 1mg was reverse transcribed with the High Capacity
complementary DNA Reverse Transcription Kit (Applied Biosystems,
Darmstadt, Germany) according to the manufacturer’s instructions.
Expression levels were detected by real-time reverse transcription–
PCR using gene expression assays for calca (Hs01100741), calcb
(Hs00265194), clr (Hs00173787_m1), flg (Hs00856927), flg2
(Hs00418578), nk1r (Hs00185530_m1), ramp1 (Hs00195288_m1),
and rcp (Hs00205544_m1), performed in a 7900HT Fast Real-Time
PCR System using the TaqMan Master Mix Reagents Kit (all from
Applied Biosystems). All assays were run in duplicate. Gene expres-
sion was quantified by comparative DDCt method, normalizing each
target gene expression to endogenous control 18s rRNA levels.
Normalized gene expression levels were compared and indicated
as relative target mRNA concentrations.
CGRP/SP ELISA
The protein concentrations of CGRP and SP in the supernatants of
mono-cultivated DRG neurons and innervated skin models, as well as
in suction blister fluids of healthy and atopic volunteers, were
measured by using the human CGRP Enzyme ImmunoAssay Kit
D Roggenkamp et al.Innervated Skin Model
1626 Journal of Investigative Dermatology (2013), Volume 133

(SPI-Bio, Montigny, France) and human Substance P Assay (R&D
Systems, Minneapolis, MN), respectively, according to the manufac-
turer’s instructions. Protein concentrations in suction blister fluids
were normalized to total protein concentrations determined by the
BCA Protein Quantification Kit Uptima (Interchim, Montlucon,
France). Absorption was measured using the Spectra Max 250
(Molecular Devices, Ismaning, Germany).
Statistical analysis
For description of the data, the arithmetic mean and the SD
were represented graphically. Data were analyzed with Graph-
Pad Prism 5 (San Diego, CA). Student’s t-test was used, which was
based on a two-sided hypothesis testing with a significance
level of 0.05.
CONFLICT OF INTERESTThe authors DR, SK, FS, HW, and GN are employees of Beiersdorf AG.
ACKNOWLEDGMENTSWe thank Elmar Forsch and Anne-Christin Worthmann for excellent technicalassistance.
SUPPLEMENTARY MATERIAL
Supplementary material is linked to the online version of the paper at http://www.nature.com/jid
REFERENCES
Albertin G, Carraro G, Parnigotto PP et al. (2003) Human skin keratinocytesand fibroblasts express adrenomedullin and its receptors, and adrenome-dullin enhances their growth in vitro by stimulating proliferation andinhibiting apoptosis. Int J Mol Med 11:635–9
Arulmani U, Maassenvandenbrink A, Villalon CM et al. (2004) Calcitoningene-related peptide and its role in migraine pathophysiology. Eur JPharmacol 500:315–30
Black PH (2002) Stress and the inflammatory response: a review of neurogenicinflammation. Brain Behav Immun 16:622–53
Botchkarev VA, Eichmuller S, Peters EM et al. (1997) A simple immunofluor-escence technique for simultaneous visualization of mast cells and nervefibers reveals selectivity and hair cycle–dependent changes in mast cell–nerve fiber contacts in murine skin. Arch Dermatol Res 289:292–302
Cady RJ, Glenn JR, Smith KM et al. (2011) Calcitonin gene-related peptidepromotes cellular changes in trigeminal neurons and glia implicated inperipheral and central sensitization. Mol Pain 7:94
Chateau Y, Dorange G, Clement JF et al. (2007) In vitro reconstruction ofneuro-epidermal connections. J Invest Dermatol 127:979–81
Chateau Y, Misery L (2004) Connections between nerve endings and epidermalcells: are they synapses? Exp Dermatol 13:2–4
Cornelissen C, Marquardt Y, Czaja K et al. (2012) IL-31 regulates differentiationand filaggrin expression in human organotypic skin models. J Allergy ClinImmunol 129:426–33. 433 e421–428
Dillon SR, Sprecher C, Hammond A et al. (2004) Interleukin 31, a cytokineproduced by activated T cells, induces dermatitis in mice. Nat Immunol5:752–60
Gibbins IL, Wattchow D, Coventry B (1987) Two immunohistochemicallyidentified populations of calcitonin gene-related peptide (CGRP)-immu-noreactive axons in human skin. Brain Res 414:143–8
Gingras M, Bergeron J, Dery J et al. (2003) In vitro development of a tissue-engineered model of peripheral nerve regeneration to study neuritegrowth. FASEB J 17:2124–6
Grando SA, Zelickson BD, Kist DA et al. (1995) Keratinocyte muscarinicacetylcholine receptors: immunolocalization and partial characterization.J Invest Dermatol 104:95–100
Grimstad O, Sawanobori Y, Vestergaard C et al. (2009) Anti-interleukin-31-antibodies ameliorate scratching behaviour in NC/Nga mice: a model ofatopic dermatitis. Exp Dermatol 18:35–43
Hendrix S, Picker B, Liezmann C et al. (2008) Skin and hair follicle innervationin experimental models: a guide for the exact and reproducible evaluationof neuronal plasticity. Exp Dermatol 17:214–27
Hilarp NA (1959) The construction and functional organization of theautonomic innervation apparatus. Acta Physiol Scand Suppl 46:1–21
Hosoi J, Murphy GF, Egan CL et al. (1993) Regulation of Langerhans cellfunction by nerves containing calcitonin gene-related peptide. Nature363:159–63
Hou Q, Barr T, Gee L et al. (2011) Keratinocyte expression of calcitonin gene-related peptide beta: implications for neuropathic and inflammatory painmechanisms. Pain 152:2036–51
Hsieh ST, Lin WM (1999) Modulation of keratinocyte proliferation by skininnervation. J Invest Dermatol 113:579–86
Huang SM, Lee H, Chung MK et al. (2008) Overexpressed transient receptorpotential vanilloid 3 ion channels in skin keratinocytes modulate painsensitivity via prostaglandin E2. J Neurosci 28:13727–37
Jarvikallio A, Harvima IT, Naukkarinen A (2003) Mast cells, nerves andneuropeptides in atopic dermatitis and nummular eczema. Arch DermatolRes 295:2–7
Joachim RA, Kuhlmei A, Dinh QT et al. (2007) Neuronal plasticity of the‘‘brain-skin connection’’: stress-triggered up-regulation of neuropeptidesin dorsal root ganglia and skin via nerve growth factor-dependentpathways. J Mol Med 85:1369–78
Katsuno M, Aihara M, Kojima M et al. (2003) Neuropeptides concentrations inthe skin of a murine (NC/Nga mice) model of atopic dermatitis.J Dermatol Sci 33:55–65
Kiistala U (1968) Suction blister device for separation of viable epidermis fromdermis. J Invest Dermatol 50:129–37
Lebonvallet N, Boulais N, Le Gall C et al. (2012) Effects of the re-innervation oforganotypic skin explants on the epidermis. Exp Dermatol 21:156–8
Li C, Lasse S, Lee P et al. (2010) Development of atopic dermatitis-like skindisease from the chronic loss of epidermal caspase-8. Proc Natl Acad SciUSA 107:22249–54
Li XM, Kleiner G, Huang CK et al. (2001) Murine model of atopic dermatitisassociated with food hypersensitivity. J Allergy Clin Immunol 107:693–702
Li Y, Hsieh ST, Chien HF et al. (1997) Sensory and motor denervation influenceepidermal thickness in rat foot glabrous skin. Exp Neurol 147:452–62
Lumpkin EA, Caterina MJ (2007) Mechanisms of sensory transduction in theskin. Nature 445:858–65
Maas-Szabowski N, Stark HJ, Fusenig NE (2000) Keratinocyte growth regula-tion in defined organotypic cultures through IL-1-induced keratinocytegrowth factor expression in resting fibroblasts. J Invest Dermatol 114:1075–1084
McLatchie LM, Fraser NJ, Main MJ et al. (1998) RAMPs regulate the transportand ligand specificity of the calcitonin-receptor-like receptor. Nature393:333–9
Mulderry PK, Ghatei MA, Spokes RA et al. (1988) Differential expression ofalpha-CGRP and beta-CGRP by primary sensory neurons and entericautonomic neurons of the rat. Neuroscience 25:195–205
Negi O, Tominaga M, Tengara S et al. (2012) Topically applied semaphorin 3Aointment inhibits scratching behavior and improves skin inflammation inNC/Nga mice with atopic dermatitis. J Dermatol Sci 66:37–43
Obreja O, Ringkamp M, Namer B et al. (2010) Patterns of activity-dependentconduction velocity changes differentiate classes of unmyelinatedmechano-insensitive afferents including cold nociceptors, in pig and inhuman. Pain 148:59–69
Ostlere LS, Cowen T, Rustin MH (1995) Neuropeptides in the skin of patientswith atopic dermatitis. Clin Exp Dermatol 20:462–7
Pavlovic S, Daniltchenko M, Tobin DJ et al. (2008) Further exploringthe brain-skin connection: stress worsens dermatitis via substanceP-dependent neurogenic inflammation in mice. J Invest Dermatol128:434–46
D Roggenkamp et al.Innervated Skin Model
www.jidonline.org 1627

Pereira U, Boulais N, Lebonvallet N et al. (2010) Development of an in vitrococulture of primary sensitive pig neurons and keratinocytes for the studyof cutaneous neurogenic inflammation. Exp Dermatol 19:931–5
Peters EM, Arck PC, Paus R (2006a) Hair growth inhibition by psychoemo-tional stress: a mouse model for neural mechanisms in hair growthcontrol. Exp Dermatol 15:1–13
Peters EM, Ericson ME, Hosoi J et al. (2006b) Neuropeptide control mechan-isms in cutaneous biology: physiological and clinical significance. J InvestDermatol 126:1937–47
Peters EM, Liezmann C, Spatz K et al. (2010) Nerve growth factor partiallyrecovers inflamed skin from stress-induced worsening in allergic inflam-mation. J Invest Dermatol 131:735–43
Pozo D (2003) VIP- and PACAP-mediated immunomodulation as prospectivetherapeutic tools. Trends Mol Med 9:211–7
Prado MA, Evans-Bain B, Dickerson IM (2002) Receptor component protein(RCP): a member of a multi-protein complex required for G-protein-coupled signal transduction. Biochem Soc Trans 30:460–4
Proksch E, Folster-Holst R, Jensen JM (2006) Skin barrier function, epidermalproliferation and differentiation in eczema. J Dermatol Sci 43:159–69
Regoli D, Drapeau G, Dion S et al. (1987) Pharmacological receptors forsubstance P and neurokinins. Life Sci 40:109–17
Reilly DM, Ferdinando D, Johnston C et al. (1997) The epidermal nerve fibrenetwork: characterization of nerve fibres in human skin by confocalmicroscopy and assessment of racial variations. Br J Dermatol 137:163–70
Roggenkamp D, Falkner S, Stab F et al. (2012) Atopic keratinocytes induceincreased neurite outgrowth in a coculture model of porcine dorsal rootganglia neurons and human skin cells. J Invest Dermatol 132:1892–900
Rukwied R, Schley M, Forsch E et al. (2010) Nerve growth factor-evokednociceptor sensitization in pig skin in vivo. J Neurosci Res 88:2066–72
Salomon J, Baran E (2008) The role of selected neuropeptides in pathogenesisof atopic dermatitis. J Eur Acad Dermatol Venereol 22:223–8
Saraceno R, Kleyn CE, Terenghi G et al. (2006) The role of neuropeptides inpsoriasis. Br J Dermatol 155:876–82
Schulze E, Witt M, Fink T et al. (1997) Immunohistochemical detection ofhuman skin nerve fibers. Acta Histochem 99:301–9
Seike M, Ikeda M, Morimoto A et al. (2002) Increased synthesis of calcitoningene-related peptide stimulates keratinocyte proliferation in murine UVB-irradiated skin. J Dermatol Sci 28:135–43
Severini C, Improta G, Falconieri-Erspamer G et al. (2002) The tachykininpeptide family. Pharmacol Rev 54:285–322
Stander S, Luger TA (2010) Itch in atopic dermatitis - pathophysiology andtreatment. Acta Dermatovenerol Croat 18:289–96
Steinhoff M, Stander S, Seeliger S et al. (2003) Modern aspects of cutaneousneurogenic inflammation. Arch Dermatol 139:1479–88
Stern RS (1997) Psoriasis. Lancet 350:349–53
Suuronen EJ, Nakamura M, Watsky MA et al. (2004) Innervated human cornealequivalents as in vitro models for nerve-target cell interactions. FASEB J18:170–2
Tanaka T, Danno K, Ikai K et al. (1988) Effects of substance P and substance Kon the growth of cultured keratinocytes. J Invest Dermatol 90:399–401
Tandrup T (1995) Are the neurons in the dorsal root ganglion pseudounipolar?A comparison of the number of neurons and number of myelinated andunmyelinated fibres in the dorsal root. J Comp Neurol 357:341–7
Tobin D, Nabarro G, Baart de la Faille H et al. (1992) Increased number ofimmunoreactive nerve fibers in atopic dermatitis. J Allergy Clin Immunol90:613–22
Vetrugno R, Liguori R, Cortelli P et al. (2003) Sympathetic skin response: basicmechanisms and clinical applications. Clin Auton Res 13:256–70
Walker CS, Conner AC, Poyner DR et al. (2010) Regulation of signaltransduction by calcitonin gene-related peptide receptors. Trends Phar-macol Sci 31:476–83
Wimalawansa SJ (1997) Amylin, calcitonin gene-related peptide,calcitonin, and adrenomedullin: a peptide superfamily. Crit Rev Neuro-biol 11:167–239
Yu XJ, Li CY, Xu YH et al. (2009) Calcitonin gene-related peptide increasesproliferation of human HaCaT keratinocytes by activation of MAP kinases.Cell Biol Int 33:1144–8
Zhang Z, Winborn CS, Marquez de Prado B et al. (2007) Sensitization ofcalcitonin gene-related peptide receptors by receptor activity-modifyingprotein-1 in the trigeminal ganglion. J Neurosci 27:2693–703
D Roggenkamp et al.Innervated Skin Model
1628 Journal of Investigative Dermatology (2013), Volume 133





























