Embed Size (px)
Citation preview

Gut 1997; 40: 196-203
Epithelium related deposition of activatedcomplement in Helicobacter pylori associatedgastritis
A E Berstad, P Brandtzaeg, R Stave, T S Halstensen
AbstractBackground and Aims-It is unknownwhether Helicobacter pyloni infectionactivates complement in vivo. Mucosaldeposition of various activation productsofthe complement system may contributeto the pathogenesis of chronic gastritisand was therefore studied by immuno-histochemistry.Patients and Methods-Ethanol fixedantrum or body gastric tissue sectionsfrom 24 patients infected with H pylori(determined by bacterial immunohisto-chemistry) and 22 uninfected patientswere examined by immunofluorescencewith monoclonal antibodies to activationneoepitopes in C3b and in the terminalcomplex (TCC). As a control group,biopsy samples from the gastric stump of23 Billroth II operated patients werestudied.Results-Patchy, bright staining for TCCoccurred below the surface epitheliumand around the glands in Hpylori positiveand negative gastritis as weli as in stumpgastritis but seldom in normal mucosa.Activated C3 was present at the apical faceof the surface epithelium, significantlymore often in the antrum and body frompatients with than without H pyloniinfection (p=0*05 and p=003 respectively),and particularly in samples with granulo-cyte infiltration (p=0 04). Many bacteriawere coated with activated C3 towards thepit openings but seldom within thefoveolae.Conclusions-Local complement activa-tion was shown to take place in simplechronic gastritis, associated as weli as un-associated with H pylori infection, andalso in stump gastritis. The fact thatactivated C3 was seldom seen on H pyloniwithin the foveolae, suggested that thebacterium evades complement attack inthis location.(Gut 1997; 40: 196-203)
Keywords: Helicobacterpylori, gastritis, immunofluor-escence, activated complement.
Helicobacter pylori colonises the gastric epi-thelium and leads to a chronic inflammatoryreaction.' It is intriguing that this organismpersists despite the induction of both humoraland cellular immunity. H pylorn specific IgGand IgA antibodies occur in peripheral blood
as well as in the gastric mucosa of infectedsubjects.2 3 The antibodies are probablyimportant in restricting the infection to themucosal surface but are not sufficient toeliminate the bacteria.4 Furthermore, it isunknown to what extent humoral or cellmediated immunopathology is involved inchronic gastritis.The systemic antibody response to H pyloni
includes all four subclasses of IgG althoughIgGI and IgG2 antibodies predominate.5Antral and body IgG and IgA antibodiesmeasured in homogenates of mucosal biopsyspecimens, are significantly increased in Hpylori positive patients - the IgG response beingparticularly noticeable in those with activechronic pangastritis.6 Externally translocatedantibodies of the IgA, IgG, and IgM classesbind in vivo to H pylori and the frequency ofcoated bacteria is higher when neutrophils arepresent in the mucosa than in inactivegastritis.7 Furthermore, H pyloni infection canapparently induce gastric autoantibodies8 thatto some extent are directed against the gastrinproducing cells9 10; antibodies to interleukin 8(IL 8) have also been reported." In addition,H pylori seems to share antigenic determinantswith foveolar epithelial cells and pyloricglands.'2 The induction of various types ofgastric autoantibodies could be one way inwhich the bacterium causes full thicknesslesions in the mucosa.H pylori specific antibodies of the IgG class
are not regularly detected in gastric juice' 6because this class is not actively transported tothe gastric lumen.'3 However, small amounts ofIgG can passively diffuse between epithelialcells,'3 particularly in inflammation where Hpylori infection can lead to a weakening of thetight junction complexes between surfacemucous cells.'4 Because IgG does not combinewith secretory component (SC), this antibodyclass is rapidly degraded in gastric juice.'5The numbers of mucosal IgA, IgM, and IgG
producing plasma cells and blasts increasesignificantly with increasing degree of gastritis;the IgG class shows the largest relative in-crease, particularly in the basal part of themucosa.16 H pylori antigens that cross thestomach epithelium may form subepithelialimmune complexes with locally producedantibodies and thereby cause complementactivation and inflammation (type III hyper-sensitivity) as recently discussed by Ernst etal." Based on the detection of neoepitopes inthe terminal complement complex (TCC) andC3b, activated complement has been observed
Laboratory forImmunohistochemistryand immunopathology(LIIPAT),Institute ofPathology,University of Oslo,The National Hospital,Rikshospitalet,Oslo, NorwayA E BerstadP Brandtzaeg
Medical Department,Lovisenberg Hospital,Oslo, NorwayR Stave
National Institute ofPublic Health,Department ofEnvironmentalMedicine,Oslo, NorwayT S HalstensenCorrespondence to:Dr A E Berstad,LIIPAT, Rikshospitalet,N-0027 Oslo, Norway.Accepted for publication28 August 1996
196
group.bmj.com on July 6, 2017 - Published by http://gut.bmj.com/Downloaded from

Complement activation in gastric mucosa
locally associated with an increased mucosalIgG response in patients with inflammatorybowel disease,'8 and related to increasedconcentrations of serum IgG and IgMantibodies to gluten in coeliac disease.'9 Inview of the large amounts of IgG producedlocally in Hpylori associated gastritis,'" it wouldbe ofinterest to look for complement activationalso in this mucosal lesion. Furthermore, invitro studies have suggested that H pylon canactivate the classic pathway of complementeven in the absence of antibodies.20
In this study we applied monoclonalantibodies (mAbs) against neoepitopes of C3b(early activation) and TCC (late activation) tosections of directly ethanol fixed mucosal tissuesamples with or without gastritis. Mechanismsof complement activation were also studied.Such in situ studies have to our knowledge notbeen performed earlier in gastritis.
Methods
Specimens and tissue preparationSpecimens of gastric antral and body mucosaobtained from nine women and 12 men witha mean age of 63 years (range, 26-81), werefixed directly in cold 96% ethanol for 24 hoursat 4°C and then embedded in paraffin wax;or they were washed for 48 hours at 4°C in0-01 M phosphate buffer (pH 7-5) containing0- 15 M NaCl (phosphate buffered saline, PBS)to extract diffusible proteins before ethanolfixation.2' Most ofthese samples had been usedin an earlier immunohistochemical study'6;they were collected surgically from 21 peoplefrom the following groups: patients operatedwith Billroth II (BID) resection for duodenal orgastric ulcer (n= 10); patients operated forduodenal neoplasia (n=2); severe kidneyfailure and gastritis (n=5); and kidney donors(n=4). Two to four additional formalin fixed,paraffin wax embedded mucosal samples fromthe antrum and body were used forimmunohistochemical detection of H pylon(see later).
Additional material consisted of endoscopicbiopsy specimens, one antrum and one bodysample, obtained from each of 25 patientsattending an outpatient gastronterology clinicfor various abdominal complaints. Thesesubjects included 13 women and 12 men witha mean age of 47 years (range, 20-79). Onespecimen from the antrum and one fromthe corpus were used for the im-munohistochemical detection of H pylori. Aseparate antral specimen from each patient wasused in the rapid urease test for detection ofHpylon (see later). Informed consent wasobtained from all these subjects, and theproject was approved by the Clinical ResearchEthics Committee ofHealth Region II. Neitherpatients with malignant diseases nor thosetaking immunosuppressive drugs or havingreceived antimicrobial treatment during thepast two months were included. A bloodsample was available from 20 ofthe patients fordetermination of serum H pylon IgG anti-bodies.
Endoscopic biopsy specimens (n=37) werealso obtained from the gastric stump of 23patients with a mean age of 64 years (range,51-75) who had been subjected to BIIresection for duodenal ulcer 27-32 years earlier(mean, 29.5). The specimens were taken fromthe lesser and the greater curvature no morethan 2 cm beyond the stoma. This groupserved solely as controls and was not includedin the statistical analysis.
All endoscopic biopsy samples werecollected from areas without macroscopicalaffection except for gastritis; they were fixeddirectly in cold 96% ethanol and embedded inparaffin wax.2'
Determination ofH pylori infectionThe H pylori infection status of patients in thestudy group was determined by immuno-histochemistry on sections (5 [um) of formalinor ethanol fixed tissue from both the antrumand body mucosa. A rabbit antiserum (1/10) toH pylori (DAKO, Glostrup, Denmark) wasapplied for 20 hours at room temperature.Antigenic retrieval by proteolytic digestion(10 g/l trypsin, 10 min, 37°C) was first per-formed for formalin fixed sections. Theantiserum distinguished H pylori from othercurved bacteria present in the stomach and hada sensitivity of 100% and a specificity of 94%compared with cultivation results.22 Fluor-escein isothiocyanate (FITC) conjugatedswine antirabbit IgG (1/160; DAKO) wasapplied for three hours as secondary reagent.Omission of the primary antiserum provided anegative control. After mounting, the tissuesections were examined by fluorescence micro-scopy (see later).To confirm the bacterial immunostaining in
the patient group providing fresh endoscopicbiopsy specimens from the antrum and body,one antral sample was tested for the presenceof H pylori urease in a urea solution at roomtemperature for three hours with phenol redpH indicator.23 A colour change from yellow topink was regarded as positive. In the samegroup of patients, serum IgG antibodies to Hpylon were tested for by an enzyme im-munoassay method (Pyloriset EIA-G, OrionDiagnostica, Espoo, Finland; sensitivity 92%,specificity 84%).24 The serum samples from 20patients attending the outpatient clinic wereexamined in duplicate at a dilution of 1/200.If the antibody level was higher than that ofreference serum no 2, the result was consideredpositive for H pylori antibodies. The same testwas used to screen the infection status of thecontrol group of stump gastritis patients.
Immunohistochemical detection of activatedcomplementThe three step two colour biotin/avidinenhanced immunofluorescence staining pro-cedure used was a slight modification of apreviously described method.'8 Serial paraffinwax sections (5,m) from directly ethanol fixedspecimens were dewaxed and incubated for 20hours with mAb to TCC (Clone aEl 1, murine
197
group.bmj.com on July 6, 2017 - Published by http://gut.bmj.com/Downloaded from

Berstad, Brandtzaeg, Stave, Halstensen
IgG2a; 1 mg/1)25 or to a C3b activationneoepitope in the C3c part of C3b/iC3b(Clone bH6, murine IgG2a; 7-5 mg/1),26 incombination with rabbit antiserum to cyto-keratin (1/100)27 or to S protein/vitronectin(1/5000).28 Selected specimens from fivepatients found to have apical complementdeposition of C3b were, in addition, examinedwith rabbit antiserum to Clq (1/500; DAKO)and C4c (1/500; DAKO) in combination withmAb to C3b. The mAbs and antisera wereappropriately diluted in PBS containing bovineserum albumin (12-5 g/l) on the basis of per-formance testing. Biotinylated horse anti-mouse IgG (2 mg/l; Vector Laboratories,Burlingame, CA) was incubated for threehours, followed by FITC-conjugated swineantirabbit IgG (1/160; DAKO) and Strep-tavidin Texas red conjugate (1/200; GibcoBRL, Gaithersburg, MD) for 30 minutes.Nine tissue samples positive for C3b apically
on the epithelium, were restained with the threestep immunofluorescence method but with anti-serum to H pylori (1/10) included instead ofantiserum to cytokeratin. By this modificationthe bacteria appeared green (FITC) and thecomplement neoepitopes red (Texas red).Dewaxed sections of 12 prewashed tissuesamples were examined for TCC and C3b.These washed tissue samples had been obtainedconcomitantly with and from the same areas asthose subjected to direct alcohol fixation inwhich complement deposits were observed.
Staining specificity was ensured by adissimilar (or negative) decoration patternobtained with different antibodies applied toserial sections. Omission of the primaryantibody also provided a negative control, asdid application of an irrelevant mAb (mouseIgG2a, 5 mg/l; Becton Dickinson, CA).To visualise neutrophilic granulocytes, mAb
to human neutrophil elastase (Clone NP57,murine IgGl, 12-6 jug/l; DAKO) was appliedin alkaline phosphatase antialkaline phospha-tase (APAAP) staining29 to parallel sections.This method also facilitated the grading ofintestinal metaplasia, as the epithelial brushborder stained bright red because of endo-genous alkaline phosphatase.
Fluorescence microscopyThe sections were examined blind by the sameinvestigator using a Leitz DMR-DXE micro-scope camera equipped with a Ploem-typevertical illuminator system (Leitz, Wetzlar,Germany). A dual filter for the simultaneousobservation of red (Texas red) and green(FITC) emission made both colours appeartogether as yellow. Pictures were recorded onEktachrome 800/1600 ISO daylight filmpushed to 800 ISO. Subepithelial TCC andapical C3b were semiquantitatively scoredfrom no (-) to intense (3) as done earlier.'9Grade 1 indicated distinct but patchy stainingin a few areas, and grade 3 continuous intensestaining. Deposits of TCC were scored separ-ately in the foveolar and glandular part of themucosa. TCC staining of elastic fibrils, mus-cular tissue, and vessels30 was not recorded.
The same investigator performed theimmunofluorescence evaluation throughoutthe study. Six weeks after the completion of thestudy, 10 sections were randomly selected forblind re-evaluation.
Grading ofgastritisParallel tissue sections from each specimenwere stained with haematoxylin and eosin andgraded for gastritis by one observer accordingto the Sydney system.3' The histologicalvariables were scored on a four point scale: 0,absent; 1, mild; 2, moderate; and 3, severe.Inflammation was examined for the presenceand density of mononuclear cells in the laminapropria; activity for the presence and density ofneutrophils in the lamina propria as well as inthe epithelium; atrophy for the loss of gastricglands; and intestinal metaplasia for thepresence of intestinal glandular tissue.Randomly selected sections were graded bytwo observers with excellent agreement.Grading of gastritis was performed withoutknowledge of the patients' clinical or infectionstatus.
Statistical analysisSamples with inflammation were combined,and complement activation was recorded aspresent or absent before statistical analysis wasperformed with Fisher's exact test, includingthe Bonferroni correction for multiple com-parisons. Reproducibility of immunofluor-escence scores was determined by the weightedK test.32 The Spearman's rank correlation testwas used to evaluate any association betweenthe degree of subepithelial TCC depositionand serum IgG antibody titres to H pylori. Avalue ofp<0 05 was considered significant.
Results
Infection status and degree ofpathology in gastritisDirectly ethanol fixed tissue samples fromantral (n=43) and body (n=43) mucosa of 46patients were studied. Of these patients, 24were considered to be infected with H pylori(52%) as determined by immunofluorescencein situ. The Table lists the results of im-munostaining, the rapid urease test, and theserum IgG antibody determination.
All gastric biopsy specimens (n=46) from Hpylori positive subjects were inflamed: 24 antralsamples showed inflammation grade 1 (n=9),grade 2 (n=13) or grade 3 (n=2); and 22 bodysamples showed grade 1 (n=14) or grade 2(n=8). Four antral samples from H pylorinegative subjects and 17 from Hpylori positivesubjects showed epithelial neutrophil in-filtration.
Directly ethanol fixed specimens (n=40)from H pylon negative subjects were lessinflamed: 19 antral samples were eitherconsidered normal (n=12), of grade 1 (n=6),or of grade 2 (n= 1); 21 body samples wereeither considered normal (n= 13) or of grade 1(n=8). One body sample from a H pylori
198
group.bmj.com on July 6, 2017 - Published by http://gut.bmj.com/Downloaded from

Complement activation in gastric mucosa
H pylori status ofpatients based on in situ immunofluorescence*
Immunofluorescence Rapid urease test H pylori antibodiesSamnple No ofcategory observations Positive Negative Positive Negative Positive Negative
Archival 21 12 9material
Outpatient 5 2 3 2 3biopsyspecimens 20 10 10 8 12 7 1 3
Total 46 24 22
*From patients providing archival material, immunofluorescence alone was performed. For fiveoutpatients, immunofluorescence was complemented by the rapid urease test, and there wascomplete agreement. In 20 outpatients, serology was also performed and this turned out to bethe least sensitive test.
negative subject and 11 from H pylori positivesubjects showed epithelial neutrophil infiltra-tion.
Biopsy samples were available from bothantrum and body mucosa in 40 subjects: 10hack predominantly antral gastritis; 18 hadpangastritis; and one had predominantly bodygastritis. Eleven patients showed no inflam-mation in either the antral or the bodysamples.
Deposition of activated complement in gastritisBright staining for the TCC neoepitope wasfound below the epithelium related to thebasement zone in 23 of 43 antral (53%) andin 20 of 43 body (47%) samples. Thesedeposits occurred in both Hpylori positive andnegative gastritis. TCC was located in a patchymanner below the surface epithelium but wasseen only seldom related to the foveolar pits ornecks (Fig 1A). TCC also occurred deep in themucosa along and below the basementmembrane zone and scattered between theglands (Fig 1B) in 26 of 43 antral samples(60%) and in 21 of 40 body samples (53%).Glands were not adequately represented inthree body samples. Bright staining of largeblood vessels, and of the mucus above thesurface epithelium as well as faint staining inmuscular tissue were seen in some specimensbut not recorded systematically.
Statistical analysis showed that subepithelialdeposition of TCC in the antrum and body(Fig 2) was significantly more often present inspecimens with than in those without
Figure 1: Two colour immunofluorescence staining (double exposure) ofcomplement deposits in sections of directly ethanolfixed antral mucosafrom patients with H pylori gastritis. (A) TCC (Texas red) is located beneath the surface epithelumthat is visualised by stainingfor cytokeratin (FITC, green). (B) Same staining combination shows TCC around glandularelements. (C) Two colour stainingfor TCC (Texas red) and S protein (FITC) shows colocalisation (yellow) aroundglandular elements. (D) Two colour stainingfor C3b (Texas red) and cytokeratin (FITC) shows luminal deposits. (E) Twocolour stainingfor C3b (Texas red) and H pylori (FITC) shows coated bacteria (yellow) on the surface epithelium,whereas bacteria in the gastric pits are uncoated (pure green). Original magnifications: A, x 1000; B, x 400; C, X 250;D, X400; E, x 1000.
199
group.bmj.com on July 6, 2017 - Published by http://gut.bmj.com/Downloaded from

Berstad, Brandtzaeg, Stave, Halstensen
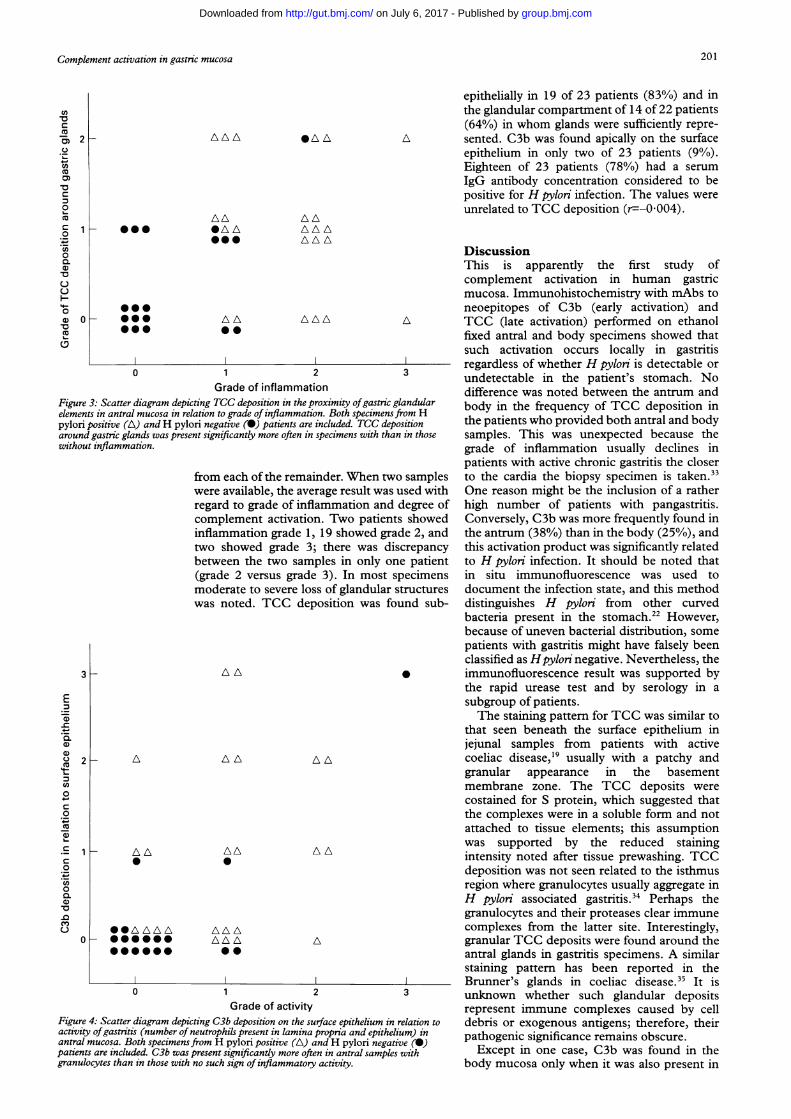
inflammation (p=0*001 and p=0 004 re-spectively). The same was true (p=0 02) forglandular TCC deposition in the antrum(Fig 3). Specimens with mild to severe lossof glandular structures were found in 15of 43 antral specimens and in six of 40body specimens. Of the 15 antrum and sixbody specimens with atrophy, TCC stainingaround and in relation to remaining glandswas found in 10 and four specimens respec-tively.
In patients who provided both antral andbody specimens (n=40), the frequency ofTCCdeposition was similar in the antrum and body.In these patients, subepithelial deposition ofTCC was found in 21 of 40 antral samples(53%) and in 20 of 40 body samples (50%).
AAz~
\ AL* * A
* A A* * A
/\ /\ /\ AA AA
A
A
AA
31 2Grade of inflammation
B
A A
I* AA
S 0A AA*A
/\/\ *A AAA
AAA
*---ILILIL0
Only faint staining for TCC was present inprewashed tissue specimens; the intensity wasclearly decreased compared with that seen indirectly alcohol fixed tissue taken fromadjacent tissue sites. This suggested that TCCwas present in a fluid phase rather than beingmembrane bound. Staining for S protein indirectly alcohol fixed tissue samples showedcolocalisation with TCC (Fig 1C).
Epithelium related deposits of C3b were notseen in prewashed tissue except in twospecimens from the same patient where thestaining was clearly reduced compared withsimilar staining after direct ethanol fixation. Insuch specimens, C3b appeared to be related tothe apical face of the surface epithelium (Fig1D) in 16 of 43 antral samples (37%) and in10 of 43 body samples (23%), a locationremarkably different from that of TCC. In thepatients who provided both antral and bodysamples (n=40), C3b was more frequentlyfound in the antrum (38%) than in the body(25%). The C3b staining varied from analmost continuous band to scattered patchesand often extended between the adjacentepithelial cells as well as into the mucus layer.It was neither found in the neck nor in theglandular epithelial areas and was likewiseabsent from the lamina propria. When biopsysamples from patients with or withoutdetectable infection were compared, apicalC3b was found more often in antral and bodysamples from the H pyloni positive group(p=0 05 and 0 03 respectively). Moreover,C3b was significantly more often present inantral samples with granulocytes present in thelamina propria and epithelium than in thosenot showing such a sign of inflammatoryactivity (p=0 04, Fig 4).Antiserum to the classic activation com-
ponents Clq and C4c produced faint co-staining with C3b in directly ethanol fixedspecimens, but considerable backgroundstaining rendered these findings inconclusive.Such costaining could not be performedreliably on prewashed tissue specimensbecause of the considerable reduction instaining intensity for C3b.H pylon produced a strong staining signal
with the polyclonal antibody reagent applied toalcohol fixed samples. Towards the openings ofthe gastric pits, bacteria were found to bepositive for C3b as shown by paired im-munofluorescence staining, while they werenegative deeper in the pits (Fig 1E). Readilyidentifiable positive bacteria were seen in fourof 16 patients with C3b staining related to theantral surface epithelium.There was good agreement between the first
and second observations of C3b and TCCdepositions (KW=0-71). Of 30 repeat observa-tions, 22 were identical, four provided anegative difference of one grade, and four apositive difference of one grade.
0 1 2Grade of inflammation
Figure 2: Scatter diagram depicting subepithelial deposition of TCC in gastric antral (A)and body (B) mucosa in relation to grade ofinflammation. Both specimensfrom H pylonpositive (A) andH pylon negative (0) patients are included. TCC in the antrum andbody was present significantly more often in specimens with than in those withoutinflammation.
Observations in gastric stumpsBiopsy specimens from 23 BII operatedpatients were studied. Two samples wereavailable from each of 14 patients, and one
A
E4-
cn
uC
Q
-
0C0
00.
0.').0Co0a,
~0Cu
2
0000*000* A A A
0
m
0
E> 2-o0.0c
00H
0
0._
a)~0
0._
.0Co0
a)
CuC.'
*0000 0000000
II~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~
I I~~~~~~~~~~~~~~~~~~~~~~~~~~~~~
200
group.bmj.com on July 6, 2017 - Published by http://gut.bmj.com/Downloaded from
Complement activation in gastric mucosa
AILAA
0@@@000000
AA*AA
A A0@
A AAAA
AAA
0 1 2Grade of inflammation
Figure 3: Scatter diagram depicting TCC deposition in the proximity ofgastric g,elements in antral mucosa in relation to grade of inflammation. Both specimens)pylori positive (A) andH pylori negative (0) patients are included. TCC deptaround gastric glands was present significantly more often in specimens with thaiwithout inflammation.
from each of the remainder. When tw(were available, the average result was i
regard to grade of inflammation and i
complement activation. Two patientinflammation grade 1, 19 showed gratwo showed grade 3; there was disbetween the two samples in only on(grade 2 versus grade 3). In most slmoderate to severe loss of glandular swas noted. TCC deposition was foi
3 AA
2 _
0
A
- AL0
* * A A A A000000
00*000
0
A A
AA
AAA
AAA
0
AA
AL
1 2Grade of activity
Figure 4: Scatter diagram depicting C3b deposition on the surface epithelium inactivity ofgastritis (number of neutrophils present in lamina propria and epitheliantral mucosa. Both specimens from H pylon positive (A) andH pylori negati
patients are included. C3b was present significandy more often in antral samplesgranulocytes than in those with no such sign ofinflammatory activity.
epithelially in 19 of 23 patients (83%) and inthe glandular compartment of 14 of 22 patients(64%) in whom glands were sufficiently repre-
A sented. C3b was found apically on the surfaceepithelium in only two of 23 patients (90/o).Eighteen of 23 patients (78%) had a serumIgG antibody concentration considered to bepositive for H pylon infection. The values wereunrelated to TCC deposition (r=--0004).
DiscussionThis is apparently the first study ofcomplement activation in human gastricmucosa. Immunohistochemistry with mAbs toneoepitopes of C3b (early activation) and
A TCC (late activation) performed on ethanolfixed antral and body specimens showed thatsuch activation occurs locally in gastritis
-0------- regardless of whether H pylon is detectable or3 undetectable in the patient's stomach. No
difference was noted between the antrum and,landular body in the frequency of TCC deposition inrsition the patients who provided both antral and bodya in those samples. This was unexpected because the
grade of inflammation usually declines inpatients with active chronic gastritis the closer
a samples to the cardia the biopsy specimen is taken.33used with One reason might be the inclusion of a ratherdegree of high number of patients with pangastritis.s showed Conversely, C3b was more frequently found inde 2, and the antrum (38%) than in the body (25%), and,crepancy this activation product was significantly relatedLe patient to H pylon infection. It should be noted thatpecimens in situ immunofluorescence was used to,tructures document the infection state, and this methodund sub- distinguishes H pylon from other curved
bacteria present in the stomach.22 However,because of uneven bacterial distribution, somepatients with gastritis might have falsely beenclassified as Hpyloni negative. Nevertheless, the
* immunofluorescence result was supported bythe rapid urease test and by serology in asubgroup of patients.The staining pattern for TCC was similar to
that seen beneath the surface epithelium injejunal samples from patients with activecoeliac disease,'9 usually with a patchy andgranular appearance in the basementmembrane zone. The TCC deposits werecostained for S protein, which suggested thatthe complexes were in a soluble form and notattached to tissue elements; this assumptionwas supported by the reduced stainingintensity noted after tissue prewashing. TCCdeposition was not seen related to the isthmusregion where granulocytes usually aggregate inH pylon associated gastritis.34 Perhaps thegranulocytes and their proteases clear immunecomplexes from the latter site. Interestingly,granular TCC deposits were found around theantral glands in gastritis specimens. A similarstaining pattern has been reported in theBrunner's glands in coeliac disease.35 It is
3 unknown whether such glandular depositsrepresent immune complexes caused by cell
relation to debris or exogenous antigens; therefore, theirurm) in pathogenic significance remains obscure.with Except in one case, C3b was found in the
body mucosa only when it was also present in
cn
cn-o
C
c.C'a
-0C0
CLl0C',-0
._
uoa)
F-0
0
U0
E-c0.._
a)0C)(00C
It
cn04-
C
0
.0_
c
0.
0.
~0.0
cnuQ,
Q)
201
1
group.bmj.com on July 6, 2017 - Published by http://gut.bmj.com/Downloaded from

202Berstad, Brandtzaeg, Stave, Halstensen
the antrum. This early activation product wasdistributed in a patchy manner apically on thesurface epithelium but not in the foveolae.Although this deposition resembled that seenapically on the colonic surface epithelium ofpatients with active ulcerative colitis, the latterwas more extensive and generally associatedwith TCC.18
Because C3b was localised to the epithelialsurface in chronic gastritis, a dynamicinteraction may exist between the bacteriumand the host - perhaps induced by H pyloriantigens. Complement activation via theclassic pathway that involves C3b, is initiatedby soluble antigen-antibody complexes or bythe binding of antibody to target antigens suchas bacteria. Antibodies of IgA, IgG, and IgMisotypes have been shown to coat H pylori invivo, the two latter classes (which arecomplement activating) usually in the presenceof epithelial neutrophil infiltration.7
In this study, we were unable to draw anyconclusion as to whether the classic oralternative activation pathway was involved inthe deposition of C3b. H pylon is not overtlyinvasive, but proinflammatory bacterial factorsmay reach the lamina propria, particularly ifthe epithelial barrier is damaged. It has alsobeen proposed that bacterial antigens mayform immune complexes with IgG and activatecomplement.' 1-7 Lipopolysaccharides fromGram negative bacteria are known to beinitiators of the alternative pathway ofcomplement activation and C3 has a labileinternal thioester bond that is activated as C3bis formed, allowing the C3b fragment to bindto free hydroxyl or amino groups on a cellmembrane.36 It is therefore likely that bothcomplement activation pathways are involvedin Hpylori associated gastritis.
It was interesting to notice that Hpylori wascostained for C3b near the openings of gastricpits and on the luminal surface. The fact thatseveral bacteria were negative in the samesection excluded unwanted antibody crossreactivity between the C3b neoepitope and Hpylori. Bacteria deeper in the pits remainedunstained, perhaps reflecting evasion ofcomplement attack; this would agree with theabsence of IgG and IgM antibody coating inthis location.7 C3b coating of the bacteria wasnot present in all Hpylori infected patients. Wecannot explain why C3b was seen to coat Hpylon in some patients, but not in others. Theethanol fixation had apparently removedbacteria and the mucus layer from the surface,which made evaluation of C3b coating evenmore difficult.
Binding of C3b to H pylon may facilitateopsonisation by cells bearing complementreceptors, thereby enhancing phagocytosis byneutrophils, monocytes, and macrophages.37In vitro studies have shown that complementis an efficient opsonin for H pylon, eitheractivation pathway being sufficient foropsonisation on its own.38 If in vivo activationof complement takes place on the surface ofHpylon, rapid hydrolysis would limit bindingof C3b to nearby host cells.36 Mammaliancells have high levels of sialic acid, which
contributes to rapid inactivation ofbound C3bmolecules.36 Microorganisms generally lackmammalian complement regulatory proteinsand can activate complement in the absence ofantibody via the alternative pathway. Thisactivation leads to accumulation of a largenumber of C3b molecules on their surface, butmany bacteria have developed strategies toevade destruction.39 However, surprisinglylittle is known about the local protectivemeasures possessed by H pylori and by thegastric epithelial cells.The actual role of chemotactic substances
(C3a, C4a, and C5a) released by complementactivation in gastritis associated with H pyloricolonisation remain undetermined, but wenoticed that the C3b deposits were sig-nificantly related to the presence of neutrophilsin the mucosa. However, these products areunlikely to be the only factors attractingneutrophils to gastric lesions; for example,increased epithelial expression of thechemokine interleukin 8 has been seen ingastritis compared with normal mucosa.40Complement could also be important inneutrophil activation in H pylori associatedgastritis. In an earlier study,20 H pylori did notinduce a significant increase in neutrophilchemiluminescence in the absence of opsonins,but this was considerably increased by theaddition of fresh agammaglobulinaemic serumas a source of complement. The complementactivating abilities of different H pylori strains(type I, type II) should be tested in futurestudies, as the ability to activate complementmight be an important virulence marker.TCC was present in mucosal samples from
most (83%) BII operated patients, whereasC3b was found in only few (9%). Preferentialdeposition ofTCC beneath the epithelium andin the glandular areas probably reflectedchronic immunopathology; these patients wererelatively old and had longstanding gastriclesions. Serum IgG antibodies to Hpylori wereoften found at a level indicating infection inthis group (78%), apparently without beingrelated to the TCC deposition. This supportedour finding in the gastritis group thatdeposition of TCC along the basementmembrane is not necessarily related to Hpyloni-infection. The fact that Hpylori is seldom foundnear the stoma in gastric stumps4' suggests thatother factors (for example, bile reflux) maintainthe inflammation in stump gastritis.
In conclusion, complement activation takesplace in chronic gastritis, both associated andunassociated with Hpylon infection, as well asin stump gastritis after BII resection. However,the pattern of complement deposition differs,which suggests separate immunopathologicalmechanisms. The role of activated comple-ment for neutrophil chemotaxis and epithelialdestruction in gastritis remains undetermined;but it is of considerable interest that activatedC3 was seen to coat H pylon and to bedeposited at the apical face of the surfaceepithelium preferentially in infected patients.We thank members of the staff at the Gastroenterology Sectionof the Medical Department at Lovisenberg Hospital, Oslo, fortheir cooperation, and Dr Kolbjorn Valnes for providing tissue
group.bmj.com on July 6, 2017 - Published by http://gut.bmj.com/Downloaded from

Complement activation in gastric mucosa
samples from his previous studies. The excellent technicalassistance of the LIIPAT laboratory staff is gratefullyacknowledged. Financial support was provided by theNorwegian Cancer Society, the Research Council of Norway,and Anders Jahre's Foundation. Glaxo Welcome A/S, Norway,is acknowledged for covering the costs of the colour plate.
1 Blaser MJ. Helicobacter pylori and the pathogenesis ofgastroduodenal inflammation. Jf Infect Dis 1990; 161:626-33.
2 Rathbone BJ, Wyatt JI, Worsley BW, Shires SE,Trejdosiewicz LK, Heatley RV, et al. Systemic and localantibody responses to gastric Campylobacter pyloridis innon-ulcer dyspepsia. Gut 1986; 27: 642-7.
3 Perez-Perez GI, Dworkin BM, Chodos JE, Blaser MJ.Campylobacter pylori antibodies in humans. Ann InternMed 1988; 109:11-7.
4 Shirley LR, Marsh WH. Campylobacter pylori gastritis inchildren with X-linked agammaglobulinemia. [Abstract]J7AlergyClin Immunol 1989; 83: 287.
5 Bontkes HJ, Veenendaal RA, Pefia AS, Goedhard JG, van
Duijn W, Kuiper J, et al. IgG subclass response to
Helicobacter pylori in patients with chronic active gastritisand duodenal ulcer. Scand Jf Gastroenterol 1992; 27:129-33.
6 Veenendaal RA, Schroijen JM, Gotz JM, Pefia AS,Roosendaal R, Veselic M, et al. Salivary, systemic, gastricjuice, and gastric mucosal IgA and IgG Helicobacterpylori antibodies in patients with active chronicHelicobacter pylori associated antral and pangastritis.Dissertation.Leiden: University of Leiden, 1994: 61-77.
7 Wyatt JI, Rathbone BJ, Heatley RV. Local immune response
to gastric Campylobacter in non-ulcer dyspepsia.JfClinPathol 1986; 39: 863-70.
8 Negrini R,LisatoL, Zanella I, Cavazzini L, GulliniS,Villanacci V, et al. Helicobacter pylori infection inducesantibodies cross-reacting with humangastric mucosa.
Gastroenterology 1991; 101: 437-45.9 Vandelli C, Bottazzo GF, Doniach D, Franceschi F.
Autoantibodies to gastrin-producing cells in antral (typeB) chronic gastritis. NEnglJMed 1979; 300: 1406-10.
10 Uibo RM, Krohn KJ. Demonstration of gastrin cellautoantibodies in antral gastritis with avidin-biotincomplex antibody technique.Clin Exp Immunol 1984; 58:341-7.
11 Crabtree JE, Peichl P, Wyatt JI, Stachl U, LindleyIJ. Gastricinterleukin-8 and IgA IL-8 autoantibodies in Helicobacterpylori infection. ScandJl1mmunol 1993; 37: 65-70.
12 Negrini R,Lisato L, Cavazzini L, Maini P, GulliniS,Basso 0, et al. Monoclonal antibodies for specificimmunoperoxidase detection of Campylobacter pylori.Gastroenterology 1989; 96: 414-20.
13 Valnes K, Brandtzaeg P, Elgjo K, Stave R. Specific andnonspecific humoral defense factors in the epithelium ofnormal and inflamed gastric mucosa. Immunohisto-chemical localization of immunoglobulins, secretory
component, lysozyme, and lactoferrin. Gastroenterology1984; 86: 402-12.
14 Bode G, Malfertheiner P, Ditschuneit H. Pathogeneticimplications of ultrastructural findings in Campylobacterpylori related gastroduodenal disease. Scand J Gastro-enterol Suppl 1988; 142: 25-39.
15 Brandtzaeg P. Immunologic basis for celiac disease,inflammatory bowel disease, and type B chronic gastritis.CurrOpin Gastroenterol 199 1; 7: 450-62.
16 Valnes K, Brandtzaeg P, Elgio K, Stave R. Quantitativedistribution of immunoglobulin-producing cells in gastricmucosa: relation to chronic gastritis and glandularatrophy. Gut 1986; 27: 505-14.
17 Ermst PB, Crowe SE, Reyes VE. The immunopathogenesisof gastroduodenal disease associated with Helicobacterpylori infection. Curr Opin Gastroenterol 1995; 11: 512-8.
18 Halstensen TS, Mollnes TE, Garred P, Fausa 0,Brandtzaeg P. Epithelial deposition of immunoglobulinGl and activated complement (C3b and terminalcomplement complex) in ulcerative colitis. Gastro-enterology 1990; 98: 1264-71.
19 Halstensen TS, Hvatum M, Scott H, Fausa 0, Brandt-zaeg P. Association of subepithelial deposition of activatedcomplement and immunoglobulin G and M response to
gluten in celiac disease. Gastroenterology 1992; 102:751-9.
20 Bematowska E, Jose P, Davies H, Stephenson M,Webster D. Interaction of campylobacter species withantibody, complement and phagocytes. Gut 1989; 30:906-11.
21 Brandtzaeg P. Mucosal and glandular distribution ofimmunoglobulin components. Immunohistochemistrywith a cold ethanol-fixation technique. Immunology 1974;26: 1101-14.
22 Andersen LP, Holck S, Povlsen CO. Campylobacter pyloridetected by indirect immunohistochemical technique.APMIS 1988; 96: 559-64.
23 Nysaeter G, Berstad K, Weberg R, Berstad A, Hardar-dottir H. Diagnosis of Helicobacter pylori infection.Rapid urease test, microscopy of smears and culture fromventricular biopsy compared with the 14C-urea breathtest. (Norwegian). Tidsskr Nor Laegeforen 1992; 112:2356-8.
24 Granberg C, Mansikka A, Lehtonen OP, Kujari H,Gronfors R, Nurmi H, et al. Diagnosis of Helicobacterpylori infection by using pyloriset EIA-G and EIA-A fordetection of serum immunoglobulin G (IgG) and IgAantibodies. JClinMicrobiol 1993; 31: 1450-3.
25 Mollnes TE, Lea T, Harboe M, Tschopp J. Monoclonalantibodies recognizing a neoantigen of poly(C9) detectthe human terminal complement complex in tissue andplasma. ScandJImmunol 1985; 22: 183-95.
26 Garred P, Mollnes TE,Lea T, Fischer E. Characterizationof a monoclonal antibody MoAb bH6 reacting with aneoepitope of human C3 expressed on C3b, iC3b, andC3c. Scandj7Immunol 1988; 27: 319-27.
27 Huitfeldt HS, Brandtzaeg P. Various keratin antibodiesproduce immunohistochemical staining of humanmyocardium and myometrium. Histochemistry1985; 83:381-9.
28 Dahlback K, Lofberg H, Dahlback B. Localization ofvitronectin (S-protein of complement) in normal humanskin. Acta Derm Venereol 1986; 66: 461-7.
29 Cordell JL, Falini B, Erber WN, Ghosh AK, Abdulaziz Z,MacDonald S, et al. Immunoenzymatic labeling ofmonoclonal antibodies using immune complexes ofalkaline phosphatase and monoclonal anti-alkalinephosphatase (APAAP complexes). J Histochem Cytochem1984; 32: 219-29.
30 Halstensen TS, Mollnes TE, Brandtzaeg P. Persistentcomplement activation in submucosal blood vessels ofactive inflammatory bowel disease: immunohistochemicalevidence. Gastroenterology 1989; 97: 10-9.
31 Price AB.The Sydney System: histological division.JfGastroenterol Hepatol 1991; 6: 209-22.
32 Altman DG. Some common problems in medical research.In:Practical statistics for medical research.1 st edn. London:Chapman and Hall, 1991: 396-9.
33 Bayerd6rffer E,Lehn N, Hatz RA, Mannes GA, Oertel H,Sauerbruch T, et al. Difference in expression ofHelicobacter pylori gastritis in antrum and body.Gastroenterology 1992; 102: 1575-82.
34 Wyatt JI. Histopathology of gastroduodenal inflammation:the impact of Helicobacter pylori. Histopathology 1995;26:1-15.
35 Gallagher RB, Kelly CP, NevilleS, Sheils 0, Weir DG,Feighery CF. Complement activation within the coeliacsmall intestine is localised to Brunner's glands. Gut 1989;30: 1568-73.
36 Kuby J. The complement system. In: Immunology. 2nd edn.New York: WH Freeman, 1994: 393-416.
37 Frank MM, Fries LF.The role of complement ininflammation and phagocytosis. Immunol Today 1991; 12:322-6.
38 McKinlay AW, Young A, Russell RI, Gemmell CG.Opsonic requirements of Helicobacter pylori. J7 MedMicrobiol 1993; 38: 209-15.
39 Cooper NR. Complement evasion strategies ofmicroorganisms. Immunol Today 1991; 12: 327-31.
40 Crabtree JE, Wyatt JI, Trejdosiewicz LK, Peichl P, NicholsPH, Ramsay N, etal. Interleukin-8 expression inHelicobacter pylori infected, normal, and neoplasticgastroduodenal mucosa.Clin Pathol 1994; 47:6 1-6.
41 O'ConnorHJ, DixonMF, Wyatt JI, Axon AT, Ward DC,Dewar EP, etal. Effect of duodenal ulcer surgery andenterogastric reflux on Campylobacter pyloridis. Lancet1986; ii: 1178-81.
203
group.bmj.com on July 6, 2017 - Published by http://gut.bmj.com/Downloaded from

gastritis.associatedcomplement in Helicobacter pylori
Epithelium related deposition of activated
A E Berstad, P Brandtzaeg, R Stave and T S Halstensen
doi: 10.1136/gut.40.2.1961997 40: 196-203 Gut
http://gut.bmj.com/content/40/2/196Updated information and services can be found at:
These include:
serviceEmail alerting
box at the top right corner of the online article. Receive free email alerts when new articles cite this article. Sign up in the
CollectionsTopic Articles on similar topics can be found in the following collections
(218)Helicobacter pylori (242)Campylobacter, Salmonella, Shigella, Escherichia coli
(1689)Stomach and duodenum
Notes
http://group.bmj.com/group/rights-licensing/permissionsTo request permissions go to:
http://journals.bmj.com/cgi/reprintformTo order reprints go to:
http://group.bmj.com/subscribe/To subscribe to BMJ go to:
group.bmj.com on July 6, 2017 - Published by http://gut.bmj.com/Downloaded from